
Submitted:
25 January 2025
Posted:
27 January 2025
You are already at the latest version
Abstract
Essential oils are widely recognized for their antifungal properties, their effectiveness against fungi fungal remains an area of active research. This study evaluates the antifungal potential of essential oils from oregano (Origanum vulgare), rosemary (Salvia rosmarinus), clove (Syzygium aromaticum), thyme (Thymus), cinnamon (Cinnamomum verum), and basil (Ocimum basilicum) in inhibiting the growth of fungi associated with banana rot, Fusarium spp., Colletotrichum spp., and Verticillium spp. Dilutions containing all the relevant fungal species were prepared and inoculated onto PDA medium supplemented with chloramphenicol for fungal growth assessment. Subcultures were performed weekly to obtain isolated and purified fungal strains, followed by macroscopic and microscopic analysis to identify the genera. Growth inhibition was quantified through in vitro inhibition and ex vivo. Results revealed that thyme exhibited the highest antifungal activity, with complete inhibition at 1000 ppm, while the cinnamon and oregano essential oils showed effective inhibition at 600 ppm. In contrast, rosemary, basil, and clove did not demonstrate significant inhibitory effects at the tested concentrations. These findings suggest that specific essential oils offer a natural alternative for controlling banana rot fungi, supporting their potential use in integrated disease management strategies.
Keywords:
oregano
; rosemary
; clove
; thyme
; basil
; cinnamon
; banana
1. Introduction
Bananas (Musa paradisiaca) are among the most significant crops globally due to their substantial economic and nutritional value, particularly in tropical and subtropical regions. Ecuador is one of the leading banana exporters, the production of bananas serves as a cornerstone of both the local and national economy [1,2]. Known for their high quality, flavor, and nutritional content (including vitamins, minerals, and carbohydrates) [3].
Despite their importance, banana production faces significant challenges, particularly in postharvest stages with fungal diseases such as crown rot, vascular wilt, and fruit decay, caused by various fungal, compromise both the quality and shelf life of the fruit [4,5,6]. These diseases are responsible for considerable economic losses each year and hinder global banana trade, especially in regions with high humidity and warm temperatures that favor fungal proliferation [7,8]. Among the fungus are Fusarium pseudocircinatum, Fusarium verticilloides, Colletotrichum tengchongense, Fusarium napiforme, and Verticillium dahliae, which affect the banana plant is vascular system, leading to reduced fruit yield and quality [9,10,11].
The control of these fungal pathogens has traditionally relied on synthetic fungicides, but growing concerns about their environmental impact, development of fungicide resistance, and potential health risks have spurred interest in finding alternative, more sustainable methods [12,13]. The essential oils (EOs), derived from plants such as Origanum vulgare (oregano), Syzygium aromaticum (clove), Cinnamomum verum (cinnamon), Thymus vulgaris (thyme), and Salvia rosmarinus (rosemary), have emerged as promising natural antifungal agents [14,15,16].
These oils contain bioactive compounds such as carvacrol, eugenol, cinnamaldehyde, and thymol, which have demonstrated potent antimicrobial and antifungal properties in various agricultural contexts [17,18]. However, while essential oils have been studied for their antimicrobial activity, there remains a significant gap in the literature regarding their effectiveness specifically against the fungus pathogens affecting bananas, particularly under postharvest conditions [1,19].
This research aims to evaluate the antifungal efficacy of these essential oils against Fusarium pseudocircinatum, Colletotrichum tengchongense, Suarium napiforme, Fusarium verticilloides, and Verticillium dahliae, isolated from banana peels. Essential oils are complex mixtures of volatile compounds, primarily terpenoids, phenolic compounds, and alcohols, each contributing to their bioactivity [20,21].
Among the essential oils considered in this study, oregano oil (Origanum vulgare) stands out due to its high carvacrol content (60-80%), a phenolic compound known for its potent antimicrobial and antioxidant properties [15,22]. Carvacrol has been shown to disrupt fungal cell membranes by integrating into the lipid bilayer, increasing membrane permeability and leading to cell death [23], thymol shares similar mechanisms of action, p-cymene and γ-terpinene, also contribute synergistically to its antifungal activity [24].
Syzygium aromaticum (clove) essential oil contains eugenol (70-85%), a compound with antifungal properties. Eugenol acts by denaturing proteins, including enzymes critical to fungal metabolism, and by inhibiting vital cellular processes such as ATP synthesis and cellular respiration, ultimately resulting in fungal cell death [20,25]. The antioxidant activity was IC50 values of 4.82 ± 0.06 × 10-2 µg/mL and EC50 values of 3.47 ± 0.2 × 10-2 µg/mL. The results indicate that essential oil exhibits significant in vitro antioxidant activity and moderate antibacterial activity [26]
Cinnamon essential oil (Cinnamomum verum), rich in cinnamaldehyde (60-70%) and eugenol (5-15%), is another promising candidate for antifungal control [17,27]. Cinnamaldehyde disrupts fungal cell membranes and inhibits ergosterol biosynthesis, a key component of the fungal cell membrane, thereby impairing fungal growth and nutrient transport, has been shown to be effective against Fusarium species wilt in bananas [28]. A study examined the antifungal activity of essential oils and their compounds on the growth of Fusarium spp. isolated from papaya rot [29]
Thyme oil (Thymus vulgaris), which contains high conntrations of thymol (30-50%) and carvacrol (10-20%), has demonstrated broad-spectrum antifungal activity by integrating into fungal cell membranes, causing membrane leakage and disrupting the integrity of the cell [30,31]. Ocimum basilicum, or basil, is also from the Lamiaceae family and is rich with bioactive compounds such as eugenol, rosmarinic acid, and flavonoids and is primarily valued for its antioxidant properties. Additionally, offers anticancer, antimicrobial, anti-inflammatory, and antioxidant benefits [32].
Finally, rosemary oil (Salvia rosmarinus), with 1,8-cinee (20-50%) and carnosol (5-15%), induces oxidative stress in fungal cells. The generation of reactive oxygen species (ROS)leads to oxidative damage to fungal proteins, lipids, and DNA, accelerating fungal cell death [14].
The antifungal effects of these essential oils are primarily explained by their ability to disrupt the fungal cell membrane, inhibit ergosterol biosynthesis, and induce oxidative stress. Disruption of the fungal cell membrane increases its permeability, allowing ions, proteins, and essential metabolites to leak out, ultimately resulting in the death of the cell [21]. Cinnamaldehyde, carvacrol, and thymol, in particular, have been shown to disrupt ergosterol biosynthesis, which is essential for maintaining the structure and function of the fungal membrane [33].
Rosmarinic acid in rosemary oil and eugenol in basil oil induce the production of reactive oxygen species ROS, which causes oxidative damage to cellular components, leading to cell death [14,16]. These mechanisms of action are particularly relevant for the control of Fusarium, Colletotrichum and Verticillium species, which are responsible for banana postharvest diseases.
While essential oils show great promise as natural antifungal agents, it is crucial to consider their potential phytotoxicity. High concentrations of essential oils can cause damage to plant tissues, leading to symptoms such as chlorosis, necrosis, and reduced growth, particularly when applied directly to the plant [12]. Previous studies have highlighted the importance of determining the optimal concentration for antifungal efficacy, ensuring that essential oils can be used effectively without harming the plant.
Emulsions containing essential oils in concentrations ranging from 0.1-1% have been reported as safe for use on postharvest fruits, is necessary specific tolerance tests on fruit of banana, are necessary to ensure the safety of these concentrations. It is essential to balance antifungal efficacy with phytotoxicity to ensure the safe and effective application of essential oils [21].
In conclusion, this study evaluates the antifungal potential of essential oils applicate in postharvest of banana, against Fusarium pseudocircinatum, Colletotrichum tengchongense, Suarium napiforme, Fusarium verticilloides, and Verticillium dahliae affecting bananas. This research will contribute valuable insights into the potential use of essential oils as natural, sustainable alternatives to synthetic fungicides.
2. Materials and Methods
2.1. Isolation and Purification of Microorganisms
In this study, Musa paradisiaca was used, specifically Ecuadorian bananas of export quality. The fruits were exposed to environmental conditions until visible signs of deterioration developed, including fungal growth on the peel covering at least 50% of the banana’s surface
The isolation and purification of pathogenic fungi from infected banana peels (Musa paradisiaca) were conducted using Potato Dextrose Agar (PDA) medium (Difco™), which is widely recognized for its ability to support fungal growth. PDA was prepared by dissolving 39 g of the medium in 1 L of distilled water, followed by sterilization in an autoclave at 121°C for 15 minutes [34,35]. To prevent bacterial contamination, chloramphenicol (Merck, Ecuador) was added at a final concentration of 0.5 g/L [36,37].
Samples were collected from 30 bananas showing visible signs of fungal infection on the peel. Approximately 20 g of infected tissue were aseptically removed from each banana. The peel fragments were immersed in sterile distilled water and manually agitated for 2 minutes. This process was repeated twice with fresh sterile water, discarding the used water after each step [38,39,40].
For spore extraction, the cleaned peel fragments were transferred to 200 mL Erlenmeyer flasks containing a 0.05% (v/v) aqueous solution of Tween 80. This surfactant facilitated the release of fungal spores from the infected tissue. The suspension was vortexed for 2 minutes to ensure homogeneity [41,42]. Four serial dilutions were prepared from the initial suspension, using a 0.1% aliquot of the previous dilution at each step to ensure proper spore distribution mixing in a vortex [43].
Aliquots of 0.1 mL from each dilution were plated onto Petri dishes containing PDA supplemented with 0.05% chloramphenicol. The plates were incubated at 25°C and monitored every 48 hours to observe fungal colony development [44]. Visible fungal colonies were carefully selected with a sterile loop to avoid cross-contamination and were subcultured weekly onto fresh PDA plates until pure cultures were obtained [43,45].
2.2. Morphological Identification
The purified fungal strains were analyzed macroscopically and microscopically to confirm their identity. Pure cultures were maintained on PDA at 25°C for short-term studies and preserved at -80°C in a 15% glycerol solution for future molecular studies.
The morphological identification of fungal pathogens involved a comprehensive analysis of macroscopic and microscopic characteristics to determine their genus [40]. Pure fungal cultures were prepared and evaluated weekly over a four-week period, with each analysis conducted in triplicate to ensure reproducibility. These evaluations were performed following inoculation on potato dextrose agar (PDA) medium. The macroscopic examination focused on key morphological traits, including colony shape, elevation, edges, color, texture, and surface appearance [46].
Observations of the upper and lower surfaces of the Petri dishes provided additional insights into fungal colony characteristics. The recorded macroscopic features were systematically compared against bibliographic information obtained from specialized literature, such as books and guides on fungal morphology, to identify the genus of the pathogens [5,47].
Microscopic analysis were conducted to observe fungal reproductive structures, including hyphae, conidia, mycelium, and spores. For this, a conventional method using adhesive tape was employed to collect the aerial mycelium, which was then mounted on microscope slides. These slides were examined for triplicate under a compound microscope with 40X and 60X objective lenses. Detailed observations focused on hyphae, spore formation, and other specialized microscopic structures. Results were cross-referenced with descriptive fungal morphology guides to confirm the findings [48].
2.3. Molecular Identification Through DNA Sequencing
Fungal species isolated from Musa paradisiaca (banana) were identified molecularly using DNA sequencing methods to ensure precise species identification. Genomic DNA was extracted from pure fungal colonies using the Invitrogen commercial kit, strictly adhering to the manufacturer’s instructions. To evaluate the quality and quantity of the extracted DNA, spectrophotometry was conducted using a Nanodrop spectrophotometer, alongside 1% agarose gel electrophoresis to verify DNA integrity [49].
The ribosomal DNA fragment corresponding to the Internal Transcribed Spacer (ITS) region was amplified using the universal primers ITS1 (5’-TCCGTAGGTGAACCTGCGG-3’) and ITS4 (5’-TCCTCCGCTTATTGATATGC-3’). The PCR amplification was optimized in a thermocycler, and the resulting amplicons were visualized on a 1.5% agarose gel stained with ethidium bromide under UV light to confirm amplification. Purified amplicons were subsequently sent to Macrogen Inc. for sequencing [41].
The obtained sequences were analyzed using BioEdit software for alignment and further comparison with publicly available databases, such as the NCBI GenBank. The BLASTn algorithm was employed to identify fungal species based on sequence similarity. Identification was confirmed with a threshold of ≥98% similarity to the deposited sequences in the database [36,41].
2.4. Ex Vivo Fungal Activity
The antifungal activity of fungal pathogens isolated from Musa paradisiaca (banana) was assessed in an ex vivo analysis under controlled conditions of approximately 13°C y 95% humidity relative. The fungi analyzed included Fusarium pseudocircinatum, Fusarium verticilloides, Colletotrichum tengchongense, Fusarium napiforme, and Verticillium dahliae, all of which were isolated from banana peel samples. These pathogens were selected based on their relevance in postharvest diseases of bananas during storage of 15 fungi total.
Fungi were stored for 7 to 10 days before analysis, and inoculum concentrations were standardized to 10⁶ conidia/mL of each inoculum, to maintain consistent infection levels [28]. Bananas harvested at physiological maturity were selected for uniform size, color, and absence of visible damage. The fruits were disinfected with a 1% sodium hypochlorite solution for 5 minutes, rinsed with sterile distilled water, and dried at room temperature [50]. Inoculation was performed using the wound method, where 100 µL of the adjusted fungal inoculum was applied. The fruits were maintained at approximately 13 °C and 95% relative humidity [51].
Observations were recorded at regular intervals to measure lesion diameter and fungal growth diameter using a millimeter ruler, with four replicates for each treatment. Fungal growth diameter was monitored weekly for up to 6 weeks post inoculation, providing a comprehensive timeline of pathogen progression. The inhibition index and severity of infection were calculated in triplicate, based on the diameter of fungal growth, this analysis enabled the classification of pathogens.
2.5. In Vitro Antifungal Activity with Essential Oils
The in vitro antifungal activity of essential oils derived from six plant species was evaluated under controlled laboratory conditions. The essential oils included oregano (Origanum vulgare), rosemary (Salvia rosmarinus), clove (Syzygium aromaticum), thyme (Thymus vulgaris), cinnamon (Cinnamomum verum), and basil (Ocimum basilicum). These oils were extracted using steam distillation, a standard method for isolating bioactive compounds from plants.
Specifically, oregano oil was obtained from dried leaves, cinnamon oil from dried bark, and clove oil from dried flower buds, while rosemary, basil, and thyme oils were extracted from fresh leaves and flowers. The oils were procured from commercial suppliers and diluted in a 0.05% Tween 80 solution to form homogeneous emulsions with concentrations of 200, 400, 600, 800, and 1000 ppm [43].
The emulsions were incorporated into cooled Potato Dextrose Agar (PDA) medium before solidification. The medium was inoculated with fungal pathogens and incubated at 25°C to observe the inhibitory effects of the essential oils on fungal growth. Negative controls included PDA medium without essential oils, allowing for baseline fungal growth comparisons.
Each experimental condition was conducted in quadruplicate, and the percentage of growth inhibition was assessed every 48 hours. Observations focused on determining the concentration of each essential oil that effectively inhibited fungal growth. The study employed a 6 × 5 mixed factor model to analyze the antifungal activity, with six types of essential oils and five concentration levels as independent variables, and the percentage of growth inhibition as the dependent variable.
Analyses identified the most effective essential oil and concentration for fungal inhibition. Results demonstrated significant variability in the antifungal efficacy of oils, providing valuable insights into their potential applications as natural antifungal agents in biocontrol and agriculture. By evaluating a diverse range of essential oils at varying concentrations, this study contributes to the development of eco-friendly alternatives for managing pathogens.
3. Results
3.1. Morphological Identification
Table 1 presents the pure fungi isolated from banana peel rot on a selective medium (PDA + Chloramphenicol), stored in PDA at approximately 25°C. It also includes macroscopic images of the front and reverse sides of Fusarium pseudocircinatum, Colletotrichum tengchongense, Fusarium napiforme, Fusarium verticilloides, and Verticillium dahliae.
Table 2 shows the aerial mycelium of the fungi. These observations offer visual details that complement the morphological analysis and microscopic images viewed under 40X and 60X magnifications.
3.2. Molecular Identification Through DNA Sequencing
Table 3 summarizes the results of the ITS region sequencing of Fusarium pseudocircinatum, Colletotrichum tengchongense, Fusarium napiforme, Fusarium verticilloides, and Verticillium dahliae fungi isolated from banana peels, listing the identified fungal organisms, the genetic fragments used, and the similarity percentages obtained through comparison with the database. The high similarity of the sequences (≥98%) ensures the reliability and accuracy of the identification of the fungal genera and species present in the analyzed samples.
3.3. Fungal Activity Ex Vivo
The severity of fungal infection in banana samples was assessed through ex vivo analysis, where 20 banana samples were monitored over a 6-week period. Fungal growth was evaluated for various species, including Fusarium pseudocircinatum, Colletotrichum tengchongense, Fusarium napiforme, Fusarium verticilloides, and Verticillium dahlia.
Figure 1 provides a visual representation of this growth over time. The analysis utilized an ANOVA approach to examine the impact of different treatments and concentrations on fungal development. Statistically significant differences were observed between treatments (p < 0.05), suggesting that both the type of treatment and its concentration have a notable influence on fungal growth.
3.4. In Vitro Antifungal Activity with Essential Oils
Figure 2, Figure 3, Figure 4, Figure 5, and Figure 6 illustrate the growth of Fusarium pseudocircinatum, Colletotrichum tengchongense, Fusarium napiforme, Fusarium verticilloides, and Verticillium dahliae, in PDA medium with essential oil concentrations of 200, 400, 600, 800, and 1000 ppm. The essential oils evaluated include oregano, basil, cinnamon, rosemary, thyme, and clove, showing their effects on fungal growth at different concentrations.
4. Discussion
4.1. Morphological identification
Regarding the macroscopic analysis in the Table 1 and Table 2, the colonies of Fusarium pseudocircinatum on PDA medium exhibited a cottony, slightly elevated growth with lobed edges. The surface of the colonies was dry, with a cottony and moderately rough texture, showing colors ranging from beige to gray. The underside of the colony also displayed a cottony texture and similar colors, predominantly white and beige tones. This macroscopic pattern is consistent with previous descriptions of Fusarium species [52,53].
In Colletotrichum tengchongense, the colonies showed globes to irregular growth, with a cottony texture and colors varying from beige to black. The colonies had a slightly moist or dry surface, with well-defined edges and a somewhat rough appearance. These characteristics are consistent with species of the Colletotrichum genus [54]. This growth pattern, with a dense texture and clear color differentiation, helps identify the species within the Colletotrichum genus.
For Fusarium napiforme, the colonies in the PDA medium exhibited crateriform growth, characterized by a cottony and rough texture on the upper surface. The shape of the colonies was crater-like, with a depressed center surrounded by slightly raised edges. The surface was white to beige, while the underside of the colonies showed darker tones, ranging from brown to dark brown. This crateriform growth pattern, combined with the cottony texture, is distinctive for Fusarium napiforme [53]. The elevated and lobed colony appearance, along with the darker coloration of the bottom side, helps in distinguishing this species from other Fusarium species [55].
For Fusarium verticilloides, the colonies on PDA displayed cottony growth with a rough and elevated texture. The colonies were typically white to beige, with central areas darkening to brown or black as the colony aged. This growth pattern is characteristic of Fusarium verticilloides [56]. Finally, for Verticillium dahliae, the colonies on PDA exhibited cottony growth with a soft texture, with a circular shape and well-defined edges. The color varied from white on the outer areas to darker tones, such as black, in the center of the colony. This color variation is typical of Verticillium dahliae [10,57].
The microscopic analysis provided additional information for the identification of these species. Fusarium pseudocircinatum showed hyaline, septate, and thin hyphae, with conidia that were cylindrical to slightly curved, organized in branched conidiophores, typical of this species [53]. In Colletotrichum tengchongense, microscopic observations revealed hyaline, septate hyphae, and conidia that were cylindrical to curved, arranged in branched conidiophores, which is a distinguishing feature of the Colletotrichum genus [58]. The arrangement of hyphae and conidia is helpful for confirming the species identity, as it is consistently seen in other species of this genus [59].
For Fusarium verticilloides, the hyphae were hyaline and septate, while the conidia had a fusiform shape, which is typical for this species [53]. The conidia were organized in branched conidiophores, a key feature for identifying Fusarium species. In Fusarium napiforme, the hyphae were hyaline and septate, and the conidia observed were fusiform or ellipsoidal, the arrangement of conidia along the hyphae and their fusiform [53]. In Verticillium dahliae, the microscopic observations revealed hyaline, septate hyphae, and conidia that were cylindrical or oval, arranged in branched structures [57]. The branched conidial structure is a distinctive feature of Verticillium.
4.2. Molecular Identification Through DNA Sequencing
Molecular sequencing of the ITS fragment provided additional confirmation for the identification of the studied species. Fusarium pseudocircinatum showed a 99.55% identity, indicating high concordance with reference sequences in the databases, confirming the molecular identification of the species [56]. This high identity percentage further strengthens the accuracy of macroscopic and microscopic analyses.
For Colletotrichum tengchongense, sequencing revealed a 99.58% identity, validating the molecular identification of this species [60]. The high degree of identity with reference sequences indicates that the macroscopic and microscopic results obtained are consistent with the molecular data. Fusarium verticilloides, the sequencing analysis of the ITS revealed a 99.11% identity, confirming the correct molecular identification of the species [56].
For Fusarium napiforme, the ITS sequencing showed a 98.92% identity, further corroborating the molecular identification of this species, the comparison with reference sequences allowed for the confirmation of its identity [61]. Finally, Verticillium dahliae showed a 99.04% identity in the ITS analysis, which also validated the correct molecular identification of the species [62].
In conclusion, the macroscopic, microscopic, and molecular analyses provided a reliable identification of the fungal species studied. The combination of these methods is important for the correct classification.
4.3. Ex Vivo Fungal Activity
This study evaluated the weekly growth of five fungal species inoculated into bananas, observing variability in both the rate of growth and the severity of colonization over time. Fusarium pseudocircinatum displayed growth reaching approximately 0.6 cm at six weeks. This suggests that, while not the most aggressive species, it remains a significant pathogen in banana storage contexts. The slow growth observed in the initial weeks, followed by a steady increase, is consistent with its ability to adapt to the environment before proliferating. The relatively high standard deviations indicate some variability in growth between the inoculated samples, which could reflect heterogeneity in storage conditions or intrinsic biological variability of the species. This behavior is also in line with previous studies, which suggest that species of Fusarium, such as Fusarium pseudocircinatum, can induce decay when environmental conditions are not adequately controlled [53].
Colletotrichum tengchongense exhibited the fastest growth among all species studied. By week six, it reached approximately 0.9 cm, standing out for its aggressiveness in colonization. This pattern is characteristic of Colletotrichum species, known for their ability to produce rapidly dispersing spores that efficiently colonize plant tissue, especially under high humidity conditions [63,64]. The growth graph shows relatively low standard deviations, suggesting that this pathogen colonized the samples more uniformly. This behavior aligns with previous research that highlights Colletotrichum as one of the most aggressive pathogens in tropical fruits like banana, due to its high ability to produce enzymes that degrade plant tissue [51,59].
Fusarium napiforme presented intermediate growth, reaching approximately 0.5 cm by the end of six weeks. Its colonization rate was not as rapid as Colletotrichum tengchongense, but it remained constant throughout the observation period. The standard deviations suggest that, while its growth was slower compared to the more aggressive species, the inoculated samples showed a relatively uniform response. Describes Fusarium napiforme as a less invasive pathogen than other Fusarium species, but one that can still cause significant damage under improper storage conditions [61].
Fusarium verticilloide exhibited the fastest growth from week two, reaching a colony size of approximately 0.8 cm by week six. Its ability to rapidly adapt to storage conditions is highlighted by its acceleration in the growth rate, making it a high-priority pathogen for banana postharvest management. The growth graph shows a marked acceleration toward the later weeks, with higher standard deviations during the initial stages, which could reflect variability in initial colonization. The growth stabilized, further underscoring its ability to rapidly adapt and occupy the available substrate [65].
Verticillium dahliae the least growth among the species, with limited colonization at approximately 0.4 cm by the end of six weeks. This reflects its reduced ability to colonize banana tissue under storage conditions. While Verticillium dahliae is a known pathogen in other crops, its slower growth in bananas could indicate that it is less efficient in colonizing tropical fruits compared to other species like Colletotrichum or Fusarium. The standard deviations suggest that the growth of Verticillium dahliae was relatively uniform among the inoculated samples, but its lower aggressiveness could make it a less immediate threat in banana [62].
The study highlights the importance of considering the specific biological characteristics of each fungal species when developing control strategies for banana storage. The more aggressive species, such as Colletotrichum tengchongense and Fusarium verticilloides, exhibited rapid and sustained growth, emphasizing the need for more stringent control measures to prevent colonization and decay. Conversely, species such as Fusarium pseudocircinatum and Fusarium napiforme, while less aggressive, remain relevant in terms of their potential for proliferation under inadequate storage conditions.
4.4. In Vitro Antifungal Activity with Essential Oils
The antifungal activity of essential oils was evaluated in vitro for oregano (Origanum vulgare), rosemary (Salvia rosmarinus), clove (Syzygium aromaticum), thyme (Thymus vulgaris), cinnamon (Cinnamomum verum), and basil (Ocimum basilicum). These essential oils of varying concentrations (200, 400, 600, 800, and 1000 ppm), were extracted using steam distillation. The most effective concentration was determined through an analysis conducted with four replicates examined every 48 hours to monitor the inhibition and the effectiveness of the essential oil at the given concentration. Table 3 presents the in vitro evaluation of antifungal activity, "+" sign indicates fungal growth, while a "-" sign denotes no fungal growth.

Table 3.
Evaluation of antifungal activity in vitro of Fusarium pseudocircinatum, Fusarium verticilloides, Colletotrichum tengchongense, Fusarium napiforme, and Verticillium dahliae using oregano, rosemary, clove, thyme, cinnamon and basil essential oils.
Table 3.
Evaluation of antifungal activity in vitro of Fusarium pseudocircinatum, Fusarium verticilloides, Colletotrichum tengchongense, Fusarium napiforme, and Verticillium dahliae using oregano, rosemary, clove, thyme, cinnamon and basil essential oils.
| Essential oil | Fungus | Concentration [ppm] | ||||
| 200 | 400 | 600 | 800 | 1000 | ||
| Cinnamon | Fusarium pseudocircinatum | + | + | - | - | - |
| Colletotrichum tengchongense | - | - | - | - | - | |
| Fusarium verticilloides | + | - | - | - | - | |
| Fusariumnapiforme | - | - | - | - | - | |
| Verticillium dahliae | - | - | - | - | - | |
| Clove | Fusarium pseudocircinatum | + | + | + | + | + |
| Colletotrichum tengchongense | + | + | - | - | - | |
| Fusarium verticilloides | + | + | - | - | - | |
| Fusariumnapiforme | - | - | - | - | - | |
| Verticillium dahliae | + | - | - | - | - | |
| Basil | Fusarium pseudocircinatum | + | + | + | + | + |
| Colletotrichum tengchongense | + | + | + | + | + | |
| Fusarium verticilloides | + | + | + | + | + | |
| Fusariumnapiforme | + | + | + | + | - | |
| Verticillium dahliae | + | + | + | + | + | |
| Oregano | Fusarium pseudocircinatum | + | - | - | - | - |
| Colletotrichum tengchongense | + | + | - | - | - | |
| Fusarium verticilloides | + | - | - | - | - | |
| Fusariumnapiforme | - | - | - | - | - | |
| Verticillium dahliae | - | - | - | - | - | |
| Rosemary | Fusarium pseudocircinatum | + | + | + | + | + |
| Colletotrichum tengchongense | + | + | + | + | + | |
| Fusarium verticilloides | + | + | + | + | + | |
| Fusariumnapiforme | + | + | + | - | - | |
| Verticillium dahliae | + | + | + | + | + | |
| Thyme | Fusarium pseudocircinatum | + | + | + | + | - |
| Colletotrichum tengchongense | + | + | - | - | - | |
| Fusarium verticilloides | + | + | - | - | - | |
| Fusariumnapiforme | - | - | - | - | - | |
| Verticillium dahlia | + | - | - | - | - | |
The results obtained with Fusarium pseudocircinatum show that oregano oils are highly effective at concentrations above 200 ppm, inhibiting the growth of the fungus in PDA medium. This finding is consistent with previous studies that have demonstrated the antifungal activity of these oils, especially in their ability to control various species of the Fusarium genus, known pathogens that cause wilting in bananas [15]. These oils could be useful not only for controlling Fusarium in bananas in general, but also as a tool in post-harvest management, where banana peels are particularly vulnerable to infections.
In the case of Colletotrichum tengchongense, which causes anthracnose in bananas, is inhibited con cinnamon, with 400 ppm of clove, oregano and thyme essential oils. This fungus primarily affects the peel, causing black spots and accelerating the fruit’s deterioration, a phenomenon that has been documented in previous studies [39,60]. Essential oils, particularly cinnamon, have been shown to be effective against various species of Colletotrichum in previous studies [66], to managing anthracnose in bananas, improving post-harvest quality.
The species Fusarium verticilloides, is sensitive to 400 ppm oregano and cinnamon oils. This fungus is responsible for the rot of the peel, a common problem in the post-harvest stage of bananas. The findings of this study coincide with previous research that has reported the effectiveness of oregano oil in inhibiting several Fusarium species [67]. Oregano and cinnamon oils also showed a positive effect against Fusarium napiforme, another pathogenic fungus associated with the banana peel. This finding is consistent with previous studies that identified oregano oil as an effective antifungal agent against various Fusarium species [53].
Verticillium dahliae showed remarkable inhibition by cinnamon, oregano at higher concentrations, and to 200 ppm of thyme and clove essential oils. Verticillium affects banana plants in the field and may continue to proliferate during storage if not properly controlled [62]. The use of essential oils such as oregano, clove, and cinnamon could offer a natural and sustainable alternative to chemical fungicides, with potential benefits for human health and the environment.
5. Conclusions
The fungal species identified (Fusarium pseudocircinatum, Colletotrichum tengchongense, Fusarium napiforme, Fusarium verticilloides, and Verticillium dahliae) exhibit varying levels of aggressiveness. Colletotrichum tengchongense and Fusarium verticilloides are the most invasive, while Fusarium pseudocircinatum and Fusarium napiforme grow more slowly but remain relevant under improper storage conditions, and Verticillium dahliae showed the most limited growth.
The essential oil of oregano (Origanum vulgare) and cinnamon (Cinnamomum verum) effectively inhibited fungal growth at a concentration of 600 ppm.
The essential oils of rosemary (Salvia rosmarinus), basil (Ocimum basilicum), and clove (Syzygium aromaticum) did not inhibit the fungal growth of Fusarium spp., Colletotrichum spp., and Verticillium spp. at the concentrations evaluated.
Autor Contributions
All the mentioned authors have significantly contributed to the development and writing of this article. All authors have read and accepted the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Data will be made available upon reasonable request.
Acknowledgments
The authors thank the support of DECAB – Escuela Politécnica Nacional.
Conflicts of Interest
The authors declare no conflicts of interest
References
- Tang Q, Liu D, Zhang J. Post-harvest control of fungal diseases in bananas using essential oils. Journal of Postharvest Biology 2019, 45, 42–49. https://doi.org/10.1016/j.postharvbio.2019.05.002.
- Mata Anchundia D, Suatunce Cunuhay P, Poveda Morán R. Análisis económico del banano orgánico y convencional en la provincia Los Ríos, Ecuador. Avances 2021, 23, 419–430.
- Borges CV, Amorim EP, Leonel M, Gomez 0Gomez HA, Santos TPR dos, Ledo CA da S, et al. Post-harvest physicochemical profile and bioactive compounds of 19 bananas and plantains genotypes. Bragantia 2018, 78, 284–296.
- Ruiz Medina MD, Ruales J. Postharvest Alternatives in Banana Cultivation. 2024. https://doi.org/10.20944/preprints202402.0783.v1.
- Aguilar-Anccota R, Arévalo-Quinde CG, Morales-Pizarro A, Galecio-Julca M, Aguilar-Anccota R, Arévalo-Quinde CG, et al. Hongos asociados a la necrosis de haces vasculares en el cultivo de banano orgánico: síntomas, aislamiento e identificación, y alternativas de manejo integrado. Scientia Agropecuaria 2021, 12, 249–256. https://doi.org/10.17268/sci.agropecu.2021.028.
- Capa Benítez LB, Alaña Castillo TP, Benítez Narváez RM. Importancia de la producción de banano orgánico. Caso: Provincia de El Oro, Ecuador. Revista Universidad y Sociedad 2016, 8, 64–71.
- Agronomía C. Tecnología Poscosecha. Agronomía Costarricense 2005, 29, 207–209.
- Mari M, Torres R, Vanneste JL. Biological control of postharvest diseases: Opportunities and challenges. Frontiers in Microbiology 2014, 5, 1–17. https://doi.org/10.3389/fmicb.2014.00688.
- Villa-Martínez A, Pérez-Leal R, Morales-Morales HA, Basurto-Sotelo M, Soto-Parra JM, Martínez-Escudero E. Situación actual en el control de Fusarium spp. y evaluación de la actividad antifúngica de extractos vegetales. Acta Agronómica 2015, 64, 194–205. https://doi.org/10.15446/acag.v64n2.43358.
- Goicoechea N. To what extent are soil amendments useful to control Verticillium wilt? Pest Management Science 2009, 65, 831–839. https://doi.org/10.1002/ps.1774.
- Pabon Montoya B, Córdova Chávez M, Alban Alcivar J, Jaramillo Robles A. Efectos antifúngicos de extractos botánicos sobre el crecimiento micelial de Colletotrichum sp. a nivel in vitro, causante de antracnosis en la fruta de aguacate. 593 Digital Publisher CEIT 2024, 9, 869–879.
- Alzate D, Mier G, Afanador L, Durango D, García C. Evaluación de la fitotoxicidad y la actividad antifúngica contra Colletotrichum acutatum de los aceites esenciales de tomillo (Thymus vulgaris), limoncillo (Cymbopogon citratus), y sus componentes mayoritarios 2009, 16.
- Bandoni AL, Retta D, Lira PMDL, Baren CM van. ¿Son realmente útiles los aceites esenciales? Boletín Latinoamericano y del Caribe de Plantas Medicinales y Aromáticas 2009, 8, 317–322.
- Oliva M de LM, Lorello IM, Baglio C, Posadaz A, Carezzano ME, Paletti Rovey MF, et al. Chemical composition, antioxidant, and antimicrobial activities of rosemary (Salvia rosmarinus Spenn.) essential oils from Argentina 2023.
- Pilozo G, Villavicencio-Vásquez M, Chóez-Guaranda I, Murillo DV, Pasaguay CD, Reyes CT, et al. Chemical, antioxidant, and antifungal analysis of oregano and thyme essential oils from Ecuador: Effect of thyme against Lasiodiplodia theobromae and its application in banana rot. Heliyon 2024, 10. https://doi.org/10.1016/j.heliyon.2024.e31443.
- Flores-Villa E, Sáenz-Galindo A, Castañeda-Facio AO, Narro-Céspedes RI, Flores-Villa E, Sáenz-Galindo A, et al. Romero (Rosmarinus officinalis L.): su origen, importancia y generalidades de sus metabolitos secundarios. TIP Revista especializada en ciencias químico-biológicas 2020, 23. https://doi.org/10.22201/fesz.23958723e.2020.0.266.
- Farias APP, Monteiro O dos S, da Silva JKR, Figueiredo PLB, Rodrigues AAC, Monteiro IN, et al. Chemical composition and biological activities of two chemotype-oils from Cinnamomum verum J. Presl growing in North Brazil. J Food Sci Technol 2020, 57, 3176–3183. https://doi.org/10.1007/s13197-020-04288-7.
- Pinto E, Silva C, Costa L. Eugenol as an antifungal agent: Mechanisms and applications. Journal of Applied Microbiology 2018, 124, 1089–1099. https://doi.org/10.1111/jam.13748.
- Abadias M, Teixidó N, Usall J, Viñas I. Evaluation of alternative strategies to control postharvest blue mould of apple caused by Penicillium expansum. International Journal of Food Microbiology 2008, 122, 25–31. https://doi.org/10.1016/j.ijfoodmicro.2007.11.048.
- Haro-González JN, Castillo-Herrera GA, Martínez-Velázquez M, Espinosa-Andrews H. Clove Essential Oil (Syzygium aromaticum L. Myrtaceae): Extraction, Chemical Composition, Food Applications, and Essential Bioactivity for Human Health. Molecules 2021, 26, 6387. https://doi.org/10.3390/molecules26216387.
- Boulanger R, Liu Y, Jiang Z. Synergistic effects of essential oils against fungal pathogens: Mechanisms and applications. Food Control 2021, 123, 107–118. https://doi.org/10.1016/j.foodcont.2021.107118.
- Kintzios SE. 21 - Oregano. In: Peter KV, editor. Handbook of Herbs and Spices (Second Edition), Woodhead Publishing; 2012, p. 417–36. https://doi.org/10.1533/9780857095688.417.
- Choi YH, Kim H, Lee S. The antifungal activity of essential oils: Mechanisms and applications in plant disease management. Phytopathology Research 2018, 44, 89–102. https://doi.org/10.1186/s42055-018-0032-2.
- Arcila-Lozano CC, Loarca-Piña G, Lecona-Uribe S, González de Mejía E. El orégano: propiedades, composición y actividad biológica de sus componentes. Archivos Latinoamericanos de Nutrición 2004, 54, 100–111.
- Prashar A, Locke IC, Evans CS. Cytotoxicity of clove (Syzygium aromaticum) oil and its major components to human skin cells. Cell Proliferation 2006, 39, 241–248. https://doi.org/10.1111/j.1365-2184.2006.00384.x.
- Selles SMA, Kouidri M, Belhamiti BT, Ait Amrane A. Chemical composition, in-vitro antibacterial and antioxidant activities of Syzygium aromaticum essential oil. Food Measure 2020, 14, 2352–2358. https://doi.org/10.1007/s11694-020-00482-5.
- Ruiz M, Ávila J, Ruales J. Diseño de un recubrimiento comestible bioactivo para aplicarlo en la frutilla (Fragaria vesca) como proceso de postcosecha 2016, 17, 276–287.
- Aquino-Martínez JG, Vázquez-García LM, Reyes-Reyes BG. Biocontrol in vitro e in vivo de Fusarium oxysporum Schlecht. f. sp. dianthi (Prill. y Delacr.) Snyder y Hans. con Hongos Antagonistas Nativos de la Zona Florícola de Villa Guerrero, Estado de México. Revista mexicana de fitopatología 2008, 26, 127–137.
- Barrera Necha LL, García Barrera LJ. Actividad antifúngica de aceites esenciales y sus compuestos sobre el crecimiento de Fusarium sp. aislado de papaya ( Carica papaya). Revista Científica UDO Agrícola 2008, 8, 33–41.
- Ultee A, Bennik MHJ, Moezelaar R. The phenolic hydroxyl group of carvacrol is essential for action against the food-borne pathogen Bacillus cereus. Applied and Environmental Microbiology 2002, 68, 1561–1568. https://doi.org/10.1128/aem.68.4.1561-1568.2002.
- Pina-Vaz C, Gonçalves Rodrigues A, Pinto E, Costa-de-Oliveira S, Tavares C, Salgueiro L, et al. Antifungal activity of Thymus oils and their major compounds. Journal of the European Academy of Dermatology and Venereology 2004, 18, 73–78. https://doi.org/10.1111/j.1468-3083.2004.00886.x.
- Shahrajabian MH, Sun W, Cheng Q. Chemical components and pharmacological benefits of Basil (Ocimum basilicum): a review. International Journal of Food Properties 2020, 23, 1961–1970. https://doi.org/10.1080/10942912.2020.1828456.
- Zhao S, Zhang Z, Wei Y. Mechanisms of antifungal action of cinnamaldehyde. Phytochemistry 2020, 173, 112112. https://doi.org/10.1016/j.phytochem.2020.112112.
- Salazar E, Hernández R, Tapia A, Gómez-Alpízar L. Identificación molecular del hongo Colletotrichum spp., aislado de banano (Musa spp) de la altura en la zona de Turrialba y determinación de su sensibilidad a fungicidas poscosecha. Agronomía Costarricense 2012, 36, 53–68.
- Tortora GJ, Funke BR, Case CL. Introducción a la microbiología. Ed. Médica Panamericana; 2007.
- Agu K, Awah N, Nnadozie A. Isolation, identification, and pathogenicity of fungi associated with cocoyam (Colocasia esculenta) spoilage. Food Science and Technology 2016.
- Suárez L, Rangel A. Aislamiento de microorganismos para control biológico de Moniliophthora roreri 2013, 62.
- Orwa P, Mugambi G, Wekesa V, Mwirichia R. Isolation of haloalkaliphilic fungi from Lake Magadi in Kenya. Heliyon 2020.
- Vilaplana R, Pazmiño L, Valencia-Chamorro S. Control of anthracnose, caused by Colletotrichum musae, on postharvest organic banana by thyme oil. Postharvest Biology and Technology 2018, 138, 56–63. https://doi.org/10.1016/j.postharvbio.2017.12.008.
- Morales R, Henríquez G. Aislamiento e identificación del moho causante de antracnosis en Musa paradisiaca l. (plátano) en cooperativa san carlos, el salvador y aislamiento de mohos y levaduras con capacidad antagonista. Crea Ciencia Revista Científica 2021, 13, 84–94. https://doi.org/10.5377/creaciencia.v13i2.11825.
- Funnell-Harris D, Prom L. Isolation and characterization of grain mold fungi Cochliobolus and Alternaria spp. from sorghum using semiselective media and DNA sequence analyses. Canadian Journal of Microbiology 2013.
- Vargas-Fernández JP, Wang-Wong A, Muñoz-Fonseca M, Vargas-Fernández JP, Wang-Wong A, Muñoz-Fonseca M. Microorganismos asociados a la enfermedad conocida como pudrición suave del fruto de banano (Musa sp.) y alternativas de control microbiológicas y químicas a nivel in vitro *. Agronomía Costarricense 2022, 46, 61–76. https://doi.org/10.15517/rac.v46i2.52046.
- Ruiz-Medina M, Ruales J. Essential Oils as an Antifungal Alternative to Control Fusarium spp., Penicillium spp., Trichoderma spp. and Aspergillus spp. 2024. https://doi.org/10.20944/preprints202408.1987.v1.
- Vanegas J Santamaría, González N Comba, Mancilla XC Pérez. Manual de Microbiología General: Principios Básicos de Laboratorio. Editorial Tadeo Lozano; 2014.
- Lopes J, Araújo G, Almeida M. Storage methods of fungi for long-term research. Mycological Studies 2021. https://doi.org/10.1016/j.mycos.2021.06.001.
- Siller-Ruiz M, Hernández-Egido S, Sánchez-Juanes F, González-Buitrago JM, Muñoz-Bellido JL. Métodos rápidos de identificación de bacterias y hongos. Enfermedades Infecciosas y Microbiología Clínica 2017, 35, 303–313.
- Acurio Vásconez RD, España Imbaquingo CK, Acurio Vásconez RD, España Imbaquingo CK. Aislamiento, caracterización y evaluación de Trichoderma spp. como promotor de crecimiento vegetal en pasturas de Raygrass (Lolium perenne) y trébol blanco (Trifolium repens). LA GRANJA Revista de Ciencias de la Vida 2017, 25, 53–61. https://doi.org/10.17163/lgr.n25.2017.05.
- Carpio-Coba CF, Noboa-Silva VF, Salazar-Castañeda EP, Lema-Saigua ER. Caracterización macroscópica y microscópica de cuatro especies forestales de la amazonia del sur de Ecuador. Polo del Conocimiento 2021, 6, 886–904. https://doi.org/10.23857/pc.v6i8.2986.
- Bellemain E, Carlsen T, Brochmann C, Coissac E, Taberlet P, Kauserud H. ITS as an environmental DNA barcode for fungi: an in silico approach reveals potential PCR biases. BMC Microbiology 2010, 10, 189.
- Chacón-Cascante A, Crespi JM. Historical overview of the European Union banana import policy. Agronomía Costarricense 2006, 30, 111–127.
- Aguilar-Anccota R, Apaza-Apaza S, Maldonado E, Calle-Cheje Y, Rafael-Rutte R, Montalvo K, et al. Control in vitro e in vivo de Thielavipsis paradoxa y Collettrichum musae cn biofungicidas en frutos de banano orgánico. Manglar 2024, 21, 57–63. https://doi.org/10.57188/manglar.2024.006.
- Leslie JF, Summerell BA. The Fusarium Laboratory Manual. Ames, Iowa: Blackwell Publishing; 2006.
- González A, others. Identification and characterization of Fusarium species from infected plants. Mycological Research 2013, 117, 189–197.
- Cai L, others. Identification of Colletotrichum species using molecular markers. Fungal Diversity 2011, 48, 25–35.
- Carrasco A, Sanfuentes E, Duran A, Valenzuela S. Cancro resinoso del pino: ¿una amenaza potencial para las plantaciones de Pinus radiata en Chile? Gayana Botánica 2016, 73, 369–380. https://doi.org/10.4067/S0717-66432016000200369.
- Marasas WFO, others. Fusarium verticilloides: Morphology and Mycotoxins. Mycological Research 2011, 115, 845–857.
- Inderbitzin P, others. Molecular and morphological characterization of Verticillium dahliae. Fungal Diversity 2011, 50, 105–118.
- Jayawardena R, others. Recent advances in molecular identification of Colletotrichum species. Mycosphere 2016, 7, 509–522.
- Rojo-Báez I, Álvarez-Rodríguez B, García-Estrada R, León-Félix J, Sañudo Barajas JA, Allende R. Situación actual de Colletotrichums spp. en México: Taxonomía, caracterización, patogénesis y control. Revista Mexicana de Fitopatología, Mexican Journal of Phytopathology 2017, 35. https://doi.org/10.18781/R.MEX.FIT.1703-9.
- Wang Y, others. Molecular and morphological characterization of Colletotrichum tengchongense. Mycological Progress 2012, 11, 423–430.
- Zhang H, others. Molecular identification of Fusarium napiforme in diverse habitats. Fungal Ecology 2015, 17, 63–69.
- Jabnoun-Khiareddine H, Mahjoub M, Daami-Remadi M. Morphological Variability Within and Among Verticillium Species Collected in Tunisia. Tunisian Journal of Plant Protection 2010, 5, 19–38.
- Sharma RR, Singh D, Singh R. Biological control of postharvest diseases of fruits and vegetables by microbial antagonists: A review. Biological Control 2009, 50, 205–221. https://doi.org/10.1016/j.biocontrol.2009.05.001.
- Bautista-Baños S, Bosquez-Molina E, Barrera-Necha LL. The use of fungal antagonists to reduce postharvest fruit decay. Mycopathologia 2003, 155, 127–132. https://doi.org/10.1023/A:1022244032461.
- Fusarium verticilloides: Morphology and Mycotoxins. Mycological Research 2011, 115, 845–857.
- Ranasinghe L, Jayawardena B, Abeywickrama K. Fungicidal activity of essential oils of Cinnamomum zeylanicum (L.) and Syzygium aromaticum (L.) Merr et L.M. Perry against crown rot and anthracnose pathogens isolated from banana. Letters in Applied Microbiology 2002, 35, 208–211. https://doi.org/10.1046/j.1472-765X.2002.01165.x.
- Burt S. Essential oils: Their antibacterial properties and potential applications in foods—A review. International Journal of Food Microbiology 2004, 94, 223–253. https://doi.org/10.1016/j.ijfoodmicro.2004.03.022.
Figure 1.
Fungal growth (cm) during 6 weeks in 20 banana samples inoculated with Fusarium pseudocircinatum, Colletotrichum tengchongense, Fusarium verticilloides, Fusarium napiforme, Verticillium dahliae stored at 13°C and 95% HR approximately.
Figure 1.
Fungal growth (cm) during 6 weeks in 20 banana samples inoculated with Fusarium pseudocircinatum, Colletotrichum tengchongense, Fusarium verticilloides, Fusarium napiforme, Verticillium dahliae stored at 13°C and 95% HR approximately.
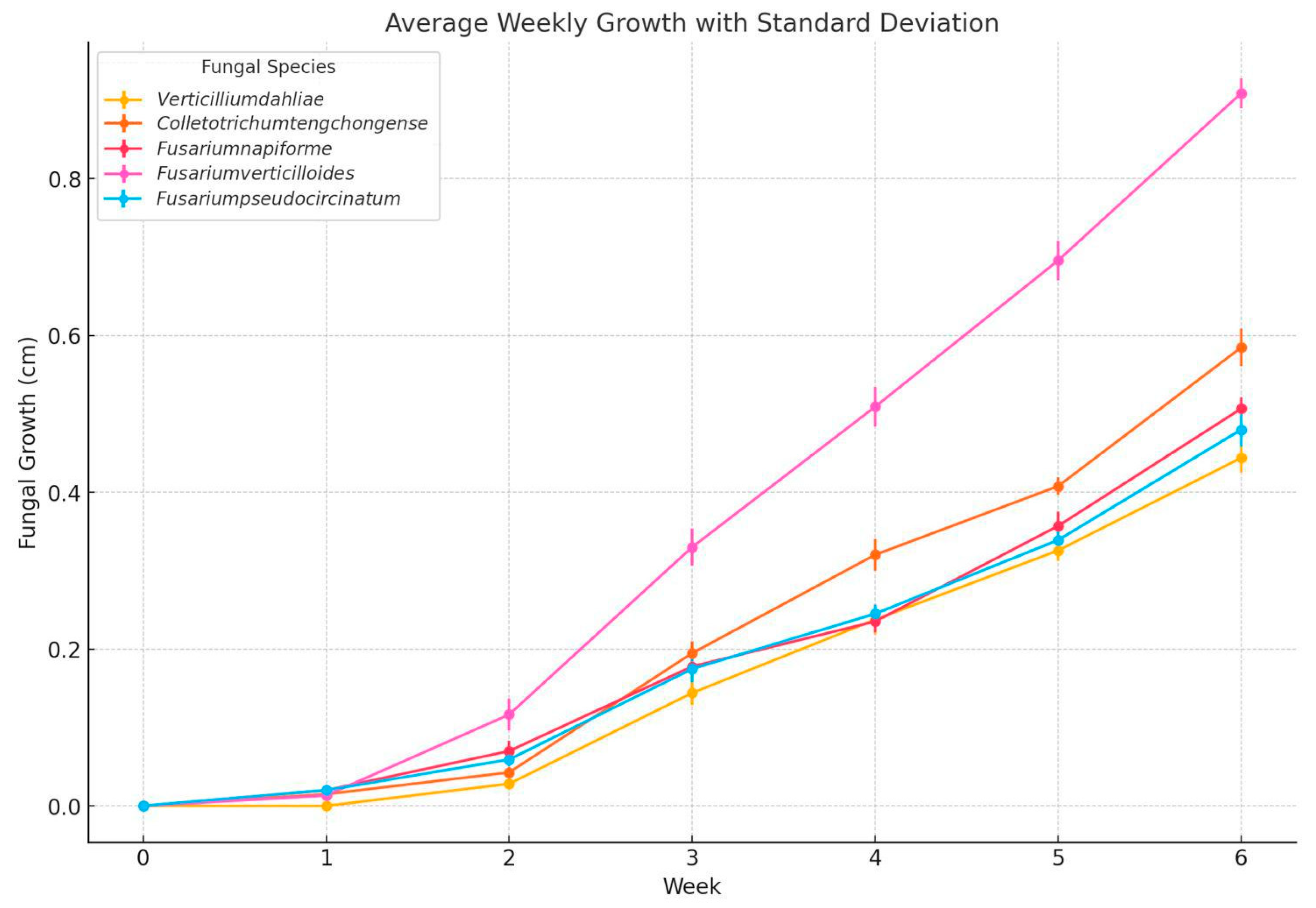
Figure 2.
Analysis in vitro of Fusarium pseudocircinatum in PDA medium with essential oils of basil, cinnamon, clove, oregano, rosemary, and thyme 200, 400, 600, 800 y 1000 ppm, stored to 25°C (n=4).
Figure 2.
Analysis in vitro of Fusarium pseudocircinatum in PDA medium with essential oils of basil, cinnamon, clove, oregano, rosemary, and thyme 200, 400, 600, 800 y 1000 ppm, stored to 25°C (n=4).
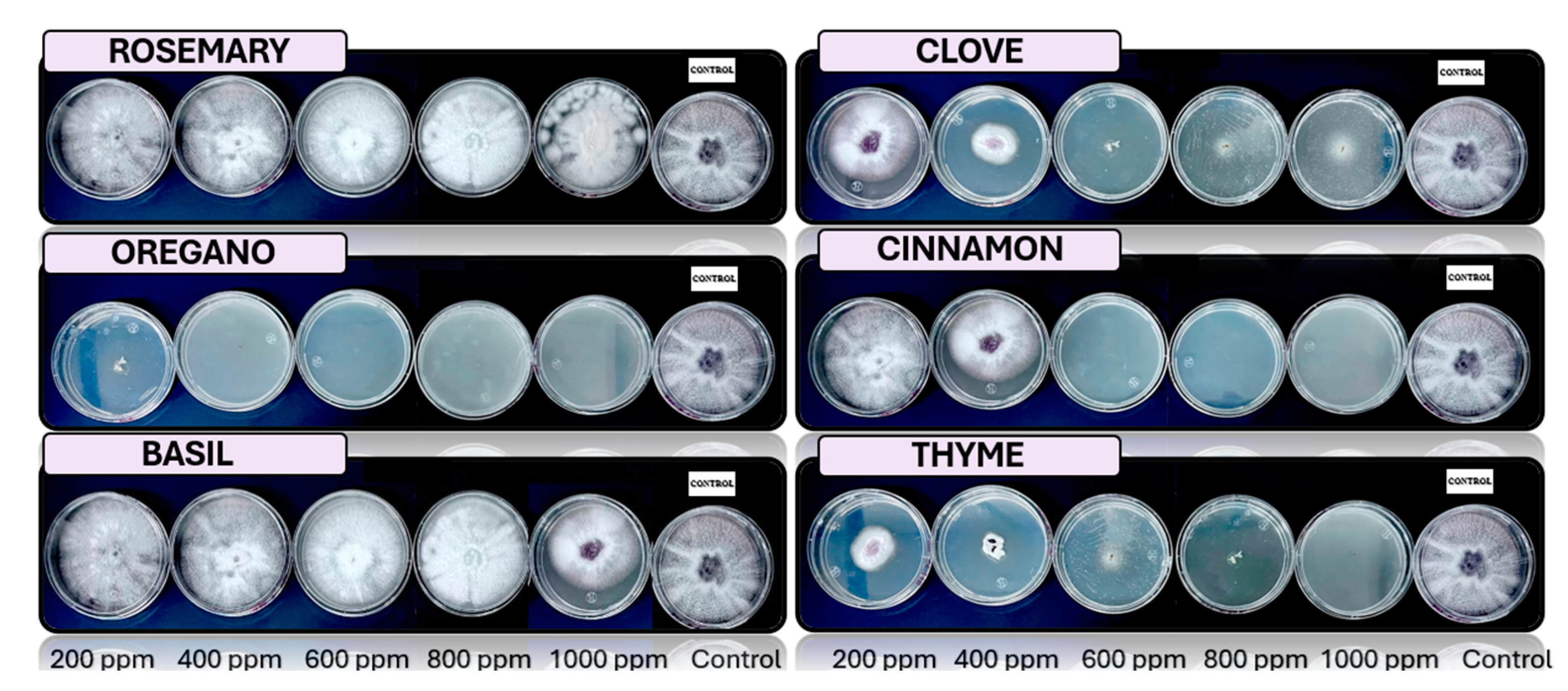
Figure 3.
Analysis in vitro of Colletotrichum tengchongense in PDA medium with essential oils of basil, cinnamon, clove, oregano, rosemary, and thyme 200, 400, 600, 800 y 1000 ppm, stored to 25°C (n=4).
Figure 3.
Analysis in vitro of Colletotrichum tengchongense in PDA medium with essential oils of basil, cinnamon, clove, oregano, rosemary, and thyme 200, 400, 600, 800 y 1000 ppm, stored to 25°C (n=4).
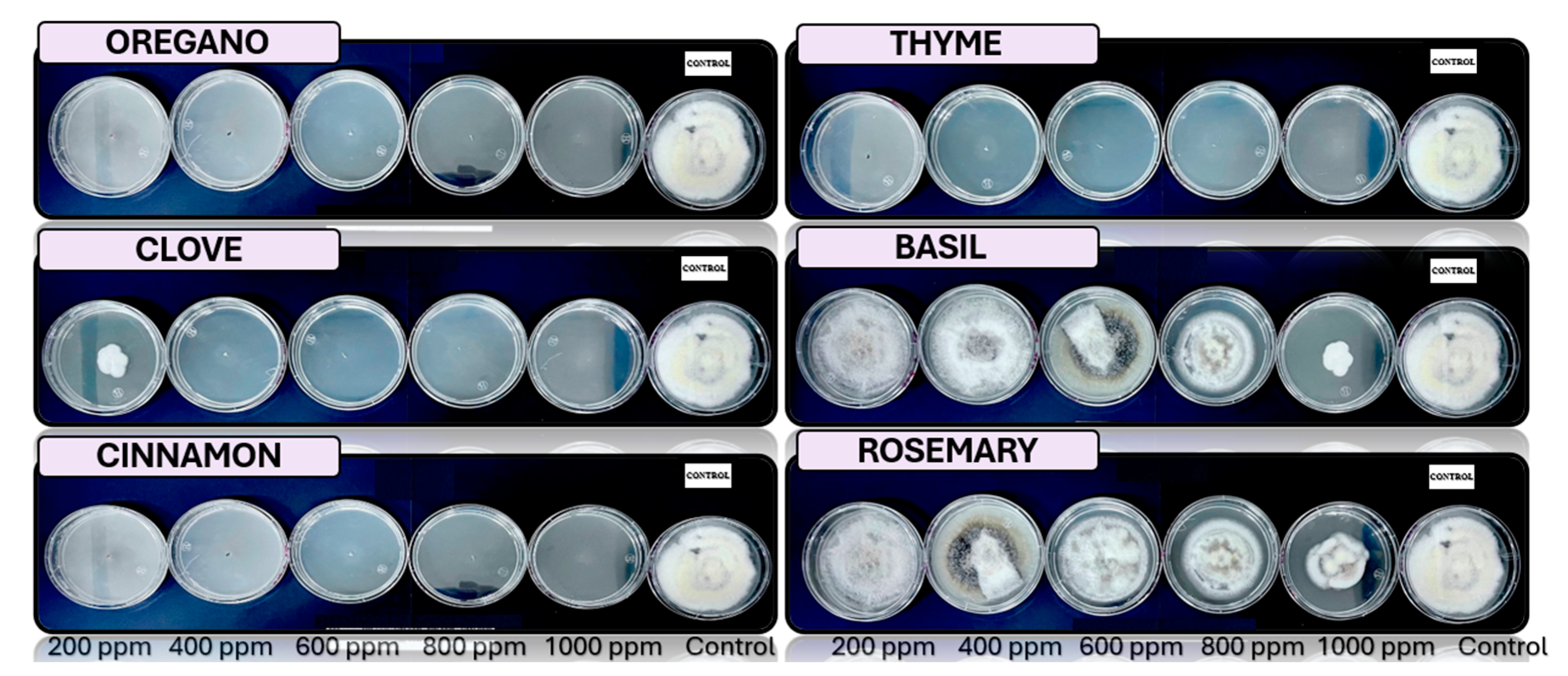
Figure 4.
Analysis in vitro of Fusarium verticilloides in PDA medium with essential oils of basil, cinnamon, clove, oregano, rosemary, and thyme 200, 400, 600, 800 y 1000 ppm, stored to 25°C (n=4).
Figure 4.
Analysis in vitro of Fusarium verticilloides in PDA medium with essential oils of basil, cinnamon, clove, oregano, rosemary, and thyme 200, 400, 600, 800 y 1000 ppm, stored to 25°C (n=4).

Figure 5.
Analysis in vitro of Fusarium napiforme in PDA medium with essential oils of basil, cinnamon, clove, oregano, rosemary, and thyme 200, 400, 600, 800 y 1000 ppm, stored to 25°C (n=4).
Figure 5.
Analysis in vitro of Fusarium napiforme in PDA medium with essential oils of basil, cinnamon, clove, oregano, rosemary, and thyme 200, 400, 600, 800 y 1000 ppm, stored to 25°C (n=4).

Figure 6.
Analysis in vitro of Verticillium dahliae in PDA medium with essential oils of basil, cinnamon, clove, oregano, rosemary, and thyme 200, 400, 600, 800 y 1000 ppm, stored to 25°C (n=4).
Figure 6.
Analysis in vitro of Verticillium dahliae in PDA medium with essential oils of basil, cinnamon, clove, oregano, rosemary, and thyme 200, 400, 600, 800 y 1000 ppm, stored to 25°C (n=4).
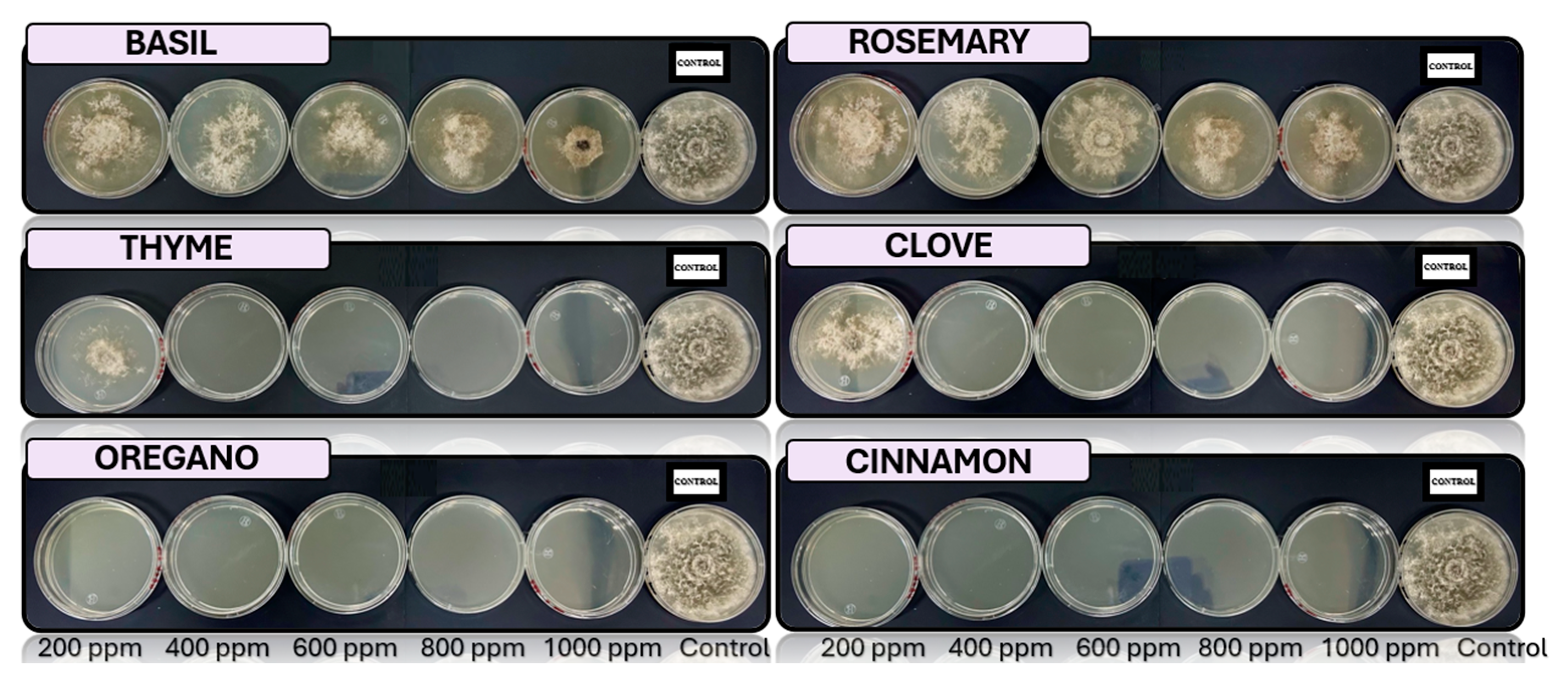
Table 1.
Macroscopy images of (a) Fusarium pseudocircinatum, (b) Colletotrichum tengchongense, (c) Fusarium verticilloides, (d) Fusarium napiforme, (e) Verticillium dahliae considering the appearance of the front side and (f) Fusarium pseudocircinatum, (g) Colletotrichum tengchongense, (h) Fusarium verticilloides, (i) Fusarium napiforme, and (i) Verticillium dahliae of reverse side.
Table 1.
Macroscopy images of (a) Fusarium pseudocircinatum, (b) Colletotrichum tengchongense, (c) Fusarium verticilloides, (d) Fusarium napiforme, (e) Verticillium dahliae considering the appearance of the front side and (f) Fusarium pseudocircinatum, (g) Colletotrichum tengchongense, (h) Fusarium verticilloides, (i) Fusarium napiforme, and (i) Verticillium dahliae of reverse side.
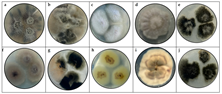 |
Table 2.
Microscopy images of (a) Fusarium pseudocircinatum, (b) Colletotrichum tengchongense, (c) Fusarium verticilloides, (d) Fusarium napiforme, (e) Verticillium dahliae to 40 X and (f) Fusarium pseudocircinatum, (g) Colletotrichum tengchongense, (h) Fusarium verticilloides, (i) Fusarium napiforme and (i) Verticillium dahliae. to 60 X.Table 3.
Table 2.
Microscopy images of (a) Fusarium pseudocircinatum, (b) Colletotrichum tengchongense, (c) Fusarium verticilloides, (d) Fusarium napiforme, (e) Verticillium dahliae to 40 X and (f) Fusarium pseudocircinatum, (g) Colletotrichum tengchongense, (h) Fusarium verticilloides, (i) Fusarium napiforme and (i) Verticillium dahliae. to 60 X.Table 3.
 |
Table 3.
Sequencing results for fungi isolated from the samples of Musa paradisiaca.
| Organism | Fragment | % identity |
| Fusarium pseudocircinatum | ITS | 99.55 % |
| Fusarium napiforme | ITS | 98.92 % |
| Colletotrichum tengchongense | ITS | 99.58 % |
| Fusarium verticilloides | ITS | 99.11 % |
| Verticillium dahliae | ITS | 99.04 % |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



