
Submitted:
23 December 2024
Posted:
26 December 2024
You are already at the latest version
Abstract
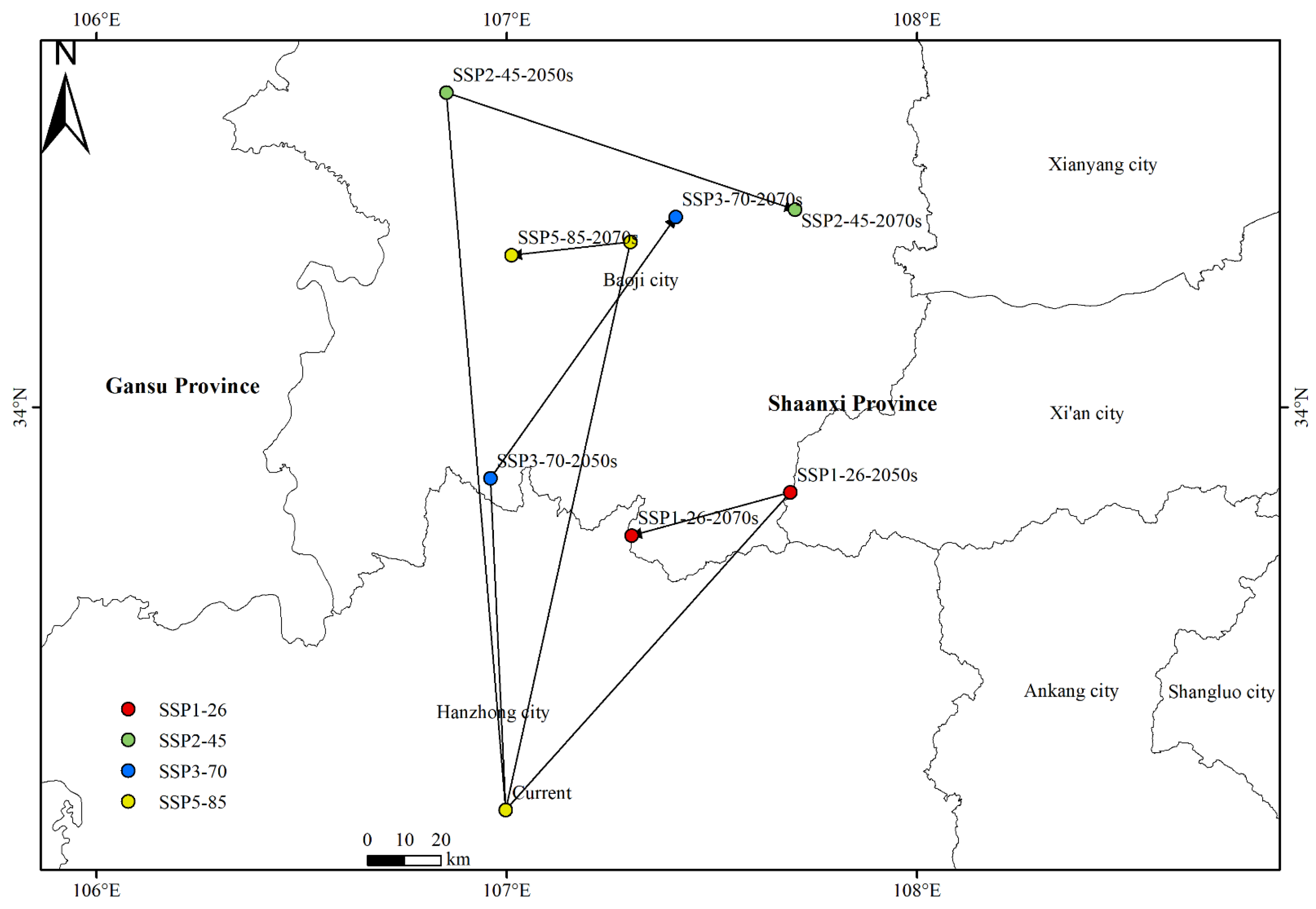
Xanthium spinosum (X. spinosum) is a highly invasive weed native to South America and distributed in 17 provinces (municipalities) of China. It has severe negative influences on ecosystems, agriculture, and husbandry. However, few studies have reported on the impact of human activity and climate change on the future distribution and centroid shift of X. spinosum. This study aimed to investigate the potential geological distribution of X. spinosum in China, as well as the distribution pattern, centroid shift, and key environmental factors influencing its distribution, under four future climate scenarios (SSP1-26, SSP2-45, SSP3-70, and SSP5-85) based on the biomod2 integrated model. The results indicated that the suitable habitats for X. spinosum would expand in the future, mainly in Inner Mongolia, Northeast China, and the plateau regions (e.g., Xinjiang and Xizang). Under future climate scenarios, the centroid would shift toward the northwest or northeast part of China, with the SSP2-45-2050s scenario showing the maximum shift distance (161.990 km). Additionally, the key environmental variables influencing the distribution of X. spinosum, including human impact index, bio5, bio7, and bio12, were determined, revealing that most of them were related to human activities, temperature, and precipitation. This study enhanced the understanding of the influence of human activity and climate change on the geographic range of X. spinosum. It provided references for early warning and management in the control of X. spinosum.
Keywords:
1. Introduction
2. Results
2.1. Evaluation of Model Accuracy
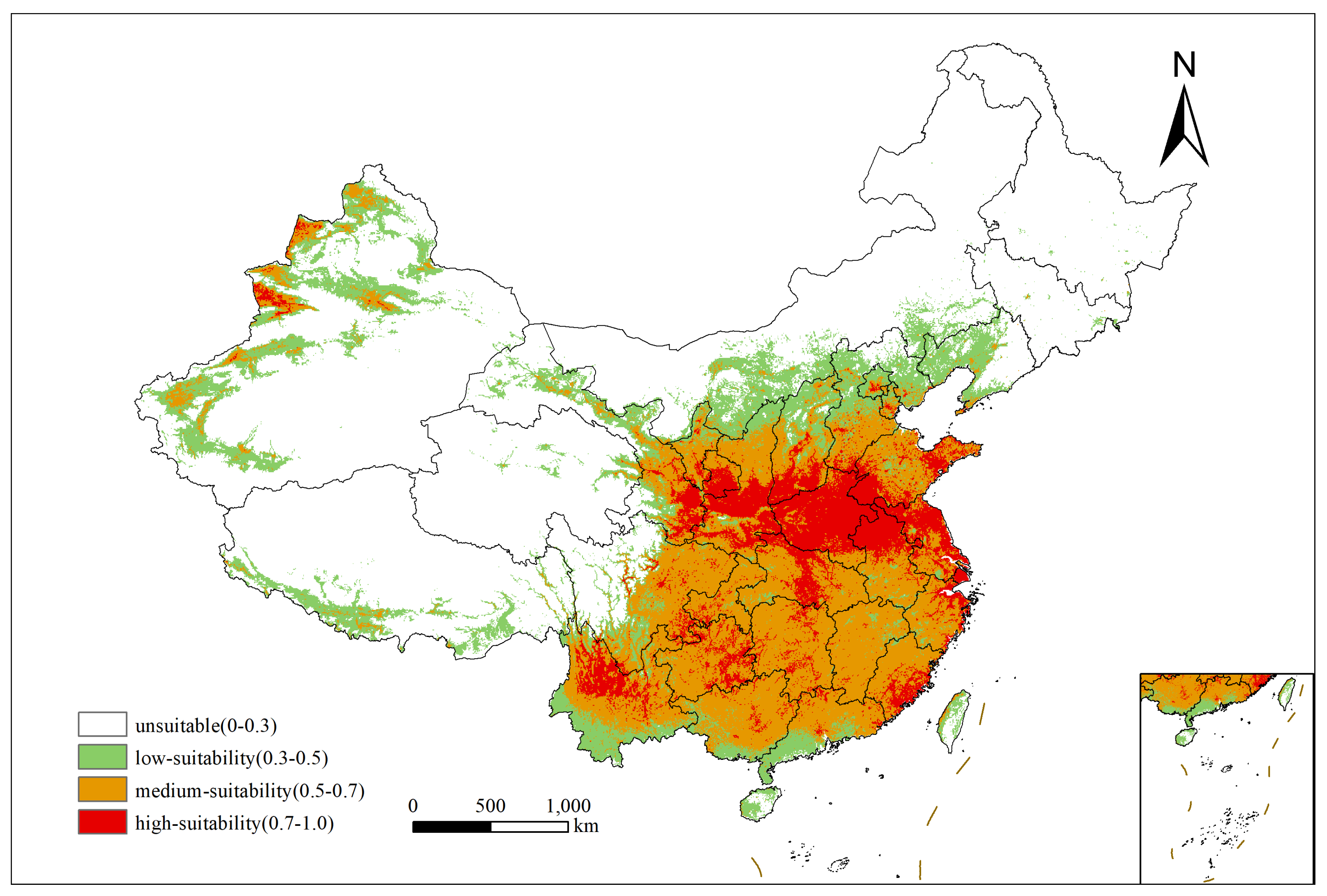
2.2. Current Distribution of the Suitable Habitats for Xanthium spinosum L. in China
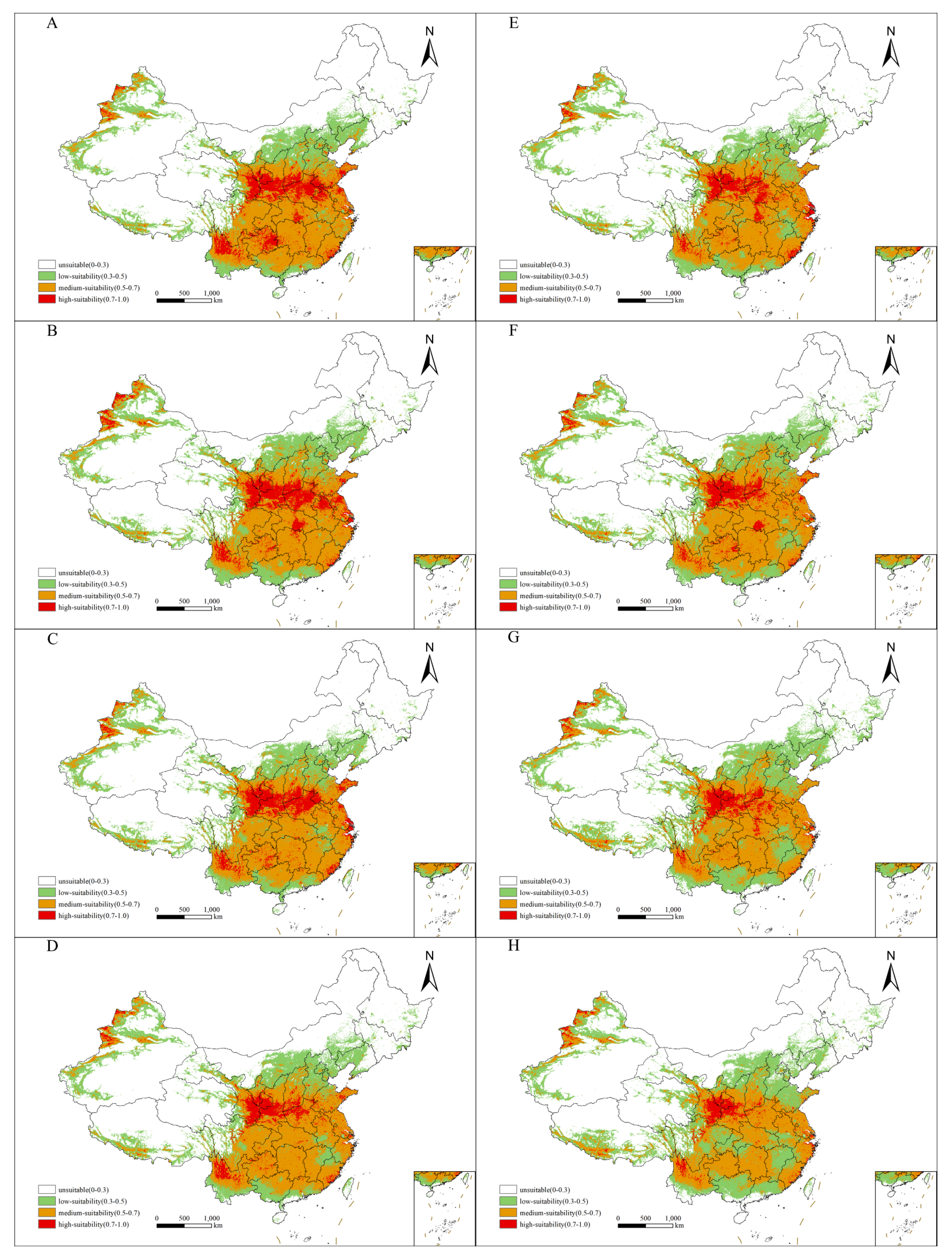
2.3. Future Distribution of Suitable Habitats for Xanthium spinosum L. in China
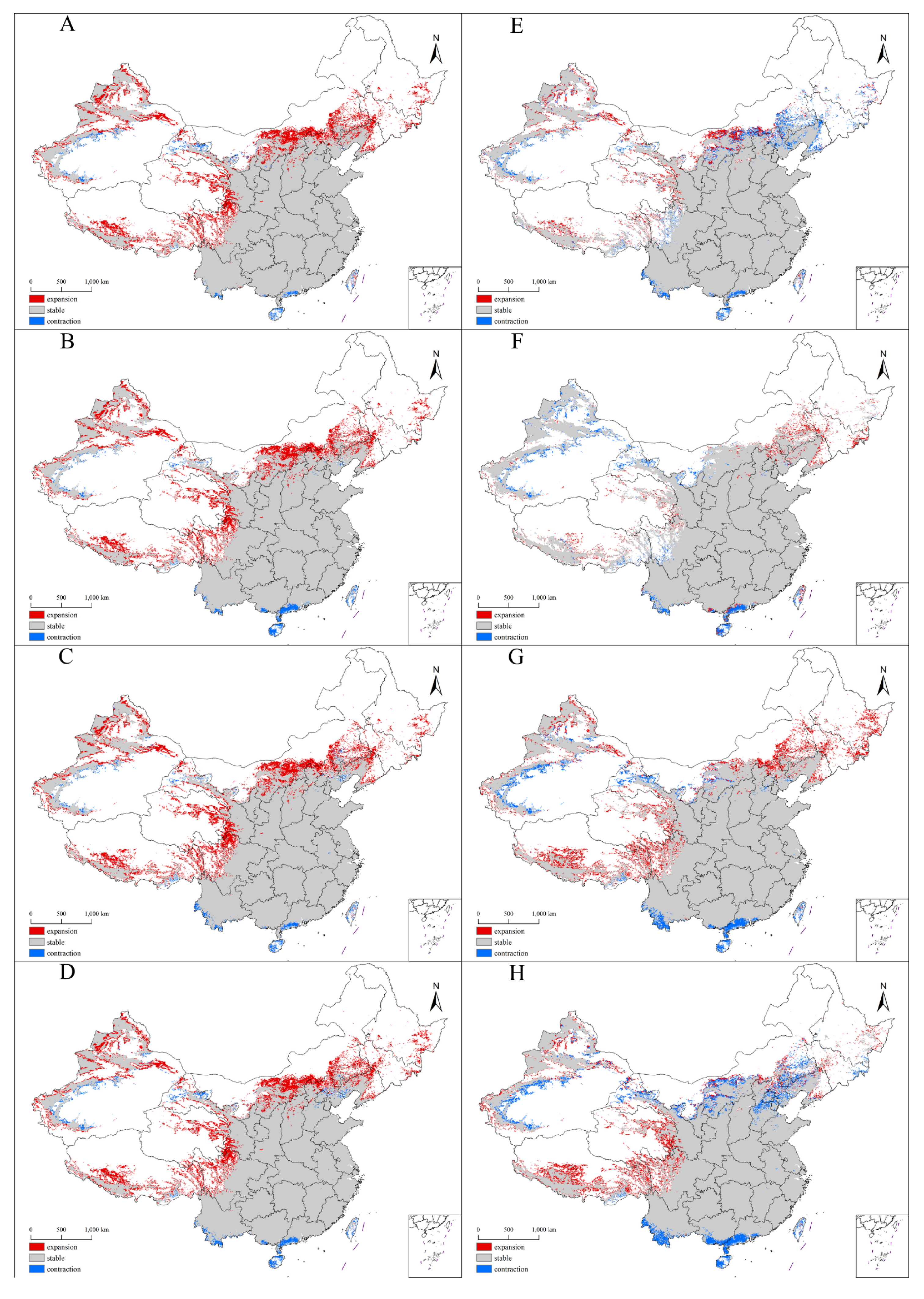
2.4. Future Distribution Patterns of the Suitable Habitats for Xanthium spinosum L. in China
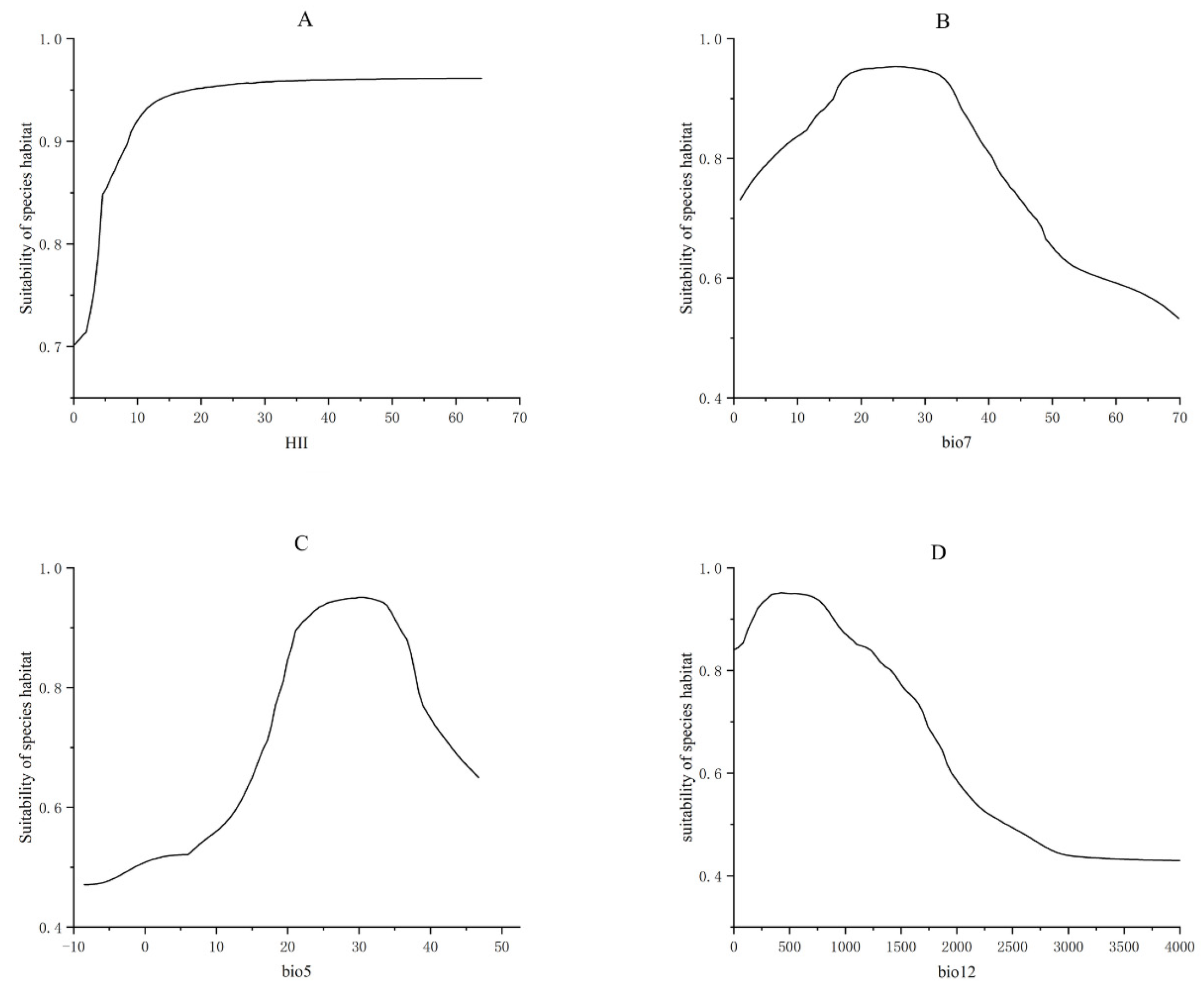
2.5. Analysis of the Environmental Factors Influencing the Distribution of Xanthium spinosum L.
3. Discussion
3.1. Assessment of Biomod2 Model Prediction Performance
3.2. Change Pattern and Shift Direction of the Suitable Habitats for Xanthium spinosum L. under Future Climate Change
3.3. Human Activities and Climatic Factors Influencing the Distribution of Xanthium spinosum L.
4. Materials and Methods
4.1. Data Collection and Processing for the Distribution of Xanthium spinosum L.
4.2. Preprocessing of Environmental Data
4.3. Model Prediction and Evaluation for the Suitable Habitats for Xanthium spinosum L.
4.4. Distribution and Area of The suitable Habitats for Xanthium spinosum L.
4.5. Changes in the Spatial Patterns of the Suitable Habitats for Xanthium spinosum L.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cheng, X.; Xu, R. Overview of exotic animal invasions in China. Bulletin of Biology 2007, 1–4, 64. [Google Scholar] [CrossRef]
- Jones, B.A.; McDermott, S.M. Health Impacts of Invasive Species through an Altered Natural Environment: Assessing Air Pollution Sinks as a Causal Pathway. Environmental and Resource Economics 2018, 71, 23–43. [Google Scholar] [CrossRef]
- Alley, R.B.; Berntsen, T.; Bindoff, N.L.; Chidthaisong, A.; Friedlingstein, P.; Gregory, J.M.; Hegerl, G.C.; Heimann, M.; Hewitson, B.; Hoskins, B.J. Summary for Policymakers. In Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L., Eds.; 2007. [Google Scholar]
- Canavan, S.; Richardson, D.M.; Visser, V.; Le Roux, J.J.; Vorontsova, M.S.; Wilson, J.R. The Global Distribution of Bamboos: Assessing Correlates of Introduction and Invasion. AoB Plants 2017, 9, plw078. [Google Scholar] [CrossRef]
- Clements, D.R.; Jones, V.L. Rapid Evolution of Invasive Weeds Under Climate Change: Present Evidence and Future Research Needs. Front. Agron. 2021, 3. [Google Scholar] [CrossRef]
- Jarvis, C. Order Out of Chaos. Linnaean Plant Types and Their Types; Linnaean Society of London in association with the Natural History Museum; 2017. [Google Scholar]
- Löve, D.; Dansereau, P. Biosystematic Studies on Xanthium: Taxonomic Appraisal and Ecological Status. Canadian Journal of Botany 1959, 37, 173–208. [Google Scholar] [CrossRef]
- Holm, L.G.; Plucknett, D.L.; Pancho, J.V.; Herberger, J.P. The World’s Worst Weeds. Distribution and Biology; 1977; ISBN 0-8248-0295-0. [Google Scholar]
- Chen, Y.; Chen, S. A new synonym of Xanthium sibiricum Patrin ex Widder. Journal of Systematics and Evolution 2004, 42, 191. [Google Scholar]
- Zhao, L.; Zang, C.; Yang, J. Distribution of a Invasive Species-Xanthium spinosum L. in Inner Mongolia and Ning Xia. Journal of Inner Mongolia University (Natural Science Edition) 2006, 308–310. [Google Scholar]
- Du, Z.; Xu, W.; Yan, P.; Wang, S.; Guo, Y. Three Newly Recorded Alien Invasive Plants of Xanthium in Xinjiang. Xinjiang Agricultural Sciences 2012, 49, 879–886. [Google Scholar]
- Song, X.; He, J. Status and Prospect of Research on Alien Invasive Plant Xanthium spinosum L. Journal of Weed Science 2023, 41, 1–7. [Google Scholar] [CrossRef]
- Li, J.; Ma, M. Seed overwinter performance of two invasive plants: Xanthium italicum Moretti and X. spinosum Linnaeus in Xinjiang. Acta Ecologica Sinica 2017, 37, 7181–7186. [Google Scholar] [CrossRef]
- Gu, W.; Ma, M. Study on reproductive biology characteristics of invasive plant Xanthium spinosum L. Journal of Shihezi University(Natural Science) 2019, 37, 332–338. [Google Scholar] [CrossRef]
- Liang, Q.; Liu, Z.; Lu, P.; Chen, W. Distribution and Growth Characteristics of Xanthium spinosum in Yili River Valley of Xinjiang. Journal of Weed Science 2017, 35, 25–29. [Google Scholar] [CrossRef]
- Dong, F.; Leng, J.; Liu, Y.; Yu, W.; Zhao, Y. Allelopathic Effects of Aqueous Extracts from Xanthium Spinosum on Seed Germination and Seedling Growth of Lactuca Sativa Var. Longifolia. Acta Prataculturae Sinica 2017, 26, 146–160. [Google Scholar]
- Yuan, Z.N.; Liu, Y.; Shao, H.; Zhao, Y.; Hu, Y. Allelopathy of Each Part of Invasive Plant Xanthium spinosum L. Journal of Henan Agricultural Sciences 2017, 46, 73–77. [Google Scholar] [CrossRef]
- Xanthium Spinosum (Bathurst Burr). CABI Compendium 2022.
- Lang, Q. Population Distribution and Ecological Characteristics of Xanthium spinosum L. in Yili Region. Rural Practical Science & Technology 2020, 78–79. [Google Scholar]
- Tao, Y.; Zhao, Y.; Hu, Y.; Shang, T.; Zhang, Z.; Lang, Q.; Liu, Y. Effects of simulated nitrogen deposition on the growth of Xanthium spinosum. Chinese Journal of Ecology 2020, 39, 3971–3978. [Google Scholar] [CrossRef]
- Xiao, Y.; He, J.; Aishan, T.; Sui, X.; Zhou, Y.; Yimingniyazi, A. Effects of Different Degrees of Xanthium Spinosum Invasion on the Invasibility of Plant Communities in the Yili Grassland of Northwest China. Biology 2023, 13, 14. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species Distribution Models: Ecological Explanation and Prediction across Space and Time. Annual review of ecology, evolution, and systematics 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Beery, S.; Cole, E.; Parker, J.; Perona, P.; Winner, K. Species Distribution Modeling for Machine Learning Practitioners: A Review. In Proceedings of the Proceedings of the 4th ACM SIGCAS Conference on Computing and Sustainable Societies; Association for Computing Machinery: Virtual Event, Australia, 2021; pp. 329–348. [Google Scholar]
- Brambilla, M.; Ficetola, G.F. Species Distribution Models as a Tool to Estimate Reproductive Parameters: A Case Study with a Passerine Bird Species. Journal of Animal Ecology 2012, 81, 781–787. [Google Scholar] [CrossRef] [PubMed]
- Moraitis, M.L.; Valavanis, V.D.; Karakassis, I. Modelling the Effects of Climate Change on the Distribution of Benthic Indicator Species in the Eastern Mediterranean Sea. Science of The Total Environment 2019, 667, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Xiao, N.; Shen, M.; Li, J. Comparison between Optimized MaxEnt and Random Forest Modeling in Predicting Potential Distribution: A Case Study with Quasipaa Boulengeri in China. Science of The Total Environment 2022, 842, 156867. [Google Scholar] [CrossRef]
- Nelder, J.A.; Wedderburn, R.W. Generalized Linear Models. Journal of the Royal Statistical Society Series A: Statistics in Society 1972, 135, 370–384. [Google Scholar] [CrossRef]
- Ho, T.K. Random Decision Forests. In Proceedings of the Proceedings of 3rd International Conference on Document Analysis and Recognition; IEEE, 1995; Volume 1, pp. 278–282. [Google Scholar]
- Bajkova, A.T. The Generalization of Maximum Entropy Method for Reconstruction of Complex Functions. Astronomical and Astrophysical Transactions 1992, 1, 313–320. [Google Scholar] [CrossRef]
- Segurado, P.; Araujo, M.B. An Evaluation of Methods for Modelling Species Distributions. Journal of biogeography 2004, 31, 1555–1568. [Google Scholar] [CrossRef]
- Araújo, M.B.; New, M. Ensemble Forecasting of Species Distributions. Trends in ecology & evolution 2007, 22, 42–47. [Google Scholar] [CrossRef]
- Thuiller, W. BIOMOD–Optimizing Predictions of Species Distributions and Projecting Potential Future Shifts under Global Change. Global change biology 2003, 9, 1353–1362. [Google Scholar] [CrossRef]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M.B. BIOMOD–a Platform for Ensemble Forecasting of Species Distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Guan, B.; Guo, H.; Chen, S.; Li, D.; Liu, X.; Gong, X.I.; Ge, G. Shifting Ranges of Eleven Invasive Alien Plants in China in the Face of Climate Change. Ecological Informatics 2020, 55, 101024. [Google Scholar] [CrossRef]
- Jia, T.; Qi, Y.; Zhao, H.; Xian, X.; Li, J.; Huang, H.; Yu, W.; Liu, W. Estimation of Climate-Induced Increased Risk of Centaurea Solstitialis L. Invasion in China: An Integrated Study Based on Biomod2. Frontiers in Ecology and Evolution 2023, 11, 1113474. [Google Scholar] [CrossRef]
- Xu, L.; Fan, Y.; Zheng, J.; Guan, J.; Lin, J.; Wu, J.; Liu, L.; Wu, R.; Liu, Y. Impacts of Climate Change and Human Activity on the Potential Distribution of Aconitum Leucostomum in China. Science of The Total Environment 2024, 912, 168829. [Google Scholar] [CrossRef] [PubMed]
- Qiao, H.; Hu, J.; Huang, J. Theoretical Basis, Future Directions, and Challenges for Ecological Niche Models. Scientia Sinica(Vitae) 2013, 43, 915–927. [Google Scholar] [CrossRef]
- Tanaka, K.R.; Torre, M.P.; Saba, V.S.; Stock, C.A.; Chen, Y. An Ensemble High-resolution Projection of Changes in the Future Habitat of American Lobster and Sea Scallop in the Northeast US Continental Shelf. Diversity and Distributions 2020, 26, 987–1001. [Google Scholar] [CrossRef]
- Zhao, Z.; Wei, H.; Guo, Y.; Gu, W. Potential distribution of Panax ginseng and its predicted responses to climate change. Chinese Journal of Applied Ecology 2016, 27, 3607–3615. [Google Scholar] [CrossRef] [PubMed]
- Zhu, N. Modelling the Suitable Habitat Distribution of Magnolia Officinalis Using Ensemble Model. Journal of Sichuan Agricultural University 2019, 37, 481–489. [Google Scholar] [CrossRef]
- McGeoch, M.A.; Butchart, S.H.; Spear, D.; Marais, E.; Kleynhans, E.J.; Symes, A.; Chanson, J.; Hoffmann, M. Global Indicators of Biological Invasion: Species Numbers, Biodiversity Impact and Policy Responses. Diversity and Distributions 2010, 16, 95–108. [Google Scholar] [CrossRef]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of Climate Change on the Future of Biodiversity. Ecology letters 2012, 15, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Díaz, S.M.; Settele, J.; Brondízio, E.; Ngo, H.; Guèze, M.; Agard, J.; Arneth, A.; Balvanera, P.; Brauman, K.; Butchart, S. The Global Assessment Report on Biodiversity and Ecosystem Services: Summary for Policy Makers. 2019. [Google Scholar]
- Bellard, C.; Thuiller, W.; Leroy, B.; Genovesi, P.; Bakkenes, M.; Courchamp, F. Will Climate Change Promote Future Invasions? Global change biology 2013, 19, 3740–3748. [Google Scholar] [CrossRef]
- Hamit, S.; Abdushalih, N.; Xu, Z.; Arman, J.; Shao, H.; Yilihar, V. Simulation of potential distribution patterns of the invasive plant species Xanthium spinosum L. (Bathurst burr) in Xinjiang under climate change. Acta Ecologica Sinica 2019, 39, 1551–1559. [Google Scholar] [CrossRef]
- BenDor, T.K.; Metcalf, S.S. The Spatial Dynamics of Invasive Species Spread. System Dynamics Review: The Journal of the System Dynamics Society 2006, 22, 27–50. [Google Scholar] [CrossRef]
- BenDor, T.K.; Metcalf, S.S.; Fontenot, L.E.; Sangunett, B.; Hannon, B. Modeling the Spread of the Emerald Ash Borer. Ecological modelling 2006, 197, 221–236. [Google Scholar] [CrossRef]
- McDermott, S.M.; Finnoff, D.C. Impact of Repeated Human Introductions and the Allee Effect on Invasive Species Spread. Ecological Modelling 2016, 329, 100–111. [Google Scholar] [CrossRef]
- Mairal, M.; Chown, S.L.; Shaw, J.; Chala, D.; Chau, J.H.; Hui, C.; Kalwij, J.M.; Münzbergová, Z.; Jansen van Vuuren, B.; Le Roux, J.J. Human Activity Strongly Influences Genetic Dynamics of the Most Widespread Invasive Plant in the sub-Antarctic. Molecular Ecology 2022, 31, 1649–1665. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.P. A Framework for Using Niche Models to Estimate Impacts of Climate Change on Species Distributions. Annals of the New York Academy of Sciences 2013, 1297, 8–28. [Google Scholar] [CrossRef]
- Marzloff, M.P.; Oliver, E.C.; Barrett, N.S.; Holbrook, N.J.; James, L.; Wotherspoon, S.J.; Johnson, C.R. Differential Vulnerability to Climate Change Yields Novel Deep-Reef Communities. Nature Climate Change 2018, 8, 873–878. [Google Scholar] [CrossRef]
- Lu, X.; Siemann, E.; Shao, X.; Wei, H.; Ding, J. Climate Warming Affects Biological Invasions by Shifting Interactions of Plants and Herbivores. Global change biology 2013, 19, 2339–2347. [Google Scholar] [CrossRef]
- Gao, C.; Guo, S.; Ma, C.; Yang, J.; Kang, X.; Li, R. Impact of Climate Change on the Potential Geographical Distribution Patterns of Luculia Pinceana Hook. f. since the Last Glacial Maximum. Forests 2024, 15, 253. [Google Scholar] [CrossRef]
- Gu, R.; Wei, S.; Li, J.; Zheng, S.; Li, Z.; Liu, G.; Fan, S. Predicting the Impacts of Climate Change on the Geographic Distribution of Moso Bamboo in China Based on Biomod2 Model. Eur J Forest Res 2024, 143, 1499–1512. [Google Scholar] [CrossRef]
- Larson, C.D.; Pollnac, F.W.; Schmitz, K.; Rew, L.J. Climate Change and Micro-Topography Are Facilitating the Mountain Invasion by a Non-Native Perennial Plant Species. NeoBiota 2021, 65, 23–45. [Google Scholar] [CrossRef]
- Wiens, J.J.; Graham, C.H. Niche Conservatism: Integrating Evolution, Ecology, and Conservation Biology. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 519–539. [Google Scholar] [CrossRef]
- Li, K.; Liu, Z.; Yan, B.; Guo, J. A multi-factor comprehensive analysis of the distribution and development potentiality of geothermal resource in Baoji region. Journalof Baoji University of Arts and Sciences (Natural Science Edition) 2022, 42, 86–92. [Google Scholar] [CrossRef]
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.-C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengård, B. Biodiversity Redistribution under Climate Change: Impacts on Ecosystems and Human Well-Being. Science 2017, 355, eaai9214. [Google Scholar] [CrossRef] [PubMed]
- Lembrechts, J.J.; Lenoir, J.; Nuñez, M.A.; Pauchard, A.; Geron, C.; Bussé, G.; Milbau, A.; Nijs, I. Microclimate Variability in Alpine Ecosystems as Stepping Stones for Non-native Plant Establishment above Their Current Elevational Limit. Ecography 2018, 41, 900–909. [Google Scholar] [CrossRef]
- Sun, H.; Wang, X.; Fan, Y.; Liu, C.; Wu, P.; Li, Q.; Yin, W. Effects of Biophysical Constraints, Climate and Phylogeny on Forest Shrub Allometries along an Altitudinal Gradient in Northeast China. Scientific Reports 2017, 7, 43769. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Li, S. Modern Coexistence Theory as a Framework for Invasion Ecology. Biodiversity Science 2020, 28, 1362. [Google Scholar] [CrossRef]
- Zhao, G.; Cui, X.; Sun, J.; Li, T.; Wang, Q.I.; Ye, X.; Fan, B. Analysis of the Distribution Pattern of Chinese Ziziphus Jujuba under Climate Change Based on Optimized Biomod2 and MaxEnt Models. Ecological Indicators 2021, 132, 108256. [Google Scholar] [CrossRef]
- Essl, F.; Biró, K.; Brandes, D.; Broennimann, O.; Bullock, J.M.; Chapman, D.S.; Chauvel, B.; Dullinger, S.; Fumanal, B.; Guisan, A. Biological Flora of the British Isles: Ambrosia Artemisiifolia. Journal of Ecology 2015, 103, 1069–1098. [Google Scholar] [CrossRef]
- Kueffer, C. Plant Invasions in the Anthropocene. Science 2017, 358, 724–725. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Li, Y.; Wu, M.; Zhang, Z.H.E.; Li, L.; Wan, S. Plant Community Responses to Nitrogen Addition and Increased Precipitation: The Importance of Water Availability and Species Traits. Global Change Biology 2011, 17, 2936–2944. [Google Scholar] [CrossRef]
- Lu, Z.; Yang, H.; Du, W.; Li, Z.; Sun, F.; Yang, J.; Chen, C.; Gao, Y.; Wang, S.; Wu, P. Preliminary Study on Control Effects of Five Herbicides on Invasive Species of Xanthium sibiricum in Ningxia. Chinese Agricultural Science Bulletin 2024, 40, 129–132. [Google Scholar] [CrossRef]
- Feijó, A.; Ge, D.Y.; Wen, Z.X.; Cheng, J.L.; Xia, L.; Yang, Q.S. Exploring GBIF Database and Extracting Climate Data from Georeferenced Localities with R Software. Bio-101 2021, e1010609. [Google Scholar] [CrossRef]
- Qin, Y.; Wang, C.; Zhao, Z.; Pan, X.; Li, Z. Climate Change Impacts on the Global Potential Geographical Distribution of the Agricultural Invasive Pest, Bactrocera Dorsalis (Hendel)(Diptera: Tephritidae). Climatic Change 2019, 155, 145–156. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, T.; Wu, Y.; Hu, R.; Huang, K.; Shao, X. Past Distribution of Epiphyllous Liverworts in China: The Usability of Historical Data. Ecology and Evolution 2018, 8, 7436–7450. [Google Scholar] [CrossRef]
- Sharma, R.; Khan, S.; Kaul, V. Predicting the Potential Habitat Suitability and Distribution of “Weed-Onion”(Asphodelus Tenuifolius Cavan.) in India under Predicted Climate Change Scenarios. Journal of Agriculture and Food Research 2023, 14, 100697. [Google Scholar] [CrossRef]
- Qin, Z.; Yang, J.H.; Gan, T.; Zhang, J.E.; Liu, Y.M.; Liu, J.M.; Yao, F.C.; Zhao, B.L. Prediction of the Breeding and Wintering Ranges of Pomacea Canaliculata in China Using Ensemble Models. Journal of Environmental Informatics 2024, 44. [Google Scholar] [CrossRef]
- Zhang, Y.; Na, X.; Li, W. Impacts of Climate Changes on the Potential Habitat Suitability of Grus Japonensis on Migration Routes. Ecological Indicators 2024, 166, 112462. [Google Scholar] [CrossRef]
- Barbet-Massin, M.; Jiguet, F.; Albert, C.H.; Thuiller, W. Selecting Pseudo-absences for Species Distribution Models: How, Where and How Many? Methods in ecology and evolution 2012, 3, 327–338. [Google Scholar] [CrossRef]
- Xian, X.; Zhao, H.; Wang, R.; Huang, H.; Chen, B.; Zhang, G.; Liu, W.; Wan, F. Climate Change Has Increased the Global Threats Posed by Three Ragweeds (Ambrosia L.) in the Anthropocene. Science of The Total Environment 2023, 859, 160252. [Google Scholar] [CrossRef]
- Fielding, A.H.; Bell, J.F. A Review of Methods for the Assessment of Prediction Errors in Conservation Presence/Absence Models. Environmental conservation 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the Accuracy of Species Distribution Models: Prevalence, Kappa and the True Skill Statistic (TSS). Journal of applied ecology 2006, 43, 1223–1232. [Google Scholar] [CrossRef]

| Models | TSS | AUC | |||||
|---|---|---|---|---|---|---|---|
| Mean | SD | CV | Mean | SD | CV | ||
| ANN | 0.7798 | 0.0132 | 0.0170 | 0.9471 | 0.0057 | 0.0060 | |
| CTA | 0.8455 | 0.0301 | 0.0356 | 0.9528 | 0.0105 | 0.0110 | |
| FDA | 0.7812 | 0.0079 | 0.0102 | 0.9516 | 0.0035 | 0.0037 | |
| GAM | 0.6821 | 0.0092 | 0.0135 | 0.9200 | 0.0039 | 0.0043 | |
| GLM | 0.7858 | 0.0075 | 0.0096 | 0.9571 | 0.0016 | 0.0017 | |
| MARS | 0.7930 | 0.0070 | 0.0089 | 0.9574 | 0.0023 | 0.0024 | |
| RF | 0.9861 | 0.0044 | 0.0044 | 0.9999 | 0.0003 | 0.0003 | |
| SRE | 0.5680 | 0.0052 | 0.0092 | 0.7841 | 0.0028 | 0.0035 | |
| Climate Scenario | Period | High-Suitability Habitat | Medium-Suitability Habitat | Low-Suitability Habitat | Total Suitable Habitat |
|---|---|---|---|---|---|
| Current | 1970–2000 | 77.8056 | 208.6410 | 128.2930 | 414.7396 |
| SSP-1-26 | 2050s | 60.3160 | 227.5660 | 161.5590 | 449.4410 |
| 2070s | 41.0694 | 220.1440 | 174.2380 | 435.4514 | |
| SSP-2-45 | 2050s | 61.2865 | 230.8140 | 174.4440 | 466.5445 |
| 2070s | 42.5226 | 243.8400 | 185.0760 | 471.4386 | |
| SSP-3-70 | 2050s | 53.9792 | 231.4700 | 171.8660 | 457.3152 |
| 2070s | 37.2483 | 226.3350 | 219.1390 | 482.7223 | |
| SSP-5-85 | 2050s | 40.1059 | 238.7590 | 179.2450 | 458.1099 |
| 2070s | 26.5990 | 207.4980 | 224.1910 | 458.2880 |
| Climate Scenario | Area (×104 km2) | Change (%) | |||||
|---|---|---|---|---|---|---|---|
| Expansion | Unchanged | Contraction | Expansion | Unchanged | Contraction | ||
| Current vs 2050s | SSP1-26 | 41.24 | 408.20 | 6.54 | 9.94 | 98.42 | 1.58 |
| SSP2-45 | 59.93 | 406.62 | 8.12 | 14.45 | 98.04 | 1.96 | |
| SSP3-70 | 50.12 | 407.20 | 7.54 | 12.08 | 98.18 | 1.82 | |
| SSP5-85 | 54.12 | 403.99 | 10.75 | 13.05 | 97.41 | 2.59 | |
| 2050s vs 2070s | SSP1-26 | 2.43 | 433.02 | 16.42 | 0.54 | 96.35 | 3.65 |
| SSP2-45 | 13.05 | 458.39 | 8.15 | 2.80 | 98.25 | 1.75 | |
| SSP3-70 | 39.81 | 442.91 | 14.41 | 8.71 | 96.85 | 3.15 | |
| SSP5-85 | 29.19 | 429.10 | 29.01 | 6.37 | 93.67 | 6.33 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



