
Submitted:
17 January 2025
Posted:
19 January 2025
You are already at the latest version
Abstract
Hookworm infections affect 500-700 million people worldwide and can lead to chronic conditions such as malnutrition and anemia. The challenge of managing these infections is heightened by the absence of effective vaccines and the potential for anthelmintic resistance to develop. A comprehensive understanding of the molecular interactions between parasite and host is vital for unraveling the complexities of infection dynamics. This study aimed to identify immune system components responsible for host specificity in hookworms by infecting immunodeficient mouse models. Findings herein indicate that innate immunity is essential in protecting against Ancylostoma ceylanicum establishment in mice. Significant differences in parasite development were noted in mice lacking the signal transducer and activator of transcription 6 (Stat6-), with female mice reliant on this Th2 pathway for protection. Secondary infections in female Stat6- mice and an immunodeficient NSG mouse reached patency, demonstrating that immunodeficient mice fail to develop protective immunity against subsequent infections, similar to human hookworm infections. In contrast, no parasite development was observed in mice infected with A. caninum, highlighting that this species survival strategies are independent of host immune landscape. These results underscore the complexity of host-parasite interactions and point to new directions for therapeutic strategies, which may differ between sex.

Keywords:
Host specificity
; Hookworm infections
; Ancylostoma ceylanicum
; Ancylostoma caninum
; Mouse models
; Stat6 pathway
; Innate immunity
1. Introduction
Hookworms are soil-transmitted nematodes (STNs) that impose a significant global health burden, affecting 500-700 million people worldwide [1]. These blood-feeding intestinal worms can lead to debilitating and sometimes fatal iron deficiency anemia [2,3]. Infections are particularly devastating in children, resulting in stunted physical and cognitive development, which may be permanent [4,5]. Both pregnant women and the elderly are at high risk for morbidity, with pregnant women facing risks such as low birth weight and increased infant mortality [6]. The substantial health impacts of hookworms highlight the urgent need for a better understanding of hookworm infections to develop more effective control strategies and interventions.
One of the most important questions in parasitology is the molecular mechanisms that determine host specificity, or the range of host species that can be infected by a particular parasite species. Parasites range from strict specialists capable of infecting only one or a few closely related hosts to generalists that can exploit numerous host taxa [7]. Hookworms cover the entire spectrum, from strict specialists like Necator americanus, capable of developing only in humans, to broad generalists like Ancylostoma ceylanicum that can exploit several hosts species. Although hookworms as a group can infect a diverse range of hosts, the factors determining their individual host specificity are poorly understood. Exposing the molecular determinants of host specificity could lead to significant breakthroughs in understanding parasite biology and identifying novel intervention targets.
While A. ceylanicum can parasitize multiple species including humans, dogs, cats, and hamsters, it notably does not establish in mice. However, studies have shown that a small fraction of the infective dose can resume development within the first 3 days in outbred mice, but they fail to establish and are expelled by day 8 post-infection (PI) [8]. Yet, when mice are immunosuppressed with hydrocortisone acetate, A. ceylanicum infective third-stage larvae (iL3s) were able to mature into gravid adults, suggesting that hookworm development may be to be due to immune-mediated factors. Furthermore, oral infection with adult hookworms leads to attachment and feeding in various mouse strains, causing anemia and weight loss before the parasites are expelled by day 14 PI [9]. This suggests that structural differences in the small intestine between permissive and non-permissive hosts do not hinder attachment and feeding.
Our previous work has demonstrated that immune mechanisms dictate host specificity in the generalist A. ceylanicum, but not in the specialist A. caninum [10]. Additionally, our comparative transcriptomic analyses of intestinal cells from permissive hamsters and non-permissive mouse hosts have revealed distinct immune responses between these two hosts. [11]. Notably, the Th2 immune pathway was significantly enhanced in non-permissive mice, highlighted by the upregulation of interleukin-4 receptor alpha (IL-4Rα) and interleukin-13 receptor alpha 2 (IL-13Rα2). In contrast, IL-4Rα was downregulated in permissive hamsters, while there were no significant changes in IL-13Rα2.
To explore which specific arms of the immune system contribute to protection, we infected Signal Transducer and Activator of Transcription 6 (Stat6) knockout mice (Stat6-) with A. ceylanicum. These mice exhibit impaired Th2 immune responses, allowing us to examine the specific role of this pathway in hookworm susceptibility. Our findings indicate a sex-dependent permissiveness in Stat6- mice, which prompted further investigation into the role of sex hormones. Observations in Stat6- and NOD scid mice suggest that innate immune mechanisms are responsible for protection against A. ceylanicum establishment. In contrast, A. caninum does not appear to rely on the host immune landscape for its establishment and may have evolved unique survival strategies, such as hypobiosis and paratenesis, to optimize transmission within its relatively narrow host range. Understanding the molecular mechanisms that mediate host specificity is critical for developing new, effective intervention strategies against hookworm infections, which, as demonstrated, may vary based on the hookworm species and host sex.
2. Materials and Methods
Five different mouse strains were used in this study. NSG mice contain the scid mutation in the DNA repair complex protein Prkdc and the complete null allele of the interleukin-2 (IL-2) receptor common gamma chain (IL2rgnull) (JAX stock #005557). These mice are completely immunodeficient as they lack mature T cells and B cells and possess defective cytokine signaling, resulting in a deficiency of natural killer cells [12]. One NOD scid gamma (NSG) mouse (kindly donated by the Fernandes Lab) at George Washington University (GW) was reinfected. This mouse had previously remained patent for 121 days during its primary infection. The remaining mice were obtained from the Jackson Laboratory, including NOD scid mice, which possess the same scid mutation in the Prkdc gene as NSG mice but lack the IL2rgnull mutation, resulting in no functional T or B cells, while retaining a normal hematopoietic environment and an intact innate immune system (Jax stock #001303) [13]. Stat6- (JAX stock #005977) exhibit lymphocytes that do not proliferate in response to IL-4, impairing Th2 immune responses [14]. Homozygous estrogen receptor 1 (Esr1) KO mice have no tissue response to estrogen and estrogen receptor alpha activity (JAX stock #026176) [15]. Finally, C57BL/6J (Black 6N) mice were used as a control for Esr1 KO as they have the same genetic background (JAX stock #005304). Syrian golden hamsters (Mesocricetus auratus) were bred in-house at GW and used for A. ceylanicum maintenance.
Ancylostoma ceylanicum (US National Parasite Collection No. 102954) was maintained in the Syrian golden hamster, while A. caninum (US National Parasite Collection No. 106970) was maintained in beagles, following established protocols [10,16]. The preparation of L3 inocula was conducted as previously described [10], and administered to the animals via oral gavage. Inocula ranged from 80-1000 infective third-stage larvae, depending on the hookworm isolate and animal species being tested. For mice, an inoculum of 50 μL was delivered, followed by 50 μL of nuclease-free water to ensure any residual iL3s were washed from the tubes. For hamsters, an inoculum of 100 μL was delivered, followed by 100 μL rinse. On the day of infection, the NOD scid mice were ~10 weeks old, the Black 6N mice were ~10-11 weeks old, the Esr1 mice were ~10-13 weeks old, and the Stat6- mice were ~6-16 weeks old. The age of the single NSG mouse used for reinfection was unknown.
Fecal samples from infected mice were obtained by housing them on wire mesh flooring over moistened alpha pads. Coprocultures were established as described previously [11] and incubated at 28°C for at least 7 days. Subsequently, iL3s were retrieved using a modified Baermann technique [17], washed, and stored in 25 mL tissue culture flasks. To track the hookworm infection over time, quantitative egg counts were performed at multiple time points using a modified McMaster method [18] until infections were no longer patent or animals were euthanized to determine worm burdens.
Stat6- mice were euthanized on day 21 PI and the small intestines were carefully excised, placed in Petri dishes containing PBS, and incubated at 37°C on a slide warmer. The intestines were cut longitudinally to expose the epithelial lining, allowing the worms to naturally detach for up to 60 minutes before being collected for counting. NOD scid mice were dissected at 7- and 11-days PI to ascertain the timing of larval expulsion compared to an immunocompetent host. To investigate whether hypobiosis occurs with A. caninum in NOD scid and Stat6- mice, one NOD scid mouse was euthanized at 17 days PI, and two Stat6- mice were euthanized at 10 days PI. Their musculoskeletal tissue, excluding the cranium, viscera, limbs, and tail, was carefully cut into fine pieces and placed in Baermann funnels overnight with PBS to extract any arrested larvae from the tissues [17].
To assess the viability of Stat6--derived A. ceylanicum relative to those derived from NSG and hamster hosts reported previously [10], 3 male hamsters were each infected with 80 Stat6--derived A. ceylanicum iL3 larvae. Prior to euthanasia on day 21 PI, quantitative fecal egg counts were performed. Following euthanasia, the small intestines of each hamster were carefully removed, placed in PBS buffer at 37°C, and dissected longitudinally to retrieve and enumerate the adult worms.
To compare worm burdens across groups, a one-way analysis of variance (ANOVA) was performed using the stats package in R (version 4.2.0). ANOVA was selected as it is suitable for comparing the means of more than two independent groups when assumptions of normality and equal variances are reasonably met. Post-hoc pairwise comparisons were conducted using Tukey's Honest Significant Difference test to identify statistically significant differences between groups. Worm burdens were compared in hamsters infected with A. ceylanicum derived from Stat6-, NSG, or hamster hosts, and separately in Stat6- and NSG mice infected with A. ceylanicum. Statistical significance was assessed at a threshold of p<0.05 for all analyses. Data visualization was performed using GraphPad Prism (version 10), as well as Python (version 3.12.1) with the pandas library (version 2.2.0) for data handling and the matplotlib library (version 3.5.2) for plotting.
3. Results
3.1. The Innate Immune System Is Responsible for Protection Against Hookworm Infection in Mice
The developmental progression of A. ceylanicum in male and female NOD scid mice infected with ~140 iL3s showed that only a few developing worms were present in the small intestine at 7 days PI, with none detected by 11 days PI. This pattern of infection is consistent with previously reported results in non-permissive, immunocompetent Swiss Webster mice (Figure 1) [10]. Three female Stat6- mice infected with ~140 A. ceylanicum iL3s had high worm burdens of 25, 35, and 38 adult worms in the small intestine at day 21 PI, which were not significantly different from the burdens observed in permissive NSG mice (p > 0.05) [10]. In contrast, three Stat6- male mice infected with the same dose of iL3s contained significantly lower burdens of 6, 6, and 3 adult worms, which were statistically different from those observed in both female Stat6- mice (p = 0.006) and NSG mice (p = 0.030). Notably, the mouse with the smallest burden had only female worms, while the other two mice had worms of both sexes. Together, these findings suggest that permissiveness to hookworm infection is mediated by the innate immune system in mice and may vary by sex.
3.2. The Stat6 Pathway Is Responsible for Hookworm Protection in Female Mice
Two male and female Stat6- mice were infected with ~140 A. ceylanicum iL3 and checked for patency beginning on day 21 PI. Pooled egg counts from the females first appeared positive on day 21, reaching a peak of 6400 eggs per gram (EPG) of feces. The infection in the female mice remained patent until day 61 PI (Figure 2). Eggs were never detected in feces from the infected male mice. Approximately 4 weeks after the female mice became egg-negative, as confirmed by three zero EPG readings over the span of 4 days, two female and two male mice were reinfected with ~140 A. ceylanicum iL3s. During this secondary infection, only the females became patent. The pooled EPG from the cage housing the female mice reached 4,500 on day 24 PI, peaking at 7,200 on day 26 PI. The infection remained patent until day 77 PI, with a final EPG of 1,500. It is important to note that the secondary infection data are derived from a different primary infection dataset than the one reported here, due to technical difficulties encountered during the original primary infection.
Two male and two female NOD scid mice were infected with ~140 A. ceylanicum iL3s; however, neither sex developed patent infections. Furthermore, six female Esr1 mice and six female Black 6N mice, each infected with ~140 A. ceylanicum iL3s, failed to develop patent infections, as confirmed by multiple EPG checks. Of these, three mice from each strain were euthanized on day 21 PI to check for the presence of adult worms, but none were detected. The remaining three mice from each strain continued to be monitored for egg output, without any evidence of patency. Finally, one male NSG mouse was reinfected to assess whether it had developed protection from the primary infection, as our previous findings indicated that naïve NSG mice are permissive. This mouse became patent on day 21 PI with an EPG of 5,300, which peaked at 37,800 EPG on day 31 PI, and remained patent until day 97 PI.
These infection outcomes were plotted and visually compared to the progression observed in permissive hamsters infected with 150 A. ceylanicum iL3s, as well as NSG mice infected with ~140 A. ceylanicum iL3s from previously published data (Figure 2) [10,19]. It is important to note that both the reinfected Stat6- female mice and the reinfected NSG mouse were not the same animals reported in the primary infection data; however, they had each been infected and had previously achieved patency.
3.3. Infectivity of Stat6- Derived A. ceylanicum Larvae for a Permissive Host
To determine if iL3 derived from Stat6- mice were viable and infectious, three male hamsters were infected with 80-100 Stat6--derived A. ceylanicum iL3s, and their infectivity was compared to that of NSG- and hamster-derived A. ceylanicum as established previously [10]. Adult worm burdens in the Stat6--derived group were 48, 51, and 75 (mean 58.0 ± 14.8 SD) (Table 1). A statistical analysis using ANOVA revealed no statistically significant differences in worm burdens among the three groups (Stat6--, NSG-, and hamster-derived A. ceylanicum) (p > 0.05). This suggests that the infectivity of the Stat6--derived larvae is similar to that of the NSG- and hamster-derived larvae, with no significant differences in their ability to establish patent infections in the host. The pooled EPG for the three hamsters infected with Stat6--derived larvae was 533, resulting in an average EPG of 178 per hamster. For the two hamsters infected with NSG-derived larvae, the pooled EPG was 4,100, with an average of 2,050 per hamster. Meanwhile, the two hamsters infected with hamster-derived larvae had a pooled EPG of 1,500, averaging 750 per hamster. Notably, the fecal output from the hamsters infected with Stat6--derived worms was lower, which may affect the accuracy of the calculated EPG. However, these EPG values confirm that all groups had reached patency by 21 days PI.
3.4. Ancylostoma Caninum Fails to Develop in Immunodeficient Stat6- and NOD Scid Mice
To investigate whether the ability to develop in immunodeficient mouse strains differs between A. ceylanicum and A. caninum, as previously observed in NSG mice, two NOD scid mice were infected with 1,000 A. caninum iL3s, and four Stat6- mice were infected with 200 A. caninum iL3s. In the two male NOD scid mice infected with A. caninum, one L3 larva was found in the small intestine on day 7 PI (Table 2). On day 17 PI, musculoskeletal tissue (8.51 g) of the second mouse was examined for hypobiotic larvae, but none were found. No worms were found in the small intestine of two male and two female Stat6- mice on day 10 PI. The musculoskeletal tissue of one male and one female was examined, revealing 2 L3 larvae in the female (8.26 g) and 1 L3 larva in the male (8.51 g) (Table 2). No adult worms were found at any time point. This supports the observation that, unlike in A. ceylanicum infections, host specificity is not mediated by the immune system in A. caninum.
4. Discussion
This study examined the development of the generalist A. ceylanicum and the specialist A. caninum across four different mouse models: two immunodeficient models (Stat6- and NOD scid mice), an Esr1 KO model, and an immunocompetent control (Black 6N). In Stat6- mice, females uniquely supported the A. ceylanicum full life cycle, reaching patency by day 21 PI and sustaining patent infections until day 61 PI, while infections in males carried significantly lower worm burdens on 21 days PI, and did not reach patency. In NOD scid mice, a few developing larvae were observed in the small intestine at day 7 PI, but none were detected by day 11 PI, indicating that innate immune mechanisms likely contribute to parasite clearance in these mice. Both male and female Esr1 KO mice were non-permissive, with no patent infections observed, suggesting that estrogen receptor activity does not influence host permissiveness to A. ceylanicum. Additionally, larvae derived from female Stat6- mice were viable and infectious to permissive hamsters, confirming that these mice serve as competent hosts for A. ceylanicum. In contrast, A. caninum did not reach patency or establish successfully in either Stat6- or NOD scid mice, implying that it may utilize alternative survival strategies independent of the host immune landscape.
The sex difference observed in the permissiveness of Stat6- mice highlights an intriguing aspect of the host response, aligning with previous reports of sex-specific immune responses to helminth infections. Studies have frequently documented higher hookworm burdens in males [20,21], a pattern that may result from behavioral factors, such as greater exposure, as well as physiological differences in immune response associated with sex hormones. This trend suggests that hookworms may have evolved to exploit male hosts more effectively, possibly due to hormonal influences on immunity. However, male-biased parasitism is not the general rule for all helminths, as females can have higher prevalence and/or intensity in selected helminth infections, or no difference may exist [22]. These physiological differences are frequently attributed to the impact of hormones on immune function. Conversely, in females, it has been documented that species like A. duodenale and A. caninum can undergo hypobiosis and vertical transmission to offspring through milk [23,24]. This may be an adaptive strategy where larvae arrest development and remain infectious, potentially influenced by the host's hormonal or immune environment. These strategies suggest a sophisticated evolutionary tactic tailored to exploit different physiological conditions within male and female hosts. However, such adaptive mechanisms vary between generalists and specialists, as vertical transmission has not been observed in A. ceylanicum, possibly due to its capability to easily exploit new host species without relying on this method.
Sex hormones, including estrogens, progestogens, and androgens, modulate immune responses through their receptors on immune cells, hence potentially impacting susceptibility to hookworm infections [25]. Experiments in latently infected lactating dogs have shown that the exogenous administration of estrogen and prolactin stimulates the resurgence of larval output in the milk, highlighting the potential for these hormones to trigger reactivation of hypobiotic larvae [26]. Moreover, both estrogen and prolactin play a role in upregulating transforming growth factor-β2 (TGF-β2), a cytokine linked to immune regulation during pregnancy [27]. The involvement of TGF-β in the activation of Caenorhabditis elegans larvae from their dormant dauer stage, along with its notable effects in stimulating canine hookworms, reinforces the theory that TGF-β originating from the host could act as a trigger for the reactivation of dormant larvae [28]. Although estrogen alone does not directly trigger feeding or reactivation in tissue-arrested A. caninum larvae, both TGF-β isoforms 1 and 2 have significant stimulatory effects on tissue-arrested larvae [29], highlighting the hormone-regulated pathway's role in their reactivation. Hence, high levels of estrogen could signal a robust immune environment, leading to unfavorable conditions for hookworm development during early infection, while these same levels may also signal the opportunity for vertical transmission.
Additionally, the findings that glucocorticoid treatment facilitates the development of A. ceylanicum iL3 in non-permissive mouse hosts [8], may reflect the underlying influence of estrogen, given that glucocorticoids have been found to inhibit the immune-stimulating properties of estrogen [30,31,32]. This inhibitory action aligns with the broader immunomodulatory role of estrogen. Notably, female mice have shown a greater dependency on Stat6 for the production of Th2 cytokines and eosinophils following IL-33 treatment compared to their male counterparts [33]. This observation led to the hypothesis that the reliance on Stat6 in females might be linked to higher levels of estrogen, especially considering that estrogen receptor-alpha (ER-α) signaling has been implicated in amplifying IL-33 release and group 2 innate lymphoid cell (ILC2)-mediated airway inflammation [34]. Estrogen typically acts as a regulator in the immune system, often suppressing Th1-mediated inflammation while enhancing Th2 responses [35]. However, the lack of permissiveness in female Esr1 KO mice to hookworm establishment, as observed herein, challenges the direct involvement of estrogen signaling through ER-α in protection against hookworms. This suggests that Stat6 might still function effectively in mediating immune responses against hookworms in the absence of direct estrogen signaling through ER-α, potentially involving other hormone receptors and their ligands.
It is essential to identify which pathways confer protective immunity in male Stat6- mice. This investigation should include exploring alternative mechanisms, such as Th1 responses which males may rely on more due to the nature of estrogen in driving Th2 responses in females. Additionally, the potential role of Stat5, capable of initiating Th2 polarization independently of Gata3, should be examined to assess its possible protective mechanism in the absence of Stat6 [36]. These pathways may provide compensatory immune responses in the absence of Stat6, a hypothesis that needs to be tested to fully understand the protective immune dynamics in male mice.
Further investigation is also necessary to elucidate the specific components of the Stat6 pathway responsible for protection, including both upstream and downstream elements such as IL-4, IL-13, and eosinophil responses. These components have been implicated in promoting worm clearance in studies involving other parasites [37,38]. These studies could lead to targeted interventions that enhance host resistance against hookworm establishment. Insights into the specific cytokine and cellular responses, specifically those governed by the innate immune system, are critical for developing effective treatment strategies. Importantly, understanding the sex-dependent differences in immune responses is essential, as treatments might need to differ between sexes based on these underlying immunological mechanisms.
In this study, no A. caninum larvae were found arrested in the male NOD scid mouse, while two larvae were detected in one female Stat6- mouse, and one larva in another male Stat6- mouse. Research involving other non-permissive mouse models has shown that larvae from A. braziliense and A. tubaeforme can survive within the anterior somatic musculature of mice following oral infection, and some larvae have even been detected in the brains of these animals [39]. Additionally, A. caninum larvae have been found in the head, thorax, and abdomen of both bred and unbred BALB/c mice following subcutaneous infection [40]. Given that the scope of our investigation of hypobiotic properties in musculoskeletal tissue was limited to a single mouse from each strain, plus a relatively low inoculum and considerably less than quantitative recovery of larvae from dissected tissue, a comprehensive examination of additional tissues, including the brain and other organs, is warranted to fully understand the hypobiotic potential of A. caninum across various mouse models, including both immunodeficient and immunocompetent mice.
5. Conclusions
Our study highlights the essential role of the innate immune system in determining host specificity in A. ceylanicum infections. The distinctions observed in hookworm development of male and female Stat6- mice underscore the complexity of host-parasite interactions. In contrast, A. caninum did not establish in either Stat6- or NOD scid mice, and minimal or no hypobiotic larvae were observed, suggesting that its survival strategies do not depend on the host's immune landscape. Moving forward, leveraging these insights will be invaluable in developing tailored approaches to control and treat hookworm infections, aiming to reduce their extensive burden on global health.
Author Contributions
Conceptualization, A.L. and J.M.H.; Methodology, A.L. and J.M.H.; Software, A.L.; Validation, A.L.; Formal Analysis, A.L.; Investigation, A.L., C.A.J., E.L.M. and H.E.; Resources, J.M.H.; Data Curation, A.L.; Writing – Original Draft Preparation, A.L. and J.M.H.; Writing – Review & Editing, A.L., J.M.H., D.M.O., E.L.M., C.A.J. and H.E.; Visualization, A.L. and C.A.J.; Supervision, J.M.H. and D.M.O.; Project Administration, J.M.H.; Funding Acquisition, J.M.H. and D.M.O.
Funding
This research was funded by grant R21AI173403 awarded to J.M.H. from the National Institute of Allergy and Infectious Diseases of the National Institutes of Health and the Harlan Endowment to the Biology department at George Washington University.
Institutional Review Board Statement
The animal study was conducted according to the guidelines of the National Institute of Health (USA) Guide for the Care and Use of Laboratory Animals and the US Department of Agriculture Animal Welfare Act [41,42]. The animal protocols were approved by The George Washington University's Institutional Animal Care and Use Committee (USA) (protocol A2021-017 approved on 9/17/21, and protocol A2021-016 approved on 9/13/21).
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
The authors would like to thank Sunny Shih, Meghan Brink, and Elodie Krawczyk for hookworm strain maintenance, and the Fernandes Lab for donating NSG mice.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| STAT6 | Signal transducer and activator of transcription 6 |
| STN | Soil transmitted nematodes |
| PI | Post-infection |
| NSG | NOD scid gamma |
| Esr | Estrogen receptor 1 |
| iL3 | Infective third-stage larvae |
| EPG | Eggs per gram |
| Black 6N | C57BL/6J |
| IL | Interleukin |
| TGF-β | Transforming growth factor-β |
| MS | Muscoskeletal |
References
- R. L. Pullan, J. L. Smith, R. Jasrasaria, and S. J. Brooker, “Global numbers of infection and disease burden of soil transmitted helminth infections in 2010,” Parasit Vectors, vol. 7, p. 37, Jan. 2014. [CrossRef]
- T. W. Gyorkos and N. L. Gilbert, “Blood Drain: Soil-Transmitted Helminths and Anemia in Pregnant Women,” PLoS Neglected Tropical Diseases, vol. 8, no. 7, p. e2912, Jul. 2014. [CrossRef]
- P. M. Jourdan, P. H. L. Lamberton, A. Fenwick, and D. G. Addiss, “Soil-transmitted helminth infections,” Lancet, vol. 391, no. 10117, pp. 252–265, Jan. 2018. [CrossRef]
- B. Lozoff, E. Jimenez, and A. W. Wolf, “Long-term developmental outcome of infants with iron deficiency,” N Engl J Med, vol. 325, no. 10, pp. 687–694, Sep. 1991. [CrossRef]
- H. Sakti et al., “Evidence for an association between hookworm infection and cognitive function in Indonesian school children,” Trop Med Int Health, vol. 4, no. 5, pp. 322–334, May 1999. [CrossRef]
- S. Brooker, P. J. Hotez, and D. A. P. Bundy, “Hookworm-Related Anaemia among Pregnant Women: A Systematic Review,” PLOS Neglected Tropical Diseases, vol. 2, no. 9, p. e291, Sep. 2008. [CrossRef]
- D. E. Manzoli, M. J. Saravia-Pietropaolo, S. I. Arce, A. Percara, L. R. Antoniazzi, and P. M. Beldomenico, “Specialist by preference, generalist by need: availability of quality hosts drives parasite choice in a natural multihost–parasite system,” International Journal for Parasitology, vol. 51, no. 7, pp. 527–534, Jun. 2021. [CrossRef]
- D. K. Ray, K. K. Bhopale, and V. B. Shrivastava, “Development of Ancylostoma ceylanicum Looss, 1911 (hamster strain) in the albino mouse, Mus musculus, with and without cortisone,” Parasitology, vol. 71, no. 2, pp. 193–197, Oct. 1975. [CrossRef]
- R. D. Bungiro Jr., B. R. Anderson, and M. Cappello, “Oral Transfer of Adult Ancylostoma ceylanicum Hookworms into Permissive and Nonpermissive Host Species,” Infect Immun, vol. 71, no. 4, pp. 1880–1886, Apr. 2003. [CrossRef]
- Langeland, E. L. McKean, D. M. O’Halloran, and J. M. Hawdon, “Immunity mediates host specificity in the human hookworm Ancylostoma ceylanicum,” Parasitology, vol. 151, no. 1, pp. 102–107, 2024. [CrossRef]
- Langeland, E. Grill, A. C. Shetty, D. M. O’Halloran, and J. M. Hawdon, “Comparative transcriptomics from intestinal cells of permissive and non-permissive hosts during Ancylostoma ceylanicum infection reveals unique signatures of protection and host specificity,” Parasitology, vol. 150, no. 6, pp. 511–523, 2023. [CrossRef]
- L. D. Shultz et al., “Human lymphoid and myeloid cell development in NOD/LtSz-scid IL2R gamma null mice engrafted with mobilized human hemopoietic stem cells,” J Immunol, vol. 174, no. 10, pp. 6477–6489, May 2005. [CrossRef]
- M. Bosma, W. Schuler, and G. Bosma, “The scid mouse mutant,” Curr Top Microbiol Immunol, vol. 137, pp. 197–202, 1988. [CrossRef]
- M. H. Kaplan, U. Schindler, S. T. Smiley, and M. J. Grusby, “Stat6 is required for mediating responses to IL-4 and for development of Th2 cells,” Immunity, vol. 4, no. 3, pp. 313–319, Mar. 1996. [CrossRef]
- S. C. Hewitt, G. E. Kissling, K. E. Fieselman, F. L. Jayes, K. E. Gerrish, and K. S. Korach, “Biological and biochemical consequences of global deletion of exon 3 from the ER alpha gene,” FASEB J, vol. 24, no. 12, pp. 4660–4667, Dec. 2010. [CrossRef]
- S. Kitchen et al., “Isolation and characterization of a naturally occurring multidrug-resistant strain of the canine hookworm, Ancylostoma caninum,” Int J Parasitol, vol. 49, no. 5, pp. 397–406, Apr. 2019. [CrossRef]
- G. Baermann, “A simple method for the detection of Ankylostomum (nematode) larvae in soil tests.,” A simple method for the detection of Ankylostomum (nematode) larvae in soil tests., pp. 41–47, 1917.
- H. M. Gordon and H. V. Whitlock, “A new technique for counting nematode eggs in sheep faeces,” Journal of the Council for Scientific and Industrial Research, vol. 12, no. 1, pp. 50–52, 1939.
- S. C. Pan et al., “Cognitive and Microbiome Impacts of Experimental Ancylostoma ceylanicum Hookworm Infections in Hamsters,” Sci Rep, vol. 9, p. 7868, May 2019. [CrossRef]
- S. Brooker, J. Bethony, and P. J. Hotez, “Human hookworm infection in the 21st century,” Adv Parasitol, vol. 58, pp. 197–288, 2004. [CrossRef]
- R. J. Quinnell et al., “Genetic and household determinants of predisposition to human hookworm infection in a Brazilian community,” The Journal of infectious diseases, vol. 202, no. 6, p. 954, Sep. 2010. [CrossRef]
- Wesołowska, “Sex—the most underappreciated variable in research: insights from helminth-infected hosts,” Veterinary Research, vol. 53, no. 1, p. 94, Nov. 2022. [CrossRef]
- G. A. Schad, “Hypobiosis and related phenomena in hookworm infection.,” Hookworm disease - current status and new directions., pp. 71–88, 1990.
- W. M. Stone and J. C. Peckham, “Infectivity of Ancylostoma caninum larvae from canine milk,” Am J Vet Res, vol. 31, no. 9, pp. 1693–1694, Sep. 1970.
- Bereshchenko, S. Bruscoli, and C. Riccardi, “Glucocorticoids, Sex Hormones, and Immunity,” Front Immunol, vol. 9, p. 1332, 2018. [CrossRef]
- M. Stoye and J. Krause, “Reactivation of inhibited larvae of Ancylostoma caninum. Effect of estradiol and progesterone,” Zentralbl Veterinarmed B, vol. 23, no. 10, pp. 822–839, Dec. 1976.
- S. L. Schneider, S. O. Gollnick, C. Grande, J. E. Pazik, and T. B. Tomasi, “Differential regulation of TGF-beta 2 by hormones in rat uterus and mammary gland,” J Reprod Immunol, vol. 32, no. 2, pp. 125–144, Dec. 1996. [CrossRef]
- H. A. Tissenbaum, J. Hawdon, M. Perregaux, P. Hotez, L. Guarente, and G. Ruvkun, “A common muscarinic pathway for diapause recovery in the distantly related nematode species Caenorhabditis elegans and Ancylostoma caninum,” Proc Natl Acad Sci U S A, vol. 97, no. 1, pp. 460–465, Jan. 2000. [CrossRef]
- P. Arasu, “In vitro reactivation of Ancylostoma caninum tissue-arrested third-stage larvae by transforming growth factor-beta,” J Parasitol, vol. 87, no. 4, pp. 733–738, Aug. 2001. [CrossRef]
- T. Bever, F. L. Hisaw, and J. T. Velardo, “Inhibitory action of desoxycorticosterone acetate, cortisone acetate, and testosterone on uterine growth induced by estradiol-17beta,” Endocrinology, vol. 59, no. 2, pp. 165–169, Aug. 1956. [CrossRef]
- Danisová, E. Seböková, and J. Kolena, “Effect of corticosteroids on estradiol and testosterone secretion by granulosa cells in culture,” Exp Clin Endocrinol, vol. 89, no. 2, pp. 165–173, Apr. 1987.
- T. Harding and N. S. Heaton, “The Impact of Estrogens and Their Receptors on Immunity and Inflammation during Infection,” Cancers (Basel), vol. 14, no. 4, p. 909, Feb. 2022. [CrossRef]
- H. Zhao et al., “Sex Differences in IL-33-Induced STAT6-Dependent Type 2 Airway Inflammation,” Front Immunol, vol. 10, p. 859, May 2019. [CrossRef]
- J.-Y. Cephus et al., “Estrogen receptor-α signaling increases allergen-induced IL-33 release and airway inflammation,” Allergy, vol. 76, no. 1, pp. 255–268, Jan. 2021. [CrossRef]
- M. L. Salem, “Estrogen, a double-edged sword: modulation of TH1- and TH2-mediated inflammations by differential regulation of TH1/TH2 cytokine production,” Curr Drug Targets Inflamm Allergy, vol. 3, no. 1, pp. 97–104, Mar. 2004. [CrossRef]
- E. Maier, A. Duschl, and J. Horejs-Hoeck, “STAT6-dependent and -independent mechanisms in Th2 polarization,” European Journal of Immunology, vol. 42, no. 11, pp. 2827–2833, 2012. [CrossRef]
- J. F. Urban et al., “IL-13, IL-4Ralpha, and Stat6 are required for the expulsion of the gastrointestinal nematode parasite Nippostrongylus brasiliensis,” Immunity, vol. 8, no. 2, pp. 255–264, Feb. 1998. [CrossRef]
- P. H. Gazzinelli-Guimaraes, S. M. Jones, D. Voehringer, K. D. Mayer-Barber, and A. E. Samarasinghe, “Eosinophils as modulators of host defense during parasitic, fungal, bacterial, and viral infections,” Journal of Leukocyte Biology, vol. 116, no. 6, pp. 1301–1323, Dec. 2024. [CrossRef]
- D. E. Norris, “The migratory behavior of the infective-stage larvae of Ancylostoma braziliense and Ancylostoma tubaeforme in rodent paratenic hosts,” J Parasitol, vol. 57, no. 5, pp. 998–1009, Oct. 1971.
- P. Arasu and D. Kwak, “Developmental Arrest and Pregnancy-Induced Transmammary Transmission of Ancylostoma caninum Larvae in the Murine Model,” The Journal of Parasitology, vol. 85, no. 5, pp. 779–784, 1999. [CrossRef]
- National Research Council, Guide for the Care and Use of Laboratory Animals. Washington, D.C.: National Academies Press (US), 2011.
- (United States Department of Agriculture) USDA, Animal Welfare Act. Washington, D.C.: U.S. Government Publishing Office., 2018.
Figure 1.
Development of Ancylostoma ceylanicum in immunocompetent and immunodeficient mouse strains over time. The x-axis shows the number of days post-infection with approximately 140 A. ceylanicum infective larvae, and the y-axis indicates the number of developed worms recovered from the small intestine. Each data point represents the mean worm burden, with error bars indicating the standard deviation for each group. Data for Swiss Webster control mice (purple) and NSG mice (green), previously published [10], are included for comparison. In this study, NOD scid mice (blue) and Stat6- female (pink) and male (red) mice each had three biological replicates per time point.
Figure 1.
Development of Ancylostoma ceylanicum in immunocompetent and immunodeficient mouse strains over time. The x-axis shows the number of days post-infection with approximately 140 A. ceylanicum infective larvae, and the y-axis indicates the number of developed worms recovered from the small intestine. Each data point represents the mean worm burden, with error bars indicating the standard deviation for each group. Data for Swiss Webster control mice (purple) and NSG mice (green), previously published [10], are included for comparison. In this study, NOD scid mice (blue) and Stat6- female (pink) and male (red) mice each had three biological replicates per time point.
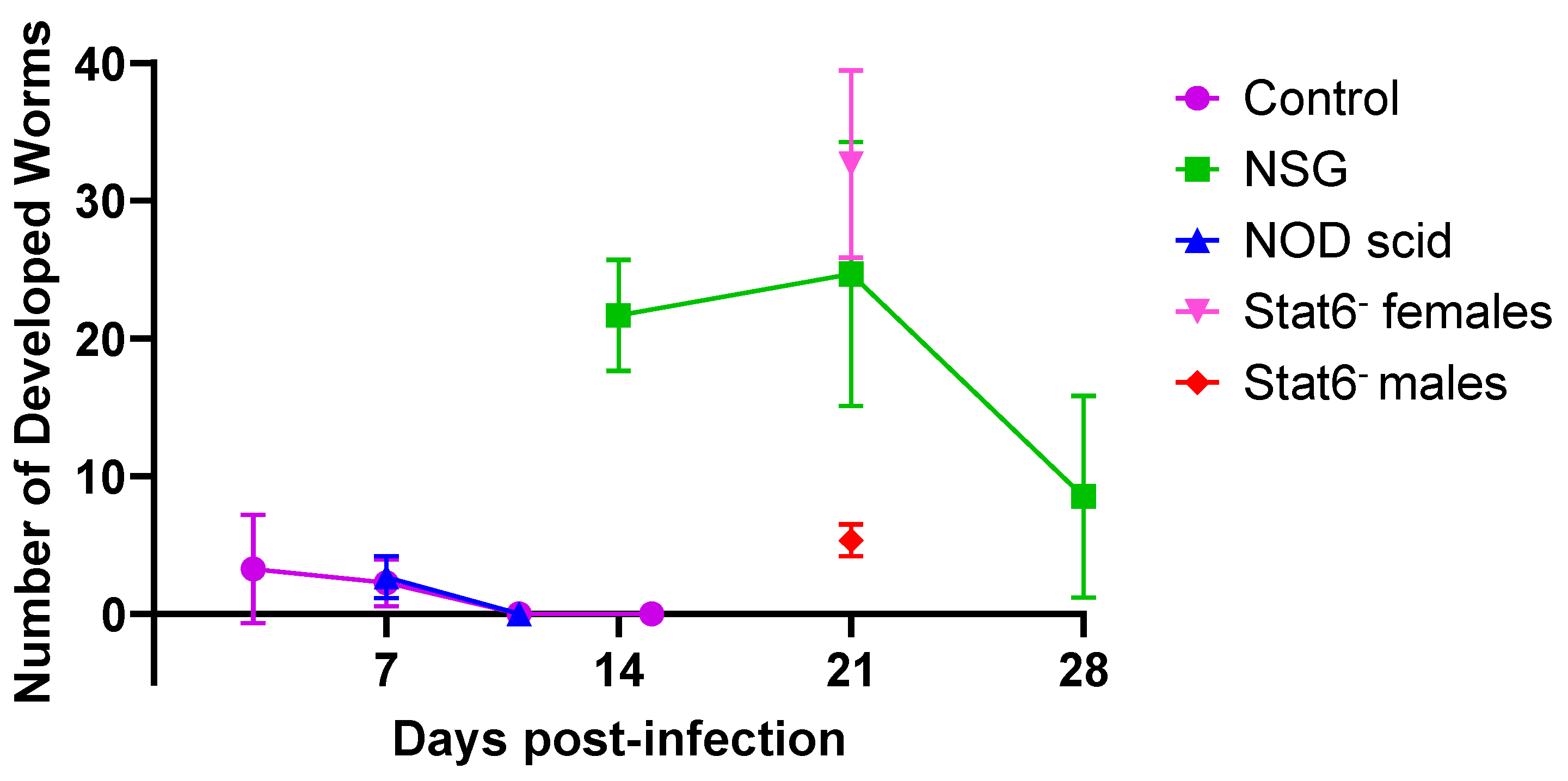
Figure 2.
Fecal egg count over time in Stat6- and NSG mice following primary and secondary Ancylostoma ceylanicum infections. Fecal pellets were collected on the indicated days post-infection (PI). Parasite egg counts are expressed as fecal eggs per gram (EPG), with values representing pooled counts from cages housing the specified animals. The green horizontal dashed line from 19 to 59 days PI indicate the presence of eggs without quantitative EPG data during the first NSG mice infection, as previously published [10]. Both Stat6- infections had EPG values pooled from two female mice housed in a single cage. For the NSG mice, the first fecal EPG assessment included three individual mice until day 77 PI, followed by a second infection with a single mouse. Hamsters were housed individually, with the number of animals ranging from n=6 to n=51 for each time point as described by Pan et al., 2019 [19].
Figure 2.
Fecal egg count over time in Stat6- and NSG mice following primary and secondary Ancylostoma ceylanicum infections. Fecal pellets were collected on the indicated days post-infection (PI). Parasite egg counts are expressed as fecal eggs per gram (EPG), with values representing pooled counts from cages housing the specified animals. The green horizontal dashed line from 19 to 59 days PI indicate the presence of eggs without quantitative EPG data during the first NSG mice infection, as previously published [10]. Both Stat6- infections had EPG values pooled from two female mice housed in a single cage. For the NSG mice, the first fecal EPG assessment included three individual mice until day 77 PI, followed by a second infection with a single mouse. Hamsters were housed individually, with the number of animals ranging from n=6 to n=51 for each time point as described by Pan et al., 2019 [19].
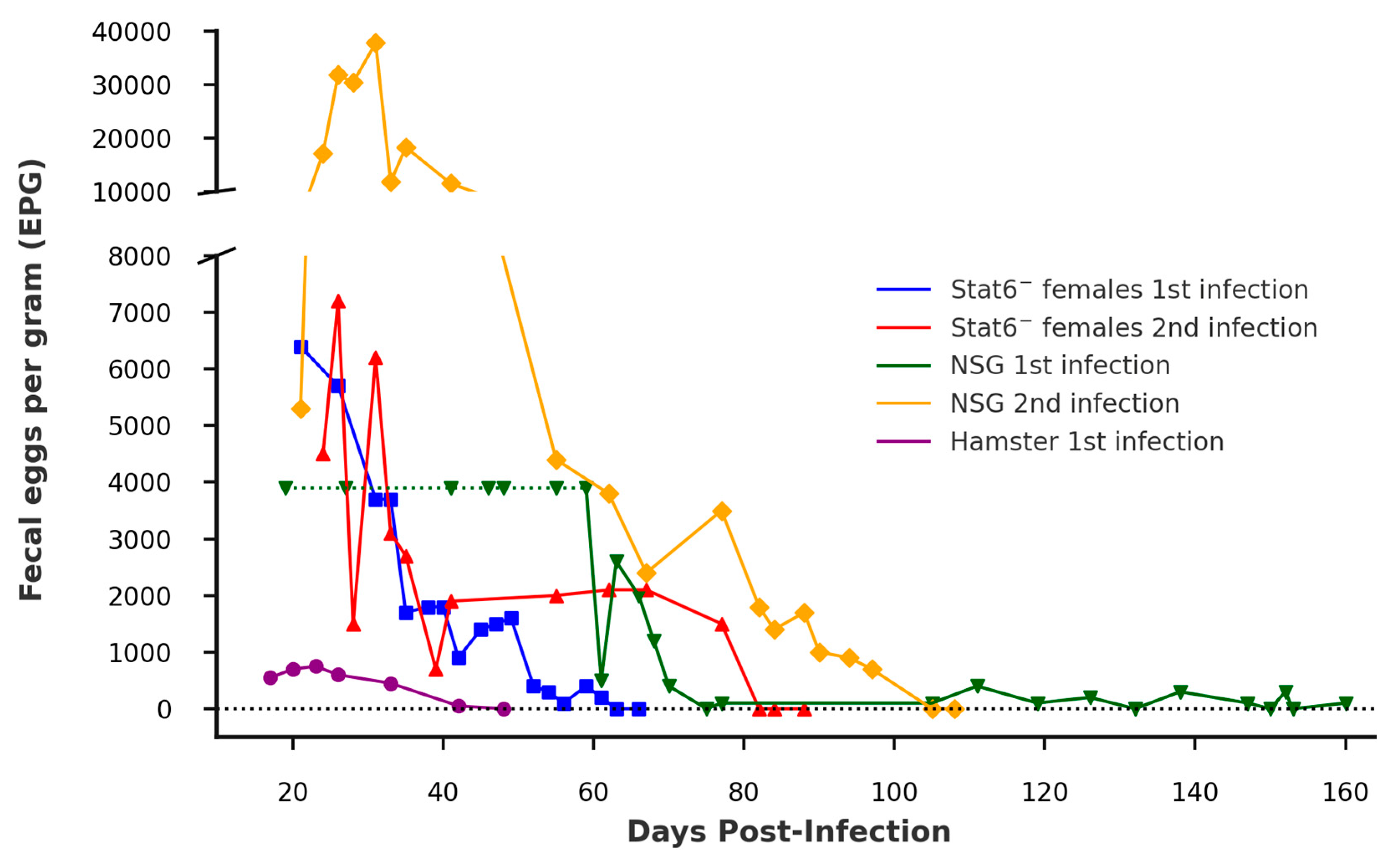

Table 1.
Adult worm burden and fecal eggs per gram in hamsters infected with different isolates of Ancylostoma ceylanicum larvae.
Table 1.
Adult worm burden and fecal eggs per gram in hamsters infected with different isolates of Ancylostoma ceylanicum larvae.
| Hamster | Sex | Infection Isolate | Worm burden | Mean ± SD | Combined EPG |
|---|---|---|---|---|---|
| 1 | M | Stat6- derived A. cey | 48 | 58.0 ± 14.8 | 533 |
| 2 | M | Stat6- derived A. cey | 51 | ||
| 3 | M | Stat6- derived A. cey | 75 | ||
| 4 | M | NSG derived A. cey* | 22 | 31 ± 12.7 | 4100 |
| 5 | M | NSG derived A. cey* | 40 | ||
| 6 | M | Hamster derived A. cey* | 35 | 36 ± 1.4 | 1500 |
| 7 | M | Hamster derived A. cey* | 37 |
SD: Standard deviation. EPG: Eggs per gram. *Data from Langeland et al., 2024.
Table 2.
Infection outcomes of Ancylostoma caninum in Stat6- and NOD scid mice.
| Mouse | Strain | Sex | Infective dose | Days PI | #L3s in SI | #L3s in MS | MS weight (g) |
|---|---|---|---|---|---|---|---|
| 1 | NOD scid | M | 1000 | 7 | 1 | N/A | N/A |
| 2 | NOD scid | M | 1000 | 17 | 0 | 0 | 8.51 |
| 3 | Stat6- | F | 200 | 10 | 0 | N/A | N/A |
| 4 | Stat6- | F | 200 | 10 | 0 | 2 | 8.26 |
| 5 | Stat6- | M | 200 | 10 | 0 | N/A | N/A |
| 6 | Stat6- | M | 200 | 10 | 0 | 1 | 8.51 |
PI: Post-infection, SI: Small intestine, MS: Musculoskeletal
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



