
Submitted:
19 December 2024
Posted:
21 December 2024
You are already at the latest version
Abstract
Pancreatic adenocarcinoma (PDAC) is an aggressive cancer with poor prognosis, primarily due to its immunosuppressive tumor microenvironment (TME), which contributes to treatment resistance. Recent research shows that the microbiome plays a key role in PDAC development, with microbial imbalances (dysbiosis) promoting inflammation, cancer progression, therapy resistance, and treatment side effects. Microbial metabolites can also affect immune cells, especially natural killer (NK) cells, which are vital for tumor surveillance, therapy response and treatment-related side effects. Dysbiosis can affect NK cell function, leading to resistance and side effects. We propose that a combined biomarker approach, integrating microbiome composition and NK cell profiles, can help predict treatment resistance and side effects, enabling more per-sonalized therapies. This review examines how dysbiosis contributes to NK cell dysfunction in PDAC and discusses strategies (e.g., antibiotics, probiotics, vaccines) to modulate the microbiome and enhance NK cell function. Targeting dysbiosis could modulate NK cell activity, improve the effectiveness of PDAC treatments, and reduce side effects. However, further research is needed to develop unified NK cell-microbiome interaction-based biomarkers for more precise and effective patient outcomes.
Keywords:
Keywords: pancreatic cancer
; dysbiosis
; NK cell
; biomarkers
; treatment resistance
; side effects
1. Introduction
Pancreatic ductal adenocarcinoma (PDAC) is the third deadliest cancer in high-income countries, with poor prognosis mainly due to its dense tumor microenvironment (TME) and immunosuppressive niche, which promote tumor progression and treatment resistance [1,2,3]. The TME also provides a favorable environment for microbial survival, leading to dysbiosis that induces immune suppression, including natural killer (NK) cell dysfunction [4]. Currently, no universal screening method exists for early PDAC detection or predicting treatment responses [5]. Moreover, there are limited diagnostic tools for assessing cancer treatment side effects, underscoring the need for new strategies.
Microbial communities, including bacteria, fungi, and viruses, play crucial roles in both health and disease [6,7]. Dysbiosis—an imbalance in these communities—has been linked to various diseases, often involving pathogenic overgrowth, loss of beneficial microbes, or reduced microbial diversity. While defining a healthy microbiota is challenging due to technical limitations, studying the mechanisms of dysbiosis provides important insights into disease processes [8].
Several studies characterized the tumor microbiota in PDAC, emphasizing the microbiome’s role in PDAC development, progression, and treatment response. Microbiota from the oral, gut, and tumor sites influence tumor biology by inducing genetic mutations, reshaping the immune landscape, and regulating cancer metabolism and oncogenic pathways. [9,10,11,12]. Research on microbiome differences between PDAC patients and healthy individuals has led to investigations on how microbial communities interact in the oral, gut, and tumor niches to affect PDAC outcomes [13]. Metagenomic, metaproteomic, and metabolomic analyses of microbiota samples have revealed that intratumoral microbiota diversity correlates with survival outcomes. Higher microbial diversity has been associated with better overall survival in resected PDAC patients. Specific microbes such as Megasphaera and Sphingomonas are linked to longer survival, and Clostridium with shorter survival [14,15,16,17]. These findings suggest that microbiome composition could serve as a prognostic biomarker for PDAC. Microbial species, particularly from the phyla Firmicutes and Proteobacteria, are predominant in PDAC, with Proteobacteria being most abundant in pancreatic tumors and associated with metastasis [18,19].
The gut microbiota also influences immune responses. Specific bacteria like Citrobacter freundii and Shigella sonnei have been shown to upregulate oncogenic pathways and contribute to immunosuppression, reprogramming the TME. Furthermore, Acidovorax ebreus (A. ebreus) is linked to reduced levels of immune cells, further promoting PDAC progression. Metabolites converted by the microbiota can regulate immune cells accumulation and activation within tumors, affecting oncogenic pathways, reducing immune cell-mediated cytotoxicity [19,20,21]. Interestingly, studies in other cancer types have shown that intratumoral microbial injections, such as Propionibacterium acnes, can accelerate tumor growth by increasing inflammatory cytokines like TNF-α and IL-1β, which activate oncogenic signaling pathways, including the Hedgehog pathway [21,22]. Since the Hedgehog pathway is a major driver in PDAC, affecting immune surveillance, further research is needed to understand how intratumoral microbes interact with these pathways in PDAC.
NK cells are crucial in immune surveillance and tumor control, targeting drug-resistant cancer cells and eliminating malignant cells without relying on major histocompatibility complex (MHC) or tumor-associated antigen presentation. [23,24,25]. They rely on a balance of activating and inhibitory receptor signals to execute cytotoxic functions, including antibody-dependent cell-mediated cytotoxicity (ADCC). NK cells can also interact with dendritic cells (DCs) to initiate adaptive immune responses, enhancing tumor immunity [26,27,28]. However, NK cell dysfunction due to cancer or treatments can impact therapy efficacy and side effects. In PDAC, where NK cells are rare in the tumor tissue and show reduced cytotoxic activity, targeting NK cell activity is an emerging strategy for improving therapeutic outcomes.
Recent findings suggest that gut microbiota plays a crucial role in promoting NK cell infiltration into tumors. For example, in a mouse tumor model colonized with Helicobacter hepaticus, improved cancer prognosis was observed, along with an increased number of NK cells within the tumor [29]. Additionally, microbiota-mediated metabolites conversion regulates NK cell cytotoxicity, further enhancing the antitumor immune response. However, PDAC patients often exhibit a CD16hiCD57hi phenotype of peripheral NK cells, with reduced cytotoxic activity, low IFN-γ, and high IL-10 levels, contributing to immune evasion. Tumor cells also limit NK cell movement to the tumors, further impairing their antitumor function [28].
Furthermore, clinical studies indicate that in tumor patients positive for Enterobacter and Enterobacteriaceae, the immune receptor TIGIT is significantly upregulated in NK cells. This upregulation has been associated with sustained clinical benefits, suggesting that the gut microbiota can modulate NK cell proliferation and function by influencing immune checkpoint receptors like TIGIT [30,31,32].
Boosting NK cells in PDAC treatment is a promising area of research. Tumor cells often block NK cell migration to the tumor, reducing their anti-tumor effects. Targeting these pathways may be crucial in enhancing NK cell responses in PDAC. While strategies like NK-cell-recruiting antibodies and engineered nanogels have shown potential, they have limitations. Therefore, targeting the microbiome to modulate NK cell activity may be a more practical and effective approach than direct NK cell therapies.
In addition to their role in cancer development, microorganisms also influence treatment-related side effects. Several studies have explored the role of microorganisms in the development of oral lesions and mucositis, a common side effect of cancer treatments [34,35,36].
This review explores how dysbiosis contributes to NK cell dysfunction in the PDAC TME. We hypothesize that microbial metabolites from dysbiosis suppress NK cell activity, promoting tumor progression and resistance to therapies. We also suggest that dysbiosis-NK cell interactions may contribute to treatment side effects. Targeting dysbiosis could restore NK cell function, improve therapeutic outcomes, and reduce side effects. This review will examine current evidence supporting these hypotheses and discuss strategies for microbiome manipulation as a novel approach to improving PDAC treatment.
2. Interaction Between Microbiota and NK Cell Function in PDAC
The interaction between the microbiota and NK cell function is important for understanding the progression of pancreatic ductal adenocarcinoma (PDAC) and treatment resistance (Figure 1). Clinically, analyzing buccal swabs and saliva, which contain leukocytes, including NK cells, in varying proportions, can provide insights into this interaction [37,38]. Additionally, using stool samples to detect microbiota can serve as a surrogate marker for natural killer cell activity (NKA) in patients. These samples help us understand the relationship between microbiota, NKA, and PDAC, including their impact on responses to cancer treatment and associated side effects [39,40,41,42,43,44,45,46,47].
2.1. Dysbiosis Induced NK Cell Dysfunction in PDAC
The microbiota profiles can differ significantly based on cancer type. In pancreatic cancer, notable changes in both oral and intrapancreatic microbiota have been observed. Oral dysbiosis is characterized by an altered abundance of microorganisms such as Porphyromonas gingivalis, Fusobacterium, Neisseria elongata, Streptococcus mitis, Bacteroides, Leptotrichia, Grabulitacetlla adiacens, and Aggregatibacter actinomycetemcomitans, while the intrapancreatic microbiota shows shifts in the levels of Gammaproteobacteria, Fusobacterium, Escherichia coli, and Bifidobacterium pseudolongum [10,17,19,49]. Additionally, the gut microbiota plays a crucial role in influencing the effectiveness of anti-cancer treatments and can significantly affect the quality of life of patients undergoing cancer therapy [40]. Therefore, the microbiome holds potential as a non-invasive tool for early cancer detection.
Many studies have investigated the role of microbiota in the initiation and progression of PDAC, with some suggesting NK cell suppression as a key factor. However, the role of dysbiosis and its interaction with NK cells remains underexplored (Figure 2).
NK cells play a key role in the early immune response to bacterial, viral, and fungal infections by activating macrophages, dendritic cells, and neutrophils before the adaptive immune system is triggered. While Toll-like receptors (TLRs) have been extensively studied in macrophages, recent research underscores their critical role in NK cells as part of the first-line defense against pathogens. TLRs on NK cells help these cells recognize and respond to infections, contributing to the body’s early defense mechanisms before the adaptive immune system becomes active. Different TLRs are expressed on NK cells, with TLR1 showing the highest expression, followed by TLR2, TLR3, TLR5, and TLR6. TLR ligands can activate NK cells directly or indirectly, though TLR9 expression is typically low or undetectable [41,42,43]. Commensal microbiotas instruct nonmucosal mononuclear phagocytes to prime NK cells. The NK cell-activating receptor NKp44 has been observed to directly interact with bacteria from the genus Mycobacterium, Nocardia farcinica, and Pseudomonas aeruginosa [42,43,44,45]. Microbiota-derived stimulator of interferon genes (STING) agonists stimulate intratumoral monocytes to produce type I interferon (IFN-I), influencing macrophage polarization and enhancing NK cell-DC communication. These findings suggest a microbiota-driven mechanism in the tumor microenvironment with potential implications for optimizing cancer therapies [44,45,46]. Lactobacillus plantarum effectively enhances the expression of the natural cytotoxicity receptor (NCR) family and IL-22 in NK cells [47]. A. muciniphila has been found to boost immune function by modulating specific cytokines and interacting with Toll-like receptors (TLRs), including TLR2 and TLR4. This bacterium also plays a protective role against pathogens, notably reducing inflammation triggered by Porphyromonas gingivalis, an opportunistic pathogen associated with periodontitis, which is implicated in the development of pancreatic cancer [46,47,48,49]. A preclinical study shows that gut microbiota influence PDAC progression by modulating NK cells and that microbiota-derived supernatant can enhance anti-tumor NKA. Thus, leveraging the gut microbiota to modulate the innate immune system offers potential for treating PDAC and could improve patient survival [39]. Another preclinical study revealed that Pseudoxanthomonas, Saccharopolyspora, and Streptomyces were associated with increased NK cell infiltration in colorectal cancer, whereas Anabaena sp. K119, an uncultured bacterium, P. putida str. KT2440, T. chromogena, and E. rectale/Roseburia showed a negative relationship with NK cell infiltration [49]. Further investigation revealed that extracts from B. pseudolongum could modify tumor-associated macrophages (TAMs), increasing tolerogenic cytokines such as IL-10 through Toll-like receptor (TLR) signaling activation. TLRs are crucial for the activation of innate immunity, as they identify specific patterns in microbial components. Previous studies have indicated that the polarization of macrophages impacts the activity of NK cells in PDAC [43,52].
Pathogenic components of Helicobacter pylori, such as ammonia, LPS, and inflammatory cytokines, contribute to pancreatic damage and carcinogenesis by activating NF-κB and AP-1 signaling, leading to KRAS mutations and persistent STAT3 activation, which all contributing to NK cell suppression [53,54,55].
Investigating how microbial imbalances affect NK cell function could reveal novel therapeutic targets and enhance the efficacy of existing treatments. By elucidating these interactions, we can develop more effective approaches to managing PDAC and ultimately improve patient outcomes.
2.2. Dysbiosis Induced Drug Resistance in PDAC
Tumor microbiome communities are significantly different between PDAC patients with long term survival and short-term survival [15,16]. Gut microbiota and tumor-associated microbes affect the anticancer therapies, highlighting their potential as novel targets for future interventions. The permeability of intestinal barrier may be a cause of bacterial dissemination into the pancreatic duct, which requires further validation. Intratumoral microbes could affect tumor susceptibility to chemotherapy and patient outcome [56,57,58,59].
Studies show that neoadjuvant chemotherapy and pre-operative biliary stenting significantly alter the microbiota composition, which can impact treatment responses and post operative complications in PDAC [58,59,60,61]. Studies have shown that gut bacteria play a crucial role in the effectiveness of PD-1-based immunotherapy, with dysbiosis linked to impaired immune cell function and reduced treatment response. In contrast, a balanced microbiome, particularly the presence of beneficial bacteria like Bifidobacterium, has been shown to enhance the efficacy of immunotherapy [64,65,66,67,68].
Many studies focus on gemcitabine-microbiome interaction. For example, E. coli influences the effectiveness of gemcitabine by promoting resistance and activating tumor cytotoxicity [67]. Gammaproteobacteria metabolize gemcitabine, inducing drug resistance via the bacterial enzyme cytidine deaminase (CDA), while CDA conversely directs 5- Fluorouracil (5-FU) to tumor cells, reducing its toxic effects and improving its pharmacokinetic properties. Interestingly the gene encoding CDA, has been identified as one of the top upregulated metabolic genes in tumors that are resistant to immunotherapy. High CDA expression in pancreatic cancer cells is associated with resistance to DNA-damaging agents. Inhibition of CDA, both in patient-derived primary cultures and orthotopic xenografts, increased replication stress and enhanced the sensitivity of pancreatic adenocarcinoma cells to oxaliplatin. The exact effects of CDA on NK cells are not fully known, but its role in nucleotide metabolism, immune regulation, and inflammation suggests it could influence NK cell activity [68,69,70].
Butyrate is a controversial substance in cancer therapy due to its complex role in treatment resistance. While gemcitabine alters the microbiota to produce butyrate, which can trigger inflammation, some studies suggest that intratumoral butyrate contributes to chemoresistance in colorectal cancer (CRC). Despite this, many recent studies have explored butyrate’s potential antitumor effects, especially in enhancing innate immune responses. However, most research has focused on CRC, so its effects on other cancer types remain unclear [72,73,74].
Platinum-based chemotherapy relies on the generation of reactive oxygen species (ROS) to exert its cytotoxic effects. The gut microbiome can enhance ROS production induced by oxaliplatin, potentially improving treatment effectiveness. However, secondary resistance to platinum-based chemotherapy may arise from interactions between ROS and NK cell activity. ROS can alter the surface charge of NK cells, making it more difficult for them to adhere to and kill cancer cells, which may reduce their effectiveness and contribute to resistance [75,76].
Hyperglycemia-induced dysbiosis can impair the response to chemotherapy drugs like gemcitabine and paclitaxel [77]. High glucose levels promote the proliferation and epithelial-mesenchymal transition of PDAC cells, while also reducing NK cell activity against these cells through the AMPK-Bmi1-GATA2-MICA/B pathway [78]. This suggests that elevated glucose helps create an immunosuppressive tumor microenvironment. PDAC tumor cells further evade IFN-I signaling, suppressing innate immune responses [79]. Metformin, an anti-hyperglycemic drug, has been shown to activate NK cells in PDAC by inhibiting AKT phosphorylation, which triggers the STING/IRF3/IFN-β pathway. This activation enhances immune responses and may improve the effectiveness of cancer treatment, potentially by modifying the duodenal microbiome [80,81].
Recent studies emphasize the need to expand microbiome research to include fungi, viruses, and protozoa, in addition to bacteria [82]. For example, fungal community has also been shown to influence the efficacy of gemcitabine-based chemotherapy. In vivo depletion of the mycobiome using antifungal treatments was found to enhance the response of orthotopic PDAC tumors to chemotherapy [83]. Malassezia fungi can suppress NK cell activity by activating the complement system, which releases molecules like C3a and C5a that reduce NK cell cytotoxicity. Chronic inflammation from complement activation may also recruit immune cells that suppress NK cells. Additionally, Malassezia may lower the levels of NKG2D receptors, supporting PDAC growth [83,84].
Further studies are needed to investigate NKA and its association with treatment resistance, particularly in the context of microbiota and NK cell interactions in PDAC.
2.3. Dysbiosis-Induced Treatment Side Effects
Recent studies have highlighted the potential of microbiota as a diagnostic tool for PDAC, but the prediction of treatment side effects remains less explored. [15,85]. Microbial dysbiosis could serve as an important marker for identifying side effects of cancer treatments [36]. Microbial communities in the oral cavity, gut, and tumor play a critical role in influencing the immune system and modulating responses to cancer therapies. Alterations in these microbial communities, known as dysbiosis, may serve as early indicators of treatment efficacy, prognosis, and potential side effects of cancer therapies [85,86]. The oral microbiota is the first point of exposure to environmental factors such as food, drinks, and medications [87]. Changes in the oral microbiota can trigger genetic and epigenetic changes in the buccal mucosa, potentially influencing the behavior of immune cells like NK cells, which are crucial in determining clinical outcomes during cancer treatment [89]. Dysbiosis in the oral microbiota may also lead to altered sensitivity of the oral mucosa to certain cancer treatments, contributing to side effects such as stomatitis (after 5-FU) or neuropathy (after oxaliplatin). Interestingly, some patients experience these side effects while others do not, suggesting that variations in the oral microbiota could predict who will develop these adverse effects [90,91].
Importantly, the oral microbiota is interconnected with both the gut and tumor microbiota. Changes in the oral microbiota can influence gut microbiota, which may, in turn, impact the tumor microenvironment. Gut microbiota composition has been linked to gastrointestinal side effects in patients treated with drugs such as irinotecan and 5-FU [90,91,92]. However, the presence or absence of these symptoms varies among patients, reinforcing the idea that microbiome composition may predict the risk of treatment-related side effects. Emerging evidence suggests that the gut microbiome and its interactions with the innate immune system play a significant role in gastrointestinal side effects development. The immune system can influence microbiome composition, while the microbiome, in turn, affects immune cell function [93]. Understanding these bidirectional interactions may improve risk prediction, personalize treatment strategies, and lead to targeted interventions to reduce or prevent chemotherapy- induced side effects in the future.
Current PDAC treatments, such as gemcitabine with nab-paclitaxel (nano albumin- bound paclitaxel) or FOLFIRINOX (a combination of 5-fluorouracil, leucovorin, irinotecan, and oxaliplatin), show objective response rates of 20-40% [93,94]. However, both oxaliplatin and nab-paclitaxel can cause neuropathy, which often leads to treatment discontinuation [96]. These chemotherapeutic agents trigger inflammatory factors that affect neurons and glial cells, contributing to the development of nociceptive pain in neuropathy [97]. 16S rDNA sequencing of rats with paclitaxel-induced peripheral neuropathy (PIPN) showed shifts in gut microbiota, with an increase in Bacteroides and UCG-005 genera and a reduction in Turicibacter, Clostridium sensu stricto 1, and Corynebacterium [98]. Despite extensive research on preventing oxaliplatin-induced neuropathy (OIPN), no definitive intervention has been proven effective. Most studies have focused on pharmacological treatments, while non- pharmacological approaches remain underexplored. More research is needed, particularly into alternative strategies such as microbiota modulation and modifying NK cell phenotypes, which could offer new solutions for mitigating OIPN [99]. Some studies have linked the occurrence of chemotherapy-induced neuropathy to changes in NK cell function.
NK cells, which are essential for immune surveillance, utilize several mechanisms to induce cytotoxicity. Key among these is the CD95/FasL system, TRAIL receptors, and NKG2D receptors on NK cells, which interact with ligands on stressed, tumor, or infected cells. However, NK cell activity is also regulated by inhibitory signals from MHC class I molecules, which can prevent the destruction of healthy cells [29,43,44].
3. Targeting Dysbiosis in PDAC for Overcoming Treatment Resistance and Managing Side Effects
Targeting dysbiosis in PDAC presents a promising strategy for improving treatment outcomes (Figure 2). Microbiome modulation can be achieved either directly—through interventions such as antibiotics or probiotics to alter microbial communities—or indirectly by modifying lifestyle factors, using organic/non-organic materials, vitamin therapy or herbal medicine [100,101,102,103,104,105,106,107,108,109].
Microbiome-based strategies have been increasingly explored in managing cancer treatment resistance and alleviating chemotherapy side effects [110,112]. These strategies aim to improve therapeutic outcomes by enhancing immune responses, particularly NK cell activation [113]. While lifestyle modifications, such as high-salt or the use of antioxidant vitamins and supplements have been suggested as non-invasive ways to modulate the microbiome and activate NK cells [39,114], this section focuses specifically on direct microbiome-targeting approaches. These include prebiotics, postbiotics, and antibiotics, which can modify the microbiome and enhance NK cell function to improve PDAC management.
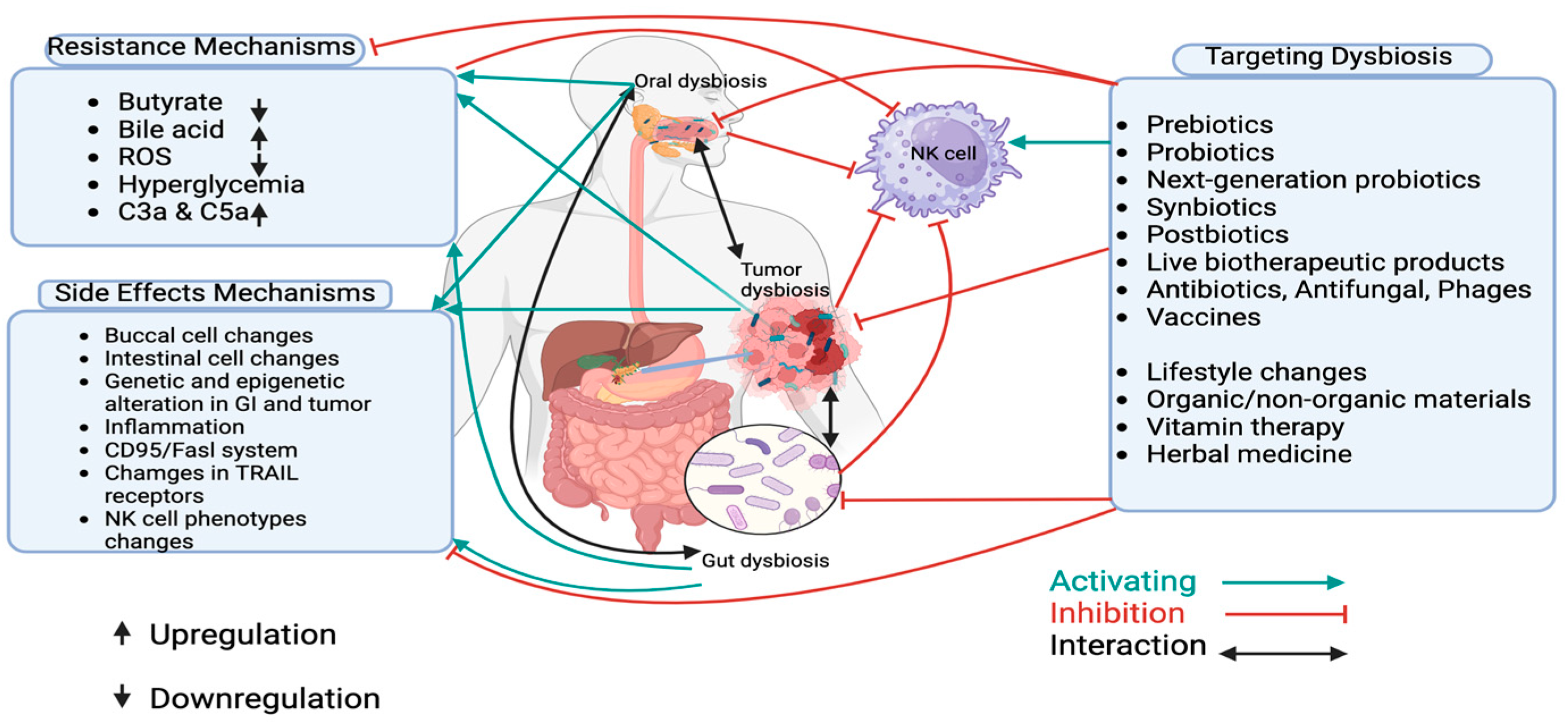
Figure 3.
Dysbiosis and Targeting Dysbiosis in PDAC. This figure summarizes the mechanisms of treatment resistance and side effects in pancreatic ductal adenocarcinoma (PDAC), focusing on the role of microbiota and NK cells. It illustrates how microbiota influence NK cell activity, contributing to both tumor immune evasion and resistance to therapy. Additionally, the figure highlights how microbiota dysbiosis can worsen treatment side effects, such as inflammation and gastrointestinal toxicity. Non-invasive strategies, including dietary changes, probiotics, and fecal microbiota transplantation (FMT), are shown as potential methods to restore microbiome balance, improve therapeutic response, and reduce adverse effects in PDAC.
Figure 3.
Dysbiosis and Targeting Dysbiosis in PDAC. This figure summarizes the mechanisms of treatment resistance and side effects in pancreatic ductal adenocarcinoma (PDAC), focusing on the role of microbiota and NK cells. It illustrates how microbiota influence NK cell activity, contributing to both tumor immune evasion and resistance to therapy. Additionally, the figure highlights how microbiota dysbiosis can worsen treatment side effects, such as inflammation and gastrointestinal toxicity. Non-invasive strategies, including dietary changes, probiotics, and fecal microbiota transplantation (FMT), are shown as potential methods to restore microbiome balance, improve therapeutic response, and reduce adverse effects in PDAC.
3.1. Prebiotics
Prebiotics are nutrients broken down by gut microbiota, impacting both the intestinal microenvironment and other organs. Prebiotics nourish probiotics and stimulate various functions in the host [115]. Many studies confirm the direct and indirect effect of prebiotics on the mediators of the immune response including NK cells [116]. A preclinical study revealed that some genes involved in PDAC tumor progression and inflammatory response were downregulated in a group of mice fed with a high level of prebiotic resistant starch diet compared to a control group [117]. Preclinical and clinical studies on the effects of prebiotics on NKA in PDAC and their correlation with overall survival would be extremely valuable, revealing potential therapeutic targets.
3.2. Probiotics
Several studies have shown that probiotics can reduce pancreaticoduodenectomy complications and inhibit cancer cell growth [118]. Many studies discussed the effects of probiotics on gastrointestinal cancers [119] and confirmed that Lactococcus lactis, a probiotic bacterium found in yogurt and cheese, can decrease various inflammatory agents that contribute to cancer, including both anti-inflammatory and pro-inflammatory molecules such as interleukin (IL)-6, IL-18, TNF-α, and NK cells [120,121]. In individuals with low NKA, consuming fermented milk containing Lactobacillus casei strain Shirota and Lactobacillus rhamnosus HN001 has significantly increased NKA, aiding in cancer prevention [122]. Ferrichrome, derived from the probiotic strain Lactobacillus casei ATCC334, has shown potential for combating PDAC [123].
Probiotics can aid in the eradication of H. pylori or in the management of H. pylori-related diseases through various mechanisms, such as enhancing the strength of the mucosal barrier, competing for adhesion sites, and modulating the immune response [124]. The role of NK cells in the local immune response to H. pylori infection has been discussed [125]. Ongoing research is exploring the role of NK cells activated by probiotics in enhancing the adaptive immune response against pathogens [126].
A meta-analysis suggested probiotics as a potential preventive measure for immune checkpoint inhibitor-induced diarrhea, especially in cases where the severity reaches grade ≥ 2 [127].
A randomized controlled trial found that probiotic supplementation may help reduce chemotherapy-induced cognitive impairment in breast cancer patients by decreasing synaptic damage, oxidative stress, and glial cell activation in the central nervous system [128]. Similarly, probiotics have shown promise in enhancing chemotherapy outcomes in PDAC. In a PDAC xenograft mouse model, the use of probiotics not only improved the effectiveness of gemcitabine but also helped mitigate chemotherapy-induced toxicity [129].
Furthermore, Bifidobacterium potentiates the efficacy of immune checkpoint inhibitor [130]. Oxaliplatin and cisplatin demonstrate enhanced efficacy in the presence of healthy microbiota [131].
Focusing on NK cell activation in PDAC patients is crucial, and employing probiotics may offer a safe and effective strategy to achieve this.
3.3. Next-Generation Probiotics
Based on comparative microbiota analyses, next-generation probiotics (NGPs) are live microorganisms that, when administered in sufficient amounts, provide health benefits to the host [132]. NGPs are identified using bioinformatics and next-generation sequencing. They meet novel food regulations for safety and toxicity and have clear modes of action. Unlike traditional probiotics, NGPs include diverse microbial species, target specific diseases, and can be used as biotherapeutics [133].
Selenium-enriched Bifidobacterium longum, a type of next-generation probiotics is a species of bacteria that belongs to the genus Bifidobacterium. This probiotic may help mitigate Irinotecan induced hepatotoxicity, reducing the inflammatory response by lowering levels of pro- inflammatory cytokines such as IL-1β and IL-18, while also promoting the expression of tight-junction proteins like occludin and ZO-1 [134].
A lower abundance of Akkermansia muciniphila causes multiple diseases, including cancer, in both mouse models and humans [135]. The use of Akkermansia as a representative example of NGPs needs further investigation.
3.4. Synbiotics
Synbiotics are combinations of prebiotics and probiotics designed to enhance the health of humans or animals [115]. Bifidobacterium lactis (B. lactis HN019) and Lactobacillus rhamnosus HN001 (L. rhamnosus HN001) have exhibited pronounced immunity-enhancing effects, particularly when combined with oligosaccharide-rich substrates, in both animal and human studies [136].
A recent study recruited ninety PDAC patients and randomly assigned them to one of three groups: placebo, probiotics, or synbiotics. Treatments were administered for 14 days pre-surgery and one-month post-surgery. The synbiotics group showed a significant increase in IFN-γ and a significant decrease in inflammatory cytokines compared to the other groups, suggesting that synbiotics enhance treatment efficacy and reduce side effects [137]. Interestingly, many studies have already suggested synbiotics as a promising adjunctive therapy for alleviating chemotherapy-associated symptoms in patients with solid tumors [138].
Emphasizing NK cell activation in PDAC through further clinical studies is essential, and utilizing synbiotics could offer a safe and effective strategy for achieving this.
3.5. Postbiotics
Postbiotics are beneficial substances like that short chain fatty acids (SCFA), exopolysaccharides, vitamins, phenols, bacterial lysates, supernatants, enzymes, and cell wall fragments derived from the gut microbiota [139]. Studies have shown that postbiotics could play a role in preventing and treating gastrointestinal cancer [140]. Although they can prevent cancer, postbiotics exhibit selective toxicity, targeting only certain tumor types [141].
The cell-free supernatant from Faecalibacterium prausnitzii A2-165 promoted the increase of anti-inflammatory cytokines (IL-10, TGF-β2, and IL-1Ra) and reduced levels of key pro-inflammatory cytokines like IL-6, TNF-α, and TNF-β [142].
3-indoleacetic acid (3-IAA) is also a type of postbiotic, produced by certain gut bacteria, and can influence various physiological processes. Tintelnot et al. revealed that the microbiota-derived metabolite 3-IAA significantly enhances the effectiveness of chemotherapy in treating PDAC [122]. Investigating whether 3-IAA can activate NK cells is an emerging area of research.
Butyrate is a postbiotic produced by gut bacteria during fiber fermentation, offering various health benefits. In PDAC, butyrate has been shown to boost gemcitabine induced apoptosis in cancer cells while also preserving intestinal mucosal integrity by reducing the abundance of pro-inflammatory microorganisms [70]. Supplementing conventional chemotherapy with butyrate or SCFA-producing bacteria, such as Faecalibacterium prausnitzii, may enhance clinical outcomes in PDAC [72,142] Interestingly, butyrate accumulates in higher concentrations and inhibits histone deacetylases, which are known to suppress NK cell activity, thereby enhancing its anticancer effects [143,144,145].
Further investigation into the application of postbiotics in the management of PDAC, particularly in combination with chemotherapy, is a very promising area of research.
3.6. Live Biotherapeutic Products
Live biotherapeutic products (LBPs) are products that contain live organisms used to prevent, treat, or cure human diseases. Unlike vaccines, they do not function by directly stimulating an immune response but rather by interacting with the body’s microbiota or other biological systems [130,146]. The effectiveness of LBPs can be influenced by various environmental factors, including diet [147]. LBPs can range from the transplantation of entire microbial communities to the introduction of individual bacterial strains, either engineered or non-engineered.
3.6.1. Fecal Microbiota Transplantation (FMT)
Fecal microbiota transplantation (FMT) is being explored as a therapeutic strategy to modulate the microbiota and treat dysbiosis, with potential benefits for cancer treatment outcomes and the reduction of treatment-related side effects [148,149].
A preclinical study involving mice with advanced PDAC, pre-treated with antibiotics, showed that fecal material from long-term PDAC survivors (over 5 years post-resection) led to smaller tumors compared to feces from healthy controls or advanced PDAC patients. The study also revealed higher levels of immune markers, IFN-γ and IL-2, in the long-term survivor group, indicating a favorable immune response. The study could identify a specific and protective bacterial signature in the stools of PDAC patients [15].
Another study found that transferring fecal material from PDAC-bearing mice (but not from healthy controls) accelerated tumor progression, highlighting the complexity of microbiome interactions in cancer [51].
FMT has also shown promise in managing infections and side effects related to cancer treatment. For example, FMT can help manage Pseudomonas aeruginosa infections resulting from chemotherapy-induced gut dysbiosis, such as 5-fluorouracil-induced mucositis [149]. Additionally, FMT using Lachnospiraceae and Roseburia species has been shown to reshape the gut microbiota, potentially alleviating chemotherapy-induced diarrhea [150]. Fur- thermore, FMT was found to reduce chemotherapy-induced peripheral neuropathy (PIPN) in rats by modulating astrocyte function and interfering with the TLR4/p38MAPK signaling pathway, suggesting its potential in managing neurological side effects [98].
It has also shown promise in reducing the adverse effects of immunotherapy and improving the efficacy of immune checkpoint inhibitors [150,151]. This suggests that FMT could potentially play a role in PDAC by addressing the loss of immunotherapy effectiveness associated with the microbiome. By modulating the microbiome, FMT may offer a strategy to improve the response to immunotherapy in PDAC, providing hope for enhancing treatment outcomes.
Given these promising findings, further research into FMT from long-term PDAC survivors or chemotherapy/Immunotherapy responders—particularly its impact on NK cell activity and clinical outcomes—is crucial for fully understanding its therapeutic potential.
3.6.2. Tumor-Colonizing Bacteria
Tumor-colonizing bacteria, a type of probiotic, prefer hypoxic environments and specific metabolites, allowing them to target tumors over healthy tissues [147]. Salmonella, Listeria, and Clostridium target tumors because they thrive in hypoxic environments and utilize specific metabolites. These bacteria can kill tumor cells directly or stimulate the immune system. For example, lipopolysaccharide triggers immune responses, and Listeria spp. increase IL-12 production, enhancing anti-tumor effects via NK cell responses [152] and Salmonella typhimurium inhibits tumor growth by inducing the pro-inflammatory cytokine interleukin-1β [153].
Another study showed that the efficacy of anti-PD-1/PD-L1 treatment in gastrointestinal cancer could be enhanced by SCFA-producing bacteria, such as Eubacterium, Lactobacillus, and Streptococcus [155].
Intratumoral pseudoxanthomas have been shown to be more prevalent in PDAC patients with long overall survival [15], suggesting that increasing their levels through tumor-colonizing probiotics could potentially improve clinical outcomes.
Further studies are needed to develop tumor-colonizing bacteria as effective NK cell stimulators in the management of PDAC. However, their effectiveness relies on successfully colonizing tumors and metastases, which can be challenging in certain tissues.
3.7. Antibiotics, Antifungal and Phage Therapy
In human PDAC, bacteria can metabolize gemcitabine, with this effect linked to intratumoral lipopolysaccharide (LPS). This can be countered using antibiotics [67]. Research in PDAC mouse models has shown that the gut and tumor microbiomes are distinct, and antibiotic treatment can alter the tumor microenvironment, boosting immune surveillance and enhancing responsiveness to immunotherapy [52,154,155,156,157].
Many studies have also discussed that depleting the gut microbiome with antibiotics can make PDAC tumors more sensitive to chemotherapy [155,156,157]. Supporting this, an independent retrospective study found that patients treated with antibiotics during gemcitabine-containing regimens had higher survival rates, suggesting that correcting dysbiosis with antibiotics could help mitigate treatment resistance [158].
However, antibiotics can also have negative effects. For example, Iida et al. found that antibiotics reduced the effectiveness of oxaliplatin and cisplatin [131]. Treatment with vancomycin decreased Firmicutes and increased Proteobacteria, leading to higher bile acid levels, which can promote PDAC carcinogenesis [160]. Various antibiotic approaches need to be investigated to deplete tumor-favoring bacteria or treat dysbiosis to enhance the treatment outcomes of PDAC. On the other hand, antifungal drugs have shown potential in improving PDAC treatment [161], with Malassezia identified as a possible target [162].
In addition to antibiotics and antifungal, phage therapy is emerging as a promising option. Phages are viruses that specifically target bacteria, with minimal impact on the surrounding microbiome. They could be used to target harmful bacteria or deliver drugs directly to tumors [163,164]. However, more research is needed to overcome the current limitations of phage therapy.
3.8. Vaccines
A Phase I clinical trial demonstrated anti-angiogenic activity of a prime-boost vaccination regimen with VXM01, an oral T cell vaccine targeting VEGFR2, in patients with advanced PDAC. An extension of the study, conducted three years later, confirmed the continued safety of the vaccine [165,166].
A phase II trial with 90 PDAC patients found that CRS-207 plus cyclophosphamide or the GVAX vaccine was safe. However, this combination did not improve OS compared to standard chemotherapy [167,168].
The use of vaccines in the management of PDAC and their effect on NK cell activities could be a promising area of research.
4. Discussion and Future Directions
Recent studies highlight the critical role of microbiome dysbiosis in PDAC, influencing clinical outcomes. Dysbiosis, characterized by microbial imbalances in the oral cavity, gut, tumor, and its microenvironment, has been implicated in PDAC progression. Specific bacteria, such as Porphyromonas gingivalis and Aggregatibacter actinomycetemcomitans, are associated with increased PDAC risk [15,16,52]. Additionally, microbial metabolites from Gammaproteobacteria can impair chemotherapy efficacy, contributing to drug resistance [15,67].
Interestingly, while Lactobacillus species are typically considered probiotics, recent research by Matsukawa suggests that their enrichment in PDAC might have a potential carcinogenic role, particularly in the tumor microenvironment [6]. This raises the question: which specific Lactobacillus species are enriched in the PDAC gut microbiome? Given the complexity of Lactobacillus strains, it is crucial to distinguish between these strains and other microbial communities, as well as to understand their differential roles in PDAC progression.
Most studies analyze microbiome data at the phylum, order, or genus level, but species- and strain-level profiling is essential for more precise insights. For instance, Listeria welshimeri (Serovar 6B SLCC 5334) provides more detailed information than genus-level data, underscoring the importance of focusing on species and strain-specific markers in PDAC management [147,153,167]. Future research should prioritize such detailed microbiome analyses to develop more accurate microbial signatures for PDAC treatment.
Although oral microbiome analysis remains underexplored, a study by Karat et al. found no significant differences in taxa abundance within the salivary microbiome but suggested stool microbiome signatures for PDAC [148]. Given the ease of saliva collection, further research into the oral microbiome is needed, and improving collection methods could provide valuable insights. Additionally, Negata et al. observed differences between the gut and oral microbiomes in PDAC [85], suggesting that these compartments, along with the intratumoral microbiome, should be studied simultaneously to better understand their dynamic interactions and role in PDAC mechanisms.
Notably, dysbiosis has also been shown to disrupt NK cell function, which is essential for immune surveillance and combating PDAC [26,27,28,29,30,39,41,42,43,44,45,46]; This dysregulation may serve as a potential biomarker for predicting treatment response, prognosis, and side effects in PDAC. Understanding how microbiota interact with NK cells could provide key insights into treatment toxicity and efficacy (Figure 4). Identifying microbial signatures linked to treatment resistance and side effects may guide personalized therapies, ultimately improving patient outcomes.
Therefore, the integration of non-invasive diagnostic tools, such as oral mucosa smears and stool samples, could offer a practical means to monitor microbiome composition and NK cell function [3,4,17,25,29]. This combined approach could help identify a unified microbial-NK cell signatures associated with treatment efficacy, failure, and side effects. Moreover, microbiome-targeting interventions, such as probiotics, synbiotics, antibiotics, and FMT, may be less invasive than direct NK cell augmentation strategies. These interventions could restore microbial balance and counteract the three main mechanisms of NK cell suppression (low NK cell proliferation, impaired cytotoxicity, and reduced tumor infiltration), thereby combating PDAC treatment resistance through a triple NK cell biomarker approach targeting tumor, microbiota, and NK cell mechanisms [169].
The complexity of microbiome-host interactions requires a combined biomarker approach, integrating microbiome and NK cell phenotyping to more accurately predict PDAC treatment outcomes. We hypothesize that changes in NK cell phenotype result from microbiome dysbiosis, and this needs to be validated. If confirmed, targeting the microbiome to modify NK cell activity could be a potential therapeutic strategy.
Microbiome-targeting interventions, including probiotics, synbiotics, antibiotics, and FMT, have the potential to enhance NK cell activation, restore microbial balance, and alleviate treatment-related side effects [126,136,137,138]. For example, FMT from long-term PDAC survivors could shift the gut microbiota towards a healthier state, potentially supporting immune function and improving treatment outcomes [15]. Antibiotics, on the other hand, can affect microbial populations, potentially altering inflammation and impacting the effectiveness of concurrent cancer therapies or their side effects [67,73,155,156,157,158,159,160].
However, several challenges remain in translating these findings into clinical practice, including individual variability in microbiota composition and difficulties in applying preclinical data to human cancer treatment. More research is needed to fully understand how dysbiosis contributes to NK cell dysfunction and treatment-related outcomes in PDAC. A promising approach is the use of non-invasive tools to monitor microbiome and NK cell phenotypes in PDAC patients. Oral mucosa smears and stool samples could be utilized to profile microbial communities and assess NK cell function, helping to identify microbial signatures linked to treatment response and treatment-related side effects. Tracking microbiota changes over time may also reveal dysbiosis patterns associated with treatment resistance or side effects.
However, one key challenge is establishing baseline microbiota profiles that distinguish normal microbial composition from cancer-associated dysbiosis. Comparative studies between healthy individuals and PDAC patients, responders versus non-responders, patients with long overall survival versus short overall survival , and those with and without side effects could identify early biomarkers of dysbiosis, leading to targeted microbiome interventions that improve treatment outcomes in PDAC. Integrating NK cell profiling with microbiome analysis overcomes the limitations of single-biomarker approaches, providing a more holistic view of the patient’s immune and microbial landscape. This integrated approach could also reduce the risks associated with microbiome-modifying therapies, such as antibiotic resistance or unintended microbial shifts, offering a better model for predicting treatment response and side effects.
Future research should focus on identifying specific microbial signatures that influence NK cell activity in PDAC. Investigating these signatures could uncover novel therapeutic targets aimed at enhancing immune responses and overcoming treatment resistance. Clinical trials validating microbiome-targeting therapies—such as prebiotics, antibiotics, and FMT—are critical to assess their safety, efficacy, and potential to improve PDAC treatment outcomes and survival.
5. Conclusion
Targeting dysbiosis in PDAC offers considerable potential for overcoming treatment resistance and managing treatment- related side effects. By modulating the microbiome and enhancing NK cell function, we can potentially improve patient outcomes and quality of life. Future research integrating NK cell phenotype profiling with microbiome diagnostics could lead to personalized treatment strategies, offering more effective and less toxic therapies for PDAC patients. Addressing the challenges of establishing baseline microbiota profiles, identifying predictive microbial signatures, and validating microbiome-targeting therapies—combined with NK cell profiling to create a unified biomarker signature—could transform PDAC management. This approach would provide safer, more tailored therapeutic options [Table 1] for patients, improving both treatment efficacy and overall outcomes.
Author Contributions
S.F contributes to the conceptualization and design of the review content. L.H.J contributes to supervision. S.F. wrote the first draft of the manuscript. Both authors contributed to the manuscript revision, read, and approved the final submitted version. L.H.J provided funding acquisition for publication.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable to this review article as it does not contain original datasets. All relevant information and sources are cited within the manuscript.
Acknowledgments
None.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Siegel, R.L.; Giaquinto, A.N.; Jemal, A. Cancer statistics, 2024. CA: A Cancer J. Clin. 2024, 74, 12–49. [CrossRef]
- Yousuf, S.; Qiu, M.; von Voithenberg, L.V.; Hulkkonen, J.; Macinkovic, I.; Schulz, A.R.; Hartmann, D.; Mueller, F.; Mijatovic, M.; Ibberson, D.; et al. Spatially Resolved Multi-Omics Single-Cell Analyses Inform Mechanisms of Immune Dysfunction in Pancreatic Cancer. Gastroenterology 2023, 165, 891–908.e14. [CrossRef]
- Wood LD, Canto MI, Jaffee EM and Simeone DM. “Pancreatic Cancer: pathogenesis, screening, diagnosis, and treatment”. Gastroenterology, vol. 163, no. 2, 386–402, 2022.
- Poggi, A.; Benelli, R.; Venè, R.; Costa, D.; Ferrari, N.; Tosetti, F.; Zocchi, M.R. Human Gut-Associated Natural Killer Cells in Health and Disease. Front. Immunol. 2019, 10, 961. [CrossRef]
- K Søreide, W Ismail, M Roalsø, J Ghotbi and C Zaharia. “Early diagnosis of pancreatic Cancer: clinical premonitions, timely precursor detection and increased curative-intent surgery”. Cancer control: journal of the Moffitt Cancer Center, vol. 30, 10732748231154711–10732748231154711, 2023.
- Matsukawa, H.; Iida, N.; Kitamura, K.; Terashima, T.; Seishima, J.; Makino, I.; Kannon, T.; Hosomichi, K.; Yamashita, T.; Sakai, Y.; et al. Dysbiotic gut microbiota in pancreatic cancer patients form correlation networks with the oral microbiota and prognostic factors. 2021, 11, 3163–+.
- Benešová, I.; Křížová, .; Kverka, M. Microbiota as the unifying factor behind the hallmarks of cancer. J. Cancer Res. Clin. Oncol. 2023, 149, 14429–14450. [CrossRef]
- Ren, Z.; Jiang, J.; Xie, H.; Li, A.; Lu, H.; Xu, S.; Zhou, L.; Zhang, H.; Cui, G.; Chen, X.; et al. Gut microbial profile analysis by MiSeq sequencing of pancreatic carcinoma patients in China. Oncotarget 2017, 8, 95176–95191. [CrossRef]
- Bracci PM. “Oral Health and the Oral Microbiome in Pancreatic Cancer: An Overview of Epidemiological Studies.”. Cancer J., vol. 23, no. 6, 2017.
- Ji, H.; Jiang, Z.; Wei, C.; Ma, Y.; Zhao, J.; Wang, F.; Zhao, B.; Wang, D.; Tang, D. Intratumoural microbiota: from theory to clinical application. Cell Commun. Signal. 2023, 21, 1–22. [CrossRef]
- Qian J, Zhang X, Wei B and Tang Z and Zhang B. “The correlation between gut and intra-tumor microbiota and PDAC: Etiology, diagnostics and therapeutics.”. Biochim Biophys Acta Rev Cancer, vol. 1878, no. 5, 2023.
- Cruz, M.S.; Tintelnot, J.; Gagliani, N. Roles of microbiota in pancreatic cancer development and treatment. Gut Microbes 2024, 16, 2320280. [CrossRef]
- Ames, N.J.; Ranucci, A.; Moriyama, B.; Wallen, G.R. The Human Microbiome and Understanding the 16S rRNA Gene in Translational Nursing Science. Nurs. Res. 2017, 66, 184–197. [CrossRef]
- Fang, Y.; Yang, G.; Yang, J.; Ren, J.; You, L.; Zhao, Y. Human microbiota colonization and pancreatic ductal carcinoma. Crit. Rev. Microbiol. 2022, 49, 455–468. [CrossRef]
- Riquelme, E.; Zhang, Y.; Zhang, L.; Montiel, M.; Zoltan, M.; Dong, W.; Quesada, P.; Sahin, I.; Chandra, V.; Lucas, A.S.; et al. Tumor Microbiome Diversity and Composition Influence Pancreatic Cancer Outcomes. Cell 2019, 178, 795–806.e12. [CrossRef]
- Huang, Y.; Zhu, N.; Zheng, X.; Liu, Y.; Lu, H.; Yin, X.; Hao, H.; Tan, Y.; Wang, D.; Hu, H.; et al. Intratumor Microbiome Analysis Identifies Positive Association Between Megasphaera and Survival of Chinese Patients With Pancreatic Ductal Adenocarcinomas. Front. Immunol. 2022, 13, 785422. [CrossRef]
- Bartolini, I.; Nannini, G.; Risaliti, M.; Matarazzo, F.; Moraldi, L.; Ringressi, M.N.; Taddei, A.; Amedei, A. Impact of microbiota-immunity axis in pancreatic cancer management. World J. Gastroenterol. 2022, 28, 4527–4539. [CrossRef]
- Narunsky-Haziza, L.; Sepich-Poore, G.D.; Livyatan, I.; Asraf, O.; Martino, C.; Nejman, D.; Gavert, N.; Stajich, J.E.; Amit, G.; González, A.; et al. Pan-cancer analyses reveal cancer-type-specific fungal ecologies and bacteriome interactions. Cell 2022, 185, 3789–3806.e17. [CrossRef]
- Chakladar, J.; Kuo, S.Z.; Castaneda, G.; Li, W.T.; Gnanasekar, A.; Yu, M.A.; Chang, E.Y.; Wang, X.Q.; Ongkeko, W.M. The Pancreatic Microbiome Is Associated with Carcinogenesis and Worse Prognosis in Males and Smokers. Cancers 2020, 12, 2672. [CrossRef]
- Wei, A.-L.; Li, M.; Li, G.-Q.; Wang, X.; Hu, W.-M.; Li, Z.-L.; Yuan, J.; Liu, H.-Y.; Zhou, L.-L.; Li, K.; et al. Oral microbiome and pancreatic cancer. World J. Gastroenterol. 2020, 26, 7679–7692. [CrossRef]
- Zhang, X.; Shi, L.; Lu, X.; Zheng, W.; Shi, J.; Yu, S.; Feng, H.; Yu, Z. Bile Acids and Liver Cancer: Molecular Mechanism and Therapeutic Prospects. Pharmaceuticals 2024, 17, 1142. [CrossRef]
- Wei X, Li W and Ma Y et al. “Endogenous Propionibacterium acnes Promotes Ovarian Cancer Progression via Regulating Hedgehog Signalling Pathway”. Cancers (Basel), vol. 14, no. 21, 2022.
- Mohelnikova-Duchonova, B.; Kocik, M.; Duchonova, B.; Brynychova, V.; Oliverius, M.; Hlavsa, J.; Honsova, E.; Mazanec, J.; Kala, Z.; Ojima, I.; et al. Hedgehog pathway overexpression in pancreatic cancer is abrogated by new-generation taxoid SB-T-1216. Pharmacogenom. J. 2017, 17, 452–460. [CrossRef]
- He, C.; Wang, D.; Shukla, S.K.; Hu, T.; Thakur, R.; Fu, X.; King, R.J.; Kollala, S.S.; Attri, K.S.; Murthy, D.; et al. Vitamin B6 Competition in the Tumor Microenvironment Hampers Antitumor Functions of NK Cells. Cancer Discov. 2023, 14, 176–193. [CrossRef]
- Wen, M.; Li, Y.; Qin, X.; Qin, B.; Wang, Q. Insight into Cancer Immunity: MHCs, Immune Cells and Commensal Microbiota. Cells 2023, 12, 1882. [CrossRef]
- Knudson, A.G., Jr. Mutation and cancer: statistical study of retinoblastoma. Proc. Natl. Acad. Sci. USA 1971, 68, 820–823. [CrossRef]
- Gautam, S.K.; Batra, S.K.; Jain, M. Molecular and metabolic regulation of immunosuppression in metastatic pancreatic ductal adenocarcinoma. Mol. Cancer 2023, 22, 1–30. [CrossRef]
- Luo W and Wen T and Qu X. “Tumor immune microenvironment-based therapies in pancreatic ductal adenocarcinoma: time to update the concept.”. J Exp Clin Cancer Res, vol. 43, no. 1, 2024.
- Marcon, F.; Zuo, J.; Pearce, H.; Nicol, S.; Margielewska-Davies, S.; Farhat, M.; Mahon, B.; Middleton, G.; Brown, R.; Roberts, K.J.; et al. NK cells in pancreatic cancer demonstrate impaired cytotoxicity and a regulatory IL-10 phenotype. OncoImmunology 2020, 9, 1845424. [CrossRef]
- Liu, C.; Fu, L.; Wang, Y.; Yang, W. Influence of the gut microbiota on immune cell interactions and cancer treatment. J. Transl. Med. 2024, 22, 1–15. [CrossRef]
- Tassone, E.; Muscolini, M.; van Montfoort, N.; Hiscott, J. Oncolytic virotherapy for pancreatic ductal adenocarcinoma: A glimmer of hope after years of disappointment?. Cytokine Growth Factor Rev. 2020, 56, 141–148. [CrossRef]
- Gur, C.; Ibrahim, Y.; Isaacson, B.; Yamin, R.; Abed, J.; Gamliel, M.; Enk, J.; Bar-On, Y.; Stanietsky-Kaynan, N.; Coppenhagen-Glazer, S.; et al. Binding of the Fap2 Protein of Fusobacterium nucleatum to Human Inhibitory Receptor TIGIT Protects Tumors from Immune Cell Attack. Immunity 2015, 42, 344–355. [CrossRef]
- Tsakmaklis, A.; Farowski, F.; Zenner, R.; Lesker, T.R.; Strowig, T.; Schlößer, H.; Lehmann, J.; von Bergwelt-Baildon, M.; Mauch, C.; Schlaak, M.; et al. TIGIT+ NK cells in combination with specific gut microbiota features predict response to checkpoint inhibitor therapy in melanoma patients. BMC Cancer 2023, 23, 1–11. [CrossRef]
- K France and A Villa. “Acute oral lesions”. Dermatol. Clin, vol. 38, 2020.
- Gasmi Benahmed, A Noor, S Menzel, A Gasmi and A. “Oral aphthous: Pathophysiology, clinical aspects and medical treatment”. Arch. Razi Inst, vol. 76, 1155–1163, 2021.
- Hong, B.-Y.; Sobue, T.; Choquette, L.; Dupuy, A.K.; Thompson, A.; Burleson, J.A.; Salner, A.L.; Schauer, P.K.; Joshi, P.; Fox, E.; et al. Chemotherapy-induced oral mucositis is associated with detrimental bacterial dysbiosis. Microbiome 2019, 7, 1–18. [CrossRef]
- Theda, C.; Hwang, S.H.; Czajko, A.; Loke, Y.J.; Leong, P.; Craig, J.M. Quantitation of the cellular content of saliva and buccal swab samples. Sci. Rep. 2018, 8, 1–8. [CrossRef]
- Farrell, J.J.; Zhang, L.; Zhou, H.; Chia, D.; Elashoff, D.; Akin, D.; Paster, B.J.; Joshipura, K.; Wong, D.T.W. Variations of oral microbiota are associated with pancreatic diseases including pancreatic cancer. Gut 2012, 61, 582–588. [CrossRef]
- Rizvi, Z.A.; Dalal, R.; Sadhu, S.; Kumar, Y.; Kumar, S.; Gupta, S.K.; Tripathy, M.R.; Rathore, D.K.; Awasthi, A. High-salt diet mediates interplay between NK cells and gut microbiota to induce potent tumor immunity. Sci. Adv. 2021, 7. [CrossRef]
- G Pourali, D Kazemi and A S Chadeganipour. “Microbiome as a biomarker and therapeutic target in pan- creatic cancer”. BMC Microbiol, vol. 24, 16–16, 2024.
- Yu, Q.; Newsome, R.C.; Beveridge, M.; Hernandez, M.C.; Gharaibeh, R.Z.; Jobin, C.; Thomas, R.M. Intestinal microbiota modulates pancreatic carcinogenesis through intratumoral natural killer cells. Gut Microbes 2022, 14, 2112881. [CrossRef]
- X Zhang, Q Liu, Q Liao and Y Zhao. “Pancreatic Cancer, gut microbiota, and therapeutic efficacy”. J Cancer, vol. 11, 2749–58, 2020.
- Noh, J.-Y.; Yoon, S.R.; Kim, T.-D.; Choi, I.; Jung, H. Toll-Like Receptors in Natural Killer Cells and Their Application for Immunotherapy. J. Immunol. Res. 2020, 2020, 1–9. [CrossRef]
- Ganal, S.C.; Sanos, S.L.; Kallfass, C.; Oberle, K.; Johner, C.; Kirschning, C.; Lienenklaus, S.; Weiss, S.; Staeheli, P.; Aichele, P.; et al. Priming of Natural Killer Cells by Nonmucosal Mononuclear Phagocytes Requires Instructive Signals from Commensal Microbiota. 2012, 37, 171–186. [CrossRef]
- Esin, S.; Batoni, G.; Counoupas, C.; Stringaro, A.; Brancatisano, F.L.; Colone, M.; Maisetta, G.; Florio, W.; Arancia, G.; Campa, M. Direct Binding of Human NK Cell Natural Cytotoxicity Receptor NKp44 to the Surfaces of Mycobacteria and Other Bacteria. Infect. Immun. 2008, 76, 1719–1727. [CrossRef]
- Lam KC, Araya RE, Huang A et al. “Microbiota triggers STING-type I IFN-dependent monocyte repro- gramming of the tumor microenvironment. Cell, 2021.
- Qiu Y, Jiang Z and Hu S, Wang L, Ma X and Yang X. “Lactobacillus plantarum enhanced IL-22 produc- tion in Natural Killer (NK) cells that protect the integrity of intestinal epithelial cell barrier damaged by enterotoxigenic Escherichia coli”. Int J Mol Sci, 2017.
- K Kaz´mierczak-Siedlecka, J Ruszkowski, K Skonieczna-Z˙ ydecka et al. “Gastrointestinal cancers: the role of microbiota in carcinogenesis and the role of probiotics and microbiota in the anti-cancer therapy efficacy”. Cent Eur J of Immunol, vol. 45, no. 4, 476–487, 2020.
- Tan, Q.; Ma, X.; Yang, B.; Liu, Y.; Xie, Y.; Wang, X.; Yuan, W.; Ma, J. Periodontitis pathogen Porphyromonas gingivalis promotes pancreatic tumorigenesis via neutrophil elastase from tumor-associated neutrophils. Gut Microbes 2022, 14, 2073785. [CrossRef]
- Seifert, L.; Werba, G.; Tiwari, S.; Ly, N.N.G.; Alothman, S.; Alqunaibit, D.; Avanzi, A.; Barilla, R.; Daley, D.; Greco, S.H.; et al. The necrosome promotes pancreatic oncogenesis via CXCL1 and Mincle-induced immune suppression. Nature 2016, 532, 245–249. [CrossRef]
- Qiu, J.; Jiang, Y.; Ye, N.; Jin, G.; Shi, H.; Qian, D. Leveraging the intratumoral microbiota to treat human cancer: are engineered exosomes an effective strategy?. J. Transl. Med. 2024, 22, 1–15. [CrossRef]
- Pushalkar, S.; Hundeyin, M.; Daley, D.; Zambirinis, C.P.; Kurz, E.; Mishra, A.; Mohan, N.; Aykut, B.; Usyk, M.; Torres, L.E.; et al. The Pancreatic Cancer Microbiome Promotes Oncogenesis by Induction of Innate and Adaptive Immune Suppression. Cancer Discov. 2018, 8, 403–416. [CrossRef]
- Hamel, Z.; Sanchez, S.; Standing, D.; Anant, S. Role of STAT3 in pancreatic cancer. Explor. Target. Anti-tumor Ther. 2024, 5, 20–33. [CrossRef]
- P B Dodhiawala, B Zhang et al. “Targeting the KRAS oncoprotein sensitizes pancreatic cancer to NK cell therapyJournal for ImmunoTherapy of”. Cancer, vol. 12, 2024.
- Schnoegl, D.; Hiesinger, A.; Huntington, N.D.; Gotthardt, D. AP-1 transcription factors in cytotoxic lymphocyte development and antitumor immunity. Curr. Opin. Immunol. 2023, 85, 102397. [CrossRef]
- Zhao L Y, Mei J X, Yu G et al. “Role of the gut microbiota in anticancer therapy: from molecular mechanisms to clinical applications. ”. Sig Transduct Target Ther, vol. 8, no. 201, 2023.
- Vogtmann, E.; Han, Y.; Caporaso, J.G.; Bokulich, N.; Mohamadkhani, A.; Moayyedkazemi, A.; Hua, X.; Kamangar, F.; Wan, Y.; Suman, S.; et al. Oral microbial community composition is associated with pancreatic cancer: A case-control study in Iran. Cancer Med. 2019, 9, 797–806. [CrossRef]
- Dong, J.; Gao, H.-L.; Wang, W.-Q.; Yu, X.-J.; Liu, L. Bidirectional and dynamic interaction between the microbiota and therapeutic resistance in pancreatic cancer. Biochim. et Biophys. Acta (BBA) - Rev. Cancer 2021, 1875, 188484. [CrossRef]
- Oar, A.; Lee, M.; Le, H.; Wilson, K.; Aiken, C.; Chantrill, L.; Simes, J.; Nguyen, N.; Barbour, A.; Samra, J.; et al. AGITG MASTERPLAN: a randomised phase II study of modified FOLFIRINOX alone or in combination with stereotactic body radiotherapy for patients with high-risk and locally advanced pancreatic cancer. BMC Cancer 2021, 21, 1–11. [CrossRef]
- N Goel, A Nadler, S Reddy, J P Hoffman and H A Pitt. “Biliary Microbiome in Pancreatic Cancer: Alter- ations with Neoadjuvant Therapy”. HPB Off. J. Int. Hepato Pancreato Biliary Assoc, vol. 21, 1753–1760, 2019.
- Nadeem, S.O.; Jajja, M.R.; Maxwell, D.W.; Pouch, S.M.; Sarmiento, J.M. Neoadjuvant chemotherapy for pancreatic cancer and changes in the biliary microbiome. Am. J. Surg. 2020, 222, 3–7. [CrossRef]
- Hank, T.; Sandini, M.; Ferrone, C.R.; Rodrigues, C.; Weniger, M.; Qadan, M.; Warshaw, A.L.; Lillemoe, K.D.; Castillo, C.F.-D. Association Between Pancreatic Fistula and Long-term Survival in the Era of Neoadjuvant Chemotherapy. JAMA Surg. 2019, 154, 943–951. [CrossRef]
- H R Shrader, A M Miller, A Tomanek-Chalkley et al. “Effect of Bacterial Contamination in Bile on Pancre- atic Cancer Cell Survival”. Surgery, vol. 169, 617–622, 2021.
- Routy, B.; le Chatelier, E.; DeRosa, L.; Duong, C.P.M.; Alou, M.T.; Daillère, R.; Fluckiger, A.; Messaoudene, M.; Rauber, C.; Roberti, M.P.; et al. Gut microbiome influences efficacy of PD-1–based immunotherapy against epithelial tumors. Science 2018, 359, 91–97. [CrossRef]
- Sivan A, Corrales L, Hubert N et al. “Commensal bifidobacterium promotes antitumor immunity and facili- tates anti-pd-L1 efficacy”. Science, vol. 350, no. 6264, 2015.
- Gnanamony, M.; Gondi, C.S. Chemoresistance in pancreatic cancer: Emerging concepts. Oncol. Lett. 2017, 13, 2507–2513. [CrossRef]
- Geller, L.T.; Barzily-Rokni, M.; Danino, T.; Jonas, O.H.; Shental, N.; Nejman, D.; Gavert, N.; Zwang, Y.; Cooper, Z.A.; Shee, K.; et al. Potential role of intratumor bacteria in mediating tumor resistance to the chemotherapeutic drug gemcitabine. Science 2017, 357, 1156–1160. [CrossRef]
- Asa El-Sayed, N Z Mohamed, M A Yassin et al. “Microbial cytosine deaminase is a programmable anti- cancer prodrug mediating enzyme: antibody, and gene directed enzyme prodrug therapy”. Heliyon, vol. 8, 9508425–9508425, 2022.
- T Scolaro, M Manco and M Pecqueux. “Nucleotide metabolism in cancer cells fuels a UDP-driven macrophage crosstalk, promoting immunosuppression and immunotherapy resistance”. Nat Cancer, vol. 5, 1206–1226, 2024.
- Audrey Lumeau et al. “Cytidine Deaminase Resolves Replicative Stress and Protects Pancreatic Cancer from DNA-Targeting Drugs.”. Cancer research, vol. 84, 1013–1028, 2024.
- Vieira, V.C.; Soares, M.A. The Role of Cytidine Deaminases on Innate Immune Responses against Human Viral Infections. BioMed Res. Int. 2013, 2013, 1–18. [CrossRef]
- A Panebianco, F Villani, F Pisati et al. “Butyrate, a postbiotic of intestinal bacteria, affects pancreatic cancer and gemcitabine response in in vitro and in vivo models ”. Biomed Pharmacother, vol. 151, 2022.
- Panebianco, C.; Adamberg, K.; Jaagura, M.; Copetti, M.; Fontana, A.; Adamberg, S.; Kolk, K.; Vilu, R.; Andriulli, A.; Pazienza, V. Influence of gemcitabine chemotherapy on the microbiota of pancreatic cancer xenografted mice. Cancer Chemother. Pharmacol. 2018, 81, 773–782. [CrossRef]
- Linsheng Xu, Bingde Hu, Jingli He, Xin Fu and Na Liu. “Intratumor microbiome-derived butyrate promotes chemo-resistance in colorectal cancer”. Front. Pharmacol., vol. 15, 2024.
- Udayasuryan, Barath et al. “Fusobacterium nucleatum induces proliferation and migration in pancreatic cancer cells through host autocrine and paracrine signaling.” Science signaling vol. 15,756 (2022): eabn4948. Iida, A Dzutsev, C A Stewart et al. “Commensal Bacteria Control Cancer Response to Therapy by Modu- lating the Tumor Microenvironment”. Science, vol. 342, 967–970, 2013. [CrossRef]
- Nakamura, K.; Matsunaga, K.-I. Susceptibility of Natural Killer (NK) Cells to Reactive Oxygen Species (ROS) and Their Restoration by the Mimics of Superoxide Dismutase (SOD). Cancer Biother. Radiopharm. 1998, 13, 275–290. [CrossRef]
- Kesh, K.; Mendez, R.; Abdelrahman, L.; Banerjee, S.; Banerjee, S. Type 2 diabetes induced microbiome dysbiosis is associated with therapy resistance in pancreatic adenocarcinoma. Microb. Cell Factories 2020, 19, 75. [CrossRef]
- Wu, S.; Zhang, H.; Gao, C.; Chen, J.; Li, H.; Meng, Z.; Bai, J.; Shen, Q.; Wu, H.; Yin, T. Hyperglycemia Enhances Immunosuppression and Aerobic Glycolysis of Pancreatic Cancer Through Upregulating Bmi1-UPF1-HK2 Pathway. Cell. Mol. Gastroenterol. Hepatol. 2022, 14, 1146–1165. [CrossRef]
- Cattolico, C.; Bailey, P.; Barry, S.T. Modulation of Type I Interferon Responses to Influence Tumor-Immune Cross Talk in PDAC. Front. Cell Dev. Biol. 2022, 10, 816517. [CrossRef]
- Dong TS, Chang H H, Hauer M et al. “Metformin alters the duodenal microbiome and decreases the inci- dence of pancreatic ductal adenocarcinoma promoted by diet-induced obesity.”. Am. J. Physiol. Gastrointest. Liver Physiol., 2019.
- D Ren, G Qin, J Zhao et al. “Metformin Activates the STING/IRF3/IFN-β Pathway by Inhibiting AKT Phosphorylation in Pancreatic Cancer”. Am. J. Cancer Res, vol. 10, no. 9, 2851–2864, 2020.
- R Vemuri, E M Shankar, M Chieppa, R Eri and K Kavanagh. “Beyond Just Bacteria: Functional Biomes in the Gut Ecosystem Including Virome, Mycobiome”. Archaeome and Helminths. Microorganisms, vol. 8, 483–483, 2020.
- Aykut, B.; Pushalkar, S.; Chen, R.; Li, Q.; Abengozar, R.; Kim, J.I.; Shadaloey, S.A.; Wu, D.; Preiss, P.; Verma, N.; et al. The fungal mycobiome promotes pancreatic oncogenesis via activation of MBL. Nature 2019, 574, 264–267. [CrossRef]
- Alam, A.; Levanduski, E.; Denz, P.; Villavicencio, H.S.; Bhatta, M.; Alhorebi, L.; Zhang, Y.; Gomez, E.C.; Morreale, B.; Senchanthisai, S.; et al. Fungal mycobiome drives IL-33 secretion and type 2 immunity in pancreatic cancer. Cancer Cell 2022, 40, 153–167.e11. [CrossRef]
- Nagata, N.; Nishijima, S.; Kojima, Y.; Hisada, Y.; Imbe, K.; Miyoshi-Akiyama, T.; Suda, W.; Kimura, M.; Aoki, R.; Sekine, K.; et al. Metagenomic Identification of Microbial Signatures Predicting Pancreatic Cancer From a Multinational Study. Gastroenterology 2022, 163, 222–238. [CrossRef]
- H Y Temel, Ö Kaymak, S Kaplan et al. “Role of microbiota and microbiota-derived short-chain fatty acids in PDAC”. Cancer Medicine, vol. 12, no. 5, 5661–75, 2023.
- Yu, Q.; Jobin, C.; Thomas, R.M. Implications of the microbiome in the development and treatment of pancreatic cancer: Thinking outside of the box by looking inside the gut. Neoplasia 2021, 23, 246–256. [CrossRef]
- Wang, S.-S.; Tang, Y.-L.; Pang, X.; Zheng, M.; Liang, X.-H. The maintenance of an oral epithelial barrier. Life Sci. 2019, 227, 129–136. [CrossRef]
- Sima Ataollahi Eshkoor and Sara Fanijavadi. “Dysbiosis-epigenetics-immune system interaction and ageing health problems”. Journal of medical microbiology, vol. 73, no. 11, 2024.
- Chander MP, C. Kartick, J. Gangadhar, P. Vijayachari, Ethno medicine and healthcare practices among Nicobarese of Car Nicobar - an indigenous tribe of Andaman and Nicobar Islands, J. Ethnopharmacol. 158 (2014) 18–24. https://doi. org/10.1016/j.jep.2014.09.046.
- Kawasaki, Y.; Kakimoto, K.; Tanaka, Y.; Shimizu, H.; Nishida, K.; Numa, K.; Kinoshita, N.; Tatsumi, Y.; Nakazawa, K.; Koshiba, R.; et al. Relationship between Chemotherapy-Induced Diarrhea and Intestinal Microbiome Composition. Digestion 2023, 104, 357–369. [CrossRef]
- A M Stringer, R J Gibson, R M Logan et al. “Faecal Microflora and Beta-Glucuronidase Expression Are Altered in an Irinotecan-Induced Diarrhea Model in Rats”. Cancer Biol. Ther, 1919.
- Secombe, K.R.; Coller, J.K.; Gibson, R.J.; Wardill, H.R.; Bowen, J.M. The bidirectional interaction of the gut microbiome and the innate immune system: Implications for chemotherapy-induced gastrointestinal toxicity. Int. J. Cancer 2018, 144, 2365–2376. [CrossRef]
- Kang, J.; Hwang, I.; Yoo, C.; Kim, K.-P.; Jeong, J.H.; Chang, H.-M.; Lee, S.S.; Park, D.H.; Song, T.J.; Seo, D.W.; et al. Nab-paclitaxel plus gemcitabine versus FOLFIRINOX as the first-line chemotherapy for patients with metastatic pancreatic cancer: retrospective analysis. Investig. New Drugs 2018, 36, 732–741. [CrossRef]
- Tong, H.; Fan, Z.; Liu, B.; Lu, T. The benefits of modified FOLFIRINOX for advanced pancreatic cancer and its induced adverse events: a systematic review and meta-analysis. Sci. Rep. 2018, 8, 1–8. [CrossRef]
- Deirdre R Pachman et al. “Comparison of oxaliplatin and paclitaxel-induced neuropathy ”. Supportive care in cancer, vol. 24, 5059–5068, 2016.
- Jaggi, A.S.; Singh, N. Mechanisms in cancer-chemotherapeutic drugs-induced peripheral neuropathy. Toxicology 2012, 291, 1–9. [CrossRef]
- Toxicology, vol. 291, 2012.
- H Shi, M Chen, C Zheng, B Yinglin and B Zhu. “Fecal Microbiota Transplantation Alleviated Paclitaxel- Induced Peripheral Neuropathy by Interfering with Astrocytes and TLR4/p38MAPK Pathway in Rats”. J Pain Res, vol. 16, 2419–2432, 2023.
- Peng, S.; Ying, A.F.; Chan, N.J.H.; Sundar, R.; Soon, Y.Y.; Bandla, A. Prevention of Oxaliplatin-Induced Peripheral Neuropathy: A Systematic Review and Meta-Analysis. Front. Oncol. 2022, 12, 731223. [CrossRef]
- Davies, A.J.; Rinaldi, S.; Costigan, M.; Oh, S.B. Cytotoxic Immunity in Peripheral Nerve Injury and Pain. Front. Neurosci. 2020, 14, 142. [CrossRef]
- Front Neurosci, vol. 14, 142–142, 2020.
- Karaliute, I.; Ramonaite, R.; Bernatoniene, J.; Petrikaite, V.; Misiunas, A.; Denkovskiene, E.; Razanskiene, A.; Gleba, Y.; Kupcinskas, J.; Skieceviciene, J. Reduction of gastrointestinal tract colonization by Klebsiella quasipneumoniae using antimicrobial protein KvarIa. Gut Pathog. 2022, 14, 1–11. [CrossRef]
- Knudson, A.G., Jr. Mutation and cancer: statistical study of retinoblastoma. Proc. Natl. Acad. Sci. USA 1971, 68, 820–823. [CrossRef]
- Li, H.; Ding, J.; Liu, C.; Huang, P.; Yang, Y.; Jin, Z.; Qin, W. Carvacrol Treatment Reduces Decay and Maintains the Postharvest Quality of Red Grape Fruits (Vitis vinifera L.) Inoculated with Alternaria alternata. Foods 2023, 12, 4305. [CrossRef]
- Nejman, D.; Livyatan, I.; Fuks, G.; Gavert, N.; Zwang, Y.; Geller, L.T.; Rotter-Maskowitz, A.; Weiser, R.; Mallel, G.; Gigi, E.; et al. The human tumor microbiome is composed of tumor type–specific intracellular bacteria. Science 2020, 368, 973–980. [CrossRef]
- Sun, H.; Zhao, X.; Zhou, Y.; Wang, J.; Ma, R.; Ren, X.; Wang, H.; Zou, L. Characterization of Oral Microbiome and Exploration of Potential Biomarkers in Patients with Pancreatic Cancer. BioMed Res. Int. 2020, 2020, 1–11. [CrossRef]
- Rinninella, E.; Cintoni, M.; Raoul, P.; Lopetuso, L.R.; Scaldaferri, F.; Pulcini, G.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. Food Components and Dietary Habits: Keys for a Healthy Gut Microbiota Composition. Nutrients 2019, 11, 2393. [CrossRef]
- Mitsuhashi, K.; Nosho, K.; Sukawa, Y.; Matsunaga, Y.; Ito, M.; Kurihara, H.; Kanno, S.; Igarashi, H.; Naito, T.; Adachi, Y.; et al. Association ofFusobacteriumspecies in pancreatic cancer tissues with molecular features and prognosis. Oncotarget 2015, 6, 7209–7220. [CrossRef]
- I J Malesza, M Malesza, J Walkowiak et al. “Western-Style Diet, Systemic Inflammation, and Gut Micro- biota: A Narrative Review”. Cells, vol. 10, 3164–3164, 2021.
- Half, E.; Keren, N.; Reshef, L.; Dorfman, T.; Lachter, I.; Kluger, Y.; Reshef, N.; Knobler, H.; Maor, Y.; Stein, A.; et al. Fecal microbiome signatures of pancreatic cancer patients. Sci. Rep. 2019, 9, 1–12. [CrossRef]
- Zhou, W.; Zhang, D.; Li, Z.; Jiang, H.; Li, J.; Ren, R.; Gao, X.; Li, J.; Wang, X.; Wang, W.; et al. The fecal microbiota of patients with pancreatic ductal adenocarcinoma and autoimmune pancreatitis characterized by metagenomic sequencing. J. Transl. Med. 2021, 19, 1–12. [CrossRef]
- Yu X, Li W and Li Z and Wu Q and Sun S. “Influence of Microbiota on Tumor Immunotherapy”. Int J Biol Sci, vol. 20, no. 6, 2024.
- Chang, C.-W.; Liu, C.-Y.; Lee, H.-C.; Huang, Y.-H.; Li, L.-H.; Chiau, J.-S.C.; Wang, T.-E.; Chu, C.-H.; Shih, S.-C.; Tsai, T.-H.; et al. Lactobacillus casei Variety rhamnosus Probiotic Preventively Attenuates 5-Fluorouracil/Oxaliplatin-Induced Intestinal Injury in a Syngeneic Colorectal Cancer Model. Front. Microbiol. 2018, 9, 983. [CrossRef]
- Lam, K.C.; Araya, R.E.; Huang, A.; Chen, Q.; Di Modica, M.; Rodrigues, R.R.; Lopès, A.; Johnson, S.B.; Schwarz, B.; Bohrnsen, E.; et al. Microbiota triggers STING-type I IFN-dependent monocyte reprogramming of the tumor microenvironment. Cell 2021, 184, 5338–5356.e21, Available from: https://www.sciencedirect.com/science/article/pii/S0092867421010667.
- Prasad, K.N.; Kumar, A.; Kochupillai, V.; Cole, W.C. High doses of multiple antioxidant vitamins: essential ingredients in improving the efficacy of standard cancer therapy.. J. Am. Coll. Nutr. 1999, 18, 13–25. [CrossRef]
- Yadav, M.K.; Kumari, I.; Singh, B.; Sharma, K.K.; Tiwari, S.K. Probiotics, prebiotics and synbiotics: Safe options for next-generation therapeutics. Appl. Microbiol. Biotechnol. 2022, 106, 505–521. [CrossRef]
- Pujari, R.; Banerjee, G. Impact of prebiotics on immune response: from the bench to the clinic. Immunol. Cell Biol. 2020, 99, 255–273. [CrossRef]
- Pujari, R.; Banerjee, G. Impact of prebiotics on immune response: from the bench to the clinic. Immunol. Cell Biol. 2020, 99, 255–273. [CrossRef]
- Trivieri N, Panebianco C, Villani A et al. “High Levels of Prebiotic Resistant Starch in Diet Modulate a Spe- cific Pattern of miRNAs Expression Profile Associated to a Better Overall Survival in Pancreatic Cancer.”. Biomolecules, vol. 11, no. 1, 2020.
- Shi Y, Cui H, Wang F et al. “Role of gut microbiota in postoperative complications and prognosis of gas- trointestinal surgery: A narrative review. ”. Medicine (Baltimore), vol. 101, no. 29, 2022.
- Davoodvandi, A.; Fallahi, F.; Tamtaji, O.R.; Tajiknia, V.; Banikazemi, Z.; Fathizadeh, H.; Abbasi-Kolli, M.; Aschner, M.; Ghandali, M.; Sahebkar, A.; et al. An Update on the Effects of Probiotics on Gastrointestinal Cancers. Front. Pharmacol. 2021, 12, 680400. [CrossRef]
- Rivai, M.I.; Lusikooy, R.E.; Putra, A.E.; Elliyanti, A. Effects of Lactococcus lactis on colorectal cancer in various terms: a narrative review. Ann. Med. Surg. 2024, 86, 3503–3507. [CrossRef]
- Tintelnot, J.; Xu, Y.; Lesker, T.R.; Schönlein, M.; Konczalla, L.; Giannou, A.D.; Pelczar, P.; Kylies, D.; Puelles, V.G.; Bielecka, A.A.; et al. Microbiota-derived 3-IAA influences chemotherapy efficacy in pancreatic cancer. Nature 2023, 615, 168–174. [CrossRef]
- Kato-Kataoka, A.; Nishida, K.; Takada, M.; Kawai, M.; Kikuchi-Hayakawa, H.; Suda, K.; Ishikawa, H.; Gondo, Y.; Shimizu, K.; Matsuki, T.; et al. Fermented Milk Containing Lactobacillus casei Strain Shirota Preserves the Diversity of the Gut Microbiota and Relieves Abdominal Dysfunction in Healthy Medical Students Exposed to Academic Stress. Appl. Environ. Microbiol. 2016, 82, 3649–3658. [CrossRef]
- Ijiri, M.; Fujiya, M.; Konishi, H.; Tanaka, H.; Ueno, N.; Kashima, S.; Moriichi, K.; Sasajima, J.; Ikuta, K.; Okumura, T. Ferrichrome identified from Lactobacillus casei ATCC334 induces apoptosis through its iron-binding site in gastric cancer cells. Tumor Biol. 2017, 39. [CrossRef]
- Goderska, K.; Pena, S.A.; Alarcon, T. Helicobacter pylori treatment: antibiotics or probiotics. Appl. Microbiol. Biotechnol. 2017, 102, 1–7. [CrossRef]
- Yun, C.H.; Lundgren, A.; Azem, J.; Sjöling, A.; Holmgren, J.; Svennerholm, A.-M.; Lundin, B.S. Natural Killer Cells andHelicobacter pyloriInfection: Bacterial Antigens and Interleukin-12 Act Synergistically To Induce Gamma Interferon Production. Infect. Immun. 2005, 73, 1482–1490. [CrossRef]
- Aziz N and Bonavida B. “Activation of Natural Killer Cells by Probiotics.”. For Immunopathol Dis Therap., 2016.
- Wang, Y.-H.; Yao, N.; Wei, K.-K.; Jiang, L.; Hanif, S.; Wang, Z.-X.; Pei, C.-X. The efficacy and safety of probiotics for prevention of chemoradiotherapy-induced diarrhea in people with abdominal and pelvic cancer: a systematic review and meta-analysis. Eur. J. Clin. Nutr. 2016, 70, 1246–1253. [CrossRef]
- Z Juan, J Chen, B Ding et al. “Probiotic supplement attenuates chemotherapy-related cognitive impairment in patients with breast cancer: a randomised, double-blind, and placebo-controlled trial ”. Eur J Cancer, vol. 161, 2022.
- C Panebianco, K Adamberg, S Adamberg et al. “Tuning Gut Microbiota through a Probiotic Blend in Gemcitabine-Treated Pancreatic Cancer Xenografted Mice”. Clin. Transl. Med., 2021.
- Lee, S.-H.; Cho, S.-Y.; Yoon, Y.; Park, C.; Sohn, J.; Jeong, J.-J.; Jeon, B.-N.; Jang, M.; An, C.; Lee, S.; et al. Bifidobacterium bifidum strains synergize with immune checkpoint inhibitors to reduce tumour burden in mice. Nat. Microbiol. 2021, 6, 277–288. [CrossRef]
- Iida, N.; Dzutsev, A.; Stewart, C.A.; Smith, L.; Bouladoux, N.; Weingarten, R.A.; Molina, D.A.; Salcedo, R.; Back, T.; Cramer, S.; et al. Commensal Bacteria Control Cancer Response to Therapy by Modulating the Tumor Microenvironment. Science 2019, 342, 967–970. [CrossRef]
- Martín, R.; Langella, P. Emerging Health Concepts in the Probiotics Field: Streamlining the Definitions. Front. Microbiol. 2019, 10, 1047. [CrossRef]
- Zhang, H.; Duan, Y.; Cai, F.; Cao, D.; Wang, L.; Qiao, Z.; Hong, Q.; Li, N.; Zheng, Y.; Su, M.; et al. Next-Generation Probiotics: Microflora Intervention to Human Diseases. BioMed Res. Int. 2022, 2022, 1–12. [CrossRef]
- Zhu, H.; Lu, C.; Gao, F.; Qian, Z.; Yin, Y.; Kan, S.; Chen, D. Selenium-enriched Bifidobacterium longum DD98 attenuates irinotecan-induced intestinal and hepatic toxicity in vitro and in vivo. Biomed. Pharmacother. 2021, 143, 112192. [CrossRef]
- Cani, P.D.; Depommier, C.; Derrien, M.; Everard, A.; de Vos, W.M. Akkermansia muciniphila: paradigm for next-generation beneficial microorganisms. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 625–637. [CrossRef]
- Li, X.; Hu, S.; Yin, J.; Peng, X.; King, L.; Li, L.; Xu, Z.; Zhou, L.; Peng, Z.; Ze, X.; et al. Effect of synbiotic supplementation on immune parameters and gut microbiota in healthy adults: a double-blind randomized controlled trial. Gut Microbes 2023, 15, 2247025. [CrossRef]
- Maher, S.; Elmeligy, H.A.; Aboushousha, T.; Helal, N.S.; Ossama, Y.; Rady, M.; Hassan, A.M.A.; Kamel, M. Synergistic immunomodulatory effect of synbiotics pre- and postoperative resection of pancreatic ductal adenocarcinoma: a randomized controlled study. Cancer Immunol. Immunother. 2024, 73, 1–12. [CrossRef]
- Singh, N.K.; Beckett, J.M.; Kalpurath, K.; Ishaq, M.; Ahmad, T.; Eri, R.D. Synbiotics as Supplemental Therapy for the Alleviation of Chemotherapy-Associated Symptoms in Patients with Solid Tumours. Nutrients 2023, 15, 1759. [CrossRef]
- Zółkiewicz J and Marzec A. “Postbiotics-A Step Beyond Pre- and Probiotics”. Nutrients, vol. 12, no. 8, 2020.
- Klemashevich, C.; Wu, C.; Howsmon, D.; Alaniz, R.C.; Lee, K.; Jayaraman, A. Rational identification of diet-derived postbiotics for improving intestinal microbiota function. Curr. Opin. Biotechnol. 2014, 26, 85–90. [CrossRef]
- I Bangolo, A.; Trivedi, C.; Jani, I.; Pender, S.; Khalid, H.; Alqinai, B.; Intisar, A.; Randhawa, K.; Moore, J.; De Deugd, N.; et al. Impact of gut microbiome in the development and treatment of pancreatic cancer: Newer insights. World J. Gastroenterol. 2023, 29, 3984–3998. [CrossRef]
- Jafari, B.; Nejad, R.A.K.; Vaziri, F.; Siadat, S.D. Evaluation of the effects of extracellular vesicles derived from Faecalibacterium prausnitzii on lung cancer cell line. Biologia 2019, 74, 889–898. [CrossRef]
- B K Sobocki, K Ka ´zmierczak Siedlecka, M Folwarski et al. “Pancreatic Cancer and Gut Microbiome- Related Aspects: A Comprehensive Review and Dietary Recommendations. ”. Nutrients, 2021.
- C Edderkaoui, B Chheda and Soufi. “An Inhibitor of GSK3B and HDACs Kills Pancreatic Cancer Cells and Slows Pancreatic Tumor Growth and Metastasis in Mice”. Gastroenterology, 2018.
- Donohoe, D.R.; Collins, L.B.; Wali, A.; Bigler, R.; Sun, W.; Bultman, S.J. The Warburg Effect Dictates the Mechanism of Butyrate-Mediated Histone Acetylation and Cell Proliferation. Mol. Cell 2012, 48, 612–626. [CrossRef]
- Schmidt, T.S.B.; Raes, J.; Bork, P. The Human Gut Microbiome: From Association to Modulation. Cell 2018, 172, 1198–1215. [CrossRef]
- Brevi, A.; Zarrinpar, A. Live Biotherapeutic Products as Cancer Treatments. Cancer Res. 2023, 83, 1929–1932. [CrossRef]
- Kartal E, Schmidt TSB, Molina-Montes E, et al. A faecal microbiota signature with high specificity for pancreatic cancer; Gut 2022;71:1359-1372.
- Chen, D.; Wu, J.; Jin, D.; Wang, B.; Cao, H. Fecal microbiota transplantation in cancer management: Current status and perspectives. Int. J. Cancer 2019, 145, 2021–2031. [CrossRef]
- Santana, A.B.; Souto, B.S.; Santos, N.C.d.M.; Pereira, J.A.; Tagliati, C.A.; Novaes, R.D.; Corsetti, P.P.; de Almeida, L.A. Murine response to the opportunistic bacterium Pseudomonas aeruginosa infection in gut dysbiosis caused by 5-fluorouracil chemotherapy-induced mucositis. Life Sci. 2022, 307, 120890. [CrossRef]
- H R Wardill, S A R Van Der Aa, A R Da Silva Ferreira et al. “Antibiotic-induced disruption of the micro- biome exacerbates chemotherapy-induced diarrhoea and can be mitigated with autologous faecal microbiota transplantation”. Eur J Cancer., vol. 153, 27–39, 2021.
- L. Derosa, B. Routy, A. Desilets et al. “Microbiota-Centered Interventions: The Next Breakthrough in Immuno-Oncology?”. Cancer Discov., pages 2396–2412, 2021.
- Selvanesan, B.C.; Chandra, D.; Quispe-Tintaya, W.; Jahangir, A.; Patel, A.; Meena, K.; Da Silva, R.A.A.; Friedman, M.; Gabor, L.; Khouri, O.; et al. Listeria delivers tetanus toxoid protein to pancreatic tumors and induces cancer cell death in mice. Sci. Transl. Med. 2022, 14, eabc1600–eabc1600. [CrossRef]
- Kim JE, Phan TX, Nguyen VH et al. “Salmonella typhimurium suppresses tumor growth via the pro- inflammatory cytokine interleukin-1beta”. Theranostics, vol. 5, no. 12, 1328–1342, 2015.
- Peng Z and Cheng S, Kou Y, Wang Z and Jin R, Hu H et al. “The gut microbiome is associated with clinical response to anti-pd-1/pd-L1 immunotherapy in gastrointestinal cancer.”. Cancer Immunol Res, vol. 8, no. 10, 1251–1261, 2020.
- Sethi, V.; Kurtom, S.; Tarique, M.; Lavania, S.; Malchiodi, Z.; Hellmund, L.; Zhang, L.; Sharma, U.; Giri, B.; Garg, B.; et al. Gut Microbiota Promotes Tumor Growth in Mice by Modulating Immune Response. Gastroenterology 2018, 155, 33–37.e6. [CrossRef]
- Thomas, R.M.; Gharaibeh, R.Z.; Gauthier, J.; Beveridge, M.; Pope, J.L.; Guijarro, M.V.; Yu, Q.; He, Z.; Ohland, C.; Newsome, R.; et al. Intestinal microbiota enhances pancreatic carcinogenesis in preclinical models. Carcinogenesis 2018, 39, 1068–1078. [CrossRef]
- Imai, H.; Saijo, K.; Komine, K.; Otsuki, Y.; Ohuchi, K.; Sato, Y.; Okita, A.; Takahashi, S.; Shirota, H.; Takahashi, M.; et al. Antibiotic therapy augments the efficacy of gemcitabine-containing regimens for advanced cancer: a retrospective study. Cancer Manag. Res. 2019, ume 11, 7953–7965. [CrossRef]
- Halle-Smith, J.M.; Pearce, H.; Nicol, S.; Hall, L.A.; Powell-Brett, S.F.; Beggs, A.D.; Iqbal, T.; Moss, P.; Roberts, K.J. Involvement of the Gut Microbiome in the Local and Systemic Immune Response to Pancreatic Ductal Adenocarcinoma. Cancers 2024, 16, 996. [CrossRef]
- Vrieze A, Out C, Fuentes S et al. “Impact of oral vancomycin on gut microbiota, bile acid metabolism, and insulin sensitivity.”. J Hepatol., vol. 60, no. 4, 824–831, 2014.
- Dambuza, I.M.; Brown, G.D. Fungi accelerate pancreatic cancer. Nature 2019, 574, 184–185. [CrossRef]
- Billamboz, M.; Jawhara, S. Anti-Malassezia Drug Candidates Based on Virulence Factors of Malassezia-Associated Diseases. Microorganisms 2023, 11, 2599. [CrossRef]
- R T Schooley, B Biswas, J J Gill et al. “Development and use of personalized bacteriophage-based therapeu- tic cocktails to treat a patient with a disseminated resistant Acinetobacter baumannii infection”. Antimicrob Agents Chemother, no. 10, 61–61, 2017.
- L K Harada, E C Silva, W F Campos et al. “Biotechnological Applications of Bacteriophages”. Microbiol. Res., 2018.
- Schmitz-Winnenthal FH, Hohmann N, Schmidt T et al. “A phase 1 trial extension to assess immunologic efficacy and safety of prime-boost vaccination with vxm01, an oral T cell vaccine against vegfr2, in patients with advanced pancreatic cancer”. Oncoimmunology, vol. 7, no. 4, 2018.
- Schmitz-Winnenthal, F.H.; Hohmann, N.; Niethammer, A.G.; Friedrich, T.; Lubenau, H.; Springer, M.; Breiner, K.M.; Mikus, G.; Weitz, J.; Ulrich, A.; et al. Anti-angiogenic activity of VXM01, an oral T-cell vaccine against VEGF receptor 2, in patients with advanced pancreatic cancer: A randomized, placebo-controlled, phase 1 trial. OncoImmunology 2015, 4, e1001217. [CrossRef]
- Le, D.T.; Wang-Gillam, A.; Picozzi, V.; Greten, T.F.; Crocenzi, T.; Springett, G.; Morse, M.; Zeh, H.; Cohen, D.; Fine, R.L.; et al. Safety and Survival With GVAX Pancreas Prime and Listeria Monocytogenes–Expressing Mesothelin (CRS-207) Boost Vaccines for Metastatic Pancreatic Cancer. J. Clin. Oncol. 2015, 33, 1325–1333. [CrossRef]
- Le DT, Picozzi VJ, Ko AH et al. “Results from a phase iib, randomized, multicenter study of gvax pancreas and crs-207 compared with chemotherapy in adults with previously treated metastatic pancreatic adenocar- cinoma (Eclipse study)”. Clin Cancer Res, vol. 25, no. 18, 2019.
- Fanijavadi, S.; Thomassen, M.; Jensen, L. H. Targeting Triple NK Cell Suppression Mechanisms: A Comprehensive Review of Biomarkers in Pancreatic Cancer Therapy. Preprints 2024, 2024121161. [CrossRef]
Figure 1.
Microbiota-NK Cell Interaction and PDAC Clinical Outcome. This figure illustrates the complex interactions between different microbiota (oral, gut, and tumor-associated) and their crosstalk with NK cells in pancreatic ductal adenocarcinoma (PDAC). The microbiota communities not only interact with NK cells but also influence each other: oral microbiota can alter the gut and, ultimately, the tumor-associated microbiota, while tumor-associated microbiota can also modify the gut and oral microbiota. Dysbiosis, a pathological disruption of the microbiota community, can suppress NK cell function, including reducing NK cell frequency, cytotoxicity, and tumor infiltration. In contrast, a balanced microbiota can enhance NK cell activity, promoting tumor surveillance and immune response. Dysbiosis, however, contributes to tumor initiation, progression, resistance to treatment by impairing NK cell-mediated immune responses, and side effects of cancer treatments.
Figure 1.
Microbiota-NK Cell Interaction and PDAC Clinical Outcome. This figure illustrates the complex interactions between different microbiota (oral, gut, and tumor-associated) and their crosstalk with NK cells in pancreatic ductal adenocarcinoma (PDAC). The microbiota communities not only interact with NK cells but also influence each other: oral microbiota can alter the gut and, ultimately, the tumor-associated microbiota, while tumor-associated microbiota can also modify the gut and oral microbiota. Dysbiosis, a pathological disruption of the microbiota community, can suppress NK cell function, including reducing NK cell frequency, cytotoxicity, and tumor infiltration. In contrast, a balanced microbiota can enhance NK cell activity, promoting tumor surveillance and immune response. Dysbiosis, however, contributes to tumor initiation, progression, resistance to treatment by impairing NK cell-mediated immune responses, and side effects of cancer treatments.
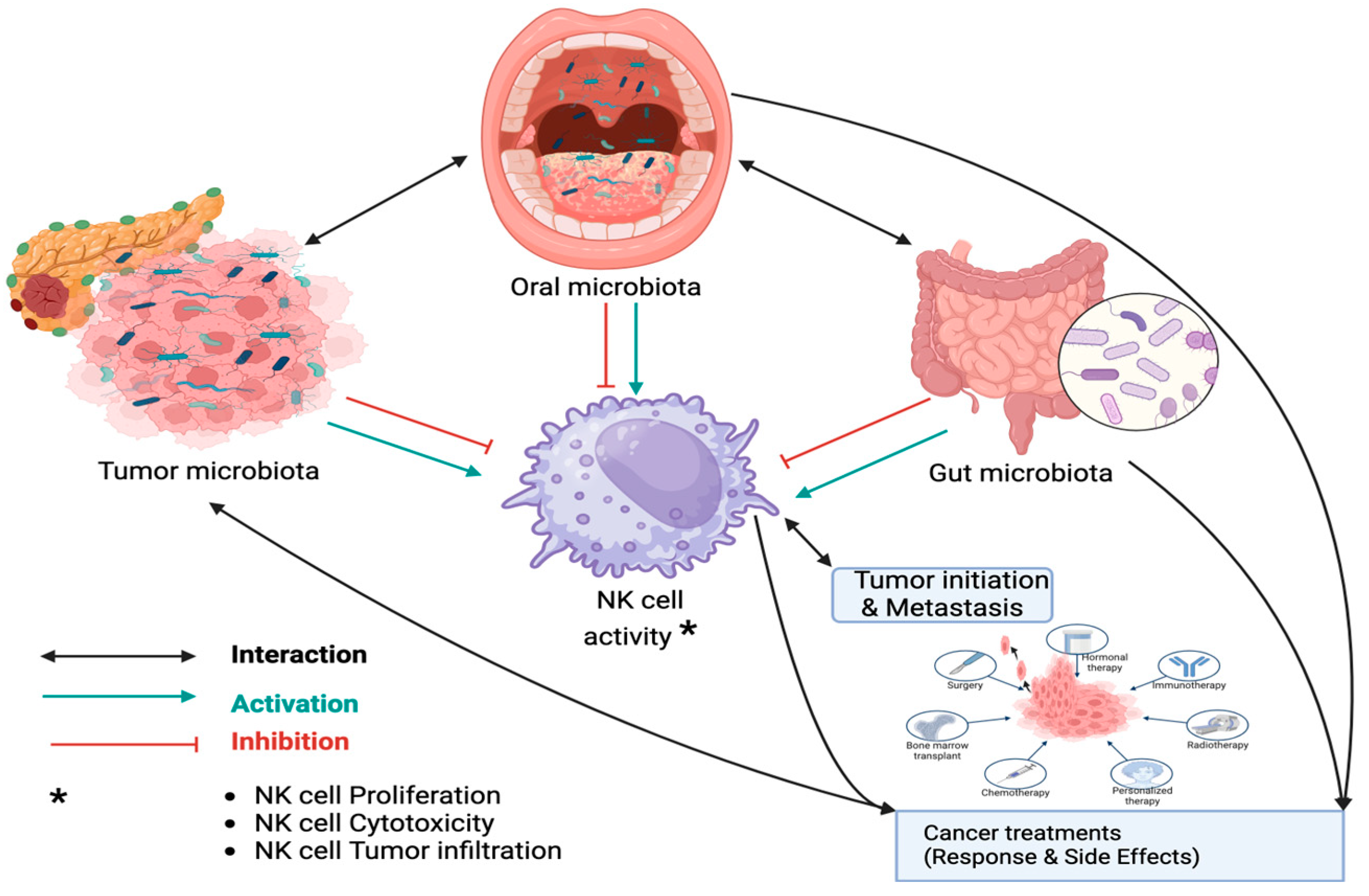
Figure 2.
Microbiota-NK Cell Interactions and Their Impact on Clinical Outcomes. This figure illustrates the various mechanisms through which the microbiota can influence NK cell activity and treatment outcomes in pancreatic ductal adenocarcinoma (PDAC). The diagram highlights the complex interplay between microbiota, NK cells, and chemotherapy. Specifically, it focuses on the AMPK-Bmi1-GATA2-MICA/B pathway, which plays a critical role in tumor immune evasion. Under conditions of energy stress, such as high glucose, AMPK is activated, leading to the upregulation of Bmi1. Bmi1, in turn, activates GATA2, which reduces the expression of MICA/B ligands. These ligands normally stimulate NK cells to recognize and attack tumor cells. By downregulating MICA/B, this pathway helps cancer cells evade NK cell-mediated immune surveillance, thereby promoting tumor growth and resistance to treatment like 5-fluorouracil (5-FU). Gut microbiota metabolites can be broadly categorized into three types based on their origin: (1) metabolites produced directly by the microbiota from dietary components, such as short-chain fatty acids (SCFAs like butyrate) and indole derivatives (like IAA), which play key roles in gut health and immune modulation (via inhibition of histone deacetylases (HDACs); (2) metabolites generated by the host and modified by the microbiota, like secondary bile acids, which influence metabolism and immune functions; and (3) metabolites produced de novo by the microbiota, such as polysaccharide A (PSA) which helps maintain immune balance. Additionally, some products like uridine, produced by cytidine deaminase (CDA) enzyme can also be assumed as de novo metabolites. These metabolites, along with other postbiotics like cell-free components (CSF), exert various effects on the host, including modulating inflammation and gut integrity.
Figure 2.
Microbiota-NK Cell Interactions and Their Impact on Clinical Outcomes. This figure illustrates the various mechanisms through which the microbiota can influence NK cell activity and treatment outcomes in pancreatic ductal adenocarcinoma (PDAC). The diagram highlights the complex interplay between microbiota, NK cells, and chemotherapy. Specifically, it focuses on the AMPK-Bmi1-GATA2-MICA/B pathway, which plays a critical role in tumor immune evasion. Under conditions of energy stress, such as high glucose, AMPK is activated, leading to the upregulation of Bmi1. Bmi1, in turn, activates GATA2, which reduces the expression of MICA/B ligands. These ligands normally stimulate NK cells to recognize and attack tumor cells. By downregulating MICA/B, this pathway helps cancer cells evade NK cell-mediated immune surveillance, thereby promoting tumor growth and resistance to treatment like 5-fluorouracil (5-FU). Gut microbiota metabolites can be broadly categorized into three types based on their origin: (1) metabolites produced directly by the microbiota from dietary components, such as short-chain fatty acids (SCFAs like butyrate) and indole derivatives (like IAA), which play key roles in gut health and immune modulation (via inhibition of histone deacetylases (HDACs); (2) metabolites generated by the host and modified by the microbiota, like secondary bile acids, which influence metabolism and immune functions; and (3) metabolites produced de novo by the microbiota, such as polysaccharide A (PSA) which helps maintain immune balance. Additionally, some products like uridine, produced by cytidine deaminase (CDA) enzyme can also be assumed as de novo metabolites. These metabolites, along with other postbiotics like cell-free components (CSF), exert various effects on the host, including modulating inflammation and gut integrity.
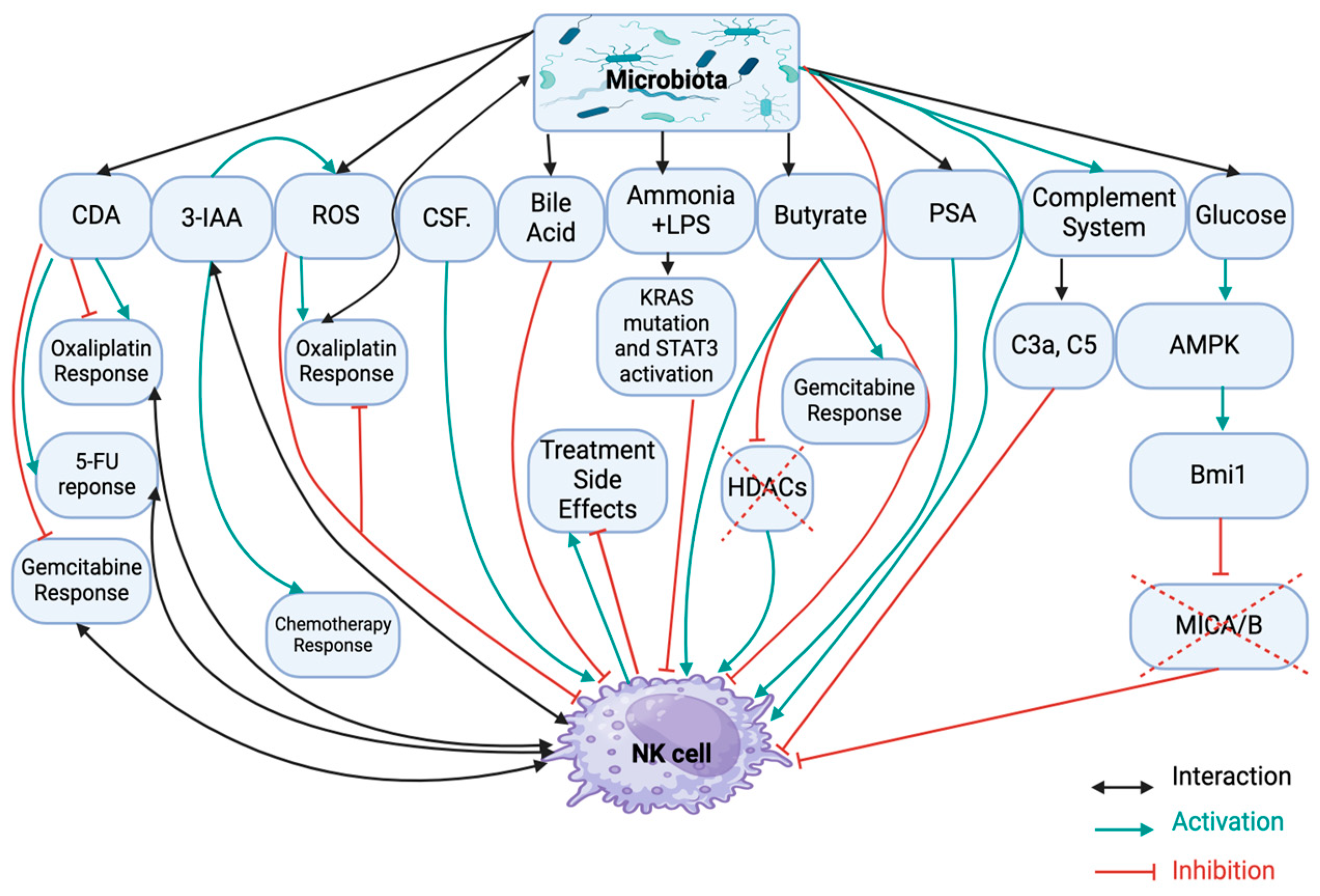
Figure 4.
Integration of Microbiome Profiling and NK Cell Analysis as Unified Biomarkers for Managing PDAC. This figure illustrates a suggested comprehensive diagnostic approach for identifying unified biomarkers to predict treatment resistance and side effects in pancreatic ductal adenocarcinoma (PDAC) patients. The approach integrates microbiota analysis from oral washing samples, stool, or tumor tissues using omics technologies, such as 16S rRNA sequencing, with NK cell profiling. 16S rRNA sequencing is a genomic technique that profiles microbial communities by analyzing the 16S ribosomal RNA gene, providing insights into microbial diversity and how the microbiome may influence PDAC treatment outcomes. The analysis can be performed in PDAC patient cohorts, categorized by long vs. short overall survival, responders vs. non-responders, and patients with vs. without side effects. By combining microbiome data with NK cell profiling, the goal is to identify combined biomarker signatures that optimize clinical outcomes. Additionally, the figure highlights the importance of metagenomics and metaproteomics, which offer complementary insights into the interactions between patient cells and the microbiome, essential for developing robust biomarkers for personalized PDAC therapies. NK cell profiling involves isolating NK cells from patient samples (e.g., peripheral blood, tumor tissues) and analyzing their phenotype, function, and gene expression. This includes flow cytometry to identify NK cell subpopulations (e.g., CD56^dim, CD56^bright), functional assays (e.g., cytotoxicity and cytokine production), and gene expression profiling (via RNA sequencing or qRT-PCR). These analyses reveal NK cell activation, exhaustion, or senescence, helping to identify biomarkers associated with treatment resistance and side effects, thereby contributing to more personalized PDAC therapies.
Figure 4.
Integration of Microbiome Profiling and NK Cell Analysis as Unified Biomarkers for Managing PDAC. This figure illustrates a suggested comprehensive diagnostic approach for identifying unified biomarkers to predict treatment resistance and side effects in pancreatic ductal adenocarcinoma (PDAC) patients. The approach integrates microbiota analysis from oral washing samples, stool, or tumor tissues using omics technologies, such as 16S rRNA sequencing, with NK cell profiling. 16S rRNA sequencing is a genomic technique that profiles microbial communities by analyzing the 16S ribosomal RNA gene, providing insights into microbial diversity and how the microbiome may influence PDAC treatment outcomes. The analysis can be performed in PDAC patient cohorts, categorized by long vs. short overall survival, responders vs. non-responders, and patients with vs. without side effects. By combining microbiome data with NK cell profiling, the goal is to identify combined biomarker signatures that optimize clinical outcomes. Additionally, the figure highlights the importance of metagenomics and metaproteomics, which offer complementary insights into the interactions between patient cells and the microbiome, essential for developing robust biomarkers for personalized PDAC therapies. NK cell profiling involves isolating NK cells from patient samples (e.g., peripheral blood, tumor tissues) and analyzing their phenotype, function, and gene expression. This includes flow cytometry to identify NK cell subpopulations (e.g., CD56^dim, CD56^bright), functional assays (e.g., cytotoxicity and cytokine production), and gene expression profiling (via RNA sequencing or qRT-PCR). These analyses reveal NK cell activation, exhaustion, or senescence, helping to identify biomarkers associated with treatment resistance and side effects, thereby contributing to more personalized PDAC therapies.

Table 1.
Association between microbiomes and NK cell function: clinical application.
| Taxonomy | Role, NK cell affection | Translational Clinical Application/Suggestion |
|---|---|---|
| Actinobacteria (phylum) | Protective in the oral microbiome. Carcinogenic in the gut microbiome)52, 67, 85. | Phylum regulation in the oral and gut microbiomes through direct or indirect dysbiosis-targeting strategies. |
| Acidovorax Ebreus; A. ebreus (species, genus: Acidovorax, phylum: proteobac) | Carcinogenic by reducing immune cells 4,10,18,19,20. | Species depletion through direct or indirect dysbiosis-targeting strategies. |
| Acinobacter baumannii (species; genus: Acinobacter, phylum: Proteobacteria) |
Carcinogenic in the intratumor microbiome 52, 67. | Intra-tumor species depletion through direct or indirect dysbiosis-targeting strategies. |
| Actinomyces (genus; phylum: Actinobacteria) | Carcinogenic in the oral and gut microbiome 20, 85, 104. Negative prognostic marker in the intratumor microbiome 16. |
Genus depletion in the intra-tumoral, intra-gut, and oral microbiomes through direct or indirect dysbiosis-targeting strategies. |
| Aggregatibacter (genus; phylum: Proteobacteria) | Negative prognostic marker in the intratumor microbiome 16. | Intra-tumor genus depletion through direct or indirect dysbiosis-targeting strategies. |
| Aggregatibacter actinomycetemcomitans (species; genus: Aggregatibacter phylum: proteobacteria) | Carcinogenic in the oral microbiome by promoting tumor favoring inflammation and NK cell exhaustion 4, 10, 16,1 8, 19, 20, 29, 52 | Species depletion in the oral microbiomes through direct or indirect dysbiosis-targeting strategies. |
| Akkermansia muciniphila (species; genus: Akkermansia, phylum: verrucomicrobia) | Carcinogenic in the intratumor microbiome. Protective in the gut microbiome by decreasing NK cell exhaustion and positive predictive marker for immunotherapy 4,10,16,18,19,20,148. | Increasing species abundance, for example, by prescribing NGPs as adjuvants for immunotherapy to enhance the efficacy of PD-1 inhibitors 133,134. |
| Alistipes shahii (species; genus: Alistipes, phylum: Bacteriodetes) | Carcinogenic in the gut microbiome 6. | Species depletion in the gut microbiome through direct or indirect dysbiosis-targeting strategies. |
| Alloprevotella (genus; phylum: Firmicutes) | Protective in the oral microbiome 104. | Increasing genus abundance, for example, by prescribing NGPs, probiotics, lifestyle changes, vitamin therapy. |
| Alloscardovia omnicolens (species; genus: Alloscardovia, phylum: Actinobacteria) | Carcinogenic in the gut microbiome 148. | Species depletion in the gut microbiome through direct or indirect dysbiosis-targeting strategies. |
| Altidipes indistinctus (species; genus: Altidipes; phylum: Bacteroidetes) | Positive predictive marker for immunotherapy and response to IL-10+CpG oligonucleotide. Protective by decreasing NK cell exhaustion 4,10,16,17,18,19,20,133,134. | Increasing species abundance, for example, by prescribing NGPs, probiotics, lifestyle changes, vitamin therapy or as adjuvants for immunotherapy or IL-10+CpG oligonucleotide to enhance the efficacy. |
| Alternaria alternata (species; genus: Alternaria, phylum: Ascomycota) | Carcinogenic by enhancing tumor favoring immunity, inducing NK cell exhaustion 4,10,19,18,20. | Species depletion through direct or indirect dysbiosis-targeting strategies. |
| Anaerostipes (genus; phylum: Firmicutes) | Protective in the gut microbiome) 8, 109. | Increasing genus abundance, for example, by prescribing NGPs, probiotics, FM, lifestyle changes, vitamin therapy. |
| Atopobium parvulum (species; genus: Atopobium; phylum: Actinobacteria) | Carcinogenic in the oral microbiome) 38. | Species depletion in the oral microbiomes through direct or indirect dysbiosis-targeting strategies. |
| Bacidiomycota (phylum of fungi) | Carcinogenic by activating mannose binding lectin, suppressing NK cells cytotoxicity 83. | Phylum depletion through direct or indirect dysbiosis-targeting strategies. |
| Bacillus clausii (species; genus: Bacillus, phylum: Firmicutes) | Protective in the intratumor microbiome 15. | Increasing species abundance, for example, by prescribing tumor-colonizing bacteria, oral probiotics, FM, lifestyle changes, vitamin therapy. |
| Bacteroidales (order; phylum: Bacteroidetes) | Carcinogenic in the gut microbiome) 109. | Order depletion in the gut microbiome through direct or indirect dysbiosis-targeting strategies. |
| Bacteroides (genus; phylum: bacteroidetes) |
Carcinogenic in the oral, gut and intratumor microbiome 105, 148. PIPN inducer, activating Astrocytes and TLR4/p38MAPK Pathway, with effects on NK cell regulation 6, 97, 98. |
Genus depletion in the intra-tumoral, intra-gut, and oral microbiomes through direct or indirect dysbiosis-targeting strategies. |
| Bacteroides coprocola (species; genus: Bacteroides, phylum: Bacteroidetes) | Protective in the gut microbiome 148. | Increasing species abundance in the gut for example, by prescribing oral probiotics, FMT, lifestyle changes, vitamin therapy. |
| Bacteroidetes (phylum) | Carcinogenic in the gut microbiome 52. | Phylum depletion in the gut microbiome through direct or indirect dysbiosis-targeting strategies. |
| Bifidobacterium (genus; phylum: Actinobacteria) | Protective in the gut microbiome 8. It can increase NKA2,64 and acts as a positive predictive biomarker for immunotherapy to potentiate the efficacy of immune checkpoint inhibitors63,64, 134. | Increasing genus abundance in the gut microbiome, for example, by prescribing FMT, NGPs, probiotics, lifestyle changes, vitamin therapy or as adjuvants for immunotherapy to potentiate its effect. |
| Bifidobacterium bifidum (species; genus: Bifidobacterium, phylum: Actinobacteria) | Protective in the gut microbiome 148. | Increasing genus abundance in the gut microbiome, for example, by prescribing FMT, NGPs, probiotics, postbiotics, lifestyle changes, vitamin therapy. |
| Bifidobacterium pseudocatenulatum(species; genus: Bifidobacterium, phylum: Actinobacteria) | Protective in the gut microbiome 6. | Increasing species abundance in the gut microbiome, for example, by prescribing FMT, NGPs, probiotics, postbiotics, lifestyle changes, vitamin therapy. |
| Blautia (genus; phylum: Firmicutes) | Protective in the gut microbiome 8. | Increasing species abundance in the gut microbiome, for example, by prescribing FMT, NGPs, probiotics, postbiotics, lifestyle changes, vitamin therapy. |
| Blautia obeum (species; genus; phylum: Firmicutes) | Protective in the gut microbiome 6. | Increasing species abundance in the gut microbiome, for example, by prescribing FMT, NGPs, probiotics, postbiotics, lifestyle changes, vitamin therapy. |
| Bradyrhizobium (genus; phylum: Proteobacteria) | Positive prognostic marker in the intratumor microbiome 16. | Increasing genus abundance in the intratumor microbiome, for example, by prescribing tumor-colonizing bacteria, FMT, NGPs, probiotics, postbiotics, lifestyle changes, vitamin therapy. |
| Capnocytophaga (genus; phylum: Bacteroidetes) | Protective in the oral microbiome 20. | Increasing genus abundance in the gut microbiome, for example, by prescribing NGPs, probiotics, postbiotics, lifestyle changes, vitamin therapy. |
| Cardiobacterium (genus; phylum: Proteobacterium) | Carcinogenic in the gut microbiome 52. | Genus depletion in the gut microbiome through direct or indirect dysbiosis-targeting strategies. |
| Citrobacter (genus; phylum: Protecobacteria) | Carcinogenic in the intratumor microbiome 66. | Genus depletion in the intratumor microbiome through direct or indirect dysbiosis-targeting strategies. |
| Citrobacter freundii (species; genus: Citrobacter, phylum: proteobacteria) |
Carcinogenic in the intratumor microbiome, contributing to immunosuppression, upregulation of oncogenic pathways, and downregulation of tumor suppressive pathways 3,19,55,104. | Species depletion in the intra-tumoral, through direct or indirect dysbiosis-targeting strategies. |
| Cladocopium symbiosum (species; genus: Cladocopium, phylum: Apicomplexa) | Carcinogenic in the gut microbiome 6. | Species depletion in the gut microbiome through direct or indirect dysbiosis-targeting strategies. |
| Clostridiaceae (family; phylum: Firmicutes) | Protective in the gut microbiome 15. | Increasing family abundance in the gut microbiome, for example, by prescribing FMT, NGPs, probiotics, postbiotics, lifestyle changes, vitamin therapy and so on. |
| Clostridiales (order; phylum: Firmicutes) | Protective in the gut microbiome 109. | Increasing order abundance in the gut microbiome, for example, by prescribing FMT, NGPs, probiotics, postbiotics, lifestyle changes, vitamin therapy. |
| Clostridium (genus; phylum: Firmicutes) |
Protective in the gut microbiome, 109 negative prognostics in gut and oral sample15,17 but protective intratumorally 147. | Reducing abundance in gut and oral samples using probiotics, while increasing intratumoral abundance with tumor-colonizing bacteria |
| Clostridium IV (group; genus: Clostridium, phylum: Firmicutes) | Protective in the gut microbiome 8. | Increasing species abundance in the gut microbiome, for example, by prescribing FMT, NGPs, probiotics, postbiotics, lifestyle changes, vitamin therapy. |
| Clostridium bolteae (species; genus: Clostridium, phylum: Firmicutes) | Carcinogenic in the gut microbiome 6. | Species depletion in the gut microbiome through direct or indirect dysbiosis-targeting strategies. |
| Clostridium_sensu stricto 1 (species; genus: Clostridiumphylum: Firmicutes) | Negative prognostic marker in the intratumor microbiome 16.; PIPN preventive in the gut microbiome, interfering with astrocytes and the TLR4/p38MAPK pathway 97,98. | Increasing species abundance in the gut through FMT to reduce PIPN, while promoting intratumoral abundance with tumor-colonizing bacteria |
| Collinsella aerofaciens (species; genus: Collnisella, phylum: Actinobacteria) | Protective in the gut microbiome 6. | Increasing species abundance in the gut microbiome through direct or indirect dysbiosis-targeting strategies. |
| Corynebacterium (genus; phylum: Actinobacteria) | PIPN preventive, by interfering with astrocytes and the TLR4/p38MAPK pathway 97,98. | Increasing genus abundance in the gut through FMT to reduce PIPN, while promoting intratumoral abundance with tumor-colonizing bacteria. |
| Coprococcus (genus; phylum: Firmicutes) | Protective in the gut microbiome 6,8. | Increasing genus abundance in the gut microbiome through direct or indirect dysbiosis-targeting strategies. |
| Desulfovibrio (genus; phylum: Proteobacteria) | Positive prognostic marker in the intratumor microbiome 16. | Increasing intratumoral abundance through direct or indirect dysbiosis-targeting strategies, such as using tumor-colonizing bacteria. |
| Dialister (genus; phylum: Firmicutes) | Carcinogenic in the oral microbiome 85. | Reducing genus abundance in the oral microbiome using direct and indirect dysbiosis-targeting strategies. |
| Escherichia coli (species; genus: Escherichia, Phylum: Proteobacteria) |
Carcinogenic in the oral microbiome 20, decreasing NK cell cytotoxicity 93, and inducing diarrhea by reducing IL-22 production in NK cells, which normally protect the integrity of the intestinal epithelial barrier damaged by enterotoxigenic Escherichia coli. Negative predictive marker for gemcitabine response 46, 67. |
Reducing species abundance in the oral microbiome using direct and indirect dysbiosis-targeting strategies. For example, Lactobacillus probiotics can be used to increase gemcitabine efficacy and prevent diarrhea |
| Elizabethkingia (genus; phylum: Bacteroidetes) | Carcinogenic 55. | Genus depletion through direct or indirect dysbiosis-targeting strategies. |
| Enhydrobacter (genus; phylum: Proteobacterai) | Positive prognostic marker in the intratumor microbiome 16. | Increasing intratumoral abundance through direct or indirect dysbiosis-targeting strategies, such as using tumor-colonizing bacteria. |
| Enterobacter (genus; phylum: Proteocteria) | Carcinogenic in the gut microbiome 8; protective intratumorally, with TIGIT upregulation31-32. | Genus depletion through direct and indirect dysbiosis-targeting strategies. Increasing intratumoral abundance using direct or indirect dysbiosis-targeting approaches, such as tumor-colonizing bacteria. |
| Enterobacter asburiae (species; genus: Enterobacter, phylum: Poteobacteria) |
Carcinogenic in the gut and intratumor microbiome28, 104. | Species depletion in the gut and intra-tumor microbiomes through direct or indirect dysbiosis-targeting strategies. |
| Enterobacteriaceae (family; phylum: Proteobacteria) |
Carcinogenic in the oral and intratumor microbiome 20, 57, 67. |
Species depletion in the gut and intra-tumor microbiomes through direct or indirect dysbiosis-targeting strategies. |
| Enterococcus hirae (species; genus: Enterococcus, phylum: Firmicutes) | Positive predictive for immunotherapy response 63. | Increasing species abundance through direct or indirect dysbiosis-targeting strategies or prescribing the species as a probiotic product to be used as adjuvant therapy for immunotherapy. |
| Erysipelotrichaeceae (family; phylum: Firmicutes) | Protective in the gut microbiome 6. | Increasing family abundance in the gut microbiome through direct or indirect dysbiosis-targeting strategies, such as using FMT. |
| Eubacterium hallii (speies; genus: Eubacterium, phylum: Firmicutes) | Protective in the gut microbiome 6. | Increasing species abundance in the gut microbiome through direct or indirect dysbiosis-targeting strategies, such as using FMT. |
| Eubacterium ventriosum (species; genus: Eubacterium, phylum: Firmicutes) | Protective in the gut microbiome85. | Increasing species abundance in the gut microbiome through direct or indirect dysbiosis-targeting strategies, such as using FMT. |
| Faecalibacterium (genus; phylum: Firmicutes) | Protective in the gut microbiome 109. | Increasing species abundance in the gut microbiome through direct or indirect dysbiosis-targeting strategies, such as using FMT. |
| Faecalibacterium prausnitzii (species; genus: Faecalibacterium; phylum: Firmicutes) | Protective in the gut microbiome 85, 148 | Increasing species abundance in the gut microbiome through direct or indirect dysbiosis-targeting strategies, such as using FMT. |
| Firmicutes (phylum) | Protective in the gut microbiota 51, reducing bile acid production and bile acid induced carcinogenesis, while increasing NK cell activity through bile acid reduction and butyrate production 73,85,1034 110, 145 | Phylum regulation in the gut and intra-tumor microbiomes through direct or indirect dysbiosis-targeting strategies |
| Flavobacterium (genus; phylum: Bacteroidetes) | Positive prognostic marker in the intratumor microbiome 16. | Increasing genus abundance in the intra-tumor microbiome through direct or indirect dysbiosis-targeting strategies, such as tumor-colonizing bacteria. |
| Flavonifractor (genus; phylum: Firmicutes) | Protective in the gut microbiome 8. | Increasing genus abundance in the gut microbiome through direct or indirect dysbiosis-targeting strategies, such as FMT. |
| Fusobacterium (genus; Fusobacteria) | Carcinogenic in the oral microbiome 105. | Species depletion in the oral microbiome through direct or indirect dysbiosis-targeting strategies. |
| Fusobacterium nucleatum (species; genus: Fusobacterium, phylum: Fusobacteria) | Carcinogenic in the gut and intratumor microbiome via cytokine secretion and decreased NK cell tumor infiltration, serving as a negative prognostic marker intratumorally 19, 27, 28, 29, 74,104,107,148. | To decrease the species using probiotics to optimize clinical outcomes, particularly to potentiate the effects of 5-FU and oxaliplatin. |
| Gammaproteobacteria (class; phylum: Proteobacteria) | Carcinogenic in the intratumor microbiome67, a negative predicative marker for gemcitabine response 28,66,67 due to bacterial CDA, and for immunotherapy, while serving as a positive predicative marker for chemotherapy (e.g., 5-FU and oxaliplatin); immune regulation via CDA102, 104, 105. | To be prescribed as tumor-colonizing bacteria alongside oxaliplatin and 5-FU-based chemotherapy to potentiate their effects. Probiotics can be used to reduce species abundance when gemcitabine is prescribed. |
| Granulicatella adiacens (species; genus: Granulicatella, Firmicutes) | Carcinogenic in the oral microbiome 38. | Species depletion in the oral microbiome through direct or indirect dysbiosis-targeting strategies. |
| Haemophilus abundance (species; genus: Haemophilus, phylum: Proteobacterai) | Carcinogenic in the oral microbiome 57. | Species depletion in the oral microbiome through direct or indirect dysbiosis-targeting strategies. |
| Helicobacter hepaticus (species; genus: Helicobacter, phylum: Proteobacteria) | Carcinogen (seroprevalence), protective intratumorally, increasing NK cell tumor infiltration 28,29. | Patients who are seropositive for H. hepaticus have a higher risk of progression and may benefit from receiving the species as tumor-colonizing bacteria to induce NK cell infiltration. |
| Helicobacter pylori (species; genus: Helicobacter, phylum: Proteobacteria) | Carcinogenic, with NK cell suppression 124, 125. | Eradicating H. pylori using antibiotics or probiotics. Alternatively, using Interleukin 12 in patients with dysbiosis positive for H. pylori to induce gamma interferon activation and enhance NK cell activity. |
| Herbaspirillum (genus; phylum: Proteobacteria) | Carcinogenic in the intratumor microbiome 49. | Genus depletion in the intra-tumor microbiome through direct or indirect dysbiosis-targeting strategies. |
| Klebsiella (genus; phylum: Proteobacteria) | Carcinogenic in the gut and intratumor microbiome 8, 52, 67. | Genus depletion in the gut and intra-tumor microbiome through direct or indirect dysbiosis-targeting strategies. |
| Klebsiella pneumoniae (species; genus: Proteobacteria, phylum: proteobacteria) | Carcinogenic in the gut and intratumor microbiome 19, 104, leading to NK cell suppression 6,125. | Species depletion in the gut and intra-tumor microbiome through direct or indirect dysbiosis-targeting strategies. |
| Lachnospiraceae (family; phylum: Firmicutes) | Protective in the gut microbiome in some studies 6, 85, but carcinogenic in the oral and gut microbiome, reported by another study 57, 109. | Further investigation is needed to specify the strain-level effects and determine the clinical relevance, as well as to regulate dysbiosis accordingly. |
| Lactobacillus (genus; phylum: Firmicutes) | Carcinogenic in the oral and gut microbiome 6,104. | Further investigation is needed to specify the strain-level effects and determine the clinical relevance, as well as to regulate dysbiosis accordingly. |
| Lactobacillus plantarum (species; genus: Lactobacillus, phylum: Firmicutes) | Protective in the gut microbiome, while carcinogenic in the intratumor microbiome 148. | Increasing genus in the gut microbiome and reducing intratumoral abundance through direct and indirect dysbiosis-targeting strategies. |
| Leptotrichia (genus; phylum: Fusobacteria) | Carcinogenic in the oral microbiome 104; however, protective in both the gut and oral microbiome, according to other studies 20, 105. | Further investigation is needed to specify the strain-level effects and determine the clinical relevance, as well as to regulate dysbiosis accordingly. |
| Listeria whelshimer I Serover 6B SLCC 5334 (strain; genus: Listeria, phylum: Firmicutes) | Protective, inducing IL-12 production and NK cell activity via LPS 147, 153, 167. | It can be used as tumor-colonizing bacteria or as a vaccination to target tumor and enhance the response to gemcitabine. |
| Malassezia globosa (species of yeast; genus: Malassezia, phylum: Ascomycota) |
Carcinogen, enhancing tumor favoring immunity by suppress NK cells through the release of C3a and C5a, thereby reducing NKG2D161. | Species depletion using antifungal drugs or other direct and indirect dysbiosis-targeting modalities |
| Megamonas (genus; phylum: Firmicutes) | Protective in the gut microbiome 52; positive prognostic marker in the intratumor microbiome 16. | Increasing genus abundance in the gut and intra-tumor microbiomes through direct or indirect dysbiosis-targeting strategies, such as tumor-colonizing bacteria. |
| Megasphaera (genus; phylum: Firmicutes) | Protective and a positive prognostic marker in the intratumor microbiome15,16, 105, while carcinogenic in the oral microbiome 105. | Increasing genus abundance in the intratumoral microbiome through strategies like tumor-colonizing bacteria, and depleting genus abundance in the oral microbiome using appropriate dysbiosis-targeting approaches. |
| Mycobacterium (genus; phylum: Actinobacteria) | Protective through NKp44 interaction 45,47. | Increasing genus abundance using direct and indirect dysbiosis-modulating approach. |
| Mycoplasma hyorhinis (species; genus: Mycoplasma, phylum: Mycoplasmatota) | Negative predictive marker for gemcitabine due to decreased cytotoxicity caused by bacterial CDA 28,66. | Decreasing species abundance using direct and indirect dysbiosis-modulating modalities, such as antibiotics and vaccination, to prevent gemcitabine resistance. |
| Neisseria (genus; phylum: Proteobacteria) | Protective in the oral microbiome 105 and a negative prognostic marker in the intratumor microbiome 16. | Decreasing genus abundance in the intratumoral microbiome and increasing genus abundance in the oral microbiome using direct and indirect dysbiosis-modulating modalities. |
| Neisseria elongata (species; genus: Neisseria, phylum: Proteobacteria) | Protective in the oral microbiome 20,38, 104,105. | Increasing species abundance in the oral microbiome through direct or indirect dysbiosis-targeting strategies. |
| Odoribacter (genus; phylum: Bacteroidetes) | Carcinogenic in the gut microbiome 109. | Genus depletion in the gut microbiome using direct or indirect dysbiosis-targeting approach. |
| Parabacteroides (genus; phylum: Bacteroidetes) | Carcinogenic in the gut microbiome 6, 52. | Genus depletion in the gut microbiome by direct or indirect dysbiosis-targeting. |
| Propionibacterium acnes (species; genus: Propionibacterium, phylum: Actinobacteria) | Carcinogenic, through cytokine modulation and Hedgehog signaling activation 21-23. | Species depletion using, for example, antibiotics, bacteriophages, or other modalities to target dysbiosis. |
| Porphyromonas (genus; phylum: Bacteroidetes) | Protective in the oral microbiome20 and a negative prognostic marker in the intratumor microbiome 16. | Increasing genus in the oral microbiome and reducing intratumoral abundance through direct and indirect dysbiosis-targeting strategies. |
| Prophyromonas gingivalis (species; genus: Prophyromonas phylum: bacteroidetes) |
Carcinogen in the oral microbiome, inducing tumor-favoring inflammation and NET induced NK cell suppression; direct effect on NK cells has not been investigated 9,16, 17, ,20 40. | Species depletion using, for example, antibiotics or other modalities to target dysbiosis. |
| Proteobacteria (phylum) | Carcinogenic in the gut and intratumor microbiomes, promoting tumor metastasis, bile acid induced carcinogenesis, and immunosuppression9,16, 17, 18, 19, 30, 37,40, 52, 104. | This phylum needs to be analyzed at the strain level, as generalizing clinical outcomes at the phylum level is not applicable due to controversial results. |
| Prevotella (genus; phylum: Bacteroidetes) |
Carcinogenic in the oral microbiome104, but protective in both oral and gut microbiomes 20, 52. | Further investigation is needed to specify the species-level effects and determine the clinical relevance, as well as to regulate dysbiosis accordingly. |
| Prevotella pallens (species; genus: Prevotella, phylum: Bacteroidetes) | Carcinogenic in the oral microbiome 85. | Species depletion in the oral microbiome using modalities to target dysbiosis. |
| Prevotella sp. C561(strain; species P.sp, genus: Prevotella, phylum: Bacteroidetes) | Carcinogenic in the oral microbiome 85. | Strain depletion in the oral microbiome using modalities to target dysbiosis. |
| Pseudomonas (genus; phylum: Proteobacteria) |
Carcinogen in the intratumor microbiome 45, 47, 49, 150. | Genus depletion in the intra-tumor microbiome through direct or indirect dysbiosis-targeting strategies. |
| Pseudomonadaceae (family; phylum: Proteobacteria) | Carcinogenic in the intratumor microbiome 66. | Family depletion in the intra-tumor microbiome through direct or indirect dysbiosis-targeting strategies; however, further investigation at the species level is recommended. |
| Pseudomonas aeroginosa (species; genus: Pseudomonas, phylum: Proteobacteria) | Carcinogenic 45, 47, 150. | Species depletion in the intra-tumor microbiome through direct or indirect dysbiosis-targeting strategies. |
| Pseudoxanthomonas (genus; phylum: Proteobacteria) | Protective as the intratumor microbiome, triggering antitumor immunity and serving as a positive prognostic marker12,15. | Increasing genus abundance in the intratumoral microbiome through strategies like tumor-colonizing bacteria or other appropriate dysbiosis-targeting approaches. |
| Romboutsia timonensis (species; genus: Romboutsia, Phylum: Firmicutes) | Protective in the gut microbiome 148. | Increasing species abundance in the gut microbiome through strategies like FMT or other appropriate dysbiosis-targeting approaches. |
| Rothia (genus; phylum: Actinobacteria) | Carcinogenic in the oral microbiome 20. | Genus depletion in the oral microbiome by targeting dysbiosis appropriately. |
| Ruminococcaceae (family; phylum; Firmicutes) | Protective in the gut microbiome 109. | Increasing family abundance in the gut microbiome through strategies like FMT or other appropriate dysbiosis-targeting approaches. However, further investigation at genus or species level is recommended. |
| Ruminococcus (genus; phylum: Firmicutes) | Positive predictive marker for immunotherapy and response to IL-10+CpG oligonucleotide 28,63, 131,155. |
To optimize clinical outcomes, using related targeting products to increase genus abundance and enhance tumor response to IL-10 + CpG oligodeoxynucleotide. |
| Streptococcus sanguinis (species; genus: Streptococcus, phylum: Firmicutes) | Protective in the gut microbiome 6. | Increasing species abundance in the gut microbiome through strategies like FMT or other appropriate dysbiosis-targeting approaches. |
| Saccharmycetes (class; phylum: Ascomycota) | Protective, triggering antitumor immunity and serving as a positive prognostic marker 15. | Increasing class abundance through appropriate dysbiosis-targeting approaches. However, further investigation at genus or species level is recommended. |
| Saccharopolyspora (genus; Phylum: Actinobacteria) | Protective and positive prognostic marker in the intratumor microbiome) 15 | Increasing genus abundance in the intratumoral microbiome through strategies like tumor-colonizing bacteria or other appropriate dysbiosis-targeting approaches. |
| Salmonella (genus; phylum: Proteobacteria) | Protective in the intratumor microbiome, activating the inflammatory cytokine interleukin-1beta and promoting NK cell activation 154. | Increasing genus abundance in the intratumoral microbiome through strategies like tumor-colonizing bacteria or other appropriate dysbiosis-targeting approaches. |
| Selenomonasb(genus; phylum: Firmicutes) | Carcinogenic in the gut microbiome 8, but protective in the oral microbiome 20. | Genus depletion in the gut microbiome, with an increase in the oral microbiome, using appropriate dysbiosis-targeting strategies. However, further investigation at the strain level is recommended. |
| Shigella sonnei (species; genus: Shigella, phylum: Proteobacteria) | Carcinogenic due to upregulation of oncogenic pathways, immunosuppression, and TME reprogramming 17. | Species depletion: however, further investigation at the strain level, particularly regarding oral-gut-intratumoral interactions and their clinical effects, is recommended. |
| Solobacterium (genus; phylum: Firmicutes) | Carcinogenic in the oral microbiome 85. | Genus depletion in the oral microbiome by targeting dysbiosis appropriately. |
| Sphingomonas (genus; phylum: Proteobacteria) |
Carcinogenic in the intratumor microbiome49; protective and a positive prognostic marker in the gut microbiome15,16. |
Genus depletion in the intra-tumor microbiome and increasing in the gut microbiome by targeting dysbiosis appropriately. |
| Spirochaeta (genus; phylum: Spirochaetes) | Protective and positive prognostic marker in the intratumor microbiome)15,16, but carcinogenic in the oral microbiome 105. | Genus depletion in the oral microbiome, with an increase in the intratumor microbiome, using appropriate dysbiosis-targeting strategies. |
| Streptococcus (genus; phylum: Firmicutes) | Carcinogenic in the oral, intratumor and gut microbiomes 20, 66. | Genus depletion in the oral, intratumor and gut microbiome, using appropriate dysbiosis-targeting strategies. However, further investigation at species and strain level is recommended. |
| Streptococcus mitis (species; genus: Streptococcus, phylum: Firmicutes) | Protective in the oral microbiome 20. | Increasing genus abundance in the oral microbiome through appropriate dysbiosis-targeting approaches. |
| Streptococcus anginosus (sepecies; genus: Streptococcus, phylum: Firmicutes) | Carcinogenic in the gut microbiome 85. | Species depletion in the gut microbiome, using appropriate dysbiosis-targeting strategies. |
| Streptococcus australis (sepecies; genus: Streptococcus, phylum: Firmicutes) | Protective in the oral microbiome 85. | Increasing species abundance in the oral microbiome through appropriate dysbiosis-targeting approaches. |
| Streptococcus mutans (sepecies; genus: Streptococcus, phylum: Firmicutes) | Carcinogenic in the gut microbiome 6. | Species depletion in the gut microbiome, using appropriate dysbiosis-targeting strategies. |
| Streptococcus oralis (sepecies; genus: Streptococcus, phylum: Firmicutes) | Carcinogenic in the gut microbiome 85. | Species depletion in the gut microbiome, using appropriate dysbiosis-targeting strategies. |
| Streptococcus salivarius (sepecies; genus: Streptococcus, phylum: Firmicutes) | Protective in the oral microbiome 85. | Increasing species abundance in the oral microbiome through appropriate dysbiosis-targeting approaches. |
| Streptococcus thermophilus (sepecies; genus: Streptococcus, phylum: Firmicutes) | Protective in the oral microbiome 85. | Increasing species abundance in the oral microbiome through appropriate dysbiosis-targeting approaches. |
| Streptococcus vestibularis(sepecies; genus: Streptococcus, phylum: Firmicutes) | Carcinogenic in the gut microbiome 85. | Species depletion in the gut microbiome, using appropriate dysbiosis-targeting strategies. |
| Streptomyces (genus; phylum: actinobacteria) | Protective in the intratumor microbiome, triggering antitumor immunity and NK cell infiltration 48; positive prognostic marker 15. | Increasing genus abundance in the intratumoral microbiome through strategies like tumor-colonizing bacteria or other appropriate dysbiosis-targeting approaches. |
| Subdoligranulum (genus; phylum: Firmicutes) | Protective in the gut microbiome 109. | Increasing genus abundance in the gut microbiome through appropriate dysbiosis-targeting approaches. |
| Tannerella (genus; phylum: Bacteroidetes) | Protective in the oral microbiome 20. | Increasing genus abundance in the oral microbiome through appropriate dysbiosis-targeting approaches. |
| Treponema (genus; phylum: Spirochaetes) | Carcinogenic in the oral microbiome 105. | Genus depletion in the oral microbiome, using appropriate dysbiosis-targeting strategies. |
| Turicibacter (genus; phylum: Firmicutes) | PIPN preventive, interfering with astrocytes and TLR4/p38MAPK pathway, and regulating NK cell function 97,98,100. | The genus needs to be increased in the gut to reduce PIPN, using, for example, FMT or other appropriate approaches to target dysbiosis. |
| UCG-005 (genus, Phylum: Firmicutes) | PIPN Inducer, activating astrocytes and the TLR4/p38MAPK pathway, and regulating NK cell 97,98,100. | The genus needs to be decreased in the gut to reduce PIPN, using, for example, antibiotics, dietary changes, or other appropriate approaches to target dysbiosis. |
| Veillonella (genus; phylum: Firmicutes) | Carcinogenic in the gut microbiome 8, 52. | Genus depletion in the gut microbiome, using appropriate dysbiosis-targeting strategies. |
| Veillonella atypica (species; genus: Veillonella, phylum: Firmicutes) | Carcinogenic in the gut microbiome 85, 148. | Species depletion in the gut microbiome, using appropriate dysbiosis-targeting strategies. |
| Veillonella parvula(species; genus: Veillonella, phylum: Firmicutes) | Carcinogen in the gut microbiome 85. | Species depletion in the gut microbiome, using appropriate dysbiosis-targeting strategies. |
5-FU: 5-Fluorouracil; CDA: Cytidine Deaminase; CpG: Unmethylated cytosine-phosphate-guanine; FMT: Fecal microbiota Transplant; IL-10: Interleukin 10; MAPK: Mitogen-activated protein kinase; NET: Neutrophil extracellular traps; NGPs: Next generation probiotics; NKA: Natural killer cell activity; OIPN: oxaliplatin-induced neuropathy; PD-1: Programmed Cell Death Protein 1; PIPN: paclitaxel-induced peripheral neuropathy; TLR4: Toll-like receptor 4.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



