
Submitted:
26 January 2025
Posted:
27 January 2025
You are already at the latest version
Abstract
Colletotrichum includes agriculturally and scientifically important pathogens that infect numerous plants. They can also adopt an endophytic lifestyle, refraining from causing disease and/or even promoting plant growth when inoculated on a non-susceptible host. In this manner, the host range of a Colletotrichum fungus can shift, depending on whether it exhibits endophytic or pathogenic lifestyles. Here, we aim to disentangle the relationship between lifestyle and host range in Colletotrichum. We discuss the ways in which pathogenic Colletotrichum species may act endophytically on alternative hosts, the molecular mechanisms underlying lifestyle, and the merits of using endophytic fungi alongside pathogenic fungi in research. In particular, we reference the Arabidopsis thaliana-Colletotrichum tofieldiae study system as a potential model for elucidating the dual roles of plant-fungus interactions, both beneficial and pathogenic, through integrative omics approaches and reverse genetics. This approach could identify key molecular targets for effective pathogen management in agriculture. Lastly, we propose an idea in which pathogenic lifestyle occupies a different host range than the endophytic lifestyle. This will enhance our understanding of pathogenicity and endophytism in a globally significant fungal genus and lays the groundwork for future research examining molecular determinants of lifestyle in plant-associated fungi.
Keywords:
Colletotrichum
; plant-microbe interactions
; endophytism
; pathogenicity
; Arabidopsis thaliana
; mycoherbicides
; fungicides
; biofertilizers
1. Introduction
In nature, plants interact with diverse microbes. These can include pathogens (plant detrimental), commensals (plant neutral), and mutualists (plant beneficial) [1]. Commensal and mutualistic microbes that live inside host tissues without visible disease symptoms for at least a portion of their lives, termed endophytes, can serve diverse functions for the host plant, including protection against pathogens [2,3], nutrient acquisition [3,4], priming of the immune system [5], and improvement of abiotic stress tolerance [6]. The term endophyte as we use it here is distinct from the term non-pathogen, which refers to any microbe that does not colonize a host pathogenically. For example, a microbe that is unable to infect a host at all may be termed a non-pathogen of that host, but it would not qualify as an endophyte unless it colonizes the host and its re-isolation and colonization within the host have been thoroughly demonstrated. Pathogens cause plants to lose biomass and nutrients and cause severe yield loss annually to important crop species [7,8,9]. At the same time, it has been gradually recognized that some pathogens can act endophytically depending on the host colonized [10,11], or within the same host but depending on certain conditions, such as the stage of the interaction [10] or the environment (e.g., temperature [12]). It has been established that the conditions of the plant-microbe interaction, such as the environment or the host genotype, can affect the outcome of the interaction [13,14]. For example, elevated temperature can suppress effector-triggered immunity (ETI) in plants and cause pathogens with avirulent effectors to become more virulent [15,16]. However, there is relatively less discussion of the fact that the conditions of the interaction can also determine the lifestyle of the microbe as it infects its host. For example, increasing light intensity transitioned a foliar endophyte into a pathogen, potentially due to increased melanization and faster growth rate associated with more light, exemplifying how the environment can impact microbial lifestyle [17]. As another example, Colletotrichum acutatum was isolated from strawberry, then inoculated onto various crop plants (strawberry, eggplant, tomato, and pepper) [11]. The fungus was able to survive and produce conidia on all plants, but only caused disease on strawberry, demonstrating the role of the host in determining the outcome of the plant-microbe interaction [11]. However, there has not been any unifying understanding of the relationship between host range (i.e., the hosts a microbe can infect) and microbial lifestyle (e.g., endophyte, pathogen). Additionally, the molecular mechanisms underlying diverse lifestyles ranging from pathogenic to endophytic remain poorly understood. Such questions are particularly important for agriculture, where distinguishing pathogens from endophytes remains an important consideration, as mis-targeting endophytes as pathogens would limit the beneficial effects of endophytes, and mis-targeting pathogens as endophytes would promote microbes that inhibit the growth of or even kill important crop species.
One particularly important plant-associated microbial genus is Colletotrichum, which comprises scientifically and agriculturally relevant fungi. Indeed, Colletotrichum species were ranked in the top ten most important fungal pathogens in molecular plant pathology, based on their uses in science as model organisms and their importance as pathogens in agriculture [18]. At the genus level, Colletotrichum fungi can infect a wide range of host plants, including a number of monocotyledonous and dicotyledonous plants [19]. Colletotrichum fungi have been isolated from various hosts such as Spinacia species (C. spinaciae), Lilium species (C. spaethianum, C. lilii), Phaseolus species (C. lindemuthianum, C. nymphaeae, C. plurivorum, C. sojae, C. spaethianum, C. phaseolorum), and Triticum species (C. karsti, C. graminicola, C. cereale) [19,20,21]. Colletotrichum fungi also have a diversity of lifestyles, including both endophytes, such as C. tofieldiae str. 61 [22] and str. 4 [12], and pathogens, such as C. higginsianum [23] and C. incanum [19], although as discussed later, lifestyle is dependent on the host colonized (Section 2; Figure 1). In this way, Colletotrichum makes an effective system for studying host-microbe interactions and microbial lifestyle because closely related endophyte-pathogen pairs may be analyzed in terms of the molecular differences between them. A single fungal species or strain may also be studied on different hosts to identify the factors that differentiate the lifestyle on one host from another. Therefore, this genus makes an excellent model for studying the determinants of lifestyle in plant-associated fungi.
In this review, we aim to disentangle the relationship between lifestyle and host range in endophytic and pathogenic fungi, with a particular focus on Colletotrichum species. We discuss how lifestyle is a consequence of the host colonized, with the same fungal species being endophytic on a largely different range of hosts than on which it is pathogenic. We then discuss progress made in elucidating the molecular determinants of lifestyle using comparative genomics. We next evaluate the benefits of applying endophytes to phytopathology research through the use of comparative omics. In particular, we reference the C. tofieldiae study system, which makes a fascinating system by which to study the determinants of lifestyle and to understand the relationship between lifestyle and host range due to the close phylogenetic relationship between pathogens and endophytes. We then note the benefits of applications of Colletotrichum research to agriculture, including the benefits of endophytes and the lessons we have learned from pathogens. Lastly, we propose an idea for the relationship between host range and lifestyle and discuss future directions for plant-fungus interactions research.
2. Lifestyle as a Result of Host Colonization
The lifestyle of plant-associated fungi can be considered in terms of the host colonized. Recent accumulating reports suggest that many fungi traditionally considered pathogens actually behave endophytically on alternative hosts [11,24,25,26,27]. Likewise, there are reports suggesting that some endophytes can behave pathogenically on alternative hosts, such as C. tofieldiae str. 4, which was originally isolated from diseased Ornithogalum umbellatum [28], but behaves endophytically on Arabidopsis thaliana [12], or foliar endophytic Colletotrichum strains, which display virulence on several plant species [29]. For the most part, fungal pathogens occupy a narrow host range, a phenomenon that has been labelled “host species specificity” [30]. From the host plant’s perspective, this is due to “non-host resistance”, a dualable resistance system that protects most plants from most pathogens [31]. While out of the scope of this review, it is nonetheless important to note that the host plant plays an active role in the plant-microbe interaction. Conversely, the host range of endophytes tends to be broader, frequently encompassing taxonomically distantly related species [32,33]. In this section, we discuss the host ranges of endophytic and pathogenic fungi.
2.1. Host Range of Pathogens
Generally, fungal plant pathogens can infect only a limited number of host species [30]. Some Colletotrichum pathogens also occupy a limited host range, whereas others actually occupy a broad host range [19,20,34,35,36] (Table 1). Indeed, some species exhibit little-to-no host specificity [e.g., 85;124], whereas others are highly host specific [e.g., [35]]. For example, C. lupini is restricted to Lupinus species pathogenically [37]; C. boninense sensu lato has broad host range as a species complex, but within this species complex, some species show restricted host ranges [38]; some isolates of C. capsici were shown to be restricted in disease-causing growth to certain legumes, although at the species-level, host range is known to be broader [39]; C. acutatum sensu lato as a species complex infects a number of host plants, but certain species within the species complex exhibit host specificity [40]; C. graminicola is restricted to Zea mays and Sorghum [41,42]; C. orbiculare sensu stricto is restricted to Cucurbitaceae and Nicotiana species [43,44]; C. higginsianum infects Brassicaceae plants pathogenically and has also been isolated from diseased Campanula and Rumex species [23,66,67]; and C. siamense pathogenically infects Capsicum species and the phylogenetically distant Pyrus pyrifolia [45]. At the same time, however, C. siamense occupies an entirely different host range as an endophyte, highlighting that one species can act endophytically on some hosts while behaving pathogenically on others [26;68]. Species-level discussion, however, is problematized when one considers the strain-level specificity of lifestyle on the same host. For example, some C. fructicola isolates are endophytic on Coffea species, whereas others are pathogenic [47]. Likewise, C. tofieldiae str. 3 is pathogenic on A. thaliana whereas C. tofieldiae str. 4 and str. 61 are endophytic [12,22]. Similarly, C. brasiliense has been isolated from Passiflora edulis as an endophyte and as a pathogen, depending on the isolate [38]. However, strain-level information is not available for all Colletotrichum species in the literature, as the taxonomy is frequently only presented down to the species or species complex level. Therefore, we continue our discussion with consideration of taxonomy down to the strain level where available, and to the species level otherwise.
2.2. Host Range of Endophytes
Endophytes, as opposed to pathogens, live within host tissues without causing disease symptoms, and may provide benefits to the host plant, observable on the macro scale by a plant growth promotion phenotype. As opposed to the narrow host range exhibited by many, but notably not all, pathogens, fungal endophytes frequently have a broader host range [32,48,49]. Because the number of studies empirically testing endophytism (i.e., isolation of a fungus from non-diseased tissue, re-infection on the host to validate lack of disease symptoms, and microscopic visualization to verify the fungus grows within host tissues) is extremely limited, especially for Colletotrichum fungi, here we consider fungi that have been isolated from surface-sterilized, non-diseased tissues as endophytes. As a rare example of a well-characterized endophyte [22], C. tofieldiae str. 61 is able to infect and promote growth of distantly related tomato and maize plants after being isolated from Arabidopsis thaliana [50,51]. However, the division between pathogens and endophytes is not evident based on species identity–as noted above (Section 2.1), some species have strain-level lifestyle differences, such as C. tofieldiae (Section 4.2). Additionally, many species are endophytic or pathogenic depending on the host colonized [11,24,25,26,27,29] (Table 1). As aforementioned, C. siamense is one example of such a pathogen, with no reported overlap between the hosts it colonizes as a pathogen compared to those it colonizes as an endophyte [32,33,34,35,36]. Another example is C. boninense sensu lato, which also has just one genus, Coffea, represented by both endophytic and pathogenic lifestyles [38]. Additionally, C. chrysophilum has been reported as a pathogen of banana, cucurbits, and cashew [52,53,54], but other isolations from symptomless plants imply that this fungus is also an endophyte of unwounded cashew, Theobroma, and Genipa [55,56]. A fourth example can be found in C. coccodes, which pathogenically infects potatoes, tomatoes, and chilis, but endophytically infects cabbage, white mustard, lettuce, and chrysanthemum [57,58,59]. Acknowledging these findings, it is apparent that lifestyle is dependent on the host colonized for many Colletotrichum fungi, with the endophytic lifestyle having a largely distinct host range compared to the pathogenic lifestyle.

Table 1.
Summarization of reported hosts of Colletotrichum fungi as pathogens and as endophytes. Underlined species are shared hosts of the fungus as a pathogen and as an endophyte.
Table 1.
Summarization of reported hosts of Colletotrichum fungi as pathogens and as endophytes. Underlined species are shared hosts of the fungus as a pathogen and as an endophyte.
| Colletotrichum species | Host range as pathogen | Host range as endophyte |
| C. lupini | Lupinus species [37] | Coffea arabica [60] |
| C. boninense s. l. | Protea species, Carica papaya, Capsicum species, Citrus species, Coffea species, Cucurbita species, Cymbidium species, Dracaena marginata, Diospyros australis, Solanum species, Passiflora edulis, Crinum asiaticum, Euonymus japonica, Persea americana, Phyllanthus acidus, Brassica oleracea [38] | Maytenus ilicifolia, Podocarpaceae, Myrtaceae, Coffea species, Mangifera indica, Pachira species Pleione bulbocodioides, Oncidium flexuosum, Quercus salicifolia, Theobroma cacao, Zamia obliqua, Parsonsia capsularis [38]; Eperua falcata, Goupia glabra, Manilkara bidentata, Mora excelsa, Catostemma fragrans, Carapa guianensis [61]; Podocarpaceae [62] |
| C. capsici | Piper betle, Vigna unguiculata, Phaseolus vulgaris, Cicer arietinum, Lupinus angustifolius, Pisum sativum, Vigna species [39] | Arachia hypogaea, Cajanus cajan, Phaseolus vulgaris, Phasolus lunatus [39] |
| C. acutatum s. l. | Malus domestica, Aspalathus linearis, Acer platanoides, Araucaria excelsa, Coffea arabica, Cyclamen species, Fragaria x ananassa, Lobelia species, Chrysanthemum coronarium, Actinidia species, Anemone coronaria, Cyclamen species, Hevea brasiliensis, Lupinus species, Nuphar luteum, Nymphaea alba, Oenothera species, Passiflora edulis, Parthenocissus species, Penstemon species, Persea americana, Prunus cerasus, Pyrus species, Solanum species, Ugni molinae, Vaccinium corymbosum, Citrus aurantium, Mahonia aquifolium [40] | Coffea robusta, Mangifera indica, Tsuga canadensis, Theobroma cacao, Dendrobium nobile [40] |
| C. graminicola | Zea mays [42]; Sorghum species [41] | Zea mays [63]; Lolium perenne [64] |
| C. orbiculare s. s. | Cucurbitaceae species [43]; Nicotiana species [65] | |
| C. higginsianum | Brassicaceae species [23]; Campanula species [66]; Rumex species [67] | Centella asiatica [69] |
| C. siamense | Capsicum species, Pyrus pyrifolia [45] | Pennisetum purpureum, Cymbopogon citratus [26]; Paullinia cupana [68] |
| C. gloeosporioides s. s. | Spondias pinnata, Persea americana, Aegle marmelos, Anacardium occidentale, Citrus species, Syzygium species, Durio zibethinus, Psidium guajava, Annona muricata, Mangifera indica, Garcinia mangosteen, Carica papaya, Passiflora edulis, Punica granatum, Nephelium lappaceum, Phyllanthus heyneanus, Achras sapota, Annona squamosa, Lycopersicon esculentum, Flacourtia inermis, Elaeocarpus glandulifer, Feronia limonia [70]; Olea europaea, Malus domestica [71] | Podocarpaceae [62]; Lolium perenne [64]; Centella asiatica [69]; Huperzia serrata [72]; Citrus species, Commelina benghalensis, Sida rhombifolia, Amaranthus deflexus [73]; Theobroma cacao, Anacardium excelsum, Genipa americana, Pentagonia macrophylla, Tetragastris panamensis, Trichilia tuberculata, Virola surinamensis, Cordia alliodora, Merremia umbellata, Zamia obliqua [74]; Uncaria rhynchophylla [75]; Camellia sinensis [76] |
| C. fructicola | Coffea species [47], Camellia sinensis, Citrus species, Malus species, Persea americana, Prunus persica, Pyrus species [77] | Coffea species [78]; Hevea species [79]; Dendrobium species [80] |
| C. tofieldiae | Arabidopsis thaliana [12], Ornithogalum umbellatum [28]; Capsella rubella [22] | Arabidopsis thaliana [12,22,51]; Zea mays, Solanum lycopersicum [50]; Cardamine hirsuta [22] |
| C. brasiliense | Passiflora edulis | Passiflora edulis |
| C. chrysophilum | Cucurbitaceae [54], Musa species [52]; Anacardium occidentale [53] | Theobroma species, Genipa species [56]; Anacardium occidentale [55] |
| C. coccodes | Capsicum species [59]; Solanum lycopersicum, Solanum tuberosum [57] | Brassica alba, Latuca sativa, Lepidum sativum, Brassica oleracea, Chrysanthemum species, Solanum nigrum [57] |
3. Comparative Genomic Approaches for Understanding the Host Range of Pathogenic Colletotrichum Fungi
Since the question of how host range is shaped in host-microbe interaction has attracted many researchers, numerous works attempting to answer this question have been conducted [81,82]. Especially in phytopathogenic fungi and oomycetes, genetic and genomic approaches have expanded our understanding of what kind of genes have evolved and are utilized to invade host plants [83,84,85,86]. As mentioned above (Section 2), Colletotrichum fungi have notable diversity in host range and lifestyle [87,88]. Comparative genomic studies have been conducted to uncover the origin and the causes of their diverse host range and lifestyle. In O’Connell et al. [89], the authors for the first time reported the genomes of two pathogenic Colletotrichum, namely C. higginsianum and C. graminicola, and compared their genomes. C. higginsianum has a larger and more diverse repertoire of candidate secreted effector genes (365 genes, 72% of the genes are species-specific) than C. graminicola (177 genes, 48% are species-specific). They discussed that the expanded effector repertoire in C. higginsianum might result from a broader host range of C. higginsianum (see also Section 2.1). Furthermore, genomic analyses showed that the genes encoding pectin-degrading enzymes among carbohydrate-active enzymes (CAZymes) are more than twice as abundant in C. higginsianum compared to C. graminicola and the majority of which are expressed in the necrotrophic infection stage. Transcriptome analyses of both fungi during infection revealed that C. graminicola upregulates cellulases and especially hemicellulases. This tendency might reflect the difference in cell wall composition between dicots, which have enriched pectin, and monocots, which contain more hemicellulose. Additionally, time-course transcriptomic analysis during C. higginsianum infection to Arabidopsis thaliana revealed that fungal gene expression is distinctive between biotrophic and necrotrophic infection stages. In this pathosystem, effector genes and secondary metabolite cluster genes were upregulated in the early infection (biotrophic) stage whereas many CAZymes were expressed in the late infection (necrotrophic) stage. Another comparative genomics study following O’Connell et al. indicated that secreted proteases, such as subtilisins, are expanded in C. orbiculare, C. fructicola (previously identified as C. gloeosporioides), and C. higginsianum, compared to other hemibiotrophic fungi such as Magnaporthe oryzae and Fusarium graminearum [89,90]. Especially in C. orbiculare and C. fructicola, metalloproteases are also highly expanded, suggesting their role in invasion into the host plant tissue [90]. Interestingly, there are slight differences in gene number encoding CAZymes between C. orbiculare and C. fructicola. These reports highlighted that Colletotrichum species show similarity in some aspects despite their relatively distant relationship within the genus; however, the expression landscape reported is dynamically different among species. Transcriptome plasticity might determine the host range and/or fungal lifestyle as a pathogen [89,90]. Supporting this idea, it has been reported in a pathosystem that transcriptional differences upon host defense may shape different host ranges between close relatives of Sclerotinia spp. [91].
Insights into the genomic evolution of Colletotrichum fungi during host adaptation have also accumulated further in recent years through genomics and/or transcriptomics approaches. For example, Gan et al. [92] sequenced C. incanum, which can infect both dicot (Raphanus sativus and A. thaliana) and monocot (lily) plants, and conducted genus-wide comparative analyses together with other five Colletotrichum species to find out hints at how this pathogen can infect both the monocot and dicot plant species. Interestingly, their analyses for identifying genes with positive selection in the C. incanum genome revealed that C. incanum genes encoding candidate secreted or nuclear-localized proteins, which are highly conserved among Colletotrichum species, are predicted to undergo diversification [92]. This analysis suggested that functional mutation, or polymorphism in these genes contributes to niche adaptation [92]. Conversely, in pathogenic Colletotrichum, excluding a few such as C. higginsianum, gene losses are predicted to occur more commonly than gene gains [92]. They also conducted comparative genomics among Colletotrichum gloeosporioides species complex and found that effector gene clusters were associated with telomeres and repeat-rich region, suggesting the host adaptation along with the evolution of accessory genomic region [93]. Baroncelli et al. [94] conducted a genus-wide comparison focusing on the difference between dicot-infecting and monocot-infecting Colletotrichum fungi. The study highlighted that monocot-infecting species evolved from dicot-infecting ancestral Colletotrichum through independent host jumps accompanied by a decrease in CAZyme repertoires. From comparative transcriptomics, they found that several core genes, including transcription factors, are potentially required commonly to adapt to plant niches whereas each gene expression profile showed high diversity. Genomic variation is even observed in intraspecies between C. higginsianum strains, particularly in gene poor regions where candidate effectors and secondary metabolism clusters are located. The variation is suggested to be associated with transposable elements [95,96]. In another research, comparisons among Colletotrichum graminicola isolates detected high polymorphism both in coding and non-coding regions of virulence-related genes [97]. Together, large-scale genomics provided significant insights into the evolution of host adaptation, leading to shape host range, and candidate genes for future investigation into the molecular mechanisms of host adaptation.
To date, how host range and specificity are formed remains a big question; however, Inoue et al. [98] identified effector genes potentially responsible for host specificity utilizing transcriptomics and generation of multiple candidate gene knockout mutants. C. orbiculare infects cucurbits and distantly related Nicotiana benthamiana. They obtained fungal gene expression profiles during cucumber infection and N. benthamiana infection, and found four cucurbit-specific effectors that were highly expressed in the early infection stage and required for virulence on cucumber and melon hosts. Notably, the expression of these effector genes was less enhanced in N. benthamiana and dispensable for infection. This study highlighted the possibility that Colletotrichum fungi use diverse sets of effector genes in response to different host plants, resulting in successful colonization. Overall, genomics and transcriptomics can be strong tools for investigating the molecular determinants of Colletotrichum host range, although which host-Colletotrichum infection system is suitable to deploy is an issue because the extensive diversity of gene repertoires and transcriptional landscape among the Colletotrichum-host interaction system provides a number of genes potentially involved in shaping host range, resulting in making it difficult to investigate.
4. Merits of Applying Endophytic Fungal Research to Phytopathology
While phytopathology has traditionally focused exclusively on pathogens, there are numerous benefits to using endophytes alongside pathogens in phytopathology research. For example, comparisons can be drawn between pathogens and endophytes to elucidate the molecular basis of lifestyle, particularly between closely related pathogens and endophytes. Subsequently, mutants can be generated, and their phenotypes screened. Such reverse genetics approaches are especially useful in Colletotrichum, where sexual reproduction is rarely observed, and there are strain-level differences in thallism (i.e., the ability to sexually reproduce within a single individual (homothallism) or whether multiple individuals are required (heterothallism)), so forward genetics approaches are difficult to accomplish [99]. Here, we discuss the merits of applying endophytes to phytopathology, with particular reference to C. tofieldiae as a potential system for studying fungal lifestyle.
4.1. Comparative Omics
One way to approach identifying molecular determinants of lifestyle is through comparative omics, as partially outlined in the preceding section with a focus on pathogenic species. Through this approach, endophytes and pathogens are directly compared, and differences between them are identified [e.g., 100] (see also Section 3). Typically, comparisons are made only between strains of similar lifestyles [e.g., 101-105, but see also 12,106], although sometimes of varying virulence levels [e.g., 107-108] or style of infection (e.g., biotrophy vs. necrotrophy) [109] (see also Section 3 for pathogen-pathogen comparisons). While this has nonetheless led to interesting findings, there is a largely unexplored plethora of information that can be uncovered through comparing fungi of different lifestyles. In particular, comparing closely related endophyte-pathogen pairs may lead to especially interesting findings about the molecular mechanisms that underlie lifestyle. For example, Hiruma et al. [12] compared the genomes and transcriptomes of two closely related C. tofieldiae strains with remarkably different lifestyles. Str. 3 is a pathogen, whereas str. 4 is an endophyte on A. thaliana. They identified a putative abscisic acid and botrydial biosynthesis cluster (ABA-BOT) uniquely activated in the pathogenic C. tofieldiae str. 3 that contributes to pathogenesis. When knock-out mutants of genes in ABA-BOT were generated, they grew endophytically and promoted plant growth under low phosphate conditions, similar to other endophytic C. tofieldiae strains, indicating a significant contribution of this gene to pathogenesis. As another example, Fu et al. [110] compared two strains of C. gloeosporioides, one pathogenic and one endophytic. They identified pathogen-specific gene clusters, which were enriched in polyketide synthase genes, potentially suggesting polyketide synthases as important regulators of lifestyle in C. gloeosporioides. As a third example, Hacquard et al. [111] compared the genomes and transcriptomes of endophytic C. tofieldiae and pathogenic C. incanum. Although the overall gene quantity was similar across C. tofieldiae and C. incanum, and the majority of genes are orthologous between the two species, C. tofieldiae-specific genes were enriched in secondary metabolism biosynthesis genes, and the number of species-specific effector genes was smaller than those of C. incanum. They then found that C. tofieldiae has more chitin-binding CAZymes than C. incanum, although during infection of A. thaliana, C. tofieldiae does not activate many CAZymes, effectors, or secondary metabolism biosynthesis genes, especially in the early stages of infection. Broadly, Hacquard et al. found genomic and transcriptomic signatures of lifestyle by comparing the genomes and transcriptomes of closely related Colletotrichum species. These studies demonstrate the potential utility of analyzing endophytes alongside pathogens in identifying determinants of lifestyle. However, there are few systems in which pathogens and endophytes are closely related enough to make meaningful comparisons. When endophytes and pathogens are too distantly related, there are numerous genes, transcripts, and proteins that differentiate them, making direct comparisons difficult. For this reason, we propose the C. tofieldiae study system as a potential model for endophyte-pathogen comparisons.
4.2. C. tofieldiae Study System
C. tofieldiae is a species of Colletotrichum belonging to the Spaethianum species complex, causing anthracnose in Agapanthus, Lupinus, Semele, and Tofieldia species [112,113]. It has also been identified as an endophyte on A. thaliana, promoting plant growth and transferring phosphate to its host under low phosphate conditions [22]. There are, however, strain-specific lifestyles in this species within the same host: C. tofieldiae str. 3 causes disease whereas strs. 4 and 61 promote plant growth in A. thaliana under low phosphate conditions [12]. Notably, strs. 3 and 4 are sister strains and share over 98% of their genomes [12]. Our recent results have revealed multiple interesting findings using the A. thaliana-C. tofieldiae study system. First, Hiruma et al. [12] found that ABA-BOT makes a significant contribution to C. tofieldiae str. 3 pathogenicity. It is uniquely activated in str. 3 and not strs. 4 or 61, although it is present in the genomes of all three strains. However, the regulatory mechanisms of ABA-BOT remained elusive until Ujimatsu et al. [114]. Ujimatsu et al. uncovered the function of BOT6, a transcription factor, in regulating ABA-BOT and other virulence-related genes. Notably, overexpression of BOT6 was sufficient to transition C. tofieldiae str. 4 from an endophyte into a pathogen. Although these findings have elucidated important mechanisms of virulence in C. tofieldiae, there are still numerous unanswered questions, such as how BOT6 is regulated, what other genetic determinants of lifestyle exist in C. tofieldiae and other Colletotrichum fungi, and the specific mechanisms by which ABA-BOT promotes fungal virulence. Because of the high similarity between C. tofieldiae str. 3 and str. 4 on a genomic level, one could readily compare the genomes of these fungi to identify putative determinants of lifestyle. These could then be compared to the genomes of other fungi, and orthologous genes could be identified, which may also be determinants of lifestyle in those fungi (e.g., the presence of ABA-BOT in other fungi apart from C. tofieldiae). Unlike many other endophyte-pathogen pairs, C. tofieldiae strains are extremely closely related to one another, so the effect of redundancy (i.e., multiple genes sharing similar functions) is likely limited, as only a small pool of genes differentiates C. tofieldiae str. 3 from str. 4. Based on our previous reports uncovering determinants of lifestyle using C. tofieldiae, we therefore suggest the C. tofieldiae study system as a model for understanding endophytism and pathogenicity.
5. Applications of Colletotrichum Research
Colletotrichum fungi have been used as models for studying plant pathogens [62,63,64]. However, there are also an increasing number of studies using Colletotrichum endophytes for their plant benefits [e.g., 12,22,50,115-116]. Here, we synthesize the literature on both endophytic and pathogenic Colletotrichum species to demonstrate that both endophytic and pathogenic fungal research can advance agricultural sciences.
5.1. Endophytic Fungi
Endophytic fungal species can be applied to agriculture for their benefits to their host plants. Indeed, Colletotrichum endophytes have been demonstrated to promote plant growth and transfer nutrients to their host [12,50]. These fungi could be applied to vulnerable crop species to promote yield while reducing the use of traditional chemical fertilizers, which can cause environmental damage [117,118]. Biofertilizers (i.e., fertilizers composed of beneficial microorganisms) can provide comparable benefits to plants as chemical fertilizers and still be profitable to growers [119,120]. Beneficial Colletotrichum endophytes could be incorporated into biofertilizers to help promote plant growth and improve crop yield. At the same time, Colletotrichum endophytes may also provide protection against pathogens [116]. Therefore, applying biofertilizers composed of Colletotrichum endophytes may provide multiple benefits to the host plants. However, strain selection must be carefully considered in order to avoid mis-applying pathogenic strains on crops, especially considering that many endophytes can behave pathogenically on certain host species. Such conditional pathogenicity may, however, be beneficial to field conditions. For example, if a fungus is pathogenic to weed species (see Section 5.2 for further discussion) or otherwise reduces the detrimental effects of weeds, but endophytic on crop species, it could provide dual benefits to the host plant by mitigating the negative effects of weeds while promoting beneficial effects for the crop [e.g., 121].
Additionally, Colletotrichum endophytes have been studied for their secondary metabolites. Secondary metabolites can be a source of beneficial substances to humans, such as compounds that inhibit cancer [122,123]. While sometimes phytotoxic, secondary metabolites can also include plant growth promoting compounds [124,125]. The role of secondary metabolites in the plant-endophyte relationship is somewhat less studied, particularly in Colletotrichum endophytes, but it stands to reason that Colletotrichum endophytes may be a source of yet undiscovered secondary metabolites that promote plant growth. In support of this hypothesis, 59 putative secondary metabolite biosynthesis clusters were found in plant growth promoting strains C. tofieldiae str. 4 and str. 61, including multiple that were upregulated in planta [126].
5.2. Lessons from Pathogenic Fungi
Much of Colletotrichum research has historically focused on phytopathogenic species, leading to many discoveries about virulence factors produced by Colletotrichum pathogens. These virulence factors can be targeted by plant breeders or bioengineers through the addition of their cognate resistance genes [127,128], although such approaches must be undertaken with caution because increasing the resistance gene load can impose fitness costs that reduce plant growth [129]. Alternatively, virulence factors identified in phytopathogenic fungi can be the target of fungicides, especially those virulence factors that are conserved across many fungi [e.g., 130]. While effective when applied appropriately, it is highly possible for resistant strains to emerge over time as a result of the selective pressure imposed by fungicides [131,132,133,134]. Therefore, fungicides may not provide an avenue for durable disease resistance, but rather a short-term management strategy, or ideally, one component of an integrated pest management strategy [135,136]. As another, potentially more sustainable option, one could develop fungicides that target core processes in fungi. One notable example is fungicides that target melanin biosynthesis, which is ultimately required by fungi to build up turgor pressure in order to penetrate the plant cell wall [137,138,139]. By developing fungicides that target core processes for expressing pathogenesis, it is possible to minimize the risk of fungi evolving fungicide resistance. Therefore, Colletotrichum fungi could be studied for their core processes, and fungicides could be developed based on these processes [140]. In these ways, the identification of virulence factors and core processes provides multiple routes for agronomists to deploy resistant crops or otherwise target pathogens in the field.
Additionally, plant pathogenic fungi have been studied for their herbicidal compounds [141]. Because plant pathogens attack their hosts using herbicidal compounds, it is possible to apply pathogens to the field of non-host species to target weeds. Colletotrichum fungi have indeed been studied as mycoherbicides, such as C. coccodes being applied to soybean and velvetweed and causing a reduction in growth and competitive ability of velvetweed compared to soybean [142]. C. truncatum has also been used as a mycoherbicide against hemp sesbania in field conditions, and caused high mortality of this important weed plant [143]. Because of the restricted host range of pathogens, it is feasible to apply mycoherbicides to the field to limit the growth of weeds without affecting crop species [144]. Colletotrichum fungi, as discussed above (Section 2.1), have limited host range too, making them potential candidates for mycoherbicide production.
6. Relationship Between Host Range and Lifestyle
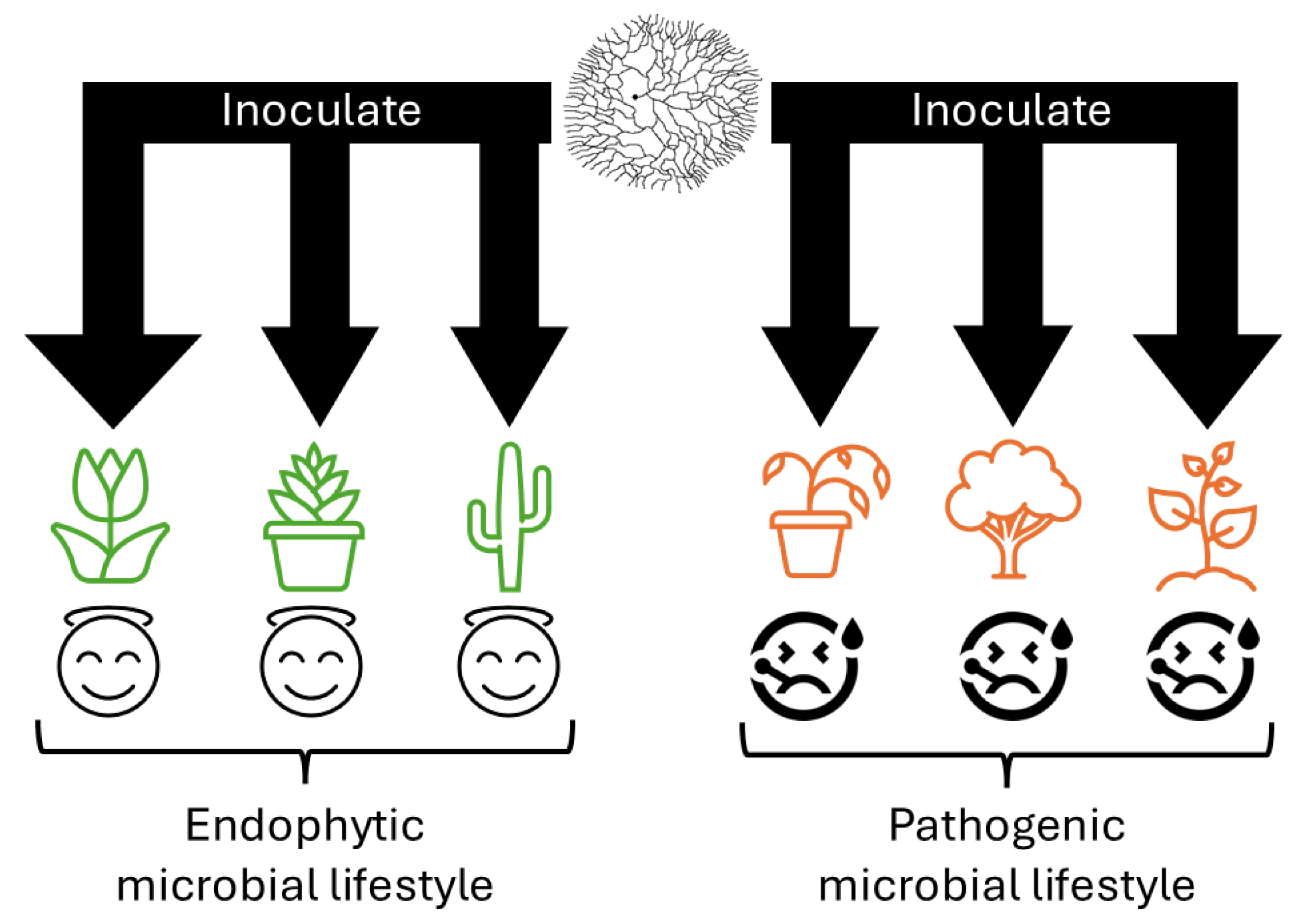
Within a single fungus with host-dependent lifestyles, the fungus will likely be endophytic on a different range of hosts than on which it is pathogenic. Thus, if it is inoculated onto a wide range of hosts, it will likely be an endophyte on different hosts than on which it is a pathogen. Note that arrows do not correlate to phylogenetic relationships among host plants.
Given the above findings, we propose the following model for the relationship between host range and lifestyle: 1) endophytism and pathogenicity are context-dependent, and may change depending on the host and environmental conditions; 2) pathogenicity and endophytism tend to occupy different host ranges compared to each other; 3) within a single fungus with context-dependent lifestyles, the pathogenic lifestyle is likely to be constrained to different hosts than the endophytic lifestyle (Figure 1). Notably, neither the host range of pathogens nor of endophytes apparently correlates with host plant phylogeny. For example, some pathogens and endophytes infect both monocots and dicots, which are phylogenetically distant from each other (e.g., C. incanum, C. fructicola). Benefits of applying fungi will therefore be context-dependent; fungi may infect some hosts endophytically and promote plant growth while growing pathogenically and inhibiting plant growth on other hosts. The use of fungi in bioproducts thus necessitates an understanding of the host range of the specific fungus used, and fungi traditionally thought to be pathogens may actually be endophytic, depending on the host plant. In this way, “pathogens” may actually make useful additions to biofertilizers, and “endophytes” may harm their plant hosts, depending on the host on which they are applied. Therefore, we suggest that host plant is a crucial consideration when developing and deploying antifungal pest management measures or pro-fungal bioproducts.
7. Concluding Remarks and Future Directions
In this review, we have demonstrated that there is a previously undiscussed relationship between host range and fungal lifestyle. Generally speaking, a pathogenic lifestyle occupies a different host range than an endophytic lifestyle, even within the same fungal species. This is to say, one fungus may be capable of colonizing a broad range of hosts, but only on a select number of those hosts will it cause disease. On the other hosts, the fungus may infect without causing any disease symptoms and may actually promote plant growth. Such lifestyle differences are driven by diverse mechanisms, but frequently include secondary metabolites, CAZymes, and effectors. Such determinants of lifestyle may make useful targets for limiting the spread of pathogens in the field, although care must be taken to avoid mis-targeting endophytes. Future studies should focus on understanding the molecular mechanisms underlying lifestyle, especially in useful study systems like C. tofieldiae. Furthermore, quantifying the host ranges of endophytic and pathogenic fungi to understand whether endophytism or pathogenicity is broader in host range could yield additional insights into the relationship between host range and fungal lifestyle. Additionally, applied work on developing bioproducts and appropriate fungicides based on the known determinants of virulence and known beneficial endophytes would be helpful in translating the current basic scientific research into agrosystems. Lastly, understanding the ways in which host plants interpret the plant-fungus interaction across fungi with different lifestyles would be helpful to understanding the ways in which host plants discriminate endophytes from pathogens. This would be particularly interesting to explore in closely related endophyte-pathogen pairs or within a single fungus that behaves endophytically or pathogenically depending on the environment, such as is the case with C. tofieldiae str. 3. Through such an improved understanding of plant-fungus interactions, we can improve crop yield and tackle food security challenges worldwide.
Acknowledgements
This work was supported in part by the JSPS KAKENHI Grant (JP23H02210,JP24K21864), the JST grant (JPMJAN23D4, JPMJFR200A), JST SPRING (JPMJSP2108 (JN, RU)). This research was funded in part by a Ben Barres Spotlight Award from eLife.
References
- Turner, T.R.; James, E.K.; Poole, P.S. The plant microbiome. Genome Biol. 2013, 14. [Google Scholar] [CrossRef]
- Berg, M.; Koskella, B. Nutrient- and dose-dependent microbiome-mediated protection against a plant pathogen. Curr. Biol. 2018, 28, 2487–2492. [Google Scholar] [CrossRef]
- Lazcano, C.; Boyd, E.; Holmes, G.; Hewavitharana, S.; Pasulka, A.; Ivors, K. The rhizosphere microbiome plays a role in the resistance to soil-borne pathogens and nutrient uptake of strawberry cultivars under field conditions. Sci. Rep. 2021, 11, 3188. [Google Scholar] [CrossRef] [PubMed]
- Sanders, F.E.; Tinker, P.B. Mechanisms of absorption of phosphate from soil by Endogone mycorrhizas. Nature 1971, 233, 278–279. [Google Scholar] [CrossRef]
- Van der Ent, S.; Van Hulten, M.; Pozo, M.J.; Czechowski, T.; Udvardi, M.K.; Pieterse, C.M.J.; Ton, J. Priming of plant innate immunity by rhizobacteria and β-aminobutyric acid: differences and similarities in regulation. New Phytol. 2009, 183, 419–431. [Google Scholar] [CrossRef] [PubMed]
- Redman, R.S.; Sheehan, K.B.; Stout, R.G.; Rodriguez, R.J.; Henson, J.M. Thermotolerance generated by plant/fungal symbiosis. Science 2002, 298, 1581. [Google Scholar] [CrossRef] [PubMed]
- Ray, R.V. Effects of pathogens and disease on plant physiology. In Agrios’ Plant Pathology, 6th ed.; Oliver, R.P., Hückelhoven, R., Del Ponte, E.M., Di Pietro, A., Eds.; Academic Press: London, United Kingdom, 2024; pp. 63–92. [Google Scholar]
- Teng, P.S.; Shane, W.W.; MacKenzie, D.R. Crop losses due to plant pathogens. Crit. Rev. Plant Sci. 1984, 2, 21–47. [Google Scholar] [CrossRef]
- Ren, P.; Sun, A.; Jiao, X.; Chen, Q.-L.; Li, F.; He, J.-Z.; Hu, H.-W. National-scale investigation reveals the dominant role of phyllosphere fungal pathogens in sorghum yield loss. Environ. Int. 2024, 185. [Google Scholar] [CrossRef]
- Dickson, S.; Smith, S.E.; Smith, F.A. Characterization of two arbuscular mycorrhizal fungi in symbiosis with Allium porrum: colonization, plant growth, and phosphate uptake. New Phytol. 1999, 144, 173–181. [Google Scholar] [CrossRef]
- Freeman, S.; Horowitz, S.; Sharon, A. Pathogenic and nonpathogenic lifestyles in Colletotrichum acutatum from strawberry and other plants. Phytopathology 2001, 91, 986–992. [Google Scholar] [CrossRef] [PubMed]
- Hiruma, K.; Aoki, S.; Takino, J.; Higa, T.; Devi Utami, Y.; Shiina, A.; Okamoto, M.; Nakamura, M.; Kawamura, N.; Ohmori, Y.; Sugita, R.; Tanoi, K.; Sato, T.; Oikawa, H.; Minami, A.; Iwasaki, W.; Saijo, Y. A fungal sesquiterpene biosynthesis gene cluster critical for mutualist-pathogen transition in Colletotrichum tofieldiae. Nature Commun. 2023, 14, 5288. [Google Scholar] [CrossRef]
- Smith, K.P.; Goodman, R.M. Host variation for interactions with beneficial plant-associated microbes. Ann. Rev. Phytopathol. 1999, 37, 473–491. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.T.; Zhang, L.; He, S.Y. Plant-microbe interactions facing environmental challenge. Cell Host Microbe 2019, 26, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Gao, X.; Feng, B.; Sheen, J.; Shan, L.; He, P. Plant immune response to pathogens differs with changing temperatures. Nat. Commun. 2013, 4, 2530. [Google Scholar] [CrossRef]
- Huot, B.; Castroverde, C.D.M.; Velásquez, A.C.; Hubbard, E.; Pulman, J.A.; Yao, J.; Childs, K.L.; Tsuda, K.; Montgomery, B.L.; He, S.Y. Dual impact of elevated temperature on plant defence and bacterial virulence in Arabidopsis. Nat. Commun. 2017, 8, 1808. [Google Scholar] [CrossRef]
- Álvarez-Loayza, P.; White, J.F., Jr.; Torres, M.S.; Balslev, H.; Kristiansen, T.; Svenning, J.-C.; Gil, N. Light converts endosymbiotic fungus to pathogen, influencing seedling survival and niche-specific filling of a common tropical tree, Iriartea deltoidea. PLoS ONE 2011, 6, e16386. [Google Scholar] [CrossRef] [PubMed]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; Foster, G.D. The top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed]
- Talhinhas, P.; Baroncelli, R. Colletotrichum species and complexes: geographic distribution, host range, and conservation status. Fungal Divers. 2021, 110, 109–198. [Google Scholar] [CrossRef]
- Damm, U.; Woudenberg, J.H.C.; Cannon, P.F.; Crous, P.W. Colletotrichum species with curved conidia from herbaceous hosts. Fung. Divers. 2009, 39, 45–87. [Google Scholar]
- Hyde, K.D.; Cai, L.; Cannon, P.F.; Crouch, J.A.; Crous, P.W.; Damm, U.; Goodwin, P.H.; Chen, H.; Johnston, P.R.; Jones, E.B.G.; Liu, Z.Y.; McKenzie, E.H.C.; Moriwaki, J.; Noireung, P.; Pennycook, S.R.; Pfenning, L.H.; Prihastuti, H.; Sato, T.; Shivas, R.G.; Tan, Y.P.; Taylor, P.W.J.; Weir, B.S.; Yang, Y.L.; Zhang, J.Z. Colletotrichum - names in current use. Fungal Divers. 2009, 39, 147–182. [Google Scholar]
- Hiruma, K.; Gerlach, N.; Sacristán, S.; Nakano, R.T.; Hacquard, S.; Kracher, B.; Neumann, U.; Ramírez, D.; Bucher, M.; O’Connell, R.J.; Schulze-Lefert, P. Root endophyte Colletotrichum tofieldiae confers plant fitness benefits that are phosphate status dependent. Cell, 2016, 165, 464–474. [Google Scholar] [CrossRef]
- Damm, U.; O’Connell, R.J.; Groenewald, J.Z.; Crous, P.W. The Colletotrichum destructivum species complex - hemibiotrophic pathogens of forage and field crops. Stud. Mycol. 2014, 79, 49–84. [Google Scholar] [CrossRef] [PubMed]
- Al-Ani, L.K.T.; Furtado, E.L. The effect of incompatible plant pathogens on the host plant. In Molecular Aspects of Plant Beneficial Microbes in Agriculture; Sharma, V., Salwan, R., Al-Ani, L.K.T., Eds.; Academic Press: London, United Kingdom, 2020; pp. 47–57. [Google Scholar]
- Coutinho, T.A.; Venter, S.N. Pantoea ananatis: an unconventional plant pathogen. Mol. Plant Pathol. 2009, 10, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Manamgoda, D.S.; Udayanga, D.; Cai, L.; Chukeatirote, E.; Hyde, K.D. Endophytic Colletotrichum from tropical grasses with a new species C. endophytica. Fungal Divers. 2013, 61, 107–115. [Google Scholar] [CrossRef]
- Lisboa, D.O.; Silva, M.A.; Pinho, D.B.; Pereira, O.L.; Furtado, G.Q. Diversity of pathogenic and endophytic Colletotrichum isolates from Licania tomentosa in Brazil. Forest Pathol. 2018, 48, e12448. [Google Scholar] [CrossRef]
- Sato, T.; Moriwaki, J.; Kaneko, S. Anthracnose fungi with curved conidia, Colletotrichum spp. Belonging to ribosomal groups 9-13, and their host ranges in Japan. Jarq Jpn Agr. Res. Q. 2015, 49, 351–362. [Google Scholar] [CrossRef]
- Yang, A.-L.; Chen, Y.-S.; Mei, L.; Guo, J.; Zhang, H.-B. Disease risk of the foliar endophyte Colletotrichum from invasive Ageratina adenophora to native plants and crops. Fungal Ecol. 2024, 72, 101386. [Google Scholar] [CrossRef]
- Li, J.; Cornelissen, B.; Rep, M. Host-specificity factors in plant pathogenic fungi. Fungal Genet. Biol. 2020, 144, 103447. [Google Scholar] [CrossRef]
- Nürnberger, T.; Lipka, V. Non-host resistance in plants: new insights into an old phenomenon. Mol. Plant Pathol. 2005, 6, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Suryanarayanan, T.S.; Devarajan, P.T.; Girivasan, K.P.; Govindarajulu, M.B.; Kumaresan, V.; Murali, T.S.; Rajamani, T.; Thirunavukkarasu, N.; Venkatesan, G. The host range of multi-host endophytic fungi. Curr. Sci. 2018, 115, 1963–1969. [Google Scholar] [CrossRef]
- U’Ren, J.M.; Lutzoni, F.; Miadlikowska, J.; Laetsch, A.D.; Arnold, A.E. Host and geographic structure of endophytic and endolichenic fungi at a continental scale. Am. J. Bot. 2012, 99, 898–914. [Google Scholar] [CrossRef]
- Jeffries, P.; Dodd, J.C.; Jeger, M.J.; Plumbley, R.A. The biology and control of Colletotrichum species on tropical fruit crops. Plant Pathol. 1990, 39, 343–366. [Google Scholar] [CrossRef]
- Crouch, J.A.; Clarke, B.B.; White Jr., J. F.; Hillman, B.I. Systematic analysis of the falcate-spored graminicolous Colletotrichum and a description of six new species from warm-season grasses. Mycologia 2009, 101, 717–732. [Google Scholar] [CrossRef] [PubMed]
- Johnston, P.R.; Jones, D. Relationships among Colletotrichum isolates from fruit-rots assessed using rDNA sequences. Mycologia 1997, 89, 420–430. [Google Scholar] [CrossRef]
- Talhinhas, P.; Baroncelli, R.; Le Floch, G. Anthracnose of Lupins caused by Colleotrichum lupini: a recent disease and successful worldwide pathogen. J. Plant Pathol. 2016, 98, 5–14. [Google Scholar]
- Damm, U.; Cannon, P.F.; Woudenberg, J.H.C.; Johnston, P.R.; Weir, B.S.; Tan, Y.P.; Shivas, R.G.; Crous, P.W. The Colletotrichum boninense species complex. Stud. Mycol. 2012, 73, 1–36. [Google Scholar] [CrossRef] [PubMed]
- Pring, R.J.; Nash, C.; Zakaria, M.; Bailey, J.A. Infection process and host range of Colletotrichum capsici. Physiol. Mol. Plant Pathol. 1995, 46, 137–152. [Google Scholar] [CrossRef]
- Damm, U.; Cannon, P.F.; Woudenberg, J.H.C.; Crous, P.W. The Colletotrichum acutatum species complex. Stud. Mycol. 2012, 73, 37–113. [Google Scholar] [CrossRef] [PubMed]
- Crouch, J.A.; Clarke, B.B.; White Jr., J. F.; Hillman, B.I. Systematic analysis of the falcate-spored graminicolous Colletotrichum and a description of six new species from warm-season grasses. Mycologia 2009, 101, 717–732. [Google Scholar] [CrossRef]
- Wheeler, H.; Politis, D.J.; Poneleit, C.G. Pathogenicity, host range, and distribution of Colletotrichum graminicola on corn. Phytopathol. 1973, 64, 293–296. [Google Scholar]
- Damm, U.; Cannon, P.F.; Liu, F.; Barreto, R.W.; Guatimosim, E.; Crous, P.W. The Colletotrichum orbiculare species complex: important pathogens of field crops and weeds. Fungal Divers. 2013, 61, 29–59. [Google Scholar] [CrossRef]
- Shen, S.; Goodwin, P.H.; Hsiang, T. Infection of Nicotiana species by the anthracnose fungus, Colletotrichum orbiculare. Eur. J. Plant Pathol. 2001, 107, 767–773. [Google Scholar] [CrossRef]
- Liu, F.; Tang, G.; Zheng, X.; Li, Y.; Sun, X.; Qi, X.; Zhou, Y.; Xu, J.; Chen, H.; Chang, X.; Zhang, S.; Gong, G. Molecular and phenotypic characterization of Colletotrichum species associated with anthracnose disease in peppers from Sichuan Province, China. Sci. Rep. 2016, 6, 32761. [Google Scholar] [CrossRef] [PubMed]
- Casas, L.L.; de Almeida, L.N.; Pereira, J.O.; Neto, P.Q.C.; de Azvedo, J.L. Colletotrichum siamense, a mycovirus-carrying endophyte, as a biological control strategy for Anthracnose in Guarana plants. Braz. Arch. Biol. Technol. 2021, 64, e21200534. [Google Scholar] [CrossRef]
- Prihastuti, H.; Cai, L.; Chen, H.; McKenzie, E.H.C.; Hyde, K.D. Characterization of Colletotrichum species associated with coffee berries in northern Thailand. Fungal Divers. 2009, 39, 89–109. [Google Scholar]
- Arnold, A.E.; Lutzoni, F. Diversity and host range of foliar fungal endophytes: are tropical leaves biodiversity hotspots? Ecology 2007, 88, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, R.J.; White Jr., J. F.; Arnold, A.E.; Redman, R.S. Fungal endophytes: diversity and functional roles. New Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef] [PubMed]
- Díaz-González, S.; Marín, P.; Sánchez, R.; Arribas, C.; Kruse, J.; González-Melendi, P.; Brunner, F.; Sacristán, S. Mutualistic Fungal Endophyte Colletotrichum tofieldiae Ct0861 colonizes and increases growth and yield of maize and tomato plants. Agronomy 2020, 10, 1493. [Google Scholar] [CrossRef]
- García, E.; Alonso, Á.; Platas, G.; Sacristán, S. The endophtyic mycobiota of Arabidopsis thaliana. Fungal Divers. 2013, 60, 71–89. [Google Scholar] [CrossRef]
- Vieira, W.A.S.; Lima, W.G.; Nascimento, E.S.; Michereff, S.J.; Câmara, M.P.S. The impact of phenotypic and molecular data on the inference of Colletotrichum diversity associated with Musa. Mycologia 2017, 109, 912–934. [Google Scholar] [CrossRef]
- Veloso, J.S. , Câmara, M.P.S.; Lima, W.G.; Michereff, S.J.; Doyl, V.P. Why species delimitation matters for fungal ecology: Colletotrichum diversity on wild and cultivated cashew in Brazil. Fungal Biol. 2018, 122, 677–691. [Google Scholar] [CrossRef]
- Vieira, W.A.S.; da Costa, C.A.; Veloso, J.S.; Lima, W.G.; Correia, K.C.; Michereff, S.J.; Pinho, D.B.; Câmara, M.P.S.; Reis, A. Diversity of Colletotrichum species causing anthracnose in Chayote in Brazil, with a description of two new species in the C. magnum complex. J. Fungi 2024, 10, 847. [Google Scholar] [CrossRef]
- Ramos, D.G.B.; Amaral, A.G.G.; Duarte, I.G.; da Silva, A.C.; Vieira, W.A.S.; Castlebury, L.A.; Câmara, M.P.S. Endophytic species of Colletotrichum associated with cashew tree in northeastern Brazil. Fungal Biol. 2024, 128, 1780–1789. [Google Scholar] [CrossRef]
- Rojas, E.I.; Rehner, S.A.; Samuels, G.J.; Van Bael, S.A.; Herre, E.A.; Cannon, P.F.; Chen, R.; Pang, J.; Wang, R.-W.; Zhang, Y.; Peng, Y.-Q.; Sha, T. Colletotrichum gloeosorioides s.l. associated with Theobroma cacao and other plants in Panamá: multilocus phylogenies distinguish host-associated pathogens from asymptomatic endophytes. Mycologia 2010, 102, 1318–1338. [Google Scholar] [CrossRef]
- Chesters, C.G.C.; Hornby, D. Studies on Colletotrichum coccodes II. Alternative host tests and tomato fruit inoculations using a typical tomato root isolate. Trans Brit. Mycol. Soc. 1965, 48, 583–594. [Google Scholar] [CrossRef]
- Chang, J.; Boer, R.; Crous, P.W.; Taylor, P.W. Biology of foliar and root infection of potato plants by Colletotrichum coccodes in Australia. Phytopathol. 2014, 104, 24. [Google Scholar]
- De Silva, D.D.; Crous, P.W.; Ades, P.K.; Hyde, K.D.; Taylor, P.W.J. Life styles of Colletotrichum species and implications for plant biosecurity. Fungal Biol. Rev. 2017, 31, 155–168. [Google Scholar] [CrossRef]
- Poma-Angamarca, R.A.; Rojas, J.R.; Sánchez-Rodríguez, A.; Ruiz-González, M.X. Diversity of fungal leaf endophytes from two Coffea arabica varieties and antagonism towards Coffee Leaf Rust. Plants 2024, 13, 814. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Cannon, P.F.; Reid, A.; Simmons, C.M. Diversity and molecular relationships of endophytic Colletotrichum isolates from the Iwokrama Forest Reserve, Guyana. Mycol. Res. 2004, 108, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Joshee, S.; Paulus, B.C.; Park, D.; Johnston, P.R. Diversity and distribution of fungal foliar endophytes in New Zealand Podocarpaceae. Mycol. Res. 2009, 113, 1003–1015. [Google Scholar] [CrossRef] [PubMed]
- Sukno, S.A.; García, V.M.; Shaw, B.D.; Thon, M.R. Root infection and systemic colonization of maize by Colletotrichum graminicola. Appl. Environ. Microbiol. 2008, 74. [Google Scholar] [CrossRef] [PubMed]
- Neill, J.C. The endophyte of Rye-Grass (Lolium perenne). N. Z. J. Sci. Tech. Section A 1940, 21, 280–291. [Google Scholar]
- Shen, S.; Goodwin, P.H.; Hsiang, T. Infection of Nicotiana species by the anthracnose fungus, Colletotrichum orbiculare. Eur. J. Plant Pathol. 2001, 107, 767–773. [Google Scholar] [CrossRef]
- Khodaei, S.; Arzanlou, M.; Torbati, M.; Eghbali, S. Novel hosts in the genus Colletotrichum and first report of C. higginsianum from Iran. Nova Hedwigia 2019, 108, 449–463. [Google Scholar] [CrossRef]
- Zhang, Y.W.; Xue, L.H.; Li, C.J. First report of Anthracnose caused by Colletotrichum higginsianum on Rumex acetosa in China. Plant Dis. 2018, 102, 1174. [Google Scholar] [CrossRef]
- Casas, L.L.; de Almeida, L.N.; Pereira, J.O.; Neto, P.Q.C.; de Azvedo, J.L. Colletotrichum siamense, a mycovirus-carrying endophyte, as a biological control strategy for Anthracnose in Guarana plants. Braz. Arch. Biol. Technol. 2021, 64, e21200534. [Google Scholar] [CrossRef]
- Rakotoniriana, E.F.; Munaut, F.; Decock, C.; Randriamampionona, D.; Andriambololoniaina, M.; Rakotomalala, T.; Rakotonirina, E.J.; Rabeemanantsoa, C.; Cheuk, K.; Ratsimamanga, S.U.; et al. Endophytic fungi from leaves of Centella asiatica: occurrence and potential interactions within leaves. Antonie van Leeuwenhoek 2007, 93, 27–36. [Google Scholar] [CrossRef]
- Alahakoon, P.W.; Brown, A.E. Host range of Colletotrichum gloeosporioides on tropical fruit crops in Sri Lanka. Int. J. Pest Manag. 2008, 40, 23–26. [Google Scholar] [CrossRef]
- Latinovic, J.; Vucinic, Z. . Cultural characteristics, pathogenicity, and host range of Colletotrichum gloeosporioides isolated from olive plants in Montenegro. Acta Hortic. 2002, 586, 753–755. [Google Scholar] [CrossRef]
- Shu, S.; Zhao, X.; Wang, W.; Zhang, G.; Cosoveanu, A.; Ahn, Y.; Wang, M. Identification of a novel endophytic fungus from Huperzia serrata which produces huperzine A. World J. Microbiol. Biotechnol. 2014, 30, 3101–3109. [Google Scholar] [CrossRef]
- Waculicz-Andrade, C.E.; Savi, D.C.; Bini, A.P.; Adamoski, D.; Goulin, E.H.; Silva Jr., G. J.; Massola Jr., N.S.; Terasawa, L.G.; Kava, V.; Glienke C. Colletotrichum gloeosporioides sensu stricto: an endophytic species or citrus pathogen in Brazil? Australas. Plant Pathol. 2017, 46, 191–203. [Google Scholar]
- Rojas, E.I.; Rehner, S.A.; Samuels, G.J.; Van Bael, S.A.; Herre, E.A.; Cannon, P.; Chen, R.; Pang, J.; Wang, R.; Zhang, P.; Peng, Y.-Q.; Sha, T. Colletotrichum gloeosporioides s.l. associated with Theobroma cacao and other plants in Panamá: multilocus phylogenies distinguish host-associated pathogens from asymptomatic endophytes. Mycologia 2017, 102, 1318–1338. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Yang, Z.-D.; Chen, X.-W.; Zhou, S.-Y.; Yu, H.-T.; Sun, J.-Y.; Yao, X.-J.; Wang, Y.-G.; Xue, H.-Y. Colletotrilactam A-D, novel lactams from Colletotrichum gloeosporioides GT-7, a fungal endophyte of Uncaria rhynchophylla. Fitoterapia 2016, 113, 158–163. [Google Scholar] [CrossRef]
- Rabha, A.J.; Naglot, A.; Sharma, G.D.; Gogoi, H.K.; Gupta, V.K.; Shreemali, D.D.; Veer, V. Morphological and molecular diversity of endophytic Colletotrichum gloeosporioides from tea plant, Camellia sinensis (L.) O. Kuntze of Assam, India. J. Genet. Eng. Biotech. 2016, 14, 181–187. [Google Scholar] [CrossRef]
- EFSA Panel on Plant Health (PLH); Bragard, C.; Dehnn-Schmutz, K.; Di Serio, F.; Gonthier, P.; Jacques, M.-A.; Miret, J.A.J.; Justesen, A.F.; MacLeod, A.; Magnusson, C.S.; et al. Pest categorisation of Colletotrichum fructicola. EFSA J. 2021, 19, e06803. [Google Scholar]
- Prihastuti, H.; Cai, L.; Chen, H.; McKenzie, E.H.C.; Hyde, K.D. Characterization of Colletotrichum species associated with coffee berries in northern Thailand. Fungal Divers. 2009, 39, 89–109. [Google Scholar]
- Amaral, A.O.; Ferreira, A.F.T.A.F.; Bentes, J.L.S. Fungal endophytic community associated with Hevea spp.: diversity, enzymatic activity, and biocontrol potential. Bra. J. Microbiol. 2022, 53, 857–872. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Nontachaiyapoom, S.; Jayawardena, R.S.; Yde, K.D.; Gentekaki, E.; Zhou, S.; Qian, Y.; Wen, T.; Kang, J. Endophytic Colletotrichum species from Dendrobium spp. in China and Northern Thailand. MycoKeys 2018, 43, 23–57. [Google Scholar] [CrossRef] [PubMed]
- Schulze-Lefert, P.; Panstruga, R. A molecular evolutionary concept connecting nonhost resistance, pathogen host range, and pathogen speciation. Trends Plant Sci. 2011, 16, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Morris, C. E.; Moury, B. Revisiting the concept of host range of plant pathogens. Annu. Rev. Phytopathol. 2019, 57, 63–90. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.-J.; van der Does, H.C.; Borkovich, K.A.; Coleman, J.J.; Daboussi, M.-J.; Di Pietro, A.; Dufresne, M.; Freitag, M.; Grabherr, M.; Henrissat, B.; et al. Comparative genomics reveals mobile pathogenicity chromosomes in Fusarium. Nature, 2010, 464, 367–373. [Google Scholar] [CrossRef]
- Raffaele, S.; Farrer, R.A.; Cano, L.M.; Studholme, D.J.; MacLean, D.; Thines, M.; Jiang, R.H.Y.; Zody, M.C.; Kunjeti, S.G.; Donofrio, N.M.; et al. . Genome evolution following host jumps in the Irish Potato Famine pathogen lineage. Science 2010, 330, 1540–1543. [Google Scholar] [CrossRef] [PubMed]
- Inoue, Y.; Vy, T.T.P.; Yoshida, K.; Asano, H.; Mitsuoka, C.; Asuke, S.; Anh, V.L.; Cumagun, C.J.R.; Chuma, I.; Terauchi, R.; et al. Evolution of the wheat blast fungus through functional losses in a host specificity determinant. Science 2017, 357, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, F.E.; Sánchez-Vallet, A.; McDonald, B.A.; Croll, D. . A fungal wheat pathogen evolved host specialization by extensive chromosomal rearrangements. ISME J. 2017, 11, 1189–1204. [Google Scholar] [CrossRef] [PubMed]
- Cannon, P.F.; Damm, U.; Johnston, P.R.; Weir, B.S. Colletotrichum - current status and future directions. Stud. Mycol. 2012, 73, 181–213. [Google Scholar] [CrossRef]
- Talhinhas, P.; Baroncelli, R. Colletotrichum species and complexes: geographic distribution, host range and conservation status. Fungal Divers. 2021, 110, 109–198. [Google Scholar] [CrossRef]
- O’Connell, R.J.; Thon, M.R.; Hacquard, S.; Amyotte, S.G.; Kleeman, J.; Torres, M.F.; Damm, U.; Buiate, E.A.; Epstein, L.; Alkan, N.; et al. Lifestyle transitions in plant pathogenic Colletotrichum fungi deciphered by genome and transcriptome analyses. Nat. Genet. 2012, 44, 1060–1065. [Google Scholar] [CrossRef] [PubMed]
- Gan, P.; Ikeda, K.; Irieda, H.; Narusaka, M.; O’Connell, R.J.; Narusaka, Y.; Takano, Y.; Kubo, Y.; Shirasu, K. . Comparative genomic and transcriptomic analyses reveal the hemibiotrophic stage shift of Colletotrichum fungi. New Phytol. 2013, 197, 1236–1249. [Google Scholar] [CrossRef] [PubMed]
- Kusch, S.; Larrouy, J.; Ibrahim, H.M.M.; Mounichetty, S.; Gasset, N.; Navaud, O.; Mbengue, M.; Zanchetta, C.; Lopez-Roques, C.; Donnadieu, C.; et al. Transcriptional response to host chemical cues underpins the expansion of host range in a fungal plant pathogen lineage. ISME J. 2022, 16, 138–148. [Google Scholar] [CrossRef]
- Gan, P.; Narusaka, M.; Kumakura, N.; Tsushima, A.; Takano, Y.; Narusaka, Y.; Shirasu, K. Genus-wide comparative genome analyses of Colletotrichum species reveal specific gene family losses and gains during adaptation to specific infection lifestyles. Genome Biol. Evol. 2016, 8, 1467–1481. [Google Scholar] [CrossRef]
- Gan, P.; Hiroyama, R.; Tsushima, A.; Masuda, S.; Shibata, A.; Ueno, A.; Kumakura, N.; Narusaka, M.; Hoat, T. X.; Narusaka, Y.; et al. . Telomeres and a repeat-rich chromosome encode effector gene clusters in plant pathogenic Colletotrichum fungi. Environ. Microbiol. 2021, 23, 6004–6018. [Google Scholar] [CrossRef] [PubMed]
- Tsushima, A.; Gan, P.; Kumakura, N.; Narusaka, M.; Takano, Y.; Yoshihiro, N.; Shirasu, K. Genomic plasticity mediated by transposable elements in the plant pathogenic fungus Colletotrichum higginsianum. Genome Biol. Evol. 2019, 11, 1487–1500. [Google Scholar] [CrossRef] [PubMed]
- Dallery, J.-F.; Lapalu, N.; Zampounis, A.; Pigné, S.; Luyten, I.; Amselem, J.; Wittenberg, A.H.J.; Zhou, S.; de Queiroz, M.V.; Robin, G.P.; et al. Gapless genome assembly of Colletotrichum higginsianum reveals chromosome structure and association of transposable elements with secondary metabolite gene clusters. BMC Genomics 2017, 18, 667. [Google Scholar] [CrossRef] [PubMed]
- Baroncelli, R.; Cobo-Díaz, J.F.; Benocci, T.; Peng, M.; Battaglia, E.; Haridas, S.; Andrepoulos, W.; LaButti, K.; Pangilinan, J.; Lipzen, A.; et al. Genome evolution and transcriptome plasticity is associated with adaptation to monocot and dicot plants in Colletotrichum fungi. GigaScience 2024, 13, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Rech, G.E.; Sanz-Martín, J.M.; Anisimova, M.; Sukno, S.A.; Thon, M.R. Natural selection on coding and noncoding DNA sequences is associated with virulence genes in a plant pathogenic fungus. Genome Biol. Evol. 2014, 6, 2368–2379. [Google Scholar] [CrossRef]
- Inoue, Y.; Phuong Vy, T.T.; Singkaravanit-Ogawa, S.; Zhang, R.; Yamada, K.; Ogawa, T.; Ishizuka, J.; Narusaka, Y.; Takano, Y. Selective deployment of virulence effectors correlates with host specificity in a fungal plant pathogen. New Phytol. 2023, 238, 1578–1592. [Google Scholar] [CrossRef]
- Wharton, P.S.; Diéguez-Uribeondo, J. The biology of Colletotrichum acutatum. Anales del Jardín Botánico de Madrid 2004, 61, 3–22. [Google Scholar] [CrossRef]
- Wei, H.; Zhong, Z.; Li, Z.; Zhang, Y.; Stukenbrock, E.H.; Tang, B.; Yang, N.; Baroncelli, R.; Peng, L.; Liu, Z.; He, X.; Yang, Y.; Yuan, Z. Loss of the accessory chromosome converts a pathogenic tree-root fungus into a mutualistic endophyte. Plant Comm. 2023, 4, 100672. [Google Scholar] [CrossRef]
- Liu, X.; Li, B.; Yang, Y.; Cai, J.; Shi, T.; Zheng, X.; Huang, G. Pathogenic adaptations revealed by comparative genome analysis of two Colletotrichum spp., the causal agent of anthracnose in rubber tree. Front. Microbiol. 2020, 11, 1484. [Google Scholar]
- Khodadadi, F.; Luciano-Rosario, D.; Gottschalk, C.; Jurick II, W.M.; Aćimović, S.G. Unveiling the arsenal of apple bitter rot fungi: comparative genomics identifies candidate effectors, CAZymes, and biosynthetic gene clusters in Colletotrichum species. J. Fungi 2024, 10, 493. [Google Scholar] [CrossRef]
- Alkemande, J.A.; Hohmann, P.; Messmer, M.M.; Barraclough, T.G. Comparative genomics reveals sources of genetic variability in the asexual plant pathogen Colletotrichum lupini. Mol. Plant Pathol. 2024, 25, e70039. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Zhou, C.; Yang, H.; Kuang, R.; Liu, K.; Huang, B.; Wei, Y. Comparative transcriptomics and genomic analyses reveal differential gene expression related to Colletotrichum brevisporum resistance in papaya (Carica papaya L.). Front. Plant Sci. 2022, 13, 1038598. [Google Scholar] [CrossRef]
- Jiang, B.; Cai, T.; Yang, X.; Dai, Y.; Yu, K.; Zhang, P.; Li, P.; Wang, C.; Liu, N.; Li, B.; Lian, S. Comparative transcriptome analysis reveals significant differences in gene expression between pathogens of apple Glomerella leaf spot and apple bitter rot. BMC Genom. 2022, 23, 256. [Google Scholar] [CrossRef]
- Morelos-Martínez, M.I.; Cano-Camacho, H.; Díaz-Tapia, K.M.; Simpson, J.; López-Romero, E.; Zavala-Páramo, M.G. Comparative genomic analyses of Colletotrichum lindemuthianum pathotypes with different virulence levels and lifestyles. J. Fungi 2024, 10, 651. [Google Scholar] [CrossRef] [PubMed]
- Dvorianinova, E.M.; Sigova, E.A.; Mallaev, T.D.; Rozhmina, T.A.; Kudryavtsveva, L.P.; Novakovskiy, R.O.; Turba, A.A.; Zhernova, D.A.; Borkhert, E.V.; Pushkova, E.N.; Melnikova, N.V.; Dmitriev, A.A. Comparative genomic analysis of Colletotrichum lini strains with different virulence on flax. J. Fungi 2024, 10, 32. [Google Scholar] [CrossRef]
- Hsieh, D.-K.; Chuang, S.-C.; Chen, C.-Y.; Chao, Y.-T.; Lu, M.-Y.J.; Lee, M.-H.; Shih, M.-C. Comparative genomics of three Colletotrichum scovillei strains and genetic analysis revealed genes involved in fungal growth and virulence on chili pepper. Front. Microbiol. 2022, 13, 818291. [Google Scholar] [CrossRef]
- O’Connell, R.J.; Thon, M.R.; Hacquard, S.; Amyotte, S.G.; Kleeman, J.; Torres, M.F.; Damm, U.; Buiate, E.A.; Epstein, L.; Alkan, N.; et al. Lifestyle transitions in plant pathogenic Colletotrichum fungi deciphered by genome and transcriptome analyses. Nat. Genet. 2012, 44, 1060–1065. [Google Scholar] [CrossRef]
- Fu, F.-F.; Hao, Z.; Wang, P.; Lu, Y.; Xue, L.-J.; Wei, G.; Tian, Y.; Hu, B.; Xu, H.; Shi, J.; Cheng, T.; Wang, G.; Yi, Y.; Chen, J. Genome sequence and comparative analysis of Colletotrichum gloeosporioides isolated from Liriodendron leaves. Phytopathol. 2020, 110, 1260–1269. [Google Scholar] [CrossRef] [PubMed]
- Hacquard, S.; Kracher, B.; Hiruma, K.; Münch, P.C.; Garrido-Oter, R.; Thon, M.R.; Weimann, A.; Damm, U.; Dallery, J.-F.; Hainaut, M.; et al. Survival trade-offs in plant roots during colonization by closely related beneficial and pathogenic fungi. Nat. Commun. 2016, 7, 11362. [Google Scholar] [CrossRef] [PubMed]
- Cannon, P.F.; Damm, U.; Johnston, P.R.; Weir, B.S. Colletotrichum - current status and future directions. Stud. Mycol. 2012, 73, 181–213. [Google Scholar] [CrossRef] [PubMed]
- Jayawardena, R.S.; Hyde, K.D.; Damm, U.; Cai, L.; Liu, M.; Li, X.H.; Zhang, W.; Zhao, W.S.; Yan, J.Y. Notes on currently accepted species of Colletotrichum. Mycosphere 2016, 7, 1192–1260. [Google Scholar] [CrossRef]
- Ujimatsu, R.; Takino, J.; Nakamura, M.; Haba, H.; Minami, A.; Hiruma, K. A fungal transcription factor BOT6 facilitates the transition of a beneficial root fungus into an adapted anthracnose pathogen. 2024, submitted.
- Lu, H.; Zou, W.X.; Meng, J.C.; Hu, J.; Tan, R.X. New bioactive metabolites produced by Colletotrichum sp., an endophytic fungus in Artemisia annua. Plant Sci. 2000, 151, 67–73. [Google Scholar] [CrossRef]
- Díaz-González, S. ; González-Sanz, C; González-Bodi, S.; Brunner, P.M.F.; Sacristán, S. Plant Growth Promoting fungal endophyte Colletotrichum tofieldiae Ct0861 reduces mycotoxigenic Aspergillus fungi in maize grains. bioRxiv.
- Chandini; Kumar, R.; Kumar, R.; Prakash, O. The impact of chemical fertilizers on our environment and ecosystem. In Research Trends in Environmental Sciences, 2nd ed.; Sharma, P., Ed.; AkiNik Publications: New Delhi, India, 2019; pp. 71–85. [Google Scholar]
- Tripathi, S.; Srivastava, P.; Devi, R.S.; Bhadouria, R. Influence of synthetic fertilizers and pesticides on soil health and soil microbiology. In Agrochemicals Detection, Treatment, and Remediation; Prasad, M.N.V., Ed.; Butterworth-Heinemann: Oxford, United Kingdom, 2020; pp. 25–54. [Google Scholar]
- Berruti, A.; Lumini, E.; Balestrini, R.; Bianciotto, V. Arbuscular mycorrhizal fungi as natural biofertilizers: let’s benefit from past successes. Front. Microbiol. 2016, 6, 1559. [Google Scholar] [CrossRef]
- Ceballos, I.; Ruiz, M.; Fernández, C.; Peña, R.; Rodríguez, A.; Sanders, I.R. The in vitro mass-produced model mycorrhizal fungus, Rhizophagus irregularis, significantly increases yields of the globally important food security crop cassava. PLoS ONE 2013, 8, e70633. [Google Scholar] [CrossRef]
- Reducing the allelopathic effect of Parthenium hysterophorus L. on wheat (Triticum aestivum L.) by Pseudomonas putida. Plant Growth Regul. 2012, 66, 155–165. [CrossRef]
- Yang, Z.-D.; Li, Z.-J.; Zhao, J.-W.; Sun, J.-H.; Yang, L.-J.; Shu, Z.-M. Secondary metabolites and PI3K inhibitory activity of Colletotrichum gloeosporioides, a fungal endophyte of Uncaria rhynchophylla. Curr. Micorbiol. 2019, 76, 904–908. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.-X.; Tan, H.-B.; Chen, Y.-C.; Li, S.-N.; Li, H.-H.; Zhang, W.-M. Secondary metabolites from the Colletotrichum gloeosporioides A12, an endophytic fungus derived from Aquilaria sinensis. Nat. Prod. Res. 2017, 32, 2360–2365. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Xue, M.; Shen, Z.; Jia, X.; Hou, X.; Lai, D.; Zhou, L. Phytotoxic secondary metabolites from fungi. Toxins, 2021, 13, 261. [Google Scholar] [CrossRef] [PubMed]
- Sirrenberg, A.; Göbel, C.; Grond, S.; Czempinski, N.; Ratzinger, A.; Karlovsky, P.; Santos, P.; Feussner, I.; Pawlowski, K. Piriformospora indica affects plant growth by auxin production. Physiol. Plant. 2007, 131, 581–589. [Google Scholar] [CrossRef]
- Nguyen, T.A.N.; Higa, T.; Shiina, A.; Devi Utami, Y.; Hiruma, K. Exploring the roles of fungal-derived secondary metabolites in plant-fungal interactions. Physiol. Mol. Plant Pathol. 2023, 125, 102021. [Google Scholar] [CrossRef]
- Tian, D.; Traw, M.B.; Chen, J.Q.; Kreitman, M.; Bergelson, J. Fitness costs of R-gene-mediated resistance in Arabidopsis thaliana. Nature 2003, 423, 74–77. [Google Scholar] [CrossRef]
- Boyd, L.A.; Ridout, C.; O’Sullivan, D.M.; Leach, J.E.; Leung, H. Plant-pathogen interactions: disease resistance in modern agriculture. Trends Genet. 2013, 29, 233–240. [Google Scholar] [CrossRef]
- Stuthman, D.D.; Leonard, K.J.; Miller-Garvin, J. Breeding crops for durable resistance to disease. Adv. Agron. 2007, 319–367. [Google Scholar]
- Jiang, B.-B.; Dong, Y.-W.; Hao, G.-F. Virulence factor xyloglucanase is a potential biochemical target for fungicide development. J. Agric. Food Chem. 2023, 71, 10483–10485. [Google Scholar] [CrossRef] [PubMed]
- Materatski, P.; Varanda, C.; Carvalho, T.; Dias, A.B.; Campos, M.D.; Gomes, L.; Nobre, T.; Rei, F.; Félix, M.R. Effect of long-term fungicide applications on virulence and diversity of Colletotrichum spp. associated to olive anthracnose. Plants 2019, 8, 311. [Google Scholar] [CrossRef]
- Karim, M.M.; Usman, H.M.; Tan, Q.; Hu, J.-J.; Fan, F.; Hussain, R.; Luo, C.-X. Fungicide resistance in Colletotrichum fructicola and Colletotrichum siamense causing peach anthracnose in China. Pestic. Biochem. Physiol. 2024, 203, 106006. [Google Scholar] [CrossRef] [PubMed]
- Rogério, F.; de Castro, R.R.L.; Massola Jr., N. S.M.; Boufleur, T.R.; dos Santos, R.F. Multiple resistance of Colletotrichum truncatum from soybean to QoI and MBC fungicides in Brazil. J. Phytopathol. 2024, 172, e13341. [Google Scholar] [CrossRef]
- Shi, N.-N.; Lian, J.-P.; Qiu, D.-. .Z.; Chen, F.-R.; Du, Y.-X. Resistance risk and molecular mechanism associated with resistance to picoxystrobin in Colletotrichum truncatum and Colletotrichum gloeosporioides. J. Integr. Agric. 2023, 22, 3681–3693. [Google Scholar] [CrossRef]
- Khan, S.M.; Ali, S.; Nawaz, A.; Bukhari, S.A.H.; Ejaz, S.; Ahmad, S. Integrated pest and disease management for better agronomic crop production. In Agronomic Crops; Hasanuzzaman, M., Ed.; Springer: Singapore, 2019; pp. 385–428. [Google Scholar]
- Kolainis, S.; Koletti, A.; Lykogianni, M.; Karamanou, D.; Gkizi, D.; Tjamos, S.E.; Paraskeuopoulos, A.; Aliferis, K.A. An integrated approach to improve plant protection against olive anthracnose caused by the Colletotrichum acutatum species complex. PLoS ONE 2020, 15, e0233916. [Google Scholar] [CrossRef]
- Wheeler, M.H.; Greenblatt, G.A. The inhibition of melanin biosynthetic reactions in Pyricularia oryzae by compounds that prevent rice blast disease. Exp. Mycol. 1988, 12, 151–160. [Google Scholar] [CrossRef]
- Howard, R.J.; Ferrari, M.A.; Roach, D.H.; Money, N.P. Penetration of hard substrates by a fungus employing enormous turgor pressure. Proc. Natl. Acad. Sci. USA, 1991, 88, 11281–11284. [Google Scholar] [CrossRef]
- de Jong, J.C.; McCormack, B.J.; Smirnoff, N.; Talbot, N.J. Glycerol generates turgor in rice blast. Nature 1997, 389, 244–245. [Google Scholar] [CrossRef]
- Kubo, Y.; Suzuki, K.; Furusawa, I.; Yamamoto, M. Melanin biosynthesis as a prerequisite for penetration by appressoria of Colletotrichum lagenarium: site of inhibition by melanin-inhibiting fungicides and their action on appressoria. Pestic. Biochem. Physiol. 1985, 23, 47–55. [Google Scholar] [CrossRef]
- Chakraborty, A.; Ray, P. Mycoherbicides for the noxious meddlesome: can Colletotrichum be a budding candidate? Front. Microbiol. 2021, 12, 754048. [Google Scholar] [CrossRef]
- Ditommaso, A.; Watson, A.K. Impact of a fungal pathogen, Colletotrichum coccodes on growth and competitive ability of Abutilon theophrasti. New Phytol. 1995, 131, 51–60. [Google Scholar] [CrossRef]
- Boyette, C.D.; Abbas, H.K.; Johnson, B.; Hoagland, R.E.; Weaver, M.A. Biological control of the weed Sesbania exaltata using a microsclerotia formulation of the bioherbicide Colletotrichum truncatum. Am. J. Plant Sci. 2014, 5, 2672–2685. [Google Scholar] [CrossRef]
- Boyette, C.D. Host range and virulence of Colletotrichum truncatum, a potential mycoherbicide for hemp sesbania (Sesbania exaltata). Plant Dis. 1991, 75, 62–64. [Google Scholar] [CrossRef]
Figure 1.
Proposed idea of lifestyle in plant-associated fungi.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



