
Submitted:
30 December 2024
Posted:
31 December 2024
You are already at the latest version
Abstract
Bacillus subtilis spore display is a versatile technology enabling presentation of heterologous proteins on the surface of recombinant spores. Undoubted benefits of such system include easiness of construction, facility of spore production and handling, enhanced stability of displayed proteins. In spite of these, the efficient spore display faces some challenges which need to be taken on in the process of design and construction. In this study, we attempted to determine a minimal anchoring motif of the crust protein CotX which can be used for display of heterologous protein on the spore surface. We succeeded in presenting mChery fluorescent protein as N-terminal fusion of CotX. Nevertheless, our results suggest that only the full-length CotX could serve as anchor in spore display, since fusions with truncated variants of this protein were not detected in the spore coat and were degraded in the cell.
Keywords:
Bacillus subtilis
; surface spore display
; spore crust
; fusion protein
; CotX
1. Introduction
Bacterial surface display is a biotechnological technology which can be used in different applications, including biocatalysis, drug and antigen delivery, or environmental remediation (Mahmodi 2024). One of the most promising variants of this method is the presentation of heterologous proteins on the surface of bacterial spores. A Gram-positive bacterium Bacillus subtilis is a leading microorganism in this field (Stulke 2023). Since the first demonstration of B. subtilis spore display (Isticato 2001) numerous applications of this techniques have been researched (Mahmodi 2024, Isticato 2023).
Major advantages of protein display on the surface of B. subtilis spores are: (i) non-pathogenicity of this microorganism, (ii) ease of genetic manipulation, (iii) facility of cultivation and spore production. While the process of spore formation belongs to B. subtilis physiology (Tan 2014) no special manipulations are required to trigger it. Bacteria initiate sporulation in response to worsening environmental conditions, e.g. nutrient exhaustion naturally occurring at the later stages of bacterial culture growth leading to formation of metabolically dormant forms. The mature spore exhibits extraordinary properties. Condensed genetic material located in the central core is surrounded by thick cortex and protective coat, ensuring spore’s resistance to various physical, chemical and biological agents (Setlow 2006, Khanna 2020). In the form of spores B. subtilis is able to survive for virtually unlimited period of time and commence its vegetative growth in the process of spore germination once environmental conditions become favorable (Christie 2020).
B. subtilis spores possess excellent properties form the point of view of biotechnology. The simplicity of genetic engineering of this microorganism allows a relatively seamless introduction of heterologous genes into its genome. A protein of interest is transcriptionally fused with one of spore coat proteins. In the physiological process of sporulation the fusion is introduced into the coat of the forming spores and hence, the recombinant protein can easily be displayed on the surface of these particles. The specific structure of the coat stabilizes the displayed protein contributing to maintenance of its structure and/or activity even in such unfavorable conditions as stomach or intestines. Due to these, recombinant B. subtilis spores prove to be useful as carriers of antigens in orally administrated vaccines (Isticato 2023). An important feature of spores is the facility of their separation from the growth medium or reaction mixture. Since spores are relatively large particles they easily sediment and can be recovered by simple centrifugation. These features make them especially useful for biocatalysis performed at the industrial scale (Zhang 2020).
Several spore coat proteins were successfully used as anchors for display of heterologous proteins. These are CotB (Isticato 2001), CotC (Mauriello 2004), CotG (Kim 2005) and CotE (Hwang 2013). Another two proteins, CotZ (Hinc 2014) and CotX (Wang 2016) which were used for that purpose belong the outermost layer of the spore coat called the crust. While the application of spore display technology is conceptually straightforward the actual success of an individual project cannot be predicted before construction of the recombinant strain and in each case an empirical approach needs to be applied. With the regard to spore structure the crust seems to be most promising in terms of the successful display of the recombinant protein. The crust consists of at least six different proteins: CotVWXYZ, which belong to the insoluble fraction of the spore coat (Zhang 1993) and CgeA (McKenney et al., 2010; Imamura et al., 2011). CotXYZ proteins were characterized as morphogens of the crust and suggested to be responsible for assembly of this layer (McKenney et al., 2010). Of special interest is the fact that these proteins are most likely glycosylated which should increase their hydrophilicity (Bartels 2019). Data obtained with CotX-GFP and CotV-GFP fusions suggest that these proteins localize on the spore poles and form a cap. CotX was suggested to support initiation and CotV propagation of the forming crust structure (Bartels 2019). Spores produced by the mutant strains lacking any of these proteins have no crust, instead they produce detached “beads on a string” crust-line structures (Shuster 2019).
In this study we wanted to use CotX protein to determine a minimal anchoring motif sufficient for presentation of heterologous proteins on spore surface. Since no crystallography data are available for this protein we relied on in silico analysis of second-order structure as well as on models obtained in homology modelling. By constructing a series of mCherry-CotX fusions we tried to visualize in the spore coat the presence of chimeric proteins. To our surprise mCherry-CotX protein was detected in the coat of spores in which a full-length CotX was used as anchor of the mCherry fluorescent protein. Moreover, results of overproduction of the mCherry-CotX chimeras suggested decreasing stability of the fusions along with shortening of the CotX. Taken together, our data suggest that only a full-length CotX can support spore display of heterologous proteins as well as provide some new insight into the structure-function relationship of this component of the spore crust.
2. Materials and Methods
2.1. Bacterial Strains and Transformation
Bacillus subtilis strains used in this study are listed in Table 1. Plasmid amplifications for nucleotide sequencing and subcloning experiments were performed with Escherichia coli strain DH5α (Sambrook 2012). Bacterial strains were transformed by previously described procedures: CaCl2 mediated transformation of E. coli competent cells (Sambrook 2012) and transformation of B. subtilis (Julkowska 2005).
2.2. Construction of Gene Fusions
DNA coding for the mCherry and CotX coat proteins was PCR amplified using the synthetic DNA fragment (Appendix A) as a template and oligonucleotide pair “mCherry forward” together with appropriate primer for CotX variant (Table 2). The amplification products ranging from 1271 bp (mCherry-CotX full-length) to 907 bp (mCherry-CotX dHSHHSH) was obtained by PCR and cloned into the pDL vector (Bacillus Genetic Stock Center) yielding pWP plasmid series (see Figure 2). Similar approach was used for construction overproducing fusion proteins plasmids, however as the plasmid backbone pHT08 was used.
2.3. Chromosomal Integration
Appropriate plasmids were linearized by digestion with a single cutting restriction enzyme. Linearized DNA was used to transform competent cells of the B. subtilis strain 168 (Anagnostopoulos 1961). Chloramphenicol-resistant (CmR) clones were the result of a double-crossover recombination event, resulting in the interruption of the non-essential amyE gene on the B. subtilis chromosome. Several CmR clones were tested by PCR using chromosomal DNA as a template and oligonucleotides AmyS and AmyA (Isticato 2004) to prime the reaction. Selected clones were called BWP53, BWP55, BWP57 or BWP59 and kept for further studies.
2.4. Preparation of Spores
Sporulation was induced by the exhaustion method in DS (Difco-Sporulation) medium as described by Nicholson and co-workers [14]. Sporulating cultures were harvested 24 h after the initiation of sporulation and purified using a lysozyme treatment to break any residual sporangial cells followed by washing steps in 1 M NaCl, 1 M KCl and water (two times), as described previously (Nicholson 1990). PMSF (0.05 M) was included to inhibit proteolysis. After the final suspension in water spores were treated at 65°C for 1 h to kill any residual cells. The spore suspension was titrated immediately for CFU/ml before freezing at −20°C. By this method we could reliably produce 6 × 1010 spores per litre of DSM culture.
2.5. Extraction of Spore Coat Proteins
Spore coat proteins were extracted from 50 μl of a suspension of spores at high density (1 × 1010 spores per ml) using a decoating extraction buffer as described elsewhere [15]. Extracted proteins were assessed for integrity by SDS-polyacrylamide gel electrophoresis (PAGE) and for concentration by two independent methods: the Pierce BCA Protein Assay (Pierce) and the BioRad DC Protein Assay kit (Bio-Rad).
2.6. Western and Dot Blotting Analyses
Extracted proteins were separated in 12% denaturing polyacrylamide gels, electro transferred onto the nitrocellulose filter Roti-NC (ROTH) and used for Western blotting analysis by standard procedures. Membrane was incubation with anti-mCherry antibodies in PBS buffer followed by secondary antibodies linked with horse radish peroxidase. The membrane was visualised by developing with tetrazolium–5-bromo-4-chloro-3-indolylphosphate according to the manufacturer’s instructions.
3. Results
3.1. In Silico Analysis of the CotX Structure
In the lack of experimental data available for the CotX structure we started our analysis applying homology modelling tools. Unfortunately, CotX present insufficient level of homology to structures of other known proteins, therefore obtained models are of low credibility. We compared a hypothetical model created with use of the threading method available in the iTasser (https://zhanglab.ccmb.med.umich.edu/I-TASSER/) with the model obtained with use of the RaptorX Structure Prediction server (http://raptorx.uchicago.edu/about/). The later predicts 3D structures for protein sequences without close homologs in the Protein Data Bank (PDB). In case of both attempts we obtained models of CotX with low credibility due to lack of proper templates for modelling process. Finally, we used AlphaFold (AF-Q08313-F1-model_v4 CotX) but also in this case the credibility of produced model also may be questioned (Fig.1A). Moreover, all obtained models did not share high structural homology. In this situation, we decided to use a second-order structure analysis of CotX protein for planning of mutant construction. We performed such investigation using genesilico (https://genesilico.pl/meta2/), MESSA (http://prodata.swmed.edu/MESSA/MESSA.cgi) methaservers as well as iTasser, RaptorX property and Robetta (http://robetta.bakerlab.org) software. Based on obtained results we built a consensus secondary structure of CotX presented on Figure 1B and used it for further investigation.
3.2. Construction and Spore Coat Analysis of Strains with Truncated Anchor CotX Proteins
The CotX protein has already been used as anchor for spore display of heterologous protein. The β-galactosidase from Bacillus stearothermophilus has been presented on the spore surface as the C-terminal fusion of this protein (Wang 2016). Therefore, we decided to explore possibilities of creating an N-terminal fusion of CotX with fluorescent protein mCherry (Shaner 2004). Moreover, such approach left the C-terminus free for manipulation, which was important regarding the predicted secondary structure of CotX. The C-terminal part of this protein was predicted to contain a series of repeating α-helical and β-sheet structures (Figure 1B) suggesting natural motifs which could be consecutively removed in construction of truncated versions of anchoring CotX protein. For convenience of cloning we decided to start with synthesis of a gene encoding CotX flanked with mCherry at both termini (Figure 2A). Such synthetic gene served as a template in series of PCR reactions in which we amplified the mCherry-encoding gene fused with truncated versions of cotX gene (Figure 2C). Obtained PCR fragments were then cloned into to pDL shuttle vector containing a physiological promoter of the cotX gene. After verification of construction correctness plasmids were used for transformation of B. subtilis 168. Obtained chloramphenicol-resistant colonies were products of double crossing-over events in which mCherry-CotX fusions were integrated with host genome in the non-essential locus amyE. Upon PCR verification of integration correctness recombinant strains were used to produce spores. The presence of mCherry-CotX fusions in spore coat was analyzed in extracts obtained from purified spores using Western blotting with mCherry-specific antibody. Surprisingly, we were able to detect mCherry only in the case of fusion with the full-length CotX (Figure 3).
Figure 2.
Construction of B. subtilis strains expressing mCherry-CotX fusion protein and its derivatives. A – A structure of synthetic fusion gene obtained de novo. Introduced restriction sites allow easy cloning its variants into pDL shuttle vector; B – schematic representation amplicon cloned as variants of mCherry-CotX fusion gene; C- pDL shuttle vector with cloned a variant of mCherry-CotX fusion gene; D – fragment of pDL shuttle vector integrated into amyE locus of B. subtilis chromosome.
Figure 2.
Construction of B. subtilis strains expressing mCherry-CotX fusion protein and its derivatives. A – A structure of synthetic fusion gene obtained de novo. Introduced restriction sites allow easy cloning its variants into pDL shuttle vector; B – schematic representation amplicon cloned as variants of mCherry-CotX fusion gene; C- pDL shuttle vector with cloned a variant of mCherry-CotX fusion gene; D – fragment of pDL shuttle vector integrated into amyE locus of B. subtilis chromosome.
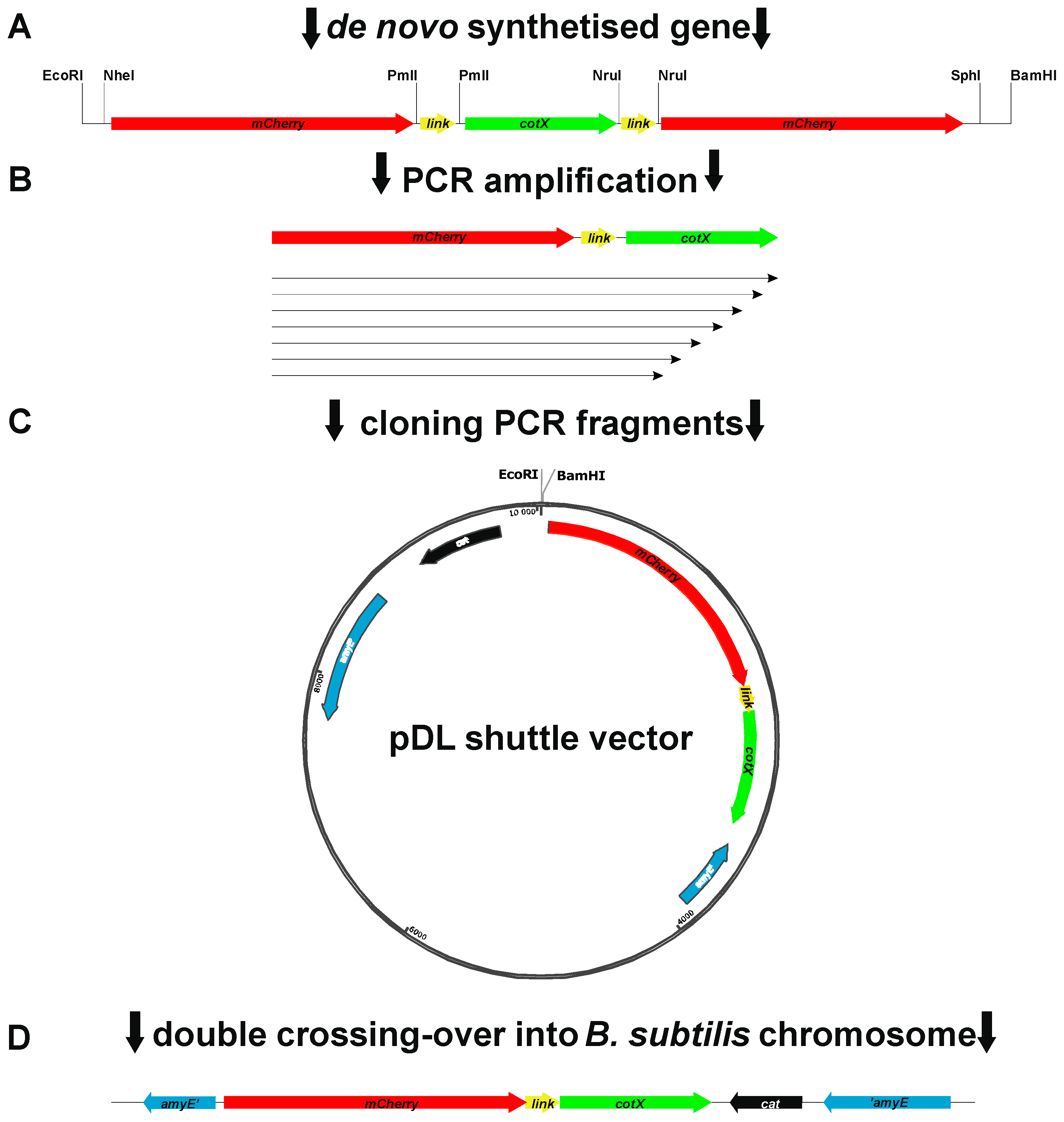
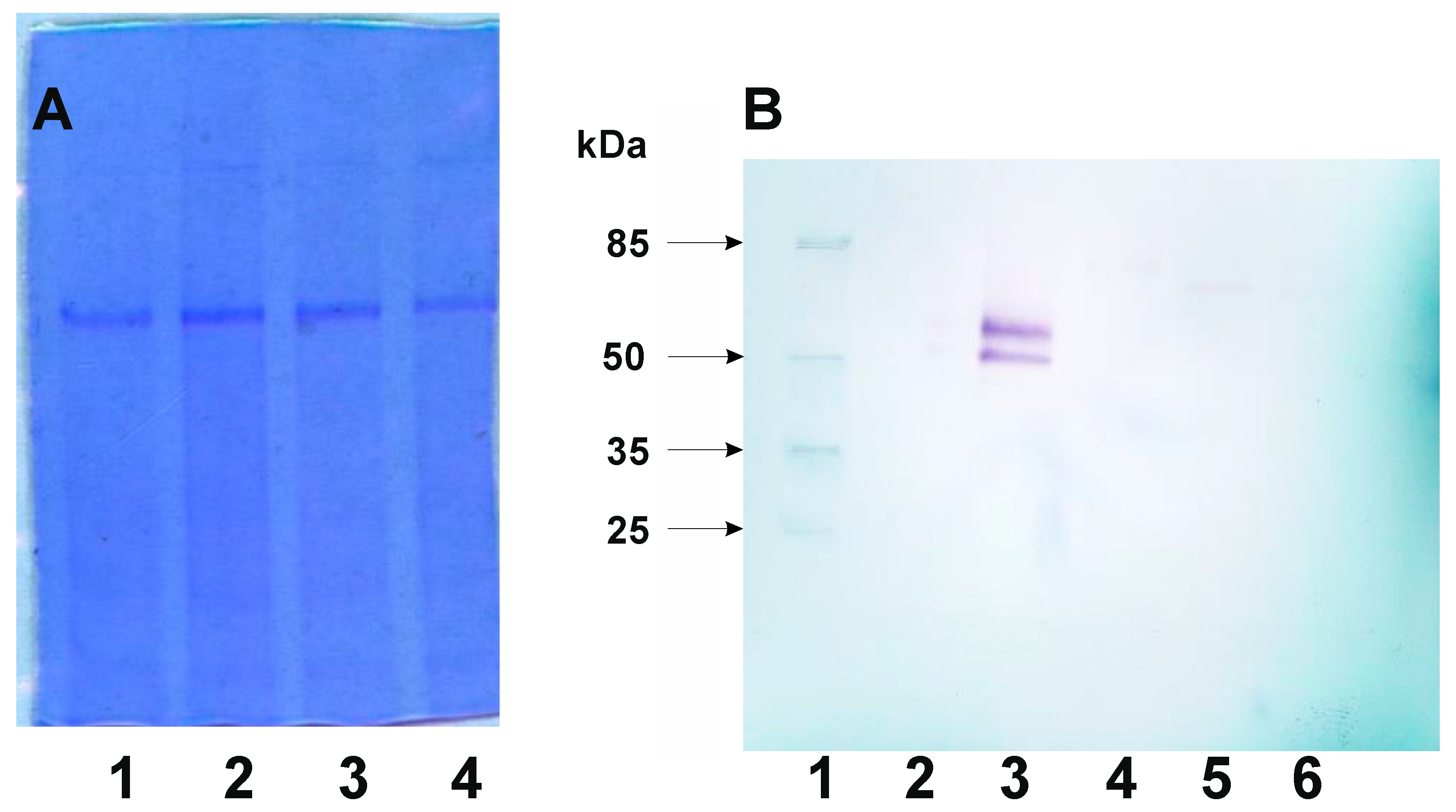
Figure 3.
Detection fusion protein CotX-mCherry and its derivatives from spore coat protein extract. A- Loading control. Lines contain 10μg of spore coat protein extract. 1 – strain BWP 53 (CotX-mCherry); 2 – strain BWP 55 (CotX ΔSH – mCherry); 3 - strain BWP – 57 (CotX ΔHHSH – mCherry); 4 - strain BWP59 (CotX ΔHSHHSH – mCherry). B- Western blotting detection of CotX-mCherry fusion protein with antibody against mCherry. 1 – prestain protein ladder; 2 – spore coat extract from B. subtilis 168 (negative control); 3 - strain BWP 53 (CotX-mCherry); 4 - strain BWP 55 (CotX ΔSH – mCherry); 5- strain BWP – 57 (CotX ΔHHSH – mCherry); 6 - strain BWP59 (CotX ΔHSHHSH – mCherry).
Figure 3.
Detection fusion protein CotX-mCherry and its derivatives from spore coat protein extract. A- Loading control. Lines contain 10μg of spore coat protein extract. 1 – strain BWP 53 (CotX-mCherry); 2 – strain BWP 55 (CotX ΔSH – mCherry); 3 - strain BWP – 57 (CotX ΔHHSH – mCherry); 4 - strain BWP59 (CotX ΔHSHHSH – mCherry). B- Western blotting detection of CotX-mCherry fusion protein with antibody against mCherry. 1 – prestain protein ladder; 2 – spore coat extract from B. subtilis 168 (negative control); 3 - strain BWP 53 (CotX-mCherry); 4 - strain BWP 55 (CotX ΔSH – mCherry); 5- strain BWP – 57 (CotX ΔHHSH – mCherry); 6 - strain BWP59 (CotX ΔHSHHSH – mCherry).
3.3. Detection of mCherry-CotX Fusions Overproduced in B. subtilis Cells
In the recombinant B. subtilis strains mCherry-CotX fusions were expressed as single genes controlled by their physiological promoter. Therefore, it was plausible to assume that detection of relatively low levels of fusion proteins could be hindered, especially if we took into consideration that the truncated versions of CotX might be incorporated into the forming crust structure with lower efficiency. To overcome such presumptive obstacle in detection of the mCherry-CotX fusion proteins we decided to express them from an inducible pgrac promoter located on the replicative vector pHT08 (MoBiTec, Germany). Upon construction of appropriate plasmids their correctness was verified by DNA sequencing. Verified plasmids were used for transformation of B. subtilis 168. Obtained strains were analyzed for mCherry-CotX overproduction induced by addition of 1mM IPTG. Upon 2 hours of induction cells were harvested by centrifugation and lysed. The presence of mCherry-CotX fusions was analyzed in prepared lysates using Western blotting with mCherry-specific antibody. We were able to observe mCherry-CotX chimeras in all tested strains which appeared as bands corresponding to proteins with decreasing molecular mass (Figure 4). Interestingly, a clear two bands of approximately 35 kDa could be visualized in each sample obtained with strains overexpressing recombinant proteins. While the mCherry itself has the molecular mass of 26.7 kDa these bands might correspond to the specific degradation products of the mCherry-CotX fusion protein, independently on the variant of the used anchoring CotX. In the case of four strains: BWP46 (mCherry-CotX, expected size 46.6 kDa), BWP48 (mCherry-CotXΔH, expected size 44.1 kDa), BWP50 (mCherry-CotXΔSH, expected size 42.3 kDa) and BWP52 (mCherry-CotXΔHSH, expected size 39.9 kDa) we observed additional single bands with molecular mass corresponding to overproduced fusions (Figure 4, lanes 4-7). It is worth noting that these bands exhibited low intensities, suggesting low level of overproduction and/or detection of overproduced mCherry-CotX chimeras. Moreover, for the strain BWP54 (mCherry-CotXΔHHSH, expected size 37.5 kDa) the band corresponding to the full-length overexpressed fusion was barely visible (Figure 4, lane 7). In the case of fusion proteins with even shorter variants of CotX: BWP54 (mCherry-CotXΔHHSH), BWP56 (mCherry-CotXΔSHHSH, expected size 35.5 kDa) and BWP58 (mCherry-CotXΔHSHHSH, expected size 33.9 kDa), we observed clear products of degradation of the fusion protein, nonetheless corresponding bands exhibited visibly higher intensity (Figure 4, lanes 8-10). To our surprise the strongest signal was observed in the case of the strain BWP56 which expressed mCherry in fusion with almost the shortest variant CotXΔSHHSH (Figure 4, lane 9).
4. Discussion
Since its invention in 2001 (Isticato 2001) the spore display technology has become an interest of various fields of biotechnology, including biocatalysis (Mahmoodi 2024), antigen delivery (Isticato 2023) or bioremediation (Hinc 2010). Regardless on application, the presentation of heterologous proteins on the surface of recombinant spores involves creation of a fusion protein in which a spore coat protein serves as an anchor. The most commonly used anchors are the crust-associated CotG (Hinc 2010b, Negri 2013, Gao 2011) and the outer coat proteins CotB (Isticato 2001, Chen 2015a, Chen 2015b) and CotC (Tavassoli 2013, Yuan 2014, Chan 2023). The outermost layer of the spore, the crust, has also been used as a target for anchoring heterologous proteins. The crust proteins CotZ and CotY (Hinc 2013, He 2016, Vetrakova 2023), as well as CotX (Wang 2016) have been used for that purpose. In spite of expanding body of research conducted on recombinant spores, there are virtually no possibilities of predicting the success of display of a given protein.
Our study aimed on establishing a minimal anchoring motif which could be useful for spore display of heterologous proteins. The choice of CotX was dictated by the fact that this is the crust protein, hence a passenger protein anchored to it should have good chances to be presented on the spore surface. The proof of our working hypothesis required analysis of CotX structure and this turned out to be not a trivial task, since no structural data was available for this protein. Our homology models created with three different programs (iTasser, RaptorX and AlphaFold) proved to share a low level of similarity. The reason for that was most probably the lack of sufficient homologs which could have been used as a modelling template. Due to that fact, in the design of our fusion proteins we had to rely on results of analysis of CotX secondary structure. Following this reasoning we chose easily distinguishable predicted structural motifs (α-helices and β-sheets) and designed a series of plasmids harboring truncated variants of CotX. The confirmation of the actual presence of predicted motifs would obviously require solving of the CotX structure or at least some structural analysis involving use of appropriate methodology and was not a part of the work plan of this study. We decided to use mCherry, a commonly used fluorescent protein (Shaner 2004), as the passenger in fusions anchored to variants of CotX.
A critical step in the creation of recombinant spores efficiently displaying heterologous proteins is the verification of their presence in the spore coat structure. In the case of our constructs we were able to detect only the fusion in which mCherry was anchored to the full-length CotX (strain BWP46; Figure 3B, lane 3). There are few possible reasons which can be proposed to explain lack of detectable fusion proteins in the coat of recombinant spores produced by other strains. (i) The instability of fusion proteins with truncated variants of CotX. While this protein is highly hydrophobic and belongs to insoluble fraction of spore coat proteins (Zhang 1993), any additional modifications might influence its physicochemical properties rendering it less stable which can result in its misfolding or degradation. The later hypothesis seems plausible especially when considering degradation products observed in the case of strains overexpressing fusions in which mCherry was attached to the shortest variants of CotX (Fig 4, lanes 8-10). (ii) The fusions with truncated CotX are not incorporated into the forming spore crust. This can be caused by the lack of structural motifs required for proper localization of the truncated CotX in the spore. (iii) The level of fusion proteins in the coat of recombinant spores might be below the limit of detection with mCherry-specific antibody used in the study. This hypothesis also seems to be possible, because we were able to detect overproduced fusion proteins (Fig 4.).
The results obtained with strains overproducing mCherry-CotX fusions are especially interesting. On the Western blotting we were able to observe bands corresponding to fusion proteins with expected molecular mass (Figure 4). Nonetheless, their intensities were not the highest, with exception of the strain BWP56 (Figure 4, lane 9) overexpressing mCherry fused to CotXΔSHHSH variant, in which the signal corresponding to the fusion protein was the strongest. In each overproducing strain we observed two approximately 35 kDa proteins detected by anti-mCherry antibody. Since no signal was visible in the control strains (Figure 4, lanes 2 and 3), we can assume that these bands correspond to the degradation products of overexpressed mCherry-CotX fusions. The important fact which should be taken under consideration is that the overexpression of genes encoding analyzed fusion proteins occurs during the vegetative growth, not in the sporulation, when expression of coat genes is expected. This can result in specific degradation of CotX protein which in the vegetative cells cannot be incorporated into the structure of the forming spore. The presence of the degradation products with the same molecular mass, independently on the variant of used CotX, seem to support this hypothesis. An important question arises regarding the degradation products visualized in strains BWP54, BWP56 and BWP58 (Figure 4, lanes 8-10). It seems that there is some minimal length of the CotX which is sufficient to prevent degradation of this protein. Starting with the strain BWP54, CotX was truncated leaving approximately half or less of the predicted α-helix–β-sheet-structured part of the protein. Up to some point, the truncated versions of the C-terminal part of CotX could be resistant to action of putative cellular proteases, which with continuing shortening of this protein could start to cleave it. It is important to emphasize that a stringent glycosylation motif present in the N-terminal part of CotX stays intact in the truncated variants. The glycosylation of CotX was suggested to increase hydrophilicity of this protein (Bartels 2019) and hence contribute to the overall hydrophilic nature of the crust which is important to allow dispersal of the spores in the aqueous environment (McKenney 2010, Imamura 2011, Abe 2014).
To summarize, in this study we constructed recombinant B. subtilis spores displaying mCherry fused to N-terminus of the crust protein CotX. Moreover, we provided evidence that only full-length CotX could be used as anchor in spore display of heterologous proteins. Our results provide also some insight into the nature of CotX, suggesting that its incorporation into the forming spore structure requires expression at appropriate time. Moreover, truncated products of e.g. prematurely terminated translation of cotX might be degraded by cellular proteases.
Author Contributions
WP – performed all wet experiments, MO – contributed discussion during the work and writing the manuscript. Authors read and approved the final manuscript.
Acknowledgments
The research was supported by the National Science Foundation grant no. 2014/13/N/NZ1/04015 for WP. We would like to thank you very much to Adam Iwanicki and Alessandro Negri for its help during manuscript preparation.
Conflicts of Interest
The authors declare that they have no competing interests.
References
- Mahmoodi, A.; Farinas, E.T. Applications of Bacillus subtilis Protein Display for Medicine, Catalysis, Environmental Remediation, and Protein Engineering. Microorganisms. 2024, 12, 97. [Google Scholar] [CrossRef]
- Stülke, J.; Grüppen, A.; Bramkamp, M.; Pelzer, S. Bacillus subtilis, a Swiss Army Knife in Science and Biotechnology. J Bacteriol. 2023, 205, e0010223. [Google Scholar] [CrossRef] [PubMed]
- Isticato, R.; Cangiano, G.; Tran, H.T.; Ciabattini, A.; Medaglini, D.; Oggioni, M.R.; De Felice, M.; Pozzi, G.; Ricca, E. Surface display of recombinant proteins on Bacillus subtilis spores. J Bacteriol. 2001, 183, 6294–301. [Google Scholar] [CrossRef]
- Isticato, R. Bacterial Spore-Based Delivery System: 20 Years of a Versatile Approach for Innovative Vaccines. Biomolecules. 2023, 13, 947. [Google Scholar] [CrossRef]
- Tan, I.S.; Ramamurthi, K.S. Spore formation in Bacillus subtilis. Environ Microbiol Rep. 2014, 6, 212–25. [Google Scholar] [CrossRef]
- Setlow, P. Spores of Bacillus subtilis: their resistance to and killing by radiation, heat and chemicals. J Appl Microbiol. 2006, 101, 514–25. [Google Scholar] [CrossRef]
- Khanna, K.; Lopez-Garrido, J.; Pogliano, K. Shaping an Endospore: Architectural Transformations During Bacillus subtilis Sporulation. Annu Rev Microbiol. 2020, 74, 361–386. [Google Scholar] [CrossRef]
- Christie, G.; Setlow, P. Bacillus spore germination: Knowns, unknowns and what we need to learn. Cell Signal. 2020, 74, 109729. [Google Scholar] [CrossRef]
- Zhang, X.; Al-Dossary, A.; Hussain, M.; Setlow, P.; Li, J. Applications of Bacillus subtilis Spores in Biotechnology and Advanced Materials. Appl Environ Microbiol. 2020, 86, e01096–20. [Google Scholar] [CrossRef]
- Mauriello, E.M.; Duc le, H.; Isticato, R.; Cangiano, G.; Hong, H.A.; De Felice, M.; Ricca, E.; Cutting, S.M. Display of heterologous antigens on the Bacillus subtilis spore coat using CotC as a fusion partner. Vaccine. 2004, 22, 1177–87. [Google Scholar] [CrossRef]
- Kim, J.H.; Lee, C.S.; Kim, B.G. Spore-displayed streptavidin: a live diagnostic tool in biotechnology. Biochem Biophys Res Commun. 2005, 331, 210–4. [Google Scholar] [CrossRef] [PubMed]
- Hwang, B.Y.; Pan, J.G.; Kim, B.G.; Kim, J.H. Functional display of active tetrameric beta-galactosidase using Bacillus subtilis spore display system. J Nanosci Nanotechnol. 2013, 13, 2313–9. [Google Scholar] [CrossRef]
- Hinc, K.; Iwanicki, A.; Obuchowski, M. New stable anchor protein and peptide linker suitable for successful spore surface display in B. subtilis. Microb Cell Fact. 2013, 12, 22. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yang, R.; Hua, X.; Zhang, W.; Zhao, W. An Approach for Lactulose Production Using the CotX-Mediated Spore-Displayed β-Galactosidase as a Biocatalyst. J Microbiol Biotechnol. 2016, 26, 1267–77. [Google Scholar] [CrossRef]
- McKenney, P.T.; Driks, A.; Eskandarian, H.A.; Grabowski, P.; Guberman, J.; Wang, K.H.; Gitai, Z.; Eichenberger, P. A distance-weighted interaction map reveals a previously uncharacterized layer of the Bacillus subtilis spore coat. Curr Biol. 2010, 20, 934–8. [Google Scholar] [CrossRef]
- Zhang, J.; Fitz-James, P.C.; Aronson, A.I. Cloning and characterization of a cluster of genes encoding polypeptides present in the insoluble fraction of the spore coat of Bacillus subtilis. J Bacteriol. 1993, 175, 3757–66. [Google Scholar] [CrossRef]
- Bartels, J.; Blüher, A.; López Castellanos, S.; Richter, M.; Günther, M.; Mascher, T. The Bacillus subtilis endospore crust: protein interaction network, architecture and glycosylation state of a potential glycoprotein layer. Mol Microbiol. 2019, 112, 1576–1592. [Google Scholar] [CrossRef]
- Shuster, B.; Khemmani, M.; Abe, K.; Huang, X.; Nakaya, Y.; Maryn, N.; Buttar, S.; Gonzalez, A.N.; Driks, A.; Sato, T.; Eichenberger, P. Contributions of crust proteins to spore surface properties in Bacillus subtilis. Mol Microbiol. 2019, 111, 825–843. [Google Scholar] [CrossRef]
- Shaner, N.C.; Campbell, R.E.; Steinbach, P.A.; Giepmans, B.N.; Palmer, A.E.; Tsien, R.Y. Improved monomeric red, orange and yellow fluorescent proteins derived from Discosoma sp. red fluorescent protein. Nat Biotechnol. 2004, 22, 1567–72. [Google Scholar] [CrossRef]
- Hinc, K.; Ghandili, S.; Karbalaee, G.; Shali, A.; Noghabi, K.A.; Ricca, E.; Ahmadian, G. Efficient binding of nickel ions to recombinant Bacillus subtilis spores. Res Microbiol. 2010, 161, 757–64. [Google Scholar] [CrossRef]
- Hinc, K.; Isticato, R.; Dembek, M.; Karczewska, J.; Iwanicki, A.; Peszyńska-Sularz, G.; De Felice, M.; Obuchowski, M.; Ricca, E. Expression and display of UreA of Helicobacter acinonychis on the surface of Bacillus subtilis spores. Microb Cell Fact. 2010, 9, 2. [Google Scholar] [CrossRef]
- Negri, A.; Potocki, W.; Iwanicki, A.; Obuchowski, M.; Hinc, K. Expression and display of Clostridium difficile protein FliD on the surface of Bacillus subtilis spores. J Med Microbiol. 2013, 62, 1379–1385. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Xu, X.; Zhang, X.; Che, B.; Ma, C.; Qiu, J.; Tao, F.; Xu, P. Chemoenzymatic synthesis of N-acetyl-D-neuraminic acid from N-acetyl-D-glucosamine by using the spore surface-displayed N-acetyl-D-neuraminic acid aldolase. Appl Environ Microbiol. 2011, 77, 7080–3. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Tian, R.; Ni, Z.; Zhang, Q.; Zhang, T.; Chen, Z.; Chen, K.; Yang, S. Surface display of the thermophilic lipase Tm1350 on the spore of Bacillus subtilis by the CotB anchor protein. Extremophiles. 2015, 19, 799–808. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, T.; Jia, J.; Vastermark, A.; Tian, R.; Ni, Z.; Chen, Z.; Chen, K.; Yang, S. Expression and display of a novel thermostable esterase from Clostridium thermocellum on the surface of Bacillus subtilis using the CotB anchor protein. J Ind Microbiol Biotechnol. 2015, 42, 1439–48. [Google Scholar] [CrossRef] [PubMed]
- Tavassoli, S.; Hinc, K.; Iwanicki, A.; Obuchowski, M.; Ahmadian, G. Investigation of spore coat display of Bacillus subtilis β-galactosidase for developing of whole cell biocatalyst. Arch Microbiol. 2013, 195, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Feng, F.; Chen, L.; et al. Surface display of Acetobacter pasteurianus AdhA on Bacillus subtilis spores to enhance ethanol tolerance for liquor industrial potential. Eur Food Res Technol 2014, 238, 285–293. [Google Scholar] [CrossRef]
- Chan, B.C.; Li, P.; Tsang, M.S.; Sung, J.C.; Kwong, K.W.; Zheng, T.; Hon, S.S.; Lau, C.P.; Cheng, W.; Chen, F.; Lau, C.B.; Leung, P.C.; Wong, C.K. Creating a Vaccine-like Supplement against Respiratory Infection Using Recombinant Bacillus subtilis Spores Expressing SARS-CoV-2 Spike Protein with Natural Products. Molecules. 2023, 28, 4996. [Google Scholar] [CrossRef]
- He, W.; Jiang, B.; Mu, W.; Zhang, T. Production of d-Allulose with d-Psicose 3-Epimerase Expressed and Displayed on the Surface of Bacillus subtilis Spores. J Agric Food Chem. 2016, 64, 7201–7. [Google Scholar] [CrossRef]
- Vetráková, A.; Chovanová, R.K.; Rechtoríková, R.; Krajčíková, D.; Barák, I. Bacillus subtilis spores displaying RBD domain of SARS-CoV-2 spike protein. Comput Struct Biotechnol J. 2023, 21, 1550–1556. [Google Scholar] [CrossRef] [PubMed]
- Imamura, D.; Kuwana, R.; Takamatsu, H.; Watabe, K. Proteins involved in formation of the outermost layer of Bacillus subtilis spores. J Bacteriol. 2011, 193, 4075–80. [Google Scholar] [CrossRef]
- Abe, K.; Kawano, Y.; Iwamoto, K.; Arai, K.; Maruyama, Y.; Eichenberger, P.; Sato, T. Developmentally-regulated excision of the SPβ prophage reconstitutes a gene required for spore envelope maturation in Bacillus subtilis. PLoS Genet. 2014, 10, e1004636. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.F. ; Maniatis, T Molecular cloning: a laboratory manual. 4th edition. Cold Spring Harbor, N.Y: Cold Spring Harbor Laboratory Press, USA. 2012.
- Julkowska, D.; Obuchowski, M.; Holland, I.B.; Séror, S.J. Comparative analysis of the development of swarming communities of Bacillus subtilis 168 and a natural wild type: critical effects of surfactin and the composition of the medium. J Bacteriol. 2005, 187, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Anagnostopoulos, C.; Spizizen, J. Requirements for transformation in Bacillus subtilis. J Bacteriol. 1961, 81, 741–746. [Google Scholar] [CrossRef]
- Isticato, R.; Cangiano, G.; De Felice, M.; Ricca, E. Display of Molecules on the Spore Surface. In Bacterial Spore Formers. Edited by Ricca, E.; Henriques, A.O.; Cutting, S.M. Norfolk, UK: Horizon Biosience. 2004, 193–200.
- Nicholson, W.L.; Setlow, P. Sporulation, germination and outgrowth. In Molecular biological methods for Bacillus. Edited by Harwood C, Cutting S. Chichester, United Kingdom: Wiley, 1990:391–450.
Figure 1.
Structure of CotX and its derivatives. A – putative three-dimensional structure of CotX protein obtained from I-TASSER (left) and AlphaFold server (right, AF-Q08313-F1-model_v4); B – predicted consensus of secondary structure of CotX, C – deletion mutants of CotX used in the study. Colors used at panel A and B represent segments which were deleted during derivative CotX construction.
Figure 1.
Structure of CotX and its derivatives. A – putative three-dimensional structure of CotX protein obtained from I-TASSER (left) and AlphaFold server (right, AF-Q08313-F1-model_v4); B – predicted consensus of secondary structure of CotX, C – deletion mutants of CotX used in the study. Colors used at panel A and B represent segments which were deleted during derivative CotX construction.
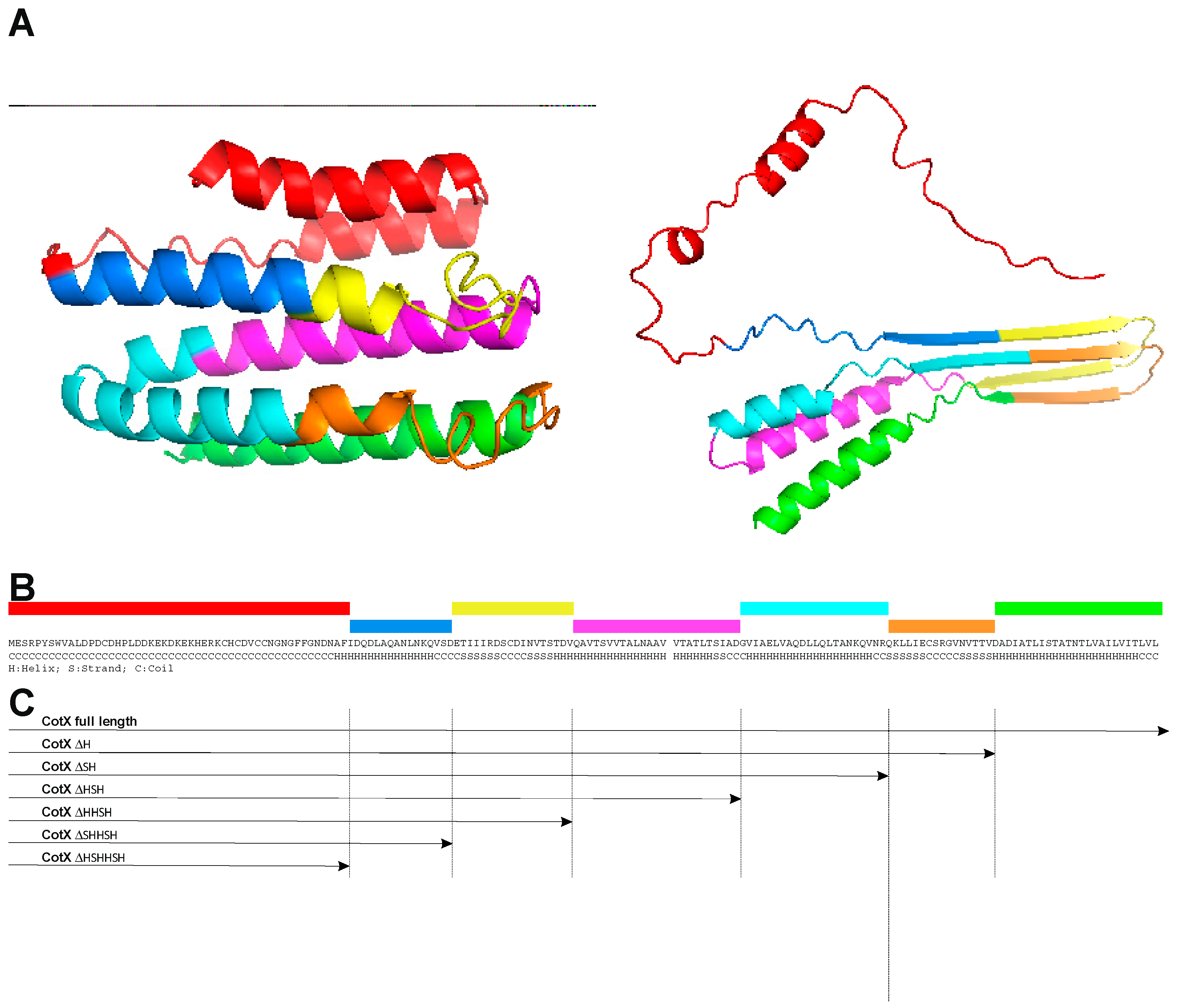
Figure 4.
Western blotting detection of CotX-mCherry and its derivatives overexpressed from pHT08 vector in B. subtilis 168 genetic background. Lines: 1 – prestainded protein ladder; 2 – protein extract from B. subtilis 168; 3 - strain BWP 46 (B. s. 168 pHT08 cotX-mCherry without IPTG induction); lines 4-11 contain protein extract from strains carrying derivatives of cotX-mCherry fusion protein induced for expression by 1mM IPTG. Line 4 - strain BWP 46 (cotX-mCherry); 5 - strain BWP 48 (cotX ΔH-mCherry); 6- strain BWP 50 (cotX ΔSH -mCherry); 7 - strain BWP 52 (cotX ΔHSH -mCherry); 8- strain BWP 54 (cotX ΔHHSH -mCherry); 9 - strain BWP 56 (cotX ΔSHHSH -mCherry); 10 - strain BWP 58 (cotX ΔHSHHSH -mCherry).
Figure 4.
Western blotting detection of CotX-mCherry and its derivatives overexpressed from pHT08 vector in B. subtilis 168 genetic background. Lines: 1 – prestainded protein ladder; 2 – protein extract from B. subtilis 168; 3 - strain BWP 46 (B. s. 168 pHT08 cotX-mCherry without IPTG induction); lines 4-11 contain protein extract from strains carrying derivatives of cotX-mCherry fusion protein induced for expression by 1mM IPTG. Line 4 - strain BWP 46 (cotX-mCherry); 5 - strain BWP 48 (cotX ΔH-mCherry); 6- strain BWP 50 (cotX ΔSH -mCherry); 7 - strain BWP 52 (cotX ΔHSH -mCherry); 8- strain BWP 54 (cotX ΔHHSH -mCherry); 9 - strain BWP 56 (cotX ΔSHHSH -mCherry); 10 - strain BWP 58 (cotX ΔHSHHSH -mCherry).
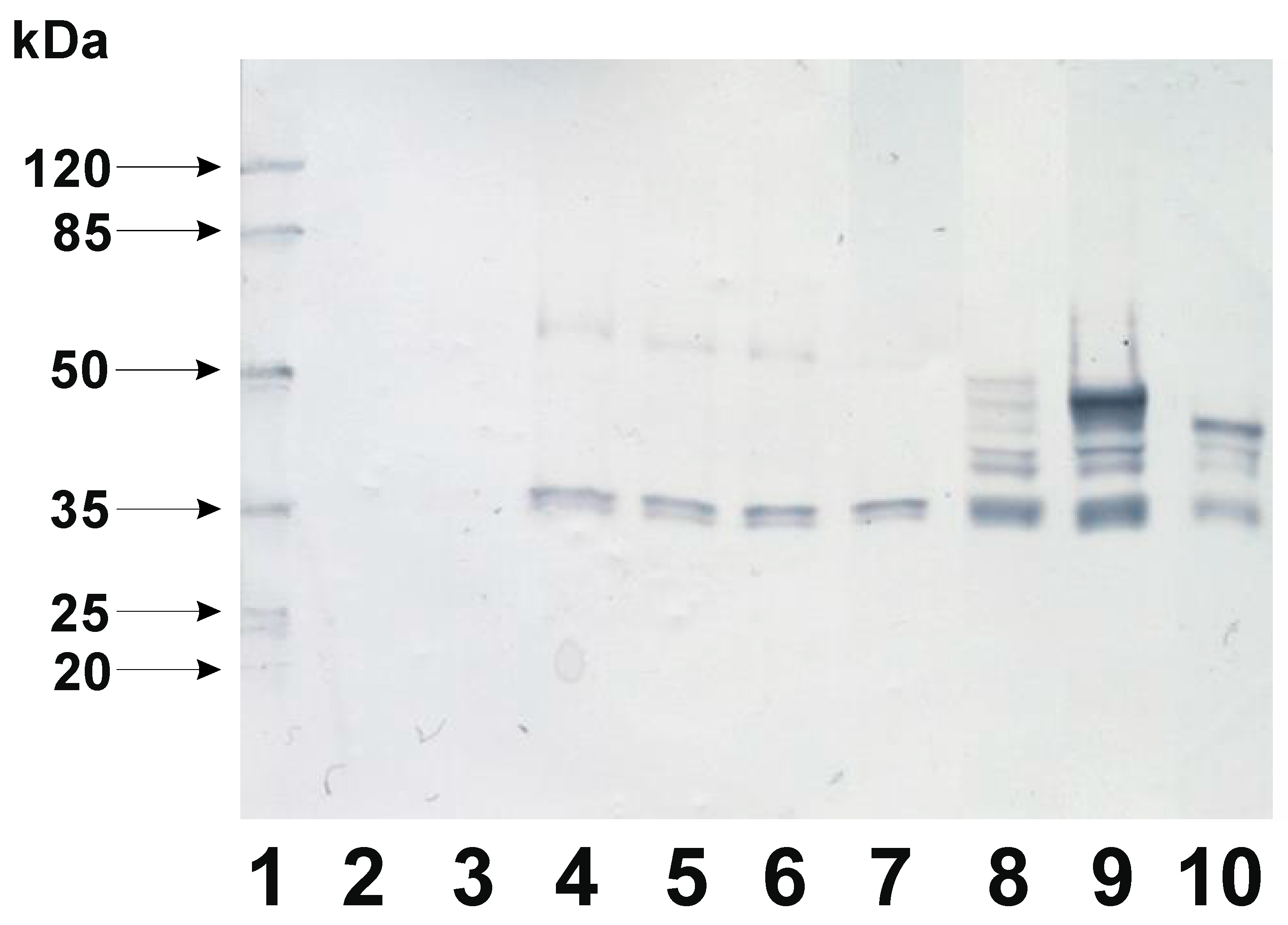

Table 1.
Strain and plasmid list.
| Strain | Relevant genotype | Source |
| 168 | trpC2 | [6] |
| BWP 46 | 168 pWP02 (cotX-mCherry) | this work |
| BWP 48 | 168 pWP04 (cotX ΔH-mCherry) | this work |
| BWP 50 | 168 pWP06 (cotX ΔSH -mCherry) | this work |
| BWP 52 | 168 pWP08 (cotX ΔHSH -mCherry) | this work |
| BWP 54 | 168 pWP010 (cotX ΔHHSH -mCherry) | this work |
| BWP 56 | 168 pWP012 (cotX ΔSHHSH -mCherry) | this work |
| BWP 58 | 168 pWP014 (cotX ΔHSHHSH -mCherry) | this work |
| BWP 53 | 168 amyE:: cotX-mCherry CmR | this work |
| BWP 55 | 168 amyE:: cotX ΔSH-mCherry CmR | this work |
| BWP 57 | 168 amyE:: cotX ΔHHSH-mCherry CmR | this work |
| BWP59 | 168 amyE:: cotX ΔHSHHSH-mCherry CmR | this work |
| Plasmid | ||
| pHT08 | IPTG inducible expression vector | MoBitec (Germany) |
| pDL | shuttle vector | Bacillus Genetic Stock Center |
| pWP01 | pDL, cotX-mCherry | this work |
| pWP02 | pHT08, cotX-mCherry | this work |
| pWP04 | pHT08, cotX ΔH-mCherry | this work |
| pWP05 | pDL, cotX ΔSH -mCherry | this work |
| pWP06 | pHT08, cotX ΔSH -mCherry | this work |
| pWP08 | pHT08, cotX ΔHSH -mCherry | this work |
| pWP09 | pDL, cotX ΔHHSH -mCherry | this work |
| pWP010 | pHT08, cotX ΔHHSH -mCherry | this work |
| pWP012 | pHT08, cotX ΔSHHSH -mCherry | this work |
| pWP013 | pDL, cotX ΔHSHHSH -mCherry | this work |
| pWP014 | pHT08, cotX ΔHSHHSH -mCherry | this work |
Table 2.
Primers used for construction mutant CotX protein.
| Forward primer | |
| Full-length CotX | 5' CAGCGGCCGCAATCACGTGGGTGGAGGAG 3' |
| mCherry | 5' CGGATCCATGGTTTCAAAAGGCG 3' |
| Reverse primers | |
| Full-length CotX | 5' CTATCTAGATTAGAGGACAAGAGTGATAAC 3' |
| CotX dH | 5' CTATCTAGATTATACTGTTGTGACGTTTAC 3' |
| CotX dSH | 5' CTATCTAGATTAGCGGTTTACTTGTTTGTT 3' |
| CotX dHSH | 5' CTATCTAGATTAGTCTGCAATTGATGTCAG 3' |
| CotX dHHSH | 5' CTATCTAGATTAGTCTGTAGATGTAACATTG 3' |
| CotX dSHHSH | 5' CTATCTAGATTATGAAACTTGTTTGTTGAG 3' |
| CotX dHSHHSH | 5' CTATCTAGATTAGAAGGCGTTGTCGTTGCC 3' |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



