
Submitted:
24 December 2024
Posted:
24 December 2024
You are already at the latest version
Abstract
Styrax is the largest genus of the family Styracaceae, with about 130 species distributed across America, Europe, and Southeast Asia. The oleo-resin of these woody shrubs, called Styrax benzoin, has a long tradition of use as incense and in therapeutics, which is stimulating research and industrial applications. Many studies have been carried out on the biological activity of different Styrax species, but only a few reviews have been published, mainly focused on traditional uses in folk medicine. This review gathers updated and valuable information on the distribution and phenology of Styrax spp., considering their phytochemicals, biological activity, and current and possible applications in medicine, animal feeding, energy production, and the food industry. Overall, Styrax obassia and Styrax japonicus are the most studied, but Styrax officinalis has been thoroughly investigated for its phytochemicals. Recent literature highlights promising applications in oncology and as an energy crop. The data described in this review could be useful to upgrade the quantity and quality of Styrax benzoin, as well as to expand the knowledge on emerging applications, for example in the development of active packaging for the food industry.
Keywords:
Economic-Importance
; Integrative-Medicine
; Phytochemicals
; Styrax
1. Introduction
The family Styracaceae of the order Ericales consists of 11 genera and about 160 species of dicotyledonous woody shrubs. The family is distributed across the regions of North America, South America, Europe, and Southeast Asia. In the American continent, the species spread from North America to Argentina, whereas in Asia they spread from Japan to Eastern India, and one species occurs in the Mediterranean region. However, other genera of Styracaceae are confined to Eastern Asia. Paleo botany suggests that the family originated in the Eurasian region and distributed across the world; fossils of various genera were obtained from the Americas, France, the Eurasian continent and Japan. The 11 genera of the family include: Styrax, Huodendron, Bruinsmia, Alniphyllum, Changiostyrax, Sinojackia, Parastyrax, Halesia, Rehderodendron and Melliodendron. During the earlier notions of traditional taxonomy, Styracaceae is placed under various families of order Ebenales, later classified into a separate family. With the evidence from molecular and morphological data, Styracaceae is known to be monophyletic in origin, with Styrax and Huodendron in one clade and the rest in another. The members of the family can grow into large trees 20-30 meters tall, and a few species occur as shrubs (1-4 meters).
The leaves are simple, alternate, petiolated, pinnately nerved, stipulated with stellate or peltate trichomes. The inflorescence is terminal with axillary cymes, racemes, or panicles with 1-2 bracteolate flowers. The flowers are bisexual, actinomorphic with hypanthium adnate to ovary walls, the calyx is synsepalous, and the corolla is sympetalous. The stamens are uniseriate, usually 2 to 4 in number, equal to lobes of the corolla; the ovary is carpellate, septate at the base, partly to completely inferior, variable according to genera, with a filiform style and a terminal stigma. The ovules are unitegmic/bitegmic, tenuinucellate, anatropous, 4-9 in number/carpel, with a parenchymatous mesotesta, a seed coat without vascular bundles, a multiplicative and sclerotic outer mesotesta, and vascular bundles in the testa [1]. Pollens are fairly uniform, solitary, radially symmetrical, tricolporate with a tectate-perforate-columellate exine. Fruits are dry with capsular, dehiscence, indehiscent, or drupaceous with persistent calyx. Seeds are globose to fusiform, rarely winged, and the endosperm is copious and oily. Further, the chromosomal number varies within genera, where Styrax has 8 chromosomes, while in Halesia, Sino-jackia and Pterostyrax chromosome number is 12.
The genus Styrax is the largest of the family Styracaceae with about 130 individual species, which represent 80% of the family species. The name Styrax is an ancient Greek word meaning “fragrant resin,” which is predominantly produced in these species [2,3,4]. The genus is widely spread across America, East Asia, the Middle East, and the Mediterranean coast. These species grow in a wide range of climates depending on the region, including humid, warm temperate, lowland, and mountain ranging from the altitude of 100 meters to 3000 meters. A few species are xerophytic and native to the Mediterranean, grow in dry climates, and the plant morphology is modified accordingly to withstand harsh conditions. Molecular and evolutionary analysis suggests that the Styrax genus originated from Eurasia, followed by initial dispersal to Southern America, and few other reports suggest that species native to sub-tropics originated from South America, followed by distribution throughout the current range. On the other hand, Styrax has only three species with entire leaf margins distributed in western Eurasia and western North America. Further, fossil records suggest that Styrax once had a wide distribution from the northern hemisphere to the Eurasian region [2,3,4,5].
The aim of this review is to provide a comprehensive and updated analysis of the scientific literature on the Styrax genus, including its classification, phenology, distribution, secondary metabolites accumulated in different tissues, antioxidant, and antimicrobial activities, nutraceutical potential, and medical and industrial applications, with particular emphasis on the Mediterranean species Styrax officinalis, known for its high-value resin. Studies on Styrax spp. published before January 2025 were collected from scientific databases (Google Scholar and Web of Science, ResearchGate, PubMed, and publishers’ websites), using the keywords “Styrax”, “resin”, “benzoin”, “biological activity”, “uses”, “properties”,” applications”, and combinations thereof.
2. Distribution
Styrax genus morphologically differs from the other genera by characteristics like a perfect flower with an inflorescence of racemes, cymes, or panicles, a superior ovary, and a drupaceous fruit [4]. This genus was initially classified into three sections [6], namely Pamphillia (South American species), Foveolaria, and Styrax. Later, Styrax was re-classified into two sections based on morphology: Valvatae and Styrax [5]. Valvatae is a predominantly tropical evergreen forest species distributed in Asia and Southern America, and it is sub-divided into two series: Valvatae and Benzoin. Valvatae is a neo-tropical clade with 77 species containing wet mesocarp and endocarp, ellipsoid seeds, whereas Benzoin, with nine species, is a paleo tropic having dry mesocarp and endocarp with globose seeds. The sections of Pamphillia and Foveolaria of Perkins classification were merged into Valvatae by Fristch. Initially, Pamphillia was considered an independent genus of Styracaceae, but then it was merged with Styrax [7] due to the similarities in floral and vegetative parts and co-occurrence of gyno-dioecy, while the Styrax section was again subdivided into Cyrta and Styrax. Cyrta series has 33 species with denticulate/serrated leaf margins and axillary/terminal inflorescence. Cyrta series accounts for 90% of species in the Styrax section, which are native to eastern North America and East Asia. Styrax series has only three species: two in the United States and one in the Mediterranean, with entire margins and terminal inflorescences.
The members of Styrax occur as shrubs or trees based on the geographical region, and have an average 180-day life cycle. Due to their widespread distribution, species of Styrax genus display morphological variations to adapt themselves to the environment. For example, Brazilian Styrax species of two regions, Cerrado (S. camporum; S. ferrugineus) and Atlantic forest (S. latifolium, S. martii, and S. leprosus), differ in the wood morphology such as length, diameter, vessels, fiber length. Further, it has been reported that Cerrado species have longer root vessels, whereas the Atlantic ones have longer stem vessels [8]. In Southeast Asia, Styrax spp. is endangered by climate changes, natural disasters and land conversion; it has been estimated that the probability of extinction of S. sumatrana in North Sumatra might increase in 2050, unless suitable conservation strategies are implemented [9] (Figure 1).
The morphological variations led to taxonomical disputes in the several species, which were reclassified several times. Huang et al. [10] re-classified 17 Styrax species of Cyrta series in the Asian region, spread from the west of Japan to Nepal and South to Sumatra; these species also display changes in morphological characteristics according to the habitat. Later, Li et al. [11] re-classified 11 species based on valvate aestivation of corolla lobes in Asia, whereas these species are spread from Eastern India to Southern China and Malesia, and grow in different habitats either as shrubs or trees. Styrax species of America vary in size: S. grandiflora often grows as a shrub with a flabellate branching system and rounded crown, S. argenteus of southern Mexico grows as a large tree that can reach 35 feet while the largest is S. glaber of the Caribbean with the height of 40 feet [12]. Apart from morphology, Styrax species also vary in the ploidy levels, as genome analysis of 15 species revealed ploidy variations ranging from diploid to hexaploid, while S. hookeri is an octaploid species [13].
3. Monophyletic Origin, Genetic Diversity and Polymorphism
Previous reports suggest that the orders Ericales and Ebenales are of a monophyletic origin, and the Styracaceae form a close relationship with Diapensiaceae based on morphological and molecular evidence [14,15]. Interestingly, the plastome analysis of Styracaceae revealed a similar structure of the plastidial genome with the exceptions of Alniphyllum and Bruinsmia, which have an inverted region of 20 kb [16]. Also, plastome analysis assisted in identifying the Styracaceae and its genera as monophyletic except for Halesia and Pterostyrax [16,17]. Yan and co-workers analyzed the plastome sequences of Styracaceae and determined Parastyrax as a sister species to the other seven genera of the family with monophyletic origin; Styrax, Huodendron, Alniphyllum-Bruinsmia are closely associated and form a sister clade within the family, whereas Halesia and Pterostyrax are of polyphyletic origin [16]. Further, the molecular analysis of the Styrax genus using ITS region (nuclear) and trnK, rpoC1, and rpoC2 regions (chloroplast) led to the identification of monophyletic origin among the four species of the genus; however, it failed to identify relations within the species. The disjunctive distribution of the genus was explained by North Eurasian origin of the genus and later migration to South America and East Asia [4].
Styrax species display genetic variations between the wild type and commercial cultivars, since Styrax is grown for economical purposes such as extraction of benzoin resin from the bark, and genetic variations affect benzoin extraction. For instance, the phylogenetic analysis of the Asian species S. sumatrana, S. suberifolius and S. chinensis revealed their closest relationship; these species are supposedly native to China [18]. In addition, S. sumatrana exhibits intra-specific differentiation; molecular analysis using the trnL-trnF chloroplast region from the three populations showed haploid nature and strong genetic differentiation, which occurred due to geographic barriers that affected free gene flow [19]. S. japonicus is a prominent member of the genus, having ornamental and medicinal properties. S. japonicus is a polymorphic species, and the commercial cultivars vary genetically. Chloroplast genome analysis of four S. japonicus sub-species/cultivars displayed the conserved nature of the genome followed by polymorphisms in the few sets of coding and non-coding genes that can be used as DNA barcodes for cultivar identification [20]. Positive selection pressure on two chloroplast genes (ycf1, ndhD) in analyzed samples of the S. japonicus indicated the adaptive evolution of the species towards novel niches [20]. In addition, 23 microsatellite regions were identified on the genomes of S. japonicus and S. confusus, which are highly polymorphic and variable in nature [21]. Transcriptome analysis and identification of expression sequence tags (EST) of S. japonicus also indicated the polymorphic nature of the species [11]. Plastome analysis identified that S. japonicus is closely related to S. grandifloras, S. confusus and S. calvescens [22]. Moreover, S. obassia and S. magnus also show high genetic diversity in the wild populations when compared to the commercial cultivars [23,24]. Interestingly, S. redivivus displays genetic polymorphism and morphological changes within the species depending on the latitude changes. Accordingly, two different groups were suggested based on the morphology, but the data was not enough to delimit the taxa [25].
The prominence of genetic diversity within Styrax species led to several discrepancies in species identification and cultivation for commercial purposes. To authenticate and understand the phylogeny, scientists elucidated the chloroplast genomes of several East Asian species for identifying potential barcodes. Species such as S. obassia, S. dasyanthus, S. zhejiangensis, S. macrocarpus, S. odoratissimus, S. calvescens, and S. japonicus were analyzed for their chloroplast genome; it was observed that these species are related to each other and share a common ancestral origin [22,26,27,28,29,30]. In addition, after analyzing 36 chloroplast genomes of Styrax species, ycf1b, and trnT-trnL regions were identified as potential barcode candidates to discriminate between the species [31]. Further, Susilowati and co-workers [18,19] used the combination of matK and rbcL genes to authenticate the Styrax species in Sumatran region. Nevertheless, the attempts were not successful. Recently, Feng et al. [32], used the ITS barcode to determine the authenticity of benzoin resin extracted from Styrax species and found that most of the analyzed samples were identified as S. japonicus and S. tonkinensis [32]. These results suggest that despite Styrax monophyletic origin, genetic variations occur within the species depending on the biogeography that affects the cultivation practices. In-depth analysis with the assistance of advanced omics techniques to understand these genetic variations and polymorphisms could lead to the development of superior varieties for escalated cultivation.
4. Economic Importance
Styrax species are predominantly grown for their resin and also as ornamental plants. The flowers are known for their fragrance, and the color ranges from white to pink, 1-2 cm in diameter, and blossoms during the mid-spring season. The plants are also used for landscaping and are known for their white bell-shaped flowers, commonly called “snow bells”. Apart from their ornamental purposes, few species are cultivated for wood, resin, and seed oil.
Resins produced by Styrax are generally termed benzoin, and the composition of benzoin resin varies according to the species. Recently, He et al. [33] have overviewed the methods used for extracting benzoin resin and controlling its quality. Benzoins are generally categorized into two classes based on the content of cinnamic acid. Benzoin resin that contains this acid is called Sumatran benzoin, which is produced by Styrax benzoin of Java and Sumatra Islands, while benzoin resin lacking cinnamic acid is called Siam benzoin or benzoe tonkinensis, produced by S. tonkinensis, native to the Indo-China belt. Free cinnamic acid, benzoic acid, benzyl cinnamate, coniferyl cinnamate, and sumaresinolic acid are the key components of Sumatra benzoin, whereas Siam benzoin has coniferyl benzoate, free benzoic acid, cinnamyl cinnamate, siaresinolic acid, sumaresinolic acid, and 3-oxosiaresinolic acid [34,35]. Analysis of different Siam benzoin and Sumatra benzoin resins by Gas Chromatography (GC-MS) and High-Performance Liquid Chromatography (HPLC-MS) coupled with mass spectrometry revealed the presence of various phenolics: benzoic acid, vanillin, isovanillin, benzyl benzoate, lubanol, benzyl cinnamate, p-coumaryl benzoate, coniferyl benzoate, p-coumaryl cinnamate, lupeol, oleanonic acid, cinnamic acid, ethyl cinnamate, coniferyl benzoate [36] (Figure 2).
Benzoin resin is a pathological exudation produced from the injured stems after the injury through the secretory ducts. To collect the resin, farmers usually make an incision, followed by exudation of transparent gum from the wound along with water and other substances; then the gum becomes hard through evaporation, and finally forms benzoin [33]. To upgrade the value of Styrax resin, Kholibrina & Aswandi [37] have proposed to process resin into an essential oil form, thus obtaining also a valuable hydrolate as a by-product, when steam distillation is used.
5. Antioxidant and Antimicrobial Activities of Styrax spp.
The use of Styrax spp. in traditional medicine often relies on their biological activity in terms of antioxidant and antimicrobial properties. Aqueous and organic solvent-based extracts of S. officinalis leaves showed potential antioxidant activity with IC50 values of 11.7 ± 0.53 and 15.5 ± 0.69 μg/mL, respectively. When S. officinalis was compared with Trolox (an IC50 value of 2.8 μg/mL), S. officinalis exhibited higher activity. Similar experiments also showed that the organic extract of leaves from S. officinalis emerged as the best antimicrobial agent [40].
Egonol, extracted from S. officinalis seeds, was demonstrated to exert antioxidant activity when applied to erythrocytes. In detail, superoxide dismutase (SOD) and glutathione-S-transferase (GST) levels were increased in erythrocytes exposed to increasing concentrations of egonol (0.2-1.0 mg/mL) for 15 minutes at 37°C, while the concentration of other enzymes was not affected. The final effect was an accelerated detoxification of superoxide radicals to hydrogen peroxide, and the boost of the antioxidant defense system of erythrocytes [41]. Antioxidant activity, with no antibacterial effect, has been reported recently for the methanolic extract of S. officinalis fruit pericarp [42]. On the contrary, only a weak antioxidant activity was observed for the crude methanolic extracts of Styrax camporum and S. ferrugineus fruits (18.47 ± 1.54 and 2.40 ± 2.01 DPPH % inhibition) with respect to gallic acid (92.41 ± 0.34 DPPH % inhibition), as reported by de Almeida Silva et al. [43]. Moreover, also the kemenyan resin obtained from S. sumatrana demonstrated radical scavenging and antioxidant activity according to DPPH assay, with IC50 values ranging from 15.28 to 31.74 mg/L [44]. Finally, egonol exhibits luminophore properties and emits at near UV region along with electron quenching properties [45]
Several researchers identified the beneficial properties of Styrax spp. extracts and it is evident with the available scientific literature. The main findings regarding the antimicrobial activities of Styrax spp. and derived molecules against pathogenic bacteria and fungi are reported in Table 1 and Table 2, respectively.
Aqueous and organic extracts of S. officinalis showed antioxidant activity as well as inhibitory against potential human pathogens such as Staphylococcus aureus, Shigella sonnei, Escherichia coli, Pseudomonas aeruginosa, Enterococcus faecium, Candida albicans, Epidermophyton floccosum with the minimum inhibitory values ranging from 12.5-25 mg/mL [46].
Also, egonol and a few derived novel compounds from the stem of S. officinalis showed antibacterial activity. Interestingly, Styrax spp. extracts display potential antifungal activity against various pathogens although effect is varies depending on the species. For example, while S. officinalis extract determined the total inhibition of Phythophtora infestans and it reduced the growth of Alternaria solani by 50.58% [50,53].
Essential oils produced from resin are used in food, as flavoring agents in beverages, and as varnishing woods. They have mind-relaxing properties [54] and could also be used for their antibacterial activity. Silver nanoparticles produced from resin essential oil possess antimicrobial activity against gram-negative and gram-positive pathogens and display radical scavenging activities. Molecular docking studies revealed the binding of essential components to the pathogenic bacterial enzymes and the reduction of their activity [55].
Further, various bacterial and fungal endophytic microorganisms were isolated from the tissues of S. officinalis, and the cell-free extracts of these endophytes were demonstrated to possess anti-microbial activity against human pathogens [56].
6. Applications of Styrax spp. in Integrative Medicine
Integrative medicine is a promising branch of evidence-based medicine that combines conventional, complementary and alternative medicine [57]. In this context, herbal plants and, most of all, medicinal plants have a long tradition of bringing medicine back to its roots and stimulating the interest of both researchers and the population. Different Styrax species have been investigated for their pharmacological potential, with S. obassia and S. japonicus emerging as the most extensively studied species [58]. Moreover, several studies [59] have been carried out by Chinese researchers using the so-called “false Styrax”, also named Storax (Liquidambar orientalis), which belongs to a different genus and therefore will not be considered in this review.
The pharmacological properties of various Styrax spp. and the uses in traditional medicine encompass a wide range of biological activities: antimicrobial, antioxidant, anti-inflammatory, antiproliferative, cytotoxic, anti-complement, promoting the biosynthesis of estrogen, inducing apoptosis, acetylcholinesterase inhibitor, antiasthmatic, anti-ulcer, inhibitory effect for interleukin, matrix metalloproteinases activity [58].
In the last decade, the scientific literature on the pharmacological properties of Styrax spp. has focused on possible applications in oncology, although previous studies already demonstrated interesting anti-proliferative effects of triterpenes extracted from S. tonkinensis, especially olenoic acid on human promyelocytic leukemia cell line HL-60 [60]. Recently, a novel hybrid dioxime ligand has been developed using egonol from S. officinalis, and the ligand has been applied to cancer cell lines to determine its efficacy. The ligands inhibited cell migration and induced apoptosis in the cell lines at 5-40µM concentration [51]. Moreover, egonol glucoside obtained from the bark of S. obassia showed cytotoxic activity in vitro against the breast cancer cell line MCF-7, the human cervical cancer cell line Hela, and the HL-60 cell line; in addition, castanoside B from the same raw material demonstrated cytotoxic activity against Hela and MCF-7 cells [61]. Similar results were observed with egonol obtained from the purification of methanol extracts of S. camporum and S. ferrugineus [43]. Finally, methanolic extracts of S. officinalis leaves were observed to be cytotoxic against human hepatocellular liver carcinoma and mammary gland carcinoma cell lines with IC50 values between 14.95µg/mL and 19.38 µg/mL [62].
7. Styrax Benzoin as an Additive for Foods and Feedstuffs
The balsamic resin of S. tonkinensis is a food and feedstuff flavoring agent. The chemical and technical assessment of the safety of this additive was initiated in 2010 within the FAO/WHO Expert Committee on food additives and ended in 2014 with the safety evaluation (FAO and WHO, 2015). Given the NOAL of 500 mg/Kg bw per day, the committee concluded that the resin would not be of safety concern when used as a flavoring agent.
In the European Union, the same resin is listed among the flavoring agents approved as feed additives, in the functional group 2b (aromatizers) included in Annex I of Regulation 1831/03/EC, although authorization expired in 2023.
In the United States, benzoin resin from Styrax spp. is approved by Food and Drug Administration (FDA) as a flavor and flavor enhancer, with the CAS (Chemical Abstract Service) No. 9000-05-09. In China, the resin obtained from Liquidambar spp. is approved as a food flavor with the FEMA (Flavor & Extract Manufacturer Association) No. 3036. Liquidambar spp. extracts or distillates are also regulated as a fragrance ingredient with a specific standard issued by the International Fragrance Association (IFRA); according to this standard, crude gums shall never be used as a fragrance ingredient for any finished product application, due to the risk of dermal sensitization (IFRA Amendment 49, available at:https://www.perfumersworld.com/documents/Project-Templates/IFRA Standards/Styrax.pdf).
Recently, benzoin oleogum resin (BOGR) has been used to develop active films for food packaging. Bathia et al. [63] loaded antioxidant gelatin/pectin films and evaluated several technological and functional properties, finding that BOGR increased the antioxidant activity and the thermal stability of the packaging material, although water permeability, opacity, and thickness were increased.
8. Styrax Officinalis
Styrax officinalis L. (Styracaceae) is a perennial deciduous small tree or shrub, famous for herbal medicine [40,64].
Among traditional medicinal plants, S. officinalis L. is considered the prescribed source of herbal medicine for antiseptic and anti-respiratory diseases. It was one of the oldest and most famous medicinal plants from ancient times. In the last two decades, the global revolution in the search for bioactive herbal products has led to a marked increase in both scientific literature and patents, in particular for the treatment of chronic human diseases in the frame of integrative medicine. In fact, conventional drugs may fail for several reasons due to narrow spectrum of activity that is used to lessen the symptoms, antimicrobial resistance, as well as limited efficacy, e.g. for the treatment of cancer, metabolic disorders, autoimmune diseases, viral infections, and many others [40].
S. officinalis L. is able to grow extensively between 0 and 1000 m altitude [64]. The plant is a deciduous shrub that grows up to 6 m in height. The leaves are simple, alternate, soft, rounded, or oval-shaped and have a greenish-white color with hairy lower surfaces. The flower is white in color and bell-shaped. The ovary of the flower is usually longer with one pattern and 8–16 stamens, twice the number of petals. The fruits are hairy ball-shaped drupes, greenish-yellow with lenient and lengthy apexes [46]. Dried fruits of S. officinalis are of different sizes ranging from 10-25 mm and occur in round/spherical shape, smooth and shiny; color is from light brown to greyish and shows distinct scar (hilum). Dry fruit is tough with 1 or 2, sometimes 3 large ovoid seeds. The tissue pericarp is different, with three distinct layers: a light brown outer layer, called the exocarp; the inner layer, called the endocarp, with a hard shell surrounding the seed; and a bright, yellowish transparent mesocarp [65]. S. officinalis is the only species identified as a drought-tolerant plant among the members of the Styrax genus. Tayoub et al [66] observed the presence of stellate hairs on the aerial parts and secretory glands in the leaf parenchyma as an adaptive strategy to survive drought conditions. Plants usually germinate from seeds by mechanical cracking of seeds, and germination occurs after 21 days during the months between May and September [67]. The species generally produce a typical benzoin resin called solid storaque, which is widely used in perfumery [34](Figure 3).
S. officinalis L. is considered native to arid regions of the Mediterranean Basin and Arabian Plateau and also found in Central America, Mexico, Southeastern Europe, Southwestern Asia and the Mediterranean region, including West and South Anatolia [4,12,40]. The focal region of the spreading of S. officinalis L. links with the coasts of the eastern Mediterranean and the islands between eastern Greece and Israel, mainly Crete and Cyprus. In Turkey, it is intense besides the west and south coasts. However, the species infiltrates deeper into central Turkey [68]. The presence of this plant has been known since ancient times. Ancient Greek geographer Strabo, while describing the city of Selge in his book “Geographika”, mentioned the rich vegetation of the region as well as the styrax tree [64] (Figure 4).
9. Phytochemicals of S. Officinalis L:
9.1. Lignans
The lignan constituents, isolated from the nucleus of S. officinalis L. stem., are: benzofuran, tetrahydrofuran, and furofuran lignans [69]. A naturally occurring benzofuran glycoside, namely 5-(3-hydroxypropyl)-7-methoxy-2- (3,4-methylenedioxyphenyl) benzofuran, egonol is widely present in S. officinalis [40,51]. Pazar et al [70] identified 5 benzofurans from the dried endocarp of S. officinalis: americanin A, egonololeat, egonol-2-metilbutanoat, egonolgentiobiside and homoegonolgentiobiside, out of which americanin and egonololeat were reported for the first time in the Styrax genus. Similarly, several lignans and triterpenes were isolated for the first time from the leaves of S. tonkinensis [71]. Akgul et al. [72] isolated several benzofurans that are derivatives of egonol from the seeds and the stem extracts of S. officinalis: egonol-B-gentiotrioside, 2-(3,4-dimethoxyphenyl)-5-(3-hydroxypropyl)-7-methoxy-benzofuran-B-gentiobioside, egonol-B-gentiobioside, egonol-B-glucoside, and 5-[3′′-(2-methylbutanoyloxy) propyl]-7-methoxy-2-(3′,4′-dimethoxyphenyl) benzofuran (Figure 5).
Hexane extracts of the S. officinalis seeds revealed the presence of two new benzofurans: (i) 5-(3"benzoyl oxy propyl)-7-methoxy-2-(3',4'-methylenedioxyphenyl)-benzofuran; (ii) 4-[3"-(1c-methylbutanoyl oxy) propyl]-2-methoxy-(3',4'-methylenedioxyphenyl)-1a, 5b-dihydrobenzo -[3,4]-cyclobuta oxirene; along with four other benzofurans: (i) 5-[3"-(1c-methylbutanoyloxy) propyl]-7-methoxy-2-(3',4'-dimeth oxy phenyl)-benzofuran; (ii) 5-[3"-(1c-methylbutanoyloxy) propyl]-7- methoxy-2-(3',4'-methyl enedioxyphenyl)-benzofuran; (iii) 5-(3"-acetoxypropyl)-7-methoxy2-(3',4'-methylenediox phenyl) -benzofuran; (iv) 5-(3"-hydroxypropyl)-7-methoxy-2-(3',4'-met hylenedioxy phenyl)-benzofuran. These four benzofurans were also reported from the seeds of S. obassia [72] . Similarly, five benzofurans, namely (i) erythrodiol-3-acetate, (ii) stigmasterol, (iii) (24R)-24-ethylcholesta-4,22-dien-3-one, (iv) 5-(3′′-hydroxy propyl)-7-methoxy-2-(3′,4′-methylenedioxyphenyl) benzofuran, and (v) 5-(3′′-hydroxy propyl)-7-methoxy-2-(3′,4′-dimethoxyphenyl) benzofuran, were isolated from the bark of S. benzoides; chemotaxonomy suggests that S. benzoides is closely associated with S. officinalis [73]. On the other hand, four benzofurans, namely (i) egonol gentiobioside, (ii) egonol gentiotrioside, (iii) egonol glucoside, and (iv) masutakeside, were isolated for the first time from the fruits and bark of S. benzoin, with structures that were quite similar to the benzofurans of S. officinalis [74]. Courel et al. [75] identified egonol, homoegonol, styraxin as significant lignans in the balsams of S. officinalis.
9.2. Terpenoids
A pentacyclic triterpenoid (Yayla et al., 2002; Hu et al., 2019) and later three other triterpenoid saponins, styrax-saponins A-C, and a deacylsaponin (124) were isolated from the extracts of S. officinalis L. Further, it has been reported that S. officinalis, native to Syria, is rich in cardioactive glycosides, flavonoids, anthraquinones, phenols, and triterpenes such as saponins and sapogenins [76]. Tayoub et al. [66] extracted essential oil (EO) from the leaf, flower, and stem tissues of S. officinalis and reported the content of terpenes. It has been identified that terpene content varied between 41-48% in all analyzed tissues. Besides terpenes, lipids, aromatic compounds, and hydrocarbons were predominant in the EO. Specifically, S. officinalis leaf EO contained terpenes like (E)-2-Hexenal, linalool, and geranial, while the stem EO contained significant amounts of α-terpineol, eugenol, and benzyl benzoate. Further, contents of linalool, dodecane, and tridecanal were higher in the flower EO [66]. In continuation to the previous study, [77] researchers evaluated the phenological changes in the EO of S. officinalis native to France. They observed the production of EO, which was yellowish with a fresh odor, had a different composition compared to the other phenological stages of the plant: lipid derivatives and monoterpenes were significantly higher in the EO, while sesquiterpenes were higher in the leaves of the vegetative stage compared to other stages. Three components, namely (E)-2-hexenal, octanol, and geraniol, were significantly higher in all the EOs. On the other hand, terpinen-4-ol, p-cymen-8-ol, geranial linalool were higher at the flowering stage, while (E,E)-2,4 heptadiene, (E,E)-2,4 decadienal, and geranylacetone were higher during the vegetative stage, and only piperanal content was significant during the fruiting stage [77]. A comparative study on the floral emissions of three Styrax species (S. japonicus, S. grandiflora and S. calvescens) resulted in the identification of various volatile terpenes, predominated by α-pinene, myrcene, linalool, germacrene D, ocimene, allo-ocimene and β-elemene; few of these compounds were also reported from the flower EOs of S. officinalis, indicating a similar chemical profile [66,78] (Figure 6).
9.3. Lipids and other Secondary Metabolites
Analysis of the fruit of S. officinalis revealed 15 compounds belonging to seven different classes of natural metabolites with chemotaxonomic relevance: tri-α-linolenoyl-sn-glycerol; 1,2-di-α-linolenoyl-3-linoleoyl-sn-glycerol; 1-α-linolenoyl-2-palmitoyl-sn-glycerol; 1,2-di-α-linolenoyl-sn-glycerol; egonol; demethylegonol; homoegonol; 1,5-anhydro-D-mannitol; glucose; sucrose; 6'''-O-benzoyl-sucrose; raffinose; lactic acid; succinic acid; glutamic acid. Furthermore, S. officinalis is a valuable source of enantiopure 1,5-anhydro-D-mannitol, which has several medicinal potentialities and is a useful building block in organic synthesis, in particular in “green” approaches of valuable and potentially biologically active molecules [79].
In plant tissue culture, treatments of stem calli with boron, niacin, and thiamine diphosphate induced the production of benzoin between 166 and 231%. Further, benzoin possesses various volatile organic compounds such as hexane, cyclohexane, and cyclopentane [80]. Seeds of S. officinalis were analyzed for lipid and protein content at different harvesting times, and seeds were rich in unsaturated fatty acids such as linoleic acid (66-73%) and oleic acid (16-22%). Interestingly, oil content increases by prolonging the harvest time; also, fatty acid contents vary between the first and second harvest. Linoleic acid content was higher during the second harvest, while oleic and palmitic acids were higher in the first harvest [81]. Further, ursolic acid, sumaresinolic acid and its derivatives were identified as predominant triterpenes in the resins of S. officinalis, along with several phenolics and flavonoids, namely cinnamyl cinnamate, methoxy eugenol, cinnamic acid, benzyl cinnamate, cinnamaldehyde, vanillin, eugenol, vanillic acid, benzyl benzoate, cinnamyl benzoate, ferulic acid, cinnamyl alcohol, propyl cinnamate, coniferyl alcohol, 3-phenylpropanyl cinnamate [36]. Dib et al. [82] analyzed the ethanolic extracts of the fruit of S. officinalis, and found that pericarp contained several active phytochemicals, such as tannins, saponins, and triterpenes, whereas alkaloids and flavonoids were absent. These extracts did not show any antimicrobial activity but displayed strong ichthyotoxic and molluscicidal effects. The toxicity tests with the snail Cornu aspersum showed a potent molluscicide activity, and the contact of snails with the extracts led to severe dehydration and foaming, ultimately leading to death in 30 min. However, ingestion of the extract did not show any toxic effects on snails
10. Specific applications of Styrax officinalis
Since ancient times, S. officinalis L has been famous for the preparation of perfumes and incense, as well as for its pharmaceutical properties [68] [83]. S. officinalis was intensively used in traditional medicine for the treatment of several diseases until the 19th century when the pharmaceutical industry started to produce officially approved drugs [40]. Crude extracts and fractions obtained from different plant parts exhibited several medicinal properties, viz., antioxidant and antimicrobial [84], anti-hemorrhoid [85], and antifungal [50] activities.
Phytochemical analysis of S. officinalis whole plant resulted in the identification of cardiac glycosides, saponin glycosides, alkaloids, phenols, volatile oils, tannins, steroids, and flavonoids. Moreover, dry fruit and seed coat extracts of S. officinalis have allelochemicals that can inhibit seedling germination. The water extracts of S. officinalis significantly inhibited Salvia sclarea seedlings growth compared to tap water controls. The allelopathic effect of S. officinalis was confirmed in another study, where scientists treated fennel, coriander, and fenugreek seeds with extracts of S. officinalis seeds, which inhibited the germination of all seeds [86].
11. Applications of Styrax officinalis as a forage crop or energy crop
S. officinalis leaves of different phenological stages have been tested as forage for ruminants. Esen et al. [87] evaluated the nutritional value of this species for goat feeding and found that metabolizable energy and degradability of dry matter and crude protein ranged from moderate to high; although affected by phenological stages, the nutritional value was interesting for application as forage until the end of the flowering stage. In this study, leaves were also used for in vitro gas production, yielding 20-27 mg/200 mg of dry matter. In another report, leaves collected at different periods (May, July, and September) were analyzed for organic matter digestibility and in vitro biogas production. It has been observed that values of crude ash, dry matter, crude fat, acid detergent fiber, and neutral detergent fiber were significantly higher in September compared with other months [88]. Also, a recent report suggests that forage of S. officinalis, for its high nutrient and fiber content, can be used as an effective food for livestock [89].
S. officinalis is considered an alternate oilseed plant in the form of perennial shrubs [64], being seeds rich in fatty acids and lipids that can be used for biodiesel production [90,91]. Palmitic acid, oleic acid and linoleic acid are the primary composite of Styrax seed oil, fatty acid and protein analysis of five Styrax seed samples revealed the uniform fatty acid composition that could be used for developing biodiesel [90]. A study conducted on S. tonkinensis, that is also used as a silviculture plant to maintain forest lands and ecosystems, revealed that its oil content generally improves post 140 days of flowering, and the precursor pool for the fatty acid biosynthesis is obtained predominantly from the pyruvate, acetaldehyde and phosphohexose pathways [92]. Both the seeds and foliage are widely used for biodiesel. Several scientific groups successfully produced biodiesel from the fresh biomass of S. confusus and S. tonkinensis through transesterification, and the yield was up to 65-70% [93,94].
Yesilyurt et al. [64] used S. officinalis L. as a novel material for synthesizing biodiesel, applying the Taguchi experimental design to identify the optimal reaction conditions. Significantly, an oil content 48.29 ± 3.81% was obtained, with a theoretical maximum yield of 89.23% biodiesel at optimal conditions: concentration of catalyst 0.6 wt%, methanol -oil molar ratio of 6:1, duration of reaction about 60 min, and the temperature of 60°C. In this study, the most significant factors for biodiesel yield were catalyst concentration and methanol to oil molar ratio, with the contributing factors of 78.07% and 20.32%, respectively. The properties of fuel -the methyl esters from S. officinalis L. oil were within the ranges of the EN 14214 specifications [64].
12. Applications of Styrax officinalis as a bio-pesticide
Styrax species, particularly Styrax officinalis, have demonstrated potential as natural pesticides due to their bioactive compounds. Hepper [95] reported for the first time the use of a powder for poisoning sea fish off Lebanon. The powder is prepared from the seeds of storax (S. officinalis L) and tubers of common cyclamen (Cyclamen persicum Mill. Another study by Dib et al. [96], investigated the bioactivity of S. officinalis fruit extract. The extract exhibited strong ichthyotoxic and molluscicidal effects. In toxicity tests, snails (Cornu aspersum) exposed to surfaces treated with a 1% (w/v) pericarp extract experienced severe dehydration and foaming, leading to death within 30 minutes. However, ingestion of treated lettuce leaves did not produce observable effects on the snails. These findings suggest that S. officinalis fruit extract is a promising natural contact molluscicide with minimal impact when ingested by non-target organisms.
Additionally, a review by Xia [97], discusses the chemical constituents and biological activities of the genus Styrax. The review highlights that various Styrax species contain compounds such as lignans and terpenoids, which exhibit significant pharmacological activities, including cytotoxic, acetylcholinesterase inhibitory, antioxidant, and antifungal effects. These properties further support the potential use of Styrax species in developing natural pesticides.
Conclusion
The interest in Styrax spp., Styrax benzoin resin and their applications is demonstrated by the growing number of papers being published in the last decade. Recent literature has been focused on possible applications in integrative medicine, especially in oncology, but also in animal feeding or energy production. Emerging applications might include the development of novel active packaging for the food industry. These studies also underscore the potential of Styrax species as sources of natural pesticides, particularly as contact molluscicides, warranting further research into their efficacy and safety in pest control applications.
Although these plants can grow in the desert, they can be endangered by climate changes and, therefore, need preservation strategies, especially in Southeast Asia. In this paper, we have highlighted the significant scientific gaps in the research on the Styrax genus to develop highly resilient and sustainable varieties for global cultivation practices. The information collected in this review can help finding new solutions to save this valuable crop from extinction, at the same time upgrading the quantity and quality of benzoin resin, which is also a source of bioactive compounds with promising and emerging industrial and medical applications.
References
- Shinke, I.; Tokuoka, T. Embryology of Styracaceae and implications for the evolution of the integument number in Ericales. Bot. J. Linn. Soc. 2020, 193, 125-139.
- Fritsch, P. Styracaceae. In Flowering Plants· Dicotyledons; Springer: 2004; pp. 434-442.
- Fritsch, P.W. Phylogeny and biogeography of the flowering plant genus Styrax (Styracaceae) based on chloroplast DNA restriction sites and DNA sequences of the internal transcribed spacer region. Mol. Phylogenet. Evol. 2001, 19, 387-408.
- Fritsch, P.W.; Morton, C.M.; Chen, T.; Meldrum, C. Phylogeny and biogeography of the Styracaceae. Int. J. Plant Sci. 2001, 162, S95-S116.
- Fritsch, P.W. Phylogeny of Styrax based on morphological characters, with implications for biogeography and infrageneric classification. Syst. Bot. 1999, 356-378.
- Perkins, J.R. Styracaceae. In: Engler, Das Pflanzenreich 1907, Heft 30; IV. 241.
- Wallnöfer, B.; ter Welle, B.; Dickison, W. A Revision of Styrax L. section Pamphilia (Mart. ex A. DC.) B. Walln.(Styracaceae). Annalen des Naturhistorischen Museums in Wien. Serie B für Botanik und Zoologie 1997, 681-720.
- Machado, S.R.; Rodella, R.A.; Angyalossy, V.; Marcati, C.R. Structural variations in root and stem wood of Styrax (Styracaceae) from Brazilian forest and cerrado. IAWA J. 2007, 28, 173-188.
- Saputra, M.H.; Lee, H.S. Evaluation of climate change impacts on the potential distribution of Styrax sumatrana in North Sumatra, Indonesia. Sustainability 2021, 13, 462.
- Huang, Y.; Fritsch, P.W.; Shi, S. A revision of the imbricate group of Styrax series Cyrta (Styracaceae) in Asia. Ann. Missouri Bot. Gard. 2003, 491-553.
- Li, G.; Fritsch, P.W. A taxonomic revision of taxa in Styrax series Cyrta (Styracaceae) with valvate corollas. J. Bot. Res. Inst. Tex. 2018, 12, 579-641.
- Gonsoulin, G.J. A revision of Styrax (Styracaceae) in north America, Central America, and the Caribbean. SIDA, Contributions to Botany 1974, 5, 191-258.
- Lobdell, M.S.; Shearer, K. Genome sizes, ploidy levels, and base compositions of Styrax species and cultivars. Hort. Science 2022, 57, 478-484.
- Morton, C.; Chase, M.; Kron, K.; Swensen, S. A molecular evaluation of the monophyly of the order Ebenales based upon rbcL sequence data. Syst. Bot. 1996, 567-586.
- Anderberg, A.A.; Rydin, C.; Källersjö, M. Phylogenetic relationships in the order Ericales sl: analyses of molecular data from five genes from the plastid and mitochondrial genomes. Am. J. Bot. 2002, 89, 677-687.
- Yan, M.-H.; Li, C.-Y.; Fritsch, P.W.; Cai, J.; Wang, H.-C. Phylogeny of the Styracaceae revisited based on whole plastome sequences, including Novel plastome data from parastyrax. Syst. Bot. 2021, 46, 162-174.
- Cai, X.-L.; Landis, J.B.; Wang, H.-X.; Wang, J.-H.; Zhu, Z.-X.; Wang, H.-F. Plastome structure and phylogenetic relationships of Styracaceae (Ericales). BMC Ecol. Evo. 2021, 21, 103.
- Susilowati, A.; Rachmat, H.H.; Fadilah, W.N.; Kusuma, Y.S. Phylogeny of kemenyan toba (Styrax sumatrana) Inferred from trnl-trnf Chloroplast DNA Sequence. In Proceedings of the Proceedings of the International Conference on Natural Resources and Technology (ICONART 2019), 2019; pp. 26-29.
- Susilowati, A.; Hendalastuti, H.; Kholibrina, C.R.; Ramadhani, R. Weak delineation of Styrax species growing in North Sumatra, Indonesia by matK+ rbcL gene. Biodiversitas 2017, 18, 1270-1274.
- Zheng, H.-Z.; Dai, W.; Xu, M.-H.; Lin, Y.-Y.; Zhu, X.-L.; Long, H.; Tong, L.-L.; Xu, X.-G. Intraspecific differentiation of Styrax japonicus (Styracaceae) as revealed by comparative chloroplast and evolutionary analyses. Genes 2024, 15.
- Wang, X.-Y.; Yu, S.; Liu, M.; Yang, Q.-S.; Chen, X.-Y. Twenty-three microsatellite loci for Styrax confusus and Styrax japonicus (Styracaceae). Conserv. Genet. Resour. 2010, 2, 51-54.
- Tong, T.; Shao, L.; Peng, Z. The complete chloroplast genome of Styrax japonicus (Styracaceae), a deciduous tree distributed in East Asia. Mitochondrial DNA Part B 2020, 5, 1863-1864.
- Wang, X.; Xing, S.-y.; Sun, L.-m.; Liu, X.-j.; Kong, Q.-q. Genetic diversity of Styrax obassia Sieb. et Zucc. based on AFLP markers. Biochem. Syst. Ecol. 2015, 61, 28-34.
- Cruz-Salazar, B.; Ruiz-Montoya, L.; Gómez-Ruiz, A.E.; Ramírez-Marcial, N. Genetic variation in natural and nursery populations of Styrax magnus (Styracaceae) for the restoration of humid mountain forests in southern Mexico. New Forests 2022, 53, 721-734.
- Fritsch, P. Population srtructuring and patterns of morphological variation in Californian Styrax (Styracaceae). Aliso: A Journal of Systematic and Floristic Botany 1996, 14, 205-218.
- Dong, F.; Liu, X.; Shen, X.; Sheng, W.; Jiang, D.; Li, Y. The complete chloroplast genome sequence of Styrax zhejiangensis, a species with an extremely small population. Mitochondrial DNA Part B 2018, 3, 998-999.
- Xu, W.-B.; Yang, P.; Huang, Y.-S.; Zhang, R.; Tang, M. Styrax hwangiae (Styracaceae), a new species endemic to limestone areas of Guangxi, China. Phytotaxa 2021, 478, 171-178.
- Xu, X.; Zhang, Y.; Tong, L.; Wang, Y.; Xia, C. The complete chloroplast genome of Styrax dasyanthus Perkins (Styracaceae). Mitochondrial DNA Part B 2020, 5, 961-962.
- Zhang, Y.; Tong, L.; Xu, X.; Zhao, Z.; Wang, Y. The complete chloroplast genome sequence of Styrax odoratissimus (Styracaceae). Mitochondrial DNA Part B 2019, 4, 4051-4052.
- Tong, L.; Xu, X.; Wang, Y.; Zhang, Y.; Zhao, Z. The complete chloroplast genome of Styrax faberi Perk.(Styracaceae). Mitochondrial DNA Part B 2020, 5, 98-99.
- Song, Y.; Zhao, W.; Xu, J.; Li, M.; Zhang, Y. Chloroplast genome evolution and species identification of Styrax (Styracaceae). Biomed Res. Int. 2022, 2022, 5364094.
- Feng, J.; He, Q.; Xie, A.; Liu, Y. Successful identification of the species of the semipetrified amber medicinal resin benzoin using molecular diagnostic technology. Sci. Rep. 2023, 13, 2943.
- He, L.; Zhou, Y.; Wan, G.; Wang, W.; Zhang, N.; Yao, L. Antinociceptive effects of flower extracts and the active fraction from Styrax japonicus. J. Ethnopharmacol. 2022, 284, 114779.
- Custódio, D.L.; Veiga-Junior, V.F. True and common balsams. Rev. Bras. Farmacogn. 2012, 22, 1372-1383.
- Burger, P.; Casale, A.; Kerdudo, A.; Michel, T.; Laville, R.; Chagnaud, F.; Fernandez, X. New insights in the chemical composition of benzoin balsams. Food Chem. 2016, 210, 613-622.
- Hovaneissian, M.; Archier, P.; Mathe, C.; Culioli, G.; Vieillescazes, C. Analytical investigation of styrax and benzoin balsams by HPLC-PAD-fluorimetry and GC-MS. Phytochem. Anal. 2008, 19, 301-310.
- Kholibrina, C.; Aswandi, A. Increasing added-value of styrax resin through post-harvesting techniques improvement and essential oil based product innovation. In Proceedings of the IOP Conference Series: Earth and Environmental Science, 2021; p. 012045.
- Lobdell, M. Styrax in cultivation: evaluation of an underrepresented ornamental genus. University of Delaware, 2013.
- Sohail Akhtar, M.; Alam, T. Chemistry, Biological Activities, and Uses of Benzoin Resin. In Gums, Resins and Latexes of Plant Origin: Chemistry, Biological Activities and Uses; Springer: 2022; pp. 1-22.
- Jaradat, N. Phytochemistry, traditional uses and biological effects of the desert plant Styrax officinalis L. J. Arid Environ. 2020, 182, 104253, doi:. [CrossRef]
- Bölükbaşı, S.Ş.; Öztürk, S.E.; Anıl, H.; Siliğ, Y. Effects of egonol that isolated from seeds of Styrax officinalis (L.) on lipid peroxidation and antioxidant defense system in human erythrocytes. Cumhur. Med. J. 2016, 38, 168-178.
- Sak, S.; Özdenefe, M.S.; Erol, Ü.H.; Takcı, A.M. Total chemical components, biological activity and chromatographic analyzes of Styrax officinalis fruit extract. JAES 2024, 9, 457-463.
- de Almeida Silva, T.; Polo, E.M.; Henrique, C.Y.; Alves, O.J.A.; Nicolella, H.D.; Gimenez, V.M.M. Styrax camporum and S. ferrugineus fruits: norneolignans, antioxidant and cytotoxic activities. J. App. Pharm. Sci. 2016, 6, 075-080.
- Hidayat, A.; Iswanto, A.H.; Susilowati, A.; Rachmat, H.H. Radical scavenging activity of kemenyan resin produced by an Indonesian native plant, Styrax sumatrana. J. Korean Wood Sci. Technol. 2018, 46, 346-354.
- İçlİ, S.; Anil, H.; Dİndar, B.; Çalişkan, Ö.A.; Yayla, Y.; Doroshenko, A.O.; Alp, S. High fluorescence emissions of some natural benzofurane derivatives isolated from Styrax officinalis. Turk. J. Chem. 2000, 24, 199-208.
- Jaradat, N.; Al-Masri, M.; Zaid, A.N.; Hussein, F.; Shadid, K.A.; Al-Rimawi, F.; Shayeb, K.; Sbeih, A.; Eid, A. Assessment of the antimicrobial and free radical scavenging activities of Moluccella spinosa, Helichrysum sanguineum, and Styrax officinalis folkloric medicinal plants from Palestine. Orient. Pharm. Exp. Med. 2018, 18, 107-114.
- Bertanha, C.S.; Utrera, S.H.; Gimenez, V.M.M.; Groppo, M.; Silva, M.L.A.; Cunha, W.R.; Martins, C.H.G.; Januário, A.H.; Pauletti, P.M. Antibacterial evaluation of Styrax pohlii and isolated compounds. Braz. J. Pharm. 2013, 49, 653-658.
- Pauletti, P.M.; Araújo, A.R.; Young, M.C.M.; Giesbrecht, A.M.; da Silva Bolzani, V. nor-Lignans from the leaves of Styrax ferrugineus (Styracaceae) with antibacterial and antifungal activity. Phytochemistry 2000, 55, 597-601.
- Öztürk, S.E.; Akgül, Y.; Anıl, H. Synthesis and antibacterial activity of egonol derivatives. Bioorg. Med. Chem. 2008, 16, 4431-4437.
- Yanar, Y.; Kadioğlu, I.; Gökçe, A.; Demirtas, I.; Gören, N.; Çam, H.; Whalon, M. In vitro antifungal activities of 26 plant extracts on mycelial growth of Phytophthora infestans (Mont.) de Bary. Afr. J. Biotechnol. 2011, 10, 2625-2629.
- Babahan-Bircan, I.; Emirdağ, S.; Özmen, A.; Abbak, M.; Ujam, O.T.; Demirkaya, I.; Günay, M.E. A new hybrid ligand and its metal complexes from a natural plant (Styrax officinalis) bearing egonol, thiosemicarbazone and oxime units, and their anti-cancer activities. Appl. Organomet. Chem. 2022, 36, e6784.
- Liu, B.-L.; Hu, X.; He, H.-L.; Qiu, L.; Li, Y.-Z.; Ding, W.-B. A new epicatechin glucopyranoside derivative from Styrax suberifolius. Nat. Prod. Res. 2020, 34, 1977-1983.
- Yanar, Y.; Gökçe, A.; Kadioglu, I.; Çam, H.; Whalon, M. IIn vitro antifungal evaluation of various plant extracts against early blight disease (Alternaria solani) of potato. Afr. J. Biotechnol. 2011, 10, 8291-8295.
- Atia Sharif, H.N.; Rehman, R.; Mushtaq, A.; Rashid, U. Review on bioactive potential of Benzoin resin. Int. J. Chem. Biochem. Sci 2016, 10, 106-110.
- Debnath, S.; Nath, M.; Sarkar, A.; Roy, G.; Chakraborty, S.K.; Debnath, B. Phytochemical characterization of Styrax benzoin resin extract, molecular docking, ADME, and antibacterial activity study. Nat. Prod. Res. 2024, 38, 1263-1268.
- Eroglu, D.; Tan, E.; Caliskan Ozdemir, S.; Kaya, I.Y.; Demiray, H.; Uzel, A. Endophytic microorganisms from Styrax officinalis and their antimicrobial activities. The 3rd International Symposium on EuroAsian Biodiversity: Belarus 2017.
- Aggarwal, M.; Aggarwal, B.; Rao, J. Integrative medicine for cardiovascular disease and prevention. Med. Clin. 2017, 101, 895-923.
- Xia, D.-D.; Han, X.-Y.; Zhang, Y.; Zhang, N. Chemical constituents and their biological activities from genus Styrax. Pharmaceuticals 2023, 16, 1043.
- Zhang, F.; Zhang, T.; Gong, J.; Fang, Q.; Qi, S.; Li, M.; Han, Y.; Liu, W.; Ge, G. The Chinese herb Styrax triggers pharmacokinetic herb-drug interactions via inhibiting intestinal CYP3A. Front. Pharmacol. 2022, 13, 974578.
- Wang, F.; Hua, H.; Pei, Y.; Chen, D.; Jing, Y. Triterpenoids from the resin of Styrax tonkinensis and their antiproliferative and differentiation effects in human leukemia HL-60 cells. J. Nat. Prod. 2006, 69, 807-810.
- Cao, T.Q.; Lee, B.M.; Jung, Y.W.; Nguyen, V.T.; Kim, J.A.; Min, B.S. Cytotoxic activity of compounds from Styrax obassia. Nat. Prod. Commun. 2017, 12, 1934578X1701200230.
- Almulla, S.I.; Diwood, N.M.; Alzbeede, A.A. Assessment of cytotoxic potentials of Styrax officinalis (Styracaceae) extracts from jordan against human cancer cell lines. J. Biomed. Sci. 2024, 1, 31-39.
- Bhatia, S.; Jawad, M.; Chinnam, S.; Al-Harrasi, A.; Shah, Y.A.; Koca, E.; Aydemir, L.Y.; Alam, T.; Mohan, S.; Zoghebi, K. Preparation and characterization of gelatin-pectin-based active films incorporated with Styrax benzoin oleo gum resin. Polym. Adv. Technol. 2024, 35, e6539.
- Yesilyurt, M.K.; Cesur, C. Biodiesel synthesis from Styrax officinalis L. seed oil as a novel and potential non-edible feedstock: A parametric optimization study through the Taguchi technique. Fuel 2020, 265, 117025.
- Hamama, Z.; Khomik, A.; Suslina, S. Morphological characterization of fruits of syrian medical plant (Styrax officinalis)-determination of saponin content. In Proceedings of the Problems of medicine in modern conditions/Collection of scientific papers based on the results of the international scientific and practical conference. , Kazan, 2016; pp. 169-173.
- Tayoub, G.; Schwob, I.; Bessiere, J.-M.; Masotti, V.; Rabier, J.; Ruzzier, M.; Viano, J. Composition of volatile oils of Styrax (Styrax officinalis L.) leaves at different phenological stages. Biochem. Syst. Ecol. 2006, 34, 705-709.
- Yücedağ, C. Effects of cracking and sowing time on germination of Styrax officinalis L. seeds. Afr. J. Biotechnol. 2011, 10, 16448-16451.
- Amigues, S. Le styrax et ses usages antiques. J. Sav. 2007, 2, 261-318.
- Pauletti, P.M.; Teles, H.L.; Silva, D.H.; Araújo, Â.R.; Bolzani, V.S. The styracaceae. Rev. Bras. Farmacogn. 2006, 16, 576-590.
- Pazar, E.; Akgül, Y. Chemical composition of the endocarps of fruits of Styrax officinalis L. Nat. Prod. Res. 2015, 29, 1466-1468.
- Hu, W.-L.; Li, Z.-L.; Chen, Q.-J.; Sun, Y.-W.; Zhai, S.; Lu, F.; Li, F.; Zhang, C.-F. Triterpenes and lignans from the leaves of Styrax tonkinensis. Biochem. Syst. Ecol. 2019, 86, 103891, doi:. [CrossRef]
- Akgul, Y.Y.; Anil, H. Benzofurans and another constituent from seeds of Styrax officinalis. Phytochemistry 2003, 63, 939-943.
- Meepowpan, P.; Winairuangrid, P.; Thangsunan, P.; Thavornyutikarn, P. Phytochemical constituents of the stems of Styrax benzoides craib and their chemosystematic significance. Chiang Mai. J. Sci. 2018, 45, 1407-1414.
- Yusof, H.; Din, L.B.; Yaacob, W.A. Benzofuran glycosides from styrax benzoin. Malay. J. Anal. Sci. 2014, 18, 676-682.
- Courel, B.; Adam, P.; Schaeffer, P. The potential of triterpenoids as chemotaxonomic tools to identify and differentiate genuine, adulterated and archaeological balsams. Microchem. J. 2019, 147, 411-421.
- Mansour, O.; Darwish, M.; Ismail, G. Phytochemical study of Stryax officinalis Lin Syria. Res. J. Pharm. Technol. 2018, 11, 1214-1216.
- Tayoub, G.; Schwob, I.; Bessière, J.M.; Rabier, J.; Masotti, V.; Mévy, J.P.; Ruzzier, M.; Girard, G.; Viano, J. Essential oil composition of leaf, flower and stem of Styrax (Styrax officinalis L.) from south-eastern France. Flavour Fragr. J. 2006, 21, 809-912.
- Chen, C.; Cao, Y.; Chen, H.; Ni, M.; Yu, F. Floral scent compounds and emission patterns of three Styrax species. Dendrobiology 2021, 85.
- Venditti, A.; Frezza, C.; Serafini, I.; Pulone, S.; Scardelletti, G.; Sciubba, F.; Bianco, A.; Serafini, M. Chemical profiling of the fruits of Styrax officinalis L. from Monti Lucretili (Latium region, Central Italy): Chemotaxonomy and nutraceutical potential. Trends Phytochem. Res. 2018, 2, 1-12.
- Demiray, H.; DEREBOYLU, A.E.; YAZICI, Z.I.; Karabey, F. Identification of benzoin obtained from calli of Styrax officinalis by HPLC. Turk. J. Bot. 2013, 37, 956-963.
- Cesur, C.; Uskutoglu, T.; Çoşge Şenkal, B.; Kiralan, S.S.; Kiralan, M.; Ramadan, M.F. Influence of harvest time on protein content and lipids profile of wild Styrax officinalis seeds from different locations in Turkey. Rend. Lincei Sci. Fis. Nat. 2019, 30, 689-698.
- Dib, R.; Makhoul, K.; Maalouf, R. Preliminary bioactivity investigation of Styrax officinalis fruit extract as potential biopesticide. J. Pharmacognosy Phytother. 2016, 8, 209-213.
- Vardar, Y.; Oflas, S. Preliminary studies on the Styrax oil. Qualitas Plantarum Et Materiae Vegetabiles 1973, 22, 145-148.
- Proestos, C.; Boziaris, I.; Nychas, G.-J.; Komaitis, M. Analysis of flavonoids and phenolic acids in Greek aromatic plants: Investigation of their antioxidant capacity and antimicrobial activity. Food Chem. 2006, 95, 664-671.
- Çakılcıoğlu, U.; Türkoğlu, İ. Plants used for hemorrhoid treatment in Elaziğ central district. In Proceedings of the I International Medicinal and Aromatic Plants Conference on Culinary Herbs 826, 2007; pp. 89-96.
- Şenkal, B.C.; Uskutoğlu, T. Impacts of extracts from Styrax officinalis . On seedling growth of Salvia sclarea. J. Agric. Sci. 2021, 5, 750-756.
- Esen, S.; Koc, F.; Özdüven, L.; Eseceli, H.; Cabi, E.; Karadağ, H. In situ and in vitro nutritive value assessment of Styrax officinalis l. as an alternative forage source for goat feeding. J. Agric. Sci. 2022, 28, 181-188.
- Balyen, İ.; Kiraz, A.B. Determination of nutritional value and methane production potential of styrax tree (Styrax officinalis L.) leaves. Heliyon 2024, 10.
- Ertekin, İ.; Atış, İ.; Yılmaz, Ş.; Can, E.; Kızılşimşek, M. Comparison of shrub leaves in terms of chemical composition and nutritive value. Kahramanmaras Sutcu. Imam. Univ. Tarim. Doga Derg. 2019, 22, 781-786.
- Wu, Q.; Zhao, X.; Chen, C.; Zhang, Z.; Yu, F. Metabolite profiling and classification of developing Styrax tonkinensis kernels. Metabolites 2020, 10, 21.
- Wu, Q.; Cao, Y.; Zhao, X.; Zhang, Z.; Yu, F.; Guy, R.D. A comparative study of seed reserve accumulation in five Styrax species with potential for biofuel production. Trees 2020, 34, 891-902.
- Wu, Q.; Cao, Y.; Chen, C.; Gao, Z.; Yu, F.; Guy, R.D. Transcriptome analysis of metabolic pathways associated with oil accumulation in developing seed kernels of Styrax tonkinensis, a woody biodiesel species. BMC Plant Biol. 2020, 20, 1-17.
- Liu, Z.X.; Nie, X.A.; Wang, Y.G. Biodiesel preparation from Styrax Confusus Hemsl. Oil catalyzed by magnetic catalyst S2O82--ZrO2/Fe3O4. Adv. Mater. Res. 2013, 805, 247-250.
- Wang, Y.; Nie, X.; Liu, Z. Biodiesel synthesis from Styrax confusus HEMSL CATALYZED by S 2 O 8 2−/ZrO 2-TiO 2-Fe 3 O 4. JAOCS 2015, 92, 813-820.
- Hepper, F.N. Two plant fish-poisons in Lebanon. Vet. Hum. Toxico. 2004, 46, 338-339.
- Dib, R.; Makhoul, K.; Maalouf, R. Preliminary bioactivity investigation of Styrax officinalis fruit extract as potential biopesticide. J. Pharmacognosy Phytother. 2016, 8, 209-213.
- Xia, D.-D.; Han, X.-Y.; Zhang, Y.; Zhang, N. Chemical constituents and their biological activities from genus Styrax. Pharm. 2023, 16, 1043.
Figure 1.
Geographic distribution of botanical species belonging to the genus Styrax. https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:327016-2#source-KBD.
Figure 1.
Geographic distribution of botanical species belonging to the genus Styrax. https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:327016-2#source-KBD.
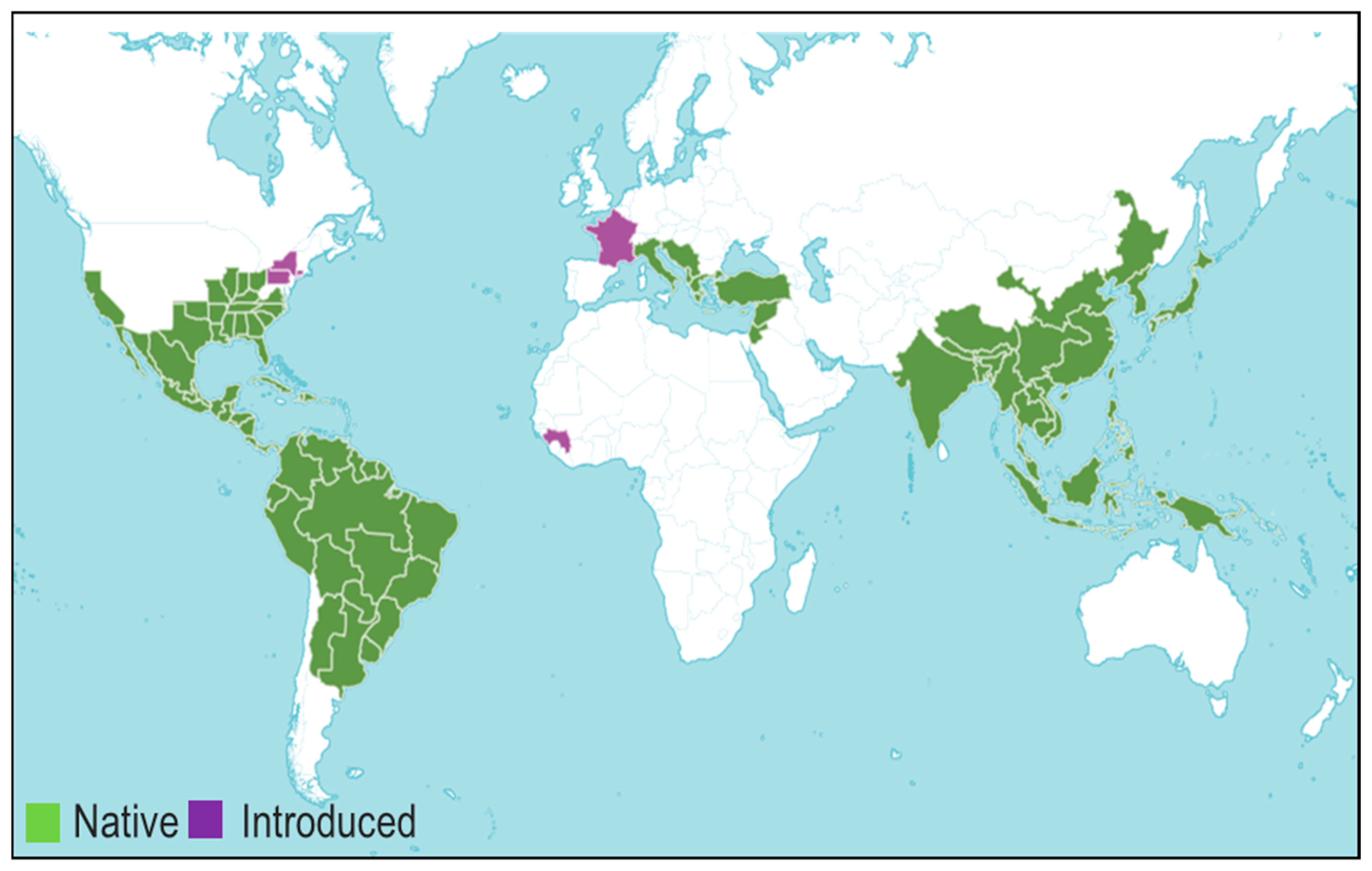
Figure 2.
Major chemical compounds found in Sumatra benzoin and Siam benzoin.
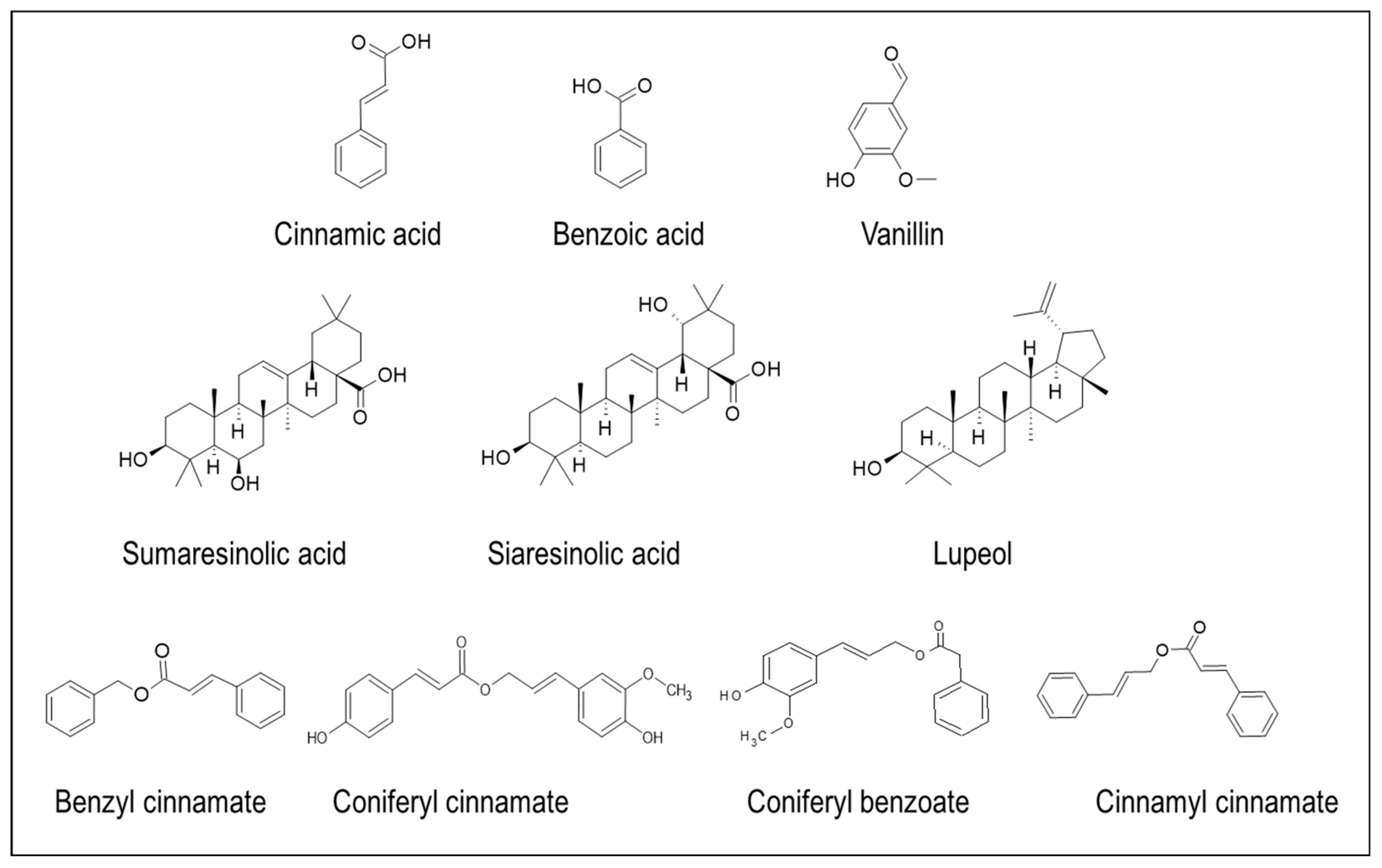
Figure 3.
Photograph of Styrax officinalis L. of the Kabul Mountain (North Israel). (A) Styrax shrub/tree; (B) flowers; (C) fruits; (D) seeds (personal photos).
Figure 3.
Photograph of Styrax officinalis L. of the Kabul Mountain (North Israel). (A) Styrax shrub/tree; (B) flowers; (C) fruits; (D) seeds (personal photos).

Figure 4.
Distribution map of Styrax officinalis. S. Officinalis distribution map in the temperate and subtropical regions of the Mediterranean Basin, Middle East, and in various parts of Western Europe.https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:327016-2#source-KBD.
Figure 4.
Distribution map of Styrax officinalis. S. Officinalis distribution map in the temperate and subtropical regions of the Mediterranean Basin, Middle East, and in various parts of Western Europe.https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:327016-2#source-KBD.
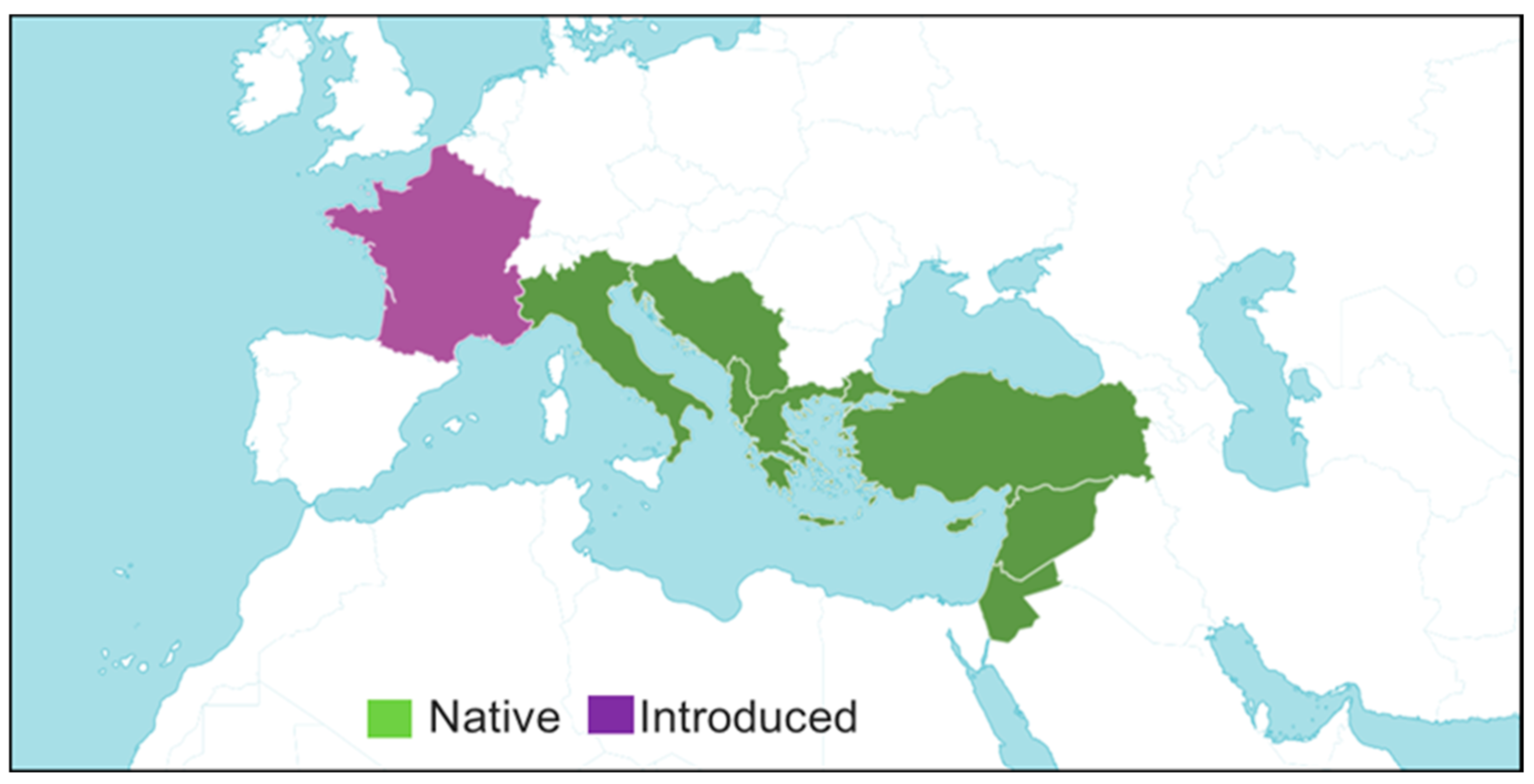
Figure 5.
Lignan compounds isolated from the nucleus of S. officinalis L. stem.
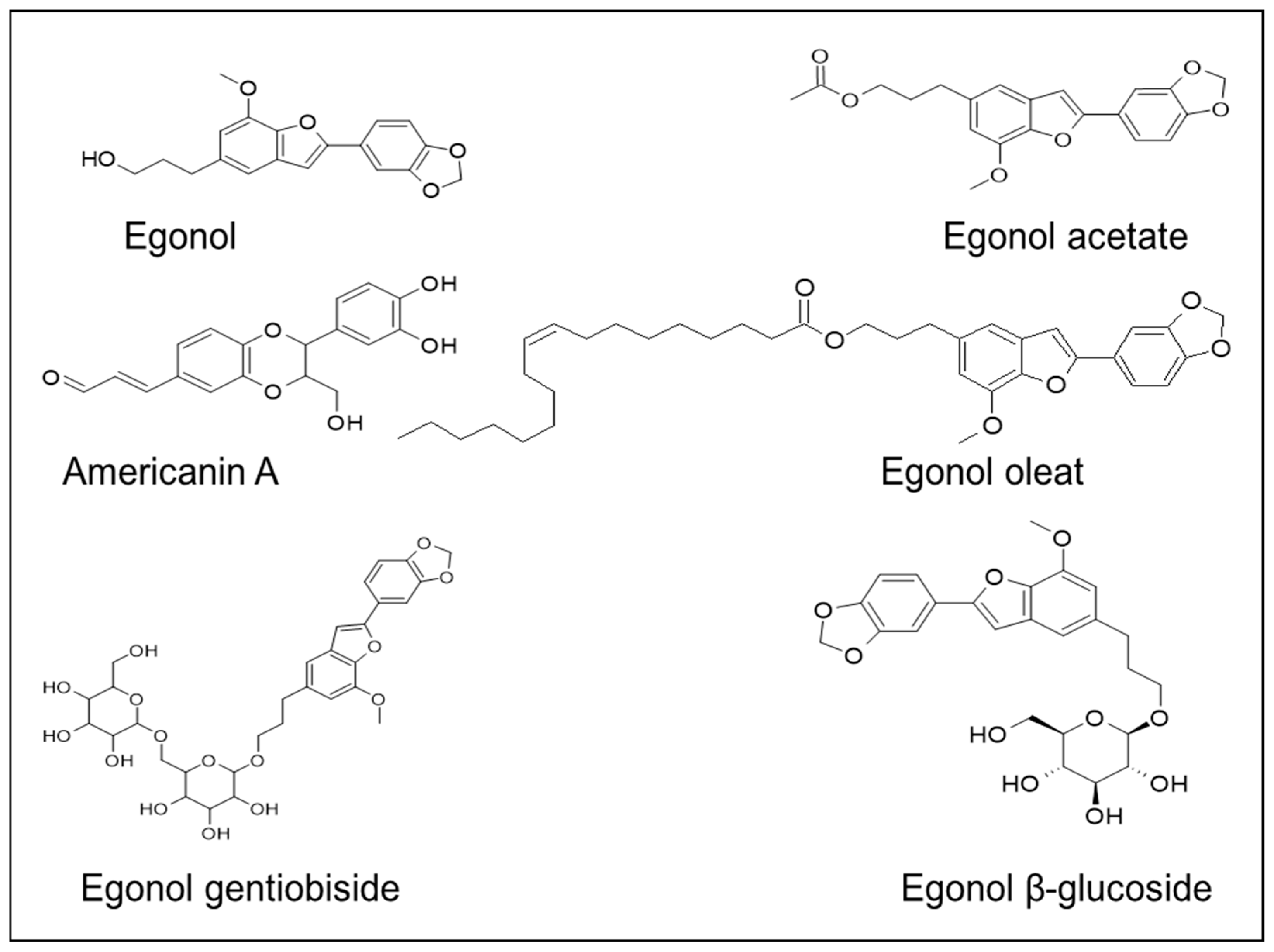
Figure 6.
Major terpene volatile compounds isolated from the S. officinalis L. essential oils of different tissues.
Figure 6.
Major terpene volatile compounds isolated from the S. officinalis L. essential oils of different tissues.
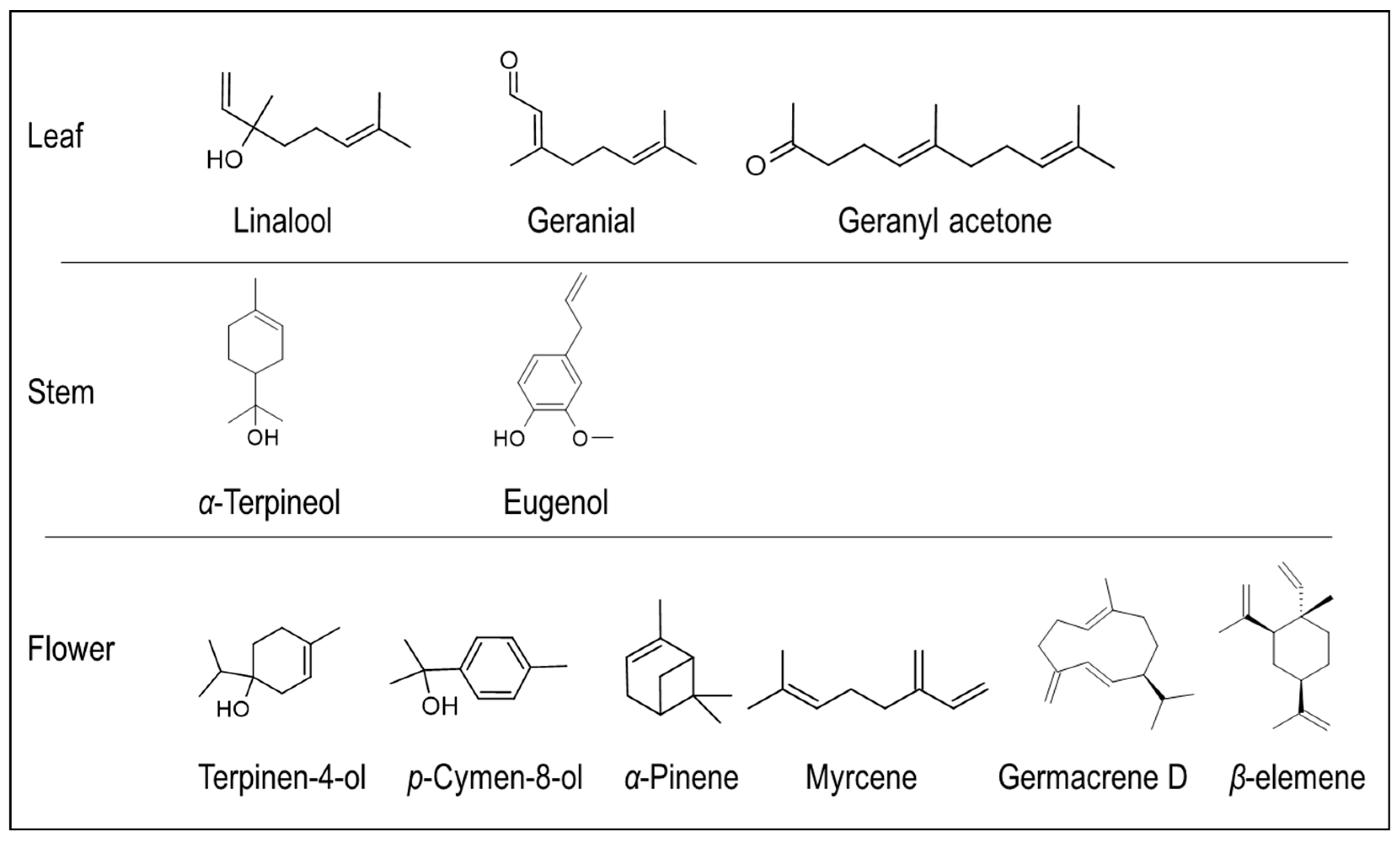

Table 1.
Antibacterial activity of Styrax spp. and derived molecules.
| Species/compound | Part of the plant | Analytical assay | Minimal Inhibitory Concentration | Target bacteria | Reference |
|---|---|---|---|---|---|
| Styrax officinalis organic extract | Aerial part | Minimum Inhibitory Concentration assay | 12.5 mg/mL 12.5 mg/mL 3.125 mg/mL 6.25 mg/mL |
Staphylococcus aureus, Enterococcus faecium, Methicillin resistant S. aureus (MRSA) Pseudomonas aeruginosa |
[46] |
| Styrax pohlii n-hexane fraction and EtOAc fraction | Aerial parts | Microdilution method | 200 μg/mL |
Streptococcus pneumoniae, Haemophilus influenzae (EtOAc fraction) |
[47] |
| Crude extract (CH2Cl2–CH3OH) of S. ferrugineus | leaves | TLC plates | 200 mg/mL |
Staphylococcus aureus | [48] |
| Egonol, egonol glycoside (S. ferrugineus) |
leaves | TLC plates | 10-20 mg/mL |
Staphylococcus aureus |
[48] |
| Egonol (S. officinalis) |
fruit | Mueller Hinton Broth method | 800 mg/mL |
Staphylococcus aureus, Bacillus subtilis, Escherichia coli |
[49] |
| Homoegonol, homoegonol glucoside (S. ferrugineus) |
leaves | TLC plates | 10-20 mg/mL | Staphylococcus aureus |
[48] |
Table 2.
Antifungal activity of Styrax spp. and derived molecules.
| Species/compound | Part of the plant | Analytical assay | Minimal Inhibitory Concentration | Target fungi | Reference |
|---|---|---|---|---|---|
| Styrax officinalis methanolic extract | Fruits | Radial growth method | 8% (w/V) | Phytophtohra infestans | [50] |
| Styrax officinalis organic extract | Aerial part | Minimum Inhibitory Concentration assay | 12.5 mg/mL |
Candida albicans |
[46] |
| Crude extract (CH2Cl2–CH3OH) (S. ferrugineus) |
leaves | TLC plates | 800 mg/mL |
Candida albicans | [48] |
| Egonol (S. ferrugineus) |
leaves | TLC plates | 5-10 μg | Cladosporium cladosporioides | [51] |
| Egonol | S. officinalis | Mueller Hinton Broth method | 25 μg/mL | Candida albicans | [49] |
| Homoegonol (S. ferrugineus) |
leaves | TLC plates | 5-10 μg | Cladosporium cladosporioides | [7] |
| Methyl β-orcinolcarboxylate (S. suberifolius) |
bark | Radial growth inhibition assay | 86.72% inhibition at 100 μg/mL | Phomopsis cytospore | [52] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



