
Submitted:
31 December 2024
Posted:
31 December 2024
You are already at the latest version
Abstract
Forensic entomology is concerned with the study of insects and other arthropods that assist in criminal investigations. Among them, necrophagous beetles (Coleoptera) play a crucial role as they are found in all stages of decomposition of corpses, which makes them valuable for forensic analysis regardless of the time period. Accurate species identification is crucial for the effective use of entomological evidence. Efficient, cost-effective and rapid methods are essential for this purpose. In the present study, we developed a novel assay that enables simple, inexpensive and rapid identification of necrophagous beetle species. Using two primer sets targeting the cytochrome c oxidase 1 (COI) gene and analysing the resulting SYBR Green I melting curves, we were able to identify fourteen beetle species: Aleochara curtula, Anoplotrupes stercorosus, Creophilus maxillosus, Dermestes undulatus, Hister unicolor, Margarinotus brunneus, Necrodes littoralis, Nicrophorus vespilloides, Ontholestes murinus, Oiceoptoma thoracicum, Philonthus cognatus, Saprinus planiusculus, Silpha tristis, and Thanatophilus rugosus. Data on the melting temperature of the amplicon can be obtained within 60–90 minutes after DNA extraction. The results contribute to the understanding of necrophagous Coleoptera in Poland and provide for the first time comprehensive data on necrophagous beetles in the vicinity of the Kampinos National Park.
Keywords:
forensic entomology
; Necrophagous Coleoptera
; species identification
; COI
; low-resolution melting real-time PCR
; Kampinos National Park
1. Introduction
Determining the time of death is a cornerstone of forensic investigations as it provides crucial insight into the circumstances of an incident. For fresh corpses, medical procedures such as measuring body temperature and assessing rigour mortis are reliable for estimating the postmortem interval (PMI) within the first 72 hours after death. Beyond this time frame, the accuracy of these methods decreases and entomological evidence becomes the most reliable approach for PMI estimation [1].
Insects are attracted to decaying remains within hours of death, with their colonisation and development providing a wealth of information for forensic analysis. However, the accuracy of insect-based PMI estimates depends on two key factors: the identification of the species present on the corpse and the temperature conditions in the environment. As ectothermic organisms, the growth and development rates of insects are temperature-dependent, a property that is utilised in the development method of PMI estimation. This method relies primarily on the immature stages of blowflies, particularly those of the family Calliphoridae, whose life cycles are well documented [1,2,3,4]. While Calliphoridae provide reliable PMI estimates for the early stages of decomposition, their utility is limited to the larval stage of their life cycle.
Forensic cases involving advanced decomposition or skeletal remains often require entomological evidence beyond that provided by blowflies. Necrophagous beetles (Coleoptera) are particularly valuable in such cases as they can be found in all stages of decomposition of corpses. Beetles offer two key forensic advantages: Certain genera, such as Thanatophilus and Necrodes, colonise remains within hours of death, and many species have a relatively slow rate of development, which increases their usefulness for PMI determination. For example, Thanatophilus species are widespread and often colonise corpses within 24 hours post mortem, making them suitable for PMI estimation [5]. Other beetle families, such as Dermestidae, Trogidae and Cleridae, are associated with later stages of decomposition or skeletal remains [6,7]. By incorporating beetle data, forensic entomologists can expand the window for PMI estimation and improve accuracy in cases of advanced decomposition.
Despite their forensic potential, the identification of necrophagous beetles - especially in the immature stage - poses a major challenge compared to blowflies. DNA barcoding has been shown to be effective in the identification of species at all stages of development [8]. However, conventional DNA barcoding methods are time-consuming, expensive and labour-intensive, making them less practical for large-scale forensic investigations.
Real-time PCR with melting curve analysis offers a promising alternative for species identification, allowing differentiation based on different melting temperatures of the PCR products. High-resolution melting PCR (HRM-PCR) provides very detailed data and is therefore suitable for the identification of various organisms, including bacteria, insects, fish and mammals, e.g., [9,10,11,12]. However, HRM-PCR requires specialised equipment, which limits its accessibility. Low-resolution melting PCR (LRM-PCR), on the other hand, has been successfully used in areas such as meat authentication, immunology, virology and the identification of pathogens in food and water [13,14,15] and can be performed with all real-time PCR devices.
With an area of 37,756 hectares, Kampinos National Park (KPN) is the second largest protected area in Poland. Its diverse habitats - including grasslands, dunes, pine forests, meadows, fields and pastures - harbour a remarkable variety of beetle species [16,17]. These environmental differences create ideal conditions for the conservation of a high beetle diversity. The KNP is one of the most valuable but (in entomological terms) lesser known national parks in Poland [18].
The aim of this study was to determine whether low-resolution real-time PCR (LRM-PCR) is suitable as a rapid, cost-effective and accessible method for the identification of necrophagous beetle species. The developed assay, based on two primer sets for COI amplification, in combination with LRM-PCR, allowed the successful identification of half of the beetle species collected in the buffer zone of Kampinos National Park. In addition, this study provides the first ever data on necrophagous beetle species living in the buffer zone of Kampinos National Park. Further details on the references can be found at the end of the document.
2. Materials and Methods
Sampling and Generation of the Standard COI Barcode
Specimens of necrophagous Coleoptera were collected between April and September 2014 from pig carcasses in the buffer zone of Kampinos National Park 30 km north of Warsaw (52°21′ N; 20°47′ E). Genomic DNA was extracted from the legs or whole bodies of the small beetles using a GeneMATRIX Tissue DNA Purification Kit (EURX, Gdańsk, Poland) according to the manufacturer’s instructions. The extracted DNA concentration was normalised to 25 ng/ml and stored at -20°C.
The COI barcode region was amplified with a combination of primers: LCO1480 (GGTCWACWAATCATAAAGATATTGG) [19]. PCR amplification of the COI barcode region was performed in 40 μl reaction volume containing: 20 μl REDTaq Ready Mix (Merck, Darmstadt, Germany), 4 μl 5 μM forward and reverse primers, 2 μl DNA extract and H2O to 40 μl. Thermal cycling was performed on a Veriti 96-well thermal cycler (Applied Biosystems, Foster City, CA, USA) with the following programme: an initial denaturation step at 94°C for 5 min, followed by 35 cycles at 95°C for 30 s, 61°C for 30 s and 72°C for 30 s and a final extension step at 72°C for 10 min. Excess dNTPs and unincorporated primers were removed using a Clean-Up Purification Kit (A&A Biotechnology, Gdynia, Poland). The purified DNA was eluted in 40 μl sterile H2O. Sequencing PCR reactions were prepared in 10 μl volumes and contained: 1 μl BigDye Terminator, 2 μl 5× sequencing buffer (BigDye Terminator v3.1 Cycle Sequencing Kit, Applied Biosystems), 1.6 μl (10 μM) forward or reverse mini barcode primer and 1.5 μl DNA and H2O to 10 μl total volume. The thermal profile for the sequencing reactions consisted of 25 cycles at 96°C for 1 min, 96°C for 10 s, 50°C for 5 s and 60°C for 1.45 min. Amplicons were sequenced using a 3500 xL Genetic Analyser (Applied Biosystems, Foster City, CA, USA) [10].
Primer Design for the Amplicon Melting Curve Analysis
The COI sequences obtained were aligned with Clustal W [20]. Regions of sequence similarity and variation were identified to design universal primers. Primer design was performed using Primer3 software [21] and primers were synthesised by Merck (Darmstadt, Germany). PCR reactions for melting curve analysis were performed using a BioRad CFX96 thermal cycler (Bio-Rad, Hercules, CA, USA). Each 20 μl reaction mixture contained: 10 μl Real Time 2 × RT PCR Mix EvaGreen (A&A Biotechnology, Gdynia, Poland), 2 μl 5 μM of each primer, 2 μl (50 ng) DNA extract and 4 μl H2O to a total volume of 20 μl. To maximise reproducibility, an equal amount (50 ng) of DNA was used for amplification and samples were taken from the outer wells of the plate. The target sequences were amplified using a thermal programme with an initial denaturation step of 95°C for 3 min, followed by 40 cycles of 95°C for 10 s, 61°C for 30 s and 72°C for 15 s. The melting curve was determined by observing the melting curve. The melting curve was analysed by observing the fluorescence changes when the temperature was increased from 65°C to 95°C.
3. Results
3.1. Preparation of Reference DNA Samples
The amplified COI products were sequenced using the Sanger sequencing method. The resulting sequences were deposited in GenBank and identified to species level by BLASTn analysis. The sequences obtained from 94 specimens were assigned to 29 Coleoptera species (Table 1). The degree of identity of these sequences, with the exception of C. maxillosus (PQ740169), ranged from 97.03% (O. murinus, PQ740150) to 100% (P. auratus, PQ740106; A. glabra, PQ740105; C. fuscus, PQ740101).
The identified species were distributed across twelve Coleoptera families, with two families being the most species-rich. The Silphidae family comprised six species: Necrodes littoralis, Nicrophorus investigator, Oiceoptoma thoracicum, Silpha tristis, Thanatophilus rugosus, and Thanatophilus sinuatus. The Histeridae family included five species: Hister unicolor, Hister rugifrons, Margarinotus brunneus, Saprinus semistriatus, and Saprinus planiusculus. The Staphylinidae were represented by four species: A. curtula, C. maxillosus, O. Murinus, and P. cognatus. The last nine families comprised of 1 – 3 species.
3.2. Analysis of the DNA Melting Profile
The COI sequences obtained were aligned, conserved regions identified and two primer pairs of designed: Coleop I (forward: CGCTAATTGGAGATGATCAAAT and reverse: CCTGTTCCTGCTCCTCTTTC) and Coleop II (forward: TAGCAACTCTTTATGGAACTCAA and reverse: GCTCATAAAGTAGCAGGGGAAT). The Coleop I primers amplify COI in eleven Coleoptera species, while the Coleop II primers amplify COI in nine species. Coleop I fails to amplify COI in D. undulatus, N. littoralis, and O. thoracicum, while Coleop II fails in A. stercorosus, C. maxillosus, H. unicolor, M. brunneus, and T. rugosus. Both primer pairs successfully amplify COI in six species: A. curtula, N. vespilloides, O. murinus, P. cognatus, S. planisculus, and S. tristis).
The melting temperature (Tm) of the Coleop I amplicons ranged from 72.1°C to 78.0°C. It is noteworthy that, the Tm values for S. planisculus and A. curtula were almost identical (76.0°C and 76.1°C, respectively). In contrast, the Tm values of Coleop II amplicons differed considerably between these species (S. planisculus: 69.4 ± 0.16°C, A. curtula: 70.0 ± 0.10°C). Similarly, the Coleop II amplicons of A. curtula and S. tristis exhibited identical Tm values (70.0°C), while the Coleop I amplicons of these species displayed significant differences in Tm (A. curtula: 76.1 ± 0.16°C, S. tristis: 76.5 ± 0.11°C).
4. Discussion
The melting temperature depends on both the length and the GC content of the amplicon, since G and C are bound by three hydrogen bonds, while A and T are bound by two. Therefore, the Tm of the amplicons differs for fragments with high and low GC content [22]. We used the EvaGreen dye to measure Tm because this fluorescent dye does not redistribute when DNA is melted, which increases the accuracy of Tm measurement [23].
The developed LRM-PCR assay allows the identification of 14 beetle species from biological samples collected from pig cadavers (Table 2). The assay is cost-effective as it does not require specific fluorescently labelled primers for each species and only two sets of primers and saturating fluorescent dye. In addition, the assay procedure is time-saving: the time required from LRM-PCR to interpretation of the results is only 2 hours. Previous methods based on real-time PCR in combination with species-specific probes are simple, inexpensive and time-saving, but the number of target species is limited by the number of detection channels of the instruments. The data obtained in this study are consistent with previously published [13,14,15].
Carrion or dead animal material provides a temporary and variable food source for a diverse and varied community of organisms [24]. Insects, especially Coleoptera and Diptera, are the main components of this community and represent a major element of the decomposition process. The order Coleoptera includes a number of forensically important families: Staphylinidae, Nitidulidae, Scarabaeidae, Silphidae, Dermestidae and Histeridae [1]. The carrion beetles (Silphidae) belong to a group of insects that are closely associated with the decomposition of animal remains and other decaying organic materials. The family Silphidae is divided into two subfamilies: Silphinae and Nicrophorinae. Worldwide, the family comprises 183 species spread over 15 genera. In north-west Europe, 28 species have been recorded: 17 from the subfamily Silphinae and 11 from the subfamily Nicrophorinae. Although the Silphidae are distributed worldwide, they are most common in temperate regions. Most species of the Silphidae family are scavengers that feed on decaying matter, but they also feed on other organisms associated with carrion, such as fly eggs, maggots and smaller carrion beetles. [25,26]. Six species of Silphidae were observed in this study (Table 1). When examining the faunal sequence of beetles on the carcasses, carrion beetles (Silphidae) are the first to be attracted, followed by weevils (Staphylinidae) and clown beetles (Histeridae) [26]. Among them, Necrodes littoralis stands out in Central Europe due to its frequent visits and broods on large vertebrate carcasses, especially in spring and summer. This species colonises carcasses in the later stages of decomposition, significantly extending the period during which the post-mortem interval (PMI) can be estimated from insect activity. More than 90 % of the documented cases occurred outdoors, especially in forests, bushes and fields [27]. Other important necrophagous species are Thanatophilus sinuatus and T. rugosus. These beetles are widely distributed throughout the Palaearctic region and are frequently found on both human and animal cadavers [28].
The second largest species-rich family is the Histeridae. Beetles of the family Histeridae are a stable component of carrion and dung communities. They are predators that live in animal dung and carrion, where they feed on other insects. Both larvae and adults are predatory and like to feed on juicy maggots and fly pupae. Adult and immature Histeridae are found in association with decaying animal or plant matter, suggesting that they are primarily scavengers [29]. Five species of Histeridae were found in our study. Saprinus spp. is usually abundant on decaying carcasses and occurs in a variety of habitats. Their occurrence and presence are related to bloating and active decomposition. Temperature and food availability appear to be the most important factors for the presence of S. semistriatus [30]. While many of the detected species (Table 1), in particular Margarinotus brunneus (Histeridae), Creophilus maxillosus (Staphylinidae), Saprinus detersus (Histeridae) and Thanatophilus sinuatus (Silphidae), have been associated with the putrefaction stages of cadaver decomposition, Dermestes frischii and Dermestes undulatus are most commonly found at outdoor and indoor crime scenes during the dry and skeletal stages of decomposition [31]. Eight species of the genus Dermestes have been identified in Europe: D. frischii, D. maculatus, D. undulatus, D. ater, D. bicolor, D. haemorrhoidalis, D. lardarius, and D. peruvianus. In arid environments, Dermestes species are probably the only necrophagous insects that feed on decaying remains [32,33].
Comprehensive surveys of the necrophagous Coleoptera of Poland in the Wielkopolskie Voivodeship in central Poland have revealed the presence of beetles of the families Silphidae [34], Staphylinidae [35,36] and Dermestidae [37]. During investigations on a military training area near Poznań, it was found that in addition to the Silphidae, representatives of the Geotrupidae and Histeridae also belonged to the dominant necrophagous beetle communities. At the same time, adult individuals of Anoplotrupes stercorosus and Hydrotaea similis reached minimum abundance on all pig carcasses in all seasons [38]. In western Poland, pine-oak, hornbeam-oak and alder forests had a similar composition of carrion beetles, including adults of N. littoralis, T. rugosus, C. maxillosus, Omalium rivulare, Oxypoda acuminata and Philonthus spp, as well as larvae of N. littoralis, C. maxillosus and Philonthus spp. However, differences were observed in the occurrence and activity times of certain taxa in the different forest types [34]. Similarly, fluctuations in the seasonal activity of Silphidae were described by Urbański and Baraniak [39]. The beetles from the family Silphidae were also frequently found on decaying pig carcasses in the forests of Subcarpathian Voivodeship (south-east Poland) [40] and in the Masurian Lake District in north-east Poland [41]. The occurrence of necrophages from the Staphylinidae family has been reported from all over Poland [2,36,42]. In addition to the Wielkopolskie Dermestidae (Dermestes haemorrhoidalis), mummified human remains were collected from a dwelling in the Lower Silesian Voivodeship, i.e. south-west Poland [43].
In Kampinos National Park and its buffer area, Coleoptera studies were limited to groups of beetles living in decaying birch wood [44] coprophagus and represented by 33 species belonging to three families: Geotrupidae, Hydrophilidae and Scarabaeidae [45], ladybird beetles [46], Scarabaeidae [47,48], Tetratomidae [49], Ptiliidae [50], and some families of Cleroidea [51,52]. However, there is only one publication dealing specifically with necrophagous beetles [53]. Moreover, Necrobia ruficollis (Fabricius, 1775) (Coleoptera: Cleridae), a species commonly found on heavily decomposed and skeletonised corpses [54], has been recorded in Poland in only one locality [55]. Our study provides new data on the occurrence of necrophagous beetles in the buffer zone of the Kampinos National Park.
5. Conclusions
Current advanced molecular methods such as DNA barcoding or HRM-PCR allow precise identification of beetle species in both the adult and larval stages, but require specialised laboratories. The currently developed diagnostic key based on amplicon melting profiles requires relatively simple real-time PCR thermocyclers, allowing a broader application of molecular techniques in forensic entomology. The future development of diagnostic identification keys for the reliable identification of necrophagous species collected as part of insect succession studies on carrion or from the remains of organic matter of deceased individuals in medico-legal investigations will further advance forensic entomology as a tool in criminal investigations and faunistic research.
Author Contributions
Conceptualization, T.M.; T.O. and J.A.N.; methodology, T.M. and J.A.N.; software, T.M.; validation, T.M.; T.O. and J.A.N.; formal analysis, T.M. and J.A.N.; investigation, K.L.; K.D.B.; resources, K.L.; K.D.B.; M.S.; data curation, T.M.; K.L.; K.D.B.; M.S. and J.A.N.; writing—original draft preparation, T.M.; K.L.; K.D.B.; M.S. and J.A.N.; writing—review and editing, T.M., T.O. and J.A.N.; visualization, M.S and K.L.; supervision, T.M. and J.A.N.; project administration, T.M.; funding acquisition, T.M.; J.A.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The sequence data presented in this study were deposited in NCBI GenBank.
Conflicts of Interest
the authors declare no conflicts of interest.
References
- Byrd, J.H.; Tomberlin, J.K. Forensic Entomology: The Utility of Arthropods in Legal Investigations; 3rd ed.; CRC Press: London, UK, 2019. [CrossRef]
- Matuszewski, S.; Bajerlein, D.; Konwerski, S.; Szpila, K. Insect succession and carrion decomposition in selected forests of Central Europe. Part 3: Succession of carrion fauna. Forensic Sci. Int. 2011, 207(1-3), 150–163. [CrossRef]
- Matuszewski, S. Matuszewski, S. Post-mortem interval estimation based on insect evidence: current challenges. Insects 2021, 12(4), 314. [Google Scholar] [CrossRef]
- Sharma, R.; Garg, R. K.; Gaur, J. R. Various methods for the estimation of the post mortem interval from Calliphoridae: A review. Egyptian J. Forensic Sci. 2015, 5(1), 1–12. [Google Scholar] [CrossRef]
- Ridgeway, J. A.; Midgley, J. M.; Collett, I. J.; Villet, M. H. Advantages of using development models of the carrion beetles Thanatophilus micans (Fabricius) and T. mutilatus (Castelneau) (Coleoptera: Silphidae) for estimating minimum post mortem intervals, verified with case data. Int. J. Legal Med. 2014, 128, 207–220. [CrossRef]
- Kočarek, P. Decomposition and Coleoptera succession on exposed carrion of small mammal in Opava, the Czech Republic. Eur. J. Soil Biol. 2003, 39, 31–45. [Google Scholar] [CrossRef]
- Castro, C. P.; García, M. D.; da Silva, P. M.; Silva, I. F.; Serrano, A. Coleoptera of forensic interest: A study of seasonal community composition and succession in Lisbon, Portugal. Forensic Sci. Int. 2013, 232(1–3), 73–83. [CrossRef]
- Ratnasingham, S.; Hebert, P. D. BOLD: The Barcode of Life Data System (http://www. barcodinglife. org). Mol. Ecol. Notes 2007, 7(3), 355–364. [CrossRef]
- Tamburro, M.; Ripabelli, G. High Resolution Melting as a rapid, reliable, accurate and cost-effective emerging tool for genotyping pathogenic bacteria and enhancing molecular epidemiological surveillance: a comprehensive review of the literature. Ann. Ig. 2017, 29(4), 293–316. [Google Scholar] [CrossRef]
- Malewski, T.; Draber-Mońko, A.; Pomorski, J.; Łoś, M.; Bogdanowicz, W. Identification of forensically important blowfly species (Diptera: Calliphoridae) by high-resolution melting PCR analysis. Int. J. Legal Med. 2010, 124, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Behrens-Chapuis, S.; Malewski, T.; Suchecka, E.; Geiger, M. F.; Herder, F.; Bogdanowicz, W. Discriminating European cyprinid specimens by barcode high-resolution melting analysis (Bar-HRM) - A cost efficient and faster way for specimen assignment? Fish Res. 2018, 204, 61–73. [Google Scholar] [CrossRef]
- Ramón-Laca, A.; Gleeson, D.; Yockney, I.; Perry, M.; Nugent, G.; Forsyth, D. M. Reliable discrimination of 10 ungulate species using high resolution melting analysis of faecal DNA. PLoS ONE 2014, 9(3), e92043. [Google Scholar] [CrossRef]
- Winder, L.; Phillips, C.; Richards, N.; Ochoa-Corona, F.; Hardwick, S.; Vink, C. J.; Goldson, S. Evaluation of DNA melting analysis as a tool for species identification. Methods in Ecology and Evolution 2011, 2(3), 312–320. [Google Scholar] [CrossRef]
- Thanakiatkrai, P.; Kitpipit, T. Meat species identification by two direct-triplex real-time PCR assays using low resolution melting. Food Chem. 2017, 233, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Mori, C.; Matsumura, S. Development and validation of simultaneous identification of 26 mammalian and poultry species by a multiplex assay. Int. J. Legal Med. 2022, 136(1), 1–12. [Google Scholar] [CrossRef] [PubMed]
- Michalska-Hejduk, D. Meadows of the “Granica” complex in the Kampinos National Park (Central Poland): geobotanical characteristics and protection proposals. Nature Conservation 2001, 58, 57–67. [Google Scholar]
- Matysiak, A.; Dembek, W. Floristic diversity of plant communities in selected post-agricultural areas of the Kampinos National Park. Woda – Środowisko – Obszary Wiejskie 2007, 6(2), 231–254.
- Banaszak, J.; Buszko, J.; Czachorowski, S.; Czechowska, W.; Hebda, G.; Liana, A.; Pawlowski, J.; Szeptycki, A.; Trojan, P.; Węgierek P. Przegląd badań inwentaryzacyjnych nad owadami w parkach narodowych Polski. Wiadomości Entomologiczne 2004, 23(2), 5–56.
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol 1994, 3, 294–299. [Google Scholar]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23(21), 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B. C.; Remm, M.; Rozen, S. G. Primer3 - new capabilities and interfaces. Nucleic Acids Res. 2012, 40(15), e115–e115. [Google Scholar] [CrossRef] [PubMed]
- Wittwer, C. T.; Hemmert, A. C.; Kent, J. O.; Rejali, N. A. DNA melting analysis. Mol. Aspects Med. 2024, 97, 101268. [Google Scholar] [CrossRef] [PubMed]
- Farrar, J.S., Reed, G.H., Wittwer, C.T., High resolution melting curve analysis for molecular diagnostics. In: Molecular Diagnostics, 2nd ed.; Patrinos, G.P.; Ansorge, W. Elsevier, London, UK, 2010, pp. 229-245. [CrossRef]
- Barton P.S.; Cunningham S.A.; Lindenmayer D.B.; Manning A.D. The role of carrion in maintaining biodiversity and ecological processes in terrestrial ecosystems. Oecologia. 2013;171, 761–772. [CrossRef]
- Dekeirsschieter, J.; Verheggen, F.; Lognay, G.; Haubruge, E. Large carrion beetles (Coleoptera, Silphidae) in Western Europe: a review. Biotechnol. Agron. Soc. Environ 2011, 15(3), 435–447. [Google Scholar]
- Sutton, P. The colonisation of stoat carrion by Nicrophorus spp.(Silphidae). The Coleopterist 2016, 25(1), 11–15. [Google Scholar] [CrossRef]
- Shayya, S.; Dégallier, N.; Nel, A.; Azar, D.; Lackner, T. Contribution to the knowledge of Saprinus Erichson, 1834 of forensic relevance from Lebanon (Coleoptera, Histeridae). ZooKeys 2018, 738, 117–152. [Google Scholar] [CrossRef] [PubMed]
- Charabidze, D.; Vincent, B.; Pasquerault, T.; Hedouin, V. The biology and ecology of Necrodes littoralis, a species of forensic interest in Europe. Int. J. Legal Med 2016, 130(1), 273–280. [Google Scholar] [CrossRef]
- Frątczak-Łagiewska, K.; Matuszewski, S. Resource partitioning between closely related carrion beetles: Thanatophilus sinuatus (F.) and Thanatophilus rugosus (L.) (Coleoptera: Silphidae). Entomol. Gen. 2018, 37, 143–156. [CrossRef]
- Bajerlein, D.; Matuszewski, S.; Konwerski, S. Insect succession on carrion: seasonality, habitat preference and residency of histerid beetles (Coleoptera: Histeridae) visiting pig carrion exposed in various forests (Western Poland). Pol. J. Ecol. 2011, 59, 787–797. [Google Scholar]
- Szelecz, I.; Feddern, N.; Seppey, C. V. W.; Amendt, J.; Mitchell, E. A. D. The importance of Saprinus semistriatus (Coleoptera: Histeridae) for estimating the minimum post-mortem interval. Leg. Med. (Tokyo). [CrossRef]
- Charabidze, D.; Colard, T.; Vincent, B.; Pasquerault, T.; Hedouin, V. Involvement of larder beetles (Coleoptera: Dermestidae) on human cadavers: a review of 81 forensic cases. Int. J. Legal Med 2014, 128(6), 1021–1030. [Google Scholar] [CrossRef] [PubMed]
- Kadej, M.; Szleszkowski, Ł.; Thannhäuser, A.; Jurek, T. Dermestes (s.str.) haemorrhoidalis (Coleoptera: Dermestidae) - The Most Frequent Species on Mummified Human Corpses in Indoor Conditions? Three Cases from Southwestern Poland. Insects 2023, 14, 23. [Google Scholar] [CrossRef] [PubMed]
- Magni, P.A.; Voss, S.C.; Testi, R.; Borrini, M.; Dadour, I.R. A Biological and Procedural Review of Forensically Significant Dermestes Species (Coleoptera: Dermestidae). J. Med. Entomol 2015, 52(5), 755–769. [Google Scholar] [CrossRef] [PubMed]
- Matuszewski, S.; Bajerlein, D.; Konwerski, S.; Szpila, K. An initial study of insect succession and carrion decomposition in various forest habitats of Central Europe. Forensic Sci. Int. [CrossRef]
- Mądra, A.; Konwerski, S.; Matuszewski, S. Necrophilous Staphylininae (Coleoptera: Staphylinidae) as indicators of season of death and corpse relocation. Forensic Sci. Int. 2014, 242, 32–37. [Google Scholar] [CrossRef]
- Frątczak-Łagiewska, K.; Grzywacz, A.; Matuszewski, S. Development and validation of forensically useful growth models for Central European population of Creophilus maxillosus L. (Coleoptera: Staphylinidae). Int. J. Legal Med. 2020, 134, 1531-1545. [CrossRef]
- Matuszewski, S.; Szafałowicz, M. Temperature-dependent appearance of forensically useful beetles on carcasses. Forensic Sci. Int. 2013, 229(1-3), 92-99. [CrossRef]
- Jarmusz, M.; Bajerlein, D. Anoplotrupes stercorosus (Scr.) and Trypocopris vernalis (L.) (Coleoptera: Geotrupidae) visiting exposed pig carrion in forests of Central Europe: seasonality, habitat preferences and influence of smell of decay on their abundances. Entomol. Gen. 2015, 35, 213-228. [CrossRef]
- Urbański, A.; Baraniak, E. Differences in early seasonal activity of three burying beetle species (Coleoptera: Silphidae: Nicrophorus F.) in Poland. Coleopt. Bull. 2015, 69, 283–292.
- Konieczna, K.; Czerniakowski, Z.; Wolański, P. The occurrence and species richnes of nicrophagous Silphidae (Coleoptera) in wooded areas in different degree of urbanization. Baltic Journal of Coleopterology, 2019, 19(2), 213-232.
- Aleksandrowicz, O.; Komosinski, K. On the fauna of carrion beetles (Coleoptera, Silphidae) of Mazurian Lakeland (north-eastern Poland). In: Protection of coleoptera in the Baltic Sea region. Skłodowski, J.; Huruk, S.; Barševskis, A.; Tarasiuk, S. Agricultural University Press, Warsaw, Poland, 2005, pp. 147–153.
- Mazur, A.; Melke, A. Staphylinina (Coleoptera: Staphylininae) of Poland. Wydawnictwo Uniwersytetu Przyrodniczego w Poznaniu, Poznań, Poland, 2022.
- Kadej, M.; Szleszkowski, Ł.; Thannhäuser, A.; Jurek, T. A mummified human corpse and associated insects of forensic importance in indoor conditions. Int. J. Legal Med. 2020, 134(5), 1963–1971. [Google Scholar] [CrossRef] [PubMed]
- Sawoniewicz, M. Beetles (Coleoptera) occurring in decaying birch (Betula spp.) wood in the Kampinos National Park. Forest Research Papers, 2013, 74 (1), 71–85. [CrossRef]
- Mroczyński, R.; Marczak, D. Coprophagous beetles (Coleoptera) found in moose (Alces alces L.) feces in Kampinos National Park. World Scientific News, 2017, 86(3), 365-370.
- Szawaryn, K.; Marczak, D. Ladybird beetles (Coleoptera: Coccinellidae) of Kampinos National Park. Entomological News (Poland), 2021, 14-29.
- Mroczyński, R.; Marczak, D. A contribution to the knowledge of the fauna of the Kampinos National Park: Scarabaeidae. Part 1. Subfamilies: Melolonthinae, Sericinae, Rutelinae, Dynastinae i Cetoninae. Entomological News, 2016, 35(3), 161–171.
- Mroczyński, R.; Marczak, D. A contribution to knowledge of fauna of Kampinos National Park: Scarabaeidae. Part 2: Subfamilies: Aphodiinae, Scarabaeinae. Entomological News, 2016, 35(4), 212–224.
- Marczak, D.; Mroczyński, R.; Masiarz, J. Contribution to the knowledge of the fauna of Kampinos National Park: Tetratomidae (Coleoptera: Tenebrionoidea). World Scientific News, 2018, 107, 196-200.
- Marczak, D.; Komosiński, K.; Masiarz, J. Contribution to the knowledge of the fauna of Kampinos National Park: Ptiliidae (Coleoptera: Staphylinoidea). World Scientific News, 2017, 83, 1-14.
- Marczak, D.; Jerzy Borowski, J.; Jędryczkowski, W. A contribution to the knowledge of the fauna of the Kampinos National Park: Dasytidae, Malachiidae (Coleoptera: Cleroidea). Entomological News, 2016, 35(2), 72-81.
- Malewski, T.; Łoś, M.; Sołtyszewski I. Application of HRM–PCR (high resolution melting PCR) for identification of forensically important Coleoptera species. Forensic Sci. Int.: Genetic Supplement Series, 2019, 7(1), 132-134. [CrossRef]
- Dekeirsschieter, J.; Frederickx, C.; Verheggen, F. J.; Boxho, P; Haubruge, E. Forensic entomology investigations from Doctor Marcel Leclercq (1924–2008): a review of cases from 1969 to 2005. J. Med. Entomol. 2013, 50(5), 935-954. [CrossRef]
- Tomsia, M.; Grzywacz, A.; Szpila, K.; Walczak, K.; Mahlerová, K.; Vaněk, D.; Matuszewski, S. Human costal cartilage, tooth cavities, and femur nutrient canals—new niches for insects used in forensic entomology. Forensic Sci. Res. 2024, owae028. [CrossRef]
Figure 1.
The workflow for the identification of necrophagous Carabidae.
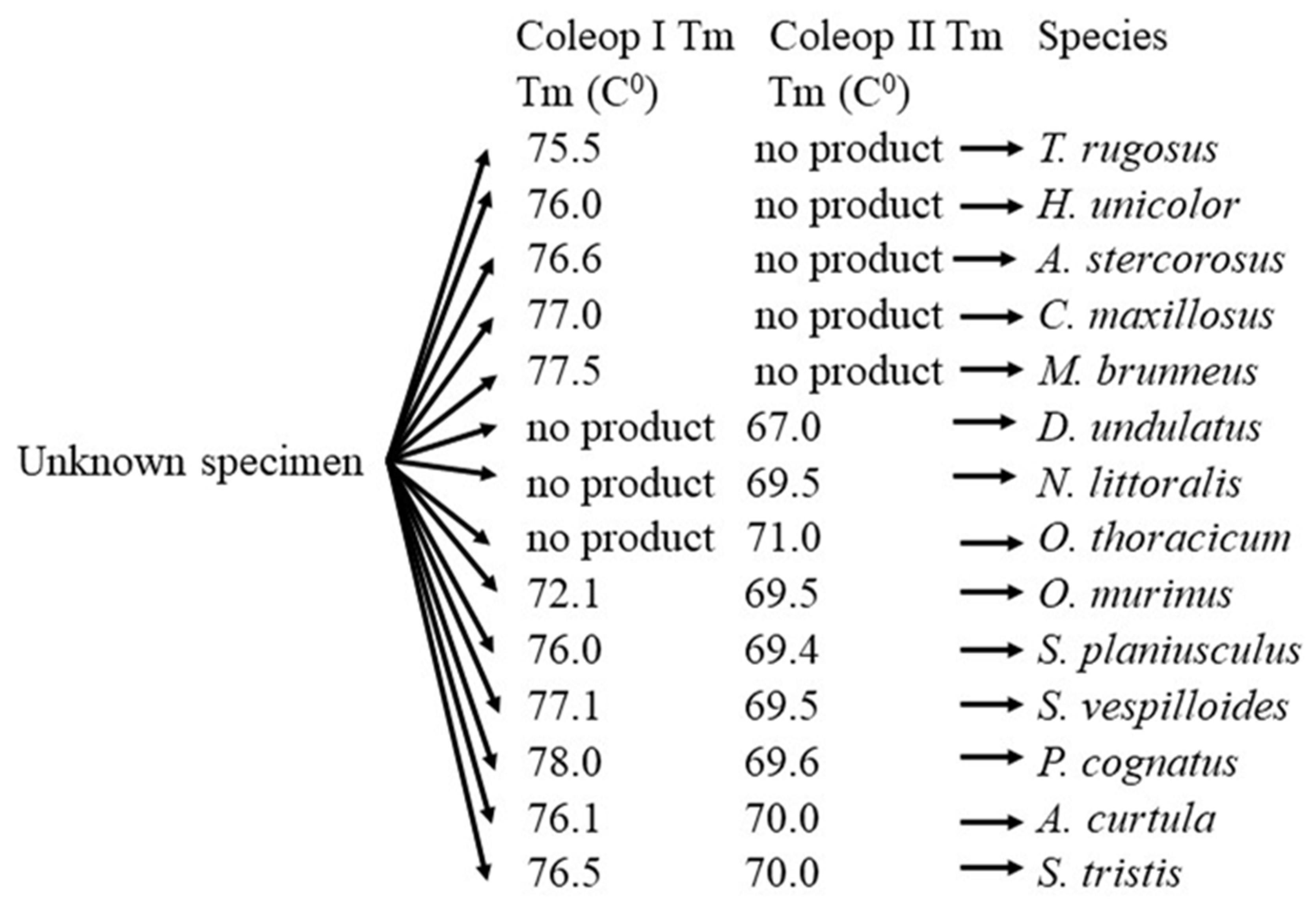

Table 1.
Minimal identity of COI sequence of the specimen.
| BLAST top-hit | Specimen GenBank | ||
|---|---|---|---|
| Identity (%) | GenBank Acc no | Closest Match | Acc no |
| 98.94 | KU906252.1 | Aleochara curtula (Goeze, 1777) | PQ740092 |
| 98.81 | KU906252.1 | A. curtula | PQ740091 |
| 98.93 | KU906252.1 | A. curtula | PQ740090 |
| 99.24 | KU906252.1 | A. curtula | PQ740089 |
| 98.96 | KU906252.1 | A. curtula | PQ740088 |
| 99.37 | KU906252.1 | A. curtula | PQ740087 |
| 100 | KM443099.1 | Anisotoma glabra (Kugelann, 1794) | PQ740105 |
| 98.72 | KM451400.1 | Anoplotrupes stercorosus (Scriba, 1791) | PQ740173 |
| 99.1 | KM451400.1 | A. stercorosus | PQ740172 |
| 98.89 | HQ165003.1 | A. stercorosus | PQ740171 |
| 99.26 | KF317270.1 | Athous subfuscus (Müller, 1764) | PQ740124 |
| Dear Colleagues, | KF317270.1 | A. subfuscus | PQ740123 |
| 99.36 | KF317270.1 | A. subfuscus | PQ740122 |
| 100 | KF317270.1 | A. subfuscus | PQ740121 |
| 99.27 | KF317270.1 | A. subfuscus | PQ740120 |
| 97.09 | HM411773.1 | Carabus nemoralis (Müller, 1764) | Dear Colleagues, |
| 98.97 | HM411773.1 | C. nemoralis | PQ740136 |
| 99.46 | HM411773.1 | C. nemoralis | PQ740135 |
| 99.16 | Dear Colleagues, | Dear Colleagues, | PQ740104 |
| 99.23 | KM449140.1 | C. fuscus | PQ740103 |
| 99.36 | KM449140.1 | C. fuscus | PQ740102 |
| 99.39 | KJ966129.1 | Creophilus maxillosus (Linnaeus, 1758) | PQ740170 |
| 96.75 | KJ966129.1 | C. maxillosus | PQ740169 |
| 99 | KJ966129.1 | C. maxillosus | PQ740168 |
| 97.4 | KU494101.1 | Dermestes undulatus (Brahm, 1790) | PQ740167 |
| 98.67 | KU494101.1 | D. undulatus | PQ740166 |
| 98.7 | KU494101.1 | D. undulatus | PQ740165 |
| 99.09 | JX064157.1 | Geotrupes stercorarius Linnaeus, 1758 | PQ740108 |
| 99.19 | JX064157.1 | G. stercorarius | PQ740107 |
| Dear Colleagues, | KU915456.1 | Hister unicolor (Linnaeus, 1758) | PQ740164 |
| 99.44 | KU915456.1 | H. unicolor | Dear Colleagues, |
| 98.82 | KU915456.1 | H. unicolor | PQ740160 |
| 99.2 | JF889776.1 | Hypocaccus rugifrons (Paykull, 1798) | PQ740114 |
| 99.42 | JF889776.1 | H. rugifrons | PQ740113 |
| 98.83 | KU908440.1 | Margarinotus brunneus (Fabricius, 1775) | PQ740162 |
| 98.49 | KU908440.1 | M. brunneus | PQ740161 |
| 98.97 | KU908440.1 | M. brunneus | PQ740159 |
| 99.65 | MG456750.1 | Necrobia ruficollis (Fabricius, 1775) | PQ740131 |
| 97.98 | KM452149.1 | Necrodes littoralis (Linnaeus, 1758) | PQ740158 |
| 98.52 | KM452149.1 | N. littoralis | PQ740157 |
| 98.65 | KM452149.1 | N. littoralis | PQ740156 |
| 98.09 | KU915489.1 | Nicrophorus investigator (Zetterstedt, 1824) | PQ740119 |
| 98.5 | KU915489.1 | N. investigator | PQ740118 |
| 99.36 | KU915489.1 | N. investigator | PQ740117 |
| 99.35 | KU915079.1 | Nitidula rufipes (Linnaeus, 1767) | PQ740128 |
| 99.16 | KU915079.1 | N. rufipes | PQ740127 |
| 99.85 | KU915079.1 | N. rufipes | PQ740126 |
| 99.48 | KU915079.1 | N. rufipes | PQ740125 |
| 99.01 | MZ659350.1 | Oiceoptoma thoracicum (Linnaeus, 1758) | PQ740155 |
| 98.68 | MZ659350.1 | O. thoracicum | PQ740154 |
| 98.19 | MZ659350.1 | O. thoracicum | PQ740153 |
| 98.74 | LR742640.1 | O. thoracicum | PQ740152 |
| 98.87 | KM440670.1 | Ontholestes murinus (Linnaeus, 1758) | PQ740151 |
| 97.03 | KM440670.1 | O. murinus | PQ740150 |
| 99.11 | KM440670.1 | O. murinus | PQ740149 |
| 98.92 | KM444600.1 | O. murinus | PQ740148 |
| 100 | OL343377.1 | Phelotrupes auratus (Motschulsky, 1857) | PQ740106 |
| 98.24 | KR485683.1 | Philonthus cognatus (Stephens, 1832) | PQ740086 |
| 98.85 | KR485683.1 | P. cognatus | PQ740085 |
| 99.32 | KR485683.1 | P. cognatus | PQ740084 |
| 99.43 | KR485683.1 | P. cognatus | PQ740083 |
| 99.37 | KR485683.1 | P. cognatus | PQ740082 |
| 99.07 | KR485683.1 | P. cognatus | PQ740081 |
| 99.01 | KM441423.1 | Ptenidium nitidum (Heer, 1841) | PQ740096 |
| 99.21 | KM441423.1 | P. nitidum | PQ740095 |
| 99.38 | KM441423.1 | P. nitidum | PQ740094 |
| 99.48 | KM441423.1 | P. nitidum | PQ740093 |
| 99.27 | MN454714.1 | Pterostichus nigrita (Paykull, 1790) | PQ740134 |
| 99.26 | MN454714.1 | P. nigrita | PQ740133 |
| 99.34 | MN454714.1 | P. nigrita | PQ740132 |
| 98.57 | MH307935.1 | Saprinus planiusculus (Motschulsky, 1849) | PQ740147 |
| 99.52 | MH307935.1 | S. planiusculus | PQ740146 |
| 99.26 | MH307935.1 | S. planiusculus | PQ740145 |
| 99.18 | KM439324.1 | Saprinus semistriatus (L.G.Scriba, 1790) | PQ740112 |
| 99.28 | KM439324.1 | S. semistriatus | PQ740111 |
| 99.16 | KM439324.1 | S. semistriatus | PQ740110 |
| 99.36 | KM439324.1 | S. semistriatus | PQ740109 |
| 99.18 | KM849301.1 | Sciodrepoides watsoni (Spence, 1813) | PQ740100 |
| 97.75 | KM849301.1 | S. watsoni | PQ740099 |
| 98.9 | KM849301.1 | S. watsoni | PQ740098 |
| 99.46 | MZ609983.1 | S. watsoni | PQ740097 |
| 98.74 | HQ559261.1 | Silpha tristis (Illiger, 1798) | PQ740116 |
| 99.25 | HQ559261.1 | S. tristis | PQ740115 |
| 99.25 | HQ559261.1 | S. tristis | PQ740174 |
| 99.04 | KU916971.1 | Stephostethus lardarius (DeGeer, 1775) | PQ740130 |
| 99.14 | KU916971.1 | S. lardarius | PQ740129 |
| 98.5 | KM441510.1 | Thanatophilus sinuatus (Fabricius, 1775) | PQ740144 |
| 98.14 | KM441510.1 | T. sinuatus | PQ740143 |
| 99.47 | KM441510.1 | T. sinuatus | PQ740142 |
| 99.05 | KM441510.1 | T. sinuatus | PQ740141 |
| 99.19 | KJ963777.1 | Thanatophilus rugosus (Linnaeus, 1758) | PQ740140 |
| 95.26 | KJ963777.1 | T. rugosus | PQ740139 |
| 99.41 | KJ963777.1 | T. rugosus | PQ740138 |
Table 2.
Melting temperature (Tm) of the COI amplicons of Coleop I and Coleop II. The Tm represent the mean value of three repplicates.
Table 2.
Melting temperature (Tm) of the COI amplicons of Coleop I and Coleop II. The Tm represent the mean value of three repplicates.
| Amplicon Tm (C0). Mean ± SD. | Species | |
|---|---|---|
| Coleop II | Coleop I | |
| 67.0 ± 0.17 | no product | Dermestes undulatus |
| 69.5 ± 0.11 | no product | Necrodes littoralis |
| 70.1 ± 0.14 | no product | Oiceoptoma thoracicum |
| no product | 75.5 ± 0.11 | Thanatophilus rugosus |
| no product | 76.0 ± 0.14 | Hister unicolor |
| no product | 76.6 ± 0.15 | Anoplotrupes stercorosus |
| no product | 77.0 ± 0.13 | Creophilus maxillosus |
| no product | 77.5 ± 0.11 | Margarinotus brunneus |
| 69.5 ± 0.12 | 72.1 ± 0.10 | Ontholestes murinus |
| 69.4 ± 0.16 | 76.0 ± 0.09 | Saprinus planiusculus |
| 69.5 ± 0.11 | 77.1 ± 0.17 | Nicrophorus vespilloides |
| 69.6 ± 0.09 | 78.0 ± 0.15 | Philonthus cognatus |
| 70.0 ± 0.10 | 76.1 ± 0.16 | Aleochara curtula |
| 70.0 ± 0.14 | 76.5 ± 0.11 | Silpha tristis |
Based on the data obtained, a workflow for the identification of necrophagous Carabidae is proposed (Figure 1).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



