
Submitted:
15 January 2025
Posted:
15 January 2025
You are already at the latest version
Abstract
This study aimed to evaluate the impact on Chlorella sorokiniana NIVA-CHL 176 growth and the plant biostimulant functions of the microalgal extract of the strigolactone mimic SL-6, in com-parison with the strigolactone analog GR24. Three molar SL-6 concentrations were tested: 10-7 M, 10-8 M, and 10-9 M, respectively. Five parameters of the microalgal growth were assessed: optical density, turbidity, biomass production, chlorophyll fluorescence, and pigment concentration. Results after 15 days of culturing revealed that the SL-6 treatments significantly enhanced bio-mass production, chlorophyll synthesis, and photosystem II activity. Treatment with SL-6 c3 (10-9 M) exhibited significant increases in chlorophyll a (27.55%), chlorophyll b (24.81%), carotenoids (12.31%), and total pigments (24.00%) compared to the solvent control (DMSO). Experimental treatment SL-6 c2 (10-8 M) followed next and recorded notable enhancements in chlorophyll a (25.80%), carotenoids (9.77%), and total pigments (20.08%). Conversely, higher doses of GR24 and SL-6 (10-7 M) inhibited the microalgal growth, reducing cell density, biomass production, and pigment synthesis. The microalgal extracts acted as a plant biostimulant stimulating root and shoot elongation and proton pump functioning in the presence and absence of salt stress. The extracts from SL-6 bio-stimulated C. sorokiniana NIVA-CHL 176 are more active as plant biostimulants.
Keywords:
microalgal biostimulant
; chlorophyll fluorescence
; light stress
; mung seedling
; proton pump
1. Introduction
Microalgae cultivation is more efficient in harvesting light and using mineral nutrients than terrestrial plants [1,2]. Their metabolic flexibility, i.e., ability to use organic nutrients in mixotrophic and heterotrophic modes [3,4] is exploited in bioremediation and wastewater treatment [5-7], and valorization of various agro-industrial by-products, such as whey [8,9], sugar cane vinasse [10,11], wine lees [12-14], ethanol thin-stillage [15], hydrolyzed lignocellulosic biomass [16] or food waste [17]. Microalgal biomass is a feedstock for energy- and material-driven biorefinery systems [18,19]. Due to its high content of bioactive ingredients, the microalgal biomass is a renewable resource for high-value-adding bioproducts, e.g., dietary supplements [20,21] and plant biostimulants [22-24].
Plant biostimulants represent a category of agricultural inputs, defined by their functions related to the improvement of “one or more of the following characteristics of the plant and/or the plant rhizosphere: (1) nutrient use efficiency, (2) tolerance resistance to (a)biotic stress, (3) quality characteristics, or (4) availability of confined nutrients in the soil or rhizosphere”[25]. Various types of extracts from microalgae demonstrated the agricultural functions specific to plant biostimulants, i.e., increased nutrient uptake and nutrient use efficiency [26], enhanced tolerance to different abiotic stress, e.g., water stress [27] or drought [28,29], improved yield quality [30], and enhanced growth and photosynthetic performance [31]. Microalgae were used as terrestrial plant models to screen the bioactive components influencing photosynthesis performance, e.g., whey hydrolysate peptides [32] or hydrophobic contaminants [33].
Strigolactones (SLs) are exo- and endo-signals produced by terrestrial plants, initially identified in root exudates as inducers of seed germination in the Striga parasitic plant [34]. The initially discovered SLs have a complex structure, with a complex ABC-ring system linked through an enol-ether bridge to the bioactiphore, a methyl-butenolide D-ring [35]. Studies from the last decade have demonstrated a complex landscape of strigolactone signal structures. Besides the strigolactone with the complex ABC-ring (called ”canonical strigolactones”), more simple molecules that maintain the bioactive D-ring were discovered [36]. As endo-signals, SLs coordinate plant response to (a)biotic stress [37,38] and integration of metabolic and nutrition signals [39]. SLs functions as exo-signals (ecomones) are both for target organisms, i.e., as pheromones (for detection of neighboring plants) and synomones (exo-signals for arbuscular mycorrhizae fungi, nitrogen-fixing bacteria, and other plant-beneficial microbes) and for eavesdropping organisms, as kairomones (exo-signals for parasitic plants, nematodes and fungal plant pathogens) [40,41].
Due to the difficulty of preparing natural SLs, which are complex and easily degradable structures, synthetic strigolactones were proposed [42]. The synthetic analog GR24 [43] is among the most used synthetic strigolactone for various studies demonstrating agricultural functions similar to plant biostimulants. The exogenous application of the strigolactone analogs GR24 enhanced plant tolerance to drought [44], salt stress [45], heat [46], low temperature [47], and toxic elements such as cadmium (Cd) [48]. Exogenous strigolactone application increased mineral nutrient uptake, including due to the stimulation of mycorrhizal symbiosis [49]. Exogenous application of GR-24 improves artemisinin production in Artemisia annua [50]. The mechanisms involved in the plant biostimulant-like effects of exogenous strigolactone are related to enhanced photosynthetic efficiency and decreased oxidative stress due to the activation of the antioxidant systems in plants [51,52].
Although strigolactones are specific endo- and exo-signals in multicellular photosynthetic organisms, with the first appearance in the green lineage on stoneworts, liverworts and mosses [53], the strigolactones were proved to be active on microalgae as well. 5-Deoxystrigol (5-DS), a canonical strigolactone, applied at 10-11 M concentration, improved the efficiency of a co-culture Scenedesmus obliquus –Ganoderma lucidum in biogas upgrading and wastewater purification [54]. The application of the strigolactone analog GR24 was demonstrated to improve the efficiency of microalgal-based technologies for biogas upgrading and removing nutrients/pollutants from digestate. GR24, applied at a concentration of 10-7 M, increased the growth rate and daily productivity of C. vulgaris FACHB-8 cultivated in a medium that simulates biogas digestate and in the presence of biogas. The increase was due to an enhanced photosynthetic performance [55].
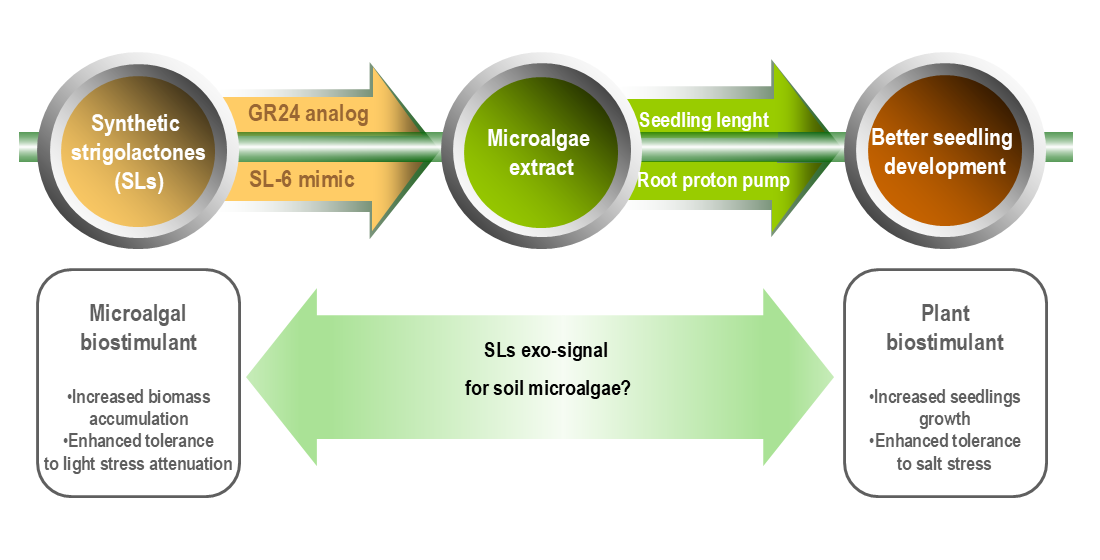
In microalgae, SLs have been reported to have functions similar to those exerted on terrestrial plants, enhanced response to abiotic stress (as endo-signals), and induction of symbiotic interactions (i.e., synomones function). SLs could be considered as “microalgal biostimulants”, a mirror term for products that have microalgal biotechnological functions analog to agricultural function of plant biostimulants. Some of these functions are increased nutrient use efficiency (including due to the enhanced photosynthesis performance), enhanced tolerance to abiotic stress, improved yield quality (bioactive compound accumulation in biomass), enhanced bioavailability of confined nutrients (due to promotion of the symbioses with others organisms) [56]. Other reported “microalgal biostimulant” products are humic substances [57,58] and protein hydrolysates prepared from waste chicken feathers [59]. Microalgal biostimulants are one of the solutions to the challenges that limit microalgal-based biotechnologies development.
The cost of synthesis of strigolactone analogs is still very high. For the scale-up applications, compounds that are easily accessible in significant amounts and safe are needed. Our group demonstrated that strigolactone mimics, synthetic molecules easier to synthesize, which keep only the butenolide active D-ring (more similar to the non-cano-nical natural strigolactone structure) are active on microalgae. The SL-F3 mimic, 3-(4-methyl-5-oxo-2,5-dihydrofuran-2-yl)-3H-benzothiazol-2-one (7), at a concentration of 10−7 M, increased the biomass accumulation due to an improved photosynthetic efficiency. The SL mimics supported microalga adaptation to the light stress determined by the continuous illumination [60].
We have previously reported the synthesis of another SL mimic, SL-6, active in inducing parasitic seed germination and modification of phytopathogenic fungi branching [61]. We improved the process for multi-gram synthesis, and we demonstrated that SL-6 has a synomone function, increasing the biofilm formation by nodulation-enhancing bacteria [62]. We recently assessed SL-6 for ecotoxicological impact on marine and freshwater organisms, including microalgae, Raphidocelis subcapitata and Skeletonema pseudocostatum. The estimated EC50 was 1.21 mg/L after 48 h exposure, corresponding to 0.4 ×10-5 M [63].
This study aimed to investigate the effect of SL-6, at concentrations significantly lower than EC50, on the growth of C. sorokiniana NIVA-CHL 17 and the plant biostimulant effect of the extract prepared from SL-6 treated microalgae in comparison to non-treated microalgae. The C. sorokiniana NIVA-CHL 17 extract was reported to stimulate the root elongation of Arabidopsis thaliana (ecotype Columbia) and the yield and photosynthetic performance of lettuce (Lactuca sativa L. cv. Finstar) [31]. We used GR24 as a positive control due to its demonstrated effect on microalgae from the Chlorella genus [64]. The microalgal extract was tested on mung seedlings in the presence and absence of salt stress to check the plant biostimulant effects. To the best of our knowledge, this is the first study to investigate (SLs) biostimulated microalgal biomass as a source of extract with (enhanced) plant biostimulant functions.
2. Results
2.1. Microalgal Growth Parameters
2.1.1. Optical Density
The optical density showed statistically significant effects of both the GR24 analogue and SL-6 mimic on C. sorokiniana compared to controls (Figure 1).
From the fifth day of culturing marginal significant differences emerged between the control group and SC group and the experimental treatments with synthetic SLs. After a week of culturing, the growth rate was significantly stimulated by c2 and c3 of GR24 and marginally significant stimulated by the lowest concentration of mimic SL-6, c3. At the end of the culturing time, at day 15, the growth rates of the variants with GR24 c2 and GR24 c3 were 12.11 ± 8.25 % and 17.68 ± 2.54 %, respectively, higher compared to SC. The growth rates of the variants with SL-6 c2 and SL-6 c3 were 7.00 ± 0.80 % and 16.26 ± 5.00 %, respectively, higher than SC. The stimulatory effect of SL-6 on C. sorokiniana was slightly lower, but similar to the stimulatory effect of GR24.
The highest concentration tested, c1, (1 × 10-7 M) of both GR24 and SL-6 induced similar or lower optical densities compared to SC, which indicates a tendency towards inhibition of microalgal growth of applied synthetic strigolactones at higher concentrations.
2.1.2. Microalgal Culture Turbidity
The turbidity measurements recorded values that followed the same trend as optical density (Figure 2).
The lower molar concentrations of 10-8 and 10-9 M stimulated the microalgal cell density with statistical significance from day 5 to day 15, compared to SC. The treatments with SL-6 c2 and c3 recorded slightly lower turbidity levels than the same concentrations of GR24, but higher than controls, starting from day 7 to the end of the culturing period. On day 15 the treatment with SL-6, 1 × 10-9 M exhibited McFarland values increased with 8.17 ± 1.3 %compared to SC.
The treatments with the highest dose of both strigolactones consistently exhibited the lowest turbidity values, lower than controls, a trend observed from day 5 to the end of the experiment, especially in the case of SL-6.
2.1.3. Biomass Quantification
The microalgal biomass resulted after two weeks of culturing was increased compared to control when the lower concentrations of SLs were applied (Figure 3). Similar to previous parameters, SL-6 induced slightly lower biomass than GR24, but higher than the controls. The lowest production of microalgal biomass was recorded in the variant with 10-7 M for both GR24 and SL-6, a little lower than the controls.
Comparing each treatment with SC, we observed distinct responses: GR24 c1 and SL-6 c1 decreased slightly the biomass by 3.96 ± 4.85 %, and 6.74 ± 4.1%, respectively; GR24 c2 and c3 increased the biomass by 26.35 ± 12.47 % and 33.17 ± 10.1 %, respectively; SL-6 c2 and c3 increased the biomass by 8.47 ± 2.55 % and 13.53 ± 5.53 %, respectively.
2.1.4. Extracted Pigments Concentration
Microalgal biomass is a source of valuable compounds such as pigments, therefore we analyzed the chlorophyll and carotenoids concentrations within the cells.
Results followed the same trend as biomass development, where higher dose treatments slightly inhibited pigment formation whereas lower concentrations stimulated chlorophyll synthesis (Figure 4).
The strigolatone analog GR24 c2 (1 × 10-8 M) and c3 (1 × 10-9 M) induced substantial increases in ChlA compared to SC, of 33.88 ±4.18 % and 39.49 ± 11.4 %, respectively (Figure 4a). SL-6 c2 and c3 showed increases in ChlA content by 25.80 ± 8.12 % and 27.55 ± 8.24 %, respectively, relative to SC. The ChlB production was stimulated the most by the treatment with GR24 c3, with a difference of approximately 86.14 ± 15.17 % compared to SC (Figure 4b). SL-6 c2 and c3 exhibited marginally significant increases in ChlB by 6.38 ± 2.4 % and 24.81 ± 4.45 %, respectively, relative to SC.
Chlorophyll a and chlorophyll b are pigments crucial for photosynthesis in microalgae. ontributing to adaptability to various environmental conditions [65]. The treatments with SL-6 c2 and SL-6 c3 increased the total carotenoids by 9.77 ± 5.2 % and 12.31 ± 5.6 %, respectively, compared to the solvent control (Figure 4c). The same concentrations of the analogue strigolactone GR24 recorded even more notable increases in total carotenoids compared to SC, with approximately 18.57 ± 6.71 % and 16.64 ± 4.11 %, respectively. SL-6 c1 (10-7 M) exhibited a decrease in total carotenoids by about 19.00 ± 6.23 % compared to SC, whereas the same concentration of GR24 did not have an effect on the total carotenoids. The total pigments followed the same trend as its components. The lower doses, c2 and c3, of GR24 used in C. sorokiniana cultures recorded significant increases in total pigments compared to SC, with differences of 26.52 ± 6.82 % and 48.89 ± 14.75 %, respectively. The mimic SL-6 showed increases in the total extracted pigment quantities, by 20.08 ± 9.60 % and 24.00 ± 9.23 %, when c2 and c3 were applied, respectively, compared to SC. The highest concentration of both the analogue GR24 and mimic SL-6 tested decreased the total pigments extracted by 8.15 ± 7.89 %, but not statistically significant (Figure 4d).
After normalization to the microalgal biomass within each treatment, the pigment content showed slightly different results compared with the non-normalized concentrations (Figure 5).
Both lowest concentrations of GR24 c2 (1 × 10-8 M) and c3 (1 × 10-9 M) induced slight increases in ChlA compared to SC, of 6.60 ± 3.00 % and 4.42 ± 1.40 %, respectively, but not statistically significant (Figure 5a). SL-6 c2 and c3 showed higher increases in the ChlA content of 15.70 ± 4.10 % and 12.30 ± 5.24 % from CS, respectively, compared to GR24 (Figure 5a).
The ChlB content was the highest upon the treatment with GR24 c3, with a 41.72 ± 18.10 % increase compared to SC (Figure 5b). Higher doses of GR24 recorded decreases in ChlB content per gram of microalgal biomass. SL-6 c1 and c3 showed marginally significant increases in ChlB by 7.85 ± 5.4 % and 12.87 ± 8.45 %, respectively, relative to SC (Figure 5b).
The total carotenoid content was slightly lower than control in the case of GR24 c3 and SL6 c1, which showed a rather opposite behaviour between the two compounds with concentration. The highest concentration of GR24 c1 (1 × 10-7 M) induced a small increase with 4.4 ± 3.4 % compared to SC. SL-6 c2 and c3 did not show a significant increase in carotenoid production (2.3 ± 1.5 % of SC for SL-6 c2).
The total pigments biosynthesized by the microalgal cultures per gram of biomass were the highest for the treatment with GR24 c3 (1 × 10-9 M) with 12.85 ± 5.46 % more than SC, followed by the treatment with SL-6 c2 and c3 with 10.78 ± 3.45 % and 9.81 ± 3.79 % more than SC, respectively. These results were marginally significant.
Considering the significance of photosystem II (PSII) and the quantum yield in microalgae photosynthesis, these results provide valuable insights into the efficiency of light energy conversion in C. sorokiniana under different treatments (Figure 6).
Treatments with lower SL doses resulted in higher chlorophyll fluorescence values compared to the solvent control at both time points, which suggests potentially enhanced PSII activity and improved quantum yield. This could indicate increased photosynthetic efficiency and confirms the stimulated biomass production in these treatments. Conversely, treatments that induce lower fluorescence values may indicate reduced PSII activity and lower quantum yield, possibly leading to decreased photosynthetic efficiency and growth. SL-6 increased significantly the C. sorokiniana quantum yield when the lower doses (c2 and c3) were applied, more than GR24, after 15 days of culturing.
We performed Pearson correlation between the parameters determined above to understand how these parameters relate to each other (Figure 7).
As expected, all the parameters of the microalgal growth correlated with each other and with the pigment concentration (per liter), as the pigment concentration is a function of the total biomass. The highest growth – pigment correlation was with ChlA and total Chl, and the lowest correlation was with ChlB. This indicates that the effect of SL on the production of pigments depends on the pigment type.
A positive correlation was observed between these parameters and the chlorophyll fluorescence as well, especially for optical density, turbidity, and ChlA that showed a statistically significant correlation. The correlation with the normalized pigments (level of expression) presented some differences compared with the pigment concentrations. The growth parameters showed statistically significant positive correlation only of the optical density with the total pigments/biomass. The total carotenoids/biomass had a negative correlation with almost all the other parameters, but not statistically significant. Other statistically significant correlations were between total pigments/biomass and ChlA, ChlB, ChlB/biomass and between the chlorophyll fluorescence and ChlA/biomass, total pigments/biomass.
In our experimental findings, treatments with SL-6, particularly at concentrations c2 (1 × 10-8 M) and c3 (1 × 10-9 M) demonstrated significant increases in C. sorokiniana's growth, biomass production, pigment synthesis, and Chl a photosynthetic yield (i.e., Chlorophyll fluorescence)
The results showed that lower concentrations of both tested synthetic strigolactones, GR24 and SL-6, stimulated microalgal growth, while higher concentrations inhibited it. Specifically, GR24 c2 (1 × 10-8 M) increased growth by 12.11 ± 8.25 %, and GR24 c3 (1 × 10-9 M) by 17.68 ± 2.54 % compared to the solvent control (SC). Similarly, SL-6 c2 increased growth by 7.00 ± 0.8 %, and SL-6 c3 by 16.26 ± 5 %. This suggests that at optimal concentrations, strigolactones enhance microalgal cell division and proliferation, which is crucial for biomass accumulation. These findings are aligning with similar growth-promoting effects of synthetic strigolactone on microalgae, reported in the literature [55,60,66]
2.2. Mung Seedlings Biotests
2.2.1. Effects on Seedlings Growth
The extract of the non-treated microalgal culture (CSk) slightly increased the root (radicle) and hypocotyl length compared to control C, without any treatment, both in the absence and presence of salt (Figure 8).
The root length of mung seedlings was further increased significantly by treatments with extracts of microalgae stimulated by 1 × 10−7, 1 × 10−8, 1 × 10−9 M SL-6, i.e., Sk-7, Sk-8, and Sk-9, respectively, compared to CSk. Sk-8 showed the highest increase, 26.46 ± 6.25 % of C whereas Sk-9 and Sk-9 induced similar values. Under stress conditions (50 mM salt), the radicle length decreased overall compared to unstressed seedlings. Treatments with Sk-7 S, Sk-8 S, and Sk-9 S determined significant increases, slightly higher than the non-treated microalgae (CSkS), reaching the control without salt and higher, indicating resilience under stress (Figure 8). The differences between the three concentrations of SL-6 were not significant.
In the case of hypocotyl, CSk, Sk-7, and Sk-9 variants had marginally significant higher length values (with approx. 5%) compared to control C and there was no statistical difference between the three treatments. Sk-8 induced a significant increase of 13.62 ± 3.54 % compared to C.
When salt stress was applied, treatments Sk-7 S, Sk-8 S, and Sk-9 S showed significant increases in hypocotyl length compared to CS, and marginally significant compared to CSkS. The differences between the three SL-6 concentrations were not statistically significant. The seedling length without salt stress varied between 4.19 ± 0.24 and 5.49 ± 0.12 cm among controls and treatments. When salt stress was applied, the seedlings length fell in between 2.91 ± 0.03 and 4.25 ± 0.06 cm. Without salt stress, the longest seedlings were recorded for variant Sk-8 and with salt stress for variant Sk-9, with 27.40 ± 5.47 % higher than C and 35.80 ± 7.60 % higher values than CS.
The SL-6 controls did not have a significant effect, neither in the absence nor in the presence of salt stress.
2.2.2. Acidification of the Growth Medium
Proton pumps are integral to plant physiology, affecting nutrient uptake, growth, pH regulation, and stress responses. Acid pH induces the activation of expansins, which induces root growth [67]. Proton pumps extrude H+ ions into the soil, lowering the pH around the roots. This acidification can help in nutrient solubilization and uptake [68].
The CSk treatment resulted in a 13.6 ± 7.5% increase in the total extracellular H+ levels compared to C, and the treatments with Sk-7 and Sk-8 exhibited a 19.7 ± 4.3% and 28.0 ± 6.7% respectively, higher extracellular H+ levels, in the absence of salt stress. Sk-9 showed a similar behavior to CSk (Figure 9a). The controls with SL-6 decreased slightly the total H+ levels compared to control C.
When salt stress was applied, there was a similar behavior, i.e., all four microalgal extract treatments (CSkS, Sk-7S, Sk-8S, Sk-9S) enhanced medium acidification in a similar trend. In particular, treatments with Sk-8S and Sk-7S had the highest effect and enhanced the H+ values with 46 ± 8.5% and 33 ± 6.4%, respectively, compared to CS. The Sk-8S treatment brought the H+ values slightly higher than the control without salt.
The specific extracellular H+ level resulted after normalizing the total H+ level to the root area (Figure 9b).
The highest specific H+ was recorded for the treatment with SK-9, both with and without salt stress, and the values were significantly higher than C(S) and CSk(S).
In order to understand better the relation between the SL effects on microalgae and the microalgal effects on the mung seedlings, we performed Pearson correlation, separately for the seedlings without stress (Figure 10a) and the salt-stressed seedlings (Figure 10b).
In both cases, eH+ was in general positively correlated with the microalga parameters, except with ChlB/biomass and total carotenoids/biomass, i.e., with the level of expression of ChlB and total carotenoids.
The seedling parameters had a tendency towards a negative correlation with ChlB/biomass and total carotenoids/biomass, especially in the absence of salt. The seedling growth parameters were positively correlated with seH+ just for the salt-stressed seedlings. The chlorophyll fluorescence parameter tends to be positively correlated with all parameters determined in mung bean seedlings, stressed or not-stressed. For the salt-stressed seedlings, seH+ of mung roots was highly statistically significant (positively) correlated with the chlorophyll fluorescence of the microalgae.
3. Discussions
3.1. Lower Doses of Synthetic Strigolactones as Microalgal Biostimulant
Synthetic SLs enhance microalgae growth parameters due to an increased photosynthetic performance. The photosynthetic yield of the photosystem II, determined by recording the chlorophyll fluorescence induced by pulse amplitude modulated (PAM) saturating light, was reported to be improved by optimal concentrations of synthetic SLs [55,60]. A similar effect of exogenously applied synthetic strigolactone on PAM-induced chlorophyll fluorescence was reported for plants submitted to abiotic stress —synthetic SLs reduced photoinhibition and improved the photosynthetic yield in plants exposed to (low) light stress [69,70]. In our present study, the increased photosynthetic yield (chlorophyll fluorescence) positively correlates with biomass accumulation. The correlations of the chlorophyll fluorescence with optical density and turbidity are statistically significant. The correlation between total biomass and pigment accumulation, Chl a, Chl b, total carotenoids, and total pigments is (highly) statistically significant. Synthetic SLs applied at optimal concentrations (in our case, 10-8 M and 10-9 M) increased the growth rate and accumulation of photosynthetic pigments. However, the expression of the photosynthetic pigments, i.e., specific pigments per total biomass, tends to be influenced differently. Chl A/ biomass and ChlB/ biomass tend to correlate positively with biomass accumulation, and total carotenoids/biomass tends to be negatively correlated with microalgae biomass accumulation.
Applying synthetic SLs seems to compensate for light attenuation along the light path due to photosynthetic active radiation (PAR) absorption by the photosynthetic pigments of the microalgae cells from the upper layers [71,72]. Due to culture shaking, microalgal cells travel in the culture flask, from the high light at the flask transparent wall to the low light/absence of PAR in the center of the flask culture. This situation involves transitioning from high light, which leads to photoinhibition and photosynthesis pigment loss, to light-limited conditions that reduce growth [73]. An argument for such a compensating effect of the light gradient in the microalgae culture is growth stimulation, which is more significant in cultures with a high cell density and a higher light attenuation rate. From the fifth day of culturing, marginal significant differences emerged between the control groups and the experimental treatments with synthetic SLs.
Various solutions were proposed to compensate for this drawback of microalgae cultivation at high cell density, such as applying flashing light [74-76], photobioreactor design that improves homogenous light distribution, e.g., internally illuminated photobioreactor [77,78], or photobioreactor internally illuminated with mirror walls [79], fluorescent dyes [80] or quantum dots [81], etc. Photosynthesis efficiency (determined by chlorophyll fluorescence/photosystem II yield) is increased in microalgal culture grown in internally illuminated photobioreactor [82] or with optimal high flashing light application [75]. Applying microalgal biostimulants based on synthetic SLs seems to represent an alternative to compensate for the stress of light attenuation in the microalgae cultivation systems. More studies are needed in this direction, including combining synthetic SLs with other solutions for homogenous light distribution and compensation of the light stress resulting from light attenuation in high cell density culture.
The ratio of total carotenoid pigments per biomass tends to be negatively correlated with biomass production and Chl b accumulation. Carotenoid expression per microalgal cell tends to decrease in the presence of synthetic SLs. The microalgae adapt to different light regimes by adjusting their photosynthetic pigment accumulation [83]. The lower accumulation of carotenoids in the presence of synthetic strigolactones seems to be related to an adaptation of C. sorokiniana to light attenuation and an enhanced tolerance to higher reactive oxygen species (ROS). The carotenoid pigments are involved in the photoprotection of the thylakoids through the xanthophyll cycle [84]. In microalgal cells from the Chlorella genus (C. vulgaris), carotenoids were demonstrated to be involved in photoprotective quenching in both photosystem I and II [85] and antioxidant protection against ROS [86]. Lower accumulation of carotenoids tends to generate more reactive oxygen species due to reduced quenching and reduced antioxidant protection. The plant biostimulants induce molecular priming [87] and enhance plant cell tolerance to oxidative stress [88,89]. A similar effect seems to be involved in microalgae cell response to microalgal biostimulant application.The detailed mechanism involved in this effect of synthetic SLs on microalgae photosynthetic performance is not yet known.
The effects of synthetic SLs application on endo-signals networks from microalgal cells are not yet completely understood. In Monoraphidium sp. QLY-1 microalgae, the application of GR24 determines an increased level of Ca2+ and nitric oxide (NO) acting as intracellular signals [66]. The treatment with 1 µM GR24 to the macrozooid stage of Haematococcus pluvialis determined a significant increase in biomass production due to increased photosynthetic efficiency. In the haematocyst stage, the same concentration of GR24 induced a higher accumulation of astaxanthin due to increased antioxidant activities and modification of fatty acids metabolism [90]. In terrestrial plants, the strigolactone endo-signaling interplay with Ca2+ [91] and NO [92].
However, the endogenous SLs were not identified in microalgae [53] despite these similarities that suggest a phytohormonal role of SLs in microalgae. The investigations done till now have demonstrated a limited presence of the following genes coding for enzymes involved in strigolactone biosynthesis in microalgae, carotenoid cleavage dioxygenase 7 (CCD7), carotenoid cleavage dioxygenase 8 (CCD8) and 9-cis/all-trans-β-carotene isomerase (D27) [93]. Microalgal genomes do not include sequences coding for receptors specific for strigolactones perception and signaling, e.g., D14 (DWARF14), an α/β-hydrolase protein acting as a non-canonical strigolactone receptor, and the other proteins involved in D14-mediated SLs perception, the F-box protein D3 and the D53 repressor protein [53].
Exo-signals that include a butenolide ring were described in various biological systems, such as those isolated from lichen-derived bacteria [94] or the quorum-sensing quenchers and biofilm inhibitors produced by seaweeds/macroalgae [95]. Perception and response systems to such exo-signals, potentially developed in microalgae, could also detect the butenolide ring from synthetic SLs. The reported effects of SLs on microalgae are mainly related to the formation of association microalgae — bacteria, microalgae — fungi, or microalgae — bacteria —fungi in aquatic systems. GR24 was reported to induce the formation of different symbiotic associations that are efficient in biogas upgrading and pollutant removal. Applying 10-9M GR24 increases the performance of C. vulgaris FACH-B—endophytic bacteria S395-2 in CO2 removal and consumption of organic and mineral nutrients [96]. GR24 applied in the same 10-9 M concentration increased the efficiency of symbiosis between C. vulgaris FACHB-8 and Pleurotus geesteranus bio-32868 and improved pollutant and CO2 removal [97]. 5-Deoxystrigol, applied at a concentration of 10−11 M, promotes the growth of symbiotic association C. vulgaris – G. lucidum – endophytic bacteria (S395-2), biogas upgrade, and nutrient removal [98].
The induction of the tolerance to the microalgae high-density cell culture could be related to a modulation of the quorum-sensing. Strigolactones modulate quorum sensing in differents systems, leading to various effects. The initial function of strigolactone in the terrestrial plants seems to be a quorum-sensing (QS) signals. The Physcomitrella patens moss regulates its colony extension and protonema branching by producing strigolactones [99]. In flowering plants, SLs are also involved in detecting neighboring plants [100]. Besides these QS functions in land plants, similar QS activities for SLs in other biological systems. The SL analog 2’-Epi-GR24 stimulated swarming motility in rhizobia [101]. SL- 6 promoted biofilm formation in P. graminis, a nodulation-helper bacteria [62]. Natural and synthetic strigolactones modulated quorum sensing in Vibrio cholerae, promoting biofilm formation and reducing toxin production [102]. The marine V. cholerae bacteria developed its quorum sensing coordinated biosynthesis of human toxin during shrimp chitin hydrolysis [103]. Hydrolyzed (shrimp) chitin was reported to promote microalgae growth [104].
Using synthetic SLs as microalgal biostimulants has implications for applied research. Increased microalgae tolerance to abiotic stress, including light (attenuation) stress, is important for higher yield and practical applications for microalgae. Elucidation of the systems from microalgae involved in perception and response to butenolide exo-signals is a fundamental research direction,which will lead to a better understanding of microalgae/phytoplankton ecology or lichen formation.
3.2. Biostimulant Effects of Microalgal Extracts Towards Mung Seedlings
The plant biostimulant functions of microalgae extracts have undoubtedly been demonstrated in the last decade [23,24,105,106]. The seed treatment with microalgae extract improved the response of resulting seedlings and/or plants to salt stress in wheat [107], bell pepper [108] and leafy lettuce [109]. Among the mechanisms involved in salt stress mitigation by microalgae extract are enhanced root growth and improved nutrient uptake [110]. In our study, the extracts from C. sorokiniana which were stimulated with different doses of SL-6 strigolactone mimic positively influenced mung seedlings development and physiology in the presence or the absence of salt stress. The effects were higher than those of the non-stimulated microalgal culture. The control treatments C-7, C-8, and C-9 were included to evaluate the baseline effects without microalgal extract. The results indicated that these controls did not significantly impact the measured parameters, including radicle length, hypocotyl length, plant height, and proton pump activity.
The same observations can also be done when salt stress was applied. This result means that the observed enhanced roots and shoots are due to the active ingredients from the microalga extracts and not from the residual effect of SL mimic. Regarding seedling measurements (Figure 8), the treatments with the microalgal extracts from the biomass grown in the presence of synthetic SL increased both radicle and hypocotyl compared to water control (C). In the absence of salt, for Sk-7 treatment, the radicle growth compared to control C was approximately 17.36% higher, from 2.4077 ± 0.133 cm to 2.8259 ± 0.108 cm, and the hypocotyl length increased by 19.22%. The treatment Sk-9 increased the root length to 2.812 ± 0.11 cm, 16.80% longer than the control and the shoot length with approximately 18.04% more than control. The Sk-8 recorded the best results, showing an increase of approx. 26.47% compared to the control C. Similar results were obtained in the presence of salt, but the highest stimulation was for Sk-9, a little higher than Sk-8.
Roots proton pump enhances nutrient bioavailability, especially phosphorus, and nutrient uptake [111] and is a marker for biostimulant action [112]. Results obtained in our study for proton pump activity showed that the treatments with the microalgal extracts enhanced the extracellular H+ level observed and measured by the medium acidification (and the resulting yellow color). The same happened for the specific extracellular H+ level (seH+), wherein the total area of the root (radicle) was considered. The chlorophyll fluorescence of microalga culture tends to be positively correlated with the root seH+ under normal conditions, and it is correlated at a highly significant statistical level with the root seH+ in salt-stressed seedlings.
This correlation suggests an involvement of the strigolactone exo-signals as a mediator of the associative interactions between edaphic (soil) benefic microalgae and plants (roots). SL-6 is a strigolactone mimic acting as exogenous signals for rhizosphere organisms [61]. Studies from the last decade spotlight the importance of microalgae as a component of the soil plant beneficial microbiome [113-115]. The microalgae from genus Chlorella (Trebouxiophyceae) have a great ecological plasticity, being adapted to both aquatic and soil environment. The strain ASIB BB67, isolated from soil alpine habitat, re-assessed as a C. vulgaris strain, has a tolerance to dessication greater than the type strain C. vulgaris SAG 211-11b, isolated from an aquatic environment [116]. C. sorokiniana strains were isolated edaphic environment, irrigated soils [117] and arid soils [118]. Populations / strain from this Chlorella genus acts as plant biostimulant / biofertilizer. The filtrate of the C. sorokiniana strain isolated from irrigated soil promote wheat plant growth [117]. The C. vulgaris strain MACC-GA0056, isolated from a sub-tropical soil, applied as crude culture, algal extract, and culture filtrate, improve wheat germination and growth [119]
Strigolactones are a “cry for help” in the rhizosphere [120], shaping plant beneficial microbiome [121,122] and recruiting microorganisms that promote plant tolerance to abiotic stress [123]. Strigolactones are released as exo-signals in the rhizosphere by salt-stressed plants in the presence of microorganisms that contribute to salt stress mitigation [124]. Chlorophyll fluorescence is a biomarker of microalgae cell health and chloroplast integrity [125]. As discussed above, the synthetic SLs (exo-signals) improved the photosynthetic performance of C. sorokiniana (measured as chlorophyll fluorescence). The extracts of microalgae (healthier) cells, from (synthetic) strigolactone challenged cultures, promote seedling development and enhance seedling tolerance to salt stress. The microalgae benefits from the presence of healthier plants – lower light stress at the soil surface due to the shadow effect, and root exudates as an alternative carbon source for mixotrophic/heterotrophic growth. Such a potential benefic interactions between the plant roots and microalgae suggest request further investigations on the role of strigolactones as exo-signals for the recruitment of beneficial microalgae by stressed plants.
4. Materials and Methods
4.1. Materials
The strigolactone analog (±) GR24 was supplied by StrigoLab (Turin, Italy). The strigolactone mimic SL-6, (2-4 –methyl – 5 – oxo - 2,5 – dihydro – furan – 2 - yloxy) – benzo [de] isoquinoline - 1,3-dione), with molecular mass of 309.28 da, was synthesized as previously described [126], starting from 1,8-naphthalic anhydride.
The chemicals for microalga cultivation were: Iron(III) chloride (FeCl3), sodium nitrate (NaNO3), calcium chloride (CaCl2·2H2O), magnesium sulfate (MgSO4·7H2O), cobalt(II) nitrate (Co(NO3)2·6H2O) were obtained from Merck (Darmstadt, Germany). Citric acid, Potassium dihydrogen phosphate (KH2PO4), zinc sulfate (ZnSO4·7H2O), boric acid (H3BO3), sodium molybdate (Na2MoO4·2H2O), were supplied by Scharlau (Barcelona, Spain). Manganese chloride (MnCl2·4H2O) was taken from Carl Roth (Karlsruhe, Germany) and EDTA disodium salt (Na2EDTA·2H2O) was purchased from Fluka (Honeywell, Morris Plains, NJ, USA). Copper sulfate (CuSO4·5H2O) was procured from Chimopar (Bucharest, Romania). The reagent for chlorophyll extraction Dimethyl sulfoxide (DMSO) was purchased from Merck (Darmstadt, Germany).
Agar and bromocresol purple, used for mung seedlings bioassay, were purchased from Scharlau (Barcelona, Spain).
The strain C. sorokiniana NIVA-CHL 17 was supplied by Norwegian Culture Collection of Algae, NORCCA, Norwegian Institute for Water Research (NIVA). Mung bean seeds (Vigna radiata (L.) R. Wilczek var. radiata) were supplied by a Romanian distributor of Vilmorin (Vilmorin, La Ménitré, France).
4.2. Testing the Effect of SL-6 on Microalgal Growth
Both GR24 and SL-6 were first solubilized in dimethyl sulfoxide (DMSO). Two stock solutions of 1 mM were prepared, considering the molar mass of each strigolactone: for GR24, MWt = 298.29 g/mol and for SL-6, MWt = 309.28 g/mol. The stock solutions were used to obtain the final tested concentrations, by serial dilutions with BG11 medium. The treatments applied were assessed in triplicate and were 1 × 10−7 M (c1), 1 × 10−8 M (c2) and, respectively 1 × 10−9 M (c3), together with a control and a solvent control (SC) with 10−7 M DMSO.
4.2.1. Microalgae Cultivation
Microalgae C. sorokiniana NIVA-CHL 17 were grown in sterilized 100 ml Erlenmeyer flasks, which contained 50 ml BG-11 growth media [127]. The medium was sterilized in an autoclave (Panasonic) at 121° C for 15 min. The final chemical concentrations were the following: 0.24 mM CaCl2·2H2O, 0.23 mM KH2PO4, 0.3 mM MgSO4·7H2O, 31 mM C6H8O7·H2O, 0.0027 mM EDTA disodium salt, 0.021 mM ammonium iron(III) citrate, 0.19 mM Na2CO3, BG-11 Trace Metals Solution 0.1% (0.046 mM H3BO3, 9 mM MnCl2·4H2O, 0.77 mM ZnSO4·7H2O, 0.17 mM Co(NO3)2·6H2O, 1.6 mM Na2MoO4·2H2O, 0.3 mM CuSO4·5H2O.
Fresh cultures were used for the test, with the final concentration of the inoculum 1% (500 µL/50 mL) of C. sorokiniana culture at the exponential phase. The sterile media distribution and flask inoculations were done in a microbiological hood Bio 2 Advantage Plus (Telstar, Barcelona, Spain), to keep axenic conditions.
The experimental treatments were incubated in a growth chamber AlgaeTron AG-230-Eco (Photon Systems Instruments, Drásov, Czech Republic) under controlled conditions of light (white, fluorescent lamp at 130 μmol/m2 ·s) and temperature (25 ± 1°C), with a photoperiod of 14/10 h of light/dark. For 15 days the vessels were continuously shaken by an orbital shaker, Unimax 1010 (Heidolph, Schwabach, Germany) set at 140 rpm.
4.2.2. Measurement of Growth Parameters
The effect of GR24 / SL-6 on C. sorokiniana was evaluated by measuring the main growth parameters such as optical density, turbidity, biomass production, and pigment concentrations for 15 days.
Optical Density: To assess the microalgal growth, the absorbance at 750 nm of a ten-fold dilution sample was measured with a UV-Vis spectrophotometer USB4000-UV-VIS Ocean Insight (Orlando, Florida, United States). This wavelength was selected to minimize measurement errors caused by chlorophyll pigment interference [128]. The parameter was measured after 3 days, 5 days, 7 days, 11 days and at the end of the experiment, after 15 days respectively.
Turbidity: The cell density of the cultures was determined by measuring turbidity using a turbidimeter Grant-Bio DEN-1B (Grant Instruments, Shepreth, Cambridgeshire United Kingdom), expressed in McFarland measurement units. The analysis was conducted with a working volume of 2 ml culture, harvested under sterile conditions.
Biomass: For biomass determination, a volume of 5 ml from each variant was aseptically pipped in pre-weighed Falcon tube and centrifuged at 9000 ×g for 10 min using a Universal 32 centrifuge (Hettich, Tuttlingen, Germany). After removing the supernatant, samples were dried to a constant weight in an oven Memmert UN 75 (Memmert, Schwabach, Germany), at 50°C [129].
Pigment concentration: 2 ml of each sample was centrifuged at 8000 ×g for 10 min at 20°C, to separate the cellular biomass from the supernatant. 2 ml of DMSO pre-heated at 60°C were added over the remaining pellet, and vortexed for 10 min. Next, the eppendorf tubes were centrifuged at 8000 ×g for 10 min. The absorbance was measured at three different wavelengths, 480 nm, 649 nm and 665 nm, necessary to calculate the content of chlorophyll a, chlorophyll b, total carotenoids and total pigments in the samples (mg/L), according to the following formulas [129]:
Additionally, the expression of pigments per gram of biomass was determined by normalizing the concentration to the microalgal biomass.
Chlorophyll fluorescence. The chlorophyll fluorescence was measured after 7 and 15 days of culturing, using a PAM fluorometer (PSI AquaPen AP 110/P, Photon Systems Instruments, Drásov). The manufacturer instructions were followed, to let the microalgal cultures adapt to dark for 10 min. The maximum quantum yield of PSII photochemistry ϕP0 was calculated according to the following formula :
where Fm—maximum chlorophyll a fluorescence (after actinic flash), Fv—maximum variable fluorescence, Fm-Fo; Fo—minimum chlorophyll a fluorescence (after dark adaptation).
4.3. Preparation of Microalgal Biomass Extract
For microalgal extracts, we selected the C. sorokiniana biomass from the treatments stimulated by the SL-6 mimic, which is more advantageous than GR24 from the economic point of view.
The microalgal biomass was concentrated by centrifugation at 9000 ×g for 10 min, and the suspension was brought to an average concentration of 0.8 g/L. The equation used to calculate the volume of supernatant that needed to be removed (ΔV) in order to achieve the final concentration was the following:
where: C1 = the concentration of microalgal biomass in each treatment (g/L); V1 = the microalga culture volume used for centrifugation (L); C2 = final concentration of microalgal biomass (g/L); V2 = final volume of microalgae culture after removing the excess supernatant (L). V2 was calculated from the equation, and the difference V2−V1 was the volume of the supernatant removed from the sample.
The suspensions were subjected to ultrasonication treatment for cell rupture and extraction of the intracellular active ingredients. Each suspension was ultrasonicated for 10 min, with 20 sec ON and 20 sec OFF, with an 80% amplitude, using a 400W ultrasonic processor UP400 st (Hielscher, Teltow, Germany) [130]. The extracts were centrifuged at 4°C and 9000 ×g for 10 min, and the supernatant was collected. Solutions were filtered first by 1.2 μm cellulose acetate filter (Sartorius Stedim Biotech GmbH) using a vacuum pump, and for sterility with a 0.2 μm PES sterile syringe filter. The final extract solutions combined both the extracellular microalgal biochemicals and the extracted intracellular active ingredients.
4.4. Plant Biostimulant Biotests on Seedlings
Mung bean seeds (Vigna radiata) were selected to test the microalgal extracts. The seeds were des-infected according to the following procedure: 3 min 90% EtOH immersion, water rinse, followed by 3 min 7% sodium hypochlorite immersion and 10 times rinsing with sterile water [131]
The biotests were carried out in 90 mm sterile petri dishes, each one containing two sterile gauze patches. On top 9 ml of each extract were aseptically pipetted and 10 sterile seeds were evenly placed using a sterilized tweezer. The experimental variants included controls with water, controls with SL-6 solutions (1 × 10-7 M, 1 × 10-8 M and 1 × 10-9 M, respectively), and treatments with the microalgal extracts, both without and with salt stress with a final molar concentration of 50 mM NaCl. The mung seeds were incubated in a growth chamber Micro Clima-SeriesTM Economic Lux Chamber (Snijders Labs, Tilburg, The Netherlands) with a temperature of 26°C for a 16 h light cycle, 240 μmol m-2 s-1 constant led light and 22°C for an 8 h dark cycle and were monitored for 5 days of growth.
4.4.1. Mung Bean Seedling Measurement
At the end of the growth period, the mung bean seedlings were photographed using a professional camera. Using the ImageJ software [132] the root (radicle) and shoot (hypocotyl) lengths were measured.
4.4.2. Medium Acidification Assay
The method applied was conducted based on [133]. On the second day of seedling growth, a solution of 0.04 g/L bromocresol purple in ddH2O was prepared and adjusted to pH 6.7 with 1 N NaOH and supplemented with agar at a final concentration of 0.75%, autoclaved at 121°C, and poured into sterile 90 mm Petri dishes. Briefly, one mung seedling of each triplicate was placed on the partially hardened agar at approximately 30°C, pushing the roots into the agar until being partially embedded into the gel. After 24 h the plates were photographed and the yellow area, the color intensity of the yellow area and the root area were measured with ImageJ. The data were used to semi-quantify the specific extracellular H+ level (seH+) and the total H+ level (eH+), using the following equations: [133]
where Ay is the yellow area, I is the intensity of the yellow area, and Ar is the root area.
4.5. Statistical Analysis and Figure Design
The data underwent one-way ANOVA analysis (with a significance level set at p ≤ 0.05), followed by a comparison of mean differences using the Tukey HSD post hoc test. Each experiment was conducted in triplicate, presenting the results as mean ± standard error. The graphs from the figures were designed using Origin Pro 2018 software (OriginLab, Northampton, MA, USA). IBM SPSS Statistics for Windows, software version 26.0.0.0 (IBM, Armonk, NY, USA) was used for statistical analysis (One-Way ANOVA) and Pearson correlation analysis (Bivariate correlations).
5. Conclusions
The study investigated the effects of a strigolactone analog (GR24) and a mimic (SL-6) on the growth and biomass production of Chlorella sorokiniana. Further studies were conducted on the effects of microalgal culture extracts on mung bean development and growth. SL-6, a cost-effective alternative to GR24, promoted microalgae growth and enhanced photosynthetic performance. Treatment with SL-6 consistently resulted in comparable or superior outcomes in biomass production, pigment synthesis, and photosystem II yield (determined as chlorophyll fluorescence) compared to GR24 treatments. These findings highlight the potential of SL-6 as a cost-efficient yet highly effective solution for enhancing microalgal cultivation and biomass yield.
The higher dose of 10-7 M inhibited microalgal growth determined as cell density, biomass production, or pigment synthesis for both tested synthetic strigolactones, GR24 and SL-6. Lower doses stimulated the growth parameters of C. sorokiniana, enhancing up to 24.00% the total pigment accumulation when SL-6 at 10-9 M was applied. These variations demonstrate a hormetic effect, with higher doses causing inhibition and lower doses have a stimulant activity.
The treatments with synthetic strigolactones, especially with SL-6, improved microalgae tolerance to the light attenuation stress in high-density culture. These results have practical implications for microalgae cultivation, offering an alternative to the solutions for compensating light stress in microalgae cultivated in agitated media.
Microalgal extracts were proved to have plant biostimulant functions, enhancing radicle elongation and overall seedling length in normal conditions and increasing mung bean tolerance to salt stress. The activity of the root proton pump, a marker for biostimulant action [112], is amplified by treatment with microalgae extracts. The extract from microalgae challenged with SL-6, especially the treatment with SK-8 (extract of C. sorokiniana culture stimulated with 10-8 M SL-6 ), demonstrated a more pronounced plant biostimulant activity than non-challenged culture. The results suggest a potential involvement of strigolactone exo-signals in establishing the mutual benefic associations between edaphic microalgae and plant roots.
Author Contributions
Conceptualization, F.O.; methodology, D.G.P., N.T., and D.C.-A.; validation, D.C.-A. and F.O.; formal analysis, D.G.P. and D.C.-A.; investigation, D.G.P., N.T., F.G.; resources, F.G., S.S. and F.O.; data curation, D.G.P. and C.L.; writing—original draft preparation, D.G.P.; writing—review and editing , D.C.-A. and F.O.; visualization, D.C.-A. and F.O.; supervision, D.C.-A. and F.O.; project administration, D.C.-A., C.L., S.S. and F.O.; funding acquisition, D.C.-A., F.G., S.S. and F.O. All authors have read and agreed to the published version of the manuscript.
Funding
The research leading to these results has received funding from NO Grants 2014-2021, under Project RO-NO-2019-540, contract no. 14/2020 and project PN.23.06.02.01 InteGral, Nucleu Program, funded by the Romanian Ministry of Research, Innovation and Digitalization.
Data Availability Statement
All data are included within the article.
Acknowledgments
We thank Marius Ghiurea for guidance in measuring the chlorophyll fluorescence of the microalgal cultures.
Conflicts of Interest
The authors declare no conflicts of interest
References
- Chen, C.; Tang, T.; Shi, Q.; Zhou, Z.; Fan, J. The potential and challenge of microalgae as promising future food sources. Trends in Food Science & Technology 2022, 126, 99–112. [Google Scholar] [CrossRef]
- Abdullah, M.; Ali, Z.; Yasin, M.T.; Amanat, K.; Sarwar, F.; Khan, J.; Ahmad, K. Advancements in sustainable production of biofuel by Microalgae: Recent insights and future directions. Environmental Research 2024, 262, 119902. [Google Scholar] [CrossRef] [PubMed]
- Abreu, A.P.; Morais, R.C.; Teixeira, J.A.; Nunes, J. A comparison between microalgal autotrophic growth and metabolite accumulation with heterotrophic, mixotrophic and photoheterotrophic cultivation modes. Renewable and Sustainable Energy Reviews 2022, 159, 112247. [Google Scholar] [CrossRef]
- Velea, S.; Oancea, F.; Fischer, F. Heterotrophic and mixotrophic microalgae cultivation. Microalgae-based biofuels and bioproducts.
- Do, C.V.T.; Pham, M.H.T.; Pham, T.Y.T.; Dinh, C.T.; Bui, T.U.T.; Tran, T.D. Microalgae and bioremediation of domestic wastewater. Current Opinion in Green and Sustainable Chemistry 2022, 34, 100595. [Google Scholar] [CrossRef]
- Geremia, E.; Ripa, M.; Catone, C.M.; Ulgiati, S. A Review about Microalgae Wastewater Treatment for Bioremediation and Biomass Production—A New Challenge for Europe. Environments 2021, 8, 136. [Google Scholar] [CrossRef]
- Kumar, P.; Perumal, P.K.; Sumathi, Y.; Singhania, R.R.; Chen, C.-W.; Dong, C.-D.; Patel, A.K. Nano-Enabled Microalgae Bioremediation: Advances in Sustainable Pollutant Removal and Value-Addition. Environmental Research 2024, 263, 120011. [Google Scholar] [CrossRef]
- Stratigakis, N.C.; Nazos, T.T.; Chatzopoulou, M.; Mparka, N.; Spantidaki, M.; Lagouvardou-Spantidaki, A.; Ghanotakis, D.F. Cultivation of a naturally resilient Chlorella sp.: A bioenergetic strategy for valorization of cheese whey for high nutritional biomass production. Algal Research 2024, 82, 103616. [Google Scholar] [CrossRef]
- Girard, J.-M.; Roy, M.-L.; Hafsa, M.B.; Gagnon, J.; Faucheux, N.; Heitz, M.; Tremblay, R.; Deschênes, J.-S. Mixotrophic cultivation of green microalgae Scenedesmus obliquus on cheese whey permeate for biodiesel production. Algal Research 2014, 5, 241–248. [Google Scholar] [CrossRef]
- Santana, H.; Cereijo, C.R.; Teles, V.C.; Nascimento, R.C.; Fernandes, M.S.; Brunale, P.; Campanha, R.C.; Soares, I.P.; Silva, F.C.; Sabaini, P.S. Microalgae cultivation in sugarcane vinasse: Selection, growth and biochemical characterization. Bioresource Technology 2017, 228, 133–140. [Google Scholar] [CrossRef]
- Silva, S.; Melo, L.B.U.; Borrego, B.B.; Gracioso, L.H.; Perpetuo, E.A.; do Nascimento, C.A.O. Sugarcane vinasse as feedstock for microalgae cultivation: from wastewater treatment to bioproducts generation. Brazilian Journal of Chemical Engineering 2023, 1–11. [Google Scholar] [CrossRef]
- Scarponi, P.; Bravi, M.; Cavinato, C. Wine Lees as Alternative Substrate for Microalgae Cultivation: New Opportunity in Winery Waste Biorefinery Application. Waste 2023, 1, 631–639. [Google Scholar] [CrossRef]
- Mastoras, P.; Zkeri, E.; Panara, A.; Dasenaki, M.E.; Maragou, N.C.; Vakalis, S.; Fountoulakis, M.S.; Thomaidis, N.S.; Stasinakis, A.S. Application of a pilot-scale solar still for wine lees management: Characterization of by-products and valorization potential. Journal of Environmental Chemical Engineering 2023, 11, 111227. [Google Scholar] [CrossRef]
- León-Vaz, A.; León, R.; Díaz-Santos, E.; Vigara, J.; Raposo, S. Using agro-industrial wastes for mixotrophic growth and lipids production by the green microalga Chlorella sorokiniana. New biotechnology 2019, 51, 31–38. [Google Scholar] [CrossRef]
- Beigbeder, J.-B.; Boboescu, I.-Z.; Lavoie, J.-M. Thin stillage treatment and co-production of bio-commodities through finely tuned Chlorella vulgaris cultivation. Journal of Cleaner Production 2019, 216, 257–267. [Google Scholar] [CrossRef]
- Miazek, K.; Remacle, C.; Richel, A.; Goffin, D. Effect of Lignocellulose Related Compounds on Microalgae Growth and Product Biosynthesis: A Review. Energies 2014, 7, 4446–4481. [Google Scholar] [CrossRef]
- Chong, J.W.R.; Khoo, K.S.; Yew, G.Y.; Leong, W.H.; Lim, J.W.; Lam, M.K.; Ho, Y.-C.; Ng, H.S.; Munawaroh, H.S.H.; Show, P.L. Advances in production of bioplastics by microalgae using food waste hydrolysate and wastewater: a review. Bioresource Technology 2021, 342, 125947. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, S.; Liu, J.; Wang, J.; Xiao, M.; Liang, Q.; Ren, X.; Wang, Y.; Mou, H.; Sun, H. Realization process of microalgal biorefinery: The optional approach toward carbon net-zero emission. Science of the Total Environment 2023, 901, 165546. [Google Scholar] [CrossRef]
- Cuevas-Castillo, G.A.; Navarro-Pineda, F.S.; Rodríguez, S.A.B.; Rivero, J.C.S. Advances on the processing of microalgal biomass for energy-driven biorefineries. Renewable and Sustainable Energy Reviews 2020, 125, 109606. [Google Scholar] [CrossRef]
- Koyande, A.K.; Chew, K.W.; Rambabu, K.; Tao, Y.; Chu, D.-T.; Show, P.-L. Microalgae: A potential alternative to health supplementation for humans. Food Science and Human Wellness 2019, 8, 16–24. [Google Scholar] [CrossRef]
- Vaz, B.d.S.; Moreira, J.B.; Morais, M.G.d.; Costa, J.A.V. Microalgae as a new source of bioactive compounds in food supplements. Current Opinion in Food Science 2016, 7, 73–77. [Google Scholar] [CrossRef]
- Chiaiese, P.; Corrado, G.; Colla, G.; Kyriacou, M.C.; Rouphael, Y. Renewable Sources of Plant Biostimulation: Microalgae as a Sustainable Means to Improve Crop Performance. Frontiers in Plant Science 2018, 9, 1782. [Google Scholar] [CrossRef] [PubMed]
- Kapoore, R.V.; Wood, E.E.; Llewellyn, C.A. Algae biostimulants: A critical look at microalgal biostimulants for sustainable agricultural practices. Biotechnology Advances 2021, 49, 107754. [Google Scholar] [CrossRef] [PubMed]
- Chabili, A.; Minaoui, F.; Hakkoum, Z.; Douma, M.; Meddich, A.; Loudiki, M. A comprehensive review of microalgae and cyanobacteria-based biostimulants for agriculture uses. Plants 2024, 13, 159. [Google Scholar] [CrossRef] [PubMed]
- EU. Regulation (EU) 2019/1009 of the European Parliament and of the Council of 5 June 2019 laying down rules on the making available on the market of EU fertilising products and amending Regulations (EC) No 1069/2009 and (EC) No 1107/2009 and repealing Regulation (EC) No 2003/2003. 2019/1009 2019.
- Mutale-Joan, C.; Redouane, B.; Najib, E.; Yassine, K.; Lyamlouli, K.; Laila, S.; Zeroual, Y.; Hicham, E.A. Screening of microalgae liquid extracts for their bio stimulant properties on plant growth, nutrient uptake and metabolite profile of Solanum lycopersicum L. Scientific Reports 2020, 10, 2820. [Google Scholar] [CrossRef]
- Oancea, F.; Velea, S.; Mincea, C.; Ilie, L. Micro-algae based plant biostimulant and its effect on water stressed tomato plants. Romanian Journal of Plant Protection 2013, 6, 104–117. [Google Scholar]
- Kusvuran, S. Microalgae (Chlorella vulgaris Beijerinck) alleviates drought stress of broccoli plants by improving nutrient uptake, secondary metabolites, and antioxidative defense system. Horticultural Plant Journal 2021, 7, 221–231. [Google Scholar] [CrossRef]
- Moon, J.; Park, Y.J.; Choi, Y.B.; Truong, T.Q.; Huynh, P.K.; Kim, Y.B.; Kim, S.M. Physiological Effects and Mechanisms of Chlorella vulgaris as a Biostimulant on the Growth and Drought Tolerance of Arabidopsis thaliana. Plants 2024, 13, 3012. [Google Scholar] [CrossRef]
- Puglisi, I.; La Bella, E.; Rovetto, E.I.; Lo Piero, A.R.; Baglieri, A. Biostimulant Effect and Biochemical Response in Lettuce Seedlings Treated with A Scenedesmus quadricauda Extract. Plants 2020, 9, 123. [Google Scholar] [CrossRef]
- Chovanček, E.; Salazar, J.; Şirin, S.; Allahverdiyeva, Y. Microalgae from Nordic collections demonstrate biostimulant effect by enhancing plant growth and photosynthetic performance. Physiologia Plantarum 2023, 175, e13911. [Google Scholar] [CrossRef]
- Brück, W.M.; Alfonso, E.; Rienth, M.; Andlauer, W. Heat Stress Resistance in Chlorella vulgaris Enhanced by Hydrolyzed Whey Proteins. Agronomy 2024, 14, 2854. [Google Scholar] [CrossRef]
- Chung, M.K.; Hu, R.; Wong, M.H.; Cheung, K.C. Comparative toxicity of hydrophobic contaminants to microalgae and higher plants. Ecotoxicology 2007, 16, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Cook, C.; Whichard, L.P.; Turner, B.; Wall, M.E.; Egley, G.H. Germination of witchweed (Striga lutea Lour.): isolation and properties of a potent stimulant. Science 1966, 154, 1189–1190. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, K.; Xie, X.; Yoneyama, K.; Takeuchi, Y. Strigolactones: structures and biological activities. Pest Management Science 2009, 65, 467–470. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, K.; Xie, X.; Yoneyama, K.; Kisugi, T.; Nomura, T.; Nakatani, Y.; Akiyama, K.; McErlean, C.S. Which are the major players, canonical or non-canonical strigolactones? Journal of experimental botany 2018, 69, 2231–2239. [Google Scholar] [CrossRef] [PubMed]
- Khalid, M.F.; Shafqat, W.; Khan, R.I.; Jawaid, M.Z.; Hussain, S.; Saqib, M.; Rizwan, M.; Ahmed, T. Unveiling the resilience mechanism: Strigolactones as master regulators of plant responses to abiotic stresses. Plant Stress 2024, 12, 100490. [Google Scholar] [CrossRef]
- Qi, J.; Mao, Y.; Cui, J.; Lu, X.; Xu, J.; Liu, Y.; Zhong, H.; Yu, W.; Li, C. The role of strigolactones in resistance to environmental stress in plants. Physiologia Plantarum 2024, 176, e14419. [Google Scholar] [CrossRef]
- Barbier, F.; Fichtner, F.; Beveridge, C. The strigolactone pathway plays a crucial role in integrating metabolic and nutritional signals in plants. Nature Plants 2023, 9, 1191–1200. [Google Scholar] [CrossRef]
- Daignan-Fornier, S.; Keita, A.; Boyer, F.-D. Chemistry of Strigolactones, Key Players in Plant Communication. ChemBioChem 2024, 25, e202400133. [Google Scholar] [CrossRef]
- Clark, J.; Bennett, T. Cracking the enigma: understanding strigolactone signalling in the rhizosphere. Journal of Experimental Botany 2024, 75, 1159–1173. [Google Scholar] [CrossRef]
- Zwanenburg, B.; Ćavar Zeljković, S.; Pospíšil, T. Synthesis of strigolactones, a strategic account. Pest Management Science 2016, 72, 15–29. [Google Scholar] [CrossRef]
- Mangnus, E.M.; Dommerholt, F.J.; De Jong, R.L.P.; Zwanenburg, B. Improved synthesis of strigol analog GR24 and evaluation of the biological activity of its diastereomers. Journal of Agricultural and Food Chemistry 1992, 40, 1230–1235. [Google Scholar] [CrossRef]
- Min, Z.; Li, R.; Chen, L.; Zhang, Y.; Li, Z.; Liu, M.; Ju, Y.; Fang, Y. Alleviation of drought stress in grapevine by foliar-applied strigolactones. Plant Physiology and Biochemistry 2019, 135, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Iftikhar, I.; Shahbaz, M.; Maqsood, M.F.; Zulfiqar, U.; Rana, S.; Farhat, F.; Farooq, H.; Ahmad, K.; Jamil, M.; Haider, F.U.; et al. Resilient mechanism of strigolactone (GR24) in regulating morphological and biochemical status of maize under salt stress. Biocatalysis and Agricultural Biotechnology 2024, 60, 103340. [Google Scholar] [CrossRef]
- Omoarelojie, L.O.; Kulkarni, M.G.; Finnie, J.F.; Pospíšil, T.; Strnad, M.; Van Staden, J. Synthetic strigolactone (rac-GR24) alleviates the adverse effects of heat stress on seed germination and photosystem II function in lupine seedlings. Plant Physiology and Biochemistry 2020, 155, 965–979. [Google Scholar] [CrossRef]
- Zhang, J.; Tang, C.; Xie, J.; Li, J.; Zhang, X.; Wang, C. Exogenous strigolactones alleviate low-temperature stress in peppers seedlings by reducing the degree of photoinhibition. BMC Plant Biology 2024, 24, 907. [Google Scholar] [CrossRef]
- Wani, K.I.; Naeem, M.; Khan, M.M.A.; Aftab, T. Insights into strigolactone (GR24) mediated regulation of cadmium-induced changes and ROS metabolism in Artemisia annua. Journal of Hazardous Materials 2023, 448, 130899. [Google Scholar] [CrossRef]
- Mitra, D.; Panneerselvam, P.; Chidambaranathan, P.; Nayak, A.K.; Priyadarshini, A.; Senapati, A.; Mohapatra, P.K.D. Strigolactone GR24-mediated mitigation of phosphorus deficiency through mycorrhization in aerobic rice. Current Research in Microbial Sciences 2024, 6, 100229. [Google Scholar] [CrossRef]
- Wani, K.I.; Zehra, A.; Choudhary, S.; Naeem, M.; Khan, M.M.A.; Khan, R.; Aftab, T. Exogenous Strigolactone (GR24) Positively Regulates Growth, Photosynthesis, and Improves Glandular Trichome Attributes for Enhanced Artemisinin Production in Artemisia annua. Journal of Plant Growth Regulation 2023, 42, 4606–4615. [Google Scholar] [CrossRef]
- Li, C.; Lu, X.; Liu, Y.; Xu, J.; Yu, W. Strigolactone Alleviates the Adverse Effects of Salt Stress on Seed Germination in Cucumber by Enhancing Antioxidant Capacity. Antioxidants 2023, 12, 1043. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, L.; Ma, C.; Su, M.; Wang, J.; Zheng, S.; Zhang, T. Exogenous strigolactones alleviate the photosynthetic inhibition and oxidative damage of cucumber seedlings under salt stress. Scientia Horticulturae 2022, 297, 110962. [Google Scholar] [CrossRef]
- Delaux, P.-M.; Xie, X.; Timme, R.E.; Puech-Pages, V.; Dunand, C.; Lecompte, E.; Delwiche, C.F.; Yoneyama, K.; Bécard, G.; Séjalon-Delmas, N. Origin of strigolactones in the green lineage. New Phytologist 2012, 195, 857–871. [Google Scholar] [CrossRef] [PubMed]
- Shu, L.; Wang, Z.; Li, Y.; Zheng, Z. Supplementation of 5-deoxystrigol for higher pollutant removal and biogas quality improvement by different microalgae-based technologies. Water Environment Research 2023, 95, e10895. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Xue, Z.; Sun, L.; Zhao, C.; Sun, S.; Liu, J.; Zhao, Y.; Liu, J. Effect of GR24 concentrations on biogas upgrade and nutrient removal by microalgae-based technology. Bioresource Technology 2020, 312, 123563. [Google Scholar] [CrossRef] [PubMed]
- Popa, D.G.; Lupu, C.; Constantinescu-Aruxandei, D.; Oancea, F. Humic Substances as Microalgal Biostimulants—Implications for Microalgal Biotechnology. Marine Drugs 2022, 20, 327. [Google Scholar] [CrossRef]
- Puglisi, I.; Barone, V.; Sidella, S.; Coppa, M.; Broccanello, C.; Gennari, M.; Baglieri, A. Biostimulant activity of humic-like substances from agro-industrial waste on Chlorella vulgaris and Scenedesmus quadricauda. European Journal of Phycology 2018, 53, 433–442. [Google Scholar] [CrossRef]
- Faraon, V.A.; Popa, D.G.; Tudor-Popa, I.; Mihăilă, E.G.; Oancea, F. Effect of Humic Acids from Biomass Biostimulant on Microalgae Growth. Chemistry Proceedings 2022, 7, 51. [Google Scholar] [CrossRef]
- Suparmaniam, U.; Lam, M.K.; Lim, J.W.; Rawindran, H.; Chai, Y.H.; Tan, I.S.; Fui Chin, B.L.; Kiew, P.L. The potential of waste chicken feather protein hydrolysate as microalgae biostimulant using organic fertilizer as nutrients source. IOP Conference Series: Earth and Environmental Science 2022, 1074, 012028. [Google Scholar] [CrossRef]
- Popa, D.G.; Georgescu, F.; Dumitrascu, F.; Shova, S.; Constantinescu-Aruxandei, D.; Draghici, C.; Vladulescu, L.; Oancea, F. Novel Strigolactone Mimics That Modulate Photosynthesis and Biomass Accumulation in Chlorella sorokiniana. Molecules 2023, 28, 7059. [Google Scholar] [CrossRef]
- Oancea, F.; Georgescu, E.; Matusova, R.; Georgescu, F.; Nicolescu, A.; Raut, I.; Jecu, M.-L.; Vladulescu, M.-C.; Vladulescu, L.; Deleanu, C. New Strigolactone Mimics as Exogenous Signals for Rhizosphere Organisms. Molecules 2017, 22, 961. [Google Scholar] [CrossRef]
- Bala, I.; Airinei, A.; Georgescu, E.; Oancea, F.; Georgescu, F.; Nicolescu, A.; Tigoianu, R.; Deleanu, C. Photophysical and biological properties of a strigolactone mimic derived from 1, 8-naphthalic anhydride. Revue Roumaine de Chimie 2022, 67, 51–62. [Google Scholar] [CrossRef]
- Brooks, S.J.; Gomes, T.; Almeida, A.C.; Christou, M.; Zheng, C.; Shaposhnikov, S.; Popa, D.G.; Georgescu, F.; Oancea, F. An ecotoxicological assessment of a strigolactone mimic used as the active ingredient in a plant biostimulant formulation. Ecotoxicology and Environmental Safety 2024, 275, 116244. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhao, C.; Sun, S.; Zhao, Y.; Liu, J. Performance of different microalgae-based technologies in nutrient removal and biogas upgrading in response to various GR24 concentrations. International Biodeterioration & Biodegradation 2021, 158, 105166. [Google Scholar] [CrossRef]
- Maltsev, Y.; Maltseva, K.; Kulikovskiy, M.; Maltseva, S. Influence of Light Conditions on Microalgae Growth and Content of Lipids, Carotenoids, and Fatty Acid Composition. Biology 2021, 10, 1060. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Zhao, Y.; Li, T.; Han, B.; Zhao, P.; Xu, J.-W.; Yu, X. Enhancement of lipid accumulation in Monoraphidium sp. QLY-1 by induction of strigolactone. Bioresource Technology 2019, 288, 121607. [Google Scholar] [CrossRef]
- Gámez-Arjona, F.M.; Sánchez-Rodríguez, C.; Montesinos, J.C. The root apoplastic pH as an integrator of plant signaling. Frontiers in Plant Science 2022, 13. [Google Scholar] [CrossRef]
- Zeng, H.; Chen, H.; Zhang, M.; Ding, M.; Xu, F.; Yan, F.; Kinoshita, T.; Zhu, Y. Plasma membrane H+-ATPases in mineral nutrition and crop improvement. Trends in Plant Science 2024. [Google Scholar] [CrossRef]
- Lu, T.; Yu, H.; Li, Q.; Chai, L.; Jiang, W. Improving Plant Growth and Alleviating Photosynthetic Inhibition and Oxidative Stress From Low-Light Stress With Exogenous GR24 in Tomato (Solanum lycopersicum L.) Seedlings. Frontiers in Plant Science 2019, 10. [Google Scholar] [CrossRef]
- Gong, J.; Wang, R.; Liu, B.; Zhu, T.; Li, H.; Long, S.; Liu, T.; Xu, Y. Regulatory mechanism of strigolactone in tall fescue to low-light stress. Plant Physiology and Biochemistry 2024, 215, 109054. [Google Scholar] [CrossRef]
- Ma, S.; Zeng, W.; Huang, Y.; Zhu, X.; Xia, A.; Zhu, X.; Liao, Q. Revealing the synergistic effects of cells, pigments, and light spectra on light transfer during microalgae growth: A comprehensive light attenuation model. Bioresource Technology 2022, 348, 126777. [Google Scholar] [CrossRef]
- Wágner, D.S.; Valverde-Pérez, B.; Plósz, B.G. Light attenuation in photobioreactors and algal pigmentation under different growth conditions – Model identification and complexity assessment. Algal Research 2018, 35, 488–499. [Google Scholar] [CrossRef]
- Serôdio, J.; Campbell, D.A. Photoinhibition in optically thick samples: Effects of light attenuation on chlorophyll fluorescence-based parameters. Journal of Theoretical Biology 2021, 513, 110580. [Google Scholar] [CrossRef] [PubMed]
- Abu-Ghosh, S.; Fixler, D.; Dubinsky, Z.; Iluz, D. Flashing light in microalgae biotechnology. Bioresource Technology 2016, 203, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Pozzobon, V. Chlorella vulgaris cultivation under super high light intensity: An application of the flashing light effect. Algal Research 2022, 68, 102874. [Google Scholar] [CrossRef]
- Khopkar, A.; Mahulkar, A.; Rajpoot, S.; Sapre, A. Identifying Opportunities for Improving the Light Utilization Efficiency of Microalgae in a Flashing Light Regime. Industrial & Engineering Chemistry Research 2023, 62, 19035–19042. [Google Scholar] [CrossRef]
- Amaral, M.S.; Loures, C.C.A.; Naves, F.L.; Baeta, B.E.L.; Silva, M.B.; Prata, A.M.R. Evaluation of cell growth performance of microalgae Chlorella minutissima using an internal light integrated photobioreactor. Journal of Environmental Chemical Engineering 2020, 8, 104200. [Google Scholar] [CrossRef]
- Zhang, H.; Gong, P.; Cai, Q.; Zhang, C.; Gao, B. Maximizing fucoxanthin production in Odontella aurita by optimizing the ratio of red and blue light-emitting diodes in an auto-controlled internally illuminated photobioreactor. Bioresource Technology 2022, 344, 126260. [Google Scholar] [CrossRef]
- Ahangar, A.K.; Yaqoubnejad, P.; Divsalar, K.; Mousavi, S.; Taghavijeloudar, M. Design a novel internally illuminated mirror photobioreactor to improve microalgae production through homogeneous light distribution. Bioresource Technology 2023, 387, 129577. [Google Scholar] [CrossRef]
- Seo, Y.H.; Lee, Y.; Jeon, D.Y.; Han, J.-I. Enhancing the light utilization efficiency of microalgae using organic dyes. Bioresource Technology 2015, 181, 355–359. [Google Scholar] [CrossRef]
- Zhang, F.; Li, Y.; Miao, X. Quantum dot-based light conversion strategy for customized cultivation of microalgae. Bioresource Technology 2024, 397, 130489. [Google Scholar] [CrossRef]
- Malapascua, J.; Ranglová, K.; Masojídek, J. Photosynthesis and growth kinetics of Chlorella vulgaris R-117 cultured in an internally LED-illuminated photobioreactor. Photosynthetica 2019, 57. [Google Scholar] [CrossRef]
- Levasseur, W.; Taidi, B.; Lacombe, R.; Perré, P.; Pozzobon, V. Impact of seconds to minutes photoperiods on Chlorella vulgaris growth rate and chlorophyll a and b content. Algal Research 2018, 36, 10–16. [Google Scholar] [CrossRef]
- Goss, R.; Latowski, D. Lipid Dependence of Xanthophyll Cycling in Higher Plants and Algae. Frontiers in Plant Science 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Girolomoni, L.; Bellamoli, F.; de la Cruz Valbuena, G.; Perozeni, F.; D'Andrea, C.; Cerullo, G.; Cazzaniga, S.; Ballottari, M. Evolutionary divergence of photoprotection in the green algal lineage: a plant-like violaxanthin de-epoxidase enzyme activates the xanthophyll cycle in the green alga Chlorella vulgaris modulating photoprotection. New Phytologist 2020, 228, 136–150. [Google Scholar] [CrossRef] [PubMed]
- Guardini, Z.; Dall’Osto, L.; Barera, S.; Jaberi, M.; Cazzaniga, S.; Vitulo, N.; Bassi, R. High Carotenoid Mutants of Chlorella vulgaris Show Enhanced Biomass Yield under High Irradiance. Plants 2021, 10, 911. [Google Scholar] [CrossRef]
- Kerchev, P.; van der Meer, T.; Sujeeth, N.; Verlee, A.; Stevens, C.V.; Van Breusegem, F.; Gechev, T. Molecular priming as an approach to induce tolerance against abiotic and oxidative stresses in crop plants. Biotechnology Advances 2020, 40, 107503. [Google Scholar] [CrossRef]
- Omoarelojie, L.O.; Kulkarni, M.G.; Finnie, J.F.; van Staden, J. Biostimulants and the modulation of plant antioxidant systems and properties. In Biostimulants for Crops from Seed Germination to Plant Development, Gupta, S., Van Staden, J., Eds.; Academic Press: 2021; pp. 333–363.
- Hasanuzzaman, M.; Parvin, K.; Bardhan, K.; Nahar, K.; Anee, T.I.; Masud, A.A.C.; Fotopoulos, V. Biostimulants for the Regulation of Reactive Oxygen Species Metabolism in Plants under Abiotic Stress. Cells 2021, 10, 2537. [Google Scholar] [CrossRef]
- Wang, X.; Mou, J.-H.; Qin, Z.-H.; Hao, T.-B.; Zheng, L.; Buhagiar, J.; Liu, Y.-H.; Balamurugan, S.; He, Y.; Lin, C.S.K.; et al. Supplementation with rac-GR24 Facilitates the Accumulation of Biomass and Astaxanthin in Two Successive Stages of Haematococcus pluvialis Cultivation. Journal of Agricultural and Food Chemistry 2022, 70, 4677–4689. [Google Scholar] [CrossRef]
- Bürger, M.; Chory, J. The Many Models of Strigolactone Signaling. Trends in Plant Science 2020, 25, 395–405. [Google Scholar] [CrossRef]
- Kolbert, Z. Strigolactone-nitric oxide interplay in plants: The story has just begun. Physiologia Plantarum 2019, 165, 487–497. [Google Scholar] [CrossRef]
- Angelo, D.M.; Annamaria, V.; Christophe, B.; Clementina, S. Bioprospecting of phytohormone biosynthetic pathways in the microalgal realm. Algal Research 2023, 76, 103307. [Google Scholar] [CrossRef]
- Motohashi, K.; Takagi, M.; Yamamura, H.; Hayakawa, M.; Shin-ya, K. A new angucycline and a new butenolide isolated from lichen-derived Streptomyces spp. The Journal of Antibiotics 2010, 63, 545–548. [Google Scholar] [CrossRef]
- Vallet, M.; Chong, Y.-M.; Tourneroche, A.; Genta-Jouve, G.; Hubas, C.; Lami, R.; Gachon, C.M.M.; Klochkova, T.; Chan, K.-G.; Prado, S. Novel α-Hydroxy γ-Butenolides of Kelp Endophytes Disrupt Bacterial Cell-to-Cell Signaling. Frontiers in Marine Science 2020, 7. [Google Scholar] [CrossRef]
- Shu, L.; Li, J.; Xu, J.; Zheng, Z. Nutrient removal and biogas upgrade using co-cultivation of Chlorella vulgaris and three different bacteria under various GR24 concentrations by induction with 5-deoxystrigol. World Journal of Microbiology and Biotechnology 2023, 39, 245. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Xue, Z.; Liu, J.; Sun, S.; Zhao, Y.; Zhang, H. Observation of few GR24 induced fungal-microalgal pellets performance for higher pollutants removal and biogas quality improvement. Energy 2022, 244, 123171. [Google Scholar] [CrossRef]
- Cheng, P.; Yang, H.; Wang, Z.; Zhao, C.; Lu, B.; Zhang, H.; Zhao, Y. The effects of influent 5-Deoxystrigol concentrations on integral biogas upgrading and nutrient removal by different algal-fungal-bacterial consortium. Algal Research 2024, 84, 103770. [Google Scholar] [CrossRef]
- Proust, H.; Hoffmann, B.; Xie, X.; Yoneyama, K.; Schaefer, D.G.; Yoneyama, K.; Nogué, F.; Rameau, C. Strigolactones regulate protonema branching and act as a quorum sensing-like signal in the moss Physcomitrella patens. Development 2011, 138, 1531–1539. [Google Scholar] [CrossRef]
- Wheeldon, C.D.; Hamon-Josse, M.; Lund, H.; Yoneyama, K.; Bennett, T. Environmental strigolactone drives early growth responses to neighboring plants and soil volume in pea. Current Biology 2022, 32, 3593–3600.e3593. [Google Scholar] [CrossRef]
- Peláez-Vico, M.A.; Bernabéu-Roda, L.; Kohlen, W.; Soto, M.J.; López-Ráez, J.A. Strigolactones in the Rhizobium-legume symbiosis: Stimulatory effect on bacterial surface motility and down-regulation of their levels in nodulated plants. Plant Science 2016, 245, 119–127. [Google Scholar] [CrossRef]
- Mozes, C.; Meijler, M.M. Modulation of Bacterial Quorum Sensing by Strigolactones. ACS Chemical Biology 2020, 15, 2055–2059. [Google Scholar] [CrossRef]
- Pruzzo, C.; Vezzulli, L.; Colwell, R.R. Global impact of Vibrio cholerae interactions with chitin. Environmental Microbiology 2008, 10, 1400–1410. [Google Scholar] [CrossRef]
- Gomaa, M.; Ali, M.M.A. Enhancement of microalgal biomass, lipid production and biodiesel characteristics by mixotrophic cultivation using enzymatically hydrolyzed chitin waste. Biomass and Bioenergy 2021, 154, 106251. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y. Microalgae: New Source of Plant Biostimulants. Agronomy 2020, 10, 1240. [Google Scholar] [CrossRef]
- Miranda, A.M.; Hernandez-Tenorio, F.; Villalta, F.; Vargas, G.J.; Sáez, A.A. Advances in the Development of Biofertilizers and Biostimulants from Microalgae. Biology 2024, 13, 199. [Google Scholar] [CrossRef] [PubMed]
- El Arroussi, H.; Elbaouchi, A.; Benhima, R.; Bendaou, N.; Smouni, A.; Wahby, I. Halophilic microalgae Dunaliella salina extracts improve seed germination and seedling growth of Triticum aestivum L. under salt stress. 2016; pp. 13–26.
- Guzmán-Murillo, M.A.; Ascencio, F.; Larrinaga-Mayoral, J.A. Germination and ROS detoxification in bell pepper (Capsicum annuum L.) under NaCl stress and treatment with microalgae extracts. Protoplasma 2013, 250, 33–42. [Google Scholar] [CrossRef]
- Nezamdoost, D.; Ghahremani, Z.; Ranjbar, M.E. Seed priming with Chlorella vulgaris extract increases salt tolerance threshold of leafy lettuce more than two times based on germination indices (a modeling approach). Journal of Applied Phycology 2024, 36, 1247–1258. [Google Scholar] [CrossRef]
- Sarsekeyeva, F.K.; Sadvakasova, A.K.; Sandybayeva, S.K.; Kossalbayev, B.D.; Huang, Z.; Zayadan, B.K.; Akmukhanova, N.R.; Leong, Y.K.; Chang, J.-S.; Allakhverdiev, S.I. Microalgae- and cyanobacteria-derived phytostimulants for mitigation of salt stress and improved agriculture. Algal Research 2024, 82, 103686. [Google Scholar] [CrossRef]
- Zeng, H.; Chen, H.; Zhang, M.; Ding, M.; Xu, F.; Yan, F.; Kinoshita, T.; Zhu, Y. Plasma membrane H+-ATPases in mineral nutrition and crop improvement. Trends in Plant Science 2024, 29, 978–994. [Google Scholar] [CrossRef]
- Zandonadi, D.B.; Santos, M.P.; Caixeta, L.S.; Marinho, E.B.; Peres, L.E.P.; Façanha, A.R. Plant proton pumps as markers of biostimulant action. Scientia Agricola 2016, 73, 24–28. [Google Scholar] [CrossRef]
- Ramakrishnan, B.; Maddela, N.R.; Venkateswarlu, K.; Megharaj, M. Potential of microalgae and cyanobacteria to improve soil health and agricultural productivity: a critical view. Environmental Science: Advances 2023, 2, 586–611. [Google Scholar] [CrossRef]
- Lee, S.-M.; Ryu, C.-M. Algae as New Kids in the Beneficial Plant Microbiome. Frontiers in Plant Science 2021, 12. [Google Scholar] [CrossRef]
- Gonçalves, J.; Freitas, J.; Fernandes, I.; Silva, P. Microalgae as Biofertilizers: A Sustainable Way to Improve Soil Fertility and Plant Growth. Sustainability 2023, 15, 12413. [Google Scholar] [CrossRef]
- Aigner, S.; Glaser, K.; Arc, E.; Holzinger, A.; Schletter, M.; Karsten, U.; Kranner, I. Adaptation to Aquatic and Terrestrial Environments in Chlorella vulgaris (Chlorophyta). Frontiers in Microbiology 2020, 11. [Google Scholar] [CrossRef]
- Kholssi, R.; Marks, E.A.N.; Miñón, J.; Montero, O.; Debdoubi, A.; Rad, C. Biofertilizing Effect of Chlorella sorokiniana Suspensions on Wheat Growth. Journal of Plant Growth Regulation 2019, 38, 644–649. [Google Scholar] [CrossRef]
- Chader, S.; Mahmah, B.; Chetehouna, K.; Amrouche, F.; Abdeladim, K. Biohydrogen production using green microalgae as an approach to operate a small proton exchange membrane fuel cell. international journal of hydrogen energy 2011, 36, 4089–4093. [Google Scholar] [CrossRef]
- Minaoui, F.; Hakkoum, Z.; Chabili, A.; Douma, M.; Mouhri, K.; Loudiki, M. Biostimulant effect of green soil microalgae Chlorella vulgaris suspensions on germination and growth of wheat (Triticum aestivum var. Achtar) and soil fertility. Algal Research 2024, 82, 103655. [Google Scholar] [CrossRef]
- López-Ráez, J.A.; Pozo, M.J.; García-Garrido, J.M. Strigolactones: a cry for help in the rhizosphere. Botany 2011, 89, 513–522. [Google Scholar] [CrossRef]
- Nasir, F.; Shi, S.; Tian, L.; Chang, C.; Ma, L.; Li, X.; Gao, Y.; Tian, C. Strigolactones shape the rhizomicrobiome in rice (Oryza sativa). Plant Science 2019, 286, 118–133. [Google Scholar] [CrossRef]
- Chen, P.; Huang, P.; Yu, H.; Yu, H.; Xie, W.; Wang, Y.; Zhou, Y.; Chen, L.; Zhang, M.; Yao, R. Strigolactones shape the assembly of root-associated microbiota in response to phosphorus availability. mSystems 2024, 9, e01124–e01123. [Google Scholar] [CrossRef]
- Soliman, S.; Wang, Y.; Han, Z.; Pervaiz, T.; El-kereamy, A. Strigolactones in Plants and Their Interaction with the Ecological Microbiome in Response to Abiotic Stress. Plants 2022, 11, 3499. [Google Scholar] [CrossRef]
- López-Ráez, J.A. How drought and salinity affect arbuscular mycorrhizal symbiosis and strigolactone biosynthesis? Planta 2016, 243, 1375–1385. [Google Scholar] [CrossRef]
- Elisabeth, B.; Rayen, F.; Behnam, T. Microalgae culture quality indicators: a review. Critical Reviews in Biotechnology 2021, 41, 457–473. [Google Scholar] [CrossRef] [PubMed]
- Bala, I.; Airinei, A.; Georgescu, E.; Oancea, F.; Georgescu, F.; Nicolescu, A.; Tigoianu, R.; Deleanu, C. Photophysical and biological properties of a strigolactone mimic derived from 1,8-naphthalic anhydride. Rev Roum Chim 2022, 67, 51–62. [Google Scholar]
- Kim, H.S.; Park, W.-K.; Lee, B.; Seon, G.; Suh, W.I.; Moon, M.; Chang, Y.K. Optimization of heterotrophic cultivation of Chlorella sp. HS2 using screening, statistical assessment, and validation. Scientific Reports 2019, 9, 19383. [Google Scholar] [CrossRef]
- MJ, G.; C, G.; RP, v.H.; ST, H. Interference by pigment in the estimation of microalgal biomass concentration by optical density. Journal of microbiological methods 2011, 85. [Google Scholar] [CrossRef]
- Chai, S.; Shi, J.; Huang, T.; Guo, Y.; Wei, J.; Guo, M.; Li, L.; Dou, S.; Liu, L.; Liu, G. Characterization of Chlorella sorokiniana growth properties in monosaccharide-supplemented batch culture. PLoS One 2018, 13, e0199873. [Google Scholar] [CrossRef] [PubMed]
- Jeon, B.-H.; Choi, J.-A.; Kim, H.-C.; Hwang, J.-H.; Abou-Shanab, R.A.; Dempsey, B.A.; Regan, J.M.; Kim, J.R. Ultrasonic disintegration of microalgal biomass and consequent improvement of bioaccessibility/bioavailability in microbial fermentation. Biotechnology for Biofuels 2013, 6, 37. [Google Scholar] [CrossRef]
- Gilbert, G.S.; Diaz, A.; Bregoff, H.A. Seed Disinfestation Practices to Control Seed-Borne Fungi and Bacteria in Home Production of Sprouts. Foods 2023, 12, 747. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nature Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Tritean, N.; Trică, B.; Dima, Ş.-O.; Capră, L.; Gabor, R.-A.; Cimpean, A.; Oancea, F.; Constantinescu-Aruxandei, D. Mechanistic insights into the plant biostimulant activity of a novel formulation based on rice husk nanobiosilica embedded in a seed coating alginate film. Frontiers in Plant Science 2024, 15. [Google Scholar] [CrossRef]
Figure 1.
Optical density of C. sorokiniana cultures during 15 days of culturing. Control - growth media without treatment; SC-DMSO 0.1%, c1: 1 × 10-7 M, c2: 1 × 10-8 M, c3: 1 × 10-9. Values represent means ± standard errors (n = 3); different letters show statistically significant differences at p < 0.05.
Figure 1.
Optical density of C. sorokiniana cultures during 15 days of culturing. Control - growth media without treatment; SC-DMSO 0.1%, c1: 1 × 10-7 M, c2: 1 × 10-8 M, c3: 1 × 10-9. Values represent means ± standard errors (n = 3); different letters show statistically significant differences at p < 0.05.
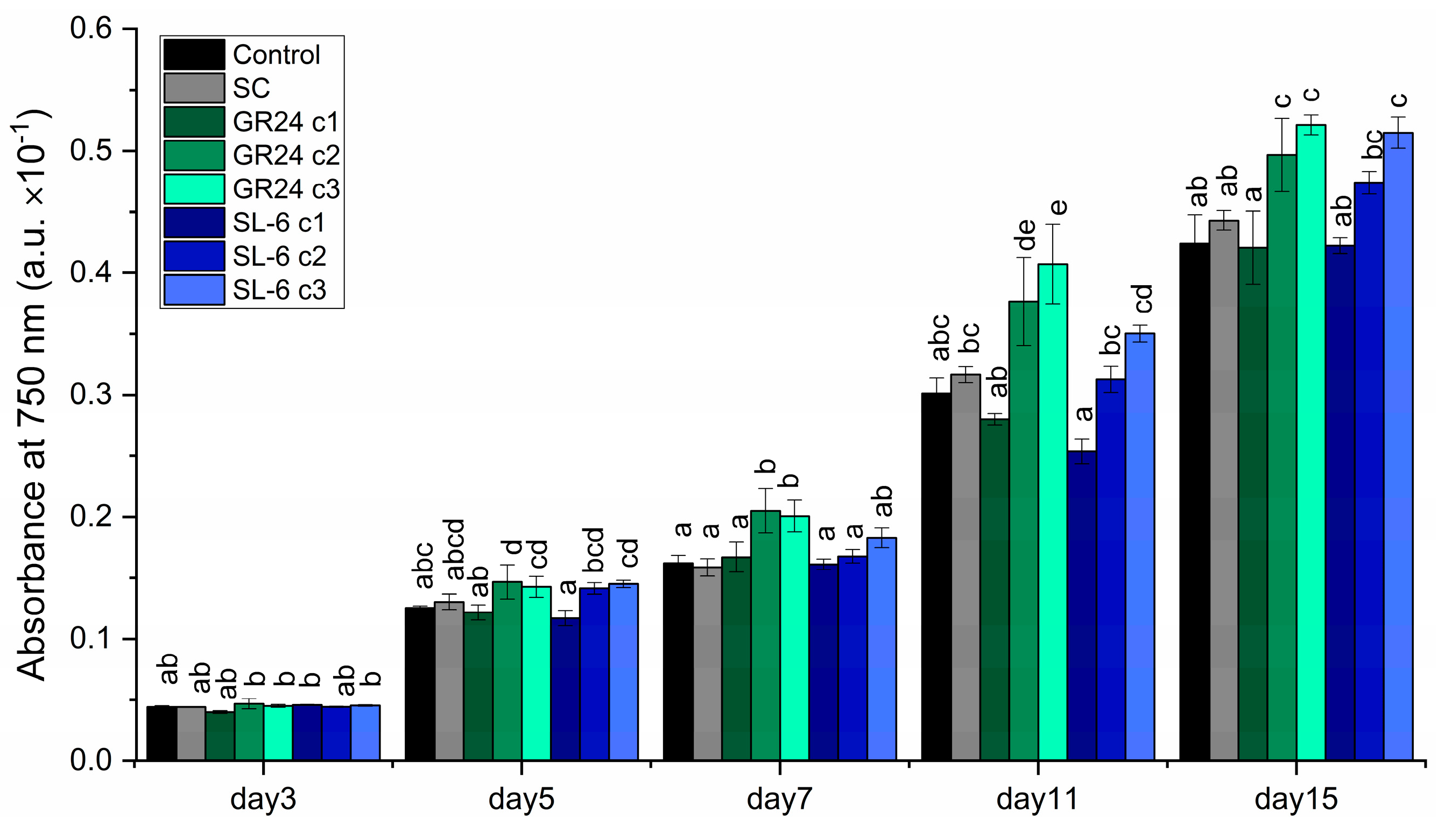
Figure 2.
Turbidity of C. sorokiniana cultures during 15 days of culturing. Control - growth media without treatment; SC-DMSO 0.1%, c1: 1 ×10-7 M, c2: 1 × 10-8 M, c3: 1 × 10-9. Values represent means ± standard errors (n = 3); different letters show statistically significant differences at p < 0.05.
Figure 2.
Turbidity of C. sorokiniana cultures during 15 days of culturing. Control - growth media without treatment; SC-DMSO 0.1%, c1: 1 ×10-7 M, c2: 1 × 10-8 M, c3: 1 × 10-9. Values represent means ± standard errors (n = 3); different letters show statistically significant differences at p < 0.05.
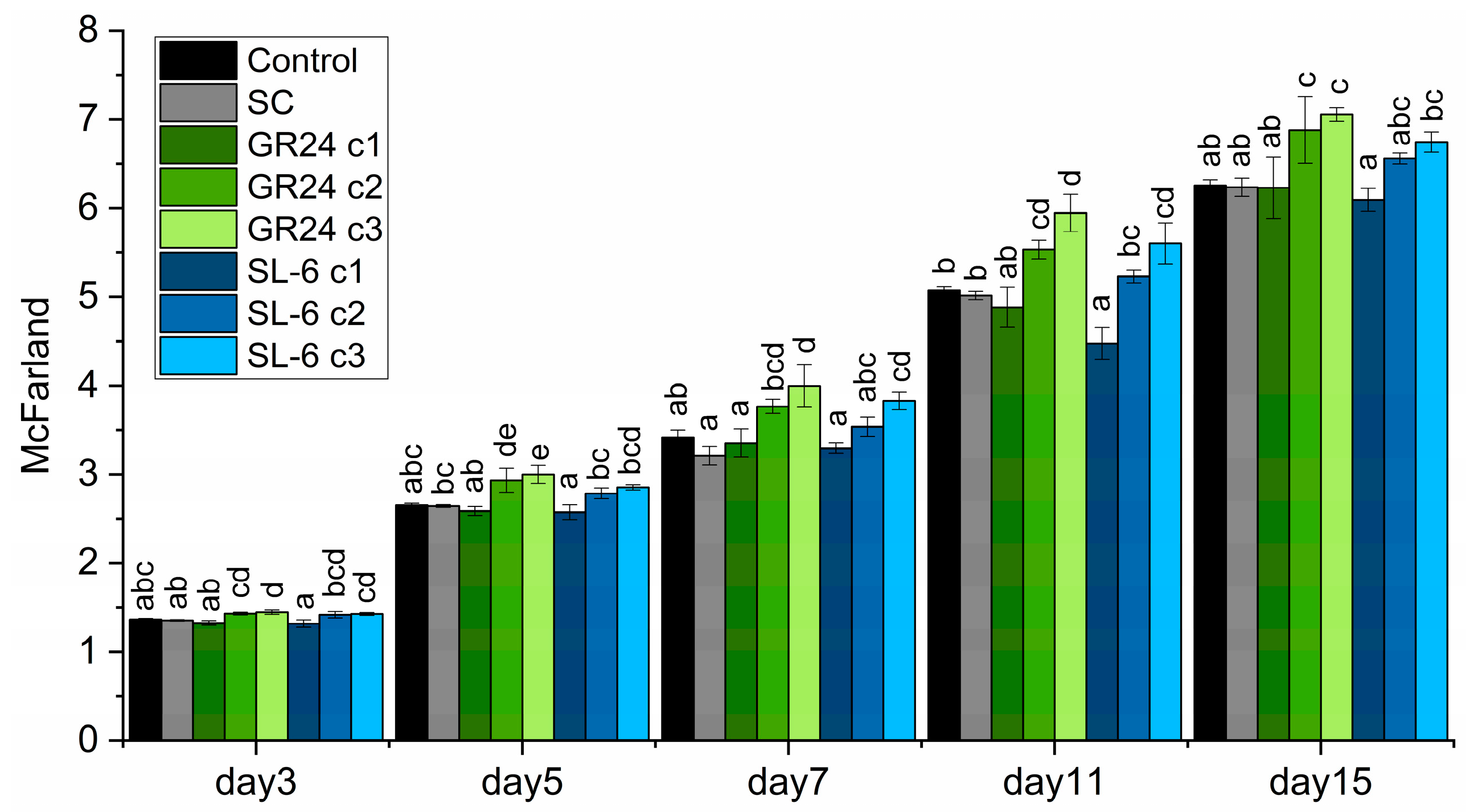
Figure 3.
Biomass quantity of C. sorokiniana cultures. Control - growth media without treatment; SC-DMSO 0.1%, c1: 1 ×10-7 M, c2: 1 × 10-8 M, c3: 1 × 10-9 M. Values represent means ± standard errors (n = 3); different letters show statistically significant differences at p < 0.05.
Figure 3.
Biomass quantity of C. sorokiniana cultures. Control - growth media without treatment; SC-DMSO 0.1%, c1: 1 ×10-7 M, c2: 1 × 10-8 M, c3: 1 × 10-9 M. Values represent means ± standard errors (n = 3); different letters show statistically significant differences at p < 0.05.

Figure 4.
Pigment concentrations. (a) Chlorophyll a, (b) Chlorophyll b, (c) Total carotenoids, (d) Total piments. Control - growth media without treatment; SC-DMSO 0.1%, c1: 1 × 10-7 M, c2: 1 × 10-8 M, c3: 1 × 10-9. Values represent means ± standard errors (n = 3); different letters show statistically significant differences at p < 0.05.
Figure 4.
Pigment concentrations. (a) Chlorophyll a, (b) Chlorophyll b, (c) Total carotenoids, (d) Total piments. Control - growth media without treatment; SC-DMSO 0.1%, c1: 1 × 10-7 M, c2: 1 × 10-8 M, c3: 1 × 10-9. Values represent means ± standard errors (n = 3); different letters show statistically significant differences at p < 0.05.
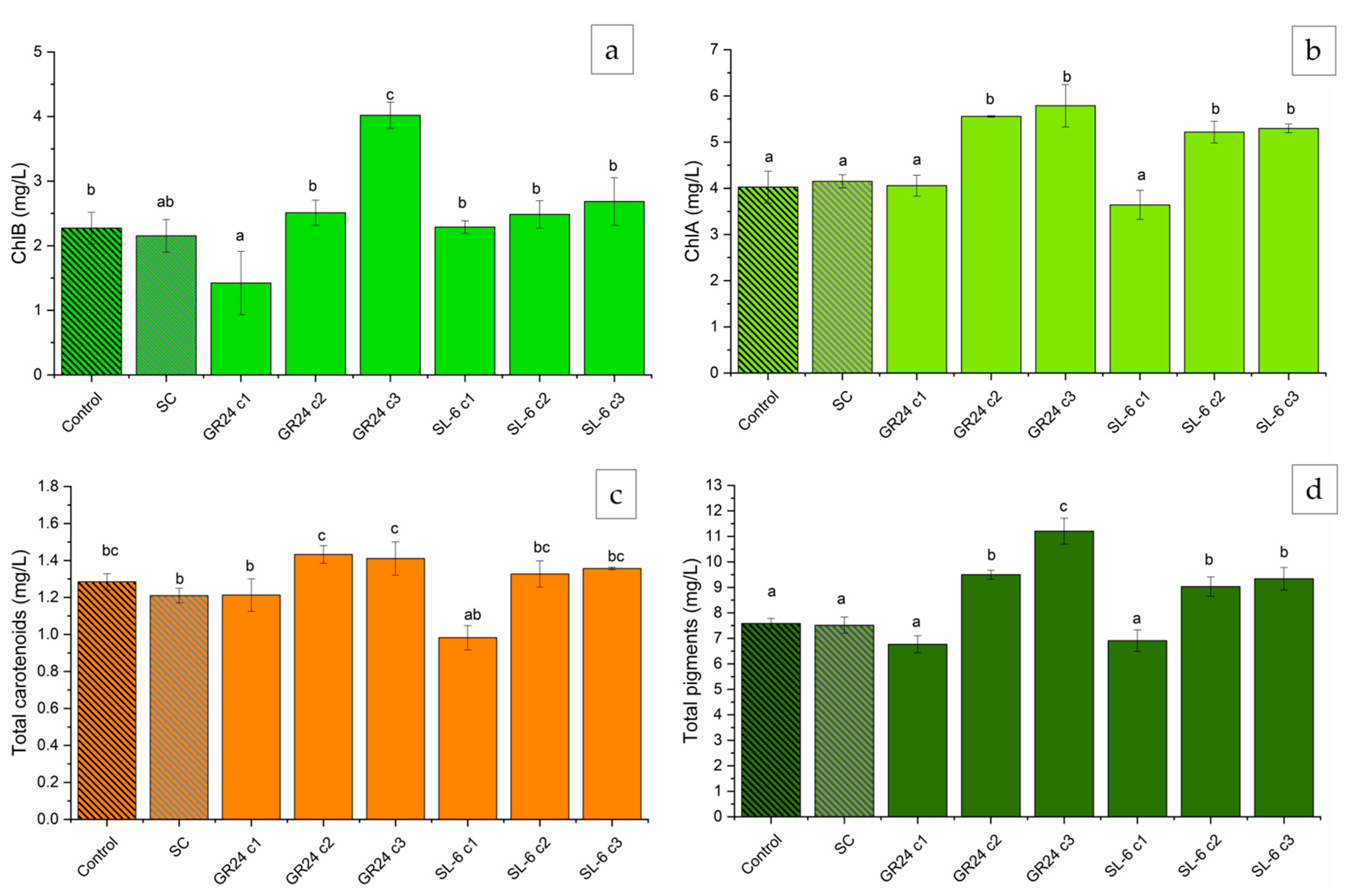
Figure 5.
Pigment concentrations in dependence of microalgal biomass. (a) Chlorophyll a, (b) Chlorophyll b, (c) Total carotenoids, (d) Total piments. Control - growth media without treatment; SC-DMSO 0.1%, c1: 1 ×10-7 M, c2: 1 × 10-8 M, c3: 1 × 10-9 M. Values represent means ± standard errors (n = 3); different letters show statistically significant differences at p < 0.05.
Figure 5.
Pigment concentrations in dependence of microalgal biomass. (a) Chlorophyll a, (b) Chlorophyll b, (c) Total carotenoids, (d) Total piments. Control - growth media without treatment; SC-DMSO 0.1%, c1: 1 ×10-7 M, c2: 1 × 10-8 M, c3: 1 × 10-9 M. Values represent means ± standard errors (n = 3); different letters show statistically significant differences at p < 0.05.
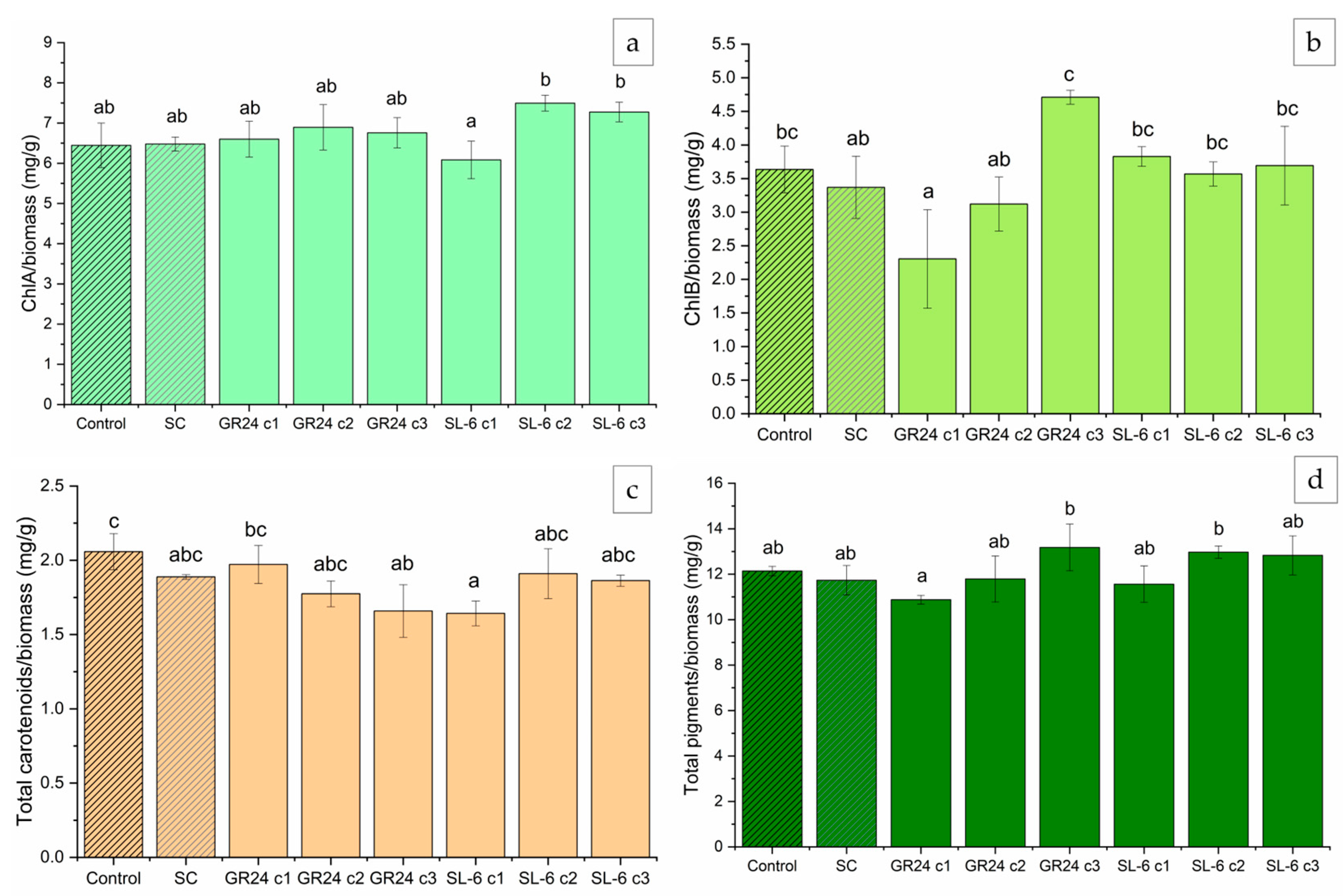
Figure 6.
Chlorophyll fluorescence of microalgal cultures. Control - growth media without treatment; SC-DMSO 0.1%, c1: 1 ×10-7 M, c2: 1 × 10-8 M, c3: 1 × 10-9 M. Values represent means ± standard errors (n = 3); different letters show statistically significant differences at p < 0.05.
Figure 6.
Chlorophyll fluorescence of microalgal cultures. Control - growth media without treatment; SC-DMSO 0.1%, c1: 1 ×10-7 M, c2: 1 × 10-8 M, c3: 1 × 10-9 M. Values represent means ± standard errors (n = 3); different letters show statistically significant differences at p < 0.05.
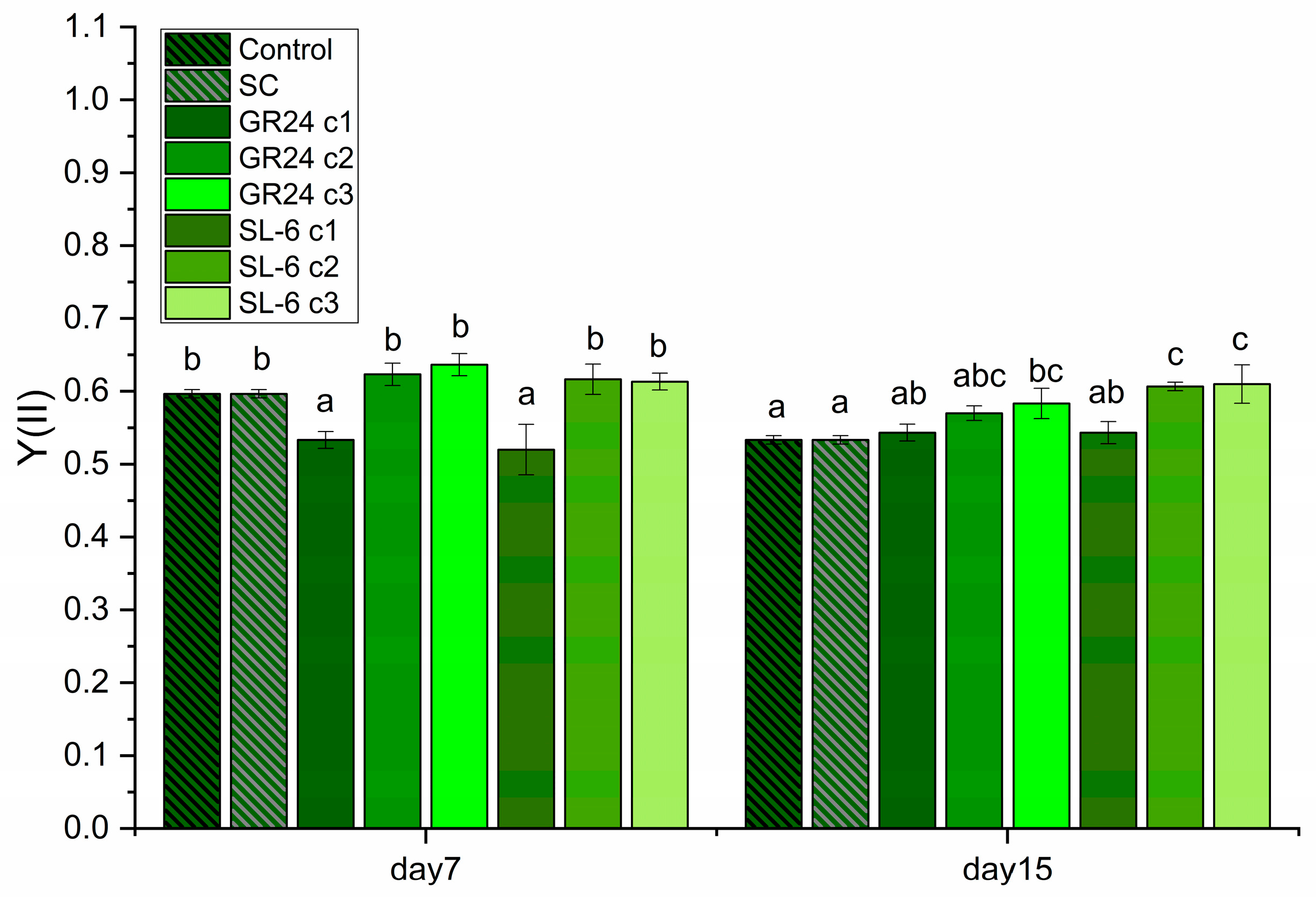
Figure 7.
Pearson correlation between the growth parameters and pigment production of C. sorokiniana grown in the absence and presence of GR24 and SL-6; ** Correlation is significant at the 0.01 level; * Correlation is significant at the 0.05 level.
Figure 7.
Pearson correlation between the growth parameters and pigment production of C. sorokiniana grown in the absence and presence of GR24 and SL-6; ** Correlation is significant at the 0.01 level; * Correlation is significant at the 0.05 level.
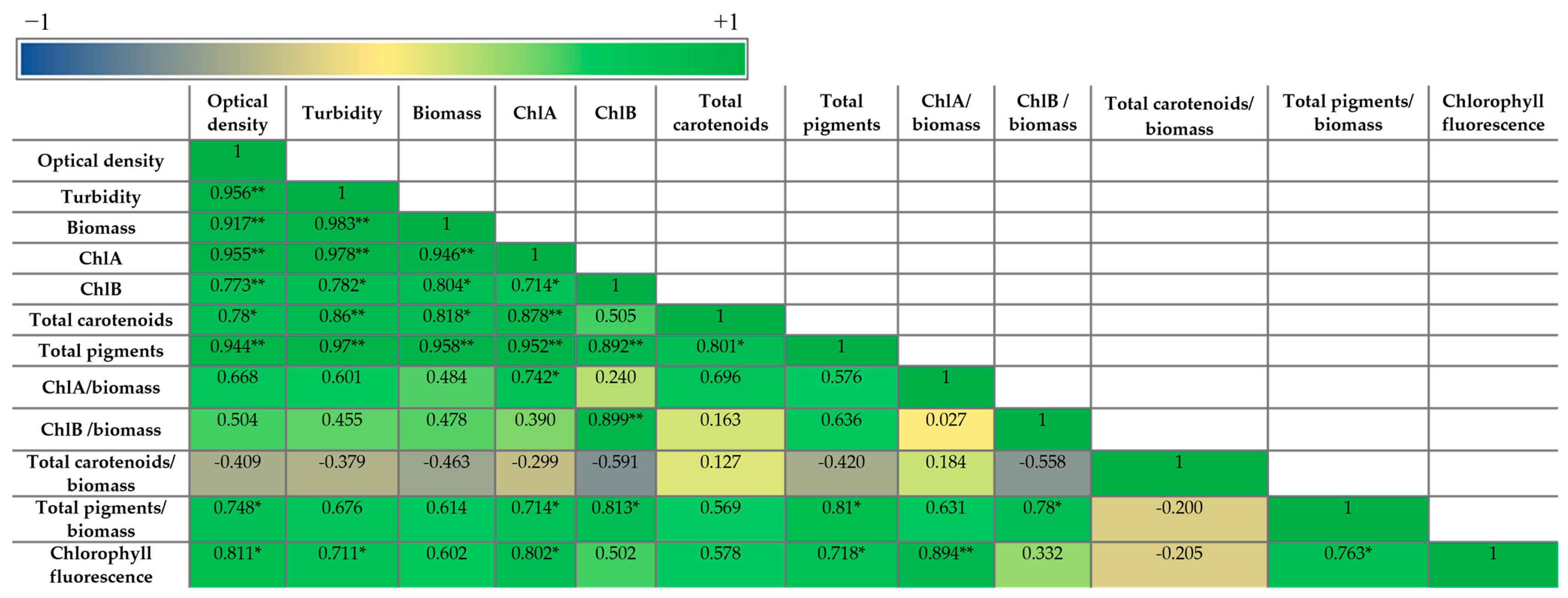
Figure 8.
Mung seedlings measurements. C, control with water; CSk, control with untreated C. sorokiniana biomass extract; Sk-7, extract from C. sorokiniana biomass treated with SL-6 c1 (1 × 10−7 M); Sk-8, extract from C. sorokiniana biomass treated with SL-6 c2(1 × 10−8 M); Sk-9, extract from C. sorokiniana biomass treated with SL-6 c3 (1 × 10−9 M); C-7, control for SL-6 c1 solution; C-8, control for SL-6 c2 solution; C-9, control for SL-6c3 solution; CS, control with water and salt stress; CSkS, control for untreated C. sorokiniana biomass extract with salt stress; Sk-7, extract from C. sorokiniana biomass treated with SL-6 c1 (1 × 10−7 M) with salt stress; Sk-8, extract from C. sorokiniana biomass treated with SL-6 c2 (1 × 10−8 M) with salt stress; Sk-9, extract from C. sorokiniana biomass treated with SL-6c3 (1 × 10−9 M) with salt stress; C-7, control for SL-6 c1 solution with salt stress; C-8, control for SL-6c2 solution with salt stress; C-9, control for SL-6 c3 solution with salt stress; Values represent means ± standard deviations. Different letters indicate statistically significant differences between samples at p < 0.05.
Figure 8.
Mung seedlings measurements. C, control with water; CSk, control with untreated C. sorokiniana biomass extract; Sk-7, extract from C. sorokiniana biomass treated with SL-6 c1 (1 × 10−7 M); Sk-8, extract from C. sorokiniana biomass treated with SL-6 c2(1 × 10−8 M); Sk-9, extract from C. sorokiniana biomass treated with SL-6 c3 (1 × 10−9 M); C-7, control for SL-6 c1 solution; C-8, control for SL-6 c2 solution; C-9, control for SL-6c3 solution; CS, control with water and salt stress; CSkS, control for untreated C. sorokiniana biomass extract with salt stress; Sk-7, extract from C. sorokiniana biomass treated with SL-6 c1 (1 × 10−7 M) with salt stress; Sk-8, extract from C. sorokiniana biomass treated with SL-6 c2 (1 × 10−8 M) with salt stress; Sk-9, extract from C. sorokiniana biomass treated with SL-6c3 (1 × 10−9 M) with salt stress; C-7, control for SL-6 c1 solution with salt stress; C-8, control for SL-6c2 solution with salt stress; C-9, control for SL-6 c3 solution with salt stress; Values represent means ± standard deviations. Different letters indicate statistically significant differences between samples at p < 0.05.
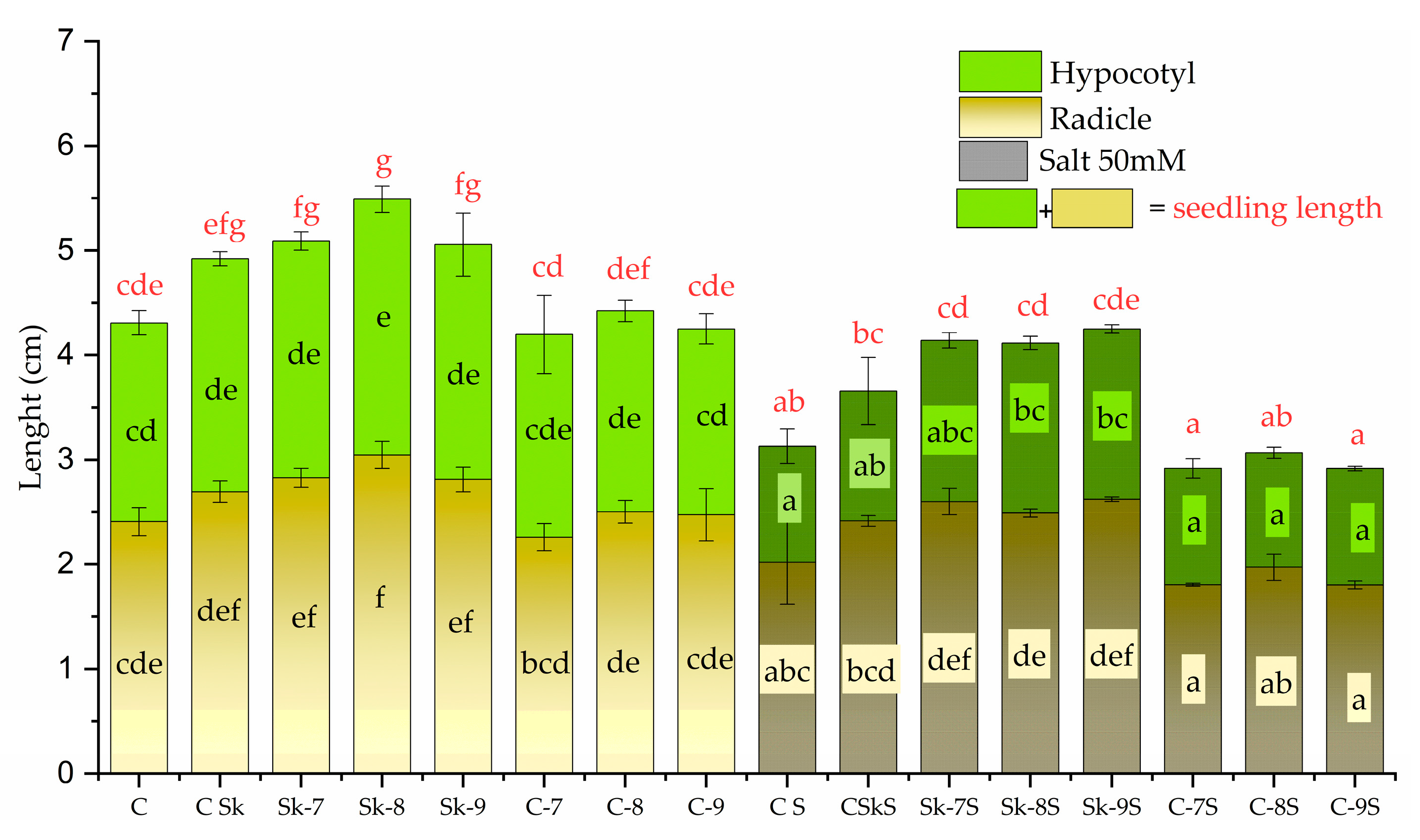
Figure 9.
Proton pump activity; (a) Total extracellular H+ level; (b) Specific extracellular H+ level; (c) petri dishes after medium acidification test; C, control with water; CSk, control for untreated C. sorokiniana biomass extract; Sk-7, extract from C. sorokiniana biomass treated with SL-6 c1(1 × 10−7 M); Sk-8, extract from C. sorokiniana biomass treated with SL-6 c2 (1 × 10−8 M); Sk-9, extract from C. sorokiniana biomass treated with SL-6 c3 (1 × 10−9 M); C-7, control for SL-6 c1 solution; C-8, control for SL-6 c2 solution; C-9, control for SL-6 c3 solution; CS, control with water and salt stress; CSkS, control for untreated C. sorokiniana biomass extract with salt stress; Sk-7, extract from C. sorokiniana biomass treated with SL-6 c1 (1 × 10−7 M) with salt stress; Sk-8, extract from C. sorokiniana biomass treated with SL-6 c2 (1 × 10−8 M) with salt stress; Sk-9, extract from C. sorokiniana biomass treated with SL-6 c3 (1 × *10−9 M) with salt stress; C-7, control for SL-6 c1 solution with salt stress; C-8, control for SL-6 c2 solution with salt stress; C-9, control for SL-6 c3 solution with salt stress; values represent means ± standard deviations. Different letters indicate statistically significant differences between samples at p < 0.05.
Figure 9.
Proton pump activity; (a) Total extracellular H+ level; (b) Specific extracellular H+ level; (c) petri dishes after medium acidification test; C, control with water; CSk, control for untreated C. sorokiniana biomass extract; Sk-7, extract from C. sorokiniana biomass treated with SL-6 c1(1 × 10−7 M); Sk-8, extract from C. sorokiniana biomass treated with SL-6 c2 (1 × 10−8 M); Sk-9, extract from C. sorokiniana biomass treated with SL-6 c3 (1 × 10−9 M); C-7, control for SL-6 c1 solution; C-8, control for SL-6 c2 solution; C-9, control for SL-6 c3 solution; CS, control with water and salt stress; CSkS, control for untreated C. sorokiniana biomass extract with salt stress; Sk-7, extract from C. sorokiniana biomass treated with SL-6 c1 (1 × 10−7 M) with salt stress; Sk-8, extract from C. sorokiniana biomass treated with SL-6 c2 (1 × 10−8 M) with salt stress; Sk-9, extract from C. sorokiniana biomass treated with SL-6 c3 (1 × *10−9 M) with salt stress; C-7, control for SL-6 c1 solution with salt stress; C-8, control for SL-6 c2 solution with salt stress; C-9, control for SL-6 c3 solution with salt stress; values represent means ± standard deviations. Different letters indicate statistically significant differences between samples at p < 0.05.
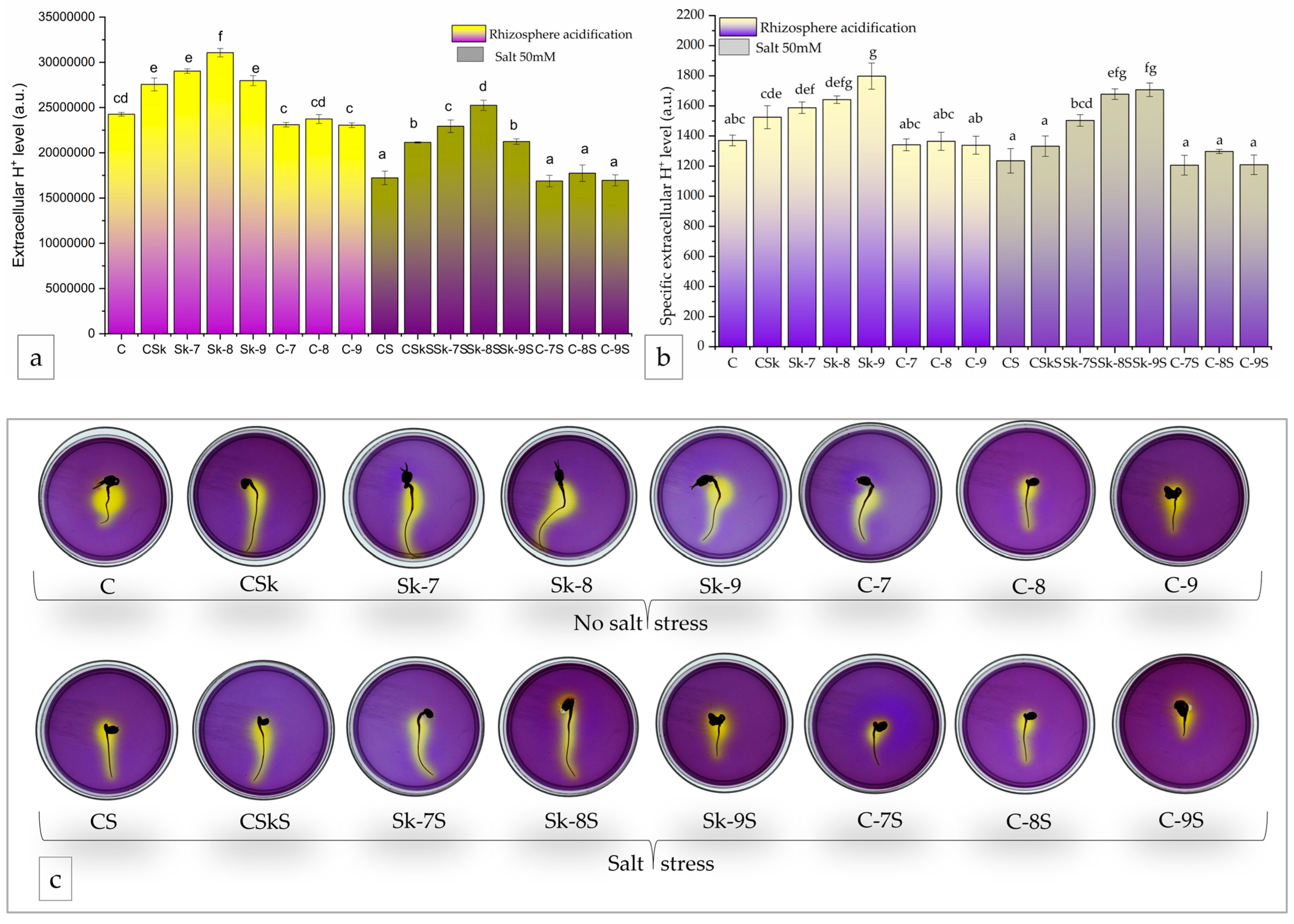
Figure 10.
Pearson correlation between the mung seedling parameters and microalga C. sorokiniana parameters in the absence (a) and presence (b) of salt stress on the mung seedlings; * Correlation is significant at the 0.05 level; ** Correlation is significant at the 0.01 level.
Figure 10.
Pearson correlation between the mung seedling parameters and microalga C. sorokiniana parameters in the absence (a) and presence (b) of salt stress on the mung seedlings; * Correlation is significant at the 0.05 level; ** Correlation is significant at the 0.01 level.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



