Submitted:
26 January 2025
Posted:
26 January 2025
You are already at the latest version
Abstract
Epilepsy is a widespread neurological disorder that remains a critical global public health challenge. While numerous antiepileptic drugs (AEDs) are available, many patients either fail to achieve adequate seizure control or experience significant side effects. One promising alternative is pure cannabidiol (CBD), but using a whole cannabis extract may be equally effective and preferred for some patients. In the current study, we employed the pentylenetetrazole (PTZ)-induced hyperactivity model in zebrafish to compare the effects of CBD with various cannabis extracts. We evaluated three cannabis strains, each subjected to three different extraction methods, and benchmarked the results against the commercially available AED valproic acid (VPA). Our findings revealed that 5.7 µg/ml of CBD and 0.01 mg/ml of different extracts significantly reduced movement compared to PTZ and VPA. In addition, effective extracts produced similar effects to pure CBD despite containing much lower molecule levels. These results reinforced and expanded previous evidence supporting the clinical potential of both CBD and whole cannabis extracts for seizure control while suggesting a possible entourage effect. Further research is necessary to determine which patients may benefit more from pure CBD versus those who might prefer the whole cannabis extracts.
Keywords:
epilepsy
; zebrafish
; cannabis extracts
; CBD
; entourage effect
1. Introduction
Epilepsy is one of the most prevalent neurological disorders, affecting approximately 50 million people worldwide, including up to 1% of children [1,2]. The disorder is marked by recurrent, unprovoked seizures that elevate the risk of injury, premature mortality, and diminished participation in daily activities, education, and employment [3]. The seizures result from excessive and synchronized electrical discharges in the brain [4], yet can occur even when no detectable brain abnormalities are present [5,6]. They manifest in diverse forms, ranging from brief lapses in consciousness or subtle involuntary movements to violent full-body convulsions, underscoring the disorder's varied nature [7]. In addition to the various forms of epilepsy, several other diseases may also present with epileptic seizures as a symptom [8].
Addressing the burden of seizures remains a global public health priority [2], although there are several antiepileptic drugs (AEDs) that target distinct mechanisms involved in seizure activity. Older AEDs, such as Carbamazepine, block sodium channels to reduce neuronal excitability, while benzodiazepines, such as Clobazam, are positive allosteric modulators of synaptic GABAA receptors [9,10,11]. Valproic acid (VPA), a commonly used AED, not only blocks sodium channels but also enhances GABA levels and reduces calcium influx, making it effective against various types of seizures, including generalized and absence seizures [12,13,14,15]. Yet, despite advancements in treatment, approximately 30% of patients do not achieve adequate seizure control with available AEDs, leading to refractory epilepsy [16,17]. Moreover, AEDs are associated with side effects that may lead to treatment discontinuation or deteriorated quality of life. Common side effects include headache, nausea, and dizziness, while more adverse effects include auditory and visual problems, skin problems, liver dysfunction, pancreatitis, and kidney disorders. Some AEDs may even result in life-threatening conditions as well as severe abnormalities, especially in patients with comorbidities or in pregnant women [18]. For example, VPA has been associated with disruptions in normal brain development, potentially leading to cognitive and behavioral deficits, an increased risk of neurodevelopmental disorders and impairments, and alterations in neurogenesis, gliogenesis, and synaptic function [19,20,21,22]. Hence, the search for safe and effective novel treatments continues.
Identifying new AEDs for treating epilepsy and seizures has been mostly done using rodent models [23]. However, using zebrafish (Danio rerio) may allow a more rapid scanning of novel therapeutics. Zebrafish offer practical benefits, including high fecundity, rapid development, and small size, which make them ideal for high-throughput, cost-effective drug screening [24]. Zebrafish models have emerged as valuable for AED research due to the genetic, physiological, and neurochemical similarities between zebrafish and humans, particularly within the central nervous system [25]. Key areas in the brain of zebrafish are homologous to the ones observed in mammals after 5 days postfertilization (dpf), hence allowing to compare the pattern of zebrafish behavior in different laboratory tests to the activity of older mice and rats [26]. Of note, the endocannabinoid system is highly conserved between zebrafish and mammals, including humans, with a high density of CB1 receptors in areas that bear signs of a functional homology to the mammalian hippocampus [27], which is the seizure-initiating zone in many epilepsy patients as well as in animal models of epilepsy [26].
In zebrafish, as in rodent models, epilepsy can be induced using convulsants like pentylenetetrazole (PTZ), leading to seizure-like behaviors. These behaviors are observable as distinct changes in swimming patterns, such as increased hyperactivity, providing a robust and efficient method for assessing the efficacy of AEDs such as VPA [28,29,30,31,32,33,34]. A recent example of such use is studies reporting that phytocannabinoids, including cannabidiol (CBD), can effectively reduce PTZ-induced hyperactivity in zebrafish models [35,36,37]. CBD is a non-psychoactive compound derived from Cannabis sativa, which demonstrated promising anticonvulsant properties in various animal seizure models, with potential mechanisms involving the endocannabinoid system and calcium signaling pathways [38,39]. Moreover, the FDA approved CBD-based AED (Epidiolex) following evidence from clinical trials involving patients with Lennox-Gastaut or Dravet syndromes [40,41].
CBD and Δ9-tetrahydrocannabinol (Δ9-THC) are the most studied compounds derived from Cannabis sativa. The plant produces these molecules in their acid form (THCA and CBDA) and decarboxylated, mainly under high temperatures, into their activated forms [42]. However, the plant produces hundreds of compounds, including many phytocannabinoids and terpenes. Several of these demonstrated therapeutic potential as anxiolytic, antidepressant, antitumor, immunomodulatory, and neuroprotective [42,43,44,45,46,47], and it was suggested that various compounds found in marijuana can enhance clinical effects [42,48,49].
Cannabidiol (CBD) has gained significant attention for its anticonvulsant properties, often being studied in isolation to delineate its specific mechanisms of action. However, the therapeutic potential of whole cannabis extracts, which contain a full spectrum of cannabinoids, terpenes, and other bioactive compounds, is increasingly recognized. These additional compounds may act synergistically with CBD, enhancing its effects through what is known as the "entourage effect." This phenomenon suggests that the combination of multiple cannabis-derived compounds may provide a broader or more potent therapeutic benefit compared to CBD alone. For this reason, many individuals and healthcare practitioners lean toward the use of whole cannabis extracts, valuing their perceived efficacy and the holistic approach they offer in managing complex conditions [50,51,52].
Here, we aimed to use the zebrafish epilepsy model, where generalized seizures are induced by PTZ, to investigate the anticonvulsant effects of whole cannabis extracts from three distinct strains. Each strain was prepared through three different extraction methods, yielding varying compositions. The therapeutic potential of the extracts was then compared to that of a positive control (VPA) and to that of pure CBD at different concentrations.
2. Materials and Methods
2.1. Zebrafish Husbandry
Zebrafish (Danio rerio) were maintained under standard animal care protocols [53]. Wildtype adult AB/ zebrafish were housed in a recirculating aquatic system (Aquazone) set to 28.5 ± 1°C with a pH range of 7.0–7.2 and a 14:10 hour light-dark cycle. Embryos from multiple breeding pairs were collected, pooled in Petri dishes containing E3 medium (5 mM NaCl, 0.17 mM KCl, 0.33 mM CaCl₂-2H₂O, 0.33 mM MgSO₄-7H₂O, 10 mM HEPES, pH 7.2), and examined to remove unfertilized ones. The fertilized embryos were then placed in an incubator maintained at 28°C.
2.2. Pure CBD
Pure CBD (obtained from Sigma-Aldrich, #C7515) was diluted in E3 media, maintaining a final DMSO (dimethyl sulfoxide, Sigma-Aldrich #472301-1L) concentration of 0.1% w/v to concentrations of 6, 9, 12, 15, and 18 µM (equivalent to 1.887, 2.830, 3.774, 4.717 and 5.660 µg/ml, respectively).
2.3. Cannabis Extractions
Inflorescences from various cannabis strains were obtained from a cannabis farm (Brlev Agricultural Crops Ltd, Israel). Three distinct extractions were performed on the inflorescences from each strain. Equal amounts of inflorescences (330 mg per strain) were distributed into three 15 ml test tubes. Two tubes were incubated at 25°C, while the third was incubated at 100°C for 1 hour to decarboxylate cannabinoid acids. Ten ml of ethyl acetate (EA) was added to the first, while 10 ml of ethanol was added to the second (E25) and third (E100) tubes, with each solution mixed using a magnetic stirrer for 90 minutes. The mixtures were centrifuged at 10,000 rpm for 10 minutes, after which the upper layers were collected. The solid phase was re-extracted twice with 3 ml of solvent each time. The upper layers from all three extractions were combined and evaporated under nitrogen. The resulting dried extracts were stored at -80°C until use.
Dried cannabis extracts were solubilized in DMSO at 200 mg/ml concentration and stored at −20°C. Immediately before seizure assays, the stored solutions of cannabis extracts were diluted in E3 media to reach the desired concentrations, maintaining a final DMSO concentration of 0.1% w/v. E3 media with 0.1% DMSO served as the vehicle control, while E3 media alone acted as the negative control.
2.4. Seizure Assay
At 5 dpf, zebrafish larvae were transferred individually to a 96-well plate, each well fitted with a 100 mesh filter insert (MultiScreen-MESH Filter Plate, Germany) containing 350 μl of buffered E3 media. Larvae were placed one per well and acclimated for 24 hours at 28°C in an incubator. On the test day, the plate was transferred to the Daniovision™ system and left for 30 minutes of acclimation. Thereafter, the insert plate containing the larvae were transferred to a "treatment plate" containing the negative and positive controls (DMSO or 5 mM VPA (Merck #PHR1061) or test substances (CBD or extracts in different concentrations) for 30 minutes. The viability of larvae after exposure to the extracts was assessed, and those that completely lost mobility were excluded from the experiment. The insert was then moved to a plate solution containing 15 mM of PTZ (#6500-25G, Sigma-Aldrich) for 30 minutes, during which motor activity was recorded. Following 30 minutes of PTZ exposure, a viability test was performed on the larvae. The experimental larvae were placed in darkness for 3 minutes to observe their movement patterns. A significant increase in movement during the dark phase, followed by a return to attenuated movement under light, indicated that the cannabis extract functioned as a suppressor of PTZ-induced hyperactivity rather than causing toxicity or sedation.
2.5. Video Tracking and Analysis
Behavioral testing was conducted using a DanioVision™ (Noldus Information Technology, Netherlands) lightproof recording chamber equipped with an infrared camera. The activity was measured with EthoVision XT13 (Noldus) software, which recorded the total distance traveled, segmented into 1-minute intervals over 30 minutes. Each group consisted of 12 larvae per treatment for the CBD experiment, with no replications. Each experimental group consisted of 12 larvae per treatment for the different extracts, with a minimum of three replicates per extract concentration. Pending no significant differences between groups, larval activity was pooled for each treatment group (for a total of n = 36/group), while all the controls were pooled to a single group (for a total of n = 108/group) for further analysis.
2.6. Characterizing Extract's Chemical Composition
Cannabinoids in the various cannabis extracts were isolated, identified, and quantified using ultra-high-performance liquid chromatography coupled with mass spectrometry (UHPLC-MS; Ultimate 3000, Thermo Fisher Scientific) and detected by a Q Exactive™ Plus Hybrid Quadrupole-Orbitrap. The sample injection volume was 5 μl. A standard solution containing 14 cannabinoid standards purchased from Merck was injected under identical conditions for comparison. Separation was achieved using an EC-C18 column (150 mm length, 3.0 mm diameter, 2.7 μm particle size; Agilent Technologies). The mobile phase consisted of 0.1% formic acid in double-distilled water (DDW) and 0.1% formic acid in acetonitrile (ACN), with a 0.4 ml/min flow rate. Gradient elution was as follows: 0–3 min, 60% ACN; 3–7 min, 60–80% ACN; 7–12 min, 80–90% ACN; 12–20 min, 90% ACN; 20–22 min, 90–60% ACN; and 22–25 min, 60% ACN. The column was maintained at 30°C, and the autosampler at 10°C. Ionization was performed in both positive and negative modes. Instrument parameters included a capillary voltage of 3500 V, capillary temperature of 350°C, and nitrogen gas flow rates of 35 ml/min for sheath gas, 10 ml/min for auxiliary gas, and 1 ml/min for sweep gas. The mass range was set from 150 to 800 m/z with a resolution of 70,000. Data analysis was conducted using Xcalibur and Freestyle software.
Terpenes from each extract were isolated and analyzed using Gas chromatography-mass spectrometry (GC-MS; Thermo Fisher TSQ 8000). Analysis was performed on an Equity-1 capillary column (60 m length, 0.25 mm internal diameter, 0.25 μm particle size). The injector temperature was set to 280°C, with a sample injection volume of 1 μl. Helium gas was used as the mobile phase at a 1 ml/min flow rate. The column temperature was programmed to increase by 5°C/min from 60°C to 275°C. Terpenes in the extracts were identified and quantified by comparing their retention times to those of standard solution (containing 34 abundant cannabis terpenes purchased from Merck) calibration curve injected in the GC-MS. Peaks with retention times differing from the standard solution were further analyzed by comparison with the NIST library.
2.7. Statistical Analysis
Data were analyzed using one-way ANOVA with a 95% confidence interval. If no significant differences were found between replicates, larval activity data were pooled by treatment and control groups for further analysis. The data was analyzed in Python using a custom in-house script. Distance moved was compared using a one-way ANOVA, and significant effects were further examined using the Tukey post-hoc test. Data is presented as Mean+SEM. In the first phase, CBD at different concentrations was compared to negative (DMSO) and positive (VPA) controls. In the 2nd phase, the same was done for extracts from the different strains, using different extraction methods and concentrations. Finally, effective treatments from the two phases were compared to identify favorable treatment options.
3. Results
3.1. Behavioral Effects
3.1.1. CBD Reduce PTZ-Induced Hyperactivity
ANOVA revealed a main Group effect (F7,88 = 47.85, P < 0.001), and post-hoc analysis revealed significantly reduced distance moved compared to PTZ following exposure to VPS and CBD at concentrations of 2.8, 3.8, 4.7, and 5.7 µg/ml (Figure 1). Further analysis revealed that, compared to VPA, there was a significantly greater distance moved following exposure to 2.8 µg/ml CBD and a significantly reduced distance moved following exposure to 5.7 µg/ml CBD. The 3.8, 4.7, and 5.7 µg/ml CBD concentrations were selected for subsequent analyses. Full descriptive statistics, effects sizes, and post-hoc results are presented in Tables S1–S3.
3.1.2. Extracts Reduce PTZ-Induced Hyperactivity
ANOVA revealed a main Group effect for all strains (CAN-1: F14,741 = 63.752, P < 0.001; CAN-2: F14,741 = 55.628, P < 0.001; CAN-3: F14,741 = 59.871, P < 0.001) and post-hoc analysis revealed significantly reduced distance moved following exposure to 0.01 mg/ml of the different extracts (Figure 2). At that concentration, across strains, movement compared to PTZ was reduced for all extracts, and compared to VPA, it was reduced for the E25 and E100 methods. For all other concentrations, there were no differences when compared to PTZ, while the distance moved was significantly higher compared to VPA. Complete descriptive statistics, effects sizes, and post-hoc results are presented in Tables S4–S6.
Hence, the concentration of 0.01 mg/ml was used for all strains and extraction methods for the following analyses.
3.1.3. Comparing Effective CBD and Extracts Concentrations
For CAN-1 (Figure 3A), ANOVA revealed a main Group effect for the cannabis extracts and CBD concentrations (F5,138 = 8.42, P < 0.001). Post-hoc analysis revealed that distance moved was significantly reduced for the E100 extract compared to 3.8 µg/ml CBD and for the E25 extract compared to 3.8 and 4.7 µg/ml CBD and EA. For CAN-2 (Figure 3B), ANOVA revealed a main Group effect for the cannabis extracts and CBD concentrations (F5,138 = 11.94, P < 0.001). Post-hoc analysis revealed that the distance moved was significantly reduced for the E100 extract compared to 3.8 and 4.7 µg/ml CBD E25 and EA. For the E25 extract, the distance moved was reduced compared to 3.8 µg/ml CBD but increased compared to 5.7 µg/ml CBD. For CAN-3 (Figure 3C), ANOVA revealed a main Group effect for the cannabis extracts and CBD concentrations (F5,138 = 13.57, P < 0.001). Again, post-hoc analysis revealed that the distance moved was significantly reduced for the E100 extract compared to 3.8 and 4.7 µg/ml CBD and E25 and EA. For the E25 and EA, the distance moved was increased compared to 5.7 µg/ml CBD. Effect sizes and post-hoc results are presented in Tables S8 and S9.
3.2. Extract profiles
The concentrations of cannabinoids in the solutions at 0.01 mg/ml of the different extracts are presented in Table 1. CAN-1 included high levels of THC and THCA, with elevated levels of THC following heating. CBD levels were considerably lower, with a minor increase following heating. CAN-2 included high levels of CBD and CBDA content following all extraction methods. THC levels were considerably lower but increased following heating. CAN-3 included relatively high levels of both THC and CBD, which further increased following heating but still were around half of their levels in CAN-1 and CAN-2, respectively.
The concentrations of terpenes in the 0.01 mg/ml solutions of the different extracts are presented in Table 2. Overall, the terpenes demonstrated a much more uniform distribution than cannabinoids. For CAN-1, there were higher levels of Beta-Caryophyllene when the extract was not heated (0.109 and 0.113% for the E25 and EA methods). For CAN-2, there were higher levels of Alpha Bisabolol for all methods (0.151, 0.126, and 0.14% for the E25, E100, and EA methods), while CAN-3 lacked any distinguished characteristic.
4. Discussion
In this work, we set to explore for the first time the anticonvulsant effects of whole cannabis extracts on PTZ-induced hyperactive larvae movement. The purpose of using two different solvents, ethanol and ethyl acetate, for cannabis extraction was to compare the relative contribution of cannabinoids and terpenes. Ethyl acetate was expected to extract a higher concentration of less polar terpenes, but the results revealed little differences between the two solvents. We found that 5.7 µg/ml of CBD and 0.01 mg/ml of different extracts significantly reduced movement compared to PTZ and VPA. For the THC-rich CAN-1, the E25 method seemed to present the best effect (with only 0.015 µg/ml CBD, 337 fold less concentrated than the best performance of pure CBD), while for the CBD-rich CAN-2 and intermittent CAN-3, the E100 method seemed to present the best effects (with 5.174 µg/ml CBD for CAN-2 and 2.661 µg/ml for CAN-3). E100 from CAN-3 demonstrated slightly better performance than the most effective pure CBD concentration, despite containing only half the CBD content. Its effects could be compared to 2.8 µg/ml of pure CBD, which exhibited a lower efficacy. These latter findings align with the historically popular methods of cannabis consumption, which primarily involve heating in various forms, with inhalation through cannabis-based cigarettes being the most common approach over time [54].
The results suggest that pure CBD and cannabis extract may be used for different applications or individuals. On the one hand, the fact that purified CBD can induce similar effects to those of the whole extract makes it a better candidate for clinical application due to the known concentration of the active ingredient, the lack of possible Δ9-THC's psychoactive effects, and the consequent reduced regulatory challenges. Moreover, studies indicated that concomitant CBD and Clobazam appear to produce additive benefits and may represent a promising approach to control seizures while reducing the side effects linked to Clobazam [54]. Similarly, while larvae treated with VPA exhibited significantly reduced survival rates compared to controls, co-treatment with VPA and CBD resulted in survival rates comparable to those of untreated controls. Notably, the combined treatment largely mitigated the adverse effects typically observed when each compound was administered individually [55]. However, purified CBD was also associated with more frequent mild and severe adverse effects compared to CBD-rich extracts [56]. On the other hand, natural compounds, such as whole cannabis extracts, are widely used as complementary natural therapies to manage seizures and other conditions [57,58]. For example, for chronic neuropathic pain, the benefit–safety profiles for cannabinoids were found to be higher than for other commonly used medications, mainly because they contribute more to quality of life and have a more favorable side effect profile [59]. Similarly, the majority of recreational and medical cannabis users report reducing or stopping their use of more conventional sleep aids after initiating cannabis use [60]. However, the response to cannabis is dose-dependent and with vast individuality in responses, and hence, the same dose may benefit some but not others [61]. For example, while most children treated with a formula that contained CBD and Δ9-THC (20:1 in olive oil) reported a reduction in seizure frequency, a minority reported aggravation of seizures [62].
Our results are relevant to the current debate in the literature regarding the possibility of an entourage effect. On the one hand, there are indications for the advantages of whole cannabis extract compared to single isolates [63]. For example, a meta-analysis found that CBD-rich extracts seem to present a better therapeutic profile than purified CBD in patients with refractory epilepsy [56]. On the other hand, recent reviews concluded that there is a lack of sound evidence supporting the existence of the entourage effect [64] and that additive enhancement of cannabinoid efficacy by terpenes remains unproven [48]. In zebrafish larvae, it was shown that the combined exposure of CBD and Δ9-THC appears to have a synergistic effect on PTZ-induced behavior [37]. However, there was also a significant reduction in the activity of control larvae, suggesting that Δ9-THC produces a general sedation that partially explains the decrease in the PTZ-induced activity pattern. Here, we showed that the effective extracts produced effects similar to pure CBD while containing a much lower concentration of this molecule. In addition, beneficial effects were observed in strains that contain low levels of Δ9-THC. Hence, our results support the entourage effect, although it is unknown whether it was supported by cannabinoids that demonstrated anti-seizure effects in former publications (e.g., [35]) or by combinations of cannabinoids and terpenes with supporting activity.
It should be noted that, compared to former studies, our results suggested the beneficial effect of pure CBD using relatively higher concentration (5.7 µg/ml or 18 µM), which may be due to protocol differences. In our study, larvae acclimated with the test solutions for 30 minutes before exposure to 15 mM PTZ, while activity was monitored for 30 minutes under light conditions. In the study by Samarut et al. [37], the exposure time was 1 h, and activity was monitored for 30 minutes after exposure to 2.5 or 5 mM PTZ. In that study, exposure to CBD at doses of 1.5-2.5 µM significantly reduced the total distance moved compared to PTZ. In the study by Thornton et al. [36], the exposure time was 24 hours, and activity was monitored for 15 minutes before and after exposure to 5 mM PTZ under light conditions. In that study, exposure to CBD at doses as low as 0.6-1.0 µM resulted in a significant reduction of total distance moved compared to PTZ. In the study by Kollipara et al. [35], the exposure time was 30 minutes, and activity was monitored for 30 minutes before and after exposure to 2.75 mM PTZ but under dark conditions. In that study, exposure to CBD at doses of 2-4 µM significantly reduced the total distance moved compared to PTZ. These results suggest that the most effective dose of CBD should be verified in the specific conditions of each study before comparison with other treatment options.
Despite the contributions of this study to the understanding of the anticonvulsant effects of cannabis extracts and cannabidiol (CBD) in a zebrafish model, several limitations warrant consideration. First, the use of the zebrafish model, while beneficial due to its genetic and physiological similarities to mammals, may not fully replicate the complexity of human epilepsy. As such, the translational relevance of these findings to human clinical practice should be carefully evaluated. Second, the study focused on a limited selection of cannabis strains and extraction methods, which may not encompass the full pharmacological diversity inherent in cannabis. The cannabis plant contains a wide array of cannabinoids and terpenes, the interactions and contributions of which to anticonvulsant effects remain incompletely understood. Future studies incorporating a broader range of cannabis cultivars, extraction techniques, and specific cannabinoid profiles are essential for elucidating their potential therapeutic properties. Additionally, this study primarily assessed short-term behavioral outcomes, limiting the ability to assess long-term efficacy and safety. Longitudinal investigations are necessary to determine therapeutic effects' sustainability and identify any delayed adverse effects. Lastly, the concentration range of CBD and cannabis extracts examined in this study was relatively narrow, which may not capture the full spectrum of their dose-dependent effects. Further dose-response studies are required to define the optimal therapeutic window better and to identify thresholds for maximal efficacy and safety. Addressing these limitations in future research will provide a more comprehensive understanding of the potential clinical applications of cannabis-based treatments for epilepsy.
In conclusion, our results validate and extend previous ones concerning the potential clinical effect of both CBD and whole cannabis extract for seizure control. Further research that will explore a broader range of strains and concentrations and different extraction methods and include more extended observation periods is needed to establish the contribution of terpenes and minor cannabinoids to seizure control and to allow personalized medicine.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, K.J. and P.A.P; methodology, K.J.; software, SPSS, CANVAS, ETHOVISION; validation, M.S.S, K.J.; formal analysis, M.S.S, R.S.; investigation, Y.B.; resources, S.K.; data curation, Y.B., S.K; writing—original draft preparation, P.A.P, K.J, M.S.S; writing—review and editing, S.T.; visualization, M.S.S.; supervision, P.A.P, K.J.; project administration, P.A.P, K.J.; funding acquisition, Tel Hai College. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The original data presented in the study are openly available in the Supplementary section.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Fine, A.; Wirrell, E.C. Seizures in Children. Pediatrics In Review 2020, 41, 321–347. [CrossRef]
- WHO. Epilepsy: A Public Health Imperative Available online: https://www.who.int/publications/i/item/epilepsy-a-public-health-imperative (accessed on 29 November 2024).
- Singh, G.; Sander, J.W. The Global Burden of Epilepsy Report: Implications for Low- and Middle-Income Countries. Epilepsy & Behavior 2020, 105, 106949. [CrossRef]
- Engel, J. Seizures and Epilepsy; OUP USA, 2013; ISBN 978-0-19-532854-7.
- Sirven, J.I. Epilepsy: A Spectrum Disorder. Cold Spring Harb Perspect Med 2015, 5, a022848. [CrossRef]
- Guerrini, R.; Marini, C.; Mantegazza, M. Genetic Epilepsy Syndromes Without Structural Brain Abnormalities: Clinical Features and Experimental Models. Neurotherapeutics 2014, 11, 269–285. [CrossRef]
- Devinsky, O.; Vezzani, A.; O’Brien, T.J.; Jette, N.; Scheffer, I.E.; de Curtis, M.; Perucca, P. Epilepsy (Primer). Nature Reviews: Disease Primers 2018, 4, 18024. [CrossRef]
- Neumann, A.-M.; Britsch, S. Molecular Genetics of Acquired Temporal Lobe Epilepsy. Biomolecules 2024, 14, 669. [CrossRef]
- Gierbolini, J.; Giarratano, M.; Benbadis, S.R. Carbamazepine-Related Antiepileptic Drugs for the Treatment of Epilepsy - a Comparative Review. Expert Opinion on Pharmacotherapy 2016, 17, 885–888. [CrossRef]
- Werner, F.-M.; Coveñas, R. Naturally Occurring and Exogenous Benzodiazepines in Epilepsy: An Update. In Naturally Occurring Benzodiazepines, Endozepines, and their Receptors; CRC Press, 2021 ISBN 978-0-367-81437-3.
- Rogawski, M.A. Reduced Efficacy and Risk of Seizure Aggravation When Cannabidiol Is Used without Clobazam. Epilepsy Behav 2020, 103. [CrossRef]
- Abdelsayed, M.; Sokolov, S. Voltage-Gated Sodium Channels: Pharmaceutical Targets via Anticonvulsants to Treat Epileptic Syndromes. Channels 2013, 7, 146–152. [CrossRef]
- Löscher, W. Basic Pharmacology of Valproate. Mol Diag Ther 2002, 16, 669–694. [CrossRef]
- Iftinca, M. Neuronal T–Type Calcium Channels: What's New? Iftinca: T–Type Channel Regulation. J Med Life 2011, 4, 126–138.
- Espinosa-Jovel, C.; Valencia, N. The Current Role of Valproic Acid in the Treatment of Epilepsy: A Glimpse into the Present of an Old Ally. Curr Treat Options Neurol 2024, 26, 393–410. [CrossRef]
- Chen, Z.; Brodie, M.J.; Liew, D.; Kwan, P. Treatment Outcomes in Patients With Newly Diagnosed Epilepsy Treated With Established and New Antiepileptic Drugs: A 30-Year Longitudinal Cohort Study. JAMA Neurology 2018, 75, 279–286. [CrossRef]
- Fattorusso, A.; Matricardi, S.; Mencaroni, E.; Dell’Isola, G.B.; Di Cara, G.; Striano, P.; Verrotti, A. The Pharmacoresistant Epilepsy: An Overview on Existent and New Emerging Therapies. Front. Neurol. 2021, 12. [CrossRef]
- Akyüz, E.; Köklü, B.; Ozenen, C.; Arulsamy, A.; Shaikh, M.F. Elucidating the Potential Side Effects of Current Anti-Seizure Drugs for Epilepsy. Current Neuropharmacology 2021, 19, 1865–1883. [CrossRef]
- Roullet, F.I.; Lai, J.K.Y.; Foster, J.A. In Utero Exposure to Valproic Acid and Autism — A Current Review of Clinical and Animal Studies. Neurotoxicology and Teratology 2013, 36, 47–56. [CrossRef]
- Verrotti, A.; Scaparrotta, A.; Cofini, M.; Chiarelli, F.; Tiboni, G.M. Developmental Neurotoxicity and Anticonvulsant Drugs: A Possible Link. Reproductive Toxicology 2014, 48, 72–80. [CrossRef]
- Zimmermann, F.F.; Gaspary, K.V.; Leite, C.E.; De Paula Cognato, G.; Bonan, C.D. Embryological Exposure to Valproic Acid Induces Social Interaction Deficits in Zebrafish (Danio Rerio): A Developmental Behavior Analysis. Neurotoxicology and Teratology 2015, 52, 36–41. [CrossRef]
- Babiec, L.; Wilkaniec, A.; Adamczyk, A. Prenatal Exposure to Valproic Acid Induces Alterations in the Expression and Activity of Purinergic Receptors in the Embryonic Rat Brain. Folia Neuropathol 2022, 60, 390–402. [CrossRef]
- Corrales-Hernández, M.G.; Villarroel-Hagemann, S.K.; Mendoza-Rodelo, I.E.; Palacios-Sánchez, L.; Gaviria-Carrillo, M.; Buitrago-Ricaurte, N.; Espinosa-Lugo, S.; Calderon-Ospina, C.-A.; Rodríguez-Quintana, J.H. Development of Antiepileptic Drugs throughout History: From Serendipity to Artificial Intelligence. Biomedicines 2023, 11, 1632. [CrossRef]
- Lessman, C.A. The Developing Zebrafish (Danio Rerio): A Vertebrate Model for High-Throughput Screening of Chemical Libraries. Birth Defects Research Part C: Embryo Today: Reviews 2011, 93, 268–280. [CrossRef]
- Saleem, S.; Kannan, R.R. Zebrafish: A Potential Preclinical Model for Neurological Research in Modern Biology. In Zebrafish Model for Biomedical Research; Bhandari, P.R., Bharani, K.K., Khurana, A., Eds.; Springer Nature: Singapore, 2022; pp. 321–345 ISBN 978-981-16-5217-2.
- Sierra, A.; Gröhn, O.; Pitkänen, A. Imaging Microstructural Damage and Plasticity in the Hippocampus during Epileptogenesis. Neuroscience 2015, 309, 162–172. [CrossRef]
- Lachowicz, J.; Szopa, A.; Ignatiuk, K.; Świąder, K.; Serefko, A. Zebrafish as an Animal Model in Cannabinoid Research. International Journal of Molecular Sciences 2023, 24, 10455. [CrossRef]
- Kundap, U.P.; Kumari, Y.; Othman, I.; Shaikh, M.F. Zebrafish as a Model for Epilepsy-Induced Cognitive Dysfunction: A Pharmacological, Biochemical and Behavioral Approach. Front. Pharmacol. 2017, 8. [CrossRef]
- Turrini, L.; Fornetto, C.; Marchetto, G.; Müllenbroich, M.C.; Tiso, N.; Vettori, A.; Resta, F.; Masi, A.; Mannaioni, G.; Pavone, F.S.; et al. Optical Mapping of Neuronal Activity during Seizures in Zebrafish. Sci Rep 2017, 7, 3025. [CrossRef]
- Liu, J.; Baraban, S.C. Network Properties Revealed during Multi-Scale Calcium Imaging of Seizure Activity in Zebrafish. eNeuro 2019, 6. [CrossRef]
- Burrows, D.R.W.; Samarut, É.; Liu, J.; Baraban, S.C.; Richardson, M.P.; Meyer, M.P.; Rosch, R.E. Imaging Epilepsy in Larval Zebrafish. European Journal of Paediatric Neurology 2020, 24, 70–80. [CrossRef]
- Milder, P.C.; Zybura, A.S.; Cummins, T.R.; Marrs, J.A. Neural Activity Correlates With Behavior Effects of Anti-Seizure Drugs Efficacy Using the Zebrafish Pentylenetetrazol Seizure Model. Front. Pharmacol. 2022, 13. [CrossRef]
- Messina, A.; Boiti, A.; Sovrano, V.A.; Sgadò, P. Micromolar Valproic Acid Doses Preserve Survival and Induce Molecular Alterations in Neurodevelopmental Genes in Two Strains of Zebrafish Larvae. Biomolecules 2020, 10, 1364. [CrossRef]
- Torres-Hernández, B.A.; Colón, L.R.; Rosa-Falero, C.; Torrado, A.; Miscalichi, N.; Ortíz, J.G.; González-Sepúlveda, L.; Pérez-Ríos, N.; Suárez-Pérez, E.; Bradsher, J.N.; et al. Reversal of Pentylenetetrazole-Altered Swimming and Neural Activity-Regulated Gene Expression in Zebrafish Larvae by Valproic Acid and Valerian Extract. Psychopharmacology 2016, 233, 2533–2547. [CrossRef]
- Kollipara, R.; Langille, E.; Tobin, C.; French, C.R. Phytocannabinoids Reduce Seizures in Larval Zebrafish and Affect Endocannabinoid Gene Expression. Biomolecules 2023, 13, 1398. [CrossRef]
- Thornton, C.; Dickson, K.E.; Carty, D.R.; Ashpole, N.M.; Willett, K.L. Cannabis Constituents Reduce Seizure Behavior in Chemically-Induced and Scn1a-Mutant Zebrafish. Epilepsy & Behavior 2020, 110, 107152. [CrossRef]
- Samarut, É.; Nixon, J.; Kundap, U.P.; Drapeau, P.; Ellis, L.D. Single and Synergistic Effects of Cannabidiol and Δ-9-Tetrahydrocannabinol on Zebrafish Models of Neuro-Hyperactivity. Front. Pharmacol. 2019, 10. [CrossRef]
- Leo, A.; Russo, E.; Elia, M. Cannabidiol and Epilepsy: Rationale and Therapeutic Potential. Pharmacological Research 2016, 107, 85–92. [CrossRef]
- Lazarini-Lopes, W.; Do Val-da Silva, R.A.; da Silva-Júnior, R.M.P.; Leite, J.P.; Garcia-Cairasco, N. The Anticonvulsant Effects of Cannabidiol in Experimental Models of Epileptic Seizures: From Behavior and Mechanisms to Clinical Insights. Neuroscience & Biobehavioral Reviews 2020, 111, 166–182. [CrossRef]
- Thiele, E.A.; Marsh, E.D.; French, J.A.; Mazurkiewicz-Beldzinska, M.; Benbadis, S.R.; Joshi, C.; Lyons, P.D.; Taylor, A.; Roberts, C.; Sommerville, K.; et al. Cannabidiol in Patients with Seizures Associated with Lennox-Gastaut Syndrome (GWPCARE4): A Randomised, Double-Blind, Placebo-Controlled Phase 3 Trial. The Lancet 2018, 391, 1085–1096. [CrossRef]
- Devinsky, O.; Cross, J.H.; Laux, L.; Marsh, E.; Miller, I.; Nabbout, R.; Scheffer, I.E.; Thiele, E.A.; Wright, S. Trial of Cannabidiol for Drug-Resistant Seizures in the Dravet Syndrome. New England Journal of Medicine 2017, 376, 2011–2020. [CrossRef]
- Ferber, S.G.; Namdar, D.; Hen-Shoval, D.; Eger, G.; Koltai, H.; Shoval, G.; Shbiro, L.; Weller, A. The "Entourage Effect": Terpenes Coupled with Cannabinoids for the Treatment of Mood Disorders and Anxiety Disorders. Current Neuropharmacology 2020, 18, 87–96. [CrossRef]
- Basavarajappa, B.S.; Subbanna, S. Unveiling the Potential of Phytocannabinoids: Exploring Marijuana’s Lesser-Known Constituents for Neurological Disorders. Biomolecules 2024, 14, 1296. [CrossRef]
- Caprioglio, D.; Amin, H.I.M.; Taglialatela-Scafati, O.; Muñoz, E.; Appendino, G. Minor Phytocannabinoids: A Misleading Name but a Promising Opportunity for Biomedical Research. Biomolecules 2022, 12, 1084. [CrossRef]
- Silva-Reis, R.; Silva, A.M.S.; Oliveira, P.A.; Cardoso, S.M. Antitumor Effects of Cannabis Sativa Bioactive Compounds on Colorectal Carcinogenesis. Biomolecules 2023, 13, 764. [CrossRef]
- Peeri, H.; Koltai, H. Cannabis Biomolecule Effects on Cancer Cells and Cancer Stem Cells: Cytotoxic, Anti-Proliferative, and Anti-Migratory Activities. Biomolecules 2022, 12, 491. [CrossRef]
- Abyadeh, M.; Gupta, V.; Paulo, J.A.; Gupta, V.; Chitranshi, N.; Godinez, A.; Saks, D.; Hasan, M.; Amirkhani, A.; McKay, M.; et al. A Proteomic View of Cellular and Molecular Effects of Cannabis. Biomolecules 2021, 11, 1411. [CrossRef]
- André, R.; Gomes, A.P.; Pereira-Leite, C.; Marques-da-Costa, A.; Monteiro Rodrigues, L.; Sassano, M.; Rijo, P.; Costa, M. do C. The Entourage Effect in Cannabis Medicinal Products: A Comprehensive Review. Pharmaceuticals 2024, 17, 1543. [CrossRef]
- Ben-Shabat, S.; Fride, E.; Sheskin, T.; Tamiri, T.; Rhee, M.-H.; Vogel, Z.; Bisogno, T.; De Petrocellis, L.; Di Marzo, V.; Mechoulam, R. An Entourage Effect: Inactive Endogenous Fatty Acid Glycerol Esters Enhance 2-Arachidonoyl-Glycerol Cannabinoid Activity. European Journal of Pharmacology 1998, 353, 23–31. [CrossRef]
- Boehnke, K.F.; Scott, J.R.; Litinas, E.; Sisley, S.; Clauw, D.J.; Goesling, J.; Williams, D.A. Cannabis Use Preferences and Decision-Making Among a Cross-Sectional Cohort of Medical Cannabis Patients with Chronic Pain. The Journal of Pain 2019, 20, 1362–1372. [CrossRef]
- Boehnke, K.F.; Gagnier, J.J.; Matallana, L.; Williams, D.A. Cannabidiol Product Dosing and Decision-Making in a National Survey of Individuals with Fibromyalgia. The Journal of Pain 2022, 23, 45–54. [CrossRef]
- Kvamme, S.L.; Pedersen, M.M.; Rømer Thomsen, K.; Thylstrup, B. Exploring the Use of Cannabis as a Substitute for Prescription Drugs in a Convenience Sample. Harm Reduct J 2021, 18, 72. [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of Embryonic Development of the Zebrafish. Developmental Dynamics 1995, 203, 253–310. [CrossRef]
- Stella, N. THC and CBD: Similarities and Differences between Siblings. Neuron 2023, 111, 302–327. [CrossRef]
- Licitra, R.; Damiani, D.; Naef, V.; Fronte, B.; Vecchia, S.D.; Sangiacomo, C.; Marchese, M.; Santorelli, F.M. Cannabidiol Mitigates Valproic Acid-Induced Developmental Toxicity and Locomotor Behavioral Impairment in Zebrafish. Journal of Biological Regulators and Homeostatic Agents 2023, 37, 4935–4946. [CrossRef]
- Pamplona, F.A.; da Silva, L.R.; Coan, A.C. Potential Clinical Benefits of CBD-Rich Cannabis Extracts Over Purified CBD in Treatment-Resistant Epilepsy: Observational Data Meta-Analysis. Front. Neurol. 2018, 9. [CrossRef]
- Sharifi-Rad, J.; Quispe, C.; Herrera-Bravo, J.; Martorell, M.; Sharopov, F.; Tumer, T.B.; Kurt, B.; Lankatillake, C.; Docea, A.O.; Moreira, A.C.; et al. A Pharmacological Perspective on Plant-Derived Bioactive Molecules for Epilepsy. Neurochem Res 2021, 46, 2205–2225. [CrossRef]
- Challal, S.; Skiba, A.; Langlois, M.; Esguerra, C.V.; Wolfender, J.-L.; Crawford, A.D.; Skalicka-Woźniak, K. Natural Product-Derived Therapies for Treating Drug-Resistant Epilepsies: From Ethnopharmacology to Evidence-Based Medicine. Journal of Ethnopharmacology 2023, 317, 116740. [CrossRef]
- Nutt, D.J.; Phillips, L.D.; Barnes, M.P.; Brander, B.; Curran, H.V.; Fayaz, A.; Finn, D.P.; Horsted, T.; Moltke, J.; Sakal, C.; et al. A Multicriteria Decision Analysis Comparing Pharmacotherapy for Chronic Neuropathic Pain, Including Cannabinoids and Cannabis-Based Medical Products. Cannabis and Cannabinoid Research 2022, 7, 482–500. [CrossRef]
- Stueber, A.; Cuttler, C. A Large-Scale Survey of Cannabis Use for Sleep: Preferred Products and Perceived Effects in Comparison to over-the-Counter and Prescription Sleep Aids. Explor Med. 2023, 4, 709–719. [CrossRef]
- Kitdumrongthum, S.; Trachootham, D. An Individuality of Response to Cannabinoids: Challenges in Safety and Efficacy of Cannabis Products. Molecules 2023, 28, 2791. [CrossRef]
- Tzadok, M.; Uliel-Siboni, S.; Linder, I.; Kramer, U.; Epstein, O.; Menascu, S.; Nissenkorn, A.; Yosef, O.B.; Hyman, E.; Granot, D.; et al. CBD-Enriched Medical Cannabis for Intractable Pediatric Epilepsy: The Current Israeli Experience. Seizure 2016, 35, 41–44. [CrossRef]
- Ross-Munro, E.; Isikgel, E.; Fleiss, B. Evaluation of the Efficacy of a Full-Spectrum Low-THC Cannabis Plant Extract Using In Vitro Models of Inflammation and Excitotoxicity. Biomolecules 2024, 14, 1434. [CrossRef]
- Christensen, C.; Rose, M.; Cornett, C.; Allesø, M. Decoding the Postulated Entourage Effect of Medicinal Cannabis: What It Is and What It Isn't. Biomedicines 2023, 11, 2323. [CrossRef]
Figure 1.
Effect of cannabidiol (CBD) concentrations on PTZ-induced larval movement. Total distance moved (mm) by zebrafish larvae during 30 minutes of PTZ exposure following pre-treatment with DMSO, VPA (positive control), or varying concentrations of CBD. ***p < 0.001 vs. PTZ; ##p < 0.01, ###p < 0.001 vs. VPA.
Figure 1.
Effect of cannabidiol (CBD) concentrations on PTZ-induced larval movement. Total distance moved (mm) by zebrafish larvae during 30 minutes of PTZ exposure following pre-treatment with DMSO, VPA (positive control), or varying concentrations of CBD. ***p < 0.001 vs. PTZ; ##p < 0.01, ###p < 0.001 vs. VPA.
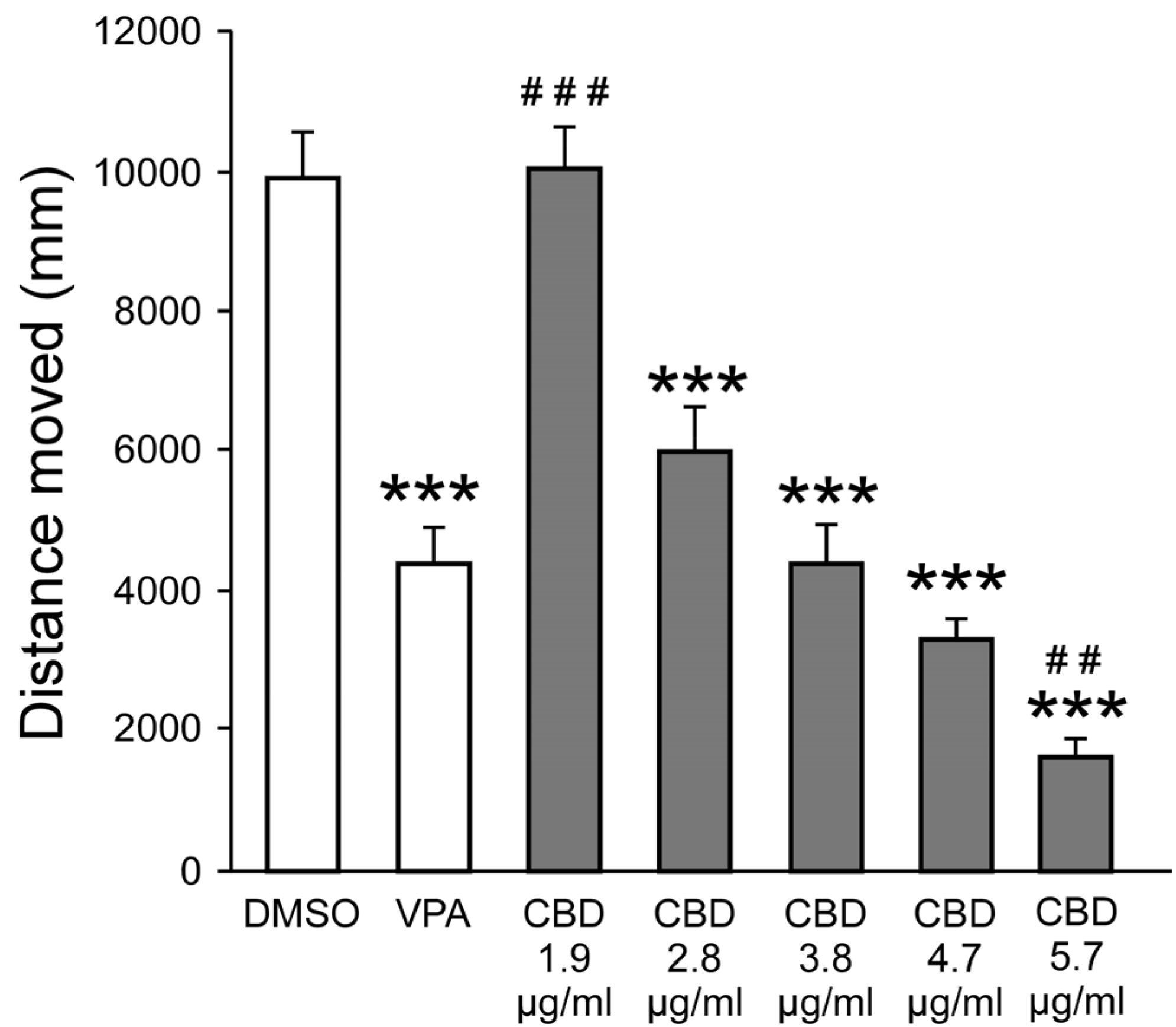
Figure 2.
Effect of CAN-1, CAN-2, and CAN-3 extracts on PTZ-induced larval movement. Total distance moved (mm) by zebrafish larvae during 30 minutes of PTZ exposure following pre-treatment with DMSO, VPA (positive control), or varying concentrations of extracts from CAN-1 (A), CAN-2 (B), and CAN-3 (C), obtained through different extraction methods (E100, E25, EA). ***p < 0.001 vs. PTZ; #p < 0.05, ##p < 0.01, ###p < 0.001 vs. VPA.
Figure 2.
Effect of CAN-1, CAN-2, and CAN-3 extracts on PTZ-induced larval movement. Total distance moved (mm) by zebrafish larvae during 30 minutes of PTZ exposure following pre-treatment with DMSO, VPA (positive control), or varying concentrations of extracts from CAN-1 (A), CAN-2 (B), and CAN-3 (C), obtained through different extraction methods (E100, E25, EA). ***p < 0.001 vs. PTZ; #p < 0.05, ##p < 0.01, ###p < 0.001 vs. VPA.
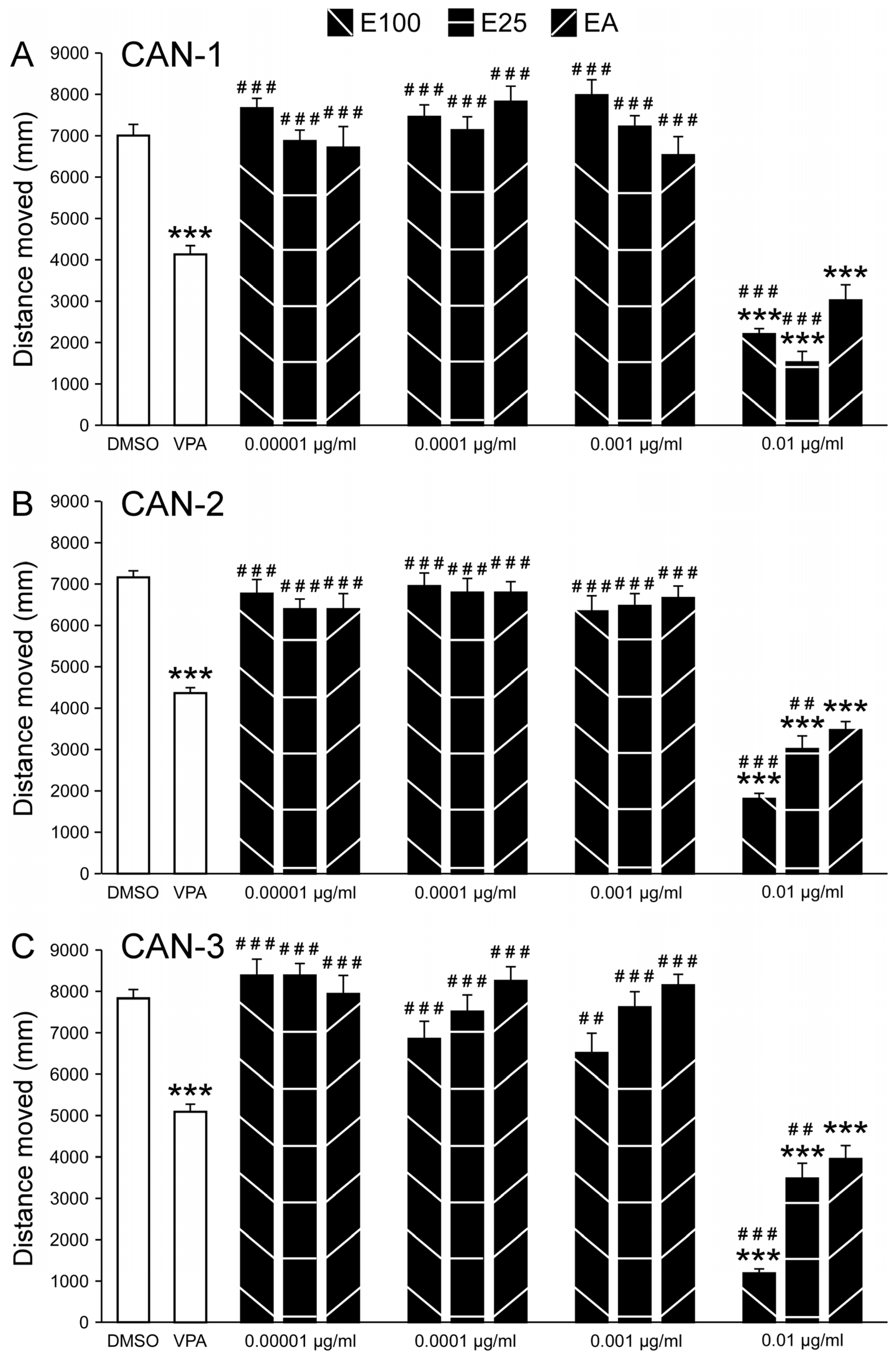
Figure 3.
Comparison of CAN-1, CAN-2, CAN-3, and CBD effects on PTZ-induced larval movement. Total distance moved (mm) by zebrafish larvae during 30 minutes of PTZ exposure following pre-treatment with CBD (n = 12/group) or 0.01 mg/mL extracts from cannabis strains CAN-1, CAN-2, and CAN-3, obtained using different extraction methods (E100, E25, EA; n = 36/group). * represents a significant difference compared to the indicated extract.
Figure 3.
Comparison of CAN-1, CAN-2, CAN-3, and CBD effects on PTZ-induced larval movement. Total distance moved (mm) by zebrafish larvae during 30 minutes of PTZ exposure following pre-treatment with CBD (n = 12/group) or 0.01 mg/mL extracts from cannabis strains CAN-1, CAN-2, and CAN-3, obtained using different extraction methods (E100, E25, EA; n = 36/group). * represents a significant difference compared to the indicated extract.
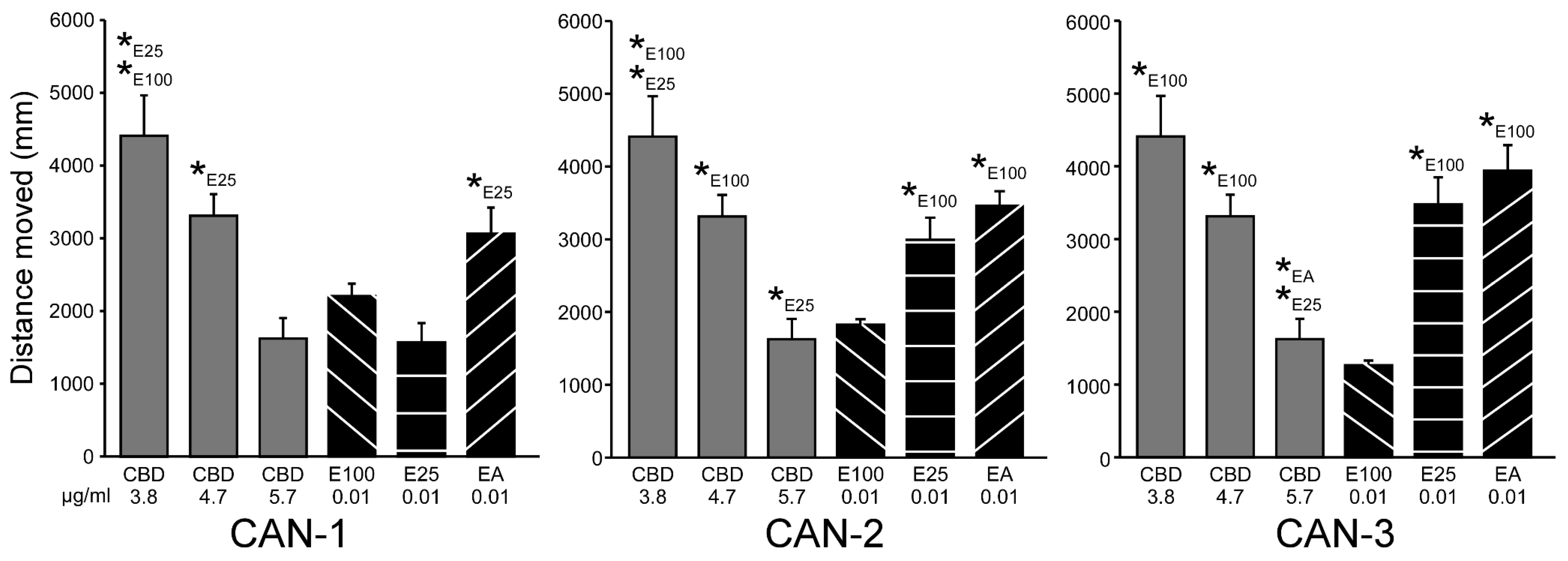

Table 1.
Cannabinoid content in the 0.01 mg/ml solutions of the different cannabis extracts (µg/ml).
Table 1.
Cannabinoid content in the 0.01 mg/ml solutions of the different cannabis extracts (µg/ml).
 |
Table 2.
Terpenes content in the 0.01 mg/ml solutions of the different cannabis extracts.
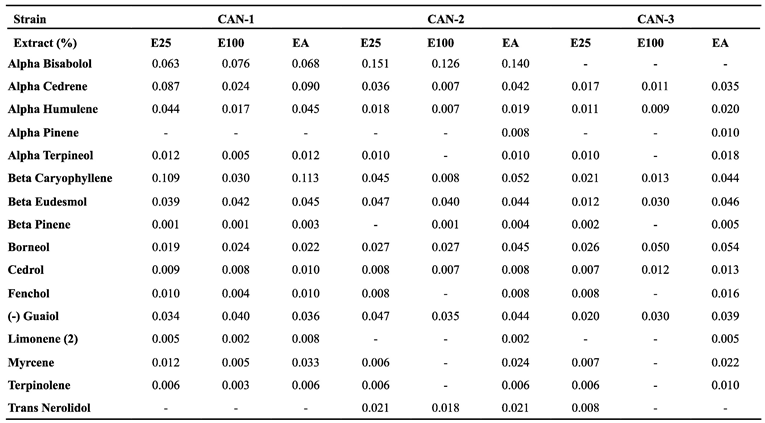 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



