
Submitted:
21 January 2025
Posted:
22 January 2025
You are already at the latest version
Abstract
A significant portion of adolescents suffer from mental illnesses and persistent pain due to repeated stress. It remains unclear which components of the nervous system are involved in the linkage between stress and pain at an early age. Prior studies in adult mice implicated the innate immune system, specifically Toll-like receptors (TLR), as critical for inducing long-term anxiety and pain-like behavior in social defeat stress (SDS) model. In this work, we investigated the pain and anxiety behavioral phenotypes of wild-type and TLR4-deficient juvenile mice subjected to repeated SDS and evaluated the engagement of TLR4 by measuring dimerization in the spinal cord, dorsal root ganglia, and prefrontal cortex. Male juvenile (4-week-old) mice (C57BL6/J or Tlr4-/-) underwent six social defeat sessions with adult CD1 aggressors. In WT mice SDS promotes chronic mechanical allodynia by von Frey testing and thermal hyperalgesia by Hargreaves test. In parallel, the stressed WT mice exhibited transient anxiety-like behavior and long-lasting locomotor activity reduction in the open-field test. Tlr4-/- stressed animals were resistant to the induction of pain-like behavior but had a remnant of anxious behavior, spending less time in the center of the arena. In WT SDS, there were concordant robust increases of TLR4 dimerization in dorsal root ganglia macrophages and spinal cord microglia, indicating activation. These results suggest that the chronic pain phenotype and locomotor impairment induced by SDS in juvenile mice depends on TLR4 engagement evidenced by dimerization in immune cells of the dorsal root ganglia and spinal cord.
Keywords:
Pain
; social stress
; TLR4
; spinal cord
; dorsal root ganglia
1. Introduction
Stress and pain are multifactorial constructs that are linked at several levels. Both activate similar pathways of the nervous system, such as the corticolimbic system, hippocampus, prefrontal cortex, and can be affected by genetic factors and psychosocial triggers (fears, emotional distress, beliefs, goals, threats) [1,2,3,4]. In acute conditions, stress may act as a protective response, inducing systemic cortisol release that provides energy to 'escape' from the stressor (a fight or flight state) [5]. However, stress becomes harmful when it occurs repeatedly for an extended period, leading to dysregulation in the neuroendocrine and immune systems [1]. Evidence shows that chronic stress is a major trigger for several mental disorders, such as anxiety and depression, and when it occurs in childhood or adolescent years, the individual becomes predisposed to the development of psychopathologies and pain sensitivity associated with increases in circulating inflammatory mediators released during stressful events [6]. In patients with depression, an increase of cytokines amounts in the peripheral blood was detected, which was also related to the participation of endogenous damage-associated molecular patterns (DAMPs) [7].
Preclinical models of physical and psychological stress have been used in rodents by applying unpleasant stressors such as repeated restraint, forced swims, sleep deprivation, maternal separation, and social defeat stress (SDS) [8]. The SDS model involves the resident-intruder paradigm in which a submissive mouse intruder is physically and sensorially exposed to an aggressor animal (resident) [9]. SDS was shown to promote behavioral alterations in adult mice, including anxiety, anhedonia, and social avoidance [10]. Interestingly, SDS also triggers widespread chronic pain phenotype, mimicking a condition similar to human fibromyalgia [10]. At present, there is an increasing interest in the study of pain phenotypes evoked by SDS in juvenile mice, given some clinical findings linking children and adolescent bullying with mental disorders and increased susceptibility to develop chronic pain in adulthood [11,12,13].
Our group has focused on studying the potential mechanisms and molecules for addressing chronic pain. In such an effort, toll-like receptor 4 (TLR4) has emerged as a key factor for neuronal sensitization [14,15,16]. These receptors are highly expressed in neural tissues, particularly in microglia and macrophages [17], and are demonstrated to participate in initiating and maintaining pain states, such as peripheral neuropathic pain [18] induced by chemotherapy and arthritic pain [19]. Despite this, there are many gaps to be filled regarding the mechanisms involved in the hyperexcitability of pain pathways as a result of chronic social stress, especially at early stages. Here, using social defeat stress (SDS), we investigated the chronic pain and anxiety-like behavior phenotype in juvenile male mice and the contribution of TLR4 activity in the dorsal root ganglia, spinal cord, and prefrontal cortex in the development of persistent pain caused by social stress.
2. Materials and Methods
2.1. Animals
Male C57Bl6/J mice at four to five weeks old were used as the experimental mice randomly divided into stressed (SDS) or no stressed (no-SDS) groups, and twelve-week-old male CD1 retired breeders as aggressors (purchased from Jackson laboratory© or ENVIGO©). They were maintained on isolated ventilated racks in a 12:12 hour light or dark cycle with controlled humidity and temperature (22°C). Food and water were available ad libitum. Tlr4-/- mice (TLR4-KO) were a gift from Dr. S. Akira (Osaka University, Japan) and backcrossed onto the C57Bl/6 background for ten generations [20]. Animal handling and behavioral experiments were performed as per the International Association for the Study of Pain (IASP) guidelines for using animals in pain research. The study was conducted in accordance with the rules established by the Institutional Animal Care and Use Committees of the University of California, San Diego, USA. All animal procedures performed in this study were approved by the Committee (IACUC – #S00137M).
2.2. Social Defeat Stress (SDS)
Social defeat stress is an ethologically relevant and widely used stress model to mimic the symptomatology of human depression and chronic pain, based on a resident-intruder paradigm [9]. We first screened several CD1 mice to categorize aggressive behavior. For this, we used a separate group of C57BL6/J mice as screeners. The screening consisted of 3 consecutive daily sessions of 3 minutes of physical contact between a CD1 with a C57 who was placed in the aggressor cage. A new screener mouse was placed in the CD1 cage in each session. The latency of CD1's first attack and the number of assaults against the screener were monitored and recorded following previous protocol standards [21]. Intermediated CD1 aggressive behavior was selected to perform the SDS protocol.
SDS protocol consisted of 6 physical sessions of 10 minutes between a C57BL6/J experimental mouse and a CD1 (resident) previously selected. They were maintained in the same cage for ten days, separated by a clear perforated plexiglass divider, allowing only sensorial contact (Supplementary Figure 1A). The physical sessions were split every two days (protocol days 1,2,5,6,9 and 10). The control group had no physical contact with the CD1 aggressor or other mice, being paired for ten days with another naive C57 separated by a perforated plexiglass divider (Supplementary Figure 1D). For acclimation, the divider was placed two days before the first stress session in the aggressor and control cages. After the end of the SDS protocol, on day 11, all experimental mice (SDS and no-SDS) were single-housed for further behavioral assessment.
2.3. Mechanical and Thermal Threshold Evaluation
The mechanical threshold was measured using Von Frey filaments (Bioseb) using the up-down method [22]. Different gauges or stiffness filaments (range from 2.44 to 4.31; 0.02-2.00 g) were applied to the plantar surface of both hind paws, and the withdrawal response was registered. Before the initiation of mechanical stimulation, mice were placed in clear, plastic, bottomless cages over a wire mesh surface and acclimated for 1 hour. Mechanical values for the left and right hind paws were measured and averaged to produce a single data point per measurement day.
Thermal hyperalgesia was evaluated by a Hargreaves-type testing device (UARDG, Department of Anesthesiology, University of California, San Diego)[23]. The device consists of a box with a glass surface on which the mice rest in plexiglass cubicles for 1 hour. A bulb is positioned below the glass surface, and when activated, it projects a focus heat light at 36-43 oC. The thermal stimulus is applied to both hind paws, and latency is defined as the time required for the mouse to produce a brisk paw withdrawal as detected by photodiode motion sensors that stop the timer and terminate the thermal stimuli. Von Frey and Hargreaves tests were performed before the stress session (day 0) and on days 7, 11, and 21 of the SDS protocol (see schema in Figure 1A). A blind investigator conducted both behavioral tests from 9:00 a.m. to 5:00 p.m.
2.4. Open-Field and Light-and-Dark Tests
Mice were subjected to the Open-Field test to evaluate locomotor activity and anxiety-like behavior. The test consisted of recording the mouse freely moving in an arena (40 × 40 × 40 cm3; 800 lx) for 10 minutes [24]. Specialized software (Ethovision 3.1, Noldus Information Technology BV, Wageningen, The Netherlands) was used to assess the real-time activity of each animal (nose, center of the body, and tail base). Each mouse was tested individually in a separate arena; 4 arenas were used per experiment. The parameters measured were: (1) number of entries to the center, (2) time spent in the center, (3) number of entries to the corners, (4) time spent in the corners, (5) total distance traveled, (6) body mobility, (7) velocity, and (8) maximum acceleration.
The light-and-dark test is performed in an apparatus with two interconnected compartments equally sized (90 × 90 × 165 cm) [25]. One compartment is illuminated with 7000 lux white light, and the other is painted dark and covered with an opaque lid. Mice started in the dark compartment and stayed for 10 minutes in the arena. The video tracker system registers the latency for the first entry into the light side and the time spent. Both behavioral tests were conducted on days 10 and 17 of the protocol between 9:00 a.m. and 5:00 p.m. Before each test, animals were placed in the room for 60 min for acclimation.
2.5. Immunostaining
Animals were euthanized eleven days after the last session of stress, day 22 of the protocol. The spinal cord (lumbar region), dorsal root ganglia (L3, L4, and L5), and prefrontal cortex were harvested and used for Flow Cytometry assay or Immunofluorescence. Tissues were dissociated using the Neural Tissue Dissociation Kit (Miltenyi Biotec, San Diego, CA, #130.092.628), following the manufacturer protocol. A debris removal solution (Kit Miltenyi Biotec, San Diego, CA, #130.109.398) was applied to the spinal cord and brain samples to eliminate debris. Single-cell suspensions were incubated for 45 min on ice with the following antibodies: APC-Cy7 live/dead Ghost dye (#18452, Cell Signaling, Danvers, MA) or 540 live/dead Ghost dye (#72086, Cell Signaling, Danvers, MA); PC7 -conjugated TLR4/MD2 (#MTS510; ThermoFisher); APC-conjugated total TLR4 (#SA15-21; Biolegend), 488-conjugated F4-80 (#123110; Biolegend), PercpCy 5.5-conjugated CD11b (#101228; Biolegend). After staining, cells were washed and analyzed using a CytoFLEX flow cytometer (Beckman Coulter, Brea, CA). To calculate TLR4 dimers, the ratio of the geometric mean fluorescence intensity of TLR4/MD2 monomer antibody over total TLR4 antibody was calculated, TLR4 dimers % = 100 − (TLR4 mono gMean × %)/(TLR4 total gMean × %) × (100%)/(highest TLR4 ratio) [14,26].
A second set of C57 mice was subjected to the SDS protocol, and the brain was harvested on day 22 for an immunofluorescence assay. The prefrontal cortex (PFC) was dissected post-fixed in a solution containing 4% formaldehyde in 0.01M PBS for 24 hours and cryoprotected using 30% sucrose until the tissue sank into the solution. Using cryocut in a Leica cryostat, 30 mm slices were performed and stored in an antifreeze solution at -20 oC. After all the PFCs were cut, 5-6 slices from each sample were processed as free-floating sections, rinsed in 0.01M PBS and blocked using 2% BSA for 60 min at room temperature. Next, the sections were incubated overnight with an anti-Iba1 antibody (1:500, Wako, 019-19741) at room temperature. The secondary antibody goat Anti-rabbit Alexa 488 (1:1000, Thermo Fisher #A11034) was applied for 1 hour. Sections were mounted in superfrost plus slides and coverslipped with ProLong™ Gold Antifade Mounting (Thermo Fisher, #P36935). Z-stacks imaging from three different ROIs of a section and three sections from each animal were executed using a Leica TCS SP5 confocal microscope with LAS AF version 2.7.3.9723 software. For quantification, the images were transformed into maximum projection, and a blind experimenter manually counted the number of positive cells for Iba1.
2.6. Statistical Analysis
Statistical analyses were performed in GraphPad Prism v10 (GraphPad Software Inc., La Jolla, CA, United States). Data are expressed as mean ± Standard Error of the Mean (SEM), and statistical differences were determined using one-way or two-way variance analysis (ANOVA) followed by Bonferroni’s multiple comparison test. Means were considered significantly different when p < 0.05. The Institutional Animal Care and Use Committees of the University of California, San Diego, approved all animal experimental procedures performed in this work (IACUC - protocol number #S00137M).
3. Results
3.1. SDS Induces Persistent Mechanical Allodynia and Thermal Hyperalgesia
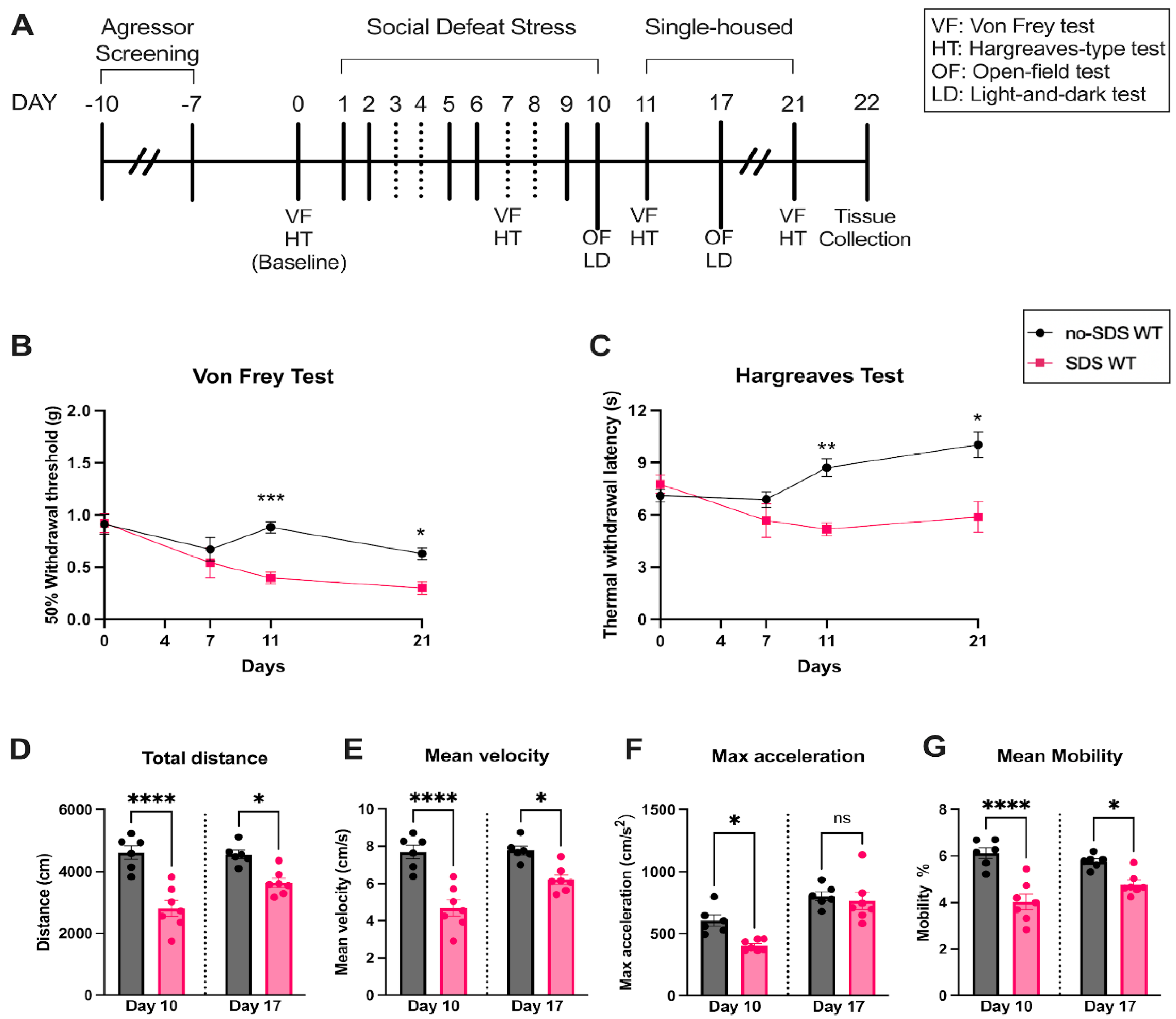
To assess whether chronic SDS can induce pain in juvenile mice, von Frey and Hargraves tests were performed at baseline, after four physical sessions (day 7), and after the end of the SDS protocol (days 11 and 21) (see experimental timeline in Figure 1A). Compared to the control group (no-SDS), stressed mice (SDS) demonstrated a slight decrease in the hind paw mechanical threshold on day 7, although not significant. From day 11, SDS animals showed a significant decrease in the paw threshold, which persisted until day 21 of the protocol, ten days after the last defeat session (Figure 1B). Similar results were observed in the thermal stimulus test, where the SDS group demonstrated a significantly reduced hind paw withdrawal latency to the heat (~43 oC) compared to controls on days 11 and 21 (Figure 1C). These findings show that SDS in juvenile male mice induces chronic mechanical allodynia and thermal hyperalgesia.
Figure 1.
Behavioral profile of WT mice. (A) Experimental timeline, SDS protocol, and behavior evaluation time points. (B) Paw mechanical threshold by von Frey filament of SDS and no-SDS groups, days 0, 7, 11, and 21. (C) Paw thermal threshold by Hargreaves test of SDS and no-SDS groups, days 0, 7, 11, and 21. (D) Open-field test: total distance traveled in the arena (cm). (E) Mean body velocity of each animal (cm/s). (F) Maximum body acceleration (cm/s2). (G) Body mean mobility (%). Data represent the mean ± SEM. Two-way ANOVA followed by Bonferroni's multiple comparisons test for (A-B). One-way ANOVA followed by Bonferroni's multiple comparisons test for (D-G). *p < 0.05, **p < 0.01, ***p < 0.001, ***p < 0.0001, ***p < 0.00001, n= 6 (SDS); n=7 (no-SDS).
Figure 1.
Behavioral profile of WT mice. (A) Experimental timeline, SDS protocol, and behavior evaluation time points. (B) Paw mechanical threshold by von Frey filament of SDS and no-SDS groups, days 0, 7, 11, and 21. (C) Paw thermal threshold by Hargreaves test of SDS and no-SDS groups, days 0, 7, 11, and 21. (D) Open-field test: total distance traveled in the arena (cm). (E) Mean body velocity of each animal (cm/s). (F) Maximum body acceleration (cm/s2). (G) Body mean mobility (%). Data represent the mean ± SEM. Two-way ANOVA followed by Bonferroni's multiple comparisons test for (A-B). One-way ANOVA followed by Bonferroni's multiple comparisons test for (D-G). *p < 0.05, **p < 0.01, ***p < 0.001, ***p < 0.0001, ***p < 0.00001, n= 6 (SDS); n=7 (no-SDS).
3.2. SDS Induces Long-Term Locomotor Changes and Short-Term Anxiety-Like Behavior
To assess anxiety-like behavior and locomotor activity, we performed the open-field and light-and-dark tests after the last stress session (day 10) and seven days after SDS (day 17). As observed in Figure 1D, stressed animals showed a significant decrease in spontaneous activity behavior (less distance traveled in the arena) during the open-field test compared to the no-SDS group on both days evaluated. Corroborating this result, the mean velocity and mobility were also decreased in SDS group (Figure 1E,F). Considering that mobility parameters indicate whether the mouse is highly mobile, moderately mobile, or immobile, our result suggests that chronic SDS resulted in a reduction of mobility, meaning less spontaneous motivation in the exploration of the arena, a profile that was not observed in the no-SDS group. Consequently, the decrease in mobility is reflected in the velocity measure since the speed is calculated by the distance traveled by the center, nose, or tail base point per unit of time. Interestingly, the max acceleration was significantly reduced on day 10 in SDS mice and reverted to a similar measurement of the control group on day 17 (Figure 1F). This behavioral profile uncovered that SDS in juvenile mice promoted persistent reduction in locomotor activity.
Furthermore, the open-field data showed that SDS triggers transient anxiety-like behavior. On day 10, the SDS group showed an anxiety-like phenotype, spending more time in the corners than in the center when compared to the non-SDS group (Figure 2A–H). This pattern was not continued after 7 days of the SDS; stressed animals show a recovery effect of the anxious state, presenting similar data to the no-SDS group at day 17 (Figure 2A–H). Despite these findings, the SDS protocol did not promote significant changes in the behavior of the stressed group in the light-and-dark test on day 10 or 17 compared to non-SDS (Figure 2I–K). Thus, an aversion to the light was not observed after SDS sessions of male juvenile mice.
3.3. TLR4 Activation in Spinal Cord, Dorsal Root Ganglia, and Prefrontal Cortex After SDS
TLR4 activation was analyzed by flow cytometry by calculating dimerization (ratio monometers to total expression) in immune cells of the DRG, spinal cord, and prefrontal cortex from the SDS and non-SDS groups. Significant changes were observed in the DRG and the spinal cord but not in the PFC. As seen in Figure 3B, TLR4 dimerization was significantly increased in microglia cells (CD11b+) of the lumbar spinal cord in the SDS group compared to the no-SDS group. Similarly, TLR4 dimerization was augmented on macrophage cells (CD11b+/F4-80+) of the DRG from stressed animals compared to non-stressed mice (Figure 3D). Besides this, analyses from Prefrontal cortex tissue in the flow cytometry or immunofluorescence assays did not show significant changes between the groups of animals regarding the number of Iba1+ cells (microglia activation marker) nor TLR4 dimerization in this population (Supplemental Figure 2A-D).
3.4. Genetic Ablation of TLR4 Expression Prevented Pain Phenotype Induced by SDS
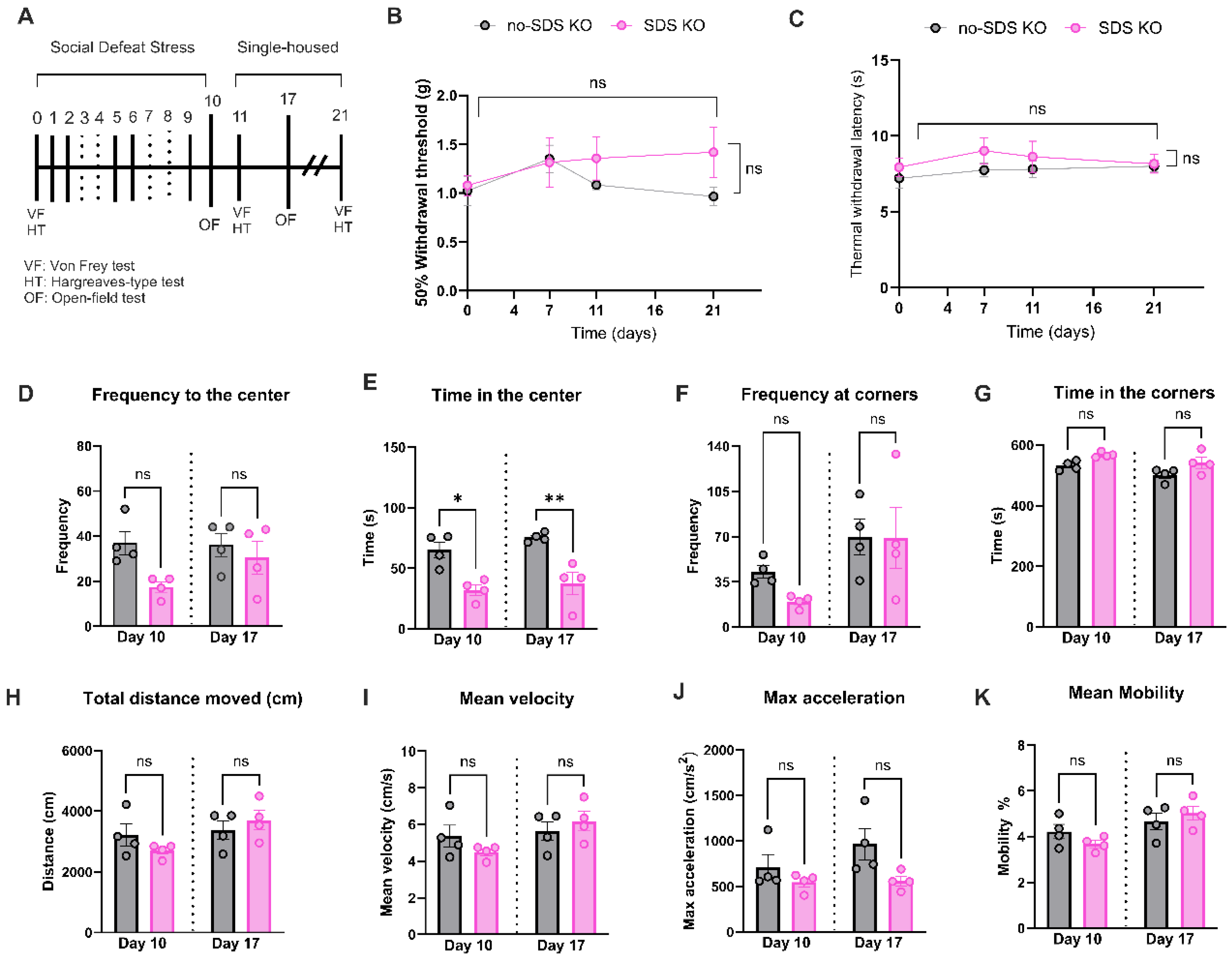
To confirm the participation of TLR4 receptors in the development of chronic pain or anxiety-like behavior phenotype in SDS, Tlr4-/- mice were subjected to the same protocol as the WT: six sessions of SDS, pain behavioral evaluation, and open-field test (see experimental timeline in Figure 4A). As reflected by the Von Frey readouts, SDS in Tlr4-/- mice did not show mechanical allodynia at any time point evaluated compared to no-SDS-KO (Figure 4B). A similar pattern was observed in thermal response after SDS in Tlr4-/- animals (Figure 4C). In the open-field test, the SDS-KO group decreased the time spent in the center of the arena in both times evaluated, days 10 and 17. However, there were no significant changes regarding the frequency to the center, the corner, and the time spent in the corners in SDS-KO compared to the no-SDS-KO group (Figure 4D,FG). With respect to locomotor activity parameters, total distance, velocity, acceleration, and mean mobility, no significant differences were detected between groups (Figure 4H–K). Together, these results suggest that TLR4 receptor depletion prevented locomotor activity reduction and chronic pain development induced by SDS.
4. Discussion
The current work reveals for the first time that SDS induces pain sensitivity in juvenile mice by involving TLR4 dimerization in the dorsal root ganglia macrophages and spinal cord microglia. In addition, SDS promotes persistent locomotor activity reduction in stressed animals compared to non-stressed animals. The lack of TLR4 expression in knock-out mice prevented the development of chronic pain phenotypes and locomotor changes induced by SDS.
A physiological stress response can be triggered by physical or psychological stressors. In these circumstances, the amygdala, in communication with the brain stem, promotes sympathetic adrenergic catecholamines, norepinephrine, and epinephrine release, resulting in typical stress symptoms: palpitation, high blood pressure, sweating, and pupil dilatation [4]. Long-lasting stress evokes hyperactivity in this system, lasting for an extended period even after the stress is discontinued [27]. In humans, chronic stressful experiences during childhood and adolescence (periods of intense social interaction) are shown to significantly impact mental health, resulting in anxiety and depression disorders, which can persist in adulthood [28]. Data from 2019 demonstrated that 970 million adults were found to be suffering from mental health disorders, and around 301 million suffer from anxiety, out of which 58 million were children and adolescents [29]. In parallel, studies proved that mental disorders caused by chronic stress exposure significantly affect the nociception pathways [27]. Animals subjected to stressful events presented an enhanced c-fos expression in the spinal cord, an increase in glutamate, and a reduction of GABA, leading to enhanced central excitability [30,31,32]. In humans, chronic stress alongside psychological disorders proved to be the primary trigger for widespread musculoskeletal pain development, known as fibromyalgia, one of the most challenging syndromes in clinics regarding pain management [33]. Collectively, these findings highlight the strong linkage between stress and pain processing. Here, we demonstrate that six sessions of SDS promote chronic pain phenotype in juvenile male mice subjugated and intimated (physically and sensorially) by the retired aggressor CD1 mice. The SDS group presented a reduction of thermal and mechanical thresholds persisting up to 21 days, 10 days after the aggressor had been removed from the cage. Similarly, previous work showed that ten consecutive days of social stress in adult mice leads to persistent hyperalgesia, long-lasting depressive-like behavior, and social avoidance [10]. In our open field data, however, SDS animals transiently exhibited depressive-like behavior; significant differences between the groups were only detected on day 10. Stressed mice presented lower distance traveled in the arena and less visited to the center (a spot considered "unsafe" and favorable to attacks) compared to non-stressed. Thus, the data suggests that SDS reduces the spontaneous interest in exploring the arena, tending to spend more time in the corners. However, this phenotype was abolished after 7 days of social stress sessions, contrary to what was observed in the behavioral pain tests where allodynia persisted until day 21. These findings indicate that juvenile mice might exhibit different anxiety behavior patterns compared to adults, recovering their interest in exploring the arena after a few days of SDS. Interestingly, the SDS protocol did not promote the natural aversion of stressed mice to brightly illuminated areas on our light-and-dark test. Nevertheless, in another study, stressed juvenile mice did not show a significant difference in exploring the arena compared to the control group in the open-field test, although they exhibited an increase in immobility behavior at tail-suspension and forced swimming tests [11]. Similar changes in locomotor activity due to stress were found in our results; the SDS group had reduced velocity and acceleration on both time points analyzed, which was reflected in lower body mobility than the non-stressed group. Accordingly, it has been reported that neonatal maternal separation-induced stress reduces exploratory activity in adult rats [34]. Unlike, the chronic restraint stress model performed for 5 days in adult rats promotes an increase in exploratory behavior [35]. Even though conflicted results, this evidence suggests that breed, age, and type or duration of the stress protocol can lead to distinct anxiety-like behavior parameters in rodents.
Recent works indicated chronic stress regulates nociception transmission through neuroinflammatory signaling involving microglia-mediated neuronal remodeling [36]. TLR4 receptors are abundantly expressed in brain and spinal microglia and typically, when activated, trigger an inflammatory cascade, leading to pro-nociceptive mediators release (e.g., TNF-α, COX-2, and interferon-β) [17]. According to our previous findings and in line with other works, TLR4 is a critical microglial receptor responsible for developing and maintaining pain [14,15,16,18,19,37,38]. In vivo TLR4 blockade attenuates hyperalgesia triggered by intrathecal LPS and neuropathic pain induced by nerve constriction injury or by chemotherapy drugs [15,16,18]. Although TLR4 is reported as an innate immune receptor, recent evidence shows that the interaction of TLR4 with N-Metil-D-Aspartate (NMDA) and Transient receptor potential vanilloid 1(TRPV1) can affect the nociceptive transmission via downstream phosphorylation of second messengers and posterior changes in primary afferent neurons transcriptome leading to changes in neuroplasticity [39] [14].
The dimerization process implies the recruitment of the TLR4 receptor at lipid rafts platforms via ligands, promoting cholesterol efflux and downstream signaling activation, which means the functionality of the receptor in the cell. In our data, TLR4 dimerization was associated with pain induced by social stress in juvenile mice since spinal microglia and dorsal root ganglia macrophages from stressed animals presented a higher level of TLR4 dimerization than the control group 10 days after the end of SDS protocol. Previously, using the social defeat stress model, a group found an increase in microglial activation and overexpression of inflammatory genes such as TLR4 in the spinal cord, which was associated with stress-induced hyperalgesia [36]. Accordingly, in this work, the global genetic depletion of TLR4 prevented the development of mechanical allodynia or thermal hyperalgesia, and no changes were observed in locomotor activity parameters, such as velocity, acceleration, or mean mobility in SDS-KO mice compared to no-SDS KO. However, a partial anxiety-like was observed in KO mice subjected to SDS since they spent less time in the center of the arena on days 10 and 17 of the protocol. Thus, the data indicated that TLR4 receptors seem to be more related to the pain phenotype than the anxiety disorder promoted by social stress in juvenile mice.
The HMGB1 (chromatin protein high-mobility group box1) is shown to be extracellularly released by immune cells such as macrophages and microglia under a stress state [40]. Into the extracellular space, through TLRs (including TLR4) signals, induce NF-κB signaling and proinflammatory cytokine production [41]. Considering the DRG is located outside the blood-brain barrier [42], circulating products released by stress from small amines (noradrenalin) to larger proteins (hormones and cytokines) can easily reach the cells of DRG, which can reflect changes in TLR4 signaling. Moreover, bulbospinal pathways are recognized to regulate dorsal horn excitability. Of note, serotonergic pathways facilitate pain signaling, activating dorsal horn neurons and microglia [43,44,45]. Serotonin receptor activation increases afferent terminal excitability [46] and activates DRG neurons [47]. Consistent with these observations, stress, as with models of conditioned fear mediated by supraspinal integration and learning, produces a profound change in spinal processing through the activation of bulbospinal serotonin projections [48]. These considerations jointly point to the possibility that higher order processes can initiate changes in downstream neuraxial signaling.
5. Conclusions
SDS in juvenile male mice promotes chronic pain phenotype and persistent locomotor activity reduction. These outcomes are related to the dimerization of TLR4 receptors in spinal microglia and dorsal root ganglia macrophages. The pain phenotype caused by SDS was prevented by genetic ablation of TLR4 receptors in stressed mice.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, Julia Borges Paes Lemes, Yury Miller and Tony Yaksh; Data curation, Julia Borges Paes Lemes, Alisa Panichkina, Kaue Franco Malange, Carlos Morado-Urbina, Sara Dochnal, Saee Jadhav and Juliana Navia-Pealez; Formal analysis, Julia Borges Paes Lemes, Alisa Panichkina, Kaue Franco Malange, Carlos Morado-Urbina and Saee Jadhav; Funding acquisition, Maripat Corr and Tony Yaksh; Investigation, Julia Borges Paes Lemes; Methodology, Julia Borges Paes Lemes, Alisa Panichkina and Maksim Dolmat; Project administration, Tony Yaksh; Resources, Yury Miller; Supervision, Yury Miller and Tony Yaksh; Validation, Alisa Panichkina; Visualization, Julia Borges Paes Lemes, Alisa Panichkina and Marco Pagliusi Jr; Writing – original draft, Julia Borges Paes Lemes, Alisa Panichkina and Sara Dochnal; Writing – review & editing, Julia Borges Paes Lemes, Alisa Panichkina, Kaue Franco Malange, Carlos Morado-Urbina, Sara Dochnal, Maksim Dolmat, Marco Pagliusi Jr, Juliana Navia-Pealez, Maripat Corr, Yury Miller and Tony Yaksh.
Funding
This project has received funding from NIH - 1R01NS131560.
Institutional Review Board Statement
The study was conducted in accordance with the rules established by the Institutional Animal Care and Use Committees of the University of California, San Diego, USA. All animal procedures performed in this study were approved by the Committee (IACUC – approval protocol number #S00137M).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available upon request from the corresponding author.
Acknowledgments
The authors thank Dr. Nicholas Webster for generously sharing access to a flow cytometer in his laboratory and thank Megan Sikkink and Dr Susan Powell for sharing the Open-field and Light-and-dark system.
Conflicts of Interest
T.L.Y and Y.I.M are inventors listed in patent applications related to the topic of this paper and scientific co-founders of Raft Pharmaceuticals LLC. The terms of this arrangement have been reviewed and approved by the University of California San Diego, in accordance with its conflict-of-interest policies. Other authors declare that they have no competing interests. All authors have read and agreed to the published version of the manuscript.
References
- McEwen, B.S. Neurobiological and Systemic Effects of Chronic Stress. Chronic Stress (Thousand Oaks) 2017, 1. [Google Scholar] [CrossRef]
- Franklin, T.B.; Silva, B.A.; Perova, Z.; Marrone, L.; Masferrer, M.E.; Zhan, Y.; Kaplan, A.; Greetham, L.; Verrechia, V.; Halman, A.; et al. Prefrontal cortical control of a brainstem social behavior circuit. Nat Neurosci 2017, 20, 260–270. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S.; Nasca, C.; Gray, J.D. Stress Effects on Neuronal Structure: Hippocampus, Amygdala, and Prefrontal Cortex. Neuropsychopharmacology 2016, 41, 3–23. [Google Scholar] [CrossRef] [PubMed]
- Hannibal, K.E.; Bishop, M.D. Chronic Stress, Cortisol Dysfunction, and Pain: A Psychoneuroendocrine Rationale for Stress Management in Pain Rehabilitation. Physical Therapy 2014, 94, 1816–1825. [Google Scholar] [CrossRef]
- Kara, E.; Hannibal, M.D.B. Chronic Stress, Cortisol Dysfunction, and Pain: A Psychoneuroendocrine Rationale for Stress Management in Pain Rehabilitation. Physical therapy 2014, 94, 1816. [Google Scholar] [CrossRef]
- Chiang, J.J.; Lam, P.H.; Chen, E.; Miller, G.E. Psychological Stress During Childhood and Adolescence and Its Association With Inflammation Across the Lifespan: A Critical Review and Meta-Analysis. Psychol Bull 2022, 148, 27–66. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.H.; Maletic, V.; Raison, C.L. Inflammation and its discontents: the role of cytokines in the pathophysiology of major depression. Biol Psychiatry 2009, 65, 732–741. [Google Scholar] [CrossRef] [PubMed]
- Atrooz, F.; Alkadhi, K.A.; Salim, S. Understanding stress: Insights from rodent models. Curr Res Neurobiol 2021, 2, 100013. [Google Scholar] [CrossRef] [PubMed]
- Golden, S.A.; Covington, H.E., 3rd; Berton, O.; Russo, S.J. A standardized protocol for repeated social defeat stress in mice. Nat Protoc 2011, 6, 1183–1191. [Google Scholar] [CrossRef]
- Piardi, L.N.; Pagliusi, M.; Bonet, I.; Brandao, A.F.; Magalhaes, S.F.; Zanelatto, F.B.; Tambeli, C.H.; Parada, C.A.; Sartori, C.R. Social stress as a trigger for depressive-like behavior and persistent hyperalgesia in mice: study of the comorbidity between depression and chronic pain. J Affect Disord 2020, 274, 759–767. [Google Scholar] [CrossRef]
- Matsumoto, K.; Takata, K.; Yamada, D.; Usuda, H.; Wada, K.; Tada, M.; Mishima, Y.; Ishihara, S.; Horie, S.; Saitoh, A.; et al. Juvenile social defeat stress exposure favors in later onset of irritable bowel syndrome-like symptoms in male mice. Sci Rep 2021, 11, 16276. [Google Scholar] [CrossRef] [PubMed]
- Lucas, R.; Talih, M.; Soares, S.; Fraga, S. Bullying Involvement and Physical Pain Between Ages 10 and 13 Years: Reported History and Quantitative Sensory Testing in a Population-Based Cohort. J Pain 2024, 25, 1012–1023. [Google Scholar] [CrossRef] [PubMed]
- Marin, T.J.; Hayden, J.A.; Lewinson, R.; Mahood, Q.; Pepler, D.; Katz, J. A Systematic Review of the Prospective Relationship Between Bullying Victimization and Pain. J Pain Res 2021, 14, 1875–1885. [Google Scholar] [CrossRef] [PubMed]
- Navia-Pelaez, J.M.; Borges Paes Lemes, J.; Gonzalez, L.; Delay, L.; Dos Santos Aggum Capettini, L.; Lu, J.W.; Goncalves Dos Santos, G.; Gregus, A.M.; Dougherty, P.M.; Yaksh, T.L.; et al. AIBP regulates TRPV1 activation in chemotherapy-induced peripheral neuropathy by controlling lipid raft dynamics and proximity to TLR4 in dorsal root ganglion neurons. Pain 2023, 164, e274–e285. [Google Scholar] [CrossRef]
- Woller, S.A.; Ravula, S.B.; Tucci, F.C.; Beaton, G.; Corr, M.; Isseroff, R.R.; Soulika, A.M.; Chigbrow, M.; Eddinger, K.A.; Yaksh, T.L. Systemic TAK-242 prevents intrathecal LPS evoked hyperalgesia in male, but not female mice and prevents delayed allodynia following intraplantar formalin in both male and female mice: The role of TLR4 in the evolution of a persistent pain state. Brain Behav Immun 2016, 56, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Bruno, K.; Woller, S.A.; Miller, Y.I.; Yaksh, T.L.; Wallace, M.; Beaton, G.; Chakravarthy, K. Targeting toll-like receptor-4 (TLR4)-an emerging therapeutic target for persistent pain states. Pain 2018, 159, 1908–1915. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Kim, H.; Lee, J.H.; Hwangbo, C. Toll-like receptor 4 (TLR4): new insight immune and aging. Immun Ageing 2023, 20, 67. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, H.; Zhang, H.; Kosturakis, A.K.; Jawad, A.B.; Dougherty, P.M. Toll-like receptor 4 signaling contributes to Paclitaxel-induced peripheral neuropathy. J Pain 2014, 15, 712–725. [Google Scholar] [CrossRef]
- Christianson, C.A.; Dumlao, D.S.; Stokes, J.A.; Dennis, E.A.; Svensson, C.I.; Corr, M.; Yaksh, T.L. Spinal TLR4 mediates the transition to a persistent mechanical hypersensitivity after the resolution of inflammation in serum-transferred arthritis. Pain 2011, 152, 2881–2891. [Google Scholar] [CrossRef]
- Hoshino, K.; Takeuchi, O.; Kawai, T.; Sanjo, H.; Ogawa, T.; Takeda, Y.; Takeda, K.; Akira, S. Cutting Edge: Toll-Like Receptor 4 (TLR4)-Deficient Mice Are Hyporesponsive to Lipopolysaccharide: Evidence for TLR4 as the Lps Gene Product. The Journal of Immunology 1999, 162, 3749–3752. [Google Scholar] [CrossRef]
- Pagliusi, M.O.F., Jr.; Sartori, C.R. Social Defeat Stress (SDS) in Mice: Using Swiss Mice as Resident. Bio Protoc 2019, 9, e3197. [Google Scholar] [CrossRef] [PubMed]
- Chaplan, S.R.; Bach, F.W.; Pogrel, J.W.; Chung, J.M.; Yaksh, T.L. Quantitative assessment of tactile allodynia in the rat paw. Journal of Neuroscience Methods 1994, 53, 55–63. [Google Scholar] [CrossRef]
- Dirig, D.M.; Salami, A.; Rathbun, M.L.; Ozaki, G.T.; Yaksh, T.L. Characterization of variables defining hindpaw withdrawal latency evoked by radiant thermal stimuli. Journal of Neuroscience Methods 1997, 76, 183–191. [Google Scholar] [CrossRef]
- Seibenhener, M.L.; Wooten, M.C. Use of the Open Field Maze to measure locomotor and anxiety-like behavior in mice. J Vis Exp 2015, e52434. [Google Scholar] [CrossRef]
- Bourin, M.; Hascoet, M. The mouse light/dark box test. Eur J Pharmacol 2003, 463, 55–65. [Google Scholar] [CrossRef]
- Navia-Pelaez, J.M.; Choi, S.H.; Dos Santos Aggum Capettini, L.; Xia, Y.; Gonen, A.; Agatisa-Boyle, C.; Delay, L.; Goncalves Dos Santos, G.; Catroli, G.F.; Kim, J.; et al. Normalization of cholesterol metabolism in spinal microglia alleviates neuropathic pain. J Exp Med 2021, 218. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, C.G.; Geha, P. Chronic Pain and Chronic Stress: Two Sides of the Same Coin? Chronic Stress (Thousand Oaks) 2017, 1. [Google Scholar] [CrossRef] [PubMed]
- Schlack, R.; Peerenboom, N.; Neuperdt, L.; Junker, S.; Beyer, A.K. The effects of mental health problems in childhood and adolescence in young adults: Results of the KiGGS cohort. J Health Monit 2021, 6, 3–19. [Google Scholar] [CrossRef]
- Organization, W.H. Mental disorders. Available online: (accessed on 01/16/2025).
- Quintero, L.; Cuesta, M.C.; Silva, J.A.; Arcaya, J.L.; Pinerua-Suhaibar, L.; Maixner, W.; Suarez-Roca, H. Repeated swim stress increases pain-induced expression of c-Fos in the rat lumbar cord. Brain Res 2003, 965, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Suarez-Roca, H.; Silva, J.A.; Arcaya, J.L.; Quintero, L.; Maixner, W.; Pinerua-Shuhaibar, L. Role of mu-opioid and NMDA receptors in the development and maintenance of repeated swim stress-induced thermal hyperalgesia. Behav Brain Res 2006, 167, 205–211. [Google Scholar] [CrossRef]
- Quintero, L.; Cardenas, R.; Suarez-Roca, H. Stress-induced hyperalgesia is associated with a reduced and delayed GABA inhibitory control that enhances post-synaptic NMDA receptor activation in the spinal cord. Pain 2011, 152, 1909–1922. [Google Scholar] [CrossRef] [PubMed]
- Sarzi-Puttini, P.; Giorgi, V.; Marotto, D.; Atzeni, F. Fibromyalgia: an update on clinical characteristics, aetiopathogenesis and treatment. Nat Rev Rheumatol 2020, 16, 645–660. [Google Scholar] [CrossRef] [PubMed]
- Ji, N.N.; Jiang, H.; Xia, M. Sex-dependent effects of postweaning exposure to an enriched environment on visceral pain and anxiety- and depression-like behaviors induced by neonatal maternal separation. Transl Pediatr 2022, 11, 1570–1576. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Ma, Y.; Dong, B.; Hu, B.; He, H.; Jia, J.; Xiong, M.; Xu, T.; Xu, B.; Xi, W. Functional magnetic resonance imaging study on anxiety and depression disorders induced by chronic restraint stress in rats. Behav Brain Res 2023, 450, 114496. [Google Scholar] [CrossRef] [PubMed]
- Sawicki, C.M.; Kim, J.K.; Weber, M.D.; Faw, T.D.; McKim, D.B.; Madalena, K.M.; Lerch, J.K.; Basso, D.M.; Humeidan, M.L.; Godbout, J.P.; et al. Microglia Promote Increased Pain Behavior through Enhanced Inflammation in the Spinal Cord during Repeated Social Defeat Stress. J Neurosci 2019, 39, 1139–1149. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, G.G.; Jimenez-Andrade, J.M.; Munoz-Islas, E.; Candanedo-Quiroz, M.E.; Cardenas, A.G.; Drummond, B.; Pham, P.; Stilson, G.; Hsu, C.C.; Delay, L.; et al. Role of TLR4 activation and signaling in bone remodeling, and afferent sprouting in serum transfer arthritis. Arthritis Res Ther 2024, 26, 212. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, Y.; Wang, J.; Fan, Q.; Zhu, J.; Yang, L.; Rong, W. TLR4 mediates upregulation and sensitization of TRPV1 in primary afferent neurons in 2,4,6-trinitrobenzene sulfate-induced colitis. Mol Pain 2019, 15, 1744806919830018. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Yu, S.; Li, Y.; Li, P.; Liu, F. Direct binding of Toll-like receptor 4 to ionotropic glutamate receptor N-methyl-D-aspartate subunit 1 induced by lipopolysaccharide in microglial cells N9 and EOC 20. Int J Mol Med 2018, 41, 1323–1330. [Google Scholar] [CrossRef]
- Wang, B.; Lian, Y.J.; Su, W.J.; Peng, W.; Dong, X.; Liu, L.L.; Gong, H.; Zhang, T.; Jiang, C.L.; Wang, Y.X. HMGB1 mediates depressive behavior induced by chronic stress through activating the kynurenine pathway. Brain Behav Immun 2018, 72, 51–60. [Google Scholar] [CrossRef]
- Andersson, U.; Tracey, K.J. HMGB1 is a therapeutic target for sterile inflammation and infection. Annu Rev Immunol 2011, 29, 139–162. [Google Scholar] [CrossRef] [PubMed]
- Berta, T.; Qadri, Y.; Tan, P.H.; Ji, R.R. Targeting dorsal root ganglia and primary sensory neurons for the treatment of chronic pain. Expert Opin Ther Targets 2017, 21, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Svensson, C.I.; Tran, T.K.; Fitzsimmons, B.; Yaksh, T.L.; Hua, X.Y. Descending serotonergic facilitation of spinal ERK activation and pain behavior. FEBS Lett 2006, 580, 6629–6634. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Kang, D.H.; Jeon, J.; Lee, H.G.; Kim, W.M.; Yoon, M.H.; Choi, J.I. Imbalance in the spinal serotonergic pathway induces aggravation of mechanical allodynia and microglial activation in carrageenan inflammation. Korean J Pain 2023, 36, 51–59. [Google Scholar] [CrossRef]
- Godinez-Chaparro, B.; Lopez-Santillan, F.J.; Orduna, P.; Granados-Soto, V. Secondary mechanical allodynia and hyperalgesia depend on descending facilitation mediated by spinal 5-HT(4), 5-HT(6) and 5-HT(7) receptors. Neuroscience 2012, 222, 379–391. [Google Scholar] [CrossRef]
- Chen, X.; Lv, R.; Li, M.; Zhang, L.; Sun, Y.; Cao, N.; Gu, B. The serotonin(5-HT)2A receptor is involved in the hypersensitivity of bladder afferent neurons in cyclophosphamide-induced cystitis. Eur J Pharmacol 2024, 982, 176909. [Google Scholar] [CrossRef] [PubMed]
- Lopez, E.R.; Carbajal, A.G.; Tian, J.B.; Bavencoffe, A.; Zhu, M.X.; Dessauer, C.W.; Walters, E.T. Serotonin enhances depolarizing spontaneous fluctuations, excitability, and ongoing activity in isolated rat DRG neurons via 5-HT(4) receptors and cAMP-dependent mechanisms. Neuropharmacology 2021, 184, 108408. [Google Scholar] [CrossRef]
- Vianna, D.M.; Allen, C.; Carrive, P. Cardiovascular and behavioral responses to conditioned fear after medullary raphe neuronal blockade. Neuroscience 2008, 153, 1344–1353. [Google Scholar] [CrossRef]
Figure 2.
Open-Field and Light-and-dark test. (A-D) Representative tracking heatmap image of four mice in the arena, color spectrum gradient, blue low intensity, and red high intensity. (A) no-SDS group (B) SDS group, on day 10. (C) no-SDS group, (D) SDS group, on day 17. (E) Frequency to the center of the arena (F) Time spent in the center. (G) Frequency of visits to the corner of the arena. (H) Time spent in the corners. (I) Cumulative time spent in the bright chamber. (J) Time until the first entrance of the bright chamber. (K) Frequency of entries to the bright chamber. Data represents the mean ± SEM. One-way ANOVA followed by Bonferroni’s multiple comparisons test, *p < 0.05, **p < 0.01, n= 6 (SDS); n=7 (no-SDS).
Figure 2.
Open-Field and Light-and-dark test. (A-D) Representative tracking heatmap image of four mice in the arena, color spectrum gradient, blue low intensity, and red high intensity. (A) no-SDS group (B) SDS group, on day 10. (C) no-SDS group, (D) SDS group, on day 17. (E) Frequency to the center of the arena (F) Time spent in the center. (G) Frequency of visits to the corner of the arena. (H) Time spent in the corners. (I) Cumulative time spent in the bright chamber. (J) Time until the first entrance of the bright chamber. (K) Frequency of entries to the bright chamber. Data represents the mean ± SEM. One-way ANOVA followed by Bonferroni’s multiple comparisons test, *p < 0.05, **p < 0.01, n= 6 (SDS); n=7 (no-SDS).
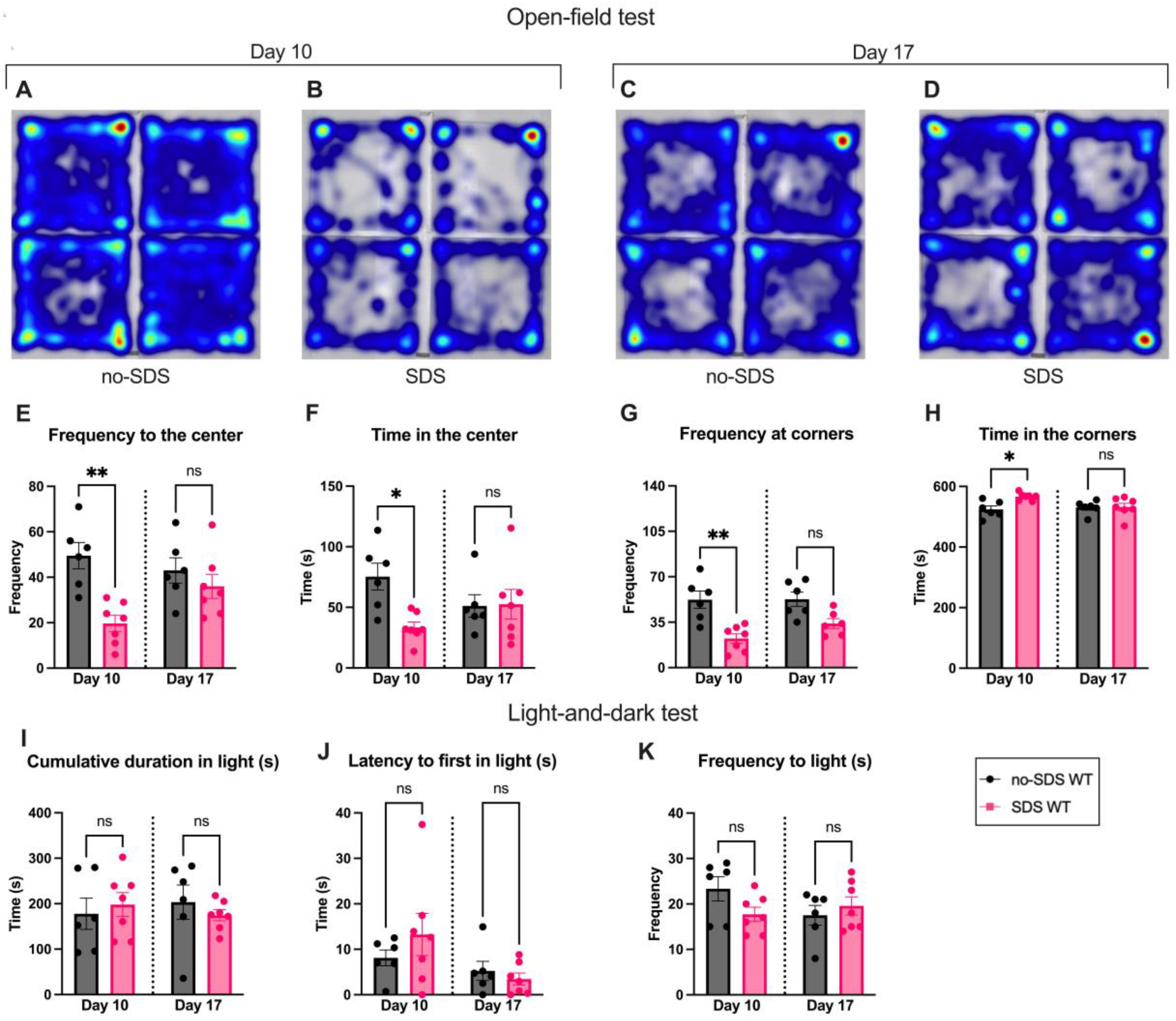
Figure 3.
Flow cytometry analysis of Spinal Cord and DRG on day 22. (A-C) Gating strategy for lumbar spinal cord and DRG cells. (B-D) TLR4 dimers (%) in SDS and no-SDS groups, microglia (CD11b+ population), and macrophages (CD11b+/F4-80+ population). Data represents mean ± SEM, t-test, SDS vs. no-SDS, *p < 0.05, **p < 0.01, n= 6 (SDS); n=6 (no-SDS).
Figure 3.
Flow cytometry analysis of Spinal Cord and DRG on day 22. (A-C) Gating strategy for lumbar spinal cord and DRG cells. (B-D) TLR4 dimers (%) in SDS and no-SDS groups, microglia (CD11b+ population), and macrophages (CD11b+/F4-80+ population). Data represents mean ± SEM, t-test, SDS vs. no-SDS, *p < 0.05, **p < 0.01, n= 6 (SDS); n=6 (no-SDS).
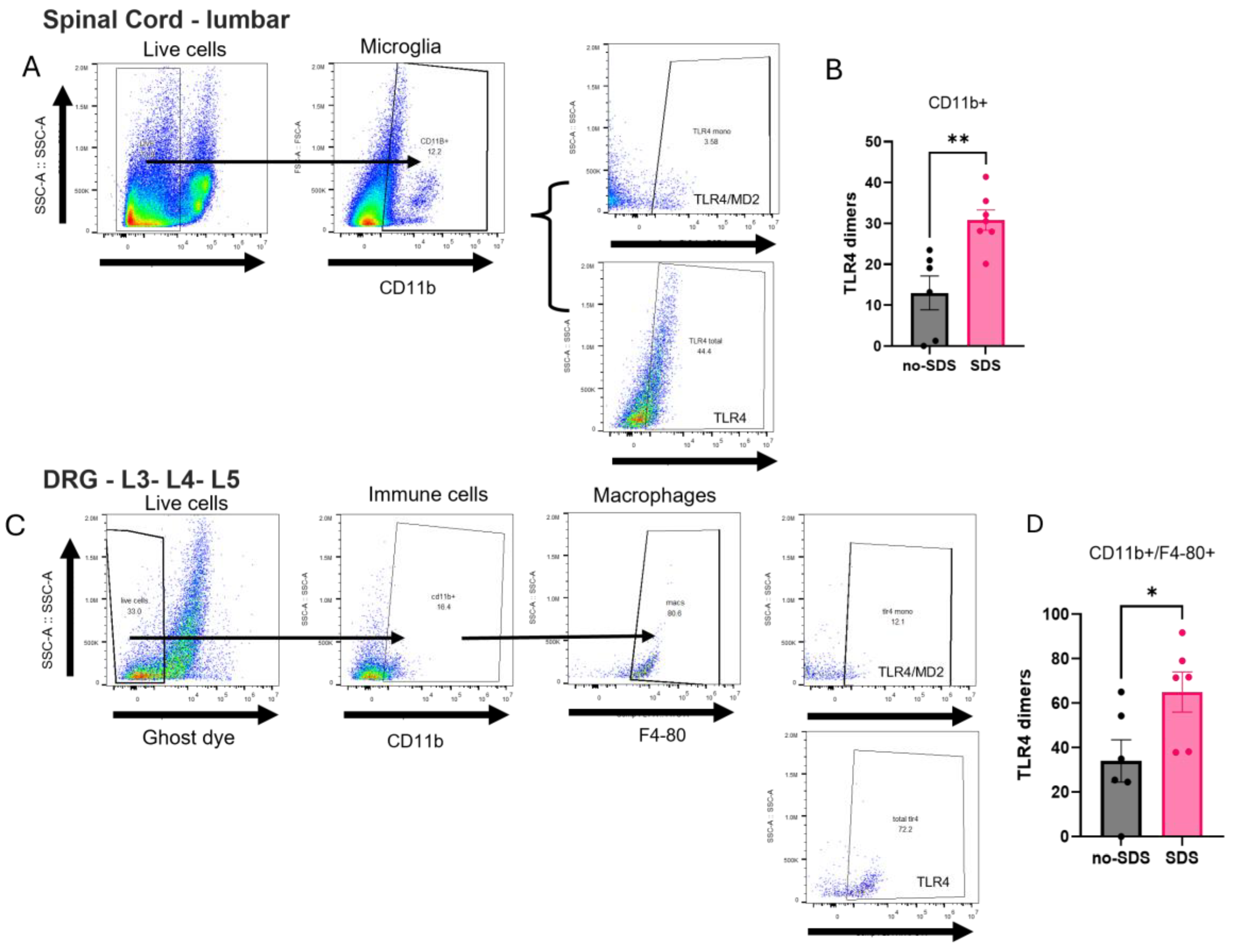
Figure 4.
Behavioral profile for Tlr4-/- mice. (A) Experimental timeline, SDS protocol, and behavior evaluation time points. (B) Paw mechanical threshold by von Frey filament of SDS and no-SDS KO mice at day 0, 7, 11, and 21. (C) Paw thermal threshold by Hargreaves test in SDS and no-SDS groups KO mice at day 0, 7, 11, and 21. n=5 (SDS); n=5 (no-SDS). Open-field test, in (D) total distance traveled in the arena (cm), in (E) Time spent in the center, in (F) Frequency of visit in corners of the arena, in (G) Frequency of visit in the center area. (H-K) Locomotor activity parameters of SDS and no-SDS in KO mice. Data represents the mean ± SEM. Two-way ANOVA followed by Bonferroni's multiple comparisons test for (B-C). One-way ANOVA followed by Bonferroni's multiple comparisons test for (D-K). *p < 0.05, **p < 0.01, n=4 (SDS); n=4 (no-SDS).
Figure 4.
Behavioral profile for Tlr4-/- mice. (A) Experimental timeline, SDS protocol, and behavior evaluation time points. (B) Paw mechanical threshold by von Frey filament of SDS and no-SDS KO mice at day 0, 7, 11, and 21. (C) Paw thermal threshold by Hargreaves test in SDS and no-SDS groups KO mice at day 0, 7, 11, and 21. n=5 (SDS); n=5 (no-SDS). Open-field test, in (D) total distance traveled in the arena (cm), in (E) Time spent in the center, in (F) Frequency of visit in corners of the arena, in (G) Frequency of visit in the center area. (H-K) Locomotor activity parameters of SDS and no-SDS in KO mice. Data represents the mean ± SEM. Two-way ANOVA followed by Bonferroni's multiple comparisons test for (B-C). One-way ANOVA followed by Bonferroni's multiple comparisons test for (D-K). *p < 0.05, **p < 0.01, n=4 (SDS); n=4 (no-SDS).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



