
Submitted:
23 January 2025
Posted:
23 January 2025
You are already at the latest version
Abstract
During the last decade, a new therapeutic revolution has involved the management of hepatocellular carcinoma (HCC). This is made possible thanks to the documented efficacy of immunotherapy for this disease. In addition, new evidence demonstrated the role of the gut-liver axis and gut microbiota in host homeostasis, tumor development, and response to therapies. In particular, intestinal dysbiosis can alter the tumor microenvironment leading to the activation of intracellular signalling pathways that promote carcinogenesis. The composition of gut microbiota proved to influence the immune checkpoint inhibitors (ICIs) efficacy and drug toxicities. Therefore, this review aims to deepen knowledge about the immunomodulatory role of gut microbiota and its possible employment as diagnostic and predictive biomarkers in diagnosis and response to immunotherapy of HCC, respectively.
Keywords:
HCC
; Immunotherapy
; Immune Checkpoints Inhibitors (ICIs)
; Gut Microbiota
1. Introduction
Hepatocellular carcinoma (HCC) is the most common primary liver tumor, corresponding to 75-85% of cases [1,2]. The diagnosis of HCC usually occurs in the advanced stage; this is the main responsible for the poor prognosis despite the new therapeutic revolution that involved HCC thanks to the documented efficacy of immunotherapy [2,3,4]. Generally, HCC develops in a liver microenvironment characterized by cirrhosis and chronic inflammation [5]. In this regard, the most relevant risk factors for HCC are HBV/HCV infection and heavy alcohol consumption which cause chronic liver inflammation [1,6]. On the other hand, the occurrence of HCC in healthy liver is becoming more common, above all in the Western world [6]. In this regard, non-alcoholic fatty liver disease (NAFLD) due to obesity has proven to be the starting point of HCC [7].
The gut microbiota consists of about a trillion microorganisms, including bacteria, viruses, protozoa, and fungi. Therefore, it is an extremely diversified micro-ecosystem, whose composition is strictly dependent on external factors [8,9,10]. Several researchers have studied the relationship between the development of HCC and intestinal dysbiosis, a condition in which atypical bacterial metabolites are produced [11,12]. This association is made possible through the gut-liver axis that consists of the biliary tract, hepatic portal vein, and biliary secretions. In this way, gut bacteria and their metabolites can reach the liver leading to hepatic inflammation, tumorigenesis, and cancer progression. Furthermore, recent data identify the gut microbiome as a key player in determining the Immune Checkpoint Inhibitors (ICIs) efficacy in the treatment of HCC [13,14,15].
This review aims to deepen knowledge about the different compositions of gut microbiota and their influence on the response to ICIs in patients affected by advanced HCC. Moreover, we explored the possibility of modulating the features of gut microbiota to obtain better therapeutic efficacy.
Immunotherapy and HCC
For many decades, HCC has been a pathology without therapeutic options, beyond loco-regional treatments [5,16]. Since 2007 we have witnessed the first therapeutic revolution thanks to the advent of Tyrosine Kinase Inhibitors (TKIs). Sorafenib, in the SHARP study, was the first TKI that demonstrated to improve survival compared to placebo for patients with advanced HCC [17]. Subsequently, other TKIs have become part of the therapeutic landscape of this disease. Among these, Lenvatinib proved to be non-inferior to Sorafenib in the REFLECT study [18]; Cabozantinib and Regorafenib have demonstrated their activity in advanced HCC patients pre-treated with other TKIs in CELESTIAL and REACH trial, respectively [19,20]. Moreover, only very few HCC patients, approximately 2.5% of them, present molecular alterations that might be amenable to targeted therapy [2,3]. However, in the last decade, we have witnessed a new therapeutic revolution due to the proven effectiveness of combination therapies including immunotherapy, in particular Immune Checkpoints Inhibitors (ICIs) [21]. These last drugs correspond to antibodies targeting programmed death-1 (PD-1), programmed death ligand-1 (PD-L1), and cytotoxic T lymphocyte antigen-4 (CTLA-4). PD-1 is a receptor expressed on the cell surface of various immune cells, and it may be up-regulated as a result of T cell activation; PD-L1 and PD-L2 are expressed on target cells and their binding to PD-1 receptor leads to the suppression of immune response favoring cancer growth [22]. CTLA-4 is another receptor with the function of downregulating the immune response [22]. Nowadays, the standard first-line treatments are either Atezolizumab (PD-L1 inhibitor) + Bevacizumab (angiogenesis inhibitor, anti-VEGF monoclonal antibody) or Durvalumab (PD-L1 inhibitor) + Tremelimumab (CTLA-4 inhibitor). The combination of Atezolizumab and Bevacizumab was tested in a phase III clinical trial, IMbrave150, compared to Sorafenib in advanced HCC patients [23]. The results showed a significant improvement in progression-free survival (PFS) and overall survival (OS). In detail, the median PFS was 6.9 versus 4.3 months (hazard ratio [HR] 0.65; 95% CI 0.53-0.81; p < 0.001) while OS was 19.2 and 13.4 months (HR 0.66; 95% CI 0.52-0.85; p <0.001) in experimental and control groups, respectively. Reported adverse events (AEs) of all grades were fewer in the combination group compared to the control one (18). On the other hand, the combination of Durvalumab plus Tremelimumab was analyzed in the phase III study, HIMALAYA, compared to Sorafenib [24]. To be specific, the patients enrolled in this trial were randomized into three groups: 1) STRIDE regimen: Tremelimumab (300 mg one dose) + Durvalumab (1500 mg every 4 weeks); 2) Durvalumab single agent (1500 mg every 4 weeks); 3) Control group: Sorafenib (400 mg twice daily). The results showed a median OS of 16.43 months for the STRIDE regimen, 16.56 months for durvalumab as a single agent, and 13.77 months for the control group. The HR for OS was 0.78 (96.02% CI, 0.65 to 0.93; P=0.0035) in the comparison between STRIDE and sorafenib arms. As regards the other comparison, the OS was non-inferior between Durvalumab monotherapy and Sorafenib groups (HR, 0.86; 95.67% CI, 0.73 to 1.03; noninferiority margin, 1.08). PFS was not significantly different among all three arms. In terms of toxicity, AEs of all grades were fewer in the STRIDE regimen arm with respect to the control arm. At ASCO 2024, the results of the CheckMate-9DW trial were reported [25]. This phase III trial assessed the combination of Nivolumab plus Ipilimumab versus Sorafenib or Lenvatinib as first-line therapy showing a significant improvement in OS (23.7 months versus 20.6 months; HR 0.79, 95% CI [0.65, 0.96]; p = 0.018). Several other ICIs, including PD-1 (Pembrolizumab, Camrelizumab, and Sintilimab), PD-L1 (Atezolizumab), are being investigated in combination with VEGF (Bevacizumab) antibodies and TKIs (Lenvatinib and Cabozantinib) in phase III clinical trials as first-line systemic therapy for advanced HCC. For example, the ORIENT-32 trial evaluated the safety, tolerability, and efficacy of Sintilimab plus IBI305 (Bevacizumab biosimilar) compared to Sorafenib as first-line therapy. The results demonstrated PFS and OS significantly longer for the experimental group [26]. By contrast, the COSMIC-312 trial investigated Cabozantinib plus Atezolizumab compared to Sorafenib while the LEAP-002 tested Pembrolizumab plus Lenvatinib versus Lenvatinib. However, in both cases, the PFS and OS were not significantly longer for the combination groups [27,28]. Patients who are considered ineligible for the standard first-line regimens can be treated with Sorafenib or Lenvatinib [17,18]. ICIs were also tested as single agents for first-line treatment of advanced HCC although without a significant survival benefit. For example, Nivolumab was compared to Sorafenib in the CheckMate 459 trial, a phase III clinical study [29]. Moreover, immunotherapy was evaluated for those patients who did not tolerate or progressed to Sorafenib. In this regard, KEYNOTE-240 was a phase III clinical trial that evaluated Pembrolizumab compared to placebo, in this setting of patients. However, the differences in the PFS and OS between the two arms were not statistically significant [30]. Second-line therapy depends on the regimen that patients have previously received. Patients who were initially treated with the standard first-line combinations should be recruited in clinical trials. In alternative, they can receive Sorafenib or Lenvatinib although no prospective clinical studies have evaluated the efficacy of TKIs after progression to the first-line therapy including ICIs. Cabozantinib is the most common drug employed as a third-line treatment [20]. For those patients that were ineligible for the combination therapies in the first line and received Sorafenib or Lenvatinib, the choice falls on Regorafenib, Cabozantinib, or Ramucirumab (if the AFP level is ≥ 400 ng/mL).
Table 1 summarizes all clinical trials about the employment of ICIs in patients with HCC.
2. Microbiota and Immunotherapy Efficacy
The gut microbiota is a heterogeneous ecosystem of microorganisms that primarily lies in the human gut [31,32]. The gut microbiota composition can be very different among individuals due to genetic, dietary and health factors. However, this diversity and the presence of various microorganisms’ species is fundamental for the maintenance of gut health [33,34,35]. The gut microbiota plays several functions including the digestion of substrates that human beings cannot directly metabolize, such as complex polysaccharides and dietary fibers. Bacteria produce short-chain fatty acids (SCFAs) through the fermentation; the main SCFAs is butyrate which provide energy to epithelial cells of intestine and have anti-inflammatory effects [35,36]. Moreover, the gut microbiota plays a key role in modulating the immune system. In detail, the interactions between intestinal immune cells (such as dendritic cells and lymphocytes) and microbiome are essential for the maintenance of immune tolerance to commensal microorganisms, preventing allergic and autoimmune responses [34]. In addition, the microbiota helps to defend the host from pathogens, limiting their invasion and colonization. Furthermore, some intestinal bacteria can synthetize essential vitamins, such as B vitamins (B1, B2, B6, B12, folates) and vitamin K [31]. The gut microbiota manages the host's metabolism through the accumulation of body fat and the regulation of blood sugar levels. The gut microbiota is also involved in the regulation of the central nervous system (CNS) through the so-called "gut-brain axis." In detail, microbiota can produce neurotransmitters that can directly influence brain function and behavior [37].
Bacteria are the main components of the gut microbiota and are represented by various species. Firmicutes are responsible for the fermentation of dietary fibers and the production of SCFAs. The most representative species of Firmicutes are Lactobacillales and Clostridia which have an important role in food digestion and the immune system modulation. Bacteroidetes are abundant in the human gut and are mainly involved in the digestion of complex polysaccharides. The most common genera include Bacteroides, which are important for the metabolism of non-digestible carbohydrates and for SCFAs production. Actinobacteria includes Bifidobacterium, which can ferment oligosaccharides, produce SCFAs, such as lactate, and inhibit the growth of pathogenic bacteria. Proteobacteria are usually potential pathogens although they are also present in a healthy microbiota. The most important genera include Salmonella and Escherichia (such as E. coli) which are not usually pathogenic but sometimes they can cause infectious disease. Fusobacteria and Verrucomicrobia are represented by a small number of species, but some of them are essential for gut health. Fusobacterium is known for its role in fermentation processes, while Akkermansia muciniphila, belonging to the phylum Verrucomicrobia, is involved in degrading intestinal mucus [31,32,36]. Archaea are microorganisms similar to bacteria but belong to a distinct domain. Although less abundant than bacteria, intestinal Archaea, particularly the genus Methanobrevibacter play an important role in intestinal metabolism. These microorganisms are involved in methane production through a process known as methanogenesis, which helps reduce hydrogen concentration in the intestine and can influence carbohydrate fermentation [31,32,36].
Intestinal fungi are less abundant with respect to bacteria although they are an important component of the gut microbiota. The most common are Candida, Saccharomyces, and Penicillium. Candida albicans is a commensal fungus that can become pathogenic in cases of microbiota imbalance, causing infections. The presence and proliferation of fungi in the gut microbiota are regulated by interactions with bacteria [31,32,36].
The virome of the human gut includes a great variety of viruses, many of which are bacteriophages. These viruses play a key role in maintaining the health microbiota, as they influence bacterial composition by regulating the growth of specific bacterial species [31,32,36].
Protozoa are unicellular organisms present in the human gut microbiota although less abundant than bacteria and fungi. Some protozoa, such as Entamoeba histolytica and Giardia lamblia, can be pathogenic and cause diseases, but other species are commensal and don’t usually cause harm to health. Protozoa can impact the gut microbiota through their interactions with bacteria and the modulation of intestinal inflammation [31,32,36].
Recent data showed that a different microbiota composition is present in ICI-responder and non-responder patients [38,39,40,41]. An example is Akkermansia muciniphila that is abundant in ICI-responder patients; this bacterium can induce a type-1 antitumor response favoring the secretion of interleukin (IL)-12 by dendritic cells (DCs). This results in an enhanced engagement of memory CD4+ T cells. Another component of microbiota, Bacteroides fragilis, is positively associated with immunotherapy efficacy since it can stimulate the activation of interferon (IFN)-γ-producing DCs and CD4+T lymphocytes [42,43]. Bifidobacterium as a single species, but also Ruminococcaceae, Bacteroides, Fusobacterium, and Eubacterium as a mixture of species, have proven to increase ICIs activity through the stimulation of immune responses mediated by IFNγ+ CD8+ T cells [38,39,40,41,42,43,44,45]. ICI responders affected by liver cancer have an enrichment of these species in their gut microbiota [46,47]. Therefore, these bacteria can promote the anticancer immune response through the activation of CD8+ T cells, which have a key role in controlling HCC development [48]. Other data suggest that the metabolites of commensal bacteria can also enhance the efficacy of antitumor treatment via the amplification of immune activity. A metabolomic investigation tested the group of metabolites produced by a specific biological network. In particular, the metabolome assessment from the faecal microbiome or serum can allow to identify those predictive biomarkers useful to foretell the long-term efficacy of immunotherapy. An example corresponds to the SCFAs produced by microbiome because they have the ability to regulate CD8+ T cell activity in ICI responder patients; moreover, a longer PFS is correlated to SCFAs presence [49,50]. Pre-clinical data on a pancreatic cancer model showed that the activity of CD8+ T cells in adoptive cell therapy (ACT) was strongly enhanced by SCFAs leading to improved ant-tumor activity and clinical results [50]. On these bases, the administration of bacteria-producing SCFAs or directly SCFAs might enhance the efficacy of ACT with CD8+T cells in HCC. Furthermore, the addition of SCFAs to an anti-PD-1 ameliorated the effectiveness of immunotherapy in a mouse model of HCC. This was obtained through the reduction of IL-17 level and the consequent suppression of cancer growth [51]. Inosine is a metabolite generally produced by Bifidobacterium species; it proved to improve ICI efficacy via the IFN-γ release and the activation of CD8+T and CD4+ T cells [52]. In contrast, ICI non-responders suffered from a severe gut dysbiosis that inhibits the triggering of type 1 immune response but stimulates immunosuppressive one. Other data demonstrated that the production of IL-10 by DCs is upregulated in ICI non-responder patients affected by HCC; this determines the amplification of T-regulatory lymphocytes (Tregs) with the consequent suppression of anti-cancer immune activity [53,54,55]. These findings suggest the need for therapeutic strategies with the aim to manage the microbiota so favoring an anti-cancer activity and avoiding the occurrence of immune resistance (Figure 1).
3. Microbiome: ICI-Responders and Non-Responders
As mentioned before, many differences in microbiota composition are present in ICI-responder and non-responder patients affected by advanced HCC. Several data support the key role of microbiome composition in the different responses to ICIs. In detail, most of studies included the collection of faecal samples at baseline and 2 months after the beginning of treatment. The enrolled patients in some of these studies had already received other therapies such as Sorafenib [26,29,30,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63].
An experimental study by Min-Woo Chug et al. documented that the microbiota of ICI-responder HCC patients had an increased representation of Enterococcus durans, Azospirillum sp. and Citrobacter freundii. On the other hand, Trichuristrichiura, Granulicatella sp., Lactobacillus reteri, Escherichia coli, Enterococcus faecium, Dialisterpneumosintes, Veillonellaatypica, Streptococcus gordonii, and Streptococcus mutans were the prevailing species in ICI non-responder patients. Moreover, a poorer response to ICIs was observed in patients with an unbalanced ratio of Firmicutes to Bacteroidetes; in contrast, a better response was described for those patients with a high ratio of Prevotella to Bacteroides [63].
Another study by Mao Jinzhu et al. showed different outcomes in 65 patients affected by advanced liver cancers (35 with biliary tract cancer, 30 with HCC) and treated with ICIs. Faecal samples from all patients were collected before the start of therapy. In detail, a great amount of Firmicutes and Bacterioides was observed in patients who experienced a clinical advantage; on the other hand, patients without clinical benefit had higher levels of bacteria from Veillonellales order and Proteobacteria. In addition, the arm with a greater amount of Veillonellaceae experienced a poorer PFS and OS while a longer PFS and OS were described in the arm with a greater amount of Bacteroides zoogleoformans, Alistipes sp. Marseille-P5997, Ruminococcuscallidus, Erysipelotrichaceae bacterium-GAM147, and Lachnospiraceae. Erysipelotrichaceae-GAM147 and Lachnospiraceae-GAM79 are commensal species and protective strains able to produce IL-10 and TGF-beta so improving the role of the epithelial barrier. These species are also implicated in the metabolism of bile acids. The genomic analysis of stool samples collected during therapy reported that the microbiota had maintained stable features in those patients with good response; otherwise, the microbiota of patients with poor response was characterized by great diversity [46].
Zheng et al. conducted a clinical trial on advanced HCC patients who progressed to Sorafenib and received Camrelizumab (an anti-PD1 agent). The design of the study established the collection of stool samples before the beginning of therapy, after 1 week, and every 3 weeks during treatment. The genomic analysis reported that a greater level of microbiome population was present in responder patients. Moreover, a higher concentration of Proteobacteria, Bacteroides, and Firmicutes was described in both responder and non-responder populations although a greater amount of Ruminococcaceae and Akkermansia muciniphila was reported in responders. A significant modification of the bacterial population was evident in non-responders during therapy. In detail, a higher level of Proteobacteria, mostly E. coli, in the period between week 3 and week 12, at which point the first species became prevalent. By contrast, Klebsiella pneumoniae was the most represented among Proteobacteria in the responder population [47].
A study by Li et al. reported an association between PFS and the oral and gut microbiome. In particular, a longer PFS was observed in HCC patients with a higher concentration of Faecalibacterium compared to those with a higher amount of Bacteroidales. Therefore, a higher level of Faecalibacterium might be a peculiarity of ICI-responders while a higher level of Bacteroidales might be a feature of ICI-non-responders [9]. Otherwise, another study by Shen et al. didn’t show any difference in microbiota between responder and non-responder HCC patients both at baseline and regarding the administered type of ICI [57]. However, it is well known that the composition and the features of microbiota are strongly influenced by external factors, such as sex, dietary, environmental, and medication. Moreover, different methods of sampling, sample storage, and analysis could affect the study outcomes.
Table 2 indicates the main bacterial species of the microbiota found in ICI-responder and non-responder HCC patients.
4. Future Perspective and Conclusions
The studies described in the previous sections evidenced the correlation between the microbiome of advanced HCC patients and the response to the treatment with ICIs. Therefore, they suggest that the modulation of microbiome composition might indirectly influence the clinical response to current therapies. Several methods could be employed to modulate the microbiome features such as antibiotics, metformin, aspirin, statins, prebiotics, probiotics, diet, and also faecal transplant [11]. Some data about patients affected by other tumor types showed that clinical outcomes can be affected by antibiotics due to their effect on the microbiome [64]. In this regard, it is well known that the chronic use of antibiotics can determine gut dysbiosis negatively affecting ICIs action. However, a non-systemic antibiotic as rifaximin proved to stimulate the development of some beneficial bacteria including Lactobacillus, Faecalibacterium, and Bifidobacterium and to favor an anti-inflammatory action. Despite the promising data, this antibiotic has yet to be analyzed in preclinical studies on HCC models. Interestingly, a retrospective study by Pinato et al. [65] tested the correlation between therapy with antibiotics and clinical outcomes in 4,098 patients suffering from advanced HCC treated with ICIs or targeted therapy. To be specific, the study evaluated the correlation between the administration of antibiotics within 1 month before or after the start of treatment and clinical outcomes (OS and PFS). Among all patients, 15% of them received antibiotics with a shorter mPFS (3.6 vs. 4.2 months; HR 1.29) and OS (8.7 vs 10.6 months; HR 1.36). In detail, among patients who received antibiotics, a shorter PFS was observed both in those treated with ICI (HR 1.52), TKIs (HR 1.29), and placebo (HR 1.23).
Other drugs including metformin, aspirin, and statins can influence the composition of gut microbiota. In particular, these drugs are associated with a reduction in the HCC incidence by means of an anti-inflammatory effect probably due to an enhanced amount of Akkermansia and Bifidobacterium [66].
Probiotics are living bacteria that usually contain Lactobacillus and Bifidobacterium. They are administered with the aim to restore and improve the gut microbiota. The last generation probiotics include other bacteria such as Akkermansia muciniphila and Clostridium. Some data put in evidence that probiotics can lead to the production of metabolites and modulate immune system with anti-inflammatory and anti-tumor actions.
Prebiotics are non-digestible food ingredients that exert similar functions to probiotics but through the stimulation of the growth and activity of bacteria in the gut microbiota [8].
Another method to modulate the microbiome is the faecal microbial transplantation (FMT). It consists of the transfer of a stool sample from a healthy donor to a host [11]. Also exists a variant of FMT, called microbial ecosystem therapeutics (MET), that corresponds to the transplantation of living cultures of intestinal bacteria deriving from a faecal sample of a healthy individual [64,67]. The transplantation can be carried out through oral capsules, enema, or endoscopy although there is no consensus regarding the dose, frequency, and duration of FMT. This technique is usually employed to eradicate Clostridium difficile infection. As regards the role of FMT in oncology, the studies examined its anti-inflammatory actions with the aim to prevent HCC carcinogenesis starting from chronic liver disease. Furthermore, other studies focused on the relationship between the alterations in microbiome composition and the consequent modification in the tumor microenvironment (TME) [11]. In detail, experimental data on mouse models and patients treated with ICIs reported that the transplantation of Ruminococcaceae family bacteria led to an enhanced amount of IFN-γ+CD8+ T lymphocytes in TME improving anti-tumor immune response. All studies on animals and humans showed that FMT enhanced the response to immunotherapy in those patients who experienced a good response to this treatment [38,39,40,41,42,43,44,45,46]. Despite the studies about FMT focused on tumors different from HCC, the results might be also relevant for HCC in consideration of the gut microbiota ability of influencing the response to ICIs in this tumor [12].
In conclusion, the currently available data suggest that different compositions of the intestinal microbiome can determine different responses to immunotherapy in patients with advanced HCC. Furthermore, preliminary data indicate that the modification of microbiota composition might improve the response to immunotherapy. Therefore, in the next future, a new therapeutic strategy could consist of modifying the microbiome features in order to positively influence the response to ICIs. In addition, the microbiome could represent a predictive factor of response to immunotherapy. However, further studies are necessary to obtain confirmatory data.
Author Contributions
Conceptualization, C.L. and G. N.; methodology, E.F.; software, G.B.; validation, G.N. and E.F.; formal analysis, C.L. investigation, F.M.M.; resources, E.L.; data curation, F.A.; writing—original draft preparation, C.L.; writing—review and editing, C.L.; visualization, F.A.; supervision, G.N.; project administration, C.L.; funding acquisition, E.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Laface, C.; Memeo, R. Clinical Updates for Gastrointestinal Malignancies. J Pers Med 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Laface, C.; Ranieri, G.; Maselli, F.M.; Ambrogio, F.; Foti, C.; Ammendola, M.; Laterza, M.; Cazzato, G.; Memeo, R.; Mastrandrea, G.; et al. Immunotherapy and the Combination with Targeted Therapies for Advanced Hepatocellular Carcinoma. Cancers (Basel) 2023, 15. [Google Scholar] [CrossRef]
- Laface, C.; Fedele, P.; Maselli, F.M.; Ambrogio, F.; Foti, C.; Molinari, P.; Ammendola, M.; Lioce, M.; Ranieri, G. Targeted Therapy for Hepatocellular Carcinoma: Old and New Opportunities. Cancers (Basel) 2022, 14. [Google Scholar] [CrossRef]
- Laface, C.; Laforgia, M.; Molinari, P.; Ugenti, I.; Gadaleta, C.D.; Porta, C.; Ranieri, G. Hepatic Arterial Infusion of Chemotherapy for Advanced Hepatobiliary Cancers: State of the Art. Cancers (Basel) 2021, 13. [Google Scholar] [CrossRef]
- Ranieri, G.; Laface, C. Loco-Regional and Systemic Chemotherapies for Hepato-Pancreatic Tumors: Integrated Treatments. Cancers (Basel) 2020, 12, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Rahimi-Farsi, N.; Bostanian, F.; Shahbazi, T.; Shamsinejad, F.S.; Bolideei, M.; Mohseni, P.; Zangooie, A.; Boustani, F.; Shoorei, H. Novel Oncogenes and Tumor Suppressor Genes in Hepatocellular Carcinoma: Carcinogenesis, Progression, and Therapeutic Targets. Gene 2025, 941, 149229. [Google Scholar] [CrossRef]
- Khare, T.; Liu, K.; Chilambe, L.O.; Khare, S. NAFLD and NAFLD Related HCC: Emerging Treatments and Clinical Trials. Int J Mol Sci 2025, 26. [Google Scholar] [CrossRef] [PubMed]
- Temraz, S.; Nassar, F.; Kreidieh, F.; Mukherji, D.; Shamseddine, A.; Nasr, R. Hepatocellular Carcinoma Immunotherapy and the Potential Influence of Gut Microbiome. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Ye, J.; Liu, M. Characterization of Gut Microbiota in Patients with Primary Hepatocellular Carcinoma Received Immune Checkpoint Inhibitors: A Chinese Population-Based Study. Medicine 2020, 99, E21788. [Google Scholar] [CrossRef]
- Luo, W.; Guo, S.; Zhou, Y.; Zhao, J.; Wang, M.; Sang, L.; Chang, B.; Wang, B. Hepatocellular Carcinoma: How the Gut Microbiota Contributes to Pathogenesis, Diagnosis, and Therapy. Front Microbiol 2022, 13. [Google Scholar] [CrossRef]
- Spanu, D.; Pretta, A.; Lai, E.; Persano, M.; Donisi, C.; Mariani, S.; Dubois, M.; Migliari, M.; Saba, G.; Ziranu, P.; et al. Hepatocellular Carcinoma and Microbiota: Implications for Clinical Management and Treatment. World J Hepatol 2022, 14, 1319–1332. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Khatiwada, S.; Behary, J.; Kim, R.; Zekry, A. Modulation of the Gut Microbiome to Improve Clinical Outcomes in Hepatocellular Carcinoma. Cancers (Basel) 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Shui, L.; Yang, X.; Li, J.; Yi, C.; Sun, Q.; Zhu, H. Gut Microbiome as a Potential Factor for Modulating Resistance to Cancer Immunotherapy. Front Immunol 2020, 10. [Google Scholar] [CrossRef]
- Oh, B.; Boyle, F.; Pavlakis, N.; Clarke, S.; Eade, T.; Hruby, G.; Lamoury, G.; Carroll, S.; Morgia, M.; Kneebone, A.; et al. The Gut Microbiome and Cancer Immunotherapy: Can We Use the Gut Microbiome as a Predictive Biomarker for Clinical Response in Cancer Immunotherapy? Cancers (Basel) 2021, 13. [Google Scholar] [CrossRef]
- Reig, M.; Forner, A.; Rimola, J.; Ferrer-Fàbrega, J.; Burrel, M.; Garcia-Criado, Á.; Kelley, R.K.; Galle, P.R.; Mazzaferro, V.; Salem, R.; et al. BCLC Strategy for Prognosis Prediction and Treatment Recommendation: The 2022 Update. J Hepatol 2022, 76, 681–693. [Google Scholar] [CrossRef]
- Ranieri, G.; Laface, C.; Fazio, V.; Ceglia, D. De; MacIna, F.; Gisone, V.; Porcelli, M.; Vinciarelli, G.; Carella, C.; Molinari, P.; et al. Local Treatment with Deep Percutaneous Electrochemotherapy of Different Tumor Lesions: Pain Relief and Objective Response Results from an Observational Study. Eur Rev Med Pharmacol Sci 2020, 24, 7764–7775. [Google Scholar] [CrossRef]
- Llovet, J.M.; Ricci, S.; Mazzaferro, V.; Hilgard, P.; Gane, E.; Blanc, J.-F.; de Oliveira, A.C.; Santoro, A.; Raoul, J.-L.; Forner, A.; et al. Sorafenib in Advanced Hepatocellular Carcinoma. New England Journal of Medicine 2008, 359, 378–390. [Google Scholar] [CrossRef] [PubMed]
- Kudo, M.; Finn, R.S.; Qin, S.; Han, K.H.; Ikeda, K.; Piscaglia, F.; Baron, A.; Park, J.W.; Han, G.; Jassem, J.; et al. Lenvatinib versus Sorafenib in First-Line Treatment of Patients with Unresectable Hepatocellular Carcinoma: A Randomised Phase 3 Non-Inferiority Trial. The Lancet 2018, 391, 1163–1173. [Google Scholar] [CrossRef]
- Li, S.; Wang, G.; Wang, C.; Gao, X.; Jin, F.; Yang, H.; Han, B.; Zhou, R.; Chen, C.; Chen, L.; et al. The REACH Trial: A Randomized Controlled Trial Assessing the Safety and Effectiveness of the Spiration® Valve System in the Treatment of Severe Emphysema. Respiration 2018, 97, 416. [Google Scholar] [CrossRef] [PubMed]
- Abou-Alfa, G.K.; Meyer, T.; Cheng, A.-L.; El-Khoueiry, A.B.; Rimassa, L.; Ryoo, B.-Y.; Cicin, I.; Merle, P.; Chen, Y.; Park, J.-W.; et al. Cabozantinib in Patients with Advanced and Progressing Hepatocellular Carcinoma. N Engl J Med 2018, 379, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Tabrizian, P.; Abdelrahim, M.; Schwartz, M. Immunotherapy and Transplantation for Hepatocellular Carcinoma. J Hepatol 2024, 80, 822–825. [Google Scholar] [CrossRef]
- Harkus, U.; Wankell, M.; Palamuthusingam, P.; McFarlane, C.; Hebbard, L. Immune Checkpoint Inhibitors in HCC: Cellular, Molecular and Systemic Data. Semin Cancer Biol 2022, 86, 799–815. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.L.; Qin, S.; Ikeda, M.; Galle, P.R.; Ducreux, M.; Kim, T.Y.; Lim, H.Y.; Kudo, M.; Breder, V.; Merle, P.; et al. Updated Efficacy and Safety Data from IMbrave150: Atezolizumab plus Bevacizumab vs. Sorafenib for Unresectable Hepatocellular Carcinoma. J Hepatol 2022, 76, 862–873. [Google Scholar] [CrossRef] [PubMed]
- Sangro, B.; Chan, S.L.; Kelley, R.K.; Lau, G.; Kudo, M.; Sukeepaisarnjaroen, W.; Yarchoan, M.; De Toni, E.N.; Furuse, J.; Kang, Y.K.; et al. Four-Year Overall Survival Update from the Phase III HIMALAYA Study of Tremelimumab plus Durvalumab in Unresectable Hepatocellular Carcinoma. Ann Oncol 2024, 35, 448–457. [Google Scholar] [CrossRef]
- Galle, P.R.; Decaens, T.; Kudo, M.; Qin, S.; Fonseca, L.; Sangro, B.; Karachiwala, H.; Park, J.-W.; Gane, E.; Pinter, M.; et al. Nivolumab (NIVO) plus Ipilimumab (IPI) vs Lenvatinib (LEN) or Sorafenib (SOR) as First-Line Treatment for Unresectable Hepatocellular Carcinoma (UHCC): First Results from CheckMate 9DW. Journal of Clinical Oncology 2024, 42, LBA4008–LBA4008. [Google Scholar] [CrossRef]
- Ren, Z.; Xu, J.; Bai, Y.; Xu, A.; Cang, S.; Du, C.; Li, Q.; Lu, Y.; Chen, Y.; Guo, Y.; et al. Sintilimab plus a Bevacizumab Biosimilar (IBI305) versus Sorafenib in Unresectable Hepatocellular Carcinoma (ORIENT-32): A Randomised, Open-Label, Phase 2-3 Study. Lancet Oncol 2021, 22, 977–990. [Google Scholar] [CrossRef]
- Llovet, J.M.; Kudo, M.; Merle, P.; Meyer, T.; Qin, S.; Ikeda, M.; Xu, R.; Edeline, J.; Ryoo, B.Y.; Ren, Z.; et al. Lenvatinib plus Pembrolizumab versus Lenvatinib plus Placebo for Advanced Hepatocellular Carcinoma (LEAP-002): A Randomised, Double-Blind, Phase 3 Trial. Lancet Oncol 2023, 24, 1399–1410. [Google Scholar] [CrossRef] [PubMed]
- Kelley, R.K.; Rimassa, L.; Cheng, A.L.; Kaseb, A.; Qin, S.; Zhu, A.X.; Chan, S.L.; Melkadze, T.; Sukeepaisarnjaroen, W.; Breder, V.; et al. Cabozantinib plus Atezolizumab versus Sorafenib for Advanced Hepatocellular Carcinoma (COSMIC-312): A Multicentre, Open-Label, Randomised, Phase 3 Trial. Lancet Oncol 2022, 23, 995–1008. [Google Scholar] [CrossRef]
- Yau, T.; Park, J.W.; Finn, R.S.; Cheng, A.L.; Mathurin, P.; Edeline, J.; Kudo, M.; Harding, J.J.; Merle, P.; Rosmorduc, O.; et al. Nivolumab versus Sorafenib in Advanced Hepatocellular Carcinoma (CheckMate 459): A Randomised, Multicentre, Open-Label, Phase 3 Trial. Lancet Oncol 2022, 23, 77–90. [Google Scholar] [CrossRef]
- Finn, R.S.; Ryoo, B.Y.; Merle, P.; Kudo, M.; Bouattour, M.; Lim, H.Y.; Breder, V.; Edeline, J.; Chao, Y.; Ogasawara, S.; et al. Pembrolizumab As Second-Line Therapy in Patients With Advanced Hepatocellular Carcinoma in KEYNOTE-240: A Randomized, Double-Blind, Phase III Trial. J Clin Oncol 2020, 38, 193–202. [Google Scholar] [CrossRef]
- Gomaa, E.Z. Human Gut Microbiota/Microbiome in Health and Diseases: A Review. Antonie Van Leeuwenhoek 2020, 113, 2019–2040. [Google Scholar] [CrossRef]
- Milani, C.; Duranti, S.; Bottacini, F.; Casey, E.; Turroni, F.; Mahony, J.; Belzer, C.; Delgado Palacio, S.; Arboleya Montes, S.; Mancabelli, L.; et al. The First Microbial Colonizers of the Human Gut: Composition, Activities, and Health Implications of the Infant Gut Microbiota. Microbiol Mol Biol Rev 2017, 81. [Google Scholar] [CrossRef]
- Mak, I.E.K.; Yao, Y.; Ng, M.T.T.; Kim, J.E. Influence of Dietary Protein and Fiber Intake Interactions on the Human Gut Microbiota Composition and Function: A Systematic Review and Network Meta-Analysis of Randomized Controlled Trials. Crit Rev Food Sci Nutr 2025, 1–19. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, N.; Su, X.; Gao, Y.; Yang, R. Gut-Microbiota-Derived Metabolites Maintain Gut and Systemic Immune Homeostasis. Cells 2023, 12. [Google Scholar] [CrossRef]
- Schoeler, M.; Caesar, R. Dietary Lipids, Gut Microbiota and Lipid Metabolism. Rev Endocr Metab Disord 2019, 20, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Adak, A.; Khan, M.R. An Insight into Gut Microbiota and Its Functionalities. Cell Mol Life Sci 2019, 76, 473–493. [Google Scholar] [CrossRef]
- Góralczyk-Bińkowska, A.; Szmajda-Krygier, D.; Kozłowska, E. The Microbiota-Gut-Brain Axis in Psychiatric Disorders. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Gopalakrishnan, V.; Spencer, C.N.; Nezi, L.; Reuben, A.; Andrews, M.C.; Karpinets, T. V.; Prieto, P.A.; Vicente, D.; Hoffman, K.; Wei, S.C.; et al. Gut Microbiome Modulates Response to Anti-PD-1 Immunotherapy in Melanoma Patients. Science 2018, 359, 97–103. [Google Scholar] [CrossRef]
- Baruch, E.N.; Youngster, I.; Ben-Betzalel, G.; Ortenberg, R.; Lahat, A.; Katz, L.; Adler, K.; Dick-Necula, D.; Raskin, S.; Bloch, N.; et al. Fecal Microbiota Transplant Promotes Response in Immunotherapy-Refractory Melanoma Patients. Science 2021, 371, 602–609. [Google Scholar] [CrossRef]
- Davar, D.; Dzutsev, A.K.; McCulloch, J.A.; Rodrigues, R.R.; Chauvin, J.M.; Morrison, R.M.; Deblasio, R.N.; Menna, C.; Ding, Q.; Pagliano, O.; et al. Fecal Microbiota Transplant Overcomes Resistance to Anti-PD-1 Therapy in Melanoma Patients. Science 2021, 371, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Routy, B.; Le Chatelier, E.; Derosa, L.; Duong, C.P.M.; Alou, M.T.; Daillère, R.; Fluckiger, A.; Messaoudene, M.; Rauber, C.; Roberti, M.P.; et al. Gut Microbiome Influences Efficacy of PD-1-Based Immunotherapy against Epithelial Tumors. Science 2018, 359, 91–97. [Google Scholar] [CrossRef]
- Vétizou, M.; Pitt, J.M.; Daillère, R.; Lepage, P.; Waldschmitt, N.; Flament, C.; Rusakiewicz, S.; Routy, B.; Roberti, M.P.; Duong, C.P.M.; et al. Anticancer Immunotherapy by CTLA-4 Blockade Relies on the Gut Microbiota. Science 2015, 350, 1079–1084. [Google Scholar] [CrossRef]
- Ayers, M.; Lunceford, J.; Nebozhyn, M.; Murphy, E.; Loboda, A.; Kaufman, D.R.; Albright, A.; Cheng, J.D.; Kang, S.P.; Shankaran, V.; et al. IFN-γ-Related MRNA Profile Predicts Clinical Response to PD-1 Blockade. J Clin Invest 2017, 127, 2930–2940. [Google Scholar] [CrossRef]
- Sivan, A.; Corrales, L.; Hubert, N.; Williams, J.B.; Aquino-Michaels, K.; Earley, Z.M.; Benyamin, F.W.; Lei, Y.M.; Jabri, B.; Alegre, M.L.; et al. Commensal Bifidobacterium Promotes Antitumor Immunity and Facilitates Anti-PD-L1 Efficacy. Science 2015, 350, 1084–1089. [Google Scholar] [CrossRef]
- Tanoue, T.; Morita, S.; Plichta, D.R.; Skelly, A.N.; Suda, W.; Sugiura, Y.; Narushima, S.; Vlamakis, H.; Motoo, I.; Sugita, K.; et al. A Defined Commensal Consortium Elicits CD8 T Cells and Anti-Cancer Immunity. Nature 2019, 565, 600–605. [Google Scholar] [CrossRef]
- Mao, J.; Wang, D.; Long, J.; Yang, X.; Lin, J.; Song, Y.; Xie, F.; Xun, Z.; Wang, Y.; Wang, Y.; et al. Gut Microbiome Is Associated with the Clinical Response to Anti-PD-1 Based Immunotherapy in Hepatobiliary Cancers. J Immunother Cancer 2021, 9. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, T.; Tu, X.; Huang, Y.; Zhang, H.; Tan, D.; Jiang, W.; Cai, S.; Zhao, P.; Song, R.; et al. Gut Microbiome Affects the Response to Anti-PD-1 Immunotherapy in Patients with Hepatocellular Carcinoma. J Immunother Cancer 2019, 7. [Google Scholar] [CrossRef]
- Fridman, W.H.; Zitvogel, L.; Sautès-Fridman, C.; Kroemer, G. The Immune Contexture in Cancer Prognosis and Treatment. Nat Rev Clin Oncol 2017, 14, 717–734. [Google Scholar] [CrossRef]
- Nomura, M.; Nagatomo, R.; Doi, K.; Shimizu, J.; Baba, K.; Saito, T.; Matsumoto, S.; Inoue, K.; Muto, M. Association of Short-Chain Fatty Acids in the Gut Microbiome With Clinical Response to Treatment With Nivolumab or Pembrolizumab in Patients With Solid Cancer Tumors. JAMA Netw Open 2020, 3, e202895. [Google Scholar] [CrossRef]
- Luu, M.; Riester, Z.; Baldrich, A.; Reichardt, N.; Yuille, S.; Busetti, A.; Klein, M.; Wempe, A.; Leister, H.; Raifer, H.; et al. Microbial Short-Chain Fatty Acids Modulate CD8+ T Cell Responses and Improve Adoptive Immunotherapy for Cancer. Nat Commun 2021, 12. [Google Scholar] [CrossRef]
- Hu, C.; Xu, B.; Wang, X.; Wan, W.H.; Lu, J.; Kong, D.; Jin, Y.; You, W.; Sun, H.; Mu, X.; et al. Gut Microbiota-Derived Short-Chain Fatty Acids Regulate Group 3 Innate Lymphoid Cells in HCC. Hepatology 2023, 77, 48–64. [Google Scholar] [CrossRef]
- Mager, L.F.; Burkhard, R.; Pett, N.; Cooke, N.C.A.; Brown, K.; Ramay, H.; Paik, S.; Stagg, J.; Groves, R.A.; Gallo, M.; et al. Microbiome-Derived Inosine Modulates Response to Checkpoint Inhibitor Immunotherapy. Science 2020, 369, 1481–1489. [Google Scholar] [CrossRef]
- Behary, J.; Amorim, N.; Jiang, X.T.; Raposo, A.; Gong, L.; McGovern, E.; Ibrahim, R.; Chu, F.; Stephens, C.; Jebeili, H.; et al. Gut Microbiota Impact on the Peripheral Immune Response in Non-Alcoholic Fatty Liver Disease Related Hepatocellular Carcinoma. Nat Commun 2021, 12. [Google Scholar] [CrossRef]
- Manuzak, J.; Dillon, S.; Wilson, C. Differential Interleukin-10 (IL-10) and IL-23 Production by Human Blood Monocytes and Dendritic Cells in Response to Commensal Enteric Bacteria. Clin Vaccine Immunol 2012, 19, 1207–1217. [Google Scholar] [CrossRef] [PubMed]
- Lurje, I.; Hammerich, L.; Tacke, F. Dendritic Cell and T Cell Crosstalk in Liver Fibrogenesis and Hepatocarcinogenesis: Implications for Prevention and Therapy of Liver Cancer. Int J Mol Sci 2020, 21, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Machida, K.; Tahara, S.M. Immunotherapy and Microbiota for Targeting of Liver Tumor-Initiating Stem-like Cells. Cancers (Basel) 2022, 14. [Google Scholar] [CrossRef]
- Shen, Y.-C.; Lee, P.-C.; Kuo, Y.-L.; Wu, W.-K.; Chen, C.-C.; Lei, C.-H.; Yeh, C.-P.; Hsu, C.; Hsu, C.-H.; Lin, Z.-Z.; et al. An Exploratory Study for the Association of Gut Microbiome with Efficacy of Immune Checkpoint Inhibitor in Patients with Hepatocellular Carcinoma. J Hepatocell Carcinoma 2021, 8, 809–822. [Google Scholar] [CrossRef]
- Muhammed, A.; D’Alessio, A.; Enica, A.; Talbot, T.; Fulgenzi, C.A.M.; Nteliopoulos, G.; Goldin, R.D.; Cortellini, A.; Pinato, D.J. Predictive Biomarkers of Response to Immune Checkpoint Inhibitors in Hepatocellular Carcinoma. Expert Rev Mol Diagn 2022, 22, 253–264. [Google Scholar] [CrossRef]
- Gok Yavuz, B.; Hasanov, E.; Lee, S.S.; Mohamed, Y.I.; Curran, M.A.; Koay, E.J.; Cristini, V.; Kaseb, A.O. Current Landscape and Future Directions of Biomarkers for Immunotherapy in Hepatocellular Carcinoma. J Hepatocell Carcinoma 2021, 8, 1195–1207. [Google Scholar] [CrossRef]
- Vaziri, F.; Colquhoun, S.D.; Wan, Y.J.Y. Hepatocellular Carcinoma Immunotherapy: The Impact of Epigenetic Drugs and the Gut Microbiome. Liver Res 2020, 4, 191–198. [Google Scholar] [CrossRef]
- Finn, R.S.; Qin, S.; Ikeda, M.; Galle, P.R.; Ducreux, M.; Kim, T.-Y.; Kudo, M.; Breder, V.; Merle, P.; Kaseb, A.O.; et al. Atezolizumab plus Bevacizumab in Unresectable Hepatocellular Carcinoma. N Engl J Med 2020, 382, 1894–1905. [Google Scholar] [CrossRef]
- Abou-Alfa, G.K.; Chan, S.L.; Kudo, M.; Lau, G.; Kelley, R.K.; Furuse, J.; Sukeepaisarnjaroen, W.; Kang, Y.-K.; Dao, T. V.; Toni, E.N. De; et al. Phase 3 Randomized, Open-Label, Multicenter Study of Tremelimumab (T) and Durvalumab (D) as First-Line Therapy in Patients (Pts) with Unresectable Hepatocellular Carcinoma (UHCC): HIMALAYA. Journal of Clinical Oncology 2022, 40, 379–379. [Google Scholar] [CrossRef]
- Chung, M.W.; Kim, M.J.; Won, E.J.; Lee, Y.J.; Yun, Y.W.; Cho, S.B.; Joo, Y.E.; Hwang, J.E.; Bae, W.K.; Chung, I.J.; et al. Gut Microbiome Composition Can Predict the Response to Nivolumab in Advanced Hepatocellular Carcinoma Patients. World J Gastroenterol 2021, 27, 7340–7349. [Google Scholar] [CrossRef]
- Giannone, G.; Ghisoni, E.; Genta, S.; Scotto, G.; Tuninetti, V.; Turinetto, M.; Valabrega, G. Immuno-Metabolism and Microenvironment in Cancer: Key Players for Immunotherapy. Int J Mol Sci 2020, 21, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Pinato, D.J.; Li, X.; Mishra-Kalyani, P.; D’Alessio, A.; Fulgenzi, C.A.M.; Scheiner, B.; Pinter, M.; Wei, G.; Schneider, J.; Rivera, D.R.; et al. Association between Antibiotics and Adverse Oncological Outcomes in Patients Receiving Targeted or Immune-Based Therapy for Hepatocellular Carcinoma. JHEP Rep 2023, 5. [Google Scholar] [CrossRef]
- Ponziani, F.R.; Nicoletti, A.; Gasbarrini, A.; Pompili, M. Diagnostic and Therapeutic Potential of the Gut Microbiota in Patients with Early Hepatocellular Carcinoma. Ther Adv Med Oncol 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Laforgia, M.; Calabrò, C.; Scattone, A.; Laface, C.; Porcelli, M.; Gadaleta, C.D.; Nardulli, P.; Ranieri, G. Pharmacotherapy in Mast Cell Leukemia. Expert Opin Pharmacother 2020, 21, 1059–1069. [Google Scholar] [CrossRef]
Figure 1.
In ICI-responder patients, the gut microbiota favors the production of IL-12 by Dendritic Cells (DC). This leads to the activation of the Th1-based immune response and citotoxicity via cytotoxic T lymphocytes. The result is the inhibition of tumor proliferation. On the other hand, in ICI-non responder patients, the gut microbiota is dysbiotic and determines the immune tolerance by DC. They leads to the proliferation of T regulatory cells which have immunosuppressive functions through the production of IL-10. The result is tumor growth.
Figure 1.
In ICI-responder patients, the gut microbiota favors the production of IL-12 by Dendritic Cells (DC). This leads to the activation of the Th1-based immune response and citotoxicity via cytotoxic T lymphocytes. The result is the inhibition of tumor proliferation. On the other hand, in ICI-non responder patients, the gut microbiota is dysbiotic and determines the immune tolerance by DC. They leads to the proliferation of T regulatory cells which have immunosuppressive functions through the production of IL-10. The result is tumor growth.
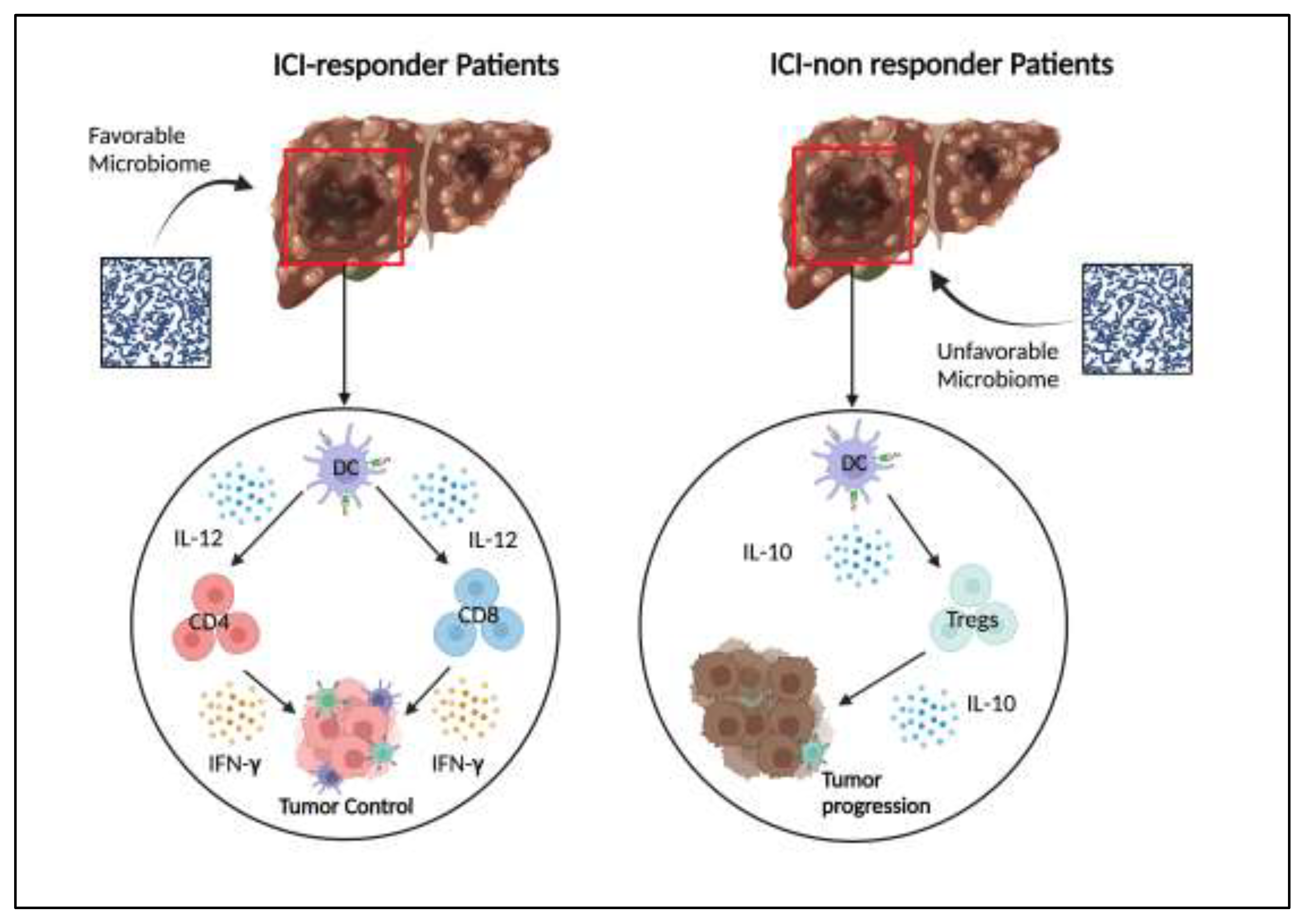

Table 1.
Summary about main clinical trials on ICIs-based therapy for HCC.
| CLINICAL TRIAL | TREATMENT | Setting | RESULTS |
|---|---|---|---|
| IMbrave 150 | Atezolizumab+Bevacizumab vs Sorafenib |
First-line | mOS: 19.2 vs 13.4 months mPFS: 6.8 vs 4.3 months ORR: 23.7% vs 11.9% |
| HIMALAYA | Durvalumab+Tremelimumab vs Sorafenib |
First-line | mOS: 16.4 vs 13.7 months mPFS: 3.78 vs 4.07 months ORR: 20.1% vs 2.1 % |
| CheckMate-9DW | Nivolumab+Ipilimumab vs Sorafenib or Lenvatinib |
First-line | mOS. 23.7 vs 20.6 months mPFS: 9.1 vs 9.2 months ORR: 36% vs 13% |
| Orient-32 | Sintilimab+IBI205 vs Sorafenib | First-line | mOS: Not reached vs 10.4 months mPFS: 4.6 vs 2.8 months |
| COSMIC-312 | Atezolizumab+Cabozantinib vs Sorafenib |
First-line | mOS: 16.5 vs 15.5 mPFS: 6.9 vs 4.3 |
| LEAP-002 | Pembrolizumab+Lenvatinib vs Lenvatinib |
First-line | mOS: 21.2 vs 19 months mPFS: 8.2 vs 8 months ORR: 26.3% vs 17.5% |
| CheckMate-459 | Nivolumab vs Sorafenib | First-line | mOS: 16.4 vs 14.7 months mPFS: 3.7 vs 3.8 months ORR: 15% vs 7% |
| Keynote-240 | Pembrolizumab vs Placebo | Second-line | mOS: 13.9 Vs 10.6 months mPFS: 3 vs 2.8 months ORR: 27.3 vs 11.9% |
HCC: hepatocellular carcinoma; mOS: median Overall Survival; mPFS: median Progression-free Survival; ORR: objective Response Rate.
Table 2.
Summary about the main species in microbiota of ICI-responder and non-responder HCC patients.
Table 2.
Summary about the main species in microbiota of ICI-responder and non-responder HCC patients.
| RESPONDERS | NON-RESPONDERS |
|---|---|
|
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



