Submitted:
25 January 2025
Posted:
27 January 2025
You are already at the latest version
Abstract
Selenium is an essential micronutrient that is critical for the maintenance of human health. It has a significant impact on immune function and is effective in the prevention of several metabolic-related illnesses. Furthermore, Se deficiency affects millions worldwide, particularly in regions such as China, Eastern Europe, and Africa. This review examines the current knowledge in Se biofortification, specifically focusing on the Se application to plants and soils, the mechanism of Se uptake and utilization, genetic variation in Se accumulation, and genetic modification to achieve high biofortification targets. Data from many studies show substantial increases in the Se levels in biofortified crops. For instance, foliar application of Se-enriched fertilizers increased Se concentration in wheat by 42-67 µg/kg and in corn by 19-36 µg/kg. Se biofortification in rice showed Se content variations ranging from 5 to 1370 ng/g, with a substantial increase observed from Se-rich soils. The qualitative analysis demonstrates that public health is improved when people have access to fortified cereals and pulses. There are many economic benefits that lead to a reduction in healthcare costs and increased productivity. Despite these promising outcomes, many challenges remain to overcome. Our findings suggest that future research should focus on developing uniform distribution of Se in major agricultural crops, optimizing the bioavailability of the nutrients and developing consumer acceptance. Furthermore, refining biofortification techniques to deliver Se into the edible parts, particularly to the grain in cereals, exploring the genetic basis of Se uptake in crops, and developing public health policies to support widespread adoption is essential. Se biofortification of agriculturally important crops is the main strategy to combat global Se malnutrition, which will have significant economic and health benefits. Continued multidisciplinary research and policy support are essential to realize this target ensuring global food and nutrient security in the long term.
Keywords:
Micronutrient deficiency
; bioavailability
; agriculture
; cereals
; food security
Introduction
Selenium is needed for all the living beings including microorganisms, plants, animals and humans (Fu et al. 2002; Novoselov et al. 2002; Lobanov et al. 2007). Micronutrient deficiency or malnutrition is severe among humans (Qaim et al. 2007). Iron, zinc, copper, calcium, magnesium, iodine, and selenium were found to be commonly insufficient in the human diet (White and Broadley 2009; Guerinot and Salt 2001; Goldhaber 2003). This is called chronic hunger (Stein 2009) or hidden hunger (FAO 2005), or micronutrient malnutrition (Yang et al. 2007). Children and breastfeeding mothers are the most affected groups to malnutrition (WHO 2002). Growing staple crops in soils with limited micronutrients (Mao et al. 2014), low income, lack of awareness of a balanced diet (Qaim et al. 2007) and increased grain yield by decreasing nutrient density in cereals and legumes (Garvin et al. 2006, Fan et al. 2008a) are identified as the main reasons for micronutrient malnutrition. Micronutrient malnutrition is common in poor households who are satisfied with their food needs through cereals and pulses (Qaim et al. 2007) which leads to loss of immunity, mental impairments, and high mortality rates (Amarya et al. 2015).
Among micronutrients, deficiency in Fe, Zn, and Se was reported in 50% of the population (Zhao and McGrath 2009), and 260 million are iodine or Se deficient (Caballero 2002), billion are iron deficient, 2.7 billion are Zn deficient, and the majority is lacking vitamins (Haddad et al. 2004, Hotz and Brown 2004). According to Combs (2001), Se is deficient in 500−1000 million people worldwide and this equals to 15% of the world population (Thavarajah et al. 2011).
The World Health Organization has identified Se-deficient, Se-low and Se-adequate countries (Yuan et al. 2012). Mombo et al. (2016) have mapped the locations according to Se availability worldwide (Figure 1).
Se Dietary Intake and Health Implications
Two independent groups discovered Se in glutathione peroxidase (GPx) in 1973 (Pedrero and Madrid 2009). GPx activity is considered a feature to evaluate Se requirement in the human body (Levander et al. 1983). According to the function of GPx, the daily requirement of Se for a woman is 55 µg and for a man is 77 µg per day (Pedrero and Madrid 2009). The recommended daily intake of Se varies by region. A daily Se intake of 55 μg is recommended for Indian subtropical regions and temperate North American regions (Thavarajah 2011), and 60–75 µg for Europe, Australia, and Asia (Giray and Hincal 2004; Lyons et al. 2005a; Spallholz et al. 2004). On the other hand, more than 300 µg of Se per day is a critical level, according to findings in Europe (Thiry et al. 2012) and according to Wu et al. (2015), the recommended nutritional intake of Se is 50-200 µg/day. Hence, as a range, 55 - 200 µg Se per day has been recommended for humans (Rayman 2000; Thomson 2004; WHO 2009). Using the average values from two studies from China and New Zealand, Goldhabar (2003) arrived at a value of 450 µg Se per day as the Estimated Average Requirement (EAR) of Se.
The required amount of a micronutrient for a daily diet can be supplemented by adding such an amount to the food during any food processing step (Graham et al. 2007). Daily Se intake for humans must be assessed carefully since the Se content and Se bioavailability differ with the soil in which plants were grown and the form of selenium ingested. It is not possible to recommend dietary items for Se intake because the soil in which they are cultivated determines the content of Se in the food. However, knowing the origin of, for example, wheat in the bread, the Se adequacy of the diet can be judged. For example, changes in blood Se in Hamilton, New Zealand were consistent with values calculated from variations in dietary intake of Se of wheat products from the blending of New Zealand wheat (11 ng/g Se) with imported Australian wheat (150 ng/g Se) (Watkinson, 1981). Furthermore, there was a positive correlation of selenium level in sunflower seeds with the concentration in the soil it was grown in an experiment with varying soil Se concentrations (Dhillon and Dhillon, 2009).
Se in the Human Body
Balancing Se concentration in our blood and cells has important health implications. The excess of Se can be toxic, and its deficiency can result in disease.
Se Toxicity in Humans
Integrating Se in selenocysteine (SeCys) and selenomethionine (SeMet) into the proteins (Pedrero and Madrid 2009), which alters protein structure, is the cause of Se toxicity in humans (Terry et al. 2000; Ellis and Salt 2003). Se is needed in a very small amount (Kieliszek and Błażejak 2013), and in humans, it can lead to health issues if consumed in excess. The main reason for Se toxicity is due to the narrow range of daily Se requirement (Thornton et al. 1983; Rosenfeld and Beath 2013; Yang et al., 1983). This is also called a “double-edged sword element” and an “essential toxin” (Levander and Burk, 2006). Due to its narrow requirement and intolerable toxic ranges, Se supplementation should be prescribed carefully to meet individual dietary needs. Plants cultivated in soils containing more than 3.0 mg kg−1 of Se are not safe for consumption (Hawkesford and Zhao, 2007). Herbivores feeding on Se hyperaccumulator plants convert methyl-SeCys to SeCys, and SeCys integrates into proteins causing Se toxicity (Freeman et al. 2006b). The daily average Se intake of 550 µg caused selenosis symptoms in a high Se region in Hubei Province of China (Huang et al. 2013). Selenosis causes skin lesions, nerve damage, and liver enlargement (McLaughlin et al. 1999). Continuous work with some electronic industries causes acute Se intoxication (Szpunar et al. 2003). Se toxicity affects not only humans, but also fish and land animals (Yang et al. 1983; Dörr et al. 2008) and such food items are not safe for consumption.
Se Deficiency in Humans
In many regions of the world, Se consumption is lower than 40 μg per day (Combs 2001; Navarro -Alarcon and Cabrera-Vique 2008). Se has the ability to prevent heart diseases and muscle disorders (Nève 1996; Fernandes et al. 2014). For example, Keshan and Kashin-Beck diseases are reported in the regions with insufficient Se in the soil (Yang et al. 2007). Keshan disease is an endemic myocardiopathy and Kashin-Beck disease reduces human working ability and well-being (Yang et al. 2007) and causes osteoarthrosis (Allander 1994). In humans, having low levels of Se has been linked to a higher likelihood of contracting HIV/AIDS and other viral diseases (Gailer et al. 2000; Rayman 2000) and male infertility (Nève 1996; Ellis and Salt 2003). Any further reduction of Se leads to the risk of arsenic (As) toxicity in humans (Kibriya et al. 2007). Most importantly, epidemiological studies have indicated an inverse relationship between Se intake and the incidence of certain cancers in humans and tumors in mammals. Blood or plasma levels of Se are usually lower in patients with cancer than healthy humans (Whanger 2004) and inclusion of methylselenocystaine (MeSeCys) in the diet of mammals reduces the incidence of certain cancers (McKenzie et al. 2009). Various organic compounds containing Se play distinct roles within the human body involving a number of metabolic processes (Table 1).
Alleviation of Se Malnutrition in Humans Through Food Intake
Se malnutrition can be addressed in five steps: finding Se-rich crop species, transformation of crop species unable to metabolize Se to enable them metabolise Se, increasing micronutrient bioavailability, raising consumer awareness of malnutrition and nutrients, and fertilizer development and application (Yang et al. 2007; Stein 2009; Brummell et al. 2011).
Food diversification is the main solution to avoid Se deficiency in poor people’s diets (Meenakshi et al. 2010), while enriched supplements are administered in developed countries (Schrauzer 2001; Finley 2005). Fifty-five micrograms of daily dietary Se intake (Bendich 2001) can be supplemented in inorganic form. Inorganic sodium selenite is a commonly used form for Se supplementation that is prepared using yeast (Saccharomyces cerevisiae) (Hilton et al. 1982; Gatlin and Wilson 1984; Pedrero and Madrid 2009; Felton et al. 1996; Schram et al. 2008).
Se is supplied in salt in treating Kashin-Beck disease (Bawa et al. 1992). The amount of Se that is available in commercially produced Se-enriched products varies (Pedrero and Madrid 2009). The toxicity of Se has been reported due to this discrepancy (Helzlsouer et al. 1985) and the factors that interfere to reduce mineral absorption in the gut must be addressed to overcome this problem (White and Broadley 2009). Increasing pro nutrients and decreasing antinutrients such as oxalate, polyphenols or phytate are some approaches for enhancing Se absorption in the human gut (Graham 2007; Brinch-Pedersen et al. 2007; Graham et al. 2007).
Organic Se forms are more bioavailable than inorganic forms in food (Pedrero and Madrid 2009). and have a higher retention power and a lower toxicity (Brummell et al. 2011; Thiry et al. 2012). Further, the bioavailability of Se is higher in plant-based food than in meat (Combs 2001). Bioavailable Se in goat and sheep milk is greater than Se in cow milk (Shen et al. 1996). Se biofortification in crops or food fortification are reliable solutions for increasing Se availability and consumption (McKenzie et al. 2009; White and Broadley 2009). Future research need to be focused on enhancing the bioavailability of Se in both animal and plant sources. Selenoprotein production in the body can be achieved by ingestion of Se as sodium selenate, selenite, and SeCys, which is mainly practiced for animal rearing for meat (Pedrero and Madrid 2009). The metabolic pathways of Se cycling within the human body are illustrated in Figure 2.
Selenium in the Environment
Se compounds in the environment are diverse (Figure 3). Se occurs both in oxidation and in organic forms in the natural environment (Qin et al. 2012). Twenty-five selenoproteins contain Se as structural proteins and enzymes (Kryukov, Castellano et al. 2003; Lu and Holmgren 2009). Glutathione peroxidases, thioredoxin reductases and iodothyronine deiodinases are Se containing enzymes (Rayman 2012; Zhang 2009; Stein 2009). Selenoprotein production in the body can be achieved by ingestion of selenium as sodium selenate, selenite, and SeCys, which is mainly practiced for animal rearing (Pedrero and Madrid 2009). Trimethyl selenonium in the urine and dimethyl selenide (DMSe) in exhaled gases are two bodily excretions of Se (Ganther 1986; Stadtman 1991). A few Se-contained compounds in plants are SeMet, SeCys, ϒ-glutamyl lenomethyl selenocystine, MeSeCys, selenocystathione, selenohomocysteine and selenomethyl selenomethionine (Reeves and Baker 2000; Montes-Bayón et al. 2002). Se containing volatile gases, namely DMSe, dimethyl diselenide (DMDSe), diethyselenide (DESe) and diethyldiselenide (DEDSe) are emitted to the environment (Dauchy et al. 1994; Gómez-Ariza, et al., 1998). Microbial activities based on oxidation, reduction, methylation, and demethylation change the status of Se (Stolz et al., 2006). Some bacterial and fungal species can convert Se substrates SeO42−, SeO32−, Se0, SeS2, H2SeO3, NaSeH into methylated products such as DMSe, DMDSe, methylselenol, dimethyl selenide sulfide (DMSeS), and DMSeDS (Eswayah et al., 2016). The predominant groups of Se-methylating organisms that can be found in soils and sediments are bacteria and fungi, while bacteria are the active Se-methylating organisms in the aquatic environments. The volatile Se compound produced by most organisms is DMSe and is non-toxic to mammals (Frankenberger and Arshad, 2001).
Se in Plants
The bioconcentration factor (BCF) in the soil for Se is calculated as the proportion of Se in the plant to Se in the soil (Wang et al. 2012). BCF is reported in a range of 0.23 to 1 (Dhillon and Dhillon 2009; Sun et al. 2010; Wang, et al. 2012; Bujdoš et al. 2005). Se mainly acts as an antioxidant in plants (Xue et al. 2001; Djanaguiraman et al. 2005). Yield increment with Se application is also evident for many crops and pastures (Qaim et al. 2007) along with iron and zinc for improved plant vigour (Graham et al., 1999; Welch 2002). It has been reported that Se promotes the carbohydrate metabolism in Solanum tuberosum (L.) (Turakainen et al. 2004).
The most bioavailable form of Se in soils is selenate, which is taken up by plants using enzymes of the sulphur assimilation pathway since most of the transporters and enzymes involved in the uptake, translocation, and assimilation cannot discriminate between the S and the Se forms of their substrates (Sors et al. 2005; Brummel et al. 2011). Selenate is reduced to selenite and converted to SeCys (Sors et al. 2005). In Se hyperaccumulators, SeCys is methylated to MeSeCys by the action of selenocysteine methyltransferase (Neuhierl et al. 1996; Brummell et al. 2011
Inorganic Se can be converted to the organic form of Se (e.g.; MeSeCys and glutamyl selenomethyl selenocysteine) in plants (Combs and Gray 1998; Tapiero et al., 2003; McKenzie et al. 2009; Brummell et al. 2011). The predominant compounds of Se in different crops are listed in Table 2. Various parts of plants accumulate distinct quantities of Se. For example, in corn, the order of Se accumulation changes as leaf >root>seed>stalk (Wang et al. 2012). The increasing order of Se in rice was arranged as husk>straw>bran>whole grain >polished rice (Sun et al. 2010). Typically, the seed contains a lower concentration of Se compared to other parts of cereals. This indicates that the Se deficiency in our diets is caused not by the poor intake of Se by the cereal crop, but rather in its transportation into the grain. Therefore, the mechanism responsible for loading Se into the grain endosperm is likely the factor that restricts its accumulation. Nevertheless, there is a lack of comprehensive understanding regarding these processes.
Se in Cereals
Around 60–80% of Se requirement in the global human diet is provided by grains (Stadlober et al. 2001). Northern Indian villagers obtain ∼90% of their Se requirement solely from grains (Hira et al. 2004). In contrast, cereals contribute around 18% of the total Se intake in Finland (Eurola et al., 1990). Cereals significantly convert selenate to SeMet (Stadlober et al., 2001). Oat has an increased Se content of around 72.5–203.08% by fertilizer application (McKevith, 2004). The foliar application Se effectively increased Se content in whole wheat (42-67 µg/kg) and corn (19-36 µg /kg) (Milovac et al., 1998). The Se content in wheat is mostly influenced by the geographical region (Hawkesford and Zhao 2007). In regions where the soil is rich in Se, cereals can contribute significantly to daily Se requirements.
The required amount of Se for the human diet can be achieved by foliar application of selenate, or soil application of selenite in rice (Boldrin et al. 2013). The Se content of rice varies even within the same country, and is as follows: China 2–1370 ng/g, India 5–371 ng/g, Japan 26–109 ng/g, Thailand 6–487 ng/g and USA 6–406 ng/g (Williams et al. 2009; Kelly et al. 2002). Around 75% of the rice samples do not reach the human Se requirements according to Williams et al. (2009). This content can be boosted approximately by double through improved management practices (Fang et al. 2008). The rice bran contains a Se content that is ten times more than that found in the endosperm. This suggests that there is an important barrier in the transfer of Se into the inner endosperm of the grain. Not only Se, but other micronutrients such as Fe and Zn also exhibit similar trends (Kamral et al., 2023). The primary type of Se found in rice bran is SeMet, as reported by Li et al. (2010). SeMet is mostly accumulated in rice bran when Se is added as a fertiliser (Stadlober et al. 2001). Other forms, such as SeCys and MeSeCys, are also present in rice (Li et al. 2010; Sun et al. 2010). However, MeSeCys and SeCys are present in lower concentrations (Sun et al. 2010). Though 80% of Se is SeMet (Guzmán et al. 2009), up to 50% of MeSeCys (∼50%) has also been reported in rice in other studies (Williams et al. 2009).
The nutritive value of Se in the crop depends on the conversion of inorganic to organic Se (Whanger 2002; Hawkesford and Zhao 2007). Inorganic Se is easily converted into organic Se by rice (Li et al. 2010). The spatial distribution pattern of Se in rice grains has been demonstrated by applying selenite, SeMet, and MeSeCys (Carey et al., 2012). It has been demonstrated that the form of Se application decides Se localization in the rice grain: In selenite-treated grains, ovular vascular trace region accumulates Se and, in the SeMet applied rice plants Se is high in seed external layers and in the endosperm. In MeSeCys applied plants, Se was high in the aleurone layer and the embryo (Carey et al. 2012). These findings suggest that the high molecular mass of MeSeCys might impede the passage into the inner endosperm of the grain.
The rice husk inorganic Se content is ∼10 times greater than the endosperm inorganic Se and outer pericarp and aleurone tissues also contain higher Se than the endosperm (Carey et al., 2012). Rice endospermic Se is mainly organic, while rice bran contains both organic and inorganic Se (Williams et al. 2009). These findings indicate that either the transfer of Se into the grain or the synthesis of organic Se in the inner endosperm is the component that limits the organic Se content in the cereal grain. Moreover, these findings also suggest that cereals, specifically grains such as wheat and rice, have a major role in fulfilling the global Se dietary needs. These needs can be partially met through agronomic fortification methods, such as applying Se fertilizers, implementing specific cultivation practices, and utilizing cultivars with increased potential to accumulate Se.
Se in Soil
The Se speciation is determined by the physical features of the soil (Frankenberger and Benson 1994) (Table 3). Se is found in soil primarily as selenide, selenite, and selenate ions, while it is typically present as selenate or selenite ions (Hopper and Parker 1999). The availability of Se species in the soil is given as selenate > SeMet > SelCys > selenite > elemental se > selenide (Kabata-Pendias, and Szteke, 2015). Selenate ions (SeO42−) can form salts with various cations (eg; Na2SeO4). Selenite is a Se-containing ion with oxygen (SeO32−), found in salts (eg; Na2SeO3), while selenide (Se2−) is a compound formed by the combination of Se with another element, typically a metal or non-metal, in the form of a selenide ion (eg; H2Se, ZnSe).
Se Content in Upland and Lowland Soils
The levels of oxidized Fe and Mn bound to Se are greater in upland soil compared to lowland soil (Wang et al. 2012). For example, exchangeable Se is around 1% in the upland soils (Wang et al. 2012). It has been demonstrated that adding selenate to aerobic soil is the best way to boost grain Se content in rice (Li et al. 2010). When Se is applied with the fertilizer to upland soils, the retention of Se in the seeds is higher than when added to submerged soils (Li et al. 2010). It has been proposed that under dryland conditions, microbial activity is enhanced in the presence of CaCO3 accelerating the release of Fluvic acid-Se into inorganic and organic Se that increases Se absorption by roots (Qin et al. 2012). Alternatively, under anaerobic conditions, particularly in rice, grain nutrient dilution occurs, resulting in increased biomass and grain yield, which in turn may lead to a decrease in Se concentration in the grains. Hence, aerobic rice (upland rice) retains a higher Se content in the seeds than in submerged soils (paddy rice) (Li et al. 2010).
Organic matter-bound Se is higher than Fe and Mn- bound Se in lowland soils (Wang et al. 2012). Releasing Se from organic matter is faster than releasing it from Fe or Mn oxide-bound Se (Munier-Lamy et al. 2007). Elemental Se and selenide are more stable in lowland soils in their less accessible forms (Elrashidi et al. 1987). For example, when lowland soil is supplied with selenite, the observed Se types were selenite, SeMet, and Se oxide (Li et al. 2010), suggesting that two species are the immediate transformed products in an organic background. Soil adsorption of Se is prominent in lowland soils since the clay fraction is comprised of Fe and Al oxides (Mouta et al. 2008). However, adding selenate or selenite to lowland soil is not effective (Li et al. 2010) since selenite (SeO32-) is thermodynamically stable (Kirk 2004). Exchangeable Se is around 16.1% in lowland soils (Wang et al. 2012), Waterlogged and submerged soil conditions with high selenium (Se) levels are more effective in mitigating excessive selenium entry of Se into plants (Williams et al. 2009). Under anaerobic conditions, soil pH decreases (Li et al., 2010), soil redox potential decreases subsequently (Mikkelsen et al., 1989b) and oxide and hydroxide forms of iron are released into the soil solutions, and Se is adsorbed to oxides and hydroxides (Grybos et al., 2007; Fendorf et al., 2008; Grybos, et al., 2007). Se is more available in sandy soils than in sandy clay soils (Fernandes et al. 2014) high in cretaceous shale (Ohlendorf et al. 1986) and in volcanic tuff and in the black carbonaceous soils (Kunli et al. 2004). Therefore, it is essential to take into account the chemical and physical features of the soil while formulating strategies for Se agronomic fortification.
Approaches to Manipulation of Se in Plants for Better Human Health, Crop Yield and Soil Remediation
As Se can be deficient in diets or excessive, manipulating its content in plants is important for better health outcomes for humans.
Improving Bioavailability of Se in Soil
The bioavailability of Se in soil was reported as 1.1–3.4% in UK soil samples by Stroud et al. (2010). The bioavailable forms of Se are less than the total Se contents in the soil and this is the case for any soil type (Zhang and Moore 1996). In general, the availability of Se for plants is primarily determined by the redox potential and pH of the soil, soil parental material, soil type, and climatic conditions, as well as Se speciation (Mikkelsen et al. 1989b; Tan et al. 1994; Terry et al. 2000; Wang and Gao 2001). Continuous irrigation also increases Se availability in the soil (Ohlendorf et al. 1986). For example, the soils of the Hubei province, Ziyang and Daba regions of South Qinling in China (Kunli et al. 2004), and Punjab region in India (Sharma et al. 2010) and South Dakota in USA (Gerla, et al. 2011) are all rich in Se. Further, the type of fertilizer and fertilizer application time also determines the Se availability in the soil (Wang et al. 1996; Kahakachchi et al. 2004; Kápolna and Fodor 2006). The bioavailability of Se can be improved through the implementation of crop management practices. The retention of Se in grain is more effective when Se is applied closer to the heading stage than when it is applied during soil preparation. Drainage during the maturity period allows increased Se accumulation in rice grains. SeO42− -enriched N-fertilizers improve Se uptake; this is particularly prominent at the heading stage (Premarathna et al. 2012). The rhizosphere is the narrow region of soil that surrounds and influenced by the plant roots. Selenobacteria in this zone can be used to enhance the bioavailability of Se in soil (Acuña et al. 2013). It has been established that Fulvic acid- Humic acid-, and organic matter-bound Se is the available Se sink in the soil (Qin et al. 2012). Consequently, the addition of chelating chemicals to soil has the potential to enhance the bioavailability of Se.
Se Uptake by Plants
Plants absorb Se from the soil and distribute it to other parts through xylem and phloem. Selenate and selenite are the two main forms for the absorption of Se by plants (Ellis and Salt, 2003). Selenite is the main form of Se found in anaerobic soils, and selenate is more common in aerobic soils (Avila et al. 2013). The most abundant form of Se for plants is selenate, as selenite tends to bond with oxides or hydroxides of iron or manganese (Barrow and Whelan 1989). The availability of Iron oxyhydroxide Se is also greater in soils (Rayman 2008). BCF is higher in selenate-treated soils than in selenite-treated soils (Munier-Lamy et al. 2007), suggesting that Se is actively, not passively, transported in the plant. Se is a sulfur analogue and selenate (SeO2-) is a sulfate (SO42−) analogue and Se transportation and accumulation follow sulfur metabolic pathway in the plant cell (Sors et al. 2005). The similarity in structure and charge between selenate and sulphate leads to competition for uptake and assimilation in plants (Ellis and Salt 2003; Hoewyk et al., 2007; White et al. 2007; Tamaoki et al., 2008; White and Broadley 2009; Eiche et al., 2015). The preference of sulphate transporters for selenate and sulphate may vary among plant species and under varying nutritional conditions (Zhu et al., 2009). The absence of sulfate transporters prevents selenate from being absorbed into the cell. (Shibagaki et al. 2002; Yoshimoto et al. 2002; Bell et al. 1992). High sulfur doses reduce the Se contents in plants when Se is applied as selenate (Avila et al. 2013). However, some plants have Se-specific transporters (Feist and Parker 2001; Schiavon et al. 2015; Pilon-Smits and Quinn 2010). SULTR1 is the dominant selenate transporter in Arabidopsis thaliana (Barberon et al. 2008).
An antagonist uptake pattern of selenate and selenite in plants has been reported (Zayed and Terry 1992). Selenate is transported to shoot, and selenite has an affinity to accumulate in the roots (Li, et al. 2008). Further, selenate is better than selenite in the accumulation process (Zhu et al. 2009). Selenite uses both passive diffusion (Sors et al. 2005; Li et al. 2010) and phosphate transporters (Li et al. 2010) to enter the plant. The adsorption affinity of selenite with the soil particles is greater than selenate (Kamei-Ishikawa et al. 2007). Further, compared to selenate, rhizosphere microbes volatilize selenite significantly (Azaizeh et al., 2003). Se translocation to shoot is limited once applied as selenite (Arvy 1993; de Souza et al. 1998; Hopper and Parker 1999). Glutathione is important to convert selenite to selenate (Ellis and Salt 2003; Whanger 2004; White et al. 2007).
The organic forms of Se are deposited in roots when selenite is applied (Zayed et al. 1998). The mobility of organic Se (SeMet or MeSeCys) is greater in the plants than inorganic Se. Organic Se species are transported exclusively through phloem whereas inorganic form of Se is transported through both phloem and xylem (Carey et al. 2012). Se enters grain through both the xylem and phloem sap (Krishnan and Dayanandan 2003). SeMet and MeSeCys are dispersed throughout the external grain layers and in the endosperm and MeSeCys also gets deposited in the embryo (Carey et al. 2012).
Se Toxicity in Plants
Substitution of cysteine (Cys) and methionine (Met) by SeCys and SeMet, respectively, results in Se toxicity (selenosis) in plants (Brown and Shrift 1981). According to Guptha and Guptha (2017) Se toxicity manifests due to either malformed selenoproteins or oxidative stress. Many studies indicate that Se toxicity is a cause of oxidative stress (Chen et al., 2008; Schiaven et al., 2012; Gupta and Gupta 2017). Se is volatilized in plants after converting SeMet to DMSe, and several other forms (Ellis and Salt 2003; Zhu et al. 2009; Rayman et al. 2008; Terry et al. 2009). Most of the metabolic activities associated with Se processing within the plant cell take place inside the chloroplast (Leustek et al. 2000) (Figure 4).
The Ability of Different Plants to Metabolize and Accumulate Se
Depending on their ability to absorb and metabolise Se, plants are divided into three groups (Terry et al. 2000; Ellis and Salt 2003). The majority of plants are ‘non-accumulators’ of Se. They can only accumulate low levels of Se from the soil; less than 100 mg Se kg−1 DW, and usually much less (McKenzie et al. 2009). These plants take up Se non-specifically via the sulphur (S) assimilation pathway (White et al. 2007), ultimately forming SeCys and SeMet, which are mis-incorporated into proteins resulting in Se toxicity symptoms such as stunting, necrotic lesions on the leaves and reduced root growth (McKenzie et al. 2009). In contrast, Se hyperaccumulators, such as Astragalus bisulcatus and Stanleya pinnata, when grown in Se-rich soils can take up and accumulate large quantities of Se (up to several g Se kg−1 DW) without apparent symptoms of toxicity (Pickering et al. 2003; Freeman et al. 2006b). A plant having non protein MeSeCys amino acid can tolerate Se (Montes-Bayón, LeDuc et al. 2002, Pedrero and Madrid 2009) and are Se hyperaccumulators (Terry et al. 2000; White et al. 2004). The third group, which is intermediate are called secondary Se accumulators. They can grow on soils with moderate Se levels and accumulate moderate quantities of Se (up to 1 g Se kg−1 DW) without showing signs of toxicity. Certain Alliaceae (e.g. garlic - Allium sativum) and Brassicaceae such as. Indian mustard (Brassica juncea), broccoli (B. oleracea var. italica) and canola (B. napus) are in this group (McKenzie et al. 2009).
Three parameters are used to identify Se-hyperaccumulators: plant Se:S ratio, shoot Se concentration compared to that in roots, and leaf Se content (Galeas et al. 2007; White et al. 2007). It has not been documented as Se hyperaccumulation being essential for the plant (Brown and Shrift 1981; Anderson 1993). However, hyperaccumulation is considered as beneficial for antioxidant properties, developing antiherbivore for arthropods and antipathogenic for fungi (Hanson et al. 2003; Hanson et al. 2004; Hartikainen 2005; Freeman et al. 2006a; Quinn et al. 2007; Quinn et al. 2008). They have the advantage of surviving in high Se soils. It is worthwhile understanding the fundamental mechanism of how plants accumulate such large amounts of Se; the process will be immensely helpful in developing Se-rich cereals and pulses as well as phytoremediation of high Se-contaminated soils (Trippe & Pilon-Smits, 2021). Se hyperaccumulating plants in the families of Asteraceae, Brassicaceae and Leguminosae (Reeves and Baker 2000) are important research materials for in-depth analysis of Se hyperaccumulation, Se fortification, and Se phytoremediation.
Prevention of Se Pollution
Se pollution is one of the growing concerns across the globe, particularly in mining and agriculture areas (Lemly, 2004). Improved water management practices, such as reducing excessive irrigation or applying Se-free water have reduced Se leaching from the soil (Dhillon and Dhillon, 2001). The application of organic amendments to Se-contaminated soil mitigates the adverse impacts of Se (Sharma et al., 2011). The incorporation of crop residue or animal manure into selenated soils leads to a significant reduction in Se accumulation by plants (Ajwa et al., 1998). Bioreactors can be developed to harness microbial activities as a biological method to remove excess Se (Staicu et al., 2017; Nkansah-Boadu et al., 2019). Selenate, a highly soluble and bioavailable form of Se, can be reduced to selenite and further to less toxic elemental Se using chemical or biological methods (Hageman et al., 2013; Zhang et al., 2020). Hence, it is essential to maintain a proper equilibrium between selenite and Se in order to prevent Se toxicity in plants.
Se Phytoremediation by Se Hyperaccumulators and Se tolerance
In phytoremediation, plant extraction (phytoextraction), filtration (hemofiltration or rhizofiltration) or volatilization (phytovolatilization) mechanisms are utilized (Dhillon and Bañuelos 2017; LeDuc and Terry 2005). For example, Indian mustard (Brassica júncea Czern L.) can be used for Se-contaminated soil phytoremediation. Subsequently, harvested mustard can be used as animal feed (Banuelos 2006; Banuelos and Lin 2009), biofuels (Searchinger et al. 2008), or bioavailable Se source (Bañuelos et al. 2015). Other plants that can be grown to reduce Se content substantially are giant reed (Arundo donax), creeping saltbush (Atriplex semibaccata R. Br. L.), old man saltbush (Atriplex nummularia Lindl. L.), tall fescue (Festuca arundinacea Schreb. L.), canola (Brassica napus L.), birdsfoot trefoil (Lotus comiculatus L.), and kenaf (Hibiscus cannabinus L.) (El-Ramady et al., 2015; Banuelos, 1996). Also, plants of the Astragulus genus and Neptunia amplexicaulis (Domin.) of the family Fabaceae, which hyperaccumulate Se as MeSeCys are efficient plants for phytoremediation of Se (Peterson and Butler 1967; Neuhierl and Böck 1996). Further investigating the genetic variability in the accumulation of Se among a wider range of plant species will open up further opportunities to remediate Se contamination through phytoremediation.
Se hyperaccumulation or Se tolerance in Astragalus bisulcatus is due to the selenocysteine methyltransferase gene expression (Ellis et al. 2004; McKenzie et al. 2009). Se uptake, hyper accumulation and emission collectively maintain Se tolerance in such plants (Pilon-Smits and LeDuc 2009).
Se Reduction by Microbes
Microbial Se transfer mechanism plays an important role in Se reduction in the environment (Huang et al., 2012; Etteieb et al., 2021; Li et al., 2022). Rhizospheric microbes accelerate Se uptake and volatilization (Pilon-Smits, 2005). Se hyperaccumulator plants colonize with a diverse community of endophytic bacteria that promote plant growth (Sura-de Jong et al., 2015; Li et al., 2022). Specific strains of soil bacteria have the ability to transform soluble forms of Se into insoluble, less toxic forms, reducing the bioavailability of Se. These microorganisms decrease the amount of Se contamination in the surface environment by converting it into a volatile form through methylation (Bañuelos et al. 2007). In general, bacteria play a more prominent role than fungi in Se volatilization, and aerobic conditions accelerate bacteria-mediated Se volatilization. For example, experiments using the antibiotic ampicillin in Indian mustard showed that bacteria facilitated 35% of plant Se volatilization and 70% of plant tissue accumulation of Se (de Souza et al. 1999). Bioaugmentation of such bacteria is important for lowering Se bioavailability in soils. These findings suggest that the Se transformation process and its rate determine Se bioavailability. Se volatilization changes from SeMet>selenite>selenate in soil (Azaizeh et al., 1997). However, there is very limited understanding of this process in different agricultural systems.
Utilizing Se Detoxification in Plants for Phytoremediation
Once Se is released from SeCys and SeMet, toxic conditions arise in plants (Brown and Shrift 1981; White et al., 2004). For their survival, plants have developed Se detoxification mechanisms. One of the main Se detoxification mechanisms is volatilization as gases. Dimethyl diselenide, diethyl selenide and dimethyl selenide are emitted by such plants (Dauchy et al. 1994). It has been suggested that plants with these traits can be utilized for Se phytoremediation and Se fortified foods (Pilon-Smits and LeDuc 2009). The enzyme cystathionine gamma synthase (cgs) that converts SeCys to SeMet in the volatilization pathway prevents Se toxicity by decreasing Se accumulation by 40% (Pilon-Smits and LeDuc 2009).
Se Biofortification
Elevating Se biofortification in crops can be achieved through a variety of effective strategies. These include soil or foliar application of Se, crop management practices, as well as advancements in plant breeding and genetic transformation and editing techniques.
Genetic Biofortification or Breeding for Biofortification
Genetic biofortification of Se implies the selection and use of genotypes that have an ability to accumulate higher Se contents. It purely depends on available genetic resources for breeding purposes (White and Broadley 2009). Varying Se content in the different genotypes is advantageous for genetic biofortification (Eurola et al. 1991; Lyons et al. 2005b). As an example, wild relatives of wheat such as Aegilops tauschii have higher Se contents than durum and bread wheat (Graham et al. 1999; Lyons et al. 2003; White & Broadley, 2005a; White et al. 2007). Hence, interbreeding elite crop cultivars with Se-rich wild-relatives or landraces is one of the strategies to produce Se-enriched staple crops. Selection of genotypes that can produce MeSeCys using inorganic Se is the basis for biofortification (Roberge et al. 2003; Ramos et al. 2011). However, Se must be available in the soil for screening the grains (or other edible parts) for Se in breeding programs (Cakmak 2008). So, Se variation across diverse environmental conditions is a barrier to breeding for biofortification (Lyons et al. 2005a).
Gene Transformation for Se Biofortification
Development of food crops intended to increase dietary Se intake is Se biofortification. This can be achieved through traditional breeding but more efficiently using gene transformation technologies. Considering its anticancer properties, it would be much more beneficial if a proportion of this supplementary Se were present as MeSeCys (Bhattacharya, 2011; Poluboyarinov et al., 2020) or other organic forms (Poluboyarinov et al., 2020). Therefore, most of the plant transformation work in this area has been directed at increasing the organic Se levels. Sulfur and Se metabolism related genes were overexpressed in Brassica juncea, resulting in increased Se accumulation and Se tolerance (Lyons, et al. 2005a; Banuelos and Meek 1990). Adenosine triphosphate sulfurylase (APS), SeCys methyltransferase (SMT), and cystathionine gamma synthase (CgS) genes are used for gene transformation for Se biofortification (Neuhierl and Böck 1996; Van Huysen et al. 2003; Banuelos et al. 2005). APS is an enzyme that catalyzes sulfate and selenate (Pilon-Smits et al. 1999) for Se assimilation (Banuelos et al. 2005). Overexpressing APS in Indian mustard has been successful for Se biofortification (Pilon-Smits, et al.1999. SMT gene from the Se hyperaccumulator plant, Astragalus bisulcatus, has often been used for inducing Se tolerance (Ellis et al. 2004; LeDuc et al. 2004; McKenzie et al 2009). Cystathionine gamma-synthase (CgS) converts SeCys to SeMet, and Se volatilization is increased by 2 to 3 folds in transgenic A. thaliana by overexpressing CgS (Van Huysen et al. 2003). Gene pyramiding of APS and SMT accelerates selenate accumulation (LeDuc et al. 2006). When a cDNA encoding the SMT from Se hyperaccumulator A. bisulcatus was overexpressed in tomato (Solanum lycopersicum), transgenic plants’ accumulation from soil-supplied selenate was found to increase the assimilation of Se into leaves and fruit. Relative to wild-type, Se accumulation from soil-supplied selenate was increased by ∼1.53-fold in both leaves and fruit but MeSeCys accumulated in the fruit but not in the leaves. Depending on the transgenic line and Se treatment, up to 16% of the total Se in the fruit was present as MeSeCys. MeSeCys was produced more effectively from selenite on a percentage conversion basis, but greater accumulation of MeSeCys could be achieved from selenate due to its better translocation from the roots (Brummell et al. 2011). Quantitative Trait Loci for Se tolerance have been identified (Agalou et al. 2005; Zhang et al. 2006) for potential dissecting of major genes. However, strict regulations for genetically modified crops and intellectual property rights problems are a hindrance to their commercialization (Cohen and Paarlberg 2002; Potrykus 2009; Stein 2009).
Agronomic Se Biofortification
Agronomic Se biofortification is adding Se to the soil through the application of Se-enriched fertilizers or soil amendments to increase Se in the edible parts of food crops. Micronutrient-enriched fertilizer application is an initial step for micronutrient fortification (Lyons et al. 2005a; Hawkesford and Zhao, 2007; White and Broadley 2009, Mao et al. 2014). Application of inorganic fertilizers in Se poor soils enhances Se absorption more than 100 times for many crops (Liu et al. 2010). Developing crop varieties that are more efficient at absorbing and accumulating Se from the soil is a long-term strategy. Se can be added to current fertilizer recommendations as an additional ingredient (Cakmak 2008; 2009). Finland achieved enough Se in food for its population after introducing such fertilization programmes since the mid-1980s, the first country to do so (Zhao and Mcgrath 2009, Alfthan et al., 2015). New Zealand also achieved Se-enriched crops through Se-enriched inorganic fertilizers (Hartikainen 2005; Premarathna et al. 2012). Adding Se with Zn and/or I gave a similar effect as added Se alone (Mao et al. 2014). Most of the time soils that contain Se as selenite exhibit less bioavailability than the presence of Se as selenate (Li, et al. 2008; Mao et al. 2014). Se is applied at the rate of 10 to 20 g Se ha-1as selenate, but for receiving the same Se increment in grains, selenite should be applied at over 100 g Se ha -1(Yläranta 1985). A 35.9% increment of grain Se in barley by applying selenate was reported in comparison to selenite application (Chen et al. 2002). However, the same effect of grain Se accumulation was reported in soil-applied selenate and foliar-applied selenite in many crops (Mao et al. 2014). The conversion ratio of Se to MeSeCys in broccoli is greater when applied as selenite compared to selenate (Avila 2013), but it was controversial in the case of rice. The difference would be anaerobic soil condition in rice (Li et al. 2010). Adding Se into the soil is more efficient in Se increment in grains than using it as a foliar application, and selenate increases grain Se while selenite increases root Se in rice (Boldrin et al. 2013).
Plants of the family Brassicaceae have the capacity to produce S-rich glucosinolates known to have significant human health benefits. Selenate application to three Brassicaceae crops; broccoli (Brassica oleracea L. var. italica), cauliflower (Brassica oleracea L. var. botrytis) and forage rape (Brassica napus) and analysing glucosinolates produced in four tissues demonstrated that the distribution of Se-containing glucosinolates is different among the three crops, and broccoli florets and forage rape roots accumulated more of these (McKenzie et al. 2015). Understanding these differences in Se accumulation is important in agronomic biofortification programmes using different crops. Increasing bioavailability of the element at the rhizosphere also facilitates greater uptake of Se (Meenakshi, 2010). Adding metal chelates increases Se availability (Zhao and McGrath, 2009). Composting and adding organic materials to the soil improve the bioavailability of Se in the soil, making it more accessible to crops (Wang et al. 2018). Organic acids act as chelates to increase Se uptake efficiency in fertilizers (Morgan et al. 2005, Lynch, 2007) and organic acids improve root volume (Kirkby and Johnston 2008). Using Se-enriched water sources can also contribute to increased Se uptake by crops (Zhou et al. 2018). Further, rotating Se-accumulating crops with other crops in the same field can help maintain soil Se levels and reduce the risk of soil depletion (Bañuelos and Dhillon 2011).
Factors Affecting Biofortification
Biofortification has been recognized as a promising approach for improving the nutritional quality of edible crop species (Yang et al. 2007). Several factors can significantly impact the success of biofortification initiatives. Among them the choice of crops is very important as many crop species do not have the same ability to effectively accumulate and enhance the specific nutrients in the edible organs (White and Broadley 2005). In addressing Se deficiency, lentils have been identified as a potential candidate for Sri Lanka (Thavarajah, et al., 2011).
The most important factor is the nutrient availability in the soil, which determines the crop's ability to take up those nutrients. Further the soil composition and the soil pH and nutrient interactions can also influence the success of biofortification targets (White and Broadley, 2005). Environmental factors like climate, temperature, and precipitation can influence nutrient uptake and crop growth. Different geographic regions may require specific biofortification strategies tailored to their environmental conditions as well as cultivated crops. Furthermore, the acceptance and willingness of people to consume nutrient rich food is crucial for the success of biofortification effort (Meenakshi et al. 2010). Special precautions are also needed in biofortification programmes. For example, micronutrients can be contaminated with heavy metals, which could pose a major challenge for biofortification. This is mainly due to some of the transporters common to other nutrients' uptake could easily facilitate heavy metal uptake. Thus, biofortification with Se could also facilitate the increase of heavy metals in food, such as cadmium (Zhao and McGrath, 2009). Se can be found excessively in polluted soils and phytoremediation and biofortification cannot be expected simultaneously (Vamerali et al. 2014). Because plants cannot be grown in Se remediated substrates though such substrates are rich with Se and other heavy metals and micronutrients. Heavy metals use the same transporters of Se (Zhao and McGrath, 2009). Further, though the seeds are fortified with Se, Se concentration is higher in the husk and aleurone layer of seed and most of the time aleurone layer is discarded during processing. Milling and processing must be minimized for higher retention of Se in the diet (White and Broadley 2009; Qaim et al. 2007).
Assessment of Biofortification
Assessing biofortification is a comprehensive process that requires interdisciplinary collaboration. Local agricultural and health authorities, as well as international organizations like the World Health Organization (WHO) and the Food and Agriculture Organization (FAO) should play important roles in guiding and monitoring biofortification programs to prevent both deficiency and excess Se intake (WHO, 2006).
Usually, enhancement of crop yield quality doesn’t ensure an economic gain in agriculture (Unnevehr, 1986). Quality improvements of field crops are rare, and one example is golden rice production for Vitamin A enrichment in rice (Dawe et al. 2002). DALYS methodology has explicated the economic benefit of golden rice (Zimmermann and Qaim 2004) and found that people were still below the micronutrient threshold after introduction of Golden rice (Fiedler et al. 2000). Success of biofortification programmes is assessed using following steps: Assess biofortified crop nutrient content and ensure their productivity and marketability, evaluate nutrient concentrations in plant tissues, test nutrient retention during food processing, study consumer acceptance, and measure nutritional impact. It is also important to monitor health outcomes, environmental effects, economic feasibility, market demand, and programme progress. Strategies have to be adapted as needed, regulatory compliance, and long-term sustainability have to be ensured. Engaging communities and collaboration among stakeholders for shared knowledge and best practices are an integral part of biofortification programmes.
When there is no consumer acceptance of biofortified food, micronutrients are neglected in the diet (Qaim et al. 2007). Most of the crop improvement programmes targeted at increasing yield have resulted in the decrease of the concentration of micronutrients in the grain. Sometimes inhibitory effects of soil reduce the Se intake of the plant For example, slight doses of sulfur increase Se uptake and extra doses of sulfur inhibit Se uptake (Lyi et al. 2005). This happens when selenate is applied (Avila et al. 2013). SeO and SO42- analogues also reduce Se accumulation in plants (Pilon-Smits and Quinn, 2010). Biofortification is not important for farmers unless crops have farmer-preferred agronomic characteristics, including high yield (Stein 2009). Widespread acceptance for Se biofortification must be earned harder (Nestel et al. 2006; Qaim and Stein 2009; Qaim et al. 2007) and community approaches must be carried out for such an effort (Stein 2009).
Economic Benefits of Se Biofortification
The economic benefits of Se biofortification have been supported by several studies around the world. Horton and Ross (2003) demonstrated that poor health in the developing world associated with micronutrient deficiency has a major impact on the economies of countries. It has been demonstrated that there is a strong relationship between dietary energy used in the population and national economic growth (Wang and Taniguchi 2003). For instance, around 30% of British per capita increment was gained due to the overall nutrition assurance of the nation (Fogel 2004). According to Horton and Ross (2003), a 4% loss of gross domestic production in 10 developing countries is due to national nutrition problems. Globally, one billion people are suffering from micronutrient malnutrition (Williams et al. 2009). In Sri Lanka, it has been documented that 24-40% of the female population in the tested regions with reported goitre is Se deficient (Fordyce et al. 2000). Se deficiencies play a role in goitre pathogenesis (Fordyce et al. 2000). Se insufficiency levels ranging from 5% to 50% have been documented in various districts across Sri Lanka (Diyabalanage et al., 2020). Nutrient insufficiency has a direct impact on the economy by increasing healthcare costs and reducing workforce productivity, as it leads to a higher prevalence of chronic illnesses. Additionally, it can strain social safety nets and public resources, diverting funds away from other critical economic development initiatives. Hence, the economic benefits of Se fortification are evident in terms of improved health outcomes, reduced healthcare costs, increased productivity, and efficient allocation of public resources. However, the specific economic impact of Se fortification may vary based on the context and the population characteristics, emphasizing the need for further research and development.
Knowledge Gap
While Se biofortification holds significant promise as a solution to combat global Se malnutrition, there are several areas within this field that require further research and investigation to refine and optimize strategies. Understanding the factors that affect the bioavailability of Se in fortified crops is important. Research should focus on improving the efficiency of Se absorption and incorporation into edible plant parts. This also involves investigating the interaction between soil, plant, and human physiology to maximize the health benefits of Se fortification. Further, research should explore the most effective forms and sources of Se for fortification. Different forms of Se, such as selenate and selenite, have varying bioavailability. Additionally, alternative Se sources, like organic Se compounds, should be investigated to determine their suitability for fortification.
Different crops accumulate Se in different forms and quantities in different tissues. Identifying and breeding crop varieties that efficiently absorb Se from the soil and accumulate in edible parts is essential. Another important aspect needs attention is developing and promoting improved Se accumulating crop varieties that can thrive in different geographic regions, considering local soil and climate conditions. Developing and optimizing agronomic practices for such varieties (varietal agronomy) is also important for Se biofortification. This includes determining the appropriate timing and method of Se application, as well as studying the effects of other soil amendments, such as pH adjustments and organic matter content on Se uptake by plants.
The threshold levels for Se requirement and toxicity are very narrow. Research should provide guidelines for safe and effective Se fortification levels in staple crops. Over-fortification can lead to Se toxicity, which can have adverse health effects. It is important to establish the maximum allowable Se levels to prevent unintended health risks. Se is not a well-known element in human health. Hence, investigating consumer attitudes and acceptance of Se-fortified crops is important. Research should address concerns related to biofortified products among different cultural and demographic groups not to waste investments on Se biofortification efforts. Se has not been added to any food item so far. Long-term epidemiological studies are necessary to assess the health outcomes over extended periods including the side-effects of Se in populations that consume Se fortified crops regularly. These studies should explore non communicable diseases over extended periods since the side-effect of Se over consumption is unknown. The socioeconomic impact of Se fortification needs to be studied to ensure that it does not inadvertently lead to social inequities or disrupt traditional farming practices. Intellectual property rights and equitable access to fortified crops should also be examined and documented in advance. The region-specific fortification strategies must be formulated to utilize them for diverse soil Se contents, soil conditions, crops and environments. Se can be toxic to some plants and aquatic life, so research should evaluate the ecological consequences of increased Se in agricultural runoff and its potential effects on ecosystems. Hence, developing regulatory frameworks for Se fortification is another essential socio-economic aspect. By addressing these research areas, scientists and policymakers can better tailor Se fortification strategies to diverse geographic regions and populations, ensuring the safe and effective delivery of this essential micronutrient to those who need it most.
Conclusions
Se is a trace element that is essential for human health, yet it often goes unnoticed in discussions about nutrition and global health. It plays an essential role in various physiological processes in the human body. Se is an essential component of selenoproteins, which act as antioxidants and have been linked to various health benefits. Inadequate Se intake in the diet (less than 15-30 µg/day) can lead to weakened immune system and is considered as an etiological factor in some cardiovascular, cancer, neurodegenerative and osteoarticular diseases in humans. In severe cases, Se deficiency can result in a condition known as Keshan disease, which affects the heart and can be fatal.
The prevalence of Se deficiency is not limited to one region or population; it is a global concern. Many areas of the world, including parts of China, Eastern Europe, UK, Canada, New Zealand, Russian Federation and parts of Africa, are known to have Se-deficient soils. As a result, crops grown in these regions are often low in Se, and the deficiency is subsequently passed on to the people who depend on these crops as their primary source of sustenance.
One of the most promising and sustainable solutions to combat global Se malnutrition is Se fortification of staple crops, particularly rice. Rice is a staple food for more than half of the world's population and fortifying it with Se has the potential to reach large populations, making it a practical and effective strategy. Se biofortification, the process of enhancing the Se content of crops through agronomic practices, has been successful in increasing Se levels in rice and other staple foods.
Comprehensive Se fortification strategies involve not only increasing Se content in crops but also ensuring that the bioavailability of Se is optimized. It's not just about adding Se to the soil but also understanding how plants absorb and incorporate it into their edible parts. This approach requires a combination of soil management techniques, plant breeding, and crop selection to achieve the desired Se levels. For instance, selecting rice varieties that efficiently accumulate Se from the soil and those that can partition more Se into endosperm can make significant improvement in diets. The potential impact of Se fortification in rice crops on public health is substantial. By increasing the Se content of a widely consumed staple food, we can address the issue of Se deficiency at its source. This has several significant benefits. Se fortification can lead to improved overall health and reduced risk of diseases associated with Se deficiency. Enhanced immune function, better cardiovascular health, and reduced risk of cancer are some of the potential benefits. Fortifying rice and other staple crops is a cost-effective and sustainable approach to combat Se deficiency. It leverages existing agricultural systems and distribution channels, making it accessible to a large portion of the population. By increasing the nutritional content of staple crops, Se fortification contributes to food security, ensuring that communities have access to a more nutritious and balanced diet. As public health improves, healthcare costs associated with Se deficiency-related diseases can decrease. This can alleviate the burden on healthcare systems and improve the overall well-being of communities. Further, healthier individuals are more productive and have better educational outcomes. 15-30 µg/day Thus, Se fortification can contribute to economic growth by creating a healthier and more capable workforce. Hence, Se fortification, particularly in rice crops, is a sustainable solution to combat global Se malnutrition.
References
- Abdulah, R., K. Miyazaki, M. Nakazawa and H. Koyama (2005) Chemical forms of Se for cancer prevention. Journal of Trace Elements in Medicine and biology 19(2): 141-150.
- Acuña, J. J., M. A. Jorquera, P. J. Barra, D. E. Crowley and M. de la Luz Mora (2013) Selenobacteria selected from the rhizosphere as a potential tool for Se biofortification of wheat crops. Biology and fertility of soils 49(2): 175-185.
- Agalou, A., A. Roussis and H. P. Spaink (2005) The Arabidopsis Se-binding protein confers tolerance to toxic levels of Se. Functional plant biology 32(10): 881-890.
- Ajwa, H.A., Bañuelos, G.S. and Mayland, H.F., (1998) Se uptake by plants from soils amended with inorganic and organic materials. American Society of Agronomy, Crop Science Society of America, and Soil Science Society of America. 27 (5):1218-1227.
- Alfthan, G.; Eurola, M.; Ekholm, P.; Venäläinen, E.R.; Root, T.; Korkalainen, K.; Hartikainen, H.; Salminene, P.; Hietaniemi, V.; Aspila, P.; et al. (2015) Effects of nationwide addition of selenium to fertilizers on foods, and animal and human health in Finland: From deficiency to optimal selenium status of the population. Journal of Trace Elements in Medicine and Biology 31:142–147.
- Allander, E. (1994) Kashin-Beck disease. An analysis of research and public health activities based on a bibliography 1849-1992. Scandinavian Journal of Rheumatology 23(99): 1-36.
- Amarya, S., K. Singh and M. Sabharwal (2015) Changes during aging and their association with malnutrition. Journal of Clinical Gerontology and Geriatrics 6(3): 78-84.
- Anderson, J. W. (1993) Se interactions in sulfur metabolism. Sulfur nutrition and assimilation in higher plants: regulatory, agricultural and environmental aspects. SPB Academic Publishing, The Hague, The Netherlands: 49-60.
- Arvy, M. (1993) Selenate and selenite uptake and translocation in bean plants (Phaseolus vulgaris). Journal of Experimental Botany 44(6): 1083-1087.
- Avila, F. W., V. Faquin, Y. Yang, S. J. Ramos, L. R. Guilherme, T. W. Thannhauser and L. Li (2013) Assessment of the anticancer compounds Se-methylselenocysteine and glucosinolates in Se-biofortified broccoli (Brassica oleracea L. var. italica) sprouts and florets. Journal Agric Food Chem 61(26): 6216-6223.
- Azaizeh, H.A., Gowthaman, S. and Terry, N., (1997) Microbial Se volatilization in rhizosphere and bulk soils from a constructed wetland. Journal of Environmental Quality. 26(3): 666-672.
- Banuelos, G. (2006) Phyto-products may be essential for sustainability and implementation of phytoremediation. Environmental Pollution 144(1): 19-23.
- Azaizeh, H.A., Salhani, N., Sebesvari, Z. and Emons, H., (2003). The potential of rhizosphere microbes isolated from a constructed wetland to biomethylate Se. Journal of Environmental Quality, 32(1): 55-62.
- Banuelos, G. and D. Meek (1990) Accumulation of Se in plants grown on Se-treated soil. Journal of Environmental Quality 19(4): 772-777.
- Banuelos, G. and Z. Lin (2009). Development and Uses of Biofortified Agricultural Products, Boca Raton, FL: CRC Press.
- Bañuelos, G., Terry, N., LeDuc, D.L., Pilon-Smits, E.A. and Mackey, B., (2005) Field trial of transgenic Indian mustard plants shows enhanced phytoremediation of Se-contaminated sediment. Environmental Science & Technology, 39(6), 1771-1777.
- Barberon, M., P. Berthomieu, M. Clairotte, N. Shibagaki, J. C. Davidian and F. Gosti (2008) Unequal functional redundancy between the two Arabidopsis thaliana high-affinity sulphate transporters SULTR1; 1 and SULTR1; 2. New Phytologist 180(3): 608-619.
- Barrow, N. and B. Whelan (1989) Testing a mechanistic model. VII. The effects of pH and of electrolyte on the reaction of selenite and selenate with a soil. Journal of Soil Science 40(1): 17-28.
- Bawa, S., K. Dhillon and S. Dhillon (1992) Screening of different fodders for Se absorption capacity. Indian Journal of Dairy Science 45: 457-457.
- Beilstein, M., P. Whanger and G. Yang (1991) Chemical forms of Se in corn and rice grown in a high Se area of China. Biomedical and environmental sciences: BES 4(4): 392-398.
- Bendich, A. (2001) Dietary reference intakes for vitamin C, vitamin E, Se, and carotenoids institute of medicine washington, DC: National Academy Press, 2000 ISBN: 0-309-06935-1. Nutrition 17(4): 364.
- Bhattacharya, A. (2011). Methylselenocysteine – a promising antiangiogenic agent for overcoming drug delivery barriers in solid malignancies for therapeutic synergy with anticancer drugs. Expert Opinion on Drug Delivery, 8(6), 749–763. [CrossRef]
- Blaylock, M. J., D. E. Salt, S. Dushenkov, O. Zakharova, C. Gussman, Y. Kapulnik, B. D. Ensley and I. Raskin (1997) Enhanced accumulation of Pb in Indian mustard by soil-applied chelating agents. Environmental Science & Technology 31(3): 860-865.
- Boldrin, P. F., V. Faquin, S. J. Ramos, K. V. F. Boldrin, F. W. Ávila and L. R. G. Guilherme (2013) Soil and foliar application of Se in rice biofortification. Journal of Food Composition and Analysis 31(2): 238-244.
- Brinch-Pedersen, H., S. Borg, B. Tauris and P. B. Holm (2007) Molecular genetic approaches to increasing mineral availability and vitamin content of cereals. Journal of Cereal Science 46(3): 308-326.
- Brown, T. A. and A. Shrift (1981) Exclusion of Se from proteins of Se-tolerant Astragalus species. Plant Physiology 67(5): 1051-1053.
- Bujdoš, M., A. Muľová, J. Kubova and J. Medveď (2005) Se fractionation and speciation in rocks, soils, waters and plants in polluted surface mine environment. Environmental geology 47(3): 353-360.
- Caballero, B. (2002) "Global patterns of child health: the role of nutrition. Annals of Nutrition and Metabolism 46(Suppl. 1): 3-7.
- Cakmak, I. (2008) Enrichment of cereal grains with zinc: agronomic or genetic biofortification? Plant and soil 302(1-2): 1-17.
- Cakmak, I. (2009) Agronomic approaches in biofortification of food crops with micronutrients. The Proceedings of the International Plant Nutrition Colloquium XVI. UC Davis: Department of Plant Sciences. Retrieved from https://escholarship.org/uc/item/3d46b1th.
- Carey, A. M., K. G. Scheckel, E. Lombi, M. Newville, Y. Choi, G. J. Norton, A. H. Price and A. A. Meharg (2012) Grain accumulation of Se species in rice (Oryza sativa L.). Environ Sci Technol 46(10): 5557-5564.
- Chaney, R. L., J. S. Angle, C. L. Broadhurst, C. A. Peters, R. V. Tappero and D. L. Sparks (2007) Improved understanding of hyperaccumulation yields commercial phytoextraction and phytomining technologies. Journal of Environmental Quality 36(5): 1429-1443.
- Chen, L., Yang, F., Xu, J., Hu, Y., Hu, Q., Zhang, Y. and Pan, G., 2002. Determination of Se concentration of rice in China and effect of fertilization of selenite and selenate on Se content of rice. Journal of Agricultural and Food Chemistry, 50(18): 5128-5130.
- Chen, Y.-W., H.-Y. T. Truong and N. Belzile (2009) Abiotic formation of elemental Se and role of iron oxide surfaces. Chemosphere 74(8): 1079-1084.
- Clark, L. C., G. F. Combs, B. W. Turnbull, E. H. Slate, D. K. Chalker, J. Chow, L. S. Davis, R. A. Glover, G. F. Graham and E. G. Gross (1996) Effects of Se supplementation for cancer prevention in patients with carcinoma of the skin: a randomized controlled trial. Jama 276(24): 1957-1963.
- Cohen, J. I. and R. Paarlberg (2002) Explaining restricted approval and availability of GM crops in developing countries. AgBiotechNet 4(097): 1-6.
- Combs, G. F. (2001) Se in global food systems. British Journal of Nutrition 85(05): 517-547.
- Combs, G. F. and W. P. Gray (1998) Chemopreventive agents: Se. Pharmacology & therapeutics 79(3): 179-192.
- Dauchy, X., M. Potin-Gautier, A. Astruc and M. Astruc (1994) Analytical methods for the speciation of Se compounds: a review. Fresenius' journal of analytical chemistry 348(12): 792-805.
- Brummell, D.A., Watson, L.M., Pathirana, R., Joyce, N.I., West, P.J., Hunter, D.A., McKenzie, M.J. (2011) Biofortification of Tomato (Solanum lycopersicum) Fruit with the Anticancer Compound Methylselenocysteine Using a Selenocysteine Methyltransferase from a Selenium Hyperaccumulator, Journal of Agricultural and Food Chemistry 2011 59 (20):10987-10994. [CrossRef]
- Dawe, D., R. Robertson and L. Unnevehr (2002) Golden rice: what role could it play in alleviation of vitamin A deficiency? Food Policy 27(5): 541-560.
- De Kok, L. J. (1993) Sulfur nutrition and assimilation in higher plants. Workshop on Sulfur Metabolism in Higher Plants 1992: Garmisch-Partenkirchen, Germany), SPB Academic.
- de Souza, M. P., E. A. Pilon-Smits, C. M. Lytle, S. Hwang, J. Tai, T. S. Honma, L. Yeh and N. Terry (1998) Rate-limiting steps in Se assimilation and volatilization by Indian mustard. Plant Physiology 117(4): 1487-1494.
- Dhillon, K.S. and Dhillon, S.K., (2001) Restoration of Se-contaminated soils. In Environmental Restoration of Metals-Contaminated Soils. Lewis Publishers Boca Raton, FL.
- Dhillon, K. S. and S. K. Dhillon (2009) Se concentrations of common weeds and agricultural crops grown in the seleniferous soils of northwestern India. Science of the total environment 407(24): 6150-6156.
- Dhillon, K.S. and Bañuelos, G.S., (2017) Overview and prospects of Se phytoremediation approaches. Se in Plants: Molecular, Physiological, Ecological and Evolutionary Aspects, pp.277-321.
- Diyabalanage, S., Dangolla, A., Mallawa, C., Rajapakse, S. and Chandrajith, R., (2020) Bioavailability of Se (Se) in cattle population in Sri Lanka based on qualitative determination of glutathione peroxidase (GSH-Px) activities. Environmental geochemistry and health, 42: 617-624.
- Djanaguiraman, M., D. D. Devi, A. K. Shanker, J. A. Sheeba and U. Bangarusamy (2005) Se–an antioxidative protectant in soybean during senescence. Plant and Soil 272(1-2): 77-86.
- Dörr, A. J. M., N. Pacini, M. C. Abete, M. Prearo and A. C. Elia (2008) Effects of a Se-enriched diet on antioxidant response in adult crayfish (Procambarus clarkii). Chemosphere 73(7): 1090-1095.
- Ellis, D. R. and D. E. Salt (2003) Plants, Se and human health. Current Opinion in Plant Biology 6(3): 273-279.
- Ellis, D. R., T. G. Sors, D. G. Brunk, C. Albrecht, C. Orser, B. Lahner, K. V. Wood, H. H. Harris, I. J. Pickering and D. E. Salt (2004) Production of Se-methylselenocysteine in transgenic plants expressing selenocysteine methyltransferase. BMC Plant Biology 4(1): 1.
- El-Ramady, H.R., Abdalla, N., Alshaal, T., Elhenawy, A.S., Shams, M.S., Faizy, S.E.D.A., Belal, E.S.B., Shehata, S.A., Ragab, M.I., Amer, M.M. and Fári, M., (2015) Giant reed for Se phytoremediation under changing climate. Environmental chemistry letters, 13: 359-380.
- Elrashidi, M., D. Adriano, S. Workman and W. Lindsay (1987) Chemical equilibria of Se in soils: a theoretical development1. Soil Science 144(2): 141-152.
- Etteieb, S., Zolfaghari, M., Magdouli, S., Brar, K.K. and Brar, S.K., (2021) Performance of constructed wetland for Se, nutrient and heavy metals removal from mine effluents. Chemosphere, 281, p.130921.
- Eswayah, A.S., Smith, T.J. and Gardiner, P.H., (2016) Microbial transformations of Se species of relevance to bioremediation. Applied and environmental microbiology, 82(16): 4848-4859.
- Eurola, M. H., P. I. Ekholm, M. E. Ylinen, P. T. Varo and P. E. Koivistoinen (1991) Se in Finnish foods after beginning the use of selenate-supplemented fertilisers. Journal of the Science of Food and Agriculture 56(1): 57-70.
- Eurola, M., Ekholm, P.A.I.V.I., Ylinen, M.A.I.J.A., Koivistoinen, P.E.K.K.A. and Varo, P.E.R.T.T.I., (1990) Effects of Se fertilization on the Se content of cereal grains, flour, and bread produced in Finland. Cereal Chem, 67(4): 334-337.
- Fan, M.-S., F.-J. Zhao, S. J. Fairweather-Tait, P. R. Poulton, S. J. Dunham and S. P. McGrath (2008a) Evidence of decreasing mineral density in wheat grain over the last 160 years. Journal of Trace Elements in Medicine and Biology 22(4): 315-324.
- Fang, Y., L. Wang, Z. Xin, L. Zhao, X. An and Q. Hu (2008) Effect of foliar application of zinc, Se, and iron fertilizers on nutrients concentration and yield of rice grain in China. Journal of Agricultural and Food Chemistry 56(6): 2079-2084.
- FAO (2005). The State of Food Insecurity in the World (2005) Eradicating World Hunger-Key to Achieving the Millennium Development Goals, Food & Agriculture Org., Rome, Italy. Available at: https://www.fao.org/4/a0200e/a0200e00.htm.
- Feist, L. J. and D. R. Parker (2001) Ecotypic variation in Se accumulation among populations of Stanleya pinnata. New Phytologist 149(1): 61-69.
- Felton, S., M. Landolt, R. Grace and A. Palmisano (1996). Effects of Se dietary enhancement on hatchery-reared coho salmon, Oncorhynchus kisutch (Walbaum), when compared with wild coho: hepatic enzymes and seawater adaptation evaluated. Aquaculture Research 27(2): 135-142.
- Fendorf, S., M. Herbel, K. Tufano and B. Kocar (2008) Biogeochemical processes controlling the cycling of arsenic in soils and sediments, Biophysico-chemical Processes of Heavy Metals and Metalloids in Soil Environments, 313-338.Wiley: Hoboken, NJ.
- Fernandes, K., R. Berton and A. Coscione (2014) Se biofortification of rice and radish: effect of soil texture and efficiency of two extractants. Plant soil and environment 60(3): 105.
- Fiedler, J. L., D. R. Dado, H. Maglalang, N. Juban, M. Capistrano and M. V. Magpantay (2000) Cost analysis as a vitamin A program design and evaluation tool: a case study of the Philippines. Social Science & Medicine 51(2): 223-242.
- Finley, J. W. (2005). Proposed criteria for assessing the efficacy of cancer reduction by plant foods enriched in carotenoids, glucosinolates, polyphenols and selenocompounds. Annals of Botany 95(7): 1075-1096.
- Fogel, R. W. (2004) The escape from hunger and premature death, 1700-2100: Europe, America, and the Third World, Cambridge University Press.
- Fordyce, F. (2005) Se deficiency and toxicity in the environment. In: Essentials of medical geology–Impacts of the natural environment on public health (Eds O Selinus, B Alloway, JA Centeno, RB Finkelman, R Fuge, U Lindh, P Smedley) pp. 373-415, Elsevier-Academic Press, London.
- Fordyce, F.M., Johnson, C.C., Navaratna, U.R., Appleton, J.D. and Dissanayake, C.B., (2000) Se and iodine in soil, rice and drinking water in relation to endemic goitre in Sri Lanka. Science of the Total Environment, 263(1-3): 127-141.
- Frankenberger Jr., W.T., & Benson, S. (1994). Se in the Environment (1st ed.). CRC Press. Pp 480. Available at: . [CrossRef]
- Frankenberger Jr, W. T., & Arshad, M. (2001). Bioremediation of selenium-contaminated sediments and water. Biofactors, 14(1-4): 241-254.
- Freeman, J. L., C. F. Quinn, M. A. Marcus, S. Fakra and E. A. Pilon-Smits (2006a) Se-tolerant diamondback moth disarms hyperaccumulator plant defense. Current Biology 16(22): 2181-2192.
- Freeman, J. L., L. H. Zhang, M. A. Marcus, S. Fakra, S. P. McGrath and E. A. Pilon-Smits (2006b) Spatial imaging, speciation, and quantification of Se in the hyperaccumulator plants Astragalus bisulcatus and Stanleya pinnata. Plant physiology 142(1): 124-134.
- Fu, L.-H., X.-F. Wang, Y. Eyal, Y.-M. She, L. J. Donald, K. G. Standing and G. Ben-Hayyim (2002) A selenoprotein in the plant kingdom mass spectrometry confirms that an opal codon (UGA) encodes selenocysteine in Chlamydomonas reinhardtii glutathione peroxidase. Journal of Biological Chemistry 277(29): 25983-25991.
- Gailer, J., S. Madden, M. F. Burke, M. B. Denton and H. V. Aposhian (2000) Simultaneous multielement-specific detection of a novel glutathione–arsenic–Se ion [(GS) 2AsSe]− by ICP AES after micellar size-exclusion chromatography. Applied organometallic chemistry 14(7): 355-363.
- Galeas, M. L., L. H. Zhang, J. L. Freeman, M. Wegner and E. A. Pilon-Smits (2007) Seasonal fluctuations of Se and sulfur accumulation in Se hyperaccumulators and related nonaccumulators. New Phytologist 173(3): 517-525.
- Ganther, H. (1986) Pathways of Se metabolism including respiratory excretory products. International Journal of Toxicology 5(1): 1-5.
- Garvin, D. F., R. M. Welch and J. W. Finley (2006) "Historical shifts in the seed mineral micronutrient concentration of US hard red winter wheat germplasm. Journal of the Science of Food and Agriculture 86(13): 2213-2220.
- Gatlin 3rd, D. and R. P. Wilson (1984) Dietary Se requirement of fingerling channel catfish. The Journal of nutrition 114(3): 627-633.
- Giray, B. and F. Hincal (2004) Se status in Turkey. Journal of radioanalytical and nuclear chemistry 259(3): 447-451.
- Goldhaber, S. B. (2003) Trace element risk assessment: essentiality vs. toxicity. Regulatory Toxicology and Pharmacology 38(2): 232-242.
- Gómez-Ariza, J.L., Pozas, J.A., Giraldez, I. and Morales, E., (1998) Speciation of volatile forms of Se and inorganic Se in sediments by gas chromatography–mass spectrometry. Journal of Chromatography A, 823(1-2): 259-277.
- Graham, R. D., R. M. Welch, D. A. Saunders, I. Ortiz-Monasterio, H. E. Bouis, M. Bonierbale, S. De Haan, G. Burgos, G. Thiele and R. Liria (2007) Nutritious subsistence food systems. Advances in Agronomy 92: 1-74.
- Graham, R., D. Senadhira, S. Beebe, C. Iglesias and I. Monasterio (1999). Breeding for micronutrient density in edible portions of staple food crops: conventional approaches. Field Crops Research 60(1): 57-80.
- Grant, T. D., M. Montes-Bayón, D. LeDuc, M. W. Fricke, N. Terry and J. A. Caruso (2004) Identification and characterization of Se-methyl selenomethionine in Brassica juncea roots. Journal of Chromatography A 1026(1): 159-166.
- Grčman, H., D. Vodnik, Š. Velikonja-Bolta and D. Leštan (2003) Ethylenediaminedissuccinate as a new chelate for environmentally safe enhanced lead phytoextraction. Journal of Environmental Quality 32(2): 500-506.
- Gregorio, G. B., D. Senadhira, H. Htut and R. D. Graham (2000) Breeding for trace mineral density in rice. Food and Nutrition Bulletin 21(4): 382-386.
- Grybos, M., M. Davranche, G. Gruau and P. Petitjean (2007) Is trace metal release in wetland soils controlled by organic matter mobility or Fe-oxyhydroxides reduction? Journal of colloid and interface science 314(2): 490-501.
- Guerinot, M. L. and D. E. Salt (2001) Fortified foods and phytoremediation. Two sides of the same coin. Plant Physiology 125(1): 164-167.
- Gupta, M. and Gupta, S., (2017) An overview of selenium uptake, metabolism, and toxicity in plants. Frontiers in plant science, 7, p.2074.
- Guzmán Mar, J. L., L. Hinojosa Reyes, G. Mizanur Rahman and H. S. Kingston (2009) Simultaneous extraction of arsenic and Se species from rice products by microwave-assisted enzymatic extraction and analysis by ion chromatography-inductively coupled plasma-mass spectrometry. Journal of agricultural and food chemistry 57(8): 3005-3013.
- Haddad, L., J. Ross, A. Oshaug, L. E. Torheim and B. Cogill (2004). 5th report on the world nutrition situation. Nutrition for improved development outcomes. (Vol. 5). United Nations System Standing Committee on Nutrition.
- Hageman, S.P., van der Weijden, R.D., Weijma, J. and Buisman, C.J., (2013) Microbiological selenate to selenite conversion for Se removal. Water research, 47(7), pp.2118-2128.
- Hansen, D., P. J. Duda, A. Zayed and N. Terry (1998) Se removal by constructed wetlands: role of biological volatilization. Environmental Science & Technology 32(5): 591-597.
- Hanson, B., G. F. Garifullina, S. D. Lindblom, A. Wangeline, A. Ackley, K. Kramer, A. P. Norton, C. B. Lawrence and E. A. Pilon-Smits (2003) Se accumulation protects Brassica juncea from invertebrate herbivory and fungal infection. New Phytologist 159(2): 461-469.
- Hanson, B., S. D. Lindblom, M. L. Loeffler and E. A. Pilon-Smits (2004) Se protects plants from phloem-feeding aphids due to both deterrence and toxicity. New Phytologist 162(3): 655-662.
- Hartikainen, H. (2005) Biogeochemistry of Se and its impact on food chain quality and human health. Journal of Trace Elements in Medicine and Biology 18(4): 309-318.
- Hawkesford, M.J. and F.J. Zhao, (2007) Strategies for increasing the Se content of wheat. Journal of Cereal Science, 46(3): 282-292.
- He, Z. L., J. Shentu and X. E. Yang (2010) Manganese and Se. Trace elements in soils. 1st ed. John Wiley & Sons, United Kingdom: Pp. 481-497.
- Helzlsouer, K., Jacobs, R. and Morris, S., (1985) Acute Se intoxication in the United States. In Fed Proc 44, No. 1670, p. 27.
- Hilton, J., P. Hodson and S. Slinger (1982) Absorption, distribution, half-life and possible routes of elimination of dietary Se in juvenile rainbow trout (Salmo gairdneri). Comparative Biochemistry and Physiology Part C: Comparative Pharmacology 71(1): 49-55.
- Hira, C. K., K. Partal and K. Dhillon (2004) Dietary Se intake by men and women in high and low Se areas of Punjab. Public health nutrition 7(01): 39-43.
- Hopper, J. L. and D. R. Parker (1999) Plant availability of selenite and selenate as influenced by the competing ions phosphate and sulfate. Plant and Soil 210(2): 199-207.
- Horton, S. and J. Ross (2003) The economics of iron deficiency. Food policy 28(1): 51-75.
- Hotz, C. and K. H. Brown (2004) Assessment of the risk of zinc deficiency in populations and options for its control, International nutrition foundation: for UNU.
- Hu, Y., G. L. Duan, Y. Z. Huang, Y. X. Liu and G. X. Sun (2014) Interactive effects of different inorganic As and Se species on their uptake and translocation by rice (Oryza sativa L.) seedlings. Environ Sci Pollut Res Int 21(5): 3955-3962.
- Huang, J. W., J. Chen, W. R. Berti and S. D. Cunningham (1997) Phytoremediation of lead-contaminated soils: role of synthetic chelates in lead phytoextraction. Environmental Science & Technology 31(3): 800-805.
- Huang, J.C., Passeport, E. and Terry, N., (2012) Development of a constructed wetland water treatment system for Se removal: use of mesocosms to evaluate design parameters. Environmental science & technology, 46(21): 12021-12029.
- Huang, Y., Wang, Q., Gao, J., Lin, Z., Banuelos, G.S., Yuan, L. and Yin, X., (2013) Daily dietary Se intake in a high Se area of Enshi, China. Nutrients, 5(3): 700-710.
- Jayaweera, G. R. and J. W. Biggar (1996) Role of redox potential in chemical transformations of Se in soils. Soil Science Society of America Journal 60(4): 1056-1063.
- Kabata-Pendias, A. (2010) Trace elements in soils and plants, CRC press.
- Kahakachchi, C., H. T. Boakye, P. C. Uden and J. F. Tyson (2004) Chromatographic speciation of anionic and neutral Se compounds in Se-accumulating Brassica juncea (Indian mustard) and in selenized yeast. Journal of Chromatography A 1054(1): 303-312.
- Kamei-Ishikawa, N., K. Tagami and S. Uchida (2007) Sorption kinetics of Se on humic acid. Journal of Radioanalytical and Nuclear Chemistry 274(3): 555-561.
- Kápolna, E. and P. Fodor (2006) Speciation analysis of Se enriched green onions (Allium fistulosum) by HPLC-ICP-MS. Microchemical journal 84(1): 56-62.
- Kelly, S., M. Baxter, S. Chapman, C. Rhodes, J. Dennis and P. Brereton (2002) The application of isotopic and elemental analysis to determine the geographical origin of premium long grain rice. European Food Research and Technology 214(1): 72-78.
- Kibriya, M. G., F. Jasmine, M. Argos, W. J. Verret, M. Rakibuz-Zaman, A. Ahmed, F. Parvez and H. Ahsan (2007) Changes in gene expression profiles in response to Se supplementation among individuals with arsenic-induced pre-malignant skin lesions. Toxicology letters 169(2): 162-176.
- Kieliszek, M. and S. Błażejak (2013) Se: significance, and outlook for supplementation. Nutrition 29(5): 713-718.
- Kirk, G. (2004) The biogeochemistry of submerged soils, John Wiley & Sons. Chichester.
- Kirkby, E. A. and A. E. J. Johnston (2008) Soil and fertilizer phosphorus in relation to crop nutrition. In: White, P.J., Hammond, J.P. (eds) The ecophysiology of plant-phosphorus interactions, vol 7. Springer, Dordrecht.
- Krishnan, S. and P. Dayanandan (2003). "Structural and histochemical studies on grain-filling in the caryopsis of rice (Oryza sativa L.). Journal of biosciences 28(4): 455-469.
- Kryukov, G. V., S. Castellano, S. V. Novoselov, A. V. Lobanov, O. Zehtab, R. Guigó and V. N. Gladyshev (2003) Characterization of mammalian selenoproteomes. Science 300(5624): 1439-1443.
- Kulp, T. R. and L. M. Pratt (2004) Speciation and weathering of Se in Upper Cretaceous chalk and shale from South Dakota and Wyoming, USA. Geochimica et Cosmochimica Acta 68(18): 3687-3701.
- Kunli, L., X. Lirong, T. Jian'an, W. Douhu and X. Lianhua (2004) Se source in the selenosis area of the Daba region, South Qinling Mountain, China. Environmental Geology 45(3): 426-432.
- LeDuc, D. L. and N. Terry (2005) Phytoremediation of toxic trace elements in soil and water. Journal of Industrial Microbiology and Biotechnology 32(11-12): 514-520.
- LeDuc, D. L., A. S. Tarun, M. Montes-Bayon, J. Meija, M. F. Malit, C. P. Wu, M. AbdelSamie, C.-Y. Chiang, A. Tagmount and B. Neuhierl (2004) Overexpression of selenocysteine methyltransferase in Arabidopsis and Indian mustard increases Se tolerance and accumulation. Plant Physiology 135(1): 377-383.
- LeDuc, D. L., M. AbdelSamie, M. Móntes-Bayon, C. P. Wu, S. J. Reisinger and N. Terry (2006). Overexpressing both ATP sulfurylase and selenocysteine methyltransferase enhances Se phytoremediation traits in Indian mustard. Environmental Pollution 144(1): 70-76.
- Lemly, A.D., (2004) Aquatic Se pollution is a global environmental safety issue. Ecotoxicology and environmental safety, 59(1): 44-56.
- Lenz, M. and P. N. Lens (2009) The essential toxin: the changing perception of Se in environmental sciences. Science of the Total Environment 407(12): 3620-3633.
- Leustek, T., M. N. Martin, J.-A. Bick and J. P. Davies (2000) Pathways and regulation of sulfur metabolism revealed through molecular and genetic studies. Annual review of plant biology 51(1): 141-165.
- Levander, O. A., G. Alfthan, H. Arvilommi, C. Gref, J. Huttunen, M. Kataja, P. Koivistoinen and J. Pikkarainen (1983) Bioavailability of Se to Finnish men as assessed by platelet glutathione peroxidase activity and other blood parameters. The American journal of clinical nutrition 37(6): 887-897.
- Li, H. F., E. Lombi, J. L. Stroud, S. P. McGrath and F. J. Zhao (2010). Se speciation in soil and rice: influence of water management and Se fertilization. J Agric Food Chem 58(22): 11837-11843.
- Li, T., Xu, H., Zhang, Y., Zhang, H., Hu, X., Sun, Y., Gu, X., Luo, J., Zhou, D. and Gao, B., (2022) Treatment technologies for Se contaminated water: A critical review. Environmental Pollution, 299, p.118858.
- Liu, Y., F. Li, X. Yin and Z. Lin (2010) Plant-based biofortification: from phytoremediation to Se-enriched agriculture products. Green chemistry for environmental sustainability. Pp. 341-356. CRC press, Boca Raton.
- Lobanov, A. V., D. E. Fomenko, Y. Zhang, A. Sengupta, D. L. Hatfield and V. N. Gladyshev (2007) Evolutionary dynamics of eukaryotic selenoproteomes: large selenoproteomes may associate with aquatic life and small with terrestrial life. Genome biology 8(9): 1.
- Low, J. W., M. Arimond, N. Osman, B. Cunguara, F. Zano and D. Tschirley (2007) A food-based approach introducing orange-fleshed sweet potatoes increased vitamin A intake and serum retinol concentrations in young children in rural Mozambique. The Journal of nutrition 137(5): 1320-1327.
- Lu, J. and A. Holmgren (2009) Selenoproteins. Journal of Biological Chemistry 284(2): 723-727.
- Lyi, S. M., L. I. Heller, M. Rutzke, R. M. Welch, L. V. Kochian and L. Li (2005) Molecular and biochemical characterization of the selenocysteine Se-methyltransferase gene and Se-methylselenocysteine synthesis in broccoli. Plant Physiology 138(1): 409-420.
- Lynch, J. P. (2007) Turner review no. 14. Roots of the second green revolution. Australian Journal of Botany 55(5): 493-512.
- Lyons, G. H., G. J. Judson, I. Ortiz-Monasterio, Y. Genc, J. C. Stangoulis and R. D. Graham (2005a) Se in Australia: Se status and biofortification of wheat for better health. Journal of Trace Elements in Medicine and Biology 19(1): 75-82.
- Lyons, G., I. Ortiz-Monasterio, J. Stangoulis and R. Graham (2005b) Se concentration in wheat grain: Is there sufficient genotypic variation to use in breeding? Plant and Soil 269 (1-2): 369-380.
- Lyons, G., J. Stangoulis and R. Graham (2003) High-Se wheat: biofortification for better health. Nutrition Research Reviews 16(01): 45-60.
- Mao, H., J. Wang, Z. Wang, Y. Zan, G. Lyons and C. Zou (2014) Using agronomic biofortification to boost zinc, Se, and iodine concentrations of food crops grown on the loess plateau in China. Journal of soil science and plant nutrition 14(2): 459-470.
- McKenzie, R.C., Rafferty, T.S., Beckett, G.J., Arthur, J.R. (2001). Effects of selenium on immunity and aging. In: Hatfield, D.L. (eds) Selenium. Springer, Boston, MA. Pp: 21257-272. [CrossRef]
- McKevith, B., (2004) Nutritional aspects of cereals. Nutrition Bulletin, 29(2): 111-142.
- McKenzie, M.J., Hunter, D.A., Pathirana, R. et al. (2009) Accumulation of an organic anticancer selenium compound in a transgenic Solanaceous species shows wider applicability of the selenocysteine methyltransferase transgene from selenium hyperaccumulators. Transgenic Res 18, 407-424 . [CrossRef]
- McLaughlin, M. J., D. Parker and J. Clarke (1999). Metals and micronutrients–food safety issues. Field crops research 60(1): 143-163.
- Meenakshi, J., N. L. Johnson, V. M. Manyong, H. DeGroote, J. Javelosa, D. R. Yanggen, F. Naher, C. Gonzalez, J. Garcia and E. Meng (2010) How cost-effective is biofortification in combating micronutrient malnutrition? An ex ante assessment. World Development 38(1): 64-75.
- Mikkelsen, R. L., D. S. Mikkelsen and A. Abshahi (1989a) Effects of soil flooding on Se transformations and accumulation by rice. Soil Science Society of America Journal 53(1): 122-127.
- Mikkelsen, R., A. L. Page and F. T. Bingham (1989b) Factors affecting Se accumulation by agricultural crops. Se in Agriculture and the Environment (Seinagric): 65-94.
- Milovac, M., Djermanović, V. and Djujić, I., (1998) Effects of cereal supplementation with Se. Journal of Environmental Pathology, Toxicology and Oncology: Official Organ of the International Society for Environmental Toxicology and Cancer, 17(3-4): 313-320.
- Mishra, R. R., S. Prajapati, J. Das, T. K. Dangar, N. Das and H. Thatoi (2011) Reduction of selenite to red elemental Se by moderately halotolerant Bacillus megaterium strains isolated from Bhitarkanika mangrove soil and characterization of reduced product. Chemosphere 84(9): 1231-1237.
- Mombo, S., Schreck, E., Dumat, C., Laplanche, C., Pierart, A., Longchamp, M., Besson, P. and Castrec-Rouelle, M., 2016. Bioaccessibility of selenium after human ingestion in relation to its chemical species and compartmentalization in maize. Environmental geochemistry and health, 38: 869-883.
- Montes-Bayón, M., D. L. LeDuc, N. Terry and J. A. Caruso (2002) Se speciation in wild-type and genetically modified Se accumulating plants with HPLC separation and ICP-MS/ES-MS detection. Journal of Analytical Atomic Spectrometry 17(8): 872-879.
- Morgan, J., G. Bending and P. White (2005) Biological costs and benefits to plant–microbe interactions in the rhizosphere. Journal of Experimental Botany 56 (417): 1729-1739.
- Mouta, E. R., W. J. d. Melo, M. R. Soares, L. R. F. Alleoni and J. C. Casagrande (2008) Adsorção de selênio em latossolos. Revista Brasileira de Ciência do Solo: 1033-1041.
- Munier-Lamy, C., S. Deneux-Mustin, C. Mustin, D. Merlet, J. Berthelin and C. Leyval (2007) Se bioavailability and uptake as affected by four different plants in a loamy clay soil with particular attention to mycorrhizae inoculated ryegrass. Journal of environmental radioactivity 97(2): 148-158.
- Navarro-Alarcon, M. and C. Cabrera-Vique (2008) Se in food and the human body: a review. Science of the total environment 400(1): 115-141.
- Nestel, P., H. E. Bouis, J. Meenakshi and W. Pfeiffer (2006) Biofortification of staple food crops. The Journal of nutrition 136(4): 1064-1067.
- Neuhierl, B. and A. Böck (1996) "On the Mechanism of Se Tolerance in Se-Accumulating Plants. European Journal of Biochemistry 239(1): 235-238.
- Nève, J. (1996). Se as a risk factor for cardiovascular diseases. Journal of cardiovascular risk 3(1): 42-47.
- Nkansah-Boadu, F. and Baldwin, S.A., (2019) Biological Removal of Se from Mining Influenced Waters: A Critical Review of Challenges. In Proceedings of the IMWA 2019 Conference-Mine Water: Technological and Ecological Challenges.
- Novoselov, S. V., M. Rao, N. V. Onoshko, H. Zhi, G. V. Kryukov, Y. Xiang, D. P. Weeks, D. L. Hatfield and V. N. Gladyshev (2002) Selenoproteins and selenocysteine insertion system in the model plant cell system, Chlamydomonas reinhardtii. The EMBO journal 21(14): 3681-3693.
- Nowack, B., R. Schulin and B. H. Robinson (2006) Critical assessment of chelant-enhanced metal phytoextraction. Environmental Science & Technology 40(17): 5225-5232.
- Ogra, Y., K. Ishiwata, J. R. Encinar, R. Łobiński and K. T. Suzuki (2004) Speciation of Se in Se-enriched shiitake mushroom, Lentinula edodes. Analytical and bioanalytical chemistry 379(5-6): 861-866.
- Ogra, Y., K. Ishiwata, Y. Iwashita and K. T. Suzuki (2005) Simultaneous speciation of Se and sulfur species in selenized odorless garlic (Allium sativum L. Shiro) and shallot (Allium ascalonicum) by HPLC–inductively coupled plasma-(octopole reaction system)-mass spectrometry and electrospray ionization-tandem mass spectrometry. Journal of Chromatography A 1093(1): 118-125.
- Ohlendorf, H. M., R. W. Lowe, P. R. Kelly and T. E. Harvey (1986) Se and heavy metals in San Francisco Bay diving ducks. The Journal of wildlife management: 64-70.
- Parada, J. and J. Aguilera (2007) Food microstructure affects the bioavailability of several nutrients. Journal of food science 72(2): R21-R32.
- Pedrero, Z. and Y. Madrid (2009) Novel approaches for Se speciation in foodstuffs and biological specimens: a review. Analytica chimica acta 634(2): 135-152.
- Peterson, P.J. and Butler, G.W., (1967) Significance of selenocystathionine in an Australian Se-accumulating plant, Neptunia amplexicaulis. Nature, 213(5076): 599-600.
- Pilon, M., J. D. Owen, G. F. Garifullina, T. Kurihara, H. Mihara, N. Esaki and E. A. Pilon-Smits (2003) Enhanced Se tolerance and accumulation in transgenic Arabidopsis expressing a mouse selenocysteine lyase. Plant Physiology 131(3): 1250-1257.
- Pilon-Smits, E. (2005) Phytoremediation. Annu. Rev. Plant Biol. 56: 15-39.
- Pilon-Smits, E. A. and D. L. LeDuc (2009) Phytoremediation of Se using transgenic plants. Curr Opin Biotechnol 20(2): 207-212.
- Pilon-Smits, E. A. H. and C. F. Quinn (2010) Se Metabolism in Plants. Plant Cell Monographs 17: 225−241.
- Pilon-Smits, E.A., Hwang, S., Mel Lytle, C., Zhu, Y., Tai, J.C., Bravo, R.C., Chen, Y., Leustek, T. and Terry, N., 1999. Overexpression of ATP sulfurylase in Indian mustard leads to increased selenate uptake, reduction, and tolerance. Plant Physiology, 119(1), pp.123-132.
- Pilon-Smits, E.A., (2019) On the ecology of Se accumulation in plants. Plants, 8(7), p.197.
- Poblaciones, M., S. Rodrigo, O. Santamaría, Y. Chen and S. McGrath (2014) Agronomic Se biofortification in Triticum durum under Mediterranean conditions: From grain to cooked pasta. Food chemistry 146: 378-384.
- Poluboyarinov, P.A., Elistratov, D.G. & Moiseeva, I.J. (2020) antitumor activity of selenium and search parameters for its new potentially active derivatives. Russian Journal of Bioorganic Chemistry 46: 989–1003. [CrossRef]
- Potrykus, I. (2009) Lessons from Golden Rice on public sector responsibility and failure. New Biotechnology 25: S321-S322.
- Pray, C. and J. Huang (2008) Biofortification for China: political responses to food fortification and GM technology, interest groups, and possible strategies. Ag Bio Forum. 10(3): 2007.
- Premarathna, L., M. J. McLaughlin, J. K. Kirby, G. M. Hettiarachchi, S. Stacey and D. J. Chittleborough (2012) Selenate-enriched urea granules are a highly effective fertilizer for Se biofortification of paddy rice grain. The Journal of Agricultural and Food Chemistry. 60(23): 6037-6044.
- Prom-u-thai, C., S. Fukai, I. D. Godwin and L. Huang (2007) Genotypic variation of iron partitioning in rice grain. Journal of the Science of Food and Agriculture 87(11): 2049-2054.
- Qaim, M. and Stein, A.J., 2009. Biofortification of staple crops: how well does it work and what does it cost Ernährungs Umschau, 56(5): 274-280.
- Qaim, M., A. J. Stein and J. Meenakshi (2007) Economics of biofortification. Agricultural Economics 37(s1): 119-133.
- Qin, H.-b., J.-m. Zhu and H. Su (2012) Se fractions in organic matter from Se-rich soils and weathered stone coal in selenosis areas of China. Chemosphere 86(6): 626-633.
- Quinn, C. F., M. L. Galeas, J. L. Freeman and E. AH (2007) Se: Deterrence, toxicity, and adaptation. Peptides 125: 6160-6164.
- Quinn, C., J. Freeman, M. Galeas, E. Klamper and E. Pilon-Smits (2008) Se protects plants from prairie dog herbivory–implications for the functional significance and evolution of Se hyperaccumulation. Oecologia 155: 267-275.
- Ramos, S. J., Y. Yuan, V. Faquin, L. R. G. Guilherme and L. Li (2011) Evaluation of genotypic variation of broccoli (Brassica oleracea var. italic) in response to Se treatment. Journal of agricultural and food chemistry 59(8): 3657-3665.
- Rayman, M. P. (2000) The importance of Se to human health. The lancet 356(9225): 233-241.
- Rayman, M. P. (2008) Food-chain Se and human health: emphasis on intake. British journal of nutrition 100(02): 254-268.
- Rayman, M. P. (2012) Se and human health. The Lancet 379(9822): 1256-1268.
- Reeves, R. D. and A. J. Baker (2000) Metal-accumulating plants. Phytoremediation of toxic metals: using plants to clean up the environment. Wiley, New York: 193-229.
- Roberge, M. T., A. J. Borgerding and J. W. Finley (2003) Speciation of Se compounds from high Se broccoli is affected by the extracting solution. Journal of agricultural and food chemistry 51(15): 4191-4197.
- Robinson, B. H., G. Bañuelos, H. M. Conesa, M. W. Evangelou and R. Schulin (2009) The phytomanagement of trace elements in soil. Critical Reviews in Plant Sciences 28(4): 240-266.
- Rosenfeld, I. and O. A. Beath (2013) Se: geobotany, biochemistry, toxicity, and nutrition, Academic Press.
- Schiavon, M., Moro, I., Pilon-Smits, E.A., Matozzo, V., Malagoli, M. and Dalla Vecchia, F., (2012) Accumulation of selenium in Ulva sp. and effects on morphology, ultrastructure and antioxidant enzymes and metabolites. Aquatic toxicology, 122: 222-231.
- Schiavon, M., M. Pilon, M. Malagoli and E. A. Pilon-Smits (2015) Exploring the importance of sulfate transporters and ATP sulphurylases for Se hyperaccumulation—a comparison of Stanleya pinnata and Brassica juncea (Brassicaceae). Frontiers in plant science 6: 2.
- Schram, E., Z. Pedrero, C. Cámara, J. W. Van Der Heul and J. B. Luten (2008) Enrichment of African catfish with functional Se originating from garlic. Aquaculture Research 39(8): 850-860.
- Schrauzer, G. N. (2001). Nutritional Se supplements: product types, quality, and safety. Journal of the American College of Nutrition 20(1): 1-4.
- Searchinger, T., R. Heimlich, R. A. Houghton, F. Dong, A. Elobeid, J. Fabiosa, S. Tokgoz, D. Hayes and T.-H. Yu (2008) Use of US croplands for biofuels increases greenhouse gases through emissions from land-use change. Science 319(5867): 1238-1240.
- Séby, F., Gautier, M.P., Lespes, G. and Astruc, M., (1997) Se speciation in soils after alkaline extraction. Science of the Total Environment, 207(2-3): 81-90.
- Sharma, S., A. Bansal, S. K. Dhillon and K. S. Dhillon (2010) Comparative effects of selenate and selenite on growth and biochemical composition of rapeseed (Brassica napus L.). Plant and Soil 329(1-2): 339-348.
- Sharma, S., Bansal, A., Dogra, R., Dhillon, S.K. and Dhillon, K.S., (2011) Effect of organic amendments on uptake of Se and biochemical grain composition of wheat and rape grown on seleniferous soils in northwestern India. Journal of Plant Nutrition and Soil Science, 174(2): 269-275.
- Shen, L., P. V. Dael, L. Luten and H. Deelstra (1996) Estimation of Se bioavailability from human, cow's, goat and sheep milk by an in vitro method. International journal of food sciences and nutrition 47(1): 75-81.
- Shibagaki, N., A. Rose, J. P. McDermott, T. Fujiwara, H. Hayashi, T. Yoneyama and J. P. Davies (2002) Selenate-resistant mutants of Arabidopsis thaliana identify SULTR1; 2, a sulfate transporter required for efficient transport of sulfate into roots. The Plant Journal 29(4): 475-486.
- Sors, T. G., D. R. Ellis and D. E. Salt (2005) Se uptake, translocation, assimilation and metabolic fate in plants. Photosynth Res 86(3): 373-389.
- Spallholz, J. E., L. M. Boylan and M. Rhaman (2004) Environmental hypothesis: is poor dietary Se intake an underlying factor for arsenicosis and cancer in Bangladesh and West Bengal, India. Science of the total environment 323(1): 21-32.
- Stadlober, M., M. Sager and K. J. Irgolic (2001) Effects of selenate supplemented fertilisation on the Se level of cereals—identification and quantification of Se compounds by HPLC–ICP–MS. Food Chemistry 73(3): 357-366.
- Stadtman, T. (1991) Biosynthesis and function of selenocysteine-containing enzymes. Journal of Biological Chemistry 266(25): 16257-16260.
- Staicu, L.C., Van Hullebusch, E.D., Rittmann, B.E. and Lens, P.N., (2017) Industrial Se pollution: sources and biological treatment technologies. Bioremediation of Se contaminated wastewater, pp.75-101.
- Stein, A. J. (2009). Global impacts of human mineral malnutrition. Plant and Soil 335(1-2): 133-154.
- Stolz, J.F., Basu, P., Santini, J.M. and Oremland, R.S., (2006) Arsenic and Se in microbial metabolism. Annu. Rev. Microbiology, 60:107-130.
- Stroud, J., M. Broadley, I. Foot, S. Fairweather-Tait, D. Hart, R. Hurst, P. Knott, H. Mowat, K. Norman and P. Scott (2010) Soil factors affecting Se concentration in wheat grain and the fate and speciation of Se fertilisers applied to soil. Plant and soil 332(1-2): 19-30.
- Sun, G.-X., X. Liu, P. N. Williams and Y.-G. Zhu (2010) Distribution and translocation of Se from soil to grain and its speciation in paddy rice (Oryza sativa L.). Environmental science & technology 44(17): 6706-6711.
- Sura-de Jong, M., Reynolds, R.J., Richterova, K., Musilova, L., Staicu, L.C., Chocholata, I., Cappa, J.J., Taghavi, S., van der Lelie, D., Frantik, T. and Dolinova, I., (2015) Se hyperaccumulators harbor a diverse endophytic bacterial community characterized by high Se resistance and plant growth promoting properties. Frontiers in plant science, 6, p.113.
- Szpunar, J., R. Lobinski and A. Prange (2003) Hyphenated techniques for elemental speciation in biological systems. Applied spectroscopy 57(3): 102A-112A.
- Tan, J., W. Wang, D. Wang and S. Hou (1994) Adsorption, volatilization, and speciation of Se in different types of soils in China. Se in the Environment: 47-68.
- Tapiero, H., D. Townsend and K. Tew (2003) The antioxidant role of Se and seleno-compounds. Biomedicine & pharmacotherapy 57(3): 134-144.
- Terry, N., A. Zayed, M. De Souza and A. Tarun (2000) Se in higher plants. Annual review of plant biology 51(1): 401-432.
- Thavarajah, D., P. Thavarajah, A. Wejesuriya, M. Rutzke, R. P. Glahn, G. F. Combs and A. Vandenberg (2011) The potential of lentil (Lens culinaris L.) as a whole food for increased Se, iron, and zinc intake: preliminary results from a 3 year study. Euphytica 180(1): 123-128.
- Thiry, C., A. Ruttens, L. De Temmerman, Y.-J. Schneider and L. Pussemier (2012) Current knowledge in species-related bioavailability of Se in food. Food Chemistry 130(4): 767-784.
- Thomson, C. (2004) Assessment of requirements for Se and adequacy of Se status: a review. European Journal of Clinical Nutrition 58(3): 391-402.
- Thornton, I., D. Kinniburgh, G. Pullen and C. Smith (1983) Geochemical aspects of Se in British soils and implications to animal health [Toxicity and deficiency diseases; Great Britain]. Trace substances in environmental health: proceedings of University of Missouri's annual conference (USA).
- Turakainen, M., H. Hartikainen and M. M. Seppänen (2004) Effects of Se treatments on potato (Solanum tuberosum L.) growth and concentrations of soluble sugars and starch. Journal of agricultural and food chemistry 52(17): 5378-5382.
- United Republic of Tanzania (2020) National Biofortification Guidelines. Ministry of Agriculture, May 2020. https://www.kilimo.go.tz/uploads/dasip/English_version_Final_new2.pdf, Retrieved on Oct 30, 2023.
- Unnevehr, L. J. (1986). Consumer demand for rice grain quality and returns to research for quality improvement in Southeast Asia. American Journal of Agricultural Economics. 68(3): 634-641.
- Vamerali, T., M. Bandiera, P. Lucchini, N. M. Dickinson and G. Mosca (2014) Long-term phytomanagement of metal-contaminated land with field crops: integrated remediation and biofortification. European Journal of Agronomy. 53: 56-66.
- Van Huysen, T., S. Abdel-Ghany, K. L. Hale, D. LeDuc, N. Terry and E. A. Pilon-Smits (2003) Overexpression of cystathionine-γ-synthase enhances Se volatilization in Brassica juncea. Planta 218(1): 71-78.
- Vonderheide, A. P., K. Wrobel, S. S. Kannamkumarath, C. B'Hymer, M. Montes-Bayón, C. Ponce de León and J. A. Caruso (2002) "Characterization of Se species in Brazil nuts by HPLC-ICP-MS and ES-MS. Journal of agricultural and food chemistry 50(20): 5722-5728.
- Wang, D., Dinh, Q.T., Thu, T.T.A., Zhou, F., Yang, W., Wang, M., Song, W. and Liang, D., (2018) Effect of Se-enriched organic material amendment on Se fraction transformation and bioavailability in soil. Chemosphere, 199: 417-426.
- Wang, S., D. Liang, D. Wang, W. Wei, D. Fu and Z. Lin (2012) Se fractionation and speciation in agriculture soils and accumulation in corn (Zea mays L.) under field conditions in Shaanxi Province, China. Science of the total Environment 427: 159-164.
- Wang, X. and K. Taniguchi (2003) Does better nutrition enhance economic growth? The economic cost of hunger. Nutrition intake and economic growth. Studies on the cost of hunger. FAO, Rome.
- Wang, Z. and Y. Gao (2001). Biogeochemical cycling of Se in Chinese environments. Applied Geochemistry 16(11): 1345-1351.
- Wang, Z., S. Xie and A. Peng (1996) Distribution of Se in soybean samples with different Se concentration. Journal of Agricultural and Food Chemistry 44(9): 2754-2759.
- Watkinson, J.H. (1981) Changes of blood selenium in New Zealand adults with time and importation of Australian wheat. The American Journal of Clinical Nutrition. 34(5): 936-942.
- Welch, R. M. (2002) Breeding strategies for biofortified staple plant foods to reduce micronutrient malnutrition globally. The Journal of nutrition. 132(3): 495S-499S.
- Whanger, P. (2002) Selenocompounds in plants and animals and their biological significance. Journal of the American College of Nutrition. 21(3): 223-232.
- Whanger P. D. (2004) Selenium and its relationship to cancer: an update. British Journal of Nutrition. 91(1):11-28. doi:10.1079/BJN20031015.
- White, P. J. and M. R. Broadley (2009) Biofortification of crops with seven mineral elements often lacking in human diets--iron, zinc, copper, calcium, magnesium, Se and iodine. New Phytol. 182(1): 49-84.
- White, P. J., M. R. Broadley, H. C. Bowen and S. E. Johnson (2007) Se and its relationship with sulfur. Sulfur in plants an ecological perspective, Springer: 225-252.
- White, P., H. Bowen, P. Parmaguru, M. Fritz, W. Spracklen, R. Spiby, M. Meacham, A. Mead, M. Harriman and L. Trueman (2004) Interactions between Se and Sulphur nutrition in Arabidopsis thaliana. Journal of Experimental Botany 55(404): 1927-1937.
- WHO (2002) The world health report 2002: reducing risks, promoting healthy life, World Health Organization.
- WHO (2009) Global health risks: mortality and burden of disease attributable to selected major risks, World Health Organization.
- Williams, P. N., E. Lombi, G.-X. Sun, K. Scheckel, Y.-G. Zhu, X. Feng, J. Zhu, A.-M. Carey, E. Adomako and Y. Lawgali (2009). Se characterization in the global rice supply chain. Environmental science & technology. 43(15): 6024-6030.
- World Health Organization, 2006. Guidelines on food fortification with micronutrients. World Health Organization. https://iris.who.int/bitstream/handle/10665/43412/9241594012_eng.pdf, Retrieved on October 31, 2023.
- Wu, Z., G. S. Banuelos, Z. Q. Lin, Y. Liu, L. Yuan, X. Yin and M. Li (2015). Biofortification and phytoremediation of Se in China. Front Plant Science 6: 136.
- Xue, T., H. Hartikainen and V. Piironen (2001). Antioxidative and growth-promoting effect of Se on senescing lettuce. Plant and Soil. 237(1): 55-61.
- Yang, G., S. Wang, R. Zhou and S. Sun (1983). Endemic Se intoxication of humans in China. The American journal of clinical nutrition. 37(5): 872-881.
- Yang, X. E., W. R. Chen and Y. Feng (2007). Improving human micronutrient nutrition through biofortification in the soil-plant system: China as a case study. Environ Geochem Health. 29(5): 413-428.
- Yläranta, T. (1985). Increasing the Se content of cereal and grass crops in Finland. Academic dissertation, Agricultural Research Centre, Institute of Soil Science, Jokioinen. Yliopistopaino: Helsinki.
- Yoshimoto, N., H. Takahashi, F. W. Smith, T. Yamaya and K. Saito (2002). Two distinct high-affinity sulfate transporters with different inducibilities mediate uptake of sulfate in Arabidopsis roots. The Plant Journal 29(4): 465-473.
- Yuan, L., X. Yin, Y. Zhu, F. Li, Y. Huang, Y. Liu and Z. Lin (2012). Se in plants and soils, and selenosis in Enshi, China: implications for Se biofortification. Phytoremediation and biofortification, Springer: 7-31.
- Zayed, A. M. and N. Terry (1992). Se volatilization in broccoli as influenced by sulfate supply. Journal of Plant Physiology. 140(6): 646-652.
- Zayed, A., C. M. Lytle and N. Terry (1998). Accumulation and volatilization of different chemical species of Se by plants. Planta 206(2): 284-292.
- Zhang, J. (2009). "Evaluation of nanotoxicity of foods and drugs: Biological roperties of red elemental Se at nano size (nano-Se) in vitro and in vivo. Nanotoxicity, from in vivo and in vitro models to health risks.97-114.
- Zhang, L., P. F. Byrne and E. A. Pilon-Smits (2006). Mapping quantitative trait loci associated with selenate tolerance in Arabidopsis thaliana. New Phytologist. 170(1): 33-42.
- Zhang, Y. and J. N. Moore (1996). Se fractionation and speciation in a wetland system. Environmental science & technology 30(8): 2613-2619.
- Zhang, Z., Xiong, Y., Chen, H. and Tang, Y., 2020. Understanding the composition and spatial distribution of biological selenate reduction products for potential Se recovery. Environmental Science: Water Research & Technology, 6(8): 2153-2163.
- Zhao, F. J. and S. P. McGrath (2009). Biofortification and phytoremediation. Curr Opin Plant Biol 12(3): 373-380.
- Zhao, F.-J., Y. Su, S. Dunham, M. Rakszegi, Z. Bedo, S. McGrath and P. Shewry (2009). Variation in mineral micronutrient concentrations in grain of wheat lines of diverse origin. Journal of cereal science 49(2): 290-295.
- Zhou, X., Li, Y. and Lai, F., 2018. Effects of different water management on absorption and accumulation of Se in rice. Saudi Journal of Biological Sciences, 25(6): 1178-1182.
- Zhu, Y. G., E. A. Pilon-Smits, F. J. Zhao, P. N. Williams and A. A. Meharg (2009). Se in higher plants: understanding mechanisms for biofortification and phytoremediation. Trends Plant Sci 14(8): 436-442.
- Zimmermann, R. and M. Qaim (2004). Potential health benefits of Golden Rice: a Philippine case study. Food Policy 29(2): 147-168.
Figure 1.
Soil Se levels in the world. Regions naturally rich in Selenium appear in red; pink areas correspond to regions with naturally low Se concentration and white areas correspond to regions with unknown concentration. Source: Mombo et al. (2016).
Figure 1.
Soil Se levels in the world. Regions naturally rich in Selenium appear in red; pink areas correspond to regions with naturally low Se concentration and white areas correspond to regions with unknown concentration. Source: Mombo et al. (2016).
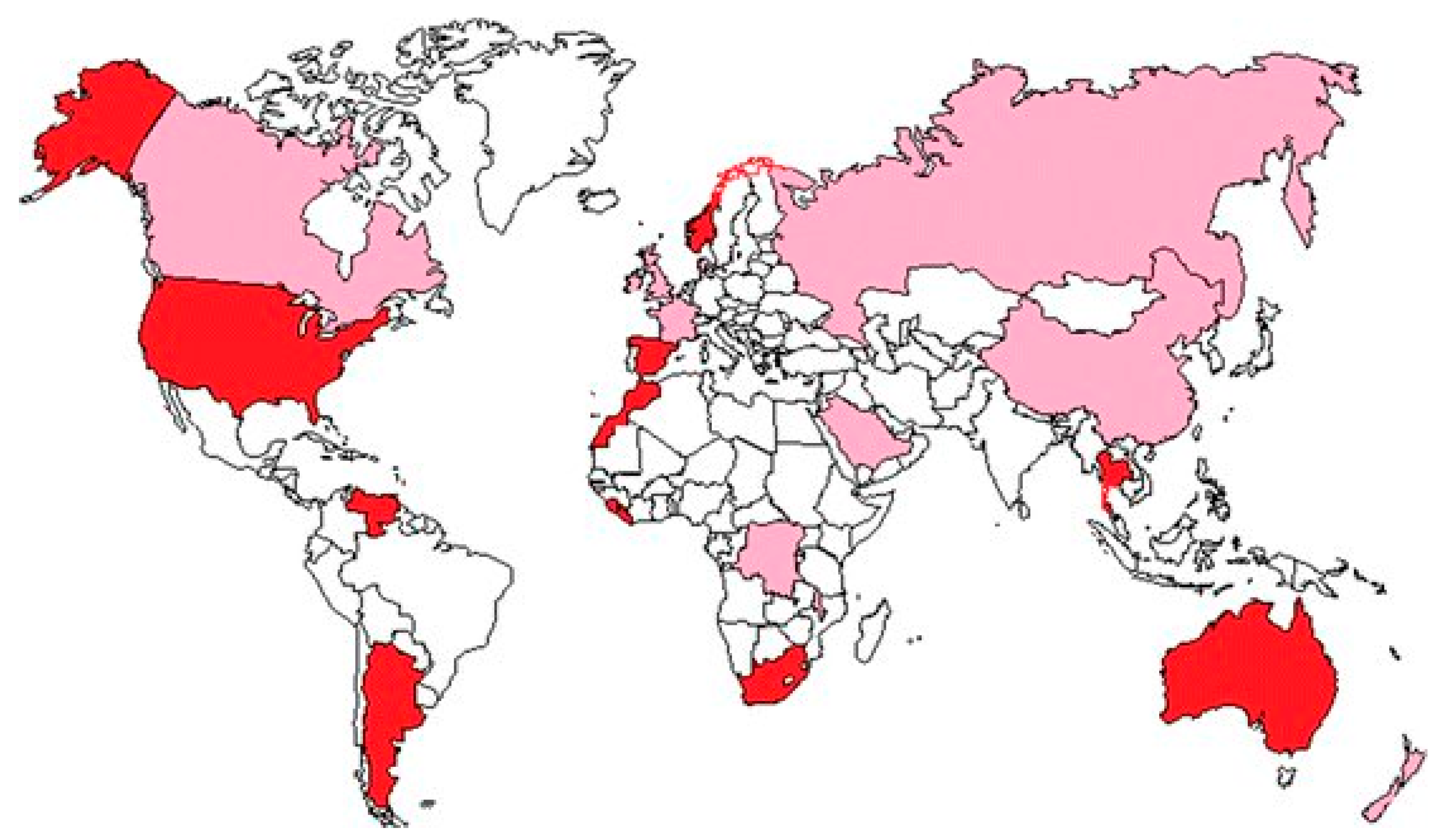
Figure 2.
Cycling of selenium in the human body.
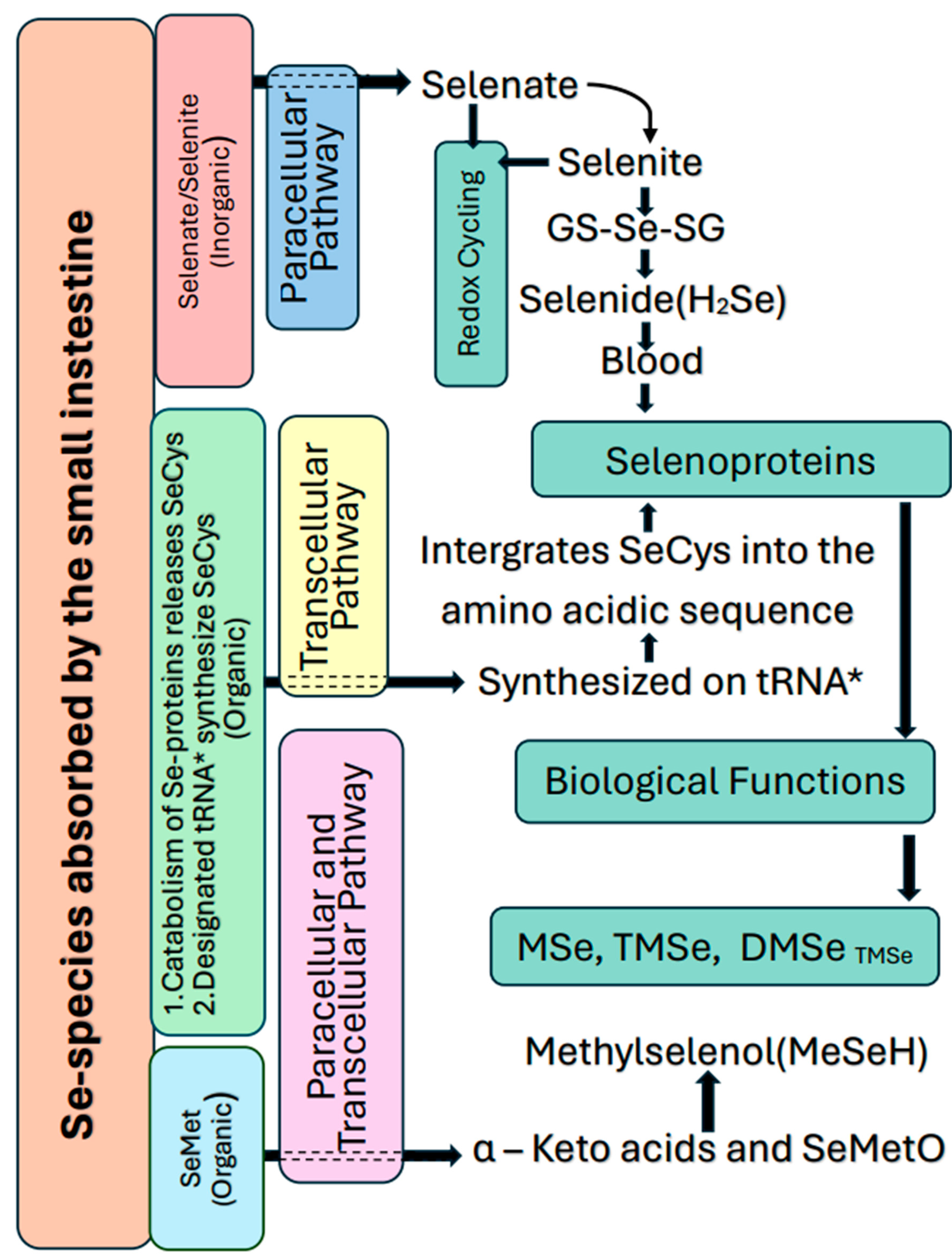
Figure 3.
Diverse forms of Se in the environment.
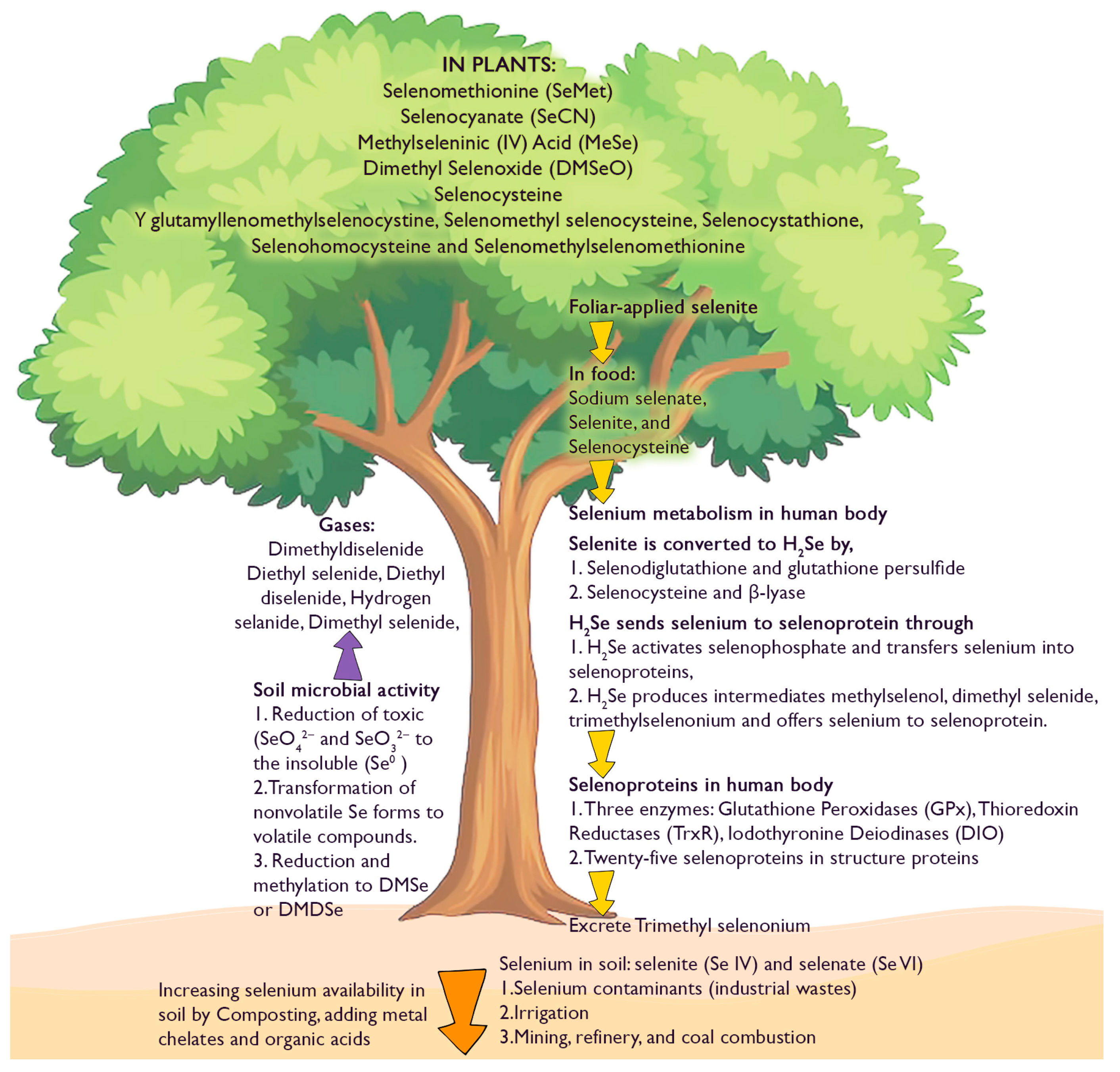
Figure 4.
Se metabolism in plant cell. DMSe: Dimethylselenide, DMDSe: Dimethyldiselenide, SeMet: Selenomethionine, SeHCys: Selenohomocysteine, SeCys: Selenocysteine, APSe: Adenosine phosphoselenate, APR: Adenosine phosphosulfate reductase, SMT: Selenomethyltransferase.
Figure 4.
Se metabolism in plant cell. DMSe: Dimethylselenide, DMDSe: Dimethyldiselenide, SeMet: Selenomethionine, SeHCys: Selenohomocysteine, SeCys: Selenocysteine, APSe: Adenosine phosphoselenate, APR: Adenosine phosphosulfate reductase, SMT: Selenomethyltransferase.
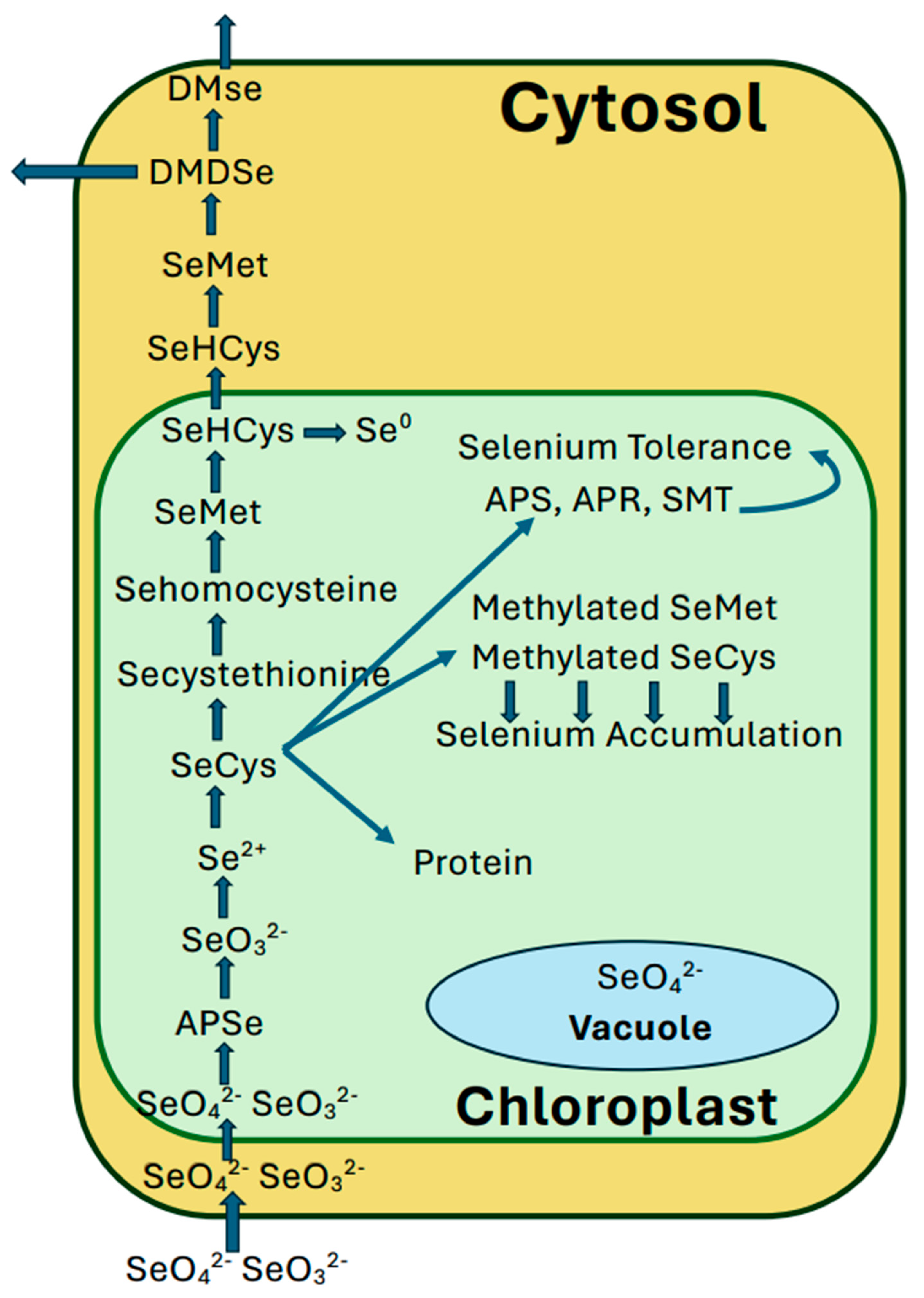

Table 1.
Se containing biological compounds and their role in the human body.
| Se in biological compound | Properties | Reference |
| Glutathione peroxidase (GPx) | Se containing enzyme, protecting cells from oxidative damage | Parada and Aguilera 2007; Thiry et al. 2012 |
| Thioredoxin reductases (TrxR) | Maintaining the cellular redox balance, important for various cellular processes, including DNA synthesis, repair, and cell growth. | Thiry et al. 2012; Lu and Holmgren 2014 |
| Iodothyronine deiodinases (DIO) | Responsible for the conversion of thyroid hormones. | Lu and Holmgren 2014 |
| Selenoprotein | Free-radical scavenging activity in the human body with cancer preventive properties | Clark et al. 1996; Rayman 2000; McKenzie et al. 2001; Abdulah et al. 2005 |
| Methylselenol | An anticancer agent | Abdulah et al. 2005 |
| Methylselenocysteine and selenomethionine | More anti-carcinogenic and more bioavailable compound** | Rayman 2008; Brummell et al. 2011 |
** Bioavailable fraction: The proportion of the available nutrient in the blood stream to ingested amount of nutrient (Parada and Aguilera 2007). *Bioactive fraction: The amount of Se that transformed into active seven metabolites are the bioactive Se (Thiry et al. 2012). Se is easily measured in biological tissues (Goldhaber 2003).
Table 2.
Prominent Se compounds in different crops.
| Crop | Prominent Se compounds | Reference |
| Garlic, onion, and leeks, Broccoli | Selenomethyl cysteine, methyl selenocysteine (MeSeCys), | Beilstein et al. 1991; Ogra et al. 2005; Avila et al. 2013 |
| Brassica and Allium | and Glu-MeSeCys |
Beilstein et al. 1991; Ogra et al. 2005; Whanger 2002; Rayman 2008 |
| Indian mustard | Selenomethionine(SeMet) SeOMet, | Grant et al. 2004 |
| Broccoli sprout | MeSeCys and glucosinolates | Avila et al. 2013 |
| Arabidopsis thaliana | Selenite | Beilstein et al. 1991; Kahakachchi et al. 2004; Pilon-Smits and Quinn 2010 |
| Wheat, barley, and rye | SeMet | Stadlober et al. 2001, Poblaciones et al. 2014 |
| Brazil nuts | SeMet | Vonderheide et al. 2002 |
| Shiitake mushrooms | SeMet | Ogra et al. 2004 |
| Rice | SeMet (Dominant), MeSeCys, SeCys | Beilstein et al. 1991; Li et al. 2010 |
| Radish | MeSeCys and -glutamyl S MeSeCys | Combs and Gray 1998; Tapiero et al. 2003 |
Table 3.
Se speciation in soil.
| Valency | Remarks | Reference |
| Se (0) | Produced by microbial activities | (Chen et al. 2009; Mishra et al. 2011). |
| Produced by the physical reduction of both Se (+4) and Se (+6) becomes oxidized forms | Chen et al. 2009; Mishra et al. 2011 | |
| Insoluble, immobile and poorly bioavailable | Kulp and Pratt 2004; Lenz and Lens 2009 | |
| Very common and found everywhere at the concentration range of 0.01 to 2.0 mg kg–1 | (Fordyce 2005). | |
| Elemental Se is prominent in anaerobic soils | Elrashidi et al. 1987 | |
| Se (+2) Selenide or Se2- |
Very slightly mobile in acid soils, forms stable minerals and organic compounds. The selenide minerals produced after biotic reduction, and selenide minerals are bioavailable. Major species: HSe−, H2Se° |
Kulp and Pratt 2004; Lenz and Lens 2009; Kabata-Pendias, and Szteke, 2015 |
| Selenide is prominent in anaerobic soils | Elrashidi et al. 1987; Wu et al. 2015 | |
| Se (+4) Selenite (SeO32-) Hydro selenite (HSeO4) |
The most toxic valence state organic matter and on Fe-, Mn- or Al oxidizes. Easily absorbed hydrous sesquioxides and organic matter Bound with oxides and oxyhydroxides of such metals is dissolved by microorganisms at later stages Major species: HSeO4–, SeO23– Selenite becomes predominant in acidic to neutral soils |
He et al. 2010; Wu et al. 2015 Wang et al. 2012; Kabata-Pendias, and Szteke, 2015; Elrashidi et al. 1987 |
| Selenite is available in acidic soils. Acid soluble fraction of Se (VI) is approximately 44% | Elrashidi et al. 1987; Wang et al. 2012 | |
| Selenite increases arsenic in rice | Hu et al. 2014 | |
| Se (+6) Selenate (SeO42-), SeO |
The dominant species that are highly and slowly absorbed by the plants. Alkaline soils solubilize Se (+6). Highly water soluble and highly bioavailable. Not adsorbed on hydrous sesquioxides (mainly Fe2O3·H2O) major species: SeO42– Selenate is the predominant in oxidized and alkaline soils |
Elrashidi et al. 1987; Séby et al., 1997; Se speciation in soils after alkaline extraction. Science of the Total Environment, 207(2-3), pp.81-90. Dhillon and Dhillon 2009 Kabata-Pendias, and Szteke, 2015 |
| Selenate species are available in aerobic neutral or alkaline soils | Wang et al. 2012 | |
| Under submerged or waterlogged conditions, selenate is converted to elemental Se, selenite or selenide | Mikkelsen et al. 1989b; Jayaweera and Biggar 1996 | |
| Se (+6) prevents arsenic penetration into rice roots | Hu et al. 2014 | |
| Acid-soluble fraction of Se (IV) is approximately 15% | Séby et al., 1997 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



