
Submitted:
12 January 2025
Posted:
13 January 2025
You are already at the latest version
Abstract
This paper presents a novel study on the microbial diversity of Nodipecten nodosus in aquaculture waters and its production. The Lion’s paw scallop (Nodipecten nodosus) is the most economically relevant organism, highly farmed in southeastern Brazil. However, the microbiological contamination of these animals can make their consumption unfeasible. This study assessed the bacterial diversity of Lion’s paw scallop gill arch tissues and the surrounding water of a scallop farm in Angra dos Reis, Rio de Janeiro, Brazil. The isolates (n = 216) were submitted to MALDI-TOF MS identification. Susceptibility to antimicrobials and the ability to produce carbapenems were analyzed by disk diffusion. Studies with isolates of Pseudomonas putida exhibited the highest percentage of resistance to ciprofloxin (42%). All Pseudomonas putida isolates were sensitive to meropenem. Vibrio alginolyticos and Vibrio fluvialis exhibited greater resistance to meropenem. The presence of resistant Vibrio spp. and Pseudomonas spp. and potentially pathogenic bacteria in the analyzed waters and gill arch tissues highlights potential antimicrobial resistance reservoirs, suggesting public health risks regarding bathing in the waters and consuming Lion's paw scallops from the analyzed farm.
Keywords:
Marine pollution
; Marine bivalves
; Antimicrobial susceptibility
; Pseudomonas spp
; Vibrio spp.
1. Introduction
The species Nodipecten nodosus [1], known as scallop, is a bivalve of the Pectinidae family with great economic value in the global market. It is essential for mariculture in several countries, including Brazil [2,3,4]. These bivalves play a fundamental role in aquatic ecosystems by filtering water and helping to maintain its quality. In addition, their sensitivity to pollutants makes them effective bioindicators for monitoring environmental health [5].
Growing environmental concerns in commercial production make it essential to ensure the health and growth of these organisms, as recorded in several studies [6,7,8,9]. Bivalves are a prominent commodity in aquaculture, with significant global production and economic importance in fishing activities worldwide [10].
Bivalves are benthic organisms that filter large volumes of water to obtain food and oxygen, which exposes them to harmful agents such as heavy metals (e.g., mercury, cadmium, lead), phytoplankton toxins, pathogenic bacteria (e., g. Vibrio spp., Escherichia coli), viruses, protozoa, and saprophytic fungi [11,12,13,14,15]. Thus, RDC No. 1, of July 7, 2022, from ANVISA, establishes microbiological standards for food and their applications. This legislation applies to bivalves since they are often consumed in nature and, therefore, can transmit etiological agents that cause diseases, representing a risk to public health [16].
The nutritional properties of fish, crustaceans, and mollusks from mariculture and aquaculture are widely recognized for their benefits to human health [17]. Mariculture can alleviate pressure on natural fish populations threatened by overfishing and ecosystem degradation [18,19,20]. In this scenario, studies have reported that high pollution caused by sewage, associated with scallop mortality, has negative effects on water quality degradation [21,22,23], which compromises the health of organisms, increasing vulnerability to diseases and reducing productivity [24,25]. Pathogens such as E. coli, Salmonella spp., and enteric viruses, such as norovirus, are frequently found in polluted waters and cause foodborne illness outbreaks [26].
Santos et al. [27] highlighted the growing concern regarding global production and trade. The World Organization for Animal Health [28] listed notifiable pathogens present in shellfish, including herpesvirus, abalone, parasites such as Bonamia exits, B. ostreae, Perkinsus marinus, P. Olsen, and Marteilia refringent, as well as bacteria such as Xenohaliotis californiensis. Although not officially listed, bacteria and fungi in shellfish pose risks to production and human health [27]. Among the microorganisms of interest to public health are bacterial species of the genera Vibrio, Pseudomonas, Aeromonas and Salmonella, in addition to diarrheagenic pathotypes of Escherichia coli and fungi of the genus Aspergillus [29,30,31,32,33,34,35].
Thompson et al. [36] warned that the release of untreated sewage can increase nutrient levels in the water, favoring the growth of pathogens such as Vibrio, which harms the health of scallops. The analyses indicated a greater abundance of microorganisms in the summer, suggesting a negative correlation with scallop production. For example, V. alginolyticus expresses several virulence factors, such as enzymes and toxins that facilitate colonization. The spread of this pathogen can cause disease outbreaks in mussel farms, threatening the health of scallops [37].
In view of this, the problems faced by Brazilian mariculture, especially in Rio de Janeiro, have their most sensitive point in the absence of an in-depth study of the biotic and abiotic parameters involved in this productive activity. One of the points of susceptibility of mariculture is the lack of knowledge of the problems that affect the cultivation lines, whether due to abiotic factors, such as the physical-chemical factors of the surrounding environment, or biotic factors, such as the development of bacterial flora that may produce changes in the stocks and generate considerable population losses for the growers [38]. For example, one of the main factors for the deterioration of water quality in Ilha Grande Bay (BIG), in the municipality of Angra dos Reis, Rio de Janeiro, is the rapid population growth that increased by approximately 73.9% between 2010 and 2021 [39], generating greater pressure on water resources and intensifying the discharge of domestic and industrial effluents [36].
Antimicrobial resistance (AMR) seriously threatens public health, resulting from increased multidrug-resistant (MDR) microorganisms resistant to existing treatments. The development of AMR occurs naturally due to genetic variability and bacterial interactions; however, it is accelerated by selective pressure from the indiscriminate use of antimicrobials in human and veterinary medicine and food production [40,41].
In Brazil, the Ministry of Agriculture, Livestock, and Supply (MAPA) is responsible for inspection through Ordinance No. 171 of 2018, issued by the Secretariat of Agricultural Defense (SDA) and endorsed by ANVISA, which imposes restrictions on the use of antimicrobials essential for human medicine as zootechnical additives but still allows their use as growth promoters [42].
Thus, due to the recurring drop in production and increase in mortality on aquaculture farms that cultivate Nodipecten nodosus at BIG in Angra dos Reis/RJ since 2018 [43], this species was chosen as the object of this study. Its high sensitivity, resulting from exclusive feeding by filtration of suspended particles, makes it vulnerable to infections by microbiological pathogens, such as Vibrio spp. and E. coli. Therefore, understanding these factors is essential to improve management practices and reduce losses on aquaculture farms.
This study aimed to isolate and identify microbiological contaminants in water samples and Nodipecten nodosus collected in three areas: Bananal Island Cove, Gipóia Island Cove, and Náutica Beach Cove, in Jacuecanga, to evaluate the sanitary conditions of the bivalve mollusk cultivation area at BIG, in Angra dos Reis/RJ, as well as to investigate the impact of these conditions on the quality and health of the cultivated mollusks. In addition, the research analyzed how these factors influence the environmental sustainability of the activity and contribute to predicting risks to public health according to the limits established by current legislation.
2. Materials and Methods
2.1. Sample Collection
Between February and December 2021, eleven collections were carried out, totaling (n = 55) liters of water and (n = 275) scallops, in the following mariculture farms on the south coast of Rio de Janeiro: (1) Jacuecanga Beach Cove, located between 23º00’29” S and 44º14’13” W; (2) Gipóia Island Cove, between 23º02’16” S and 44º23’08” W; and (3) Bananal Island Cove, between 23º06’04” S and 44º15’07” W, at a depth of 15-20 m (Figure 1). The collections were carried out in a pre-established partnership with the Eco-Development Institute of Ilha Grande Bay (IED-BIG), located at Rua EAP s/nº, Vila da Petrobrás, Angra dos Reis, RJ.
The Nodipecten nodosus scallops were collected manually randomly and subjected to the flow shown in Figure 2. All collected samples underwent strict control of removing bivalve mollusks by the criteria established by the Interministerial Normative Instruction MPA/MAPA No. 7 of May 8, 2012. This regulation determines that the results of monitoring contaminating microorganisms and biotoxins produced by marine microalgae should be used to define strategies for removing bivalve mollusks, especially those intended for human consumption.
Five scallops were collected monthly at each collection point, totaling 275 samples (n = 275). The scallop samples, with the valves closed, were packaged in sterilized polyethylene bags and adequately identified with the location and date of collection. They were then stored in a cooler with ice to maintain the temperature between 6 and 10°C until they arrived at the laboratory, where they were processed.
All monthly water samples collected for this study were obtained at each specifiedpoint (1 L per point), as shown in Figure 1, using one-liter bottles previously sterilized to avoid biological contamination by RDC No. 724/2022. Collection points: (1) Gipóia Island Cove; (2) Bananal Island Cove; and (3) Jacuecanga Beach Cove, in addition to raw water from scallop farming and ultraviolet-treated water from scallop farming. Thus, 5 L of water was collected monthly, totaling 55 L of samples over the period (n = 55).
2.2. Sample Preparation
All collected samples were sent to the Reference Microorganism Laboratory of the National Institute for Quality Control in Health (INCQS) and to the Center for Technological Development in Health (CDTS) of the Oswaldo Cruz Foundation (Fiocruz). The samples underwent an initial external washing to remove surface marine materials and were then immersed in a chlorinated water solution (5 ppm of Free Residual Chlorine – FRC) by health inspection legislation [44].
Adult scallops (n = 275), with an average size of 6.25 ± 1 cm, had their valves washed externally to remove residues by legislation. Under aseptic conditions, the valves were opened with a sterile spatula to access the soft tissues, and the adductor muscle was disconnected. The branchial arch tissue was removed with forceps and a scalpel. Then, 25 g of each sample was added to 225 ml of buffered saline solution and homogenized in the Stomacher for 60 seconds [45,46].
The filtration of the homogenized tissues of the scallops and the collected seawater was performed in a vacuum system, using cellulose acetate membranes with a porosity of 0.22 µm (EMD Millipore). The seawater, the gill arch tissues of the scallops, and the filtered membranes were inoculated in BHI (Brain Heart Infusion) broth with 1-3% sodium chloride (NaCl). Then, the samples were plated on plates with solid BHI media to isolate microorganisms. The isolated strains were cultivated in BHI agar at 35°C for 24 h.
2.3. Sample Processing
Then, one colony of each isolate was chosen, and, using the direct transfer procedure, the samples were applied to each spot of the microplate 96 MSP, Bruker® – Billerica, USA. After drying, 1 µl of 70% formic acid (Sigma-Aldrich, St Louis, MO, USA) was added over the bacterial layer to cover it and optimize cell lysis. After drying the lysis solution, 1 µl of matrix solution (α-cyano-4-hydroxycinnamic acid diluted in 50% acetonitrile and 2.5% trifluoroacetic acid+ (Sigma-Aldrich, St Louis, MO, USA) was added.
The microplates were left at room temperature until completely dry. Then, they were inserted into the Bruker MALDI-TOF LT Microflex mass spectrometer (Bruker Optic GMBH, Ettlingen, Germany) equipped with a 337 nm nitrogen laser in linear mode. The operation was controlled by the FlexControl 3.3 software (Bruker Optic GMBH, Ettlingen, Germany). The spectra were collected in the mass range between 2,000 and 20,000 m/z and subsequently analyzed by the MALDI Biotyper 2.0 software (Bruker Optic GMBH, Ettlingen, Germany), using standardized settings for bacterial identification.
The MALDI Biotyper 2.0 software analyzed the spectra of the unknown samples and compared them with reference samples in the database, categorizing the results on a scale of zero to three, where higher values indicate more accurate and reliable identification. The isolates were identified by proteomic analysis using the MALDI-TOF (Matrix-Assisted Laser Desorption/Ionization—Time of Flight) mass spectrometer in partnership with the Bacteriology and Bioassays Laboratory of the Evandro Chagas National Institute of Infectiology (INI/FIOCRUZ).
2.4. Criteria for Classifying Samples
Antimicrobial susceptibility was determined by the disk diffusion technique (Kirby-Bauer method) according to the criteria established by the European Committee on Antimicrobial Susceptibility Testing [47]. The isolates were initially plated on Brain Hearth Infusion (BHI) agar for 24h at 37°C. The microbial growth was suspended in a sterile saline solution (0.85% NaCl) to obtain a turbidity standard 0.5 on the McFarland scale. The suspension was plated on Mueller-Hinton agar media with a swab, where the antimicrobial disks were deposited. The inhibition zones were read after incubation at 37°C for 16-18h.
Isolates that showed resistance to up to two antimicrobials of the same class were classified as non-MDR (non-multidrug-resistant); resistance to up to three antimicrobials of different courses, as MDR (Multidrug-resistant); resistance to at least one antimicrobial in all classes – except ≤ 2, as XDR (Extensively drug-resistant); and resistance to all antimicrobials tested, were classified as PDR (Pandrug-resistant).
3. Results and Discussion
This study evaluated the microbial quality of the bivalves Nodipecten nodosus and water in mariculture farms in the coves of Ilha Grande Bay, Angra dos Reis, RJ (Figure 1), focusing on the prevalence of Pseudomonas spp. and Vibrio spp. over a year.
From the collected samples (55 L of water and 275 scallops), 216 species (100%) were isolated, all identified by MALDI-TOF MS. The prevalence was of the genus Pseudomonas spp. (n = 62; 29%), followed by the genus Vibrio spp. (n = 52; 24%), as shown in Figure 3.
3.1. Determination of the Antimicrobial Susceptibility Profile
AMR poses a serious global threat to public health due to the increasing spread of new resistance mechanisms in several regions [48]. The scenario is worrying, with estimates of around 700,000 annual deaths from drug-resistant infections and projections that this number could exceed 10 million deaths per year by 2050 [49]
In this study, 114 isolates were selected to evaluate antimicrobial susceptibility, of which 62 were Pseudomonas spp. (52 Pseudomonas putida and 10 Pseudomonas monteilii) and 52 were Vibrio spp. (37 V. alginolyticus, 11 V. fluvialis and 4 V. harveyi). These isolates were obtained from collected scallops and analyzed water samples.
In recent decades, Pseudomonas aeruginosa has emerged as one of the top three critical amd priority pathogens contributing significantly to AMR [50]. Infections caused by P. aeruginosa are often fatal and difficult to treat due to their intrinsic resistance to several antimicrobial agents.
A new-found study investigated the presence of pharmaceutical compounds in samples collected from a wastewater treatment plant in India. The plant received effluent from approximately 90 regional bulk drug manufacturers. The results revealed that ciprofloxacin concentrations in the samples were significantly higher than those typically found in the blood of patients receiving treatment with this antimicrobial. This finding highlights the concern about potential environmental contamination by pharmaceutical residues and its implications for public health and the environment.
Bacteria recovered from water collected from the treatment plant were tested for resistance to 39 antibiotics. The results showed that approximately 30% of the bacteria were resistant to 29 and 32 antibiotics, while approximately 20% were resistant to between 33 and 36 antibiotics [50]. This significant level of resistance highlights the concern about environmental contamination by pharmaceutical residues since bacteria present in soil can transfer resistance genes to clinically relevant pathogens [51]. The diversity in resistance observed reflects the complexity of the spread of multidrug-resistant pathogens, often observed in hospital settings. It may be attributed to factors such as the bacteria's natural habitats and the competition in these niches.
Furthermore, certain bacterial strains demonstrate a remarkable ability to maintain their MDR phenotype and spread through the community. This persistence is likely facilitated by additional genetic content that minimizes the relative cost of expressing antibacterial resistance genes. The community spread of MDR bacteria poses a significant threat to public health and demands an urgent and effective approach to its control and mitigation.
Bacterial resistance results from the ability of these populations to adapt to aggression over time, combined with the inappropriate use of antimicrobials. Such conditions can lead to a significant increase in the severity of infections caused by several bacterial species. Pettibone et al. [52] and Patha and Gopal [53] raised potential public health concerns about co-resistance to metals and antibiotics. These authors observed that bacterial isolates obtained from fish tissues commonly consumed by humans exhibited simultaneous resistance to several metals and antibiotics. This natural process is created to defend bacteria that produce antibiotics from their compounds and improve their survival ability in highly competitive microbial environments [54].
Accordingly, monitoring bivalve mollusks intended for consumption by the Brazilian population becomes indispensable and highly relevant. This monitoring is essential to guarantee food safety, covering both mariculture farms, where the mollusks are cultivated, and natural environments, where these animals are collected in an exploratory manner. Effective control on both fronts is essential to prevent contamination risks by pathogens and toxins, ensuring food quality and consumer health.
3.2. Antimicrobial Susceptibility: Pseudomonas spp.
Resistance to anti-pseudomonas antibiotics has intensified in recent years due to the emergence of a considerably high frequency of antimicrobial resistance. Most non-fermenting Gram-negative bacteria of the genus Pseudomonas spp. are saprophytes; however, they can contaminate human clinical samples. It has been shown that these microorganisms can also act as pathogens under specific conditions.
In this study, the prevalence of Pseudomonas spp. was identified in 62 isolates using the MALDI-TOF MS methodology, of which 84% were classified as Pseudomonas putida and 16% as Pseudomonas monteilii. Among these species, a higher percentage of resistance to several antibiotics was observed, respectively: ciprofloxacin (42%, 10%), cefepime (37%, 10%), aztreonam (35%, 40%), ceftazidime (6%, 0%), amikacin (4%, 0%), piperacillin/tazobactam (2%, 0%), meropenem (2%, 0%) and imipenem (2%, 0%) (Figure 4).
Pseudomonas aeruginosa is a common Gram-negative opportunistic pathogen responsible for several hospital-acquired infections, often associated with high rates of antimicrobial resistance. This microorganism can rapidly develop multiple (MDR), extensive (XDR), and even total (PDR) resistance in a short period of time [48]. Due to its versatile metabolism and ability to survive adverse environmental conditions and physicochemical stress [55], it has aroused great interest as a host cell for synthetic biology and metabolic engineering.
Pseudomonas putida has considerable potential for biotechnological applications in agriculture, biocatalysis, bioremediation, and bioplastics production [56]. Among Pseudomonas species, intrinsic resistance to antimicrobials is largely attributed to mechanisms such as low membrane permeability, synthesis of β-lactamases, and efflux systems. However, due to remarkable genetic plasticity, members of the genus Pseudomonas spp. have demonstrated the ability to acquire virtually all known mechanisms of antimicrobial resistance [57].
According to Mohammed et al. [48], the results of this study are consistent with research carried out in Iran, Sudan, Egypt, and South Africa that analyzed 289 clinical isolates of Pseudomonas aeruginosa for susceptibility to 14 antimicrobial agents belonging to 8 categories. A high prevalence of resistance was observed, especially to ticarcillin-clavulanic acid (89.3%), meropenem (50.9%), and fosfomycin (37.7%). Resistance to antipseudomonal antimicrobials ranged from 37.7% to 14.5%, with the highest rates observed for cefepime (37.7%), imipenem (37.4%), aztreonam (25.3%), levofloxacin (23.5%), piperacillin-tazobactam (22.1%), and tobramycin (14.5%). Global resistance to these drugs poses a significant therapeutic challenge in treating P. aeruginosa infections. However, polymyxins and aminoglycosides have shown greater efficacy, with a susceptibility of 90.3% colistin, 86.5% amikacin, and 82.4% tobramycin.
In the study presented, the prevalence of Pseudomonas spp. was identified in 62 isolates using the MALDI-TOF MS methodology, of which 84% were classified as Pseudomonas putida and 16% as P. monteilii. Among these species, a higher percentage of resistance to several antibiotics was observed, respectively: ciprofloxacin (42%, 10%), cefepime (37%, 10%), aztreonam (35%, 40%), ceftazidime (6%, 0%), amikacin (4%, 0%), piperacillin/ tazobactam (2%, 0%), meropenem (2%, 0%) and imipenem (2%, 0%). P. putida, found at all collection points, is a saprophytic bacterium widely in the rhizosphere.
The definition of nom-MDR, MDR, XDR, and PDR for P. monteilli and P. putida was established based on a standardized international document [58], following the recommendations of CLSI and EUCAST. However, isolates of P. monteilli and P. putida classified as MDR demonstrated resistance to at least one agent in three antimicrobial categories. Those identified as XDR exhibited resistance to at least one agent in six categories. Isolates that did not show susceptibility to all commercially available antimicrobial agents were classified as PDR [58].
The nom-MDR profile was observed in isolates, in this study, of Pseudomonas monteilli and Pseudomonas putida from the three points: Gipóia Island Cove, Bananal Island Cove, and Jacuecanga Beach Cove. The isolates of P. putida presented the highest percentage of resistance to ciprofloxacin (42%, 22/52) – antipseudomonal fluoroquinolones, followed by cefepime (37%, 19/52), aztreonam (35%, 18/52) ceftazidime (6%, 3/52), amikacin, aminoglycosides (4%, 2/52) and piperacillin/tazobactam and imipenem – antipseudomonal carbapenems (2%, 1/52), antipseudomonal penicillin + β-lactamase inhibitors. Most P. putida isolates were sensitive to meropenem and antipseudomonal carbapenems. Meanwhile, P. monteilli isolates showed resistance only to aztreonam (40%, 4/10), followed by cefepime and ciprofloxacin (10%, 1/10), being sensitive to the other antimicrobials evaluated.
MDR bacteria are recognized as one of the most serious public health problems today [59]. The Infectious Diseases Society of America considers AMR one of the greatest threats to human health globally. The increase in MDR bacteria represents a critical danger for several reasons. P. aeruginosa is known for its multidrug resistance and resistance to many types of antibiotics [60,61]; that is, given its ability to develop resistance to several antimicrobials, it is one of the main causes of hospital infections, being associated with diseases of respiratory and urinary tracts, surgical wounds and bloodstream infections [62]. Among the main carbapenems genes found in this species are blaSPM-1, blaKPC-1, blaNDM-1, and blaVIM-1. Until 2017, the blaSPM-1 gene was one of the only carbapenems genes detected in Brazil [63].
However, true community-acquired infections with multidrug-resistant strains of Pseudomonas aeruginosa are still relatively rare. In a study of sixty patients with community-acquired bloodstream infection with P. aeruginosa, 100% of the isolates were susceptible to meropenem, and 95% were susceptible to piperacillin/tazobactam and ceftazidime. Furthermore, in a specific case reported in Turkey, a young man with no healthcare exposure presented with an abscess caused by a strain of P. aeruginosa that was susceptible only to imipenem, amikacin, and colistin [64].
Therefore, one of the most pressing public health challenges is to prevent the increasing spread of MDR bacteria in the community. Unfortunately, regional and national data on antibiotic susceptibility is often limited. Furthermore, epidemiological data are limited when available, and a clear determination of isolates associated with community transmission is needed. However, clinical cases are increasingly visible, attracting the attention of policymakers, while subclinical spread is already occurring [59].
An effective precautionary strategy must take a multifaceted approach and involve all stakeholders. In addition to clinical stewardship practices and responsible use of antibiotics in humans, removing antibiotics from the food chain is essential. It is also important to limit the number of xenobiotics, such as quaternary ammonium compounds, released into the environment [65]. Another crucial step in limiting bacterial exposure to antibiotics is properly treating contaminated wastewater, especially from pharmaceutical plants and medical facilities.
3.3. Susceptibilidade aos Antimicrobianos: Vibrio spp.
Using the MALDI-TOF MS technique, 52 isolates of Vibrio spp. were identified from samples collected at Gipóia Island Cove, Bananal Island Cove, and Jacuecanga Beach Cove (Brazil). These included V. harveyi (8%), V. fluvialis (21%), and V. alginolyticus (71%). All identified species are of clinical significance and associated with opportunistic infections in humans, primarily through exposure to contaminated water or consumption of contaminated seafood, such as Nodipecten nodosus [66]. Among these pathogens, V. alginolyticus was notably prevalent; its contamination is linked to considerable morbidity and, in severe cases, fatal outcomes [66,67,68]. Additionally, V. fluvialis was identified, a pathogen known to cause symptoms such as bloody diarrhea and, in immunosuppressed individuals, skin wound infections that can lead to primary septicemia [69].
Previous research [70] explored the occurrence and distribution of Vibrio spp. in aquatic environments to evaluate the risks of human exposure, identifying ten pathogenic species, including Vibrio cholerae O1, in Georgian waters over a two-year period. Bacteria of the Vibrio genus are associated with cholera, a disease caused by V. cholerae, which has been responsible for seven pandemics throughout history. Cholera remains a significant public health concern today, particularly in developing nations, with an estimated 4 million cases annually and an average of 21,000 to 143,000 deaths each year [71]
Vibrio spp. is commonly found in natural aquatic environments and can pose a threat to human health as a foodborne disease agent. Canellas [72] studied its presence at three locations in Guanabara Bay (Rio de Janeiro, Brazil). Hackbusch et al. [73] examined three pathogenic Vibrio species (V. parahaemolyticus, V. vulnificus, and V. cholerae) over a 14-month period in the North Sea (Europe). Their findings suggest potential seasonal trends in the occurrence of virulence genes, emphasizing the value of long-term research to better understand bacterial behavior in the environment and associated risks to human health. Nevertheless, the impact of this genus on public health remains inadequately understood [73].
The epidemiology of Vibrio alginolyticus infection has revealed a significant increase in incidence and severity over the past ten years [74]. As an opportunistic pathogen, V. alginolyticus has caused major economic losses to mariculture [75] and contributed to the overall increase in vibriosis cases. Although V. alginolyticus is the second most common Vibrio species worldwide, other species have also increased considerably [76]. In southern China, V. alginolyticus is the third most common Vibrio type isolated from diseased marine fish, after V. harveyi and V. vulnificus [77].
In this study, the isolates of V. alginolyticus and V. fluvialis showed high percentages of resistance to tetracycline (70% and 82%, respectively), followed by piperacillin/tazobactam, where V. harveyi is also found (54%, 91%, and 25%). Resistance to ciprofloxacin was 41%, 73%, and 75%, and to ceftazidime, 32%, 36%, and 0%. V. alginolyticus isolates showed the highest percentage of resistance to tetracycline (70%, 26/37), followed by piperacillin/tazobactam (54%, 20/37), ciprofloxacin (41%, 15/37), azithromycin and ceftazidime (32%, 12/37), meropenem (24%, 9/37) and sulfamethoxazole/trimethoprim (11%, 4/37). V. fluvialis isolates showed predominant resistance to piperacillin/tazobactam (91%, 10/11), followed by tetracycline (82%, 9/11), ciprofloxacin (73%, 8/11), meropenem (64%, 7/11), ceftazidime (36%, 4/11), azithromycin and sulfamethoxazole/trimethoprim (27%, 3/11). In turn, V. harveyi isolates showed resistance only to ciprofloxacin (75%, 3/4), piperacillin/tazobactam, and azithromycin (25%, 1/4) (Figure 5).
The presence of antimicrobial-resistant organisms was observed at all collection points evaluated in the study, indicating that these microorganisms had already arrived at the sites with resistance genes. These results highlight the importance of sanitation and the urgent need to implement effective wastewater management strategies to prevent the spread of MDR bacteria in the Bananal, Gipóia, and Náutica coves in Angra dos Reis.
The presence of Vibrio spp. Resistant to third-generation cephalosporins is frequently reported in the literature and is usually associated with species isolated from seafood. Lee et al. [78] observed the presence of Vibrio parahaemolyticus in fish sold in markets and fairs in Malaysia, with resistance rates of 52% and 28% to cefotaxime and ceftazidime, respectively. In addition, 2.4% of the isolates presented the trh gene (thermostable hemolysin gene) related to direct hemolysin, indicating its pathogenic potential.
Another study performed in South Korea reported that V. parahaemolyticus species isolated from fish farms showed resistance rates of 5.7% and 8.6% to cefotaxime and ceftazidime, respectively. More than 90% of these isolates' genes encoded pathogenic islands [79]. Some authors suggest that resistance to third-generation cephalosporins in Vibrio spp. is prevalent in Asia due to the extensive use of antimicrobials in aquaculture, a particularly developed sector in this region [80,81].
Antimicrobials in the fluoroquinolone class are among the most prescribed worldwide [82]. Other studies have also reported high rates of resistance to amino and carboxypenicillins in certain species, notably Vibrio parahaemolyticus and Vibrio cholerae [83].
Studies conducted with water samples have also reported high susceptibility rates to ciprofloxacin in several Vibrio species [84,85]. However, there is growing concern about the occurrence of Vibrio spp. Resistant to this class of antimicrobials, especially in aquaculture systems. A recent study demonstrated that biofertilizers in aquatic environments can significantly increase the resistance rate of V. parahaemolyticus strains to fluoroquinolones through the horizontal transfer of resistance genes [86].
Researchers from China, such as Yu et al. [87] reported that Vibrio alginolyticus can trigger epidemic outbreaks in fish, shrimp, mollusks, and other marine organisms, causing serious economic losses to the aquaculture industry, affecting species such as groupers, shrimps, and oysters [88,89,90,91].
Epidemic diseases caused by Vibrio alginolyticus are most likely when water temperatures range from 25 to 35°C. Previous studies have reported V. alginolyticus infections in Europe, Asia, South America, and North America [92,93,94]. Ingestion of contaminated raw seafood or exposure of wounds to warm saltwater containing V. alginolyticus has been associated with bacterial infections [94].
3.4. Assessment of Antimicrobial Susceptibility Profile
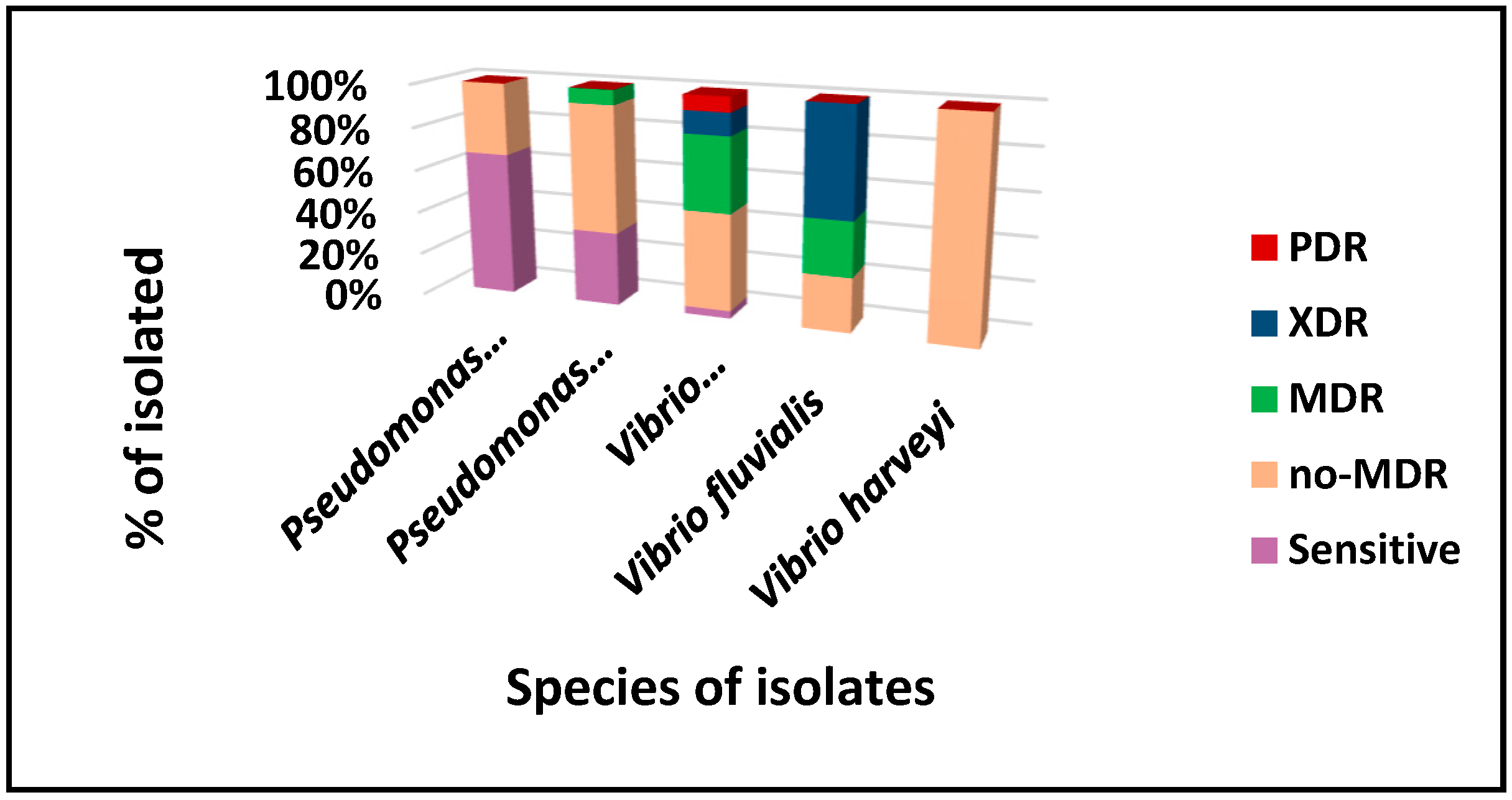
In this study, the three species of the genus Vibrio spp. and the two species of Pseudomonas spp. MDR, PDR, XDR, and non-MDR resistance profiles (Figure 6) are shown, which is concerning considering their medical importance. These isolates were obtained from samples collected in Bananal and Gipóia coves, in raw water within the farms, as well as from ultraviolet-treated water where the scallops are kept, and from seawater in Jacuecanga, an area frequently used for recreation and fishing. These data raise concerns about the public health risks for individuals exposed to these waters.
The results highlight the importance of sanitation and the urgent need to implement effective strategies for wastewater management. This is crucial to prevent the spread of bacteria with MDR, PDR, XDR, and non-MDR resistance profiles in the areas mentioned in Angra dos Reis.
Despite presenting a resistance profile to non-MDR antimicrobials, the strains P. putida, P. monteilli, Vibrio alginolyticus, V. fluvialis, and V. harveyi presented resistance to cefepime, ceftazidime and ciprofloxacin. The MDR profile was observed in P. putida, V. alginolyticus, and V. fluvialis isolates. It is also worth mentioning that the XRD profile was only observed in isolates of V. fluvialis and V. alginolyticus, and the PDR profile was only observed in V. alginolyticus.
4. Conclusions
The present study evaluated the occurrence, distribution, and microbial diversity of the gill arch tissue of N. nodosus and marine culture waters, with a prevalence of Pseudomonas spp., and Vibrio spp. over one year at three different culture sites in BIG, Angra dos Reis, Rio de Janeiro, Brazil.
Vibrio alginolyticus was observed at all collection points analyzed, constituting a threat to aquatic animals and human health with high percentages of resistance to antimicrobials and potentially pathogenic. The marine environment can collaborate to disseminate resistance genes in Vibrio spp. Species; aquatic monitoring of the analyzed regions is relevant to estimating possible risks to public health.
The species analyzed can be assessed for the presence of genes encoding virulence factors, antimicrobial resistance genes, and heavy metals, as well as for detecting other genetic elements that may suggest the transmissibility of these phenotypes, such as plasmids and integrons. In addition, the capacity to produce industrially relevant enzymes and assess biofilm formation are pertinent characteristics that could be explored in these species.
The study of the genus Vibrio spp. Offers great potential and should be beneficial, especially when dealing with bacteria isolated from an ecosystem with great ecological, economic, social, and clinical value on the BIG, Angra dos Reis, Rio de Janeiro. Thus, microbiological control of mollusks produced in different areas is a strategic and crucial condition for health surveillance, as it can prevent waterborne diseases in communities close to breeding sites.
Author Contributions
Conceptualization, A.L.S., M.B.M.C., and K.C.B.F.; methodology, A.L.S., M.B.M., G.S.L.C. and K.C.B.F.; formal analysis, R.R., J.L.Z.; writing—original draft preparation, A.L.S., M.B.M. and K.C.B.F; writing—review and editing, S.G.D-S.; supervision, M.B.M., K.C.B.F.; project administration, A.L.S., M.B.M.; funding acquisition, S.G.D-S. All authors have read and agreed to the published version of the manuscript.
Funding
S.G.D-S received fund from Carlos Chagas Filho Foundation of Research Support of the State of Rio de Janeiro (FAPERJ #010.101.029/2018) and the Brazilian Council for Scientific Research (CNPq #301744/2019-0).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data presented in this study are available upon request from the corresponding author.
Acknowledgments
We extend our sincere thanks to the team (especially, R.R.E. Silva. and J.L. Zaganelli) at the Ilha Grande Bay Eco-Development Institute for their assistance and partnership with the reference laboratory of Microorganisms, from the National Institute for Quality Control in Health (INCQS), Oswaldo Cruz Foundation, Rio de Janeiro, Brazil. We also acknowledge Vilma dos Santos for her significant laboratory assistance, for which we are truly grateful.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Linnaeus C (1758) Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Editio decima, reformata [10th revised edition], Laurentius Salvius: Holmiae 1758, 824. [CrossRef]
- Rupp. G.S.; Parsons, G.J. Aquaculture of the Scallop Nodipecten nodosus in Brazil. Develop Aquacult Fisheries Sci 2016, 40, 999–1017. [Google Scholar] [CrossRef]
- Thompson C, Bacha L, Paz PHC, de Assis Passos Oliveira M, Oliveira BCV, Omachi C, Chueke C, de Lima Hilário M, Lima M, Leomil L, et al., Collapse of scallop Nodipecten nodosus production in the tropical Southeast Brazil as a possible consequence of global warming and water pollution. Sci Total Environ 2023, 904, 166873. 904. [CrossRef]
- de Abreu Corrêa, A.; Huaman, M.E.D.; Siciliano, G.M.; Silva, R.R.E.; Zaganelli, J.L.; Pinto, A.M.V.; Dos Santos, A.L.; Vieira, C.B. First investigation of Ostreid herpesvirus-1 and human enteric viruses in a major scallop production area in Brazil. Environ Monit Assess 2024, 196, 1186. [Google Scholar] [CrossRef] [PubMed]
- Beeby, A. What do sentinels stand for? Environ Pollut. 2001, 112, 285–298. [Google Scholar] [CrossRef]
- Zhu, Z.; Qu, P.; Fu, F.; Tennenbaum, N.; Tatters, A. O.; Hutchins, D.A. Understanding the blob bloom: warming increases toxicity and abundance of the harmful bloom diatom Pseudo-nitzschia in California coastal waters. Harmful Algae 2017, 67, 36–43. [Google Scholar] [CrossRef]
- Blanco, J.; Moroño, Á.; Arévalo, F.; Correa, J.; Salgado, C.; Rossignoli, A. E.; Lamas, J. P. Twenty-five years of domoic acid monitoring in Galicia (NW Spain): Spatial, temporal and interspecific variations. Toxins 2021, 13, 756. [Google Scholar] [CrossRef]
- Scanes, E.; Byrne, M. Warming and hypoxia threaten a valuable scallop fishery: a warning for commercial bivalve ventures in climate change hotspots. Glob Chang Biol 2023, 29, 2043–2045. [Google Scholar] [CrossRef]
- Tomasetti, S.J.; Hallinan, B.D.; Tettelbach, S.T.; Volkenborn, N.; Doherty, O.W.; Allam, B.; Gobler, C.J. Warming and hypoxia reduce the performance and survival of northern bay scallops (Argopecten irradians irradians) amid a fishery collapse. Glob Chang Biol 2023, 9, 2092–2107. [Google Scholar] [CrossRef]
- Lattos, A.; Chaligiannnis, I.; Papadopoulos, D.; Giantsis, I. A.; Petridou, E.I.; et al. How safe to eat are raw bivalves? Host pathogenic and public health concern microbes within mussels, oysters, and clams in Greek markets. Foods 2021, 10, 279. [Google Scholar] [CrossRef]
- Sallenave-Namont, C.; Pouchus, Y. F.; Du Pont, R. T.; Lassus, P.; Verbist, J. F. Toxigenic saprophytic fungi in marine shellfish farming areas. Mycopathologia 2000, 149, 21–25. [Google Scholar] [CrossRef]
- Potasman, I.; Paz, A.; Odeh, M. Infectious outbreaks associated with bivalve shellfish consumption: a worldwide perspective. Clin Infect Dis 2002, 35, 921–928. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.S.; Possas, C. de A.; Viana, C.M.; Rodrigues, D. dos P. Vibrio spp. Isolated from fresh and pre-cooked mussels (Perna perna) from the Experimental Cultivation Station, Rio de Janeiro, RJ, Brazil. Food Sci Technol 2007, 27, 387–390. [Google Scholar] [CrossRef]
- Leal, D.A.G; Franco, R.M.B. Bivalve mollusks intended for human consumption with vectors of pathogenic protozoa: Detection methodologies and control standards. Rev Panam Infectol 2018, 10, 48–57. [Google Scholar]
- Lévesque, B.; Barthe, C.; Dixon, B. R.; Parrington, L. J.; Martin, D.; Doidge, B. , Proulx, J. F.; Murphy, D. Microbiological quality of blue mussels (Mytilus edulis) in Nunavik, Quebec: a pilot study. Can J Microbiol 2010, 56, 968–977. [Google Scholar] [CrossRef]
- Santos, A.L. dos, Hauser-Davis, R.A., Santos, M.J.S., De-Simone, S.G. Potentially toxic filamentous fungi associated to the economically important Nodipecten nodosus (Linnaeus, 1758) scallop farmed in southeastern Rio de Janeiro, Brazil. Mar Pollut Bull 2017, 115, 75–79. [Google Scholar] [CrossRef]
- Mau, A.; JHA, R. Aquaculture of two commercially important mollusks (abalone and limpet): existing knowledge and future prospects. Rev Aquicul 2018, 10, 611–625. [Google Scholar] [CrossRef]
- Wijsman, J.W.M.; Troost, K.; Fang, J.; Roncarati, A. Global Production of Marine Bivalves. Trends and Challenges. In: Smaal, A.; Ferreira, J.; Grant, J.; Petersen, J.; Strand, Ø. (eds) Goods and Services of Marine Bivalves. Springer, Cham. 2019. [CrossRef]
- FAO. Food and Agriculture Organization of the United Nations. The State of World Fisheries and Aquaculture 2022: Towards Blue transformation. p. 236. Rome, 2022. Available online: https://openknowledge.fao.org/server/api/core/bitstreams/a2090042-8cda-4f35-9881-16f6302ce757/content.
- Garbossa, L.H.; Souza, R.V.; Campos, C.J.; Vanz, A.; Vianna, L.F.; Rupp, G.S. Thermotolerant coliform loadings to coastal areas of Santa Catarina (Brazil) evidence the effect of growing urbanization and insufficient provision of sewerage infrastructure. Environ Monit Assess 2017, 189, 27. [Google Scholar] [CrossRef]
- De Souza, R.V.; De Campos C., J.A.; Garbossa, L.H.P.; Vianna, L.F.N.; Seiffert, W.Q. Optimizing statistical models to predict faecal pollution in coastal areas based on geographic and meteorological parameters. Mar Pollut Bull 2018, 129, 284–292. [Google Scholar] [CrossRef]
- De Souza, R.V.; De Campos, C.J.A.; Garbossa, L.H.P.; Seiffert, W.Q. Developing, cross-validating and applying regression models to predict the concentrations of faecal indicator organisms in coastal waters under different environmental scenarios. Sci Total Environ 2018, 630, 20–31. [Google Scholar] [CrossRef]
- Santos, M.M.; Soares, H.K.S.S.; Souza, A.M.; Bezerra, N.P.C.; Cantanhede, S.P.D.; Souza Serra, I.M.R. Mapping of the world scientific production on bacterial and fungal microbiota in mollusks. Lat Am J Aquat Res 2023, 51, 268–281. [Google Scholar] [CrossRef]
- Lino, A.S.; Galvão, P.M.A.; Longo, R.T.L.; Azevedo-Silva, C.E; Dorneles, P.R.; Torres, J.P.M.; Malm, O. Metal bioaccumulation in consumed marine bivalves in Southeast Brazilian coast. J Trace Elem Med Biol, 2016, 34, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Coleman, S.; Kiffney, T.; Tanaka, KR.; Morse, D.; Brady, D.C. Meta-Analysis of growth and mortality rates of net cultured sea scallops across the North Atlantic. Aquaculture 2022, 546, 737392. [Google Scholar] [CrossRef]
- Butler AJ, Thomas MK, Pintar KD. Expert elicitation as a means to attribute 28 enteric pathogens to foodborne, waterborne, animal contact, and person-to-person transmission routes in Canada. Foodborne Pathog Dis 2015, 12, 335–344. [Google Scholar] [CrossRef]
- Santos, A.L.; Medeiros, J.V.F.; Grault, C.E.; Santos, M.J.S.; Souza, A.L.A.; Carvalho, R.W. The fungus Pestalotiopsis sp., isolated from Perna perna (Bivalvia: Mytilidae) cultured on marine farms in Southeastern Brazil and destined for human consumption. Marine Pollut Bull 2020, 153, 110976. [Google Scholar] [CrossRef] [PubMed]
- World Organization for Animal Health (WOAH). Manual of diagnostic tests for aquatic animals. (2021). Available online: https://www.oie.int/en/what-we-do/standards/codesand-manuals/aquatic-manual-online-access/.
- Garnier, M.; Labreuche, Y; Nicolas, J. L. Molecular and phenotypic characterization of Vibrio aestuarianus subsp. Francensis subsp nov, a pathogen of the oyster Crassostrea gigas. Syst Appl Microbiol 2008, 31, 358–365. [Google Scholar] [CrossRef]
- Ottaviani, D.; Parlani, C.; Citterio, B.; Masini, L.; Leoni, F.; Canonico, C.; Sabatini, L.; Bruscolini, F.; Pianetti, A. Putative virulence properties of Aeromonas strains isolated from food, environmental and clinical sources in Italy: a comparative study. Int J Food Microbiol 2011, 144, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Borzykh, O.G. , Zvereva, L.V. Comparison of fungal complexes of Japanese scallop Mizuhopecten yessoensis (Jay, 1856) from different areas in the Peter the Great Bay of the Sea of Japan. Microbiology 2014, 83, 684–689. [Google Scholar] [CrossRef]
- Kwan, T.N.; Bolch, C.J. Genetic diversity of culturable Vibrio in an Australian blue mussel Mytilus galloprovincialis hatchery. Dis Aquat Organ 2015, 116, 37–46. [Google Scholar] [CrossRef]
- Dos Santos, A.L.; de Medeiros, J.V.F.; Grault, C.E.; Santos, M.J.S.; Souza, A.L.A.; de Carvalho, R.W. The fungus Pestalotiopsis sp., isolated from Perna perna (Bivalvia:Mytilidae) cultured on marine farms in Southeastern Brazil and destined for human consumption. Mar Pollut Bull 2020, 153, 110976. [Google Scholar] [CrossRef]
- Silveira, C.S.D.; Sousa, O.V.D.; Evangelista-Barreto, N.S. Propagation of antimicrobial resistant Salmonella spp. in bivalve mollusks from estuary areas of Bahia, Brazil. Rev Caatinga 2016, 29, 450–457. [Google Scholar] [CrossRef]
- Eliseikina, M.G.; Beleneva, I.A.; Kukhlevsky, A.D.; Shamshurina, E.V. Identification and analysis of the biological activity of the new strain of Pseudoalteromonas piscicida isolated from the hemal fluid of the bivalve Modiolus kurilensis (F. R. Bernard, 1983). Arch Microbiol 2021, 203, 4461–4473. [Google Scholar] [CrossRef] [PubMed]
- Thompson, C.; Bacha, L.; Paz, P. H. C.; Oliveira, M. de A. P.; Oliveira, B. C. V.; Chueke, C. O. C.; Hilário, M. de L.; Lima, M.; Leomil, L.; Felix-Cordeiro, T.; et al., Collapse of scallop Nodipecten nodosus production in the tropical Southeast Brazil as a possible consequence of global warming and water pollution. Sci Total Environm 2023, 904, 166873. [CrossRef]
- Cai, S.-H.; Wu, Z.-H.; Jian, J.-C; Lu, Y.-S. Cloning and expression of gene encoding the thermostable direct hemolysin from Vibrio alginolyticus strain HY9901, the causative agent of vibriosis of crimson snapper (Lutjanus erythopterus). J Appl Microbiol 2007, 103, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Amézquita-López, B.A.; Soto-Beltrán, M.; Lee, B.G.; Yambao, J.C.; Quiñones, B. Isolation, genotyping and antimicrobial resistance of Shiga toxin-producing Escherichia coli. J Microbiol Immunol Infect 2018, 51, 425–434. [Google Scholar] [CrossRef]
- Instituto Brazileiro de Geografia e Estatística (IBGE), 2021. Favelas e Comunidades Urbanas: aglomerados subnormais. Available online: https://www.ibge.gov.br/geociencias/organizacao-do-territorio/tipologiasdo-territorio/15788-aglomerados-subnormais.html.
- World Organization for Animal Health (WOAH). Manual of diagnostic tests for aquatic animals. 2021. Available online: https://www.oie.int/en/what-we-do/standards/codesand-manuals/aquatic-manual-online-access/.
- Brazil, Ministério da Saúde, Microrganismos resistentes aos carbapenêmicos e sua distribuição no Brasil, 2015 a 2022. Secretária de Vigilância em Saúde e Ambiente. Boletim Epidemiológico 2024, 55. Availabre at: Boletim_epidemiologico_SVSA_2_2024.pdf.
- BraziL. Ministry of Agriculture, Livestock and Food Supply. Secretariat of Agricultural Defense (MAPA/DAS). Ordinance No. 171, of December 13, 2018. Informs about the intention to prohibit the use of antimicrobials for the purpose of additives to improve food performance and opens a period for manifestation. Official Gazette of the Federative Republic of Brazil. Brasília, 2018.
- Thompson C, Bacha L, Paz PHC, de Assis Passos Oliveira M, Oliveira BCV, Omachi C, Chueke C, de Lima Hilário M, Lima M, Leomil L, et al., Collapse of scallop Nodipecten nodosus production in the tropical Southeast Brazil as a possible consequence of global warming and water pollution. Sci Total Environ 2023, 904, 166873. [CrossRef]
- MAPA 2012. Brazil. Ministry of Fisheries and Aquaculture. Interministerial Normative Instruction MPA/MAPA No. 7, of May 8, 2012. Institutes the National Program for Hygienic-Sanitary Control of Bivalve Molluscs (PNCMB) establishes the Procedures for its implementation and contains other provisions. Official Gazette [of the] Federative Republic of Brazil. Brasília, 2012.
- APHA. Standard methods for the examination of water and wastewater, 22nd Ed. American Public Health Association: Washington, DC. 2012.
- AOAC. Official methods of analysis. 16th Edition, Association of Official Analytical Chemists: Washington, DC. 1995.
- EUCAST. The European Committee on Antimicrobial Susceptibility Testing. Breakpoint tables for interpretation of MICs and zone diameters. Version 10.0, 2020 (valid from 2020-01-01). Available online: http://www.eucast.org/fileadmin/src/media/DFs/EUCASTfiles/Breakpoint.tables/v_10.0_Breakpoint_Tables.pdf.
- Mohammed, S.E.; Hamid, O.M.; Ali, S.S.; Allam, M.; Elhussein, A.M. Prevalence of multidrug-resistant, extensively drug-resistant and pandrug-resistant Pseudomonas aeruginosa clinical isolates in Khartoum State, Sudan. Am J Infect Dis Microbiol 2023, 11, 1–7. [Google Scholar] [CrossRef]
- O’Neill, J. Tackling drug-resistant Infections globally: Final report and recommendations. Review on antimicrobial resistance. Welcome Trust and HM Government. Arch Pharm Practice 2016, 7, 110, Available at: https://www.scirp.org/reference/references papers? referenceid= 2618400.. [Google Scholar]
- WHO. Cholera. 2017. Available online: https://www.who.int/news-room/fact-sheets/detail/cholera.
- Marathe, N.P.; Regina, V.R.; Walujkar, S.A.; Charan, S.S.; Moore, E.R.; Larsson, D.G.; Shouche, Y.S. A treatment planreceiving wastewater from multiple bulk drug manufacturers is a reservoir for highly multidrug resistant integron-bearing bacteria. PLoS One 2013, 8, e77310. [Google Scholar] [CrossRef]
- Pettibone, G.W.; Mear, J.P.; Sampsell, B.M. Incidence of antibiotic and metal resistance and plasmid carriage in Aeromonas isolated from brown bullhead (ictalurus nebulosus). Lett Appl Microbiol 1996, 23, 234–240. [Google Scholar] [CrossRef]
- Pathak, S.P.; Gopal, K. Occurrence of antibiotic and metal resistance in bacteria from organs of river fish. Environ Res 2005, 98, 100–103. [Google Scholar] [CrossRef]
- Rolain, J.M. Food, and human gut as reservoirs of transferable antibiotic resistance encoding genes. Front Microbiol 2013, 4, 173. [Google Scholar] [CrossRef]
- Volke, D.C.; Calero, P.; Nikel, P. I. Pseudomonas putida . Trends Microbiol 2020, 28, 512–513. [Google Scholar] [CrossRef] [PubMed]
- Nelson, K.E.; Weinel, C.; Paulsen, I.T.; Dodson, R.J.; Hilbert, H.; Martins dos Santos, V.A.P.; Fouts, D. E.; Gill, S.R.; Pop, M.; Holmes, M.; et al. Complete genome sequence and comparative analysis of the metabolically versatile Pseudomonas putida KT2440. Environ Microbiol 2002, 4, 799-808. doi: 10.1046/ j.1462-2920.2002.00366.x. Erratum in: Environ Microbiol 2003, 5, 630. [CrossRef] [PubMed]
- Livermore, D. M. Multiple mechanisms of antimicrobial resistance in Pseudomonas aeruginosa: our worst nightmare? Clin Infect Dis 2002, 34, 634–640. [Google Scholar] [CrossRef]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. , Multidrug-resistant, extensively drug-resistant and pan drug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin Microbiol Infect 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Van Duin, D.; Paterson, D.L. Multidrug-resistant bacteria in the community: trends and lessons learned. Infect Dis Clin North Am 2016, 30, 377–390. [Google Scholar] [CrossRef]
- Rodríguez-Baño, J.; Alcalá, J.C.; Cisneros, J.M.; Grill, F.; Oliver, A.; Horcajada, J.P.; Tórtola, T.; Mirelis, B.; Navarro, G.; Cuenca, M.; et al. , Community infections caused by extended-spectrum beta-lactamase-producing Escherichia coli. Arch Intern Med 2008, 168, 1897–1902. [Google Scholar] [CrossRef]
- Anderson, D.J.; Moehring, R.W.; Sloane, R.; Schmader, K.E.; Weber, D.J.; Fowlerm-JR, V.G.; Smathers, E.; Sexton, D.J. Bloodstream infections in community hospitals in the 21st century: a multicenter cohort study. PLoS One 2014, 9, e91713. [Google Scholar] [CrossRef]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.J.; Cheng, Z. Antibiotic resistance in Pseudomonas aeruginosa: mechanisms and alternative therapeutic strategies. Biotechnol Adv 2019, 37, 177–192. [Google Scholar] [CrossRef]
- Kalluf, K. O, Arend, L.N, Wuicik, T.E, Pilonetto, M, Tuon, F.F. Molecular epidemiology of SPM-1-producing Pseudomonas aeruginosa by rep-PCR in hospitals in Parana. Brazil. Infect Genet Evol 2017, 49, 130–133. [Google Scholar] [CrossRef]
- Ulug, M.; Gedik, E.; Girgin, S.; Celen, M. K.; Ayaz, C. Pyogenic liver abscess caused by community-acquired multidrug resistance Pseudomonas aeruginosa. Braz J Infect Dis 2010, 14, 218. [Google Scholar] [CrossRef]
- Hawkey, P.M.; Jones, A.M. The changing epidemiology of resistance. J Antimicrob Chemother 2009, 64, i3–i10. [Google Scholar] [CrossRef] [PubMed]
- Newton, A.; Kendall, M.; Vugia, D.J.; Henao, O.L.; Mahon, B.E. Increasing rates of vibriosis in the United States, 1996-2010: review of surveillance data from 2 systems. Clin Infect Dis 2012, 54s5, S391–395. [Google Scholar] [CrossRef] [PubMed]
- Morris-JR, J.G.; Black, R.E. Cholera and other vibrioses in the United States. New Engl J Med 1985, 312, 343–350. [Google Scholar] [CrossRef]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, RV.; Widdowson, M.A.; ROY, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States--major pathogens. Emerg Infect Dis 2011, 17, 7–15. [Google Scholar] [CrossRef]
- Okoh, A.I.; Igbinosa, E.O. Antibiotic susceptibility profiles of some Vibrio strains isolated from wastewater final effluents in a rural community of the Eastern Cape Province of South Africa. BMC Microbiol 2010, 10, 143. [Google Scholar] [CrossRef]
- Kokashvili, T.; Whitehouse, C.A.; Tskhvediani, A.; Grim, C. J.; Elbaidze, T.; Mitaishvili, N.; Janelidze, N.; Jaiani, E.; Jaley, B. J.; Lashhi, N.; et al. , Occurrence and diversity of clinically important Vibrio species in the aquatic environment of Georgia. Front Public Health 2015, 3, 232. [Google Scholar] [CrossRef]
- WHO. World Health Organization Global Strategy for Containment of Antimicrobial Resistance 2001. Scanes, E, Bryne, M., Warming and hypoxia threaten a valuable scallop fishery: a warning for commercial bivalve ventures in climate change hotspots. Glob. Chang. Biol. 2023. [CrossRef]
- Canellas, A.L.B. Isolation and characterization of bacteria of the genus Vibrio from the waters of Guanabara Bay, Rio de Janeiro, Brazil. Advisor: Marinella Silva Laport. 2020. 150f. Undergraduate Course Completion Work (Bachelor’s degree in Biological Sciences) – Paulo de Goés Institute of Microbiology, Federal University of Rio de Janeiro. Rio de Janeiro, 2020. Available online: https://pantheon.ufrj.br/bitstream/11422/17503/a/ALBCanellas.pdf.
- Hackbusch, S.; Wichels, A.; Gimenez, L.; Döpke, H.; Gerdts, G. Potentially human pathogenic Vibrio spp. in a coastal transect: Occurrence and multiple virulence factors. Sci Total Environ 2020, 707, 136113. [Google Scholar] [CrossRef]
- Mechri, B.; Monastiri, A.; Medhioub, A.; Medhioub, M.N.; Aouni, M. Molecular characterization and phylogenetic analysis of highly pathogenic vibrio alginolyticus strains isolated during mortality outbreaks in cultured ruditapes decussatus juvenile. Microb Pathog 2017, 111, 487–496. [Google Scholar] [CrossRef]
- Yu, Q.; Liu, M.; Xiao, H.; Wu, S.; Qin, X.; Ke, K.; Li, S.; Mi, H.; Shi, D.; Li, P. Development of novel aptamer-based enzyme-linked apta-sorbent assay (ELISA) for rapid detection of mariculture pathogen Vibrio alginolyticus. J Fish Dis 2019, 42, 1523–1529. [Google Scholar] [CrossRef]
- Zuo, Y.; Zhao, L.; Xu, X.; Zhang, J.; Zhang, J.; Yan, Q.; Huang, L. Mechanisms underlying the virulence regulation of new Vibrio alginolyticus ncRNA Vvrr1 with a comparative proteomic analysis. Emerg Microbes Infect 2019, 8, 1604–1618. [Google Scholar] [CrossRef]
- Deng, Y.; Xu, L.; Chen, H.; Liu, S.; Guo, Z.; Cheng, C.; Ma, H.; Feng, J. Prevalence, virulence genes, and antimicrobial resistance of Vibrio species isolated from diseased marine fish in South China. Sci Rep 2020, 10, 14329. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.H.; Mutabilib, N.S.A.; Law, J.W.F.; Wong, S.H.; Letchumanan, V. Discovery on antibiotic resistance patterns of Vibrio parahaemolyticus in Selangor reveals carbapenemase-producing Vibrio parahaemolyticus in marine and freshwater fish. Front Microbiol 2018, 25, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.H.; Kin, J.A.; Jeon, S.J.; Choi, S.S.; Kim, M.K.; YI, H.J.; CHO, S.J.; Kim, I.Y.; Chon, J.W.; Kim, D.H.; et al. , Prevalence, antibiotic-resistance, and virulence characteristics of Vibrio parahaemolyticus in restaurant fish tanks in Seoul, South Korea. Foodborne Pathog Dis 2020, 17, 209–214. [Google Scholar] [CrossRef]
- FAO (Food and Agriculture Organization of the United Nations). The State of World Fisheries and Aquaculture 2020: sustainability in action. p. 224. Rome, 2020. Available online: https://openknowledge.fao.org/server/API/core/bitstreams/170b89c1-7946-4f4d-914a-fc56e54769de/content.
- Loo, K.-Y.; Letchumanan, V.; Law, J.W.-F.; Pusparajah, P.; Goh, B.-H.; AB Mutalib, N.-S.; He, Y.-W.; Lee, L.-H. Incidence of antibiotic resistance in Vibrio spp. Rev Aquacult 2020, 12, 2590–2608. [Google Scholar] [CrossRef]
- Van Boeckel, T.P.; Gandra, S.; Ashok, A.; Caudron, Q.; Grenfell, B.T.; Levin, S.A.; Laxminarayan, R. Global antibiotic consumption 2000 to 2010: an analysis of national pharmaceutical sales data. Lancet Infect Dis 2014, 14, 742–750. [Google Scholar] [CrossRef]
- Lei, T.; Jiang, F.; He, M.; Zhang, J.; Zeng, H.; Chen, M.; Pang, R. U.; Wei, L.; Wang, J.; Ding, Y.; Wu, Q. Prevalence, virulence, antimicrobial resistance, and molecular characterization of fluoroquinolone resistance of Vibrio paraemolyticus from different types of food samples in China. Int J Food Microbiol 2020, 317, 108461. [Google Scholar] [CrossRef]
- Shah, S.Q.; Cabello, F.C.; L'Abée-Lund, T.M.; Tomova, A.; Godfrey, H.P.; Buschmann, A.H.; Sørum, H. Antimicrobial resistance and antimicrobial resistance genes in marine bacteria from salmon aquaculture and non-aquaculture sites. Environ Microbiol 2014, 16, 1310–1320. [Google Scholar] [CrossRef]
- Håkonsholm, F.; Lunestad, B.T.; Aguirre Sánchez, J.R.; Martinez-Urtaza, J.; Marathe, N.P.; Svanevik, C.S. Vibrios from the Norwegian marine environment: Characterization of associated antibiotic resistance and virulence genes. Microbiologyopen 2020, 9, e1093–2020. [Google Scholar] [CrossRef]
- Zhao, L.; HE, K.; Luo, J.; Sun, J.; Liao, L.; Tang, X.; Liu, Q.; Yang, S. Co-modulation of liver genes and intestinal microbiome of largemouth bass larvae (Micropterus salmoides) during weaning. Front Microbiol 2020, 11, 1332. [Google Scholar] [CrossRef]
- Yu, Y.; LI, H.; Wang, Y.; Zhang, Z.; Liao, M.; Rong, X.; LI, B.; Wang, C.; GE, J.; Zhang, X. Antibiotic resistance, virulence and genetic characteristics of Vibrio alginolyticus isolates from the aquatic environment in coastal mariculture areas in China. Mar Pollut Bull 2022, 185, 114219. [Google Scholar] [CrossRef]
- Liu, C.H.; Cheng, W.; Hsu, J.P.; Chen, J.C. Vibrio alginolyticus infection in the white shrimp Litopenaeus vannamei confirmed by polymerase chain reaction and 16S rDNA sequencing. Dis Aquat Org 2004, 61, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Mohamad, N.; Mohd Roseli, F.A.; Azmai, M.N.A.; Saad, M. Z.; MD Yasin, I.S.; Zuliply, N.A.; Nasruddin, N.S. Natural concurrent infection of Vibrio harveyi and V. alginolyticus in cultured hybrid groupers in Malaysia. J Aquat Anim Health 2019, 31, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, B.; Chen, G.; Jian, J.; Lu, Y.; Xu, Y.; Wu, Z. Transcriptome analysis of the pearl oyster (Pinctada fucata) hemocytes in response to Vibrio alginolyticus infection. Gene 2016, 575, 421–428. [Google Scholar] [CrossRef]
- Wang, J.; Ding, Q.; Yang, Q.; Fan, H.; Yu, G.; Liu, F.; Bello, B.K.; Zhang, X.; Zhang, T.; Dong, J.; et al. , Vibrio alginolyticus triggers inflammatory response in mouse peritoneal macrophages via activation of NLRP3 Inflammasome. Front Cell Infect Microbiol 2021, 11, 769777. [Google Scholar] [CrossRef]
- Hernández-Robles, M.F.; Álvarez-Contreras, A.K.; Juárez-García, P.; Natividad-Bonifacio, I.; Curiel-Quesada, E.; Vázquez-Salinas, C.; Quiñones-Ramírez, E. I. Virulence factors and antimicrobial resistance in environmental strains of Vibrio alginolyticus. Int Microbiol 2016, 19, 191–198. [Google Scholar] [CrossRef]
- Kang, C.H.; Shin, Y.; Jang, S.; Jung, Y.; So, J.S. Antimicrobial susceptibility of vibrio alginolyticus isolated from oysters in Korea. Environ Sci Pollut Res Int 2016, 23, 21106–21112. [Google Scholar] [CrossRef]
- Zavala-Norzagaray, A.A.; Aguirre, A.A.; Velazquez-Roman, J.; Flores-Villaseñor, H.; León-Sicairos, N.; Ley-Quiñonez, C.P.; Hernández-Díaz, L. de J.; Canizalez-Roman, A. Isolation, characterization, and antibiotic resistance of Vibrio spp. in sea turtles from Northwestern Mexico. Front Microbiol 2015, 6, 635. [Google Scholar] [CrossRef]
Figure 1.
– Study site and collection of water and scallop samples. Designed by Sergio Carvalho Moreira | Date: 03/31/2023. Source: Map of Brazil – IBGE, 2020. World map – ESRI database, 2022. The program used ArcGIS 10.8.
Figure 1.
– Study site and collection of water and scallop samples. Designed by Sergio Carvalho Moreira | Date: 03/31/2023. Source: Map of Brazil – IBGE, 2020. World map – ESRI database, 2022. The program used ArcGIS 10.8.
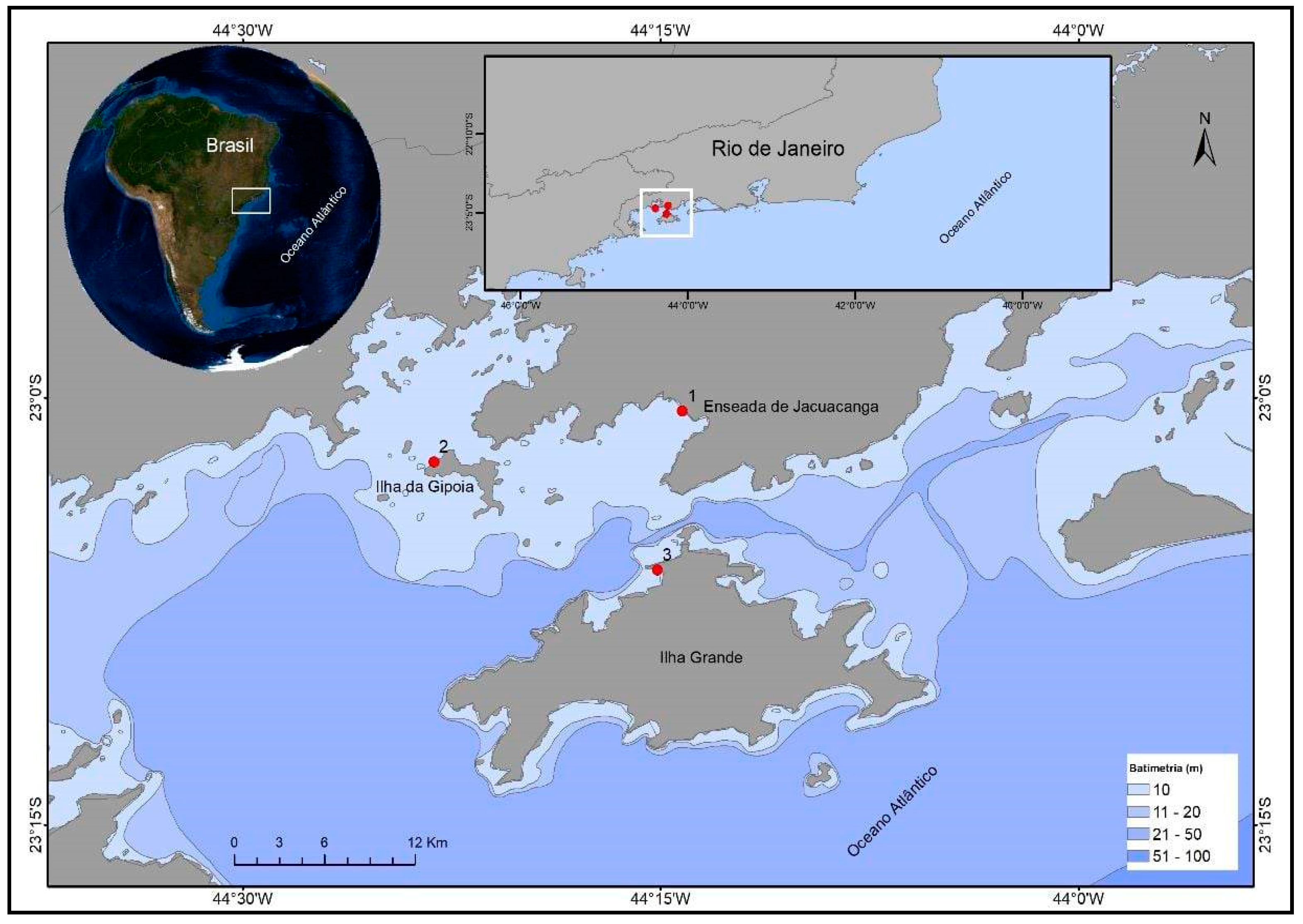
Figure 2.
– Flowchart established for sample collection, preparation, and processing.
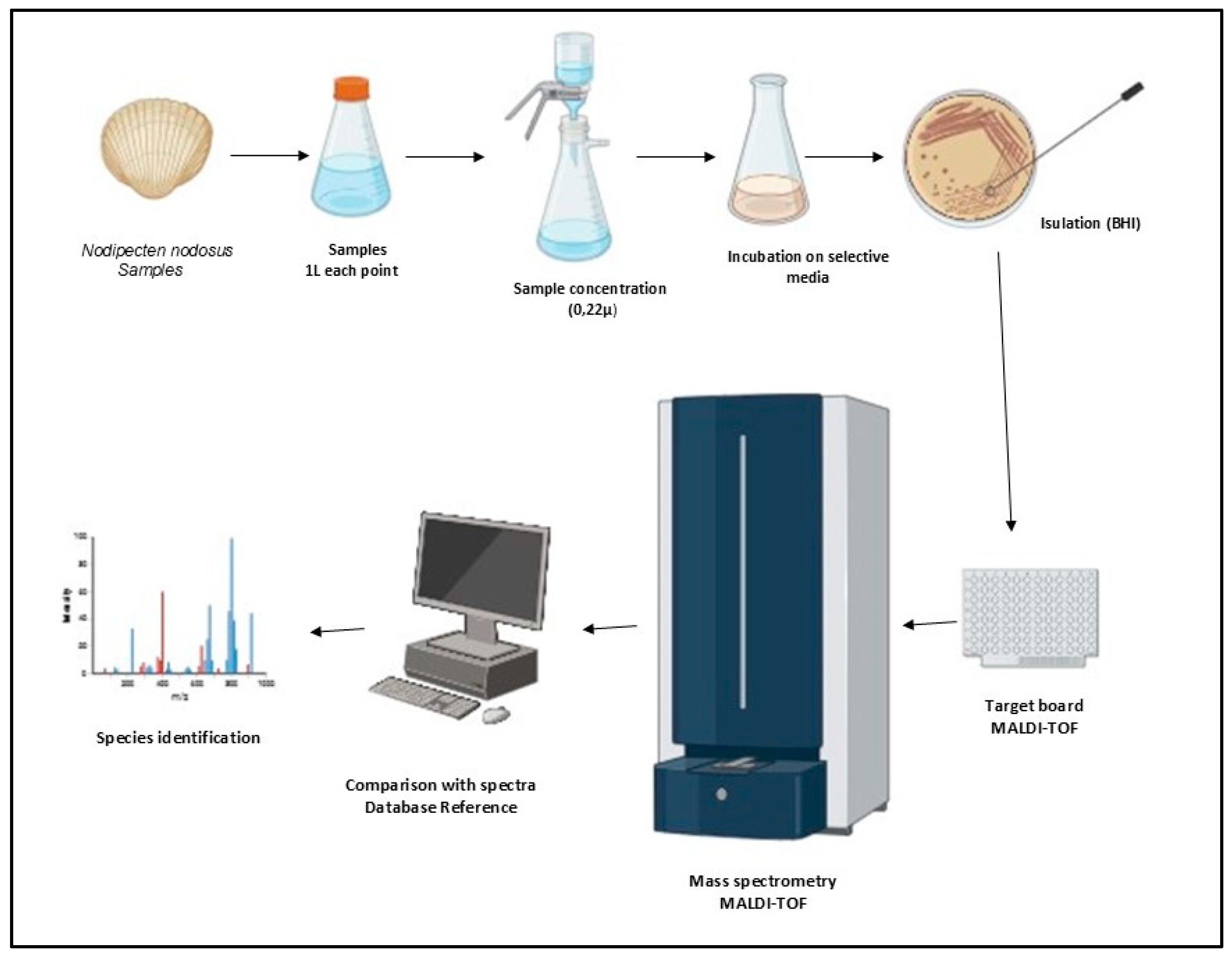
Figure 3.
Number of isolates about isolated species.
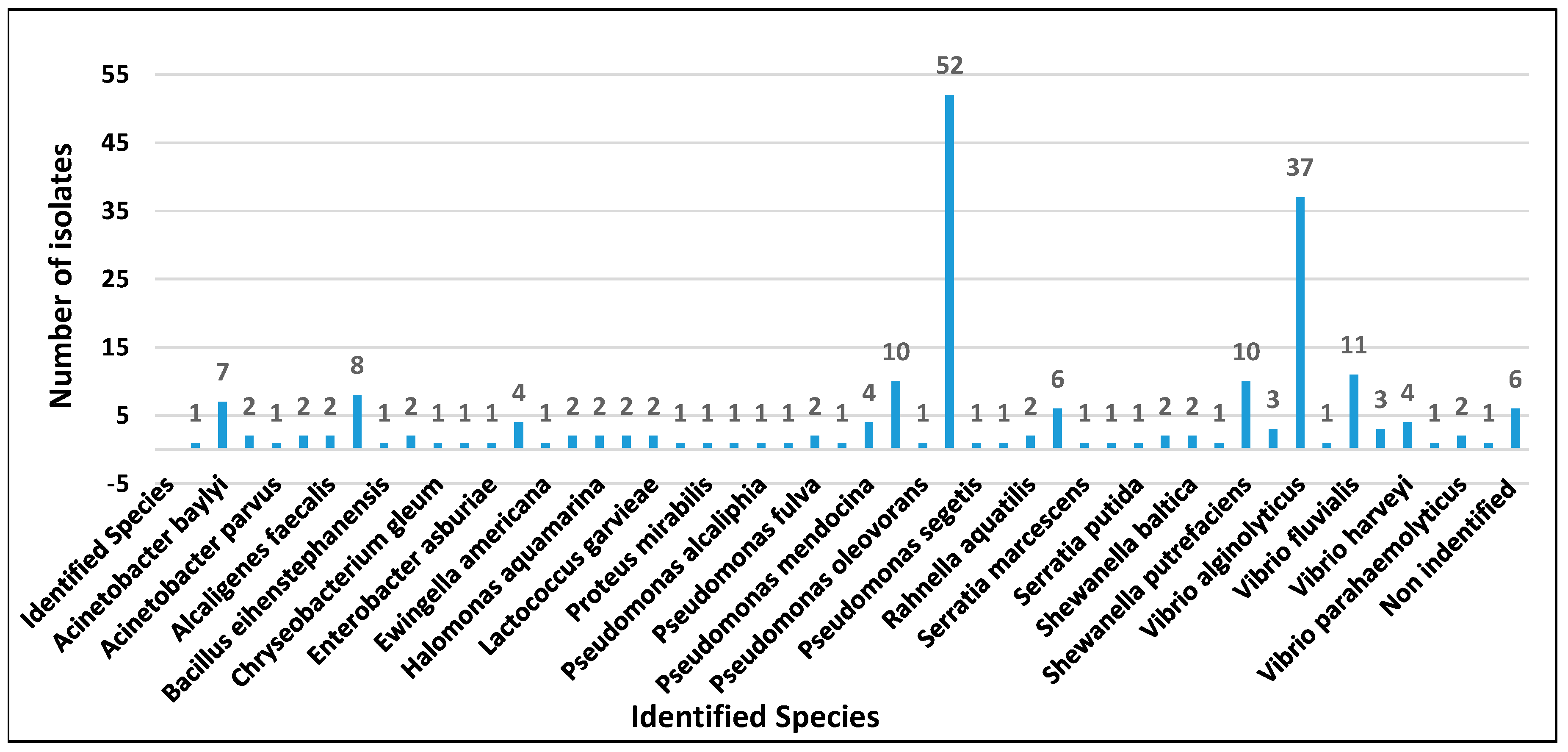
Figure 4.
Antimicrobial susceptibility profile of Pseudomonas putida, Pseudomonas monteilii isolates recovered from scallops and water.
Figure 4.
Antimicrobial susceptibility profile of Pseudomonas putida, Pseudomonas monteilii isolates recovered from scallops and water.
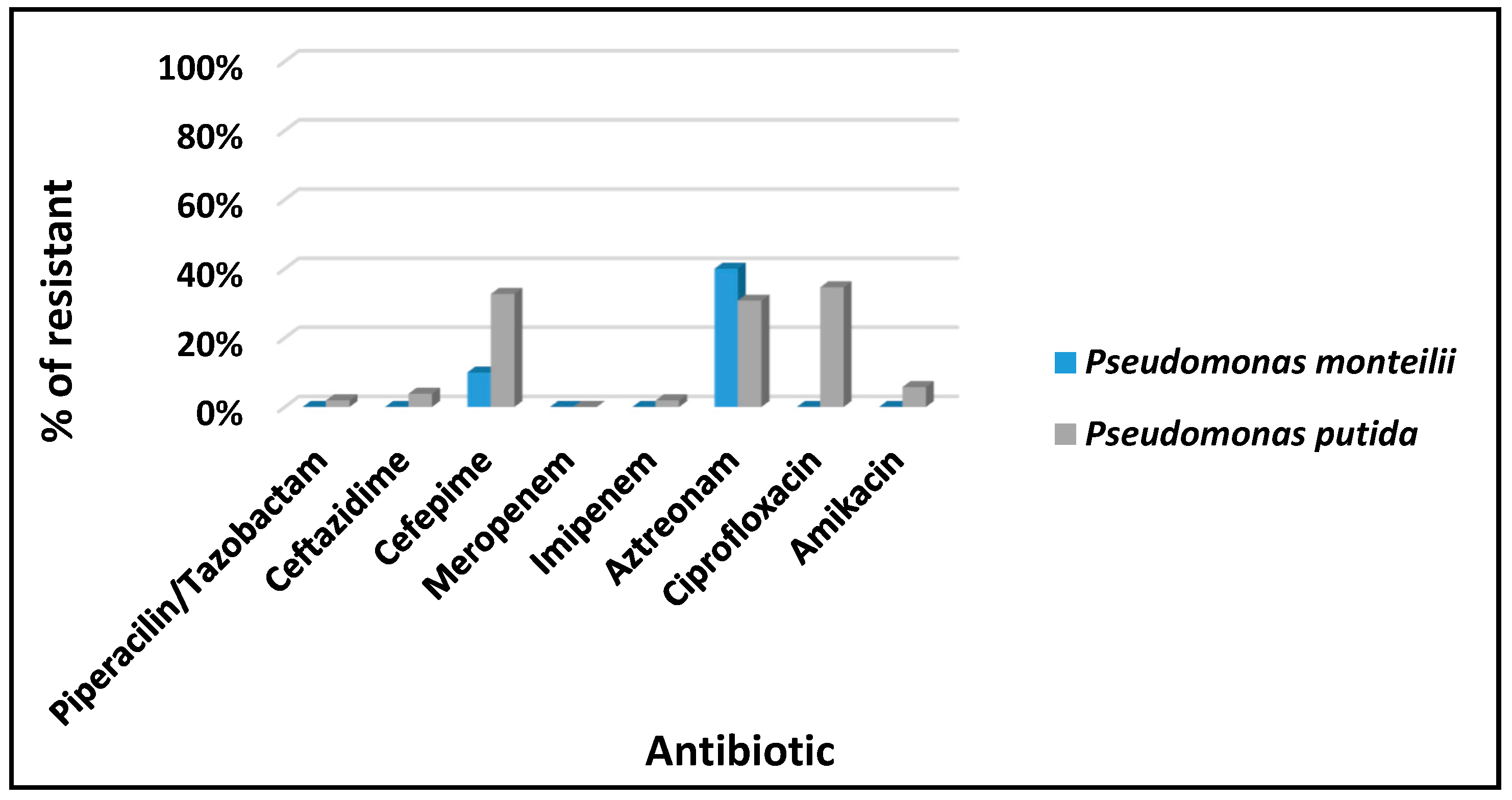
Figure 5.
Antimicrobial susceptibility profile of Vibrio alginolyticus, V. fluvialis, and V. harveyi isolates recovered from scallops and water.
Figure 5.
Antimicrobial susceptibility profile of Vibrio alginolyticus, V. fluvialis, and V. harveyi isolates recovered from scallops and water.
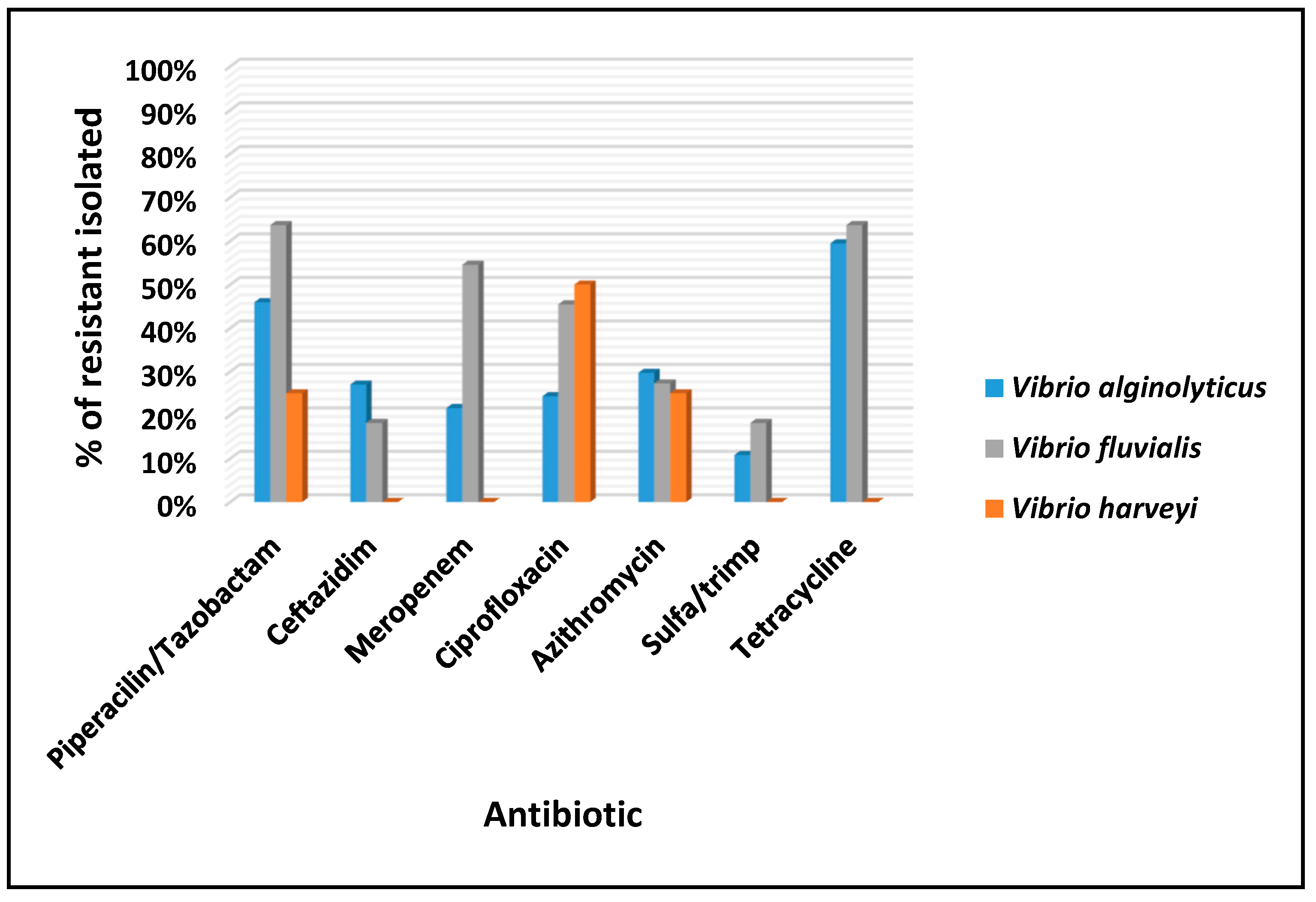
Figure 6.
Classification of antimicrobial susceptibility profiles concerning evaluated species.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



