
Submitted:
24 January 2025
Posted:
27 January 2025
You are already at the latest version
Abstract
Seed longevity, which holds paramount importance for agriculture and biodiversity conservation, continues to represent a formidable frontier in plant biology research. While advances have been made in identifying regulatory elements, the precise mechanisms behind seed lifespan determination remain intricate and context-specific. This comprehensive review compiles extensive findings on seed longevity across plant species, focusing on the genetic and environmental underpinnings. Inter-species differences in seed lifespan are tied to genetic traits, with numerous SEED LONGEVITY-ASSOCIATED GENEs (SLAGs) uncovered. These SLAGs encompass transcription factors and enzymes involved in stress responses, repair pathways, and hormone signaling. Environmental factors, particularly seed developmental conditions, significantly modulate seed longevity. Moreover, this review deliberates on the prospects of genetically engineering seed varieties with augmented longevity by precise manipulation of crucial genetic components, exemplifying the promising trajectory of seed science and its practical applications within agriculture and biodiversity preservation contexts. Collectively, our manuscript offers insights for improving seed performance and resilience in agriculture's evolving landscape.
Keywords:
seed longevity
; transcription factor
; molecular breeding
1. Introduction
Seed longevity, the inherent ability of seeds to remain viable during storage, plays a pivotal role in the perpetuation of successful plant reproduction [1]. Gradual loss of viability over time is an inherent aspect of seed aging, driven by degradation processes that ultimately reduce seedling emergence and vigor [2]. Since seeds serve as the primary vehicle for plant propagation, maintaining seed longevity is essential not only for sustaining agricultural productivity but also for conserving plant genetic diversity [3]. For cultivated crops specifically, enhanced seed longevity ensures higher probabilities of achieving optimal germination rates and sturdy seedling establishment, thereby contributing to improved crop output [2,3]. Against the backdrop of climate change, which imposes shifting selection pressures and drives changes in plant population genetics, seed conservation becomes a vital strategy in preventing the extinction of species or loss of plant communities that may not adapt or migrate at equal speeds [4]. Both in situ and ex situ seed conservation practices are thus recognized as indispensable for protecting global plant biodiversity [5]. Therefore, a thorough understanding of the complex factors influencing seed longevity carries profound ecological, agronomic, and economic implications.
Many plant species demonstrate impressive resilience to harsh environmental conditions when stored in a desiccated state [6]. Under such conditions, seeds enter a dormant phase where metabolic activity is drastically curtailed, yet their potential to germinate remains intact over extended periods [7,8]. Notable examples from botanical history abound, showcasing extraordinary seed longevity: date palm seeds (Phoenix dactylifera) have been carbon-dated to around 2000 years old [9], sacred lotus (Nelumbo nucifera) seeds have retained viability after 1300 years [10], and canna (Canna compacta) seeds have germinated after 600 years [11]. Inspired by seminal experiments like William Beal’s seed burial test initiated over a century ago, researchers continue to explore the mysteries of seed longevity [12], seeking answers to why certain seeds can survive for centuries longer than others [7].
Accumulating research delves into the molecular underpinnings of seed longevity, investigating how specific genes might confer this exceptional durability. Studies in Arabidopsis, rice, barley, maize, wheat, lettuce, oilseed rape, and tobacco, among other species, have uncovered genetic determinants of seed longevity [13,14,15,16,17,18,19,20,21,22]. An extensive body of work, particularly in Arabidopsis, a widely used model organism, has pinpointed SEED LONGEVITY ASSOCIATED GENEs (SLAGs) (Table 1). Manipulating these genes using molecular techniques has shown promise in altering seed longevity under experimental settings.
To systematize research efforts and facilitate further exploration, a comprehensive literature review has led to the assembly of information on SLAGs and their corresponding mutants across multiple species. This collective endeavor has resulted in the development of a dedicated database (https://ngdc.cncb.ac.cn/lsd/slag_mutant.php) [23], providing a rich resource and a firm groundwork for advancing knowledge into the molecular intricacies of seed longevity. This valuable tool enables researchers to delve deeper into the mechanisms that allow certain seeds to defy time, enduring for centuries, and paves the way for targeted interventions to enhance seed survival and preserve biodiversity.
2. Molecular Genetics Governing Seed Longevity
2.1. Transcription Factors in Regulating Seed Longevity
Transcriptional regulation serves as a pivotal coordinator governing diverse developmental processes and adaptive responses to a wide array of environmental challenges in plants [24,25]. At the epicenter of this regulatory mechanism are transcription factors (TFs), which exert profound control over fundamental facets of plant physiology [25,26]. Within the context of seed longevity, TFs constitute key regulators that dictate the expression of genes involved in maintaining seed vigor and viability over extended periods of storage and dormancy (Figure 1). Through their ability to bind to specific DNA sequences and modulate gene expression patterns, these TFs orchestrate a complex network that contributes to seed longevity and resilience.
The plant-specific TF ABSCISIC ACID-INSENSITIVE3 (ABI3) has emerged as a critical player in orchestrating seed dormancy and longevity via ABA-dependent pathways, as evidenced by extensive studies in Arabidopsis [27]. Mutations in the ABI3 gene lead to aberrant seed maturation, compromising dormancy, desiccation tolerance, and longevity, often accompanied by impaired chlorophyll breakdown [28]. ABI3 exerts its regulatory influence by binding to the evolutionarily conserved RY motif [CATGCA(TG)] prevalent within the promoter regions of numerous seed-specific genes [29]. Notably, AtHSFA9 and TONOPLAST INTRINSIC PROTEIN 3;1 (TIP3;1), both bearing RY motifs in their promoter sequences, serve as downstream targets of ABI3 and contribute significantly to seed longevity enhancement. While AtHSFA9 is a seed-specific heat shock factor that bolsters longevity upon activation [30], TIP3;1, a seed-specific aquaporin, also contributes positively to longevity under ABI3 regulation [31]. Loss of function of AtHSFA2 or AtHSFA9 significantly reduces seed longevity in Arabidopsis, whereas overexpresion of AtHSFA2 or AtHSFA9 leads to the increased accumulation of heat shock proteins (HSPs) and superior seed longevity [32]. As a chaperone of HSFA2, HSP90 interacts with ROTAMASE FKBP 1 (ROF1) and ROF2 to bolster seed longevity [33] (Figure 1). Accordingly, disruption of ROF1/ROF2 results in increased sensitivity to accelerated aging and poor germination under adverse conditions [33]. Moreover, the orthologs of AtHSFA9 across various plant species consistently demonstrate their ability to augment seed longevity. For instance, the overexpression of Helianthus annuus HSFA9 (HaHSFA9) or Medicago truncatula HSFA9 (MtHSFA9) results in enhanced seed thermo-tolerance and longevity, thus presenting promising candidates for molecular breeding interventions [34,35]. The interplay between TFs further underscores the complexity of seed longevity regulation. Helianthus annuus DROUGHT RESPONSIVE ELEMENT BINDING FACTOR 2 (HaDREB2), an AP2/ERBP family member, amplifies the seed longevity effects of HaHSFA9 when co-expressed, potentially by disrupting the suppressive interaction between HaHSFA9 and AUXIN-RESPONSIVE PROTEIN 27 (HaIAA27), a protein encoded by the AUXIN/INDOLE-3-ACETIC ACID (Aux/IAA) gene, thereby liberating HaHSFA9’s function to promote longevity [36,37]. However, HaDREB2 alone does not increase seed longevity without HaHSFA9.
Expanding the scope, research continues to reveal the multifaceted roles of additional TF families in regulating seed longevity. Members of the DNA BINDING WITH ONE FINGER (DOF) family, which are plant-specific TFs with a broad spectrum of biological functions [38], have been implicated in modulating seed longevity. Genetic evidence from the Arabidopsis dof4.1 loss-of-function mutant shows enhanced seed viability following artificial aging treatments, suggesting that DOF4.1 operates as a negative regulator of seed longevity [39]. Transcriptomic analysis unveiled that the expression of DELAY OF GERMINATION (DOG1), a positive regulator of seed maturation and longevity [40,41], is upregulated in the dof4.1 mutant compared to wild type plants, suggesting that DOF4.1 may negatively regulate seed longevity by repressing DOG1 [39]. Conversely, COGWHEEL1 (COG1/DOF1.5) and CYCLING DOF FACTOR 4 (CDF4/DOF2.3) serve as positive regulators, with their overexpression conferring resistance to seed deterioration in Arabidopsis [42,43]. COG1 enhances seed longevity possibly by increasing expressions of peroxidase genes based on the transcriptomic analysis of cog1-2D, a gain-of-function mutant with increased seed longevity [42]. Co-expression network analysis identified the TFs, WRKY3 and NFXL1, as components involved in seed longevity, as loss-of-function mutants of wrky3 and nflx1 exhibit reduced seed longevity [44]. Genome-wide association study (GWAS) revealed several TFs, including MYB TF (MYB47), MADS box TF (SEPALLATE 3, SPE3), and homeodomain (HB) TF (KNOTTED-LIKE HOMEOBOX OF ARABIDOPSIS THALIANA 7, KNAT7), as positive regulators of seed longevity [45]. The athb25-1D dominant Arabidopsis mutant, with higher expression of HOMEOBOX PROTEIN 25, displays improved seed longevity [46], further supporting the involvement of HB TFs in seed longevity regulation.
High-throughput RNA sequencing has identified SEUSS, a transcriptional corepressor linked to embryonic development [47], as being significantly upregulated in aged seeds of Astronium fraxinifolium [48]. While this suggests a potential role in seed longevity, the exact regulatory mechanisms remain unclear. In the floral meristem, SEUSS is known to interact with APETALA1 (AP1) and SHORT VEGETATIVE PHASE (SVP) to repress homeotic gene expression, thereby preventing premature differentiation of the floral meristem [47]. Given this, it is plausible that in the context of seed longevity, SEUSS may be recruited by specific transcription factors to form a complex that fine-tunes the expression of genes related to seed longevity.
Seed vigor and longevity are pivotal for enhancing grain quality and germplasm conservation in crops. RNA-seq co-expression regulatory network analyses identified the bZIP transcription factors bZIP23 and bZIP42 as candidate genes for seed longevity in rice [49]. The overexpression of bZIP23 robustly elevates seed vigor, a process linked to the activation of PEROXIDASE 1A (PER1A), unveiling a bZIP23-PER1A-mediated detoxification pathway that fortifies seed vigor [49]. This finding underscores the potential of targeted manipulation of key TFs through genome editing as a viable strategy to boost seed vigor, overall seed quality, and ultimately crop yield. In conclusion, transcriptional regulation, particularly through TFs like ABI3, DOF, MYB, MADS box, HB, and bZIP family proteins, constitutes a sophisticated network that deeply impacts seed longevity and vigor. These discoveries provide fertile ground for innovative agricultural advancements and germplasm conservation practices in crop species.
2.2. Impact of DNA Damage Repair on Seed Longevity
Seeds in their desiccated state possess extraordinary survival capabilities, yet they face a significant challenge as substantial DNA damage accumulates during storage, accelerating seed aging and impairing vigor [50]. To unravel the intricate defense mechanisms against this damage, researchers have turned to mutants with altered DNA repair-related genes [1,51]. This line of inquiry has shed light on the essential contribution of specific elements within the DNA repair pathway to seed longevity (Figure 1). Key players in this context include the proteins ATAXIA TELANGIECTASIA MUTATED (ATM) and ATM AND RAD3-RELATED (ATR) [52], SUPPRESSOR OF GAMMA 1 (SOG1) [53], DNA LIGASE 4 and 6 (LIG4/6) [14,54], KU70, X-RAY REPAIR CROSS COMPLEMENTING 2 (XRCC2), POLY(ADP-RIBOSE) POLYMERASE 1 (PARP1) and PARP3, EXCISION REPAIR CROSS COMPLEMENTING-GROUP 1 (ERCC1) [53], 8-OXOGUANINE (8-OXOG) and DNA GLYCOSYLASE 1 (OGG1)[55], as well as WHIRLY 1 (WHY1) and WHY3 [56].
Among the various forms of DNA damage, double-strand breaks (DSBs) are particularly detrimental [57]. The recognition of DSBs sparks intricate intracellular signaling cascades governed by the protein kinases ATM and ATR [58,59]. Mutant plants lacking functional ATM are more sensitive to DSBs and exhibit early-onset leaf senescence [60]. An intriguing observation is that seeds from ATM mutants germinate more rapidly than wild-type seeds following accelerated aging under harsh conditions of high temperature and humidity [52]. Despite this faster germination, aged ATM mutant seeds show a high prevalence of chromosomal abnormalities. Moreover, seedlings arising from these aged seeds experience reduced survival rates and slower development of true leaves compared to wild-type seedlings, emphasizing ATM’s vital role in maintaining the genomic integrity of the germinating embryo [52].
Organisms combat DSBs using two primary repair pathways: homologous recombination (HR) and non-homologous end-joining (NHEJ) [61]. When subjected to accelerated aging, mutant lines deficient in either HR (xrcc2-1, why1 why3) or NHEJ (ku70-1, ku80-3, lig4 lig6) pathways exhibit a marginally delayed germination [53,54,56]. Additionally, base excision repair (BER) and nucleotide excision repair (NER) pathways, exemplified by mutants arp1 and ogg1 (BER) and ercc1 (NER), also play a role in maintaining seed viability under stress, showing slightly delayed germination under similar conditions [53]. In summary, seed longevity is critically dependent on the proper functioning of DNA repair pathways, especially those involving ATM, ATR, and other associated components. Mutations in these genes can impact seed vigor and chromosomal stability. The ATM mutants, although demonstrating accelerated germination, suffer from compromised chromosomal integrity, revealing a delicate balance between rapid germination and genomic fidelity [52]. By deepening our understanding of the role DNA repair plays in seed aging, we can develop informed strategies to enhance seed viability and bolster crop resilience.
2.3. Role of Protein Repair or Homeostasis in Maintaining Seed Longevity
Reactive oxygen species (ROS) cause oxidative damage to seed proteins during storage, affecting their structural integrity and functionality, a typical hallmark of aging [62]. Among the amino acids, Methionine (Met) is notably susceptible to oxidation, transforming into S- and R-diastereomers of methionine sulfoxide (MetSO) [63,64]. Methionine sulfoxide reductase (MSR), consisting of MSRA and MSRB subtypes that specifically reduce Met-S-SO and Met-R-SO back to Met, respectively [65], counteracts this oxidation. MSR activity positively correlates with seed longevity across various plant species and its expression levels [66,67,68]. In aged rice seeds, reduced MSR activity and elevated MetSO content correlate with decreased seed vigor [66]. Overexpression of the seed-specific enzyme, OsMSRB5, effectively diminishes MetSO formation, thereby enhancing seed vigor and longevity by optimizing ROS balance [66], underscoring the critical role of MSR in sustaining seed longevity.
Simultaneously, cellular proteins inherently incur covalent damage that contributes to aging [69]. During seed aging, damaged proteins accrue abnormal isoaspartate (isoAsp) residues, degrading seed vigor and longevity. This issue is addressed by Protein-L-isoaspartyl methyltransferase (PIMT), which repairs these residues and positively affects seed longevity across several plant taxa [70,71,72,73]. PIMT activity, predominantly observed in seeds, is instrumental for seed vigor and longevity. Elevating PIMT gene expression boosts seed longevity and germination vigor in Arabidopsis and chickpea [70,73]. Analogously, overexpression of OsPIMT1 in rice reduces isoAsp accumulation, thus improving embryo viability and extending seed longevity [72]; conversely, loss of PIMT function leads to decreased seed vigor under stress conditions. The pivotal part played by PIMT in extending seed longevity underscores its critical importance in combating the detrimental effects of isoAsp accumulation during seed senescence.
Seeds can survive extreme desiccation for millennia by entering a state of quiescence. This involves accumulating protective storage proteins and lipids through intricate adjustments in protein homeostasis. Recently, reseachers found that disruption of proteostasis tiggerred by mutations of type-II metacaspase (MCA-II) proteases compromises seed longevity in Arabidopsis [74]. MCA-II mutant seeds fail to confine the AAA ATPase CDC48 (CELL DIVISION CYCLE 48) to the endoplasmic reticulum, leading to the accumulation of misfolded proteins and compromised seed viability. The localization of CDC48 to the endoplasmic reticulum is contingent upon MCA-II-mediated cleavage of PUX10 (ubiquitination regulatory X domain–containing 10), an adaptor protein that regulates the association of CDC48 with lipid droplets. PUX10 cleavage facilitates the dynamic shuttling of CDC48 between lipid droplets and the endoplasmic reticulum, a critical regulatory mechanism for maintaining spatiotemporal proteolysis, lipid droplet dynamics, and overall protein homeostasis. Interestingly, removing the PUX10 adaptor in MCA-II mutant seeds partially restores proteostasis, CDC48 localization, and lipid droplet dynamics, thereby extending seed lifespan [74]. This work reveals a novel proteolytic module essential for seed longevity.
2.4. Role of RFOs in Regulating Seed Longeivity
Raffinose family oligosaccharides (RFOs), including raffinose, stachyose and verbascose, play a critical role in seed longevity and vigor [75,76,77,78,79]. During seed maturation, RFOs accumulate alongside other compounds such as sucrose and LEA proteins, contributing to desiccation tolerance and the preservation of cellular integrity. This, in turn, enhances seed longevity and vigor [75,76,77,78,79].
Galactinol synthase (GolS), a key enzyme in RFO biosynthesis, is upregulated during seed development, leading to increased RFO levels [75]. Overexpression of Cicer arietinum CaGolS1/2 or Arachis duranensis AdGolS3 in Arabidopsis has been shown to improve seed vigor and longevity [75,80]. Additionally, mutations in GolS genes can lead to reduced galactinol levels and decreased seed lifespan [79]. The critical role of RFOs in longevity is further underscored by the zmdreb2a mutant in maize, which exhibits decreased seed longevity due to reduced expression of ZmRS (raffinose synthase), a gene responsible for raffinose synthesis, and consequently lower RFO accumulation [77]. Interestingly, the relationship between RFOs and seed vigor is complex and may vary between plant species. In Arabidopsis, the total RFO content and RFO/sucrose ratio, rather than individual RFO amounts, are positively correlated with seed vigor [76]. In contrast, in maize, raffinose appears to be the primary RFO associated with seed vigor [76]. The expression of GolS is regulated by various factors. Heat shock cis-elements (HSEs) have been identified in the promoter of BnGolS1 in Brassica napus [21], suggesting regulation by heat shock factors (HSFs). BnHSFA4a, a heat shock transcription factor, binds to these HSEs and activates BnGolS1 expression. Additionally, BnHSFA4a can directly regulate the expression of other genes involved in RFO biosynthesis, such as BnGolS2 and BnRS2, further enhancing RFO production and improving seed longevity and stress tolerance [21].
RFOs are hydrolyzed during seed germination, but the specific genes involved in this process are not fully understood. Maize alkaline α-galactosidase 1 (ZmAGA1) is a key enzyme responsible for RFO hydrolysis [81]. Overexpression of ZmAGA1 enhances seed germination under stress conditions but may also negatively impact seed aging tolerance [81], suggesting a potential trade-off between seed germination and longevity. In support of this observation, integrated quantitative trait locus (QTL) analyses for seed longevity in Arabidopsis reveal a negative correlation between seed longevity and seed dormancy [14]. In summary, RFOs play a crucial role in seed longevity and vigor. Their biosynthesis is regulated by factors such as GolS and HSFs, while the hydrolysis of RFOs during germination is mediated by enzymes like ZmAGA1. Understanding the complex interplay between RFO biosynthesis, hydrolysis, and seed quality is essential for developing strategies to improve seed longevity and vigor.
2.5. Hormonal Regulation of Seed Longevity
Phytohormones play a central role in orchestrating the complex series of events during seed maturation, profoundly impacting essential quality attributes such as germination potential, dormancy, and longevity [82]. Advances in molecular-genetic, biochemical, and pharmacological research have progressively uncovered the detailed contributions of phytohormones to seed longevity and the underlying regulatory mechanisms (Figure 1).
2.5.1. ABA: A Central Regulator of Seed Longevity
Among the phytohormones, abscisic acid (ABA) is a pivotal regulator intricately involved in controlling seed longevity, dormancy, and desiccation tolerance [27]. Deficiencies in ABA synthesis and signaling components significantly impinge on seed longevity, as demonstrated in several studies [1,13,83,84]. For example, the aba1 mutant, unable to produce epoxy-carotenoid precursors necessary for ABA biosynthesis, exhibits drastically reduced ABA levels compared to wild-type plants in Arabidopsis. Dominant mutations like abi1-1 and abi2-1, which affect genes coding for type 2C protein phosphatases (PP2C), interfere with ABA signaling by inhibiting SUCROSE NON-FERMENTING 1-RELATED PROTEIN KINASE 2 (SnRK2), leading to attenuated ABA responsiveness. Notably, ABA-deficient mutants (aba1) and ABA-insensitive mutants (abi1-1 and abi2-1) display reduced desiccation tolerance and longevity in Arabidopsis [85].
The perception of ABA begins with the engagement of intracellular receptors, specifically pyrabactin resistance 1 (PYR1) and PYR1-like (PYL) proteins, which form complexes with clade A PP2Cs, ultimately activating SnRK2 protein kinases [86]. Activated SnRK2s then modulate the expression of ABA-responsive genes by phosphorylating transcription factors like ABA-responsive element-binding factors (ABFs) [86]. Accordingly, mutants devoid of functional PYR/PYL, SnRK2, or ABF2/3/4 display compromised longevity relative to wild-type plants. Interestingly, SnRK2.6 regulates seed longevity by phosphorylating aquaporin PIP2;1 [86], suggesting that ABA controls seed longevity through regulating aquaporin function at both transcriptional and post-translational levels.
2.5.2. Impact of Auxin on Seed Longevity
Auxin, a pivotal plant hormone, exerts a sophisticated influence on the attainment of seed longevity [37,87,88]. During the maturation of Arabidopsis seeds, there is a concurrent escalation and spatial distribution of auxin signaling inputs and outputs within the embryo, tightly aligned with the seed’s journey towards achieving longevity [88]. Experimental supplementation of auxin during the maturation phase has been demonstrated to enhance seed longevity. Mutants with dysfunctional auxin biosynthesis pathways consistently exhibit altered longevity, reflecting a clear dose-response relationship that is tied to the intensity of auxin signaling activity [88]. The identification of a conserved gene network related to seed longevity, enriched with the cis-regulatory element ARFAT, an auxin response factor binding site, underscores the direct link between auxin signaling and the acquisition of longevity [44]. Moreover, biochemical evidence reveals that auxins enhance seed longevity by destabilizing the HaIAA27 protein and thus stimulating HSFA9 expression [37], providing additional insights into the molecular mechanisms underlying auxin regulation of seed longevity.
Auxin’s downstream actions intersect with the ABA signaling cascade within the embryo. It has been found that auxin promotes the expression of ABI3 and its LEA protein target, EARLY METHIONINE1 (EM1), with ABI3 activity shown to be dysregulated in the auxin biosynthesis mutant cyp79b2 [88]. More importantly, the beneficial influence of external auxin application on seed longevity during development is negated in abi3-1 mutants, underscoring the synergy between auxin and ABA pathways [88]. Beyond its interaction with the ABA signaling pathway, auxin may also directly modulate genes pertinent to seed longevity, implicating its involvement through both ABA-dependent and independent routes. This dual regulatory mechanism suggests that auxin plays a multifaceted role in the complex regulatory web governing seed longevity.
2.5.3. Influence of Gibberellins (GA) on Seed Longevity
GAs are known for their prominent role in triggering seed germination and subsequent growth. Comparative analyses of higher longevity (HL) and lower longevity (LL) varieties after natural aging have led to the identification of specific long-lived mRNAs in rice, including the gibberellin receptor gene GID1. Seeds store various long-lived mRNAs, some of which are crucial for the early stages of germination and, consequently, for seed longevity. These findings suggest that gibberellin signaling plays a role in seed longevity [89]. Genetic analysis has shown that overexpression of ARABIDOPSIS THALIANA HOMEOBOX 25 (AtHB25) in an Arabidopsis ‘activation tagging’ line collection resulted in elevated levels of active gibberellins and increased transcripts of the gibberellin biosynthesis gene GIBBERELLIN 3-OXIDASE 2 [46]. This augmented gibberellin activity was correlated with improved resistance to controlled deterioration tests (CDT). Of note, GA3-treated plants and the quintuple DELLA mutant, characterized by persistent gibberellin responses, displayed stronger CDT resistance, pointing to a potentially positive role of gibberellins in enhancing seed longevity [46]. However, conflicting evidence arises from mutants like ga1-3, which is defective in gibberellin synthesis, and the gibberellin-insensitive gai mutant, neither of which showed decreased germination following prolonged dry storage when compared to wild-type plants [90]. This inconsistency highlights that while there is suggestive evidence for gibberellins’ participation in seed longevity, more research is needed to definitively establish their precise role.
Despite the wealth of research affirming ABA’s role in seed longevity, the contribution of other hormones remains less clear-cut [91]. Studies have shown that mutants resistant to ethylene and jasmonic acid do not significantly lose viability after long-term storage, implying a limited role for these hormones in regulating longevity [83]. On a separate note, recent findings indicate that brassinosteroids (BR) might negatively affect seed longevity during the priming process, a controlled treatment designed to improve germination performance [92]. Seeds from BR-deficient mutants such as cyp85a1/a2 and det2 demonstrate prolonged longevity post-priming in Arabidopsis [92], suggesting a possible connection between BR signaling and seed longevity, an area that merits further investigation.
2.6. Seed Dormancy and Longevity: Positive and Negative Correlations
Seed dormancy and longevity are both critical traits for plant survival and agricultural productivity. Ideally, a favorable correlation between these traits would benefit both natural ecosystems and crop cultivation. However, studies have reported both positive and negative correlations between seed dormancy and longevity [93].
The testa of higher plant seeds protects the embryo against adverse environmental conditions. Mutations in the TRANSPARENT TESTA genes have been shown to decrease seed dormancy and longevity in Arabidopsis due to increased seed coat permeability and altered flavonoid deposition [93]. Disruptions in cutin biosynthesis and deposition, caused by mutations in genes such as GPAT4/8, DCR, LACS2, or BDG1, also compromise seed dormancy and longevity in Arabidopsis [94]. Mutants in the VTE1 gene, which is involved in tocopherol-mediated antioxidant activity, exhibit decreased levels of both dormancy and longevity in Arabidopsis [95]. Interestingly, a loss-of-function mutation in RBOHD, which encodes an NADPH oxidase, results in increased dormancy and longevity in Arabidopsis compared to the wild type [45,96]. Additionally, using a tetragenic system, researchers discovered that natural genes controlling seed dormancy are also involved in the regulation of soil seed bank longevity in rice [97]. These findings suggest a positive correlation between seed dormancy and longevity.
Higher-order DELLA mutants in a gibberellin-deficient background (ga1-3) exhibit reduced dormancy due to the constitutive activation of gibberellin signaling in Arabidopsis [98]. Conversely, the DELLA quintuple mutant demonstrates increased resistance to accelerated aging, likely attributable to enhanced seed coat mucilage production [46]. Disruption of CYP707A1/A2, which are involved in ABA catabolism, results in enhanced dormancy but reduced longevity in Arabidopsis [99]. Similarly, the aspartic protease ASPG1, which is responsible for seed reserve mobilization, shows increased dormancy but decreased longevity in mutants with reduced proteolytic activity [100]. Auxin biosynthesis mutants, such as taa1, tar1, and yuc1, display reduced dormancy but increased longevity in Arabidopsis [88]. Additionally, quantitative trait locus (QTL) analyses in recombinant inbred line populations have revealed a negative correlation between seed dormancy and seed longevity in Arabidopsis [14]. These findings suggest that ABA catabolism and auxin biosynthesis play pivotal roles in the observed negative correlation between seed dormancy and longevity.
In summary, existing genetic analyses have demonstrated both positive and negative correlations between seed dormancy and longevity. However, it is challenging to interpret these contradictory findings, given that few studies have explicitly examined the relationship between these two traits. Nonetheless, based on the limited research available, it is possibly reasonable to hypothesize that the regulation of seed dormancy and longevity involves both shared and independent signaling pathways.
3. Environmental Regulation of Seed Longevity
Seed longevity is intricately influenced by a multitude of environmental factors in conjunction with genetic determinants [101,102] (Figure 1). The environment experienced by the maternal plant during seed maturation, along with the conditions following harvest and throughout storage, plays a critical role in dictating seed viability [17,103]. Key environmental parameters that significantly affect seed longevity include temperature, humidity, light exposure, and oxygen concentration [104]. Soil attributes, such as pH levels and mineral content, also have profound effects on seed survival within the soil seed bank [105,106]. Additionally, the seed microbiome, composed of endophytes and pathogens, subtly adjusts the seed microenvironment and defense mechanisms, thus influencing seed longevity [107]. Our understanding of seed longevity in cultivated crops primarily stems from studies employing wet aging conditions to assess seed vigor. By contrast, a broader range of dry storage conditions has been applied to wild species. Despite this, our understanding of how environmental factors interact with molecular regulators of longevity remains incomplete.
3.1. Influence of Temperature on Seed Longevity
Storage temperature primarily impacts seed longevity through its modulation of enzymatic activities within the seed [108]. Elevated temperatures and moisture levels intensify seed metabolism, whereas lower temperatures modify the phenylpropanoid composition and permeability of the seed coat, as observed in Arabidopsis mutants transparent testa and reduce longevity [93]. Consequently, precise temperature management is essential to optimize seed longevity during storage. The temperature-sensitive DOG1, a key regulator of seed dormancy in Arabidopsis, also influences seed longevity [40]. DOG1 promotes seed longevity by upregulating a variety of genes, including HSPs and LEAs, partly through the activation of ABI5 expression and in concert with ABI3 signaling [109]. Moreover, DOG1 protein levels fluctuate with seed maturation temperature, and the loss of DOG1 function hampers seed dormancy induction at low maturation temperatures [110]. It is plausible that DOG1 orchestrates seed longevity responses to temperature via similar mechanisms.
Furthermore, the effect of temperature on seed longevity varies across species and genotypes [13,105,111]. Warm temperatures during seed development tend to boost longevity in alpine species and Arabidopsis, yet they might be detrimental to rice and Medicago truncatula [108,112]. On the other hand, low temperatures during seed development might diminish longevity in Arabidopsis, but do not seem to have a noticeable effect on Medicago truncatula, potentially due to species-specific adaptations [108].
3.2. Water Availability
Water availability has a nuanced and context-dependent impact on seed longevity, which varies according to species and the intensity of water stress [113]. Soybean seeds experiencing water stress during maturation produce mature green seeds that correlate with decreased longevity [114]. Drought similarly reduces seed longevity in Medicago truncatula, even without an obvious chlorophyll retention phenotype [108]. However, in Brassica rapa, withholding irrigation during early seed filling stages actually accelerates the accrual of seed longevity, enabling longer viability under dry storage [104]. Peanuts exemplify the species-specific response where drought stress during seed development can lead to increased longevity [101,113].
The effect of water availability on seed longevity is complex and stage-dependent. Developing seeds possess a remarkable ability to adjust their maturation processes in response to water availability, thereby mitigating potential losses in longevity [113]. Field experiments on wheat, simulating varying rainfall patterns across growth stages, demonstrate the adaptability of seed development in response to water availability [115]. Field experiments on wheat, simulating rainfall fluctuations at different developmental stages, demonstrate the plasticity of seed development programs in response to water availability [116]. An increase in seed water content due to wetting during development can decrease post-harvest longevity, yet allowing seeds to re-dry naturally restores much of this lost longevity [116], underscoring the intricate interaction among water availability, seed development, and longevity.
Although the exact molecular mechanisms behind water-regulated seed longevity are not fully understood, ABA signaling pathways appear to play a pivotal role in mediating seed maturation and dormancy under water stress [117]. Further exploration into these pathways is expected to shed light on the intricate regulatory networks that govern seed longevity in variable environmental contexts.
3.3. Light Exposure
During seed filling, embryos actively absorb 20-30% of light, particularly enriched in green and far-red wavelengths [118]. Photosynthesis is essential for energy production required for seed reserve accumulation, with chloroplasts adapting their pigment composition and photosystem activity to cope with shade conditions [104]. As seeds mature and gain longevity, chloroplasts disintegrate, and chlorophyll molecules undergo specialized degradation mechanisms differing from those seen in senescent leaves [101,119].
To ensure longevity, it’s crucial to manage photochemical reactions during chloroplast breakdown to avoid the build-up of toxic compounds [120]. Photoperiod and light intensity during seed development both influence seed longevity, with evidence pointing to their regulatory roles in pathways related to longevity [14]. In Arabidopsis, light perception involves genes underlying the DOG3 and DOG6 loci, hinting at a genetic basis for light-mediated longevity control [117]. Future research focused on cloning and characterizing these genes will likely uncover more about the intricate interplay between light perception and seed longevity regulation.
3.4. Nutrient Supply
Nutrient availability in the soil and the mother plant’s nutritional state profoundly influences seed yield and germination traits, reflecting a sophisticated interplay between genetics and environmental elements like temperature and light [13,101]. Various plant species, including tomato, Arabidopsis, and oilseed rape, provide evidence for the impact of nitrate, phosphate, and sulfate availability on seed traits [104]. Although the direct connection between nutrient availability and seed longevity has been less studied, new evidence suggests a potential link.
In barley, seeds harvested from plants grown under optimal nutrient conditions demonstrated enhanced longevity compared to those from nutrient-limited environments [101]. Nitrogen availability has been found to affect seed longevity in Arabidopsis, with higher nitrate levels corresponding to longer seed lifespans [121]. Changes in amino acid and glucoronate contents, along with alterations in gene transcripts linked to cell wall metabolism, emphasize the impact of nutrient availability on seed composition and longevity [122]. Metabolic sensors like the Target of Rapamycin (TOR) complex and SnRK1 complex are crucial for integrating nutrient and energy signals to regulate seed development and longevity [123]. Mutant plants lacking these sensors exhibit decreased resistance to aging, highlighting the importance of metabolic sensing pathways in regulating seed longevity [101]. Thus, a deeper understanding of the intricate balance between nutrient availability, metabolic sensing, and seed longevity necessitates further investigation into the underlying molecular mechanisms.
3.5. Oxygen Level
Oxygen levels during storage significantly affect seed longevity by modulating the formation of reactive oxygen species (ROS) and consequent oxidative damage to macromolecules [124]. High oxygen levels are associated with increased chromosomal abnormalities and reductions in seed viability and vigor [125,126]. Research on Vicia faba and soybean seeds stored under elevated oxygen pressures demonstrates the harmful effects of high oxygen levels, leading to rapid loss of germination capacity [127].
On the other hand, reduced oxygen levels can extend seed longevity during storage. Ultra-dried Brassica seeds maintained viability for over three decades when kept in a modified atmosphere with lowered oxygen levels [128]. These results highlight the significance of oxygen regulation for maintaining seed viability during storage and point to the promise of modified atmosphere storage techniques for seed preservation.
4. Strategies to Enhance Seed Longevity
Current research highlights that seed longevity is determined not only by external environmental influences but also by intricate genetic factors [84]. The revelation of numerous genes deeply involved in seed longevity pathways raises the possibility of engineering seed varieties with enhanced longevity by targeting specific genetic elements. Additionally, considering that seeds often encounter suboptimal storage conditions that undermine their viability, exploring feasible methods to restore vigor to aged seeds is crucial for agriculture, laboratory research, and plant biodiversity conservation.
4.1. Extension of Seed Longevity Through Molecular Genetics
Genetic regulation plays a significant role in controlling seed longevity, thus offering a route to manipulate this trait by altering gene expression using molecular genetics or genome editing technologies. Researchers have probed the molecular foundations of seed longevity, pinpointing essential genes (SEED LONGEVITY-ASSOCIATED GENE, SLAG) and pathways. For example, genes involved in antioxidant defense systems, such as SUPEROXIDE DISMUTASE (SOD), CATALASE (CAT), and PEROXIDASES (POD), help mitigate oxidative stress and preserve seed viability during storage [129]. Genes responsible for synthesizing and managing storage compounds, like LEAs and HSPs, have also been shown to significantly affect seed longevity [130].
With recent breakthroughs in genome editing, such as CRISPR-Cas9, scientists can now make precise adjustments to the genetic makeup of seeds. For example, editing the FUSCA3 (FUS3) gene, which acts as a key regulator of seed maturation and longevity [44,131,132,133], has proven effective in enhancing seed storability and germination vigor in Arabidopsis. Similarly, CRISPR/Cas9-mediated knockout of the LIPOXYGENASE 10 (OsLOX10) gene in rice led to increased seed longevity compared to wild-type under artificial aging conditions [134]. Knockout of type-II metacaspase (MCA-II) proteases via CRISPR in Arabidopsis disrupts proteostasis in seeds, thereby compromising seed longevity [74]. Leveraging molecular genetics and genome editing opens up the possibility of designing seeds with improved longevity characteristics tailored to specific environmental conditions or storage regimens. However, more research is needed to fully understand the complex genetic networks that govern seed longevity and to optimize the application of these techniques in crop improvement projects. Additionally, it’s crucial to remember that gene editing can have unexpected consequences due to the complex nature of gene function. For example, although a knockout of the RBOHD gene can positively impact seed dormancy and longevity [45,96], it may also lead to undesirable phenotypes like reduced growth rate and weakened immunity in rbohd mutants [135].
4.2. Extending Seed Longevity Through Seed Priming
Seed priming, a technique that partially activates germination processes without fully initiating germination, is known to improve seed performance and enhance stress tolerance in crop plants [136,137,138,139,140,141,142]. This process aids in facilitating cellular repair mechanisms, which contributes to improved seedling vigor and crop yield [51]. Studies on leek (Allium porrum) and Brassica oleracea seeds have shown that priming is linked to heightened rates of DNA synthesis and accelerated cell division, leading to more rapid germination and quicker establishment of seedlings [143,144]. Moreover, priming induces changes in gene expression, such as the upregulation of DNA repair pathways and the increased activity of the protein repair enzyme L-ISOASPARTYL METHYLTRANSFERASE [145]. These molecular adjustments help reduce chromosomal abnormalities and enhance the overall quality of germination [51].
However, priming can occasionally diminish the storability or longevity of seeds. For example, the beneficial effects of seed priming were evident only for the first 15 days of storage at 25°C in rice [146]. Beyond this period, the performance of the primed seeds declined, becoming even poorer than that of the non-primed seeds. The detrimental effects of storing the primed seeds at 25°C were associated with impaired starch metabolism within the rice seeds [146,147]. To tackle this challenge, researchers have developed an innovative priming method aimed at enhancing seed survival rates and maintaining seed longevity through the use of biologically active compounds, tested using Arabidopsis seeds [148]. Their findings indicated that priming with cell cycle inhibitors, such as mimosine, aphidicolin, hydroxyurea, and oryzalin, significantly improved both the survival rate and storability of seeds [148]. This suggests that the progression of the cell cycle during priming serves as a critical checkpoint affecting seed storability. By modulating this checkpoint through the inhibition of cell cycle progression, it may be possible to develop priming methods that preserve seed longevity while simultaneously boosting other aspects of seed performance.
4.3. Revitalizing Old Seeds
Despite the importance of optimal storage, real-world constraints such as infrastructure limitations, natural disasters, and logistical issues can hinder the provision of perfect seed storage environments. This is especially true for rare or experimentally conserved germplasm that may have suffered from subpar storage conditions, reducing their viability. Therefore, developing methods to revive even a portion of these seeds carries immense value.
Aged or poorly stored seeds often suffer from energy depletion, reduced enzyme activities, and hypoxic conditions during germination [101,149], necessitating targeted interventions to restore vital components. Hydrogen peroxide, for instance, has been successfully used to resuscitate four-year-old squash (Cucurbita pepo) seeds by acting as an oxygen supplement [150]. Beyond direct chemical treatments, in vitro tissue culture techniques present a promising avenue to rescue the germination potential of immature and aged cucurbit seeds. This method extracts embryos from deteriorating seeds and cultivates them in a nutrient-rich, sterile environment conducive to germination, with success partially hinging on the selection, concentration, and synergistic combination of specific plant growth regulators. Ethylene supplementation, for example, has been shown to expedite germination in aged Brassica napus seeds [151], while cucumber seed regeneration has been effectively achieved with the combined use of 1-naphthaleneacetic acid and 6-benzylaminopurine [152,153]. Low doses of epibrassinolide have also been found to improve germination rates in pepper (Capsicum annuum) seeds [154]. GA3 holds potential for enhancing seed germination when applied externally, though its effectiveness is highly dependent on dosage - lower concentrations can stimulate germination, while higher amounts can inhibit it [155,156] Interestingly, alternative gibberellin compounds like GA4/7 may outperform GA3 in promoting cucurbit seed germination, suggesting potential benefits over conventional GA3 usage [155]. Notably, the Zn-specific chelator TPEN (N, N, N’, N’-Tetrakis (2-pyridylmethyl) ethylenediamine) can significantly delay the aging process of the seeds by regulating the levels of glutathione [157], suggesting that free metal ions released due to the loss of membrane integrity may be both a consequence and a contributing factor to seed aging.
While rescue strategies play a vital role in rejuvenating aged seeds, the fundamental importance of proper seed storage cannot be underestimated. To optimally preserve laboratory seeds, meticulous attention must be paid to keeping seed density low in centrifuge tubes, using hermetically sealed containers for cold storage, maintaining ideal relative humidity levels to prevent moisture damage (Weigel and Glazebrook, 2002). When it comes to preserving laboratory seeds, optimal storage necessitates meticulous attention to several key aspects: first, ensuring minimal seed density within centrifuge tubes, thereby minimizing the risk of accelerated deterioration; second, employing hermetically sealed containers for long-term preservation in cold environments, and maintaining a relative humidity level to prevent moisture-induced damage, and incorporating silica gel pellets to act as a desiccant, absorbing excess moisture and prolonging seed lifespan and viability (Weigel and Glazebrook, 2002). Moreover, the strategic integration of silica gel pellets into the storage system serves as an effective desiccant measure, which actively absorbs excess moisture and thereby extends the lifespan and viability of the seeds (Weigel and Glazebrook, 2002). By combining stringent storage practices with these revitalization techniques, researchers can maximize the likelihood of aged seeds germinating successfully while safeguarding against the multiple factors contributing to seed deterioration and loss of vigor over time.
5. Challenges, Questions and Approaches
While advancements in biotechnology and molecular biology have led to significant insights into the mechanisms governing seed vigor and longevity, numerous challenges persist in translating these discoveries into practical applications. One of the primary obstacles in enhancing seed vigor and longevity is the complexity of the underlying biological processes. These processes involve intricate networks of genetic, metabolic, and environmental interactions that are not yet fully understood [13]. For instance, the role of ROS in seed aging is well-documented, but the precise mechanisms by which ROS damage cellular components and the ways to mitigate this damage remain areas of ongoing research [62]. Another challenge is the variability among different plant species and genotypes. Seeds of various species exhibit different sensitivities to environmental stressors and storage conditions, making it difficult to develop universal strategies for improving seed quality [158]. Additionally, there is a need for more comprehensive understanding of how abiotic stresses, such as temperature and humidity, interact with seed physiology to affect longevity [101]. Moreover, economic constraints and the lack of standardized protocols for seed testing and evaluation pose significant barriers to progress[159]. There is a continuous need for cost-effective and reliable methods to assess seed quality, which can be applied globally, from small-scale farmers to large agribusinesses [160,161].
To address these challenges, an interdisciplinary approach is required, combining genetics, biochemistry, and agronomy with biotechnology and precision agriculture [13,162]. Strategies could include the use of genetic engineering to introduce or enhance protective mechanisms against ROS, the development of species-specific storage guidelines based on detailed physiological studies, and the creation of robust seed quality assessment tools [160]. Furthermore, global collaboration and sharing of resources and knowledge could accelerate the discovery of effective solutions.
While enhancing seed longevity offers clear benefits, such as improved storage stability and extended viability, it is essential to consider the potential downsides. One significant concern is the possible trade-off between seed longevity and germination vigor. Seeds engineered for extended shelf life might exhibit reduced field performance due to alterations in metabolic pathways that affect germination efficiency and early seedling establishment [2]. Additionally, increasing seed longevity could inadvertently select for traits that delay germination, potentially leading to uneven crop stands and reduced uniformity in planting populations, which are detrimental to crop management and yield consistency [14,84].
Furthermore, the genetic modifications required to achieve enhanced longevity might have unintended consequences on plant health and susceptibility to diseases. For instance, changes in seed composition could impact the plant’s natural defense mechanisms against pathogens and pests [163]. Lastly, there is an ecological consideration; seeds with extended viability might persist in the soil longer, potentially outcompeting native species and disrupting local ecosystems if they are not properly managed [164].
Author Contributions
Conceptualization, Z.L.; writing—original draft preparation, S.T., J.C. and S.L.; writing—reviewing and editing, S.T. and Z.L.; supervision, Z.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was generously supported by several funding sources, namely the National Natural Science Foundation of China (32470367 and 32170345 to Z.L.), the Beijing Municipal Natural Science Foundation (5232015 to Z.L.), the Open Research Fund of the esteemed National Center for Protein Sciences at Peking University in Beijing (KF-202304), and the High-level Talent Introduction Programme of Beijing Forestry University.
Data Availability Statement
All the data in this review paper are available online.
Acknowledgments
We sincerely apologize to those authors whose valuable contributions were unable to be included in this review due to space limitations.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| SLAG | SEED LONGEVITY-ASSOCIATED GENE |
| ABI3 | ABSCISIC ACID-INSENSITIVE3 |
| TIP3;1 | TONOPLAST INTRINSIC PROTEIN 3;1 |
| ROF1 | ROTAMASE FKBP 1 |
| DREB2 | DROUGHT RESPONSIVE ELEMENT BINDING FACTOR 2 |
| IAA27 | AUXIN-RESPONSIVE PROTEIN 27 |
| DOG1 | DELAY OF GERMINATION |
| COG1 | COGWHEEL1 |
| CDF4 | CYCLING DOF FACTOR 4 |
| PER1A | PEROXIDASE 1A |
| ATM | ATAXIA TELANGIECTASIA MUTATED |
| ATR | ATM AND RAD3-RELATED |
| SOG1 | SUPPRESSOR OF GAMMA 1 |
| LIG4 | DNA LIGASE 4 |
| XRCC2 | X-RAY REPAIR CROSS COMPLEMENTING 2 |
| PARP1 | POLY(ADP-RIBOSE) POLYMERASE 1 |
| ERCC1 | EXCISION REPAIR CROSS COMPLEMENT-ING-GROUP 1 |
| 8-OXOG | 8-OXOGUANINE |
| DSBS | double-strand breaks |
| HR | homologous recombination |
| ROS | Reactive oxygen species |
| SnRK2 | SUCROSE NON-FERMENTING 1-RELATED PROTEIN KINASE 2 |
| ABA | abscisic acid |
| EM1 | EARLY METHIONINE1 |
| AtHB25 | ARABIDOPSIS THALIANA HOMEOBOX 25 |
| TOR | Target of Rapamycin |
| SOD | SUPEROXIDE DISMUTASE |
| CAT | CATALASE |
| POD | PEROXIDASES |
References
- Sano, N.; Rajjou, L.; North, H.M.; Debeaujon, I.; Marion-Poll, A.; Seo, M. Staying Alive: Molecular Aspects of Seed Longevity. Plant Cell Physiol 2016, 57, 660–674. [Google Scholar] [CrossRef] [PubMed]
- Reed, R.C.; Bradford, K.J.; Khanday, I. Seed germination and vigor: ensuring crop sustainability in a changing climate. Heredity (Edinb) 2022, 128, 450–459. [Google Scholar] [CrossRef]
- Ramtekey, V.; Cherukuri, S.; Kumar, S.; V, S.K.; Sheoran, S.; K, U.B.; K, B.N.; Kumar, S.; Singh, A.N.; Singh, H.V. Seed Longevity in Legumes: Deeper Insights Into Mechanisms and Molecular Perspectives. Front Plant Sci 2022, 13, 918206. [Google Scholar] [CrossRef] [PubMed]
- Guan, B.; Gao, J.; Chen, W.; Gong, X.; Ge, G. The Effects of Climate Change on Landscape Connectivity and Genetic Clusters in a Small Subtropical and Warm-Temperate Tree. Front Plant Sci 2021, 12, 671336. [Google Scholar] [CrossRef]
- Walters, C.; Pence, V.C. The unique role of seed banking and cryobiotechnologies in plant conservation. Plants People Planet 2021, 3, 83–91. [Google Scholar] [CrossRef]
- Saatkamp, A.; Cochrane, A.; Commander, L.; Guja, L.K.; Jimenez-Alfaro, B.; Larson, J.; Nicotra, A.; Poschlod, P.; Silveira, F.A.O.; Cross, A.T.; et al. A research agenda for seed-trait functional ecology. New Phytol 2019, 221, 1764–1775. [Google Scholar] [CrossRef]
- Rajjou, L.; Debeaujon, I. Seed longevity: survival and maintenance of high germination ability of dry seeds. C R Biol 2008, 331, 796–805. [Google Scholar] [CrossRef]
- Buitink, J.; Leprince, O. Intracellular glasses and seed survival in the dry state. C R Biol 2008, 331, 788–795. [Google Scholar] [CrossRef] [PubMed]
- Sallon, S.; Solowey, E.; Cohen, Y.; Korchinsky, R.; Egli, M.; Woodhatch, I.; Simchoni, O.; Kislev, M. Germination, genetics, and growth of an ancient date seed. Science 2008, 320, 1464. [Google Scholar] [CrossRef]
- Shen-Miller, J.; Lindner, P.; Xie, Y.; Villa, S.; Wooding, K.; Clarke, S.G.; Loo, R.R.; Loo, J.A. Thermal-stable proteins of fruit of long-living Sacred Lotus Nelumbo nucifera Gaertn var. China Antique. Trop Plant Biol 2013, 6. [Google Scholar] [CrossRef] [PubMed]
- Lerman, J.C.; Cigliano, E.M. New carbon-14 evidence for six hundred years old Canna compacta seed. Nature 1971, 232, 568–570. [Google Scholar] [CrossRef]
- Brown, K. Botany. Patience yields secrets of seed longevity. Science 2001, 291, 1884–1885. [Google Scholar] [CrossRef] [PubMed]
- Arif, M.A.R.; Afzal, I.; Borner, A. Genetic Aspects and Molecular Causes of Seed Longevity in Plants-A Review. Plants (Basel) 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.P.; Keizer, P.; van Eeuwijk, F.; Smeekens, S.; Bentsink, L. Natural variation for seed longevity and seed dormancy are negatively correlated in Arabidopsis. Plant Physiol 2012, 160, 2083–2092. [Google Scholar] [CrossRef] [PubMed]
- Nagel, M.; Kodde, J.; Pistrick, S.; Mascher, M.; Borner, A.; Groot, S.P. Barley Seed Aging: Genetics behind the Dry Elevated Pressure of Oxygen Aging and Moist Controlled Deterioration. Front Plant Sci 2016, 7, 388. [Google Scholar] [CrossRef]
- Guzzon, F.; Gianella, M.; Velazquez Juarez, J.A.; Sanchez Cano, C.; Costich, D.E. Seed longevity of maize conserved under germplasm bank conditions for up to 60 years. Ann Bot 2021, 127, 775–785. [Google Scholar] [CrossRef] [PubMed]
- Agacka-Moldoch, M.; Arif, M.A.; Lohwasser, U.; Doroszewska, T.; Qualset, C.O.; Borner, A. The inheritance of wheat grain longevity: a comparison between induced and natural ageing. J Appl Genet 2016, 57, 477–481. [Google Scholar] [CrossRef] [PubMed]
- Schwember, A.R.; Bradford, K.J. Quantitative trait loci associated with longevity of lettuce seeds under conventional and controlled deterioration storage conditions. J Exp Bot 2010, 61, 4423–4436. [Google Scholar] [CrossRef] [PubMed]
- Arif, M.A.; Nagel, M.; Lohwasser, U.; Borner, A. Genetic architecture of seed longevity in bread wheat (Triticum aestivum L.). J Biosci 2017, 42, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Gao, Y.; Lin, C.; Pan, R.; Ma, W.; Zheng, Y.; Guan, Y.; Hu, J. Suppression of LOX activity enhanced seed vigour and longevity of tobacco (Nicotiana tabacum L.) seeds during storage. Conserv Physiol 2018, 6, coy047. [Google Scholar] [CrossRef] [PubMed]
- Lang, S.; Liu, X.; Xue, H.; Li, X.; Wang, X. Functional characterization of BnHSFA4a as a heat shock transcription factor in controlling the re-establishment of desiccation tolerance in seeds. J Exp Bot 2017, 68, 2361–2375. [Google Scholar] [CrossRef]
- Lee, J.S.; Velasco-Punzalan, M.; Pacleb, M.; Valdez, R.; Kretzschmar, T.; McNally, K.L.; Ismail, A.M.; Cruz, P.C.S.; Sackville Hamilton, N.R.; Hay, F.R. Variation in seed longevity among diverse Indica rice varieties. Ann Bot 2019, 124, 447–460. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, Y.; Li, S.; Tan, S.; Cao, J.; Wang, H.L.; Luo, J.; Guo, H.; Zhang, Z.; Li, Z. Leaf Senescence Database v5.0: A Comprehensive Repository for Facilitating Plant Senescence Research. J Mol Biol, 6853. [Google Scholar] [CrossRef]
- Kaufmann, K.; Pajoro, A.; Angenent, G.C. Regulation of transcription in plants: mechanisms controlling developmental switches. Nat Rev Genet 2010, 11, 830–842. [Google Scholar] [CrossRef]
- Ng, D.W.; Abeysinghe, J.K.; Kamali, M. Regulating the Regulators: The Control of Transcription Factors in Plant Defense Signaling. Int J Mol Sci 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.Y.; Kagale, S.; Ferrie, A.M.R. Multifaceted roles of transcription factors during plant embryogenesis. Front Plant Sci 2023, 14, 1322728. [Google Scholar] [CrossRef] [PubMed]
- Clerkx, E.J.; El-Lithy, M.E.; Vierling, E.; Ruys, G.J.; Blankestijn-De Vries, H.; Groot, S.P.; Vreugdenhil, D.; Koornneef, M. Analysis of natural allelic variation of Arabidopsis seed germination and seed longevity traits between the accessions Landsberg erecta and Shakdara, using a new recombinant inbred line population. Plant Physiol 2004, 135, 432–443. [Google Scholar] [CrossRef] [PubMed]
- Parcy, F.; Valon, C.; Kohara, A.; Misera, S.; Giraudat, J. The ABSCISIC ACID-INSENSITIVE3, FUSCA3, and LEAFY COTYLEDON1 loci act in concert to control multiple aspects of Arabidopsis seed development. Plant Cell 1997, 9, 1265–1277. [Google Scholar] [CrossRef] [PubMed]
- Monke, G.; Altschmied, L.; Tewes, A.; Reidt, W.; Mock, H.P.; Baumlein, H.; Conrad, U. Seed-specific transcription factors ABI3 and FUS3: molecular interaction with DNA. Planta 2004, 219, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Kotak, S.; Vierling, E.; Baumlein, H.; von Koskull-Doring, P. A novel transcriptional cascade regulating expression of heat stress proteins during seed development of Arabidopsis. Plant Cell 2007, 19, 182–195. [Google Scholar] [CrossRef]
- Mao, Z.; Sun, W. Arabidopsis seed-specific vacuolar aquaporins are involved in maintaining seed longevity under the control of ABSCISIC ACID INSENSITIVE 3. J Exp Bot 2015, 66, 4781–4794. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhu, Y.; Tang, L.; Wang, Y.; Sun, R.; Deng, X. Arabidopsis HSFA9 acts as a regulator of heat response gene expression and the acquisition of thermotolerance and seed longevity. Plant Cell Physiol 2023. [Google Scholar] [CrossRef] [PubMed]
- Meiri, D.; Breiman, A. Arabidopsis ROF1 (FKBP62) modulates thermotolerance by interacting with HSP90.1 and affecting the accumulation of HsfA2-regulated sHSPs. Plant J 2009, 59, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Almoguera, C.; Personat, J.M.; Prieto-Dapena, P.; Jordano, J. Heat shock transcription factors involved in seed desiccation tolerance and longevity retard vegetative senescence in transgenic tobacco. Planta 2015, 242, 461–475. [Google Scholar] [CrossRef]
- Verdier, J.; Lalanne, D.; Pelletier, S.; Torres-Jerez, I.; Righetti, K.; Bandyopadhyay, K.; Leprince, O.; Chatelain, E.; Vu, B.L.; Gouzy, J.; et al. A regulatory network-based approach dissects late maturation processes related to the acquisition of desiccation tolerance and longevity of Medicago truncatula seeds. Plant Physiol 2013, 163, 757–774. [Google Scholar] [CrossRef]
- Almoguera, C.; Prieto-Dapena, P.; Diaz-Martin, J.; Espinosa, J.M.; Carranco, R.; Jordano, J. The HaDREB2 transcription factor enhances basal thermotolerance and longevity of seeds through functional interaction with HaHSFA9. BMC Plant Biol 2009, 9, 75. [Google Scholar] [CrossRef] [PubMed]
- Carranco, R.; Espinosa, J.M.; Prieto-Dapena, P.; Almoguera, C.; Jordano, J. Repression by an auxin/indole acetic acid protein connects auxin signaling with heat shock factor-mediated seed longevity. Proc Natl Acad Sci U S A 2010, 107, 21908–21913. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Sun, H. DOF transcription factors: Specific regulators of plant biological processes. Front Plant Sci 2023, 14, 1044918. [Google Scholar] [CrossRef]
- Ninoles, R.; Ruiz-Pastor, C.M.; Arjona-Mudarra, P.; Casan, J.; Renard, J.; Bueso, E.; Mateos, R.; Serrano, R.; Gadea, J. Transcription Factor DOF4.1 Regulates Seed Longevity in Arabidopsis via Seed Permeability and Modulation of Seed Storage Protein Accumulation. Front Plant Sci 2022, 13, 915184. [Google Scholar] [CrossRef] [PubMed]
- Bentsink, L.; Jowett, J.; Hanhart, C.J.; Koornneef, M. Cloning of DOG1, a quantitative trait locus controlling seed dormancy in Arabidopsis. Proc Natl Acad Sci U S A 2006, 103, 17042–17047. [Google Scholar] [CrossRef] [PubMed]
- Carrillo-Barral, N.; Rodriguez-Gacio, M.D.C.; Matilla, A.J. Delay of Germination-1 (DOG1): A Key to Understanding Seed Dormancy. Plants (Basel) 2020, 9. [Google Scholar] [CrossRef]
- Renard, J.; Martinez-Almonacid, I.; Sonntag, A.; Molina, I.; Moya-Cuevas, J.; Bissoli, G.; Munoz-Bertomeu, J.; Faus, I.; Ninoles, R.; Shigeto, J.; et al. PRX2 and PRX25, peroxidases regulated by COG1, are involved in seed longevity in Arabidopsis. Plant Cell Environ 2020, 43, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Bueso, E.; Munoz-Bertomeu, J.; Campos, F.; Martinez, C.; Tello, C.; Martinez-Almonacid, I.; Ballester, P.; Simon-Moya, M.; Brunaud, V.; Yenush, L.; et al. Arabidopsis COGWHEEL1 links light perception and gibberellins with seed tolerance to deterioration. Plant J 2016, 87, 583–596. [Google Scholar] [CrossRef] [PubMed]
- Righetti, K.; Vu, J.L.; Pelletier, S.; Vu, B.L.; Glaab, E.; Lalanne, D.; Pasha, A.; Patel, R.V.; Provart, N.J.; Verdier, J.; et al. Inference of Longevity-Related Genes from a Robust Coexpression Network of Seed Maturation Identifies Regulators Linking Seed Storability to Biotic Defense-Related Pathways. Plant Cell 2015, 27, 2692–2708. [Google Scholar] [CrossRef]
- Renard, J.; Ninoles, R.; Martinez-Almonacid, I.; Gayubas, B.; Mateos-Fernandez, R.; Bissoli, G.; Bueso, E.; Serrano, R.; Gadea, J. Identification of novel seed longevity genes related to oxidative stress and seed coat by genome-wide association studies and reverse genetics. Plant Cell Environ 2020, 43, 2523–2539. [Google Scholar] [CrossRef] [PubMed]
- Bueso, E.; Munoz-Bertomeu, J.; Campos, F.; Brunaud, V.; Martinez, L.; Sayas, E.; Ballester, P.; Yenush, L.; Serrano, R. ARABIDOPSIS THALIANA HOMEOBOX25 uncovers a role for Gibberellins in seed longevity. Plant Physiol 2014, 164, 999–1010. [Google Scholar] [CrossRef]
- Gregis, V.; Sessa, A.; Colombo, L.; Kater, M.M. AGL24, SHORT VEGETATIVE PHASE, and APETALA1 redundantly control AGAMOUS during early stages of flower development in Arabidopsis. Plant Cell 2006, 18, 1373–1382. [Google Scholar] [CrossRef]
- Pereira Neto, L.G.; Rossini, B.C.; Marino, C.L.; Toorop, P.E.; Silva, E.A.A. Comparative Seeds Storage Transcriptome Analysis of Astronium fraxinifolium Schott, a Threatened Tree Species from Brazil. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Wang, W.Q.; Xu, D.Y.; Sui, Y.P.; Ding, X.H.; Song, X.J. A multiomic study uncovers a bZIP23-PER1A-mediated detoxification pathway to enhance seed vigor in rice. Proc Natl Acad Sci U S A 2022, 119. [Google Scholar] [CrossRef] [PubMed]
- Cheah, K.S.; Osborne, D.J. DNA lesions occur with loss of viability in embryos of ageing rye seed. Nature 1978, 272, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Waterworth, W.; Balobaid, A.; West, C. Seed longevity and genome damage. Biosci Rep 2024, 44. [Google Scholar] [CrossRef] [PubMed]
- Waterworth, W.M.; Footitt, S.; Bray, C.M.; Finch-Savage, W.E.; West, C.E. DNA damage checkpoint kinase ATM regulates germination and maintains genome stability in seeds. Proc Natl Acad Sci U S A 2016, 113, 9647–9652. [Google Scholar] [CrossRef]
- Waterworth, W.M.; Latham, R.; Wang, D.; Alsharif, M.; West, C.E. Seed DNA damage responses promote germination and growth in Arabidopsis thaliana. Proc Natl Acad Sci U S A 2022, 119, e2202172119. [Google Scholar] [CrossRef]
- Waterworth, W.M.; Masnavi, G.; Bhardwaj, R.M.; Jiang, Q.; Bray, C.M.; West, C.E. A plant DNA ligase is an important determinant of seed longevity. Plant J 2010, 63, 848–860. [Google Scholar] [CrossRef]
- Chen, H.; Chu, P.; Zhou, Y.; Li, Y.; Liu, J.; Ding, Y.; Tsang, E.W.; Jiang, L.; Wu, K.; Huang, S. Overexpression of AtOGG1, a DNA glycosylase/AP lyase, enhances seed longevity and abiotic stress tolerance in Arabidopsis. J Exp Bot 2012, 63, 4107–4121. [Google Scholar] [CrossRef]
- Taylor, R.E.; Waterworth, W.; West, C.E.; Foyer, C.H. WHIRLY proteins maintain seed longevity by effects on seed oxygen signalling during imbibition. Biochem J 2023, 480, 941–956. [Google Scholar] [CrossRef]
- Kaye, J.A.; Melo, J.A.; Cheung, S.K.; Vaze, M.B.; Haber, J.E.; Toczyski, D.P. DNA breaks promote genomic instability by impeding proper chromosome segregation. Curr Biol 2004, 14, 2096–2106. [Google Scholar] [CrossRef]
- Culligan, K.M.; Robertson, C.E.; Foreman, J.; Doerner, P.; Britt, A.B. ATR and ATM play both distinct and additive roles in response to ionizing radiation. Plant J 2006, 48, 947–961. [Google Scholar] [CrossRef]
- Falck, J.; Coates, J.; Jackson, S.P. Conserved modes of recruitment of ATM, ATR and DNA-PKcs to sites of DNA damage. Nature 2005, 434, 605–611. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Kim, J.H.; Kim, J.; Lyu, J.I.; Zhang, Y.; Guo, H.; Nam, H.G.; Woo, H.R. ATM suppresses leaf senescence triggered by DNA double-strand break through epigenetic control of senescence-associated genes in Arabidopsis. New Phytol 2020, 227, 473–484. [Google Scholar] [CrossRef]
- Haber, J.E. Partners and pathwaysrepairing a double-strand break. Trends Genet 2000, 16, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Jeevan Kumar, S.P.; Rajendra Prasad, S.; Banerjee, R.; Thammineni, C. Seed birth to death: dual functions of reactive oxygen species in seed physiology. Ann Bot 2015, 116, 663–668. [Google Scholar] [CrossRef]
- Stadtman, E.R.; Berlett, B.S. Reactive oxygen-mediated protein oxidation in aging and disease. Chem Res Toxicol 1997, 10, 485–494. [Google Scholar] [CrossRef]
- Yermolaieva, O.; Xu, R.; Schinstock, C.; Brot, N.; Weissbach, H.; Heinemann, S.H.; Hoshi, T. Methionine sulfoxide reductase A protects neuronal cells against brief hypoxia/reoxygenation. Proc Natl Acad Sci U S A 2004, 101, 1159–1164. [Google Scholar] [CrossRef]
- Moskovitz, J.; Berlett, B.S.; Poston, J.M.; Stadtman, E.R. The yeast peptide-methionine sulfoxide reductase functions as an antioxidant in vivo. Proc Natl Acad Sci U S A 1997, 94, 9585–9589. [Google Scholar] [CrossRef] [PubMed]
- Hazra, A.; Varshney, V.; Verma, P.; Kamble, N.U.; Ghosh, S.; Achary, R.K.; Gautam, S.; Majee, M. Methionine sulfoxide reductase B5 plays a key role in preserving seed vigor and longevity in rice (Oryza sativa). New Phytol 2022, 236, 1042–1060. [Google Scholar] [CrossRef] [PubMed]
- Chatelain, E.; Satour, P.; Laugier, E.; Ly Vu, B.; Payet, N.; Rey, P.; Montrichard, F. Evidence for participation of the methionine sulfoxide reductase repair system in plant seed longevity. Proc Natl Acad Sci U S A 2013, 110, 3633–3638. [Google Scholar] [CrossRef] [PubMed]
- Buitink, J.; Leger, J.J.; Guisle, I.; Vu, B.L.; Wuilleme, S.; Lamirault, G.; Le Bars, A.; Le Meur, N.; Becker, A.; Kuster, H.; et al. Transcriptome profiling uncovers metabolic and regulatory processes occurring during the transition from desiccation-sensitive to desiccation-tolerant stages in Medicago truncatula seeds. Plant J 2006, 47, 735–750. [Google Scholar] [CrossRef]
- Castellión, M.; Matiacevich, S.; Buera, P.; Maldonado, S. Protein deterioration and longevity of quinoa seeds during long-term storage. Food Chem 2010, 121, 952–958. [Google Scholar] [CrossRef]
- Oge, L.; Bourdais, G.; Bove, J.; Collet, B.; Godin, B.; Granier, F.; Boutin, J.P.; Job, D.; Jullien, M.; Grappin, P. Protein repair L-isoaspartyl methyltransferase 1 is involved in both seed longevity and germination vigor in Arabidopsis. Plant Cell 2008, 20, 3022–3037. [Google Scholar] [CrossRef] [PubMed]
- Mudgett, M.B.; Lowenson, J.D.; Clarke, S. Protein repair L-isoaspartyl methyltransferase in plants. Phylogenetic distribution and the accumulation of substrate proteins in aged barley seeds. Plant Physiol 1997, 115, 1481–1489. [Google Scholar] [CrossRef] [PubMed]
- Petla, B.P.; Kamble, N.U.; Kumar, M.; Verma, P.; Ghosh, S.; Singh, A.; Rao, V.; Salvi, P.; Kaur, H.; Saxena, S.C.; et al. Rice PROTEIN l-ISOASPARTYL METHYLTRANSFERASE isoforms differentially accumulate during seed maturation to restrict deleterious isoAsp and reactive oxygen species accumulation and are implicated in seed vigor and longevity. New Phytol 2016, 211, 627–645. [Google Scholar] [CrossRef]
- Verma, P.; Kaur, H.; Petla, B.P.; Rao, V.; Saxena, S.C.; Majee, M. PROTEIN L-ISOASPARTYL METHYLTRANSFERASE2 is differentially expressed in chickpea and enhances seed vigor and longevity by reducing abnormal isoaspartyl accumulation predominantly in seed nuclear proteins. Plant Physiol 2013, 161, 1141–1157. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Hatzianestis, I.H.; Pfirrmann, T.; Reza, S.H.; Minina, E.A.; Moazzami, A.; Stael, S.; Gutierrez-Beltran, E.; Pitsili, E.; Dormann, P.; et al. Seed longevity is controlled by metacaspases. Nat Commun 2024, 15, 6748. [Google Scholar] [CrossRef]
- Salvi, P.; Saxena, S.C.; Petla, B.P.; Kamble, N.U.; Kaur, H.; Verma, P.; Rao, V.; Ghosh, S.; Majee, M. Differentially expressed galactinol synthase(s) in chickpea are implicated in seed vigor and longevity by limiting the age induced ROS accumulation. Sci Rep 2016, 6, 35088. [Google Scholar] [CrossRef]
- Li, T.; Zhang, Y.; Wang, D.; Liu, Y.; Dirk, L.M.A.; Goodman, J.; Downie, A.B.; Wang, J.; Wang, G.; Zhao, T. Regulation of Seed Vigor by Manipulation of Raffinose Family Oligosaccharides in Maize and Arabidopsis thaliana. Mol Plant 2017, 10, 1540–1555. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Qi, J.; Hao, G.; Zhang, C.; Wang, C.; Dirk, L.M.A.; Downie, A.B.; Zhao, T. ZmDREB1A Regulates RAFFINOSE SYNTHASE Controlling Raffinose Accumulation and Plant Chilling Stress Tolerance in Maize. Plant Cell Physiol 2020, 61, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Salvi, P.; Varshney, V.; Majee, M. Raffinose family oligosaccharides (RFOs): role in seed vigor and longevity. Biosci Rep 2022, 42. [Google Scholar] [CrossRef]
- de Souza Vidigal, D.; Willems, L.; van Arkel, J.; Dekkers, B.J.W.; Hilhorst, H.W.M.; Bentsink, L. Galactinol as marker for seed longevity. Plant Sci 2016, 246, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Vinson, C.C.; Mota, A.P.Z.; Porto, B.N.; Oliveira, T.N.; Sampaio, I.; Lacerda, A.L.; Danchin, E.G.J.; Guimaraes, P.M.; Williams, T.C.R.; Brasileiro, A.C.M. Characterization of raffinose metabolism genes uncovers a wild Arachis galactinol synthase conferring tolerance to abiotic stresses. Sci Rep 2020, 10, 15258. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, D.; Dirk, L.M.A.; Downie, A.B.; Zhao, T. ZmAGA1 Hydrolyzes RFOs Late during the Lag Phase of Seed Germination, Shifting Sugar Metabolism toward Seed Germination Over Seed Aging Tolerance. J Agric Food Chem 2021, 69, 11606–11615. [Google Scholar] [CrossRef] [PubMed]
- Locascio, A.; Roig-Villanova, I.; Bernardi, J.; Varotto, S. Current perspectives on the hormonal control of seed development in Arabidopsis and maize: a focus on auxin. Frontiers in Plant Science 2014, 5. [Google Scholar] [CrossRef]
- Clerkx, E.J.; Vries, H.B.; Ruys, G.J.; Groot, S.P.; Koornneef, M. Characterization of green seed, an enhancer of abi3-1 in Arabidopsis that affects seed longevity. Plant Physiol 2003, 132, 1077–1084. [Google Scholar] [CrossRef] [PubMed]
- Rehmani, M.S.; Aziz, U.; Xian, B.; Shu, K. Seed Dormancy and Longevity: A Mutual Dependence or a Trade-Off? Plant Cell Physiol 2022, 63, 1029–1037. [Google Scholar] [CrossRef]
- Finkelstein, R.R.; Gampala, S.S.; Rock, C.D. Abscisic acid signaling in seeds and seedlings. Plant Cell 2002, 14 Suppl, S15–45. [Google Scholar] [CrossRef]
- Chen, K.; Li, G.J.; Bressan, R.A.; Song, C.P.; Zhu, J.K.; Zhao, Y. Abscisic acid dynamics, signaling, and functions in plants. J Integr Plant Biol 2020, 62, 25–54. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, H.; Zhao, Y.; Feng, Z.; Li, Q.; Yang, H.Q.; Luan, S.; Li, J.; He, Z.H. Auxin controls seed dormancy through stimulation of abscisic acid signaling by inducing ARF-mediated ABI3 activation in Arabidopsis. Proc Natl Acad Sci U S A 2013, 110, 15485–15490. [Google Scholar] [CrossRef] [PubMed]
- Pellizzaro, A.; Neveu, M.; Lalanne, D.; Ly Vu, B.; Kanno, Y.; Seo, M.; Leprince, O.; Buitink, J. A role for auxin signaling in the acquisition of longevity during seed maturation. New Phytol 2020, 225, 284–296. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Wang, S.; Tang, Y.; Jiang, L.; He, W.; Lin, Q.; Yu, F.; Wang, L. Transcriptome-Wide Characterization of Seed Aging in Rice: Identification of Specific Long-Lived mRNAs for Seed Longevity. Front Plant Sci 2022, 13, 857390. [Google Scholar] [CrossRef]
- Debeaujon, I.; Koornneef, M. Gibberellin requirement for Arabidopsis seed germination is determined both by testa characteristics and embryonic abscisic acid. Plant Physiol 2000, 122, 415–424. [Google Scholar] [CrossRef]
- Pirredda, M.; Fananas-Pueyo, I.; Onate-Sanchez, L.; Mira, S. Seed Longevity and Ageing: A Review on Physiological and Genetic Factors with an Emphasis on Hormonal Regulation. Plants (Basel) 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Sano, N.; Kim, J.S.; Onda, Y.; Nomura, T.; Mochida, K.; Okamoto, M.; Seo, M. RNA-Seq using bulked recombinant inbred line populations uncovers the importance of brassinosteroid for seed longevity after priming treatments. Sci Rep 2017, 7, 8095. [Google Scholar] [CrossRef]
- Debeaujon, I.; Leon-Kloosterziel, K.M.; Koornneef, M. Influence of the testa on seed dormancy, germination, and longevity in Arabidopsis. Plant Physiol 2000, 122, 403–414. [Google Scholar] [CrossRef]
- De Giorgi, J.; Piskurewicz, U.; Loubery, S.; Utz-Pugin, A.; Bailly, C.; Mene-Saffrane, L.; Lopez-Molina, L. An Endosperm-Associated Cuticle Is Required for Arabidopsis Seed Viability, Dormancy and Early Control of Germination. PLoS Genet 2015, 11, e1005708. [Google Scholar] [CrossRef]
- Sattler, S.E.; Gilliland, L.U.; Magallanes-Lundback, M.; Pollard, M.; DellaPenna, D. Vitamin E is essential for seed longevity and for preventing lipid peroxidation during germination. Plant Cell 2004, 16, 1419–1432. [Google Scholar] [CrossRef]
- Leymarie, J.; Vitkauskaite, G.; Hoang, H.H.; Gendreau, E.; Chazoule, V.; Meimoun, P.; Corbineau, F.; El-Maarouf-Bouteau, H.; Bailly, C. Role of reactive oxygen species in the regulation of Arabidopsis seed dormancy. Plant Cell Physiol 2012, 53, 96–106. [Google Scholar] [CrossRef]
- Pipatpongpinyo, W.; Korkmaz, U.; Wu, H.; Kena, A.; Ye, H.; Feng, J.; Gu, X.Y. Assembling seed dormancy genes into a system identified their effects on seedbank longevity in weedy rice. Heredity (Edinb) 2020, 124, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Penfield, S.; Gilday, A.D.; Halliday, K.J.; Graham, I.A. DELLA-mediated cotyledon expansion breaks coat-imposed seed dormancy. Curr Biol 2006, 16, 2366–2370. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, M.; Kuwahara, A.; Seo, M.; Kushiro, T.; Asami, T.; Hirai, N.; Kamiya, Y.; Koshiba, T.; Nambara, E. CYP707A1 and CYP707A2, which encode abscisic acid 8′-hydroxylases, are indispensable for proper control of seed dormancy and germination in Arabidopsis. Plant Physiol 2006, 141, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Yao, X.; Ye, T.; Ma, S.; Liu, X.; Yin, X.; Wu, Y. Arabidopsis Aspartic Protease ASPG1 Affects Seed Dormancy, Seed Longevity and Seed Germination. Plant Cell Physiol 2018, 59, 1415–1431. [Google Scholar] [CrossRef] [PubMed]
- Zinsmeister, J.; Leprince, O.; Buitink, J. Molecular and environmental factors regulating seed longevity. Biochem J 2020, 477, 305–323. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Chen, F.; Luo, X.; Dai, Y.; Yang, Y.; Zheng, C.; Yang, W.; Shu, K. A matter of life and death: Molecular, physiological, and environmental regulation of seed longevity. Plant Cell Environ 2020, 43, 293–302. [Google Scholar] [CrossRef]
- Wiebach, J.; Nagel, M.; Borner, A.; Altmann, T.; Riewe, D. Age-dependent loss of seed viability is associated with increased lipid oxidation and hydrolysis. Plant Cell Environ 2020, 43, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Walters, C.; Ballesteros, D.; Vertucci, V.A. Structural mechanics of seed deterioration: Standing the test of time. Plant Science 2010, 179, 565–573. [Google Scholar] [CrossRef]
- Probert, R.J.; Daws, M.I.; Hay, F.R. Ecological correlates of ex situ seed longevity: a comparative study on 195 species. Ann Bot 2009, 104, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Daws, M.I.; Kabadajic, A.; Manger, K.; Kranner, I. Extreme thermo-tolerance in seeds of desert succulents is related to maximum annual temperature. S Afr J Bot 2007, 73, 262–265. [Google Scholar] [CrossRef]
- Nelson, E.B. The seed microbiome: Origins, interactions, and impacts. Plant Soil 2018, 422, 7–34. [Google Scholar] [CrossRef]
- Ramtekey, V.; Cherukuri, S.; Kumar, S.; Sripathy, K.V.; Sheoran, S.; Udaya, B.K.; Bhojaraja, N.K.; Kumar, S.; Singh, A.N.; Singh, H.V. Seed Longevity in Legumes: Deeper Insights Into Mechanisms and Molecular Perspectives. Frontiers in Plant Science 2022, 13. [Google Scholar] [CrossRef]
- Dekkers, B.J.; He, H.; Hanson, J.; Willems, L.A.; Jamar, D.C.; Cueff, G.; Rajjou, L.; Hilhorst, H.W.; Bentsink, L. The Arabidopsis DELAY OF GERMINATION 1 gene affects ABSCISIC ACID INSENSITIVE 5 (ABI5) expression and genetically interacts with ABI3 during Arabidopsis seed development. Plant J 2016, 85, 451–465. [Google Scholar] [CrossRef]
- Murphey, M.; Kovach, K.; Elnacash, T.; He, H.Z.; Bentsink, L.; Donohue, K. -imposed dormancy mediates germination responses to temperature cues. Environ Exp Bot 2015, 112, 33–43. [Google Scholar] [CrossRef]
- Walters, C.; Wheeler, L.M.; Grotenhuis, J.M. Longevity of seeds stored in a genebank: species characteristics. Seed Sci Res 2005, 15, 1–20. [Google Scholar] [CrossRef]
- Bernareggi, G.; Carbognani, M.; Petraglia, A.; Mondoni, A. Climate warming could increase seed longevity of alpine snowbed plants. Alpine Bot 2015, 125, 69–78. [Google Scholar] [CrossRef]
- Reed, R.C.; Bradford, K.J.; Khanday, I. Seed germination and vigor: ensuring crop sustainability in a changing climate. Heredity 2022, 128, 450–459. [Google Scholar] [CrossRef]
- Rao, P.J.M.; Pallavi, M.; Bharathi, Y.; Priya, P.B.; Sujatha, P.; Prabhavathi, K. Insights into mechanisms of seed longevity in soybean: a review. Frontiers in Plant Science 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros, D.; Pritchard, H.W.; Walters, C. Dry architecture: towards the understanding of the variation of longevity in desiccation-tolerant germplasm. Seed Sci Res 2020, 30, 142–155. [Google Scholar] [CrossRef]
- Ellis, R.H.; Yadav, G. Effect of simulated rainfall during wheat seed development and maturation on subsequent seed longevity is reversible. Seed Sci Res 2016, 26, 67–76. [Google Scholar] [CrossRef]
- Weitbrecht, K.; Muller, K.; Leubner-Metzger, G. First off the mark: early seed germination. J Exp Bot 2011, 62, 3289–3309. [Google Scholar] [CrossRef] [PubMed]
- Shackira, A.M.; Sarath, N.G.; Aswathi, K.P.R.; Pardha-Saradhi, P.; Puthur, J.T. Green seed photosynthesis: What is it? What do we know about it? Where to go? Plant Physiology Reports 2022, 27, 573–579. [Google Scholar] [CrossRef]
- Pereira Lima, J.J.; Buitink, J.; Lalanne, D.; Rossi, R.F.; Pelletier, S.; da Silva, E.A.A.; Leprince, O. Molecular characterization of the acquisition of longevity during seed maturation in soybean. PLoS One 2017, 12, e0180282. [Google Scholar] [CrossRef]
- Griffo, A.; Bosco, N.; Pagano, A.; Balestrazzi, A.; Macovei, A. Noninvasive Methods to Detect Reactive Oxygen Species as a Proxy of Seed Quality. Antioxidants (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Nagel, M.; Kranner, I.; Neumann, K.; Rolletschek, H.; Seal, C.E.; Colville, L.; Fernandez-Marin, B.; Borner, A. Genome-wide association mapping and biochemical markers reveal that seed ageing and longevity are intricately affected by genetic background and developmental and environmental conditions in barley. Plant Cell Environ 2015, 38, 1011–1022. [Google Scholar] [CrossRef] [PubMed]
- He, H.; de Souza Vidigal, D.; Snoek, L.B.; Schnabel, S.; Nijveen, H.; Hilhorst, H.; Bentsink, L. Interaction between parental environment and genotype affects plant and seed performance in Arabidopsis. J Exp Bot 2014, 65, 6603–6615. [Google Scholar] [CrossRef] [PubMed]
- Rosnoblet, C.; Aubry, C.; Leprince, O.; Vu, B.L.; Rogniaux, H.; Buitink, J. The regulatory gamma subunit SNF4b of the sucrose non-fermenting-related kinase complex is involved in longevity and stachyose accumulation during maturation of Medicago truncatula seeds. Plant J 2007, 51, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.P.J.; Chintagunta, A.D.; Reddy, Y.M.; Rajjou, L.; Garlapati, V.K.; Agarwal, D.K.; Prasad, S.R.; Simal-Gandara, J. Implications of reactive oxygen and nitrogen species in seed physiology for sustainable crop productivity under changing climate conditions. Curr Plant Biol 2021, 26. [Google Scholar] [CrossRef]
- Buijs, G.; Willems, L.A.J.; Kodde, J.; Groot, S.P.C.; Bentsink, L. Evaluating the EPPO method for seed longevity analyses in Arabidopsis. Plant Sci 2020, 301, 110644. [Google Scholar] [CrossRef] [PubMed]
- Fleming, M.B.; Hill, L.M.; Walters, C. The kinetics of ageing in dry-stored seeds: a comparison of viability loss and RNA degradation in unique legacy seed collections. Ann Bot 2019, 123, 1133–1146. [Google Scholar] [CrossRef] [PubMed]
- Rao, P.J.M.; Pallavi, M.; Bharathi, Y.; Priya, P.B.; Sujatha, P.; Prabhavathi, K. Insights into mechanisms of seed longevity in soybean: a review. Front Plant Sci 2023, 14, 1206318. [Google Scholar] [CrossRef] [PubMed]
- González-Benito, M.E.; Pérez-García, F.; Tejeda, G.; Gómez-Campo, C. Effect of the gaseous environment and water content on seed viability of four Brassicaceae species after 36 years storage. Seed Sci Technol 2011, 39, 443–451. [Google Scholar] [CrossRef]
- Bailly, C.; El-Maarouf-Bouteau, H.; Corbineau, F. From intracellular signaling networks to cell death: the dual role of reactive oxygen species in seed physiology. C R Biol 2008, 331, 806–814. [Google Scholar] [CrossRef]
- Battaglia, M.; Olvera-Carrillo, Y.; Garciarrubio, A.; Campos, F.; Covarrubias, A.A. The enigmatic LEA proteins and other hydrophilins. Plant Physiol 2008, 148, 6–24. [Google Scholar] [CrossRef]
- Wang, F.; Perry, S.E. Identification of direct targets of FUSCA3, a key regulator of Arabidopsis seed development. Plant Physiol 2013, 161, 1251–1264. [Google Scholar] [CrossRef] [PubMed]
- Sugliani, M.; Rajjou, L.; Clerkx, E.J.; Koornneef, M.; Soppe, W.J. Natural modifiers of seed longevity in the Arabidopsis mutants abscisic acid insensitive3-5 (abi3-5) and leafy cotyledon1-3 (lec1-3). New Phytol 2009, 184, 898–908. [Google Scholar] [CrossRef]
- Roscoe, T.J.; Vaissayre, V.; Paszkiewicz, G.; Clavijo, F.; Kelemen, Z.; Michaud, C.; Lepiniec, L.C.; Dubreucq, B.; Zhou, D.X.; Devic, M. Regulation of FUSCA3 Expression During Seed Development in Arabidopsis. Plant Cell Physiol 2019, 60, 476–487. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Xu, H.; Zhang, L.; Shi, Y.; Song, Y.; Wang, X.; Cai, Q.; He, W.; Xie, H.; Zhang, J. The lipoxygenase OsLOX10 affects seed longevity and resistance to saline-alkaline stress during rice seedlings. Plant Mol Biol 2023, 111, 415–428. [Google Scholar] [CrossRef]
- Lee, D.; Lal, N.K.; Lin, Z.D.; Ma, S.; Liu, J.; Castro, B.; Toruno, T.; Dinesh-Kumar, S.P.; Coaker, G. Regulation of reactive oxygen species during plant immunity through phosphorylation and ubiquitination of RBOHD. Nat Commun 2020, 11, 1838. [Google Scholar] [CrossRef] [PubMed]
- Marthandan, V.; Geetha, R.; Kumutha, K.; Renganathan, V.G.; Karthikeyan, A.; Ramalingam, J. Seed Priming: A Feasible Strategy to Enhance Drought Tolerance in Crop Plants. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, P.; Gupta, M. Micronutrient seed priming: new insights in ameliorating heavy metal stress. Environ Sci Pollut Res Int 2022, 29, 58590–58606. [Google Scholar] [CrossRef]
- Donia, D.T.; Carbone, M. Seed Priming with Zinc Oxide Nanoparticles to Enhance Crop Tolerance to Environmental Stresses. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Nile, S.H.; Thiruvengadam, M.; Wang, Y.; Samynathan, R.; Shariati, M.A.; Rebezov, M.; Nile, A.; Sun, M.; Venkidasamy, B.; Xiao, J.; et al. Nano-priming as emerging seed priming technology for sustainable agriculture-recent developments and future perspectives. J Nanobiotechnology 2022, 20, 254. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, E.A. Seed priming to alleviate salinity stress in germinating seeds. J Plant Physiol 2016, 192, 38–46. [Google Scholar] [CrossRef]
- Paparella, S.; Araujo, S.S.; Rossi, G.; Wijayasinghe, M.; Carbonera, D.; Balestrazzi, A. Seed priming: state of the art and new perspectives. Plant Cell Rep 2015, 34, 1281–1293. [Google Scholar] [CrossRef]
- Rhaman, M.S.; Imran, S.; Rauf, F.; Khatun, M.; Baskin, C.C.; Murata, Y.; Hasanuzzaman, M. Seed Priming with Phytohormones: An Effective Approach for the Mitigation of Abiotic Stress. Plants (Basel) 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Hourston, J.E.; Perez, M.; Gawthrop, F.; Richards, M.; Steinbrecher, T.; Leubner-Metzger, G. The effects of high oxygen partial pressure on vegetable Allium seeds with a short shelf-life. Planta 2020, 251, 105. [Google Scholar] [CrossRef]
- Soeda, Y.; Konings, M.C.; Vorst, O.; van Houwelingen, A.M.; Stoopen, G.M.; Maliepaard, C.A.; Kodde, J.; Bino, R.J.; Groot, S.P.; van der Geest, A.H. Gene expression programs during Brassica oleracea seed maturation, osmopriming, and germination are indicators of progression of the germination process and the stress tolerance level. Plant Physiol 2005, 137, 354–368. [Google Scholar] [CrossRef]
- Forti, C.; Ottobrino, V.; Bassolino, L.; Toppino, L.; Rotino, G.L.; Pagano, A.; Macovei, A.; Balestrazzi, A. Molecular dynamics of pre-germinative metabolism in primed eggplant (Solanum melongena L.) seeds. Hortic Res 2020, 7, 87. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Zheng, M.; Khan, F.; Khaliq, A.; Fahad, S.; Peng, S.; Huang, J.; Cui, K.; Nie, L. Benefits of rice seed priming are offset permanently by prolonged storage and the storage conditions. Sci Rep 2015, 5, 8101. [Google Scholar] [CrossRef] [PubMed]
- Ren, M.; Tan, B.; Xu, J.; Yang, Z.; Zheng, H.; Tang, Q.; Zhang, X.; Wang, W. Priming methods affected deterioration speed of primed rice seeds by regulating reactive oxygen species accumulation, seed respiration and starch degradation. Front Plant Sci 2023, 14, 1267103. [Google Scholar] [CrossRef] [PubMed]
- Sano, N.; Seo, M. Cell cycle inhibitors improve seed storability after priming treatments. J Plant Res 2019, 132, 263–271. [Google Scholar] [CrossRef]
- Finkelstein, R.; Reeves, W.; Ariizumi, T.; Steber, C. Molecular aspects of seed dormancy. Annu Rev Plant Biol 2008, 59, 387–415. [Google Scholar] [CrossRef]
- Liu, G.D.; Porterfield, D.M.; Li, Y.C.; Klassen, W. Increased Oxygen Bioavailability Improved Vigor and Germination of Aged Vegetable Seeds. Hortscience 2012, 47, 1714–1721. [Google Scholar] [CrossRef]
- Takayanagi, K.; Harrington, J.F. Enhancement of germination rate of aged seeds by ethylene. Plant Physiol 1971, 47, 521–524. [Google Scholar] [CrossRef]
- Mazzoni-Putman, S.M.; Brumos, J.; Zhao, C.; Alonso, J.M.; Stepanova, A.N. Auxin Interactions with Other Hormones in Plant Development. Cold Spring Harb Perspect Biol 2021, 13. [Google Scholar] [CrossRef]
- Matilla, A.J. Auxin: Hormonal Signal Required for Seed Development and Dormancy. Plants (Basel) 2020, 9. [Google Scholar] [CrossRef]
- Barboza da Silva, C.; Marcos, J. Storage performance of primed bell pepper seeds with 24-Epibrassinolide. Agron J 2020, 112, 948–960. [Google Scholar] [CrossRef]
- Evensen, K.B.; Loy, J.B. Effects of gibberellic Acid and gold light on germination, enzyme activities, and amino Acid pool size in a dwarf strain of watermelon. Plant Physiol 1978, 62, 6–9. [Google Scholar] [CrossRef]
- Zhao, H.; Zhang, Y.; Zheng, Y. Integration of ABA, GA, and light signaling in seed germination through the regulation of ABI5. Front Plant Sci 2022, 13, 1000803. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, Y.; He, Y.Q.; Ye, T.T.; Huang, X.; Wu, H.; Ma, T.X.; Pritchard, H.W.; Wang, X.F.; Xue, H. Glutathionylation of a glycolytic enzyme promotes cell death and vigor loss during aging of elm seeds. Plant Physiol 2024, 195, 2596–2616. [Google Scholar] [CrossRef] [PubMed]
- T, V.N.; S, R.; R, L.R. Evaluation of diverse soybean genotypes for seed longevity and its association with seed coat colour. Sci Rep 2023, 13, 4313. [Google Scholar] [CrossRef]
- Hay, F.R.; Valdez, R.; Lee, J.S.; Sta Cruz, P.C. Seed longevity phenotyping: recommendations on research methodology. J Exp Bot 2019, 70, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Buitink, J.; Leprince, O. A Seed Storage Protocol to Determine Longevity. Methods Mol Biol 2024, 2830, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Fenollosa, E.; Jene, L.; Munne-Bosch, S. A rapid and sensitive method to assess seed longevity through accelerated aging in an invasive plant species. Plant Methods 2020, 16, 64. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, P.; Pramitha, L.; Aggarwal, P.R.; Rana, S.; Vetriventhan, M.; Muthamilarasan, M. Biotechnological interventions for improving the seed longevity in cereal crops: progress and prospects. Crit Rev Biotechnol 2023, 43, 309–325. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Solanki, M.K.; Wang, Z.; Solanki, A.C.; Singh, V.K.; Divvela, P.K. Revealing the seed microbiome: Navigating sequencing tools, microbial assembly, and functions to amplify plant fitness. Microbiol Res 2024, 279, 127549. [Google Scholar] [CrossRef] [PubMed]
- Moravcova, L.; Carta, A.; Pysek, P.; Skalova, H.; Gioria, M. Long-term seed burial reveals differences in the seed-banking strategies of naturalized and invasive alien herbs. Sci Rep 2022, 12, 8859. [Google Scholar] [CrossRef]
Figure 1.
Influence of Endogenous and Exogenous Signals on Seed Longevity and Underlying Regulatory Mechanisms. Seed longevity is influenced by a variety of environmental cues such as nutrient status, temperature, moisture, and light, as well as internal signals like plant hormones. Endogenous and exogenous stress factors have the ability to trigger diverse types of damage, such as DNA damage and protein damage, which in turn compromise seed longevity or vigor. Multiple plant hormones, such as ABA and GA, regulate seed longevity through modulation of transcription factors. ABA, Abscisic Acid; GA, Gibberellic Acid; BR, Brassinosteroids; ROS, Reactive Oxygen Species; PMIT, Protein-L-isoaspartate (D-aspartate) O-methyltransferase; ATM, ATAXIA TELANGIECTASIA MUTATED; ATR, ATAXIA TELANGIECTASIA AND RAD3-RELATED; CDF, CYCLING DOF FACTOR; COG1, COGWHEEL1; TOR, TARGET OF RAPAMYCIN; DOF, DNA BINDING WITH ONE FINGER; DOG1, Delay of Germination 1; HSFA9, heat shock factor A9; ROF1, ROTAMASE FKBP 1; HSP, heat shock protein; TIP3;1, ALPHA-TONOPLAST INTRINSIC PROTEIN; TIR1, TRANSPORT INHIBITOR RESPONSE 1. The arrow represents promotion, while the vertical line stands for inhibition. P, Phosphorylation.
Figure 1.
Influence of Endogenous and Exogenous Signals on Seed Longevity and Underlying Regulatory Mechanisms. Seed longevity is influenced by a variety of environmental cues such as nutrient status, temperature, moisture, and light, as well as internal signals like plant hormones. Endogenous and exogenous stress factors have the ability to trigger diverse types of damage, such as DNA damage and protein damage, which in turn compromise seed longevity or vigor. Multiple plant hormones, such as ABA and GA, regulate seed longevity through modulation of transcription factors. ABA, Abscisic Acid; GA, Gibberellic Acid; BR, Brassinosteroids; ROS, Reactive Oxygen Species; PMIT, Protein-L-isoaspartate (D-aspartate) O-methyltransferase; ATM, ATAXIA TELANGIECTASIA MUTATED; ATR, ATAXIA TELANGIECTASIA AND RAD3-RELATED; CDF, CYCLING DOF FACTOR; COG1, COGWHEEL1; TOR, TARGET OF RAPAMYCIN; DOF, DNA BINDING WITH ONE FINGER; DOG1, Delay of Germination 1; HSFA9, heat shock factor A9; ROF1, ROTAMASE FKBP 1; HSP, heat shock protein; TIP3;1, ALPHA-TONOPLAST INTRINSIC PROTEIN; TIR1, TRANSPORT INHIBITOR RESPONSE 1. The arrow represents promotion, while the vertical line stands for inhibition. P, Phosphorylation.
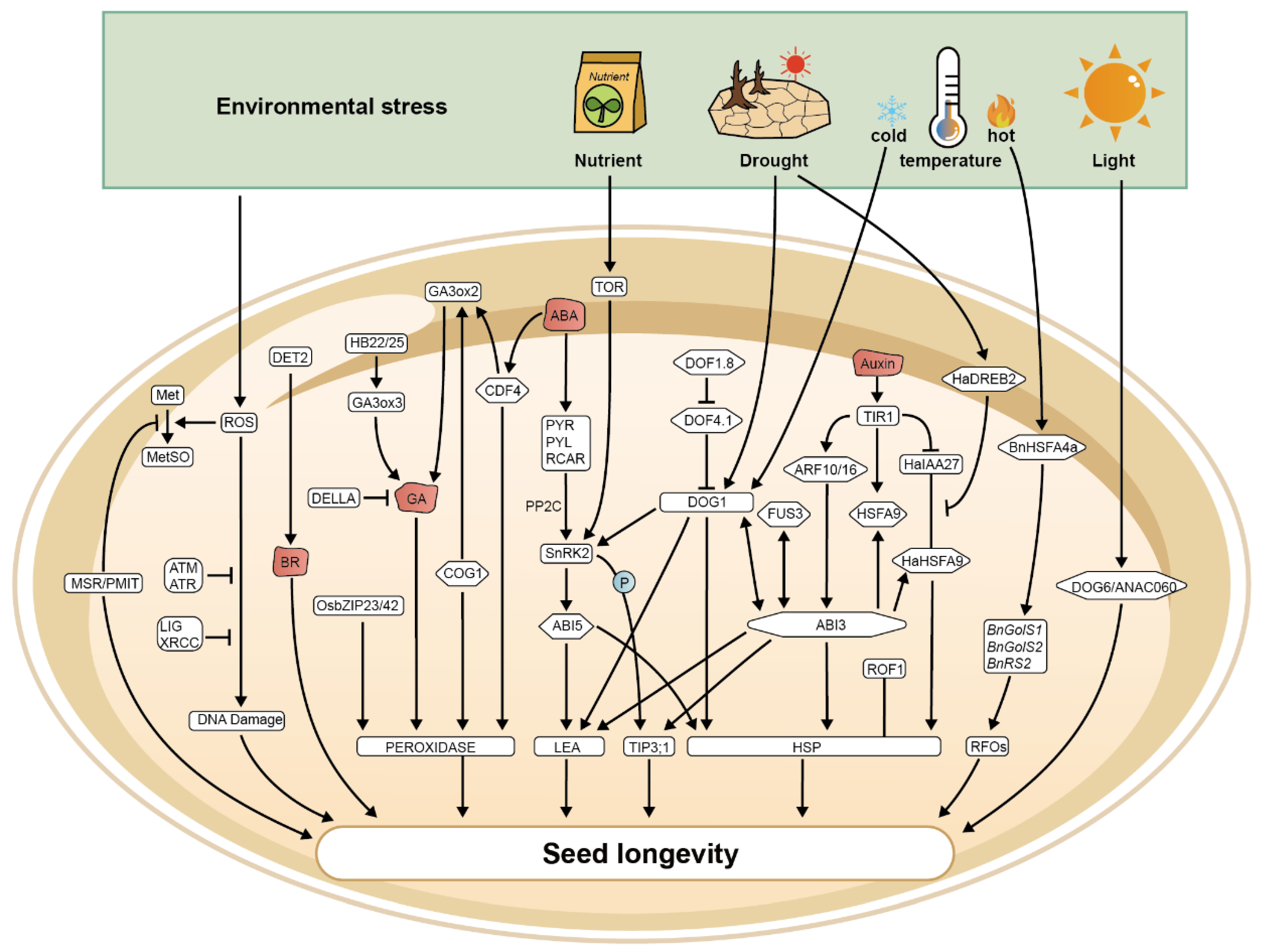

Table 1.
Genes Involved in Regulating Seed Longevity in Arabidopsis.
| Locus | Gene | Effect | Pathway | References (PubMed ID) |
| AT4G13250 | NYC1 | enhance | Chlorophyll degradation | 22751379 |
| AT3G48190 | ATM | decrease | DNA repair | 27503884 |
| AT5G40820 | ATR | decrease | DNA repair | 27503884 |
| AT3G05210 | ERCC1 | enhance | DNA repair | 35858436 |
| AT1G16970 | KU70 | enhance | DNA repair | 35858436 |
| AT5G57160 | LIG4 | enhance | DNA repair | 20584150 |
| AT1G66730 | LIG6 | enhance | DNA repair | 20584150 |
| AT1G25580 | SOG1 | decrease | DNA repair | 35858436 |
| AT1G21710 | OGG1 | enhance | DNA repair | 22473985 |
| AT2G31320 | PARP1 | enhance | DNA repair | 35858436 |
| AT5G22470 | PARP3 | enhance | DNA repair | 24533577 |
| AT1G14410 | WHY1 | enhance | DNA repair | 37351567 |
| AT2G02740 | WHY3 | enhance | DNA repair | 37351567 |
| AT5G64520 | XRCC2 | enhance | DNA repair | 35858436 |
| AT1G34790 | TT1 | enhance | Flavonoid biosynthesis | 10677433 |
| AT5G48100 | TT10 | enhance | Flavonoid biosynthesis | 10677433 |
| AT5G42800 | TT3 | enhance | Flavonoid biosynthesis | 10677433 |
| AT3G55120 | TT5 | enhance | Flavonoid biosynthesis | 10677433 |
| AT5G07990 | TT7 | enhance | Flavonoid biosynthesis | 10677433 |
| AT4G09820 | TT8 | enhance | Flavonoid biosynthesis | 10677433 |
| AT3G28430 | TT9 | enhance | Flavonoid biosynthesis | 10677433 |
| AT3G24650 | ABI3 | enhance | Hormone, ABA | 12231895 |
| AT3G18490 | ASPG1 | enhance | Hormone, ABA | 29648652 |
| AT5G45830 | DOG1 | enhance | Hormone, ABA | 17065317 |
| AT2G36610 | ATHB22 | enhance | Hormone, GA | 24335333 |
| AT5G65410 | ATHB25 | enhance | Hormone, GA | 24335333 |
| AT1G14440 | ATHB31 | enhance | Hormone, GA | 24335333 |
| AT1G80340 | GA3OX2 | enhance | Hormone, GA | 24335333 |
| AT2G01570 | RGA1 | decrease | Hormone, GA | 24335333 |
| AT1G14920 | RGA2 | decrease | Hormone, GA | 24335333 |
| AT1G66350 | RGL1 | decrease | Hormone, GA | 24335333 |
| AT3G03450 | RGL2 | decrease | Hormone, GA | 24335333 |
| AT5G17490 | RGL3 | decrease | Hormone, GA | 24335333 |
| AT1G09570 | PHYA | decrease | Light | 27227784 |
| AT2G18790 | PHYB | decrease | Light | 27227784 |
| AT2G45970 | CYP86A8 | enhance | Lipid biosynthesis | 32519347 |
| AT3G47860 | AtCHL | enhance | Lipid peroxidation | 23837879 |
| AT5G58070 | AtTIL | enhance | Lipid peroxidation | 23837879 |
| AT1G55020 | LOX1 | decrease | Lipid peroxidation | 28371855 |
| AT1G28440 | AtHSL1 | enhance | LRR-RLK | 35763091 |
| AT2G27500 | BG14 | enhance | Metabolism Carbohydrate | 36625794 |
| AT2G47180 | GOLS1 | enhance | Metabolism Galactose | 26993241 |
| AT1G56600 | GOLS2 | enhance | Metabolism Galactose | 26993241 |
| AT1G30370 | AtDLAH | enhance | Metabolism Lipid | 21856645 |
| AT2G19900 | NADP-ME | enhance | Metabolism Malate | 29744896 |
| AT4G15940 | AtFAHD1a | decrease | Metabolism Oxoacid | 33804275 |
| AT4G02770 | PSAD1 | enhance | PHOTOSYSTEM | 32519347 |
| AT1G62710 | β-VPE | enhance | Protein catabolism | 30782971 |
| AT2G26130 | RSL1 | enhance | Protein degradation | 24388521 |
| AT5G45360 | SKIP31 | enhance | Protein degradation | 37462265 |
| AT5G53000 | TAP46 | enhance | Protein dephosphorylation | 25399018 |
| AT3G25230 | ROF1 | enhance | Protein isomerization | 22268595 |
| AT5G48570 | ROF2 | enhance | Protein isomerization | 22268595 |
| AT3G48330 | PIMT1 | enhance | Protein repair | 19011119 |
| AT3G57520 | AtSIP2 | decrease | Raffinose catabolism | 34553917 |
| AT4G02750 | SSTPR | enhance | RNA modification | 32519347 |
| AT1G19570 | DHAR1 | enhance | ROS detoxification | 32519347 |
| AT1G05250 | PRX2 | enhance | ROS detoxification | 31600827 |
| AT2G41480 | PRX25 | enhance | ROS detoxification | 31600827 |
| AT5G64120 | PRX71 | enhance | ROS detoxification | 31600827 |
| AT5G47910 | RBOHD | decrease | ROS production | 32519347 |
| AT1G19230 | RBOHE | decrease | ROS production | 32519347 |
| AT1G64060 | RBOHF | decrease | ROS production | 32519347 |
| AT3G17520 | LEA | enhance | Seed development | 32519347 |
| AT5G44120 | CRUA | enhance | Seed storage protein | 26184996 |
| AT1G03880 | CRUB | enhance | Seed storage protein | 26184996 |
| AT4G28520 | CRUC | enhance | Seed storage protein | 26184996 |
| AT4G36920 | AtAP2 | enhance | TF AP2/EREBP | 10677433 |
| AT5G53210 | SPCH1 | enhance | TF bHLH | 32519347 |
| AT2G34140 | CDF4 | enhance | TF DOF | 27227784 |
| AT1G29160 | COG1 | enhance | TF DOF | 31600827 |
| AT4G00940 | DOF4.1 | decrease | TF DOF | 35845633 |
| AT5G42630 | ATS | enhance | TF G2-LIKE | 10677433 |
| AT1G79840 | GL2 | enhance | TF HB | 10677433 |
| AT1G62990 | KNAT7 | decrease | TF HB | 32519347 |
| AT5G54070 | AtHSFA9 | enhance | TF HSF | 32683703 |
| AT5G15800 | AGL2 | decrease | TF MADS | 32519347 |
| AT1G18710 | MYB47 | enhance | TF MYB | 32519347 |
| AT1G21970 | LEC1 | enhance | TF NF-YB | 19754639 |
| AT2G38470 | WRKY33 | enhance | TF WRKY | 26410298 |
| AT4G32770 | VTE1 | enhance | Tocopherol biosynthesis | 15155886 |
| AT2G18950 | VTE2 | enhance | Tocopherol biosynthesis | 15155886 |
| AT1G73190 | TIP3.1 | enhance | Transmembrane transport | 26019256 |
| AT1G17810 | TIP3.2 | enhance | Transmembrane transport | 26019256 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



