
Submitted:
16 December 2024
Posted:
17 December 2024
You are already at the latest version
Abstract
This study presents the second phase of a year-long investigation comparing multiple PCR anal-yses and histological examinations to confirm the presence of characteristic lesions for each path-ogen in three different regions of Latin America. More than 20 agents, including DNA and RNA viruses, bacteria and microsporidia, have been targeted. In addition to wild P. vannamei, which was studied previously, samples of wild P. stylirostris and P. monodon were included. Notably, a positive PCR test result alone does not confirm the presence of a viable pathogen or a disease state. Similarly, positive PCR results do not necessarily correlate with the presence of histological le-sions characteristic of the targeted pathogen. Wenzhou shrimp virus 8 (WzSV8) was found to be widespread among shrimp in all regions, including both farm-raised and wild populations. Histo-pathological analysis indicated that shrimp typically presented coinfections, such as WzSV8, dec-apod hepanhamaparvovirus (DHPV), chronic midgut inflammation, and tubule disten-sion/epithelial atrophy, consistent with the toxicity of Pir A/B or another bacterial toxin. Bacterial muscle necrosis was also found in some regions. In general, bacterial infection was the dominant pathology in all three regions during the year. We also postulate that both WzSV8 and DHPV can infect not only hepatopancreatic cells but also cells in the ceca and intestine.
Keywords:
Wenzhou shrimp virus 8 - WzSV8
; Hepanhamaparvovirus – DHPV
; Postlarvae – PL. Viral inclusion-VIN
1. Introduction
This study is the second phase of a year-long investigation comparing multiple PCR analyses and histological examinations to confirm the presence of characteristic lesions for each pathogen in three different regions of Latin America (Intriago et al., 2024). More than 20 agents, including DNA and RNA viruses, bacteria and microsporidia, have been targeted. The following DNA viruses were used: DHPV, MHBV, DIV1, WSSV and IHHNV. RNA viruses: WzSV8/PvSV, PvNV, CMNV, IMNV, YHV, TSV and MrNV. Bacteria and others: Spiroplasma, Rickettsia-like bacteria (RLB), necrotizing hepatopancreatitis bacteria (NHP-B), Vibrio spp., AHPND, EHP and other non-EHP microsporidia.
The initial phase of the study revealed the critical importance of primer selection in obtaining reliable results (Intriago et al., 2024). They found significant variations in outcomes with different primers, leading to the conclusion that seasonal evaluations are necessary to identify the most effective primer combinations. The lack of consistency found in several primers for viruses could be attributed to the genetic diversity of pathogens affected by the geographical distribution of the host, environmental factors, and adaptations of the viruses to different environmental factors in different geographical areas. There is also the possibility of false positive PCR test results arising from EVE that include the sequence of the PCR target. WzSV8 was the most common pathogen detected by PCR and by histology in all regions sampled regardless of the size of the animal and/or the environment, including wild animals. WzSV8 was rarely found alone. Generally, WzSV8 was accompanied by lesions in the midgut that were presumed to be due to bacteria and to a lower prevalence of DHPV, WSSV or BP. WzSV8 was found in cultured larvae as well as in wild animals (Region 1), the latter at 100% prevalence. Therefore, it is not wrong to assume that this virus is endemic to this region and is already widespread in the world.
Intriago et al. (2024) also found differences in the level of coinfection between the 3 regions, which was related to the culture system or the health status of the broodstock/source of the postlarvae. The midgut histopathology results were compatible with SHPN, chronic AHPND and/or RLB/NHP. These findings are also consistent with those previously described that Vibrio species-associated diseases are predominant in the region. (Morales-Covarrubias et al.,2018, Aranguren Caro et al., 2020),
Similarly, most samples were collected by clients monitoring shrimp health or addressing disease outbreaks in their ponds. Notably, most broodstock specimens appeared healthy, serving as reference points for assessing the potential transmission of pathogens into the production cycle via postlarvae used for pond stocking. Additionally, wild penaeid shrimp (P. vannamei, P. stylirostris and P. monodon) from various regions were included to evaluate potential agents and their interconnection with the aquaculture sector.
1.1. Rationale of the Study
This study was not an epidemiological investigation but rather a prevalence analysis of various shrimp pathogens in randomly collected samples from hatcheries, farms, maturation units, and wild animals across three different regions in Latin America from May 2023 to October 2023. More than 20 agents, including DNA and RNA viruses, bacteria and microsporidia, have been targeted. This work represents the second phase of a one-year study aiming to identify potential pathological variations using histology and PCR in several Latin American countries, following the initial report by Intriago et al. (2024).
2. Materials and Methods
2.1. Sample Collection
More than 140 samples from surveillance sampling of P. vannamei originating from three different regions in Latin America were analyzed throughout the period spanning from May 2023 to October 2023. One region draws its culture water from the Atlantic Ocean, and the other two from the Pacific Ocean. Regions 1 and 3 were the same regions as described in the first surveillance described in (Intriago et al., 2024), whereas the third region was a different region; for this reason, we named it region 4. Regions 1 and 4 are in the Pacific Ocean, and region 3 is in the Atlantic Ocean.
Sampling when possible included animals from hatcheries, broodstock centers, farms, and wild animals. It should be noted that shrimp sampled for PCR and histology were different individuals from the same populations. To protect client privacy, the countries or exact locations from which the samples were obtained will not be revealed here. However, the clients from World Organization for Animal Health (WOAH) member countries were informed of their responsibility to notify the competent authority of their country regarding positive test results for any shrimp pathogens listed by WOAH or arising from any unusual incidences of mortality. It would then be the responsibility of the relevant competent authorities from those member countries to report to the WOAH.
2.2. PCR Methods Used
DNA was extracted from whole larvae, tissue or organs fixed in 90% alcohol following the manufacturer’s protocol (Omega, Bio-Tek E.Z.N.A. tissue DNA kit). In brief, each sample was minced with sterilized scissors and then ground using a microcentrifuge pestle. Approximately 200 mg of tissue was then transferred to a clean 1.5 mL Eppendorf tube. To this end, 500 μL of tissue lysis buffer (TL) and 25 μL of Omega Biotek (OB) protease solution were added, and the mixture was vortexed and then incubated in a thermoblock at 55°C for approximately 3 h with vortexing every 30 minutes. RNA was removed by adding 4 μL of RNase A (100 mg/mL), and after mixing, the sample was kept at room temperature for 2 min. The sample was then centrifuged at 13,500 RPM for 5 min, and the supernatant was carefully transferred to a new 1.5 mL Eppendorf tube. Then, 220 μL of BL buffer was added, and the mixture was vortexed and incubated at 70°C for 10 mins. Next, 220 μL of 100% ethanol was added, the mixture was vortexed, and the contents were passed through a HiBind® DNA Mini Column into a 2 mL collection tube. The columns were then centrifuged at 13,500 RPM for 1 min, after which the filtrate was discarded. Subsequently, 500 μL of HBC buffer (diluted with 100% isopropanol) was added to the column, and the sample was spun at 13,500 RPM for 30 seconds. The filtrate was discarded, the column was washed twice with 700 μL of DNA wash buffer diluted with 100% ethanol, and the sample was centrifuged at 13,500 RPM for 30 seconds. The filtrate was discarded. This step was repeated. The column was then centrifuged at 13,500 RPM for 2 min to dry it. The dried column was placed in a new nuclease-free 1.5 mL Eppendorf tube, and 100 μL of elution buffer, which was heated to 70°C, was added to the column. The sample was allowed to sit for 2 min before being centrifuged at 13,500 RPM for 1 min. This elution step was repeated. The eluted DNA was then stored at -20°C until needed.
RNA was extracted from whole larvae, tissue or organs fixed in 90% alcohol following the manufacturer’s protocol (Omega, Bio-Tek E.Z.N.A. Total RNA Kit). In brief, each sample was minced with sterilized scissors and then ground using a microcentrifuge pestle. Approximately 200 mg of tissue was then transferred to a clean 1.5 mL Eppendorf tube. Then, 700 μL of TRK Lysis Buffer was added, and the tube was left at room temperature for approximately 3 h with vortexing every 30 minutes. The sample was then centrifuged at 13,500 RPM for 5 min, and the supernatant was carefully transferred to a new 1.5 mL Eppendorf tube to which 420 μL of 70% ethanol was added. After vortexing to mix thoroughly, the contents were passed through a HiBind® RNA Mini Column into a 2 mL collection tube. The columns were then centrifuged at 13,500 RPM for 1 min, after which the filtrate was discarded. Subsequently, 500 μL of RNA Wash Buffer I was added to the column, and the sample was spun at 13,500 RPM for 30 seconds. The filtrate was discarded, and the column was washed twice with 500 μL of RNA Wash Buffer II and diluted with 100% ethanol. The column was then centrifuged at 13,500 RPM for 1 min to dry it. The filtrate was discarded. This step was repeated. The column was then centrifuged at 13,500 RPM for 2 min to dry it. The dried column was placed in a new nuclease-free 1.5 mL Eppendorf tube, and 70 μL of nuclease-free water was added to the column. The sample was centrifuged at 13,500 RPM for 2 min. This elution step was repeated. The eluted RNA was then stored at -70°C until needed. The samples used for extraction were as follows:
2.3. Samples Used for Extraction
| IHHNV: | 2 pleopods per animal pool of 5 animals. |
| PvNV: | 2 pleopods per animal pool of 5 animals. |
| Spiroplasma: | DNA pool of 2 pleopods per animal, pool of 5 animals; 10 |
| gill pools of animals; whole hepatopancreas pools of 5 animals | |
| WSSV: | 10 gills per animal pool of 5 animals |
| TSV: | 10 gills per animal pool of 5 animals |
| MrNV and XSV: | 10 gills per animal pool of 5 animals |
| IMNV: | 10 gills per animal pool of 5 animals |
| YHV-GAV: | 10 gills per animal pool of 5 animals |
| DHPV: | Whole hepatopancreas pool of 5 animals |
| MHBV: | Whole hepatopancreas pool of 5 animals |
| DIV1: | Whole hepatopancreas pool of 5 animals |
| WzSV8: | Whole hepatopancreas pool of 5 animals |
| PvSV: | Whole hepatopancreas pool of 5 animals |
| RLB: | Whole hepatopancreas pool of 5 animals |
| NHPB: | Whole hepatopancreas pool of 5 animals |
| EHP: | Whole hepatopancreas pool of 5 animals |
| AHPND: | Whole hepatopancreas pools of 5 animals |
| Microsporidia: | 0.5 grams of tail muscle per animal pool of 5 animals |
| CMNV: | 0.5 grams of tail muscle per animal pool of 5 animals |
| Vibrio Community: | DNA pool of 2 pleopods per animal pool of 5 animals. 0.5 grams of tail muscle per animal pool 5 of animals |
- The Following Pathogens Were Screened with the Methods Tested.
Wenzhou shrimp virus 8 (WzSV8) was identified by Lanza (2022), Cruz-Flores et al. (2022), and Srisala et al. (2023). Hepanhamaparvovirus (DHPV) Phromjai et al. (2002), Umesha et al. (2006), Pantoja and Lightner (2000), Srisala et al. (2021). Macrobrachium Bidnavirus (MrBdv) Gangnonngiw et al. (2023), Rickettsia-like bacteria (RLB) Nunan et al., (2003), Potts et al. (2020). Necrotizing hepatopancreatitis bacteria (NHP-B) Aranguren et al. (2010). Spiroplasma Nunan et al. (2004), Bastian et al. (2004), Ding et al. (2007). Non-EHP Microsporida Pasharawipas and Flegel (1994) Sokolova et al. (2015), Weiss and Vossbrinck (1999). Infectious hypodermal and hematopoietic necrosis virus (IHHNV) Nunan et al. (2000), Tang et al. (2000), Tang and Lightner (2006), Tang et al. (2007a). Enterocytozoon hepatopenaei (EHP) Tourtip et al. (2009), Tangprasittipap et al. (2013), Tang et al. (2015), Jaroenlak et al., (2016), Tang et al., (2017). Vibrio spp. Thompson et al. (2004). Acute Hepatopancreatic Necrosis (AHPND) Dangtip et al. (2015). Decapod iridescent virus 1 (DIV1) Qiu et al., (2017). White spot syndrome virus (WSSV) Lo et al. (1996). Penaeus vannamei Noda virus (PvNV) Tang et al. (2007b). Covert mortality Nodavirus (CMNV) Zhang et al. (2017). Infectious Myonecrosis Virus (IMNV) Poulos and Lightner (2006). Yellow Head Virus (YHV) Mohr et al., (2015). Taura Syndrome Virus (TSV) Nunan et al. (1998), Navarro et al. (2009). Machrobrachium Nodavirus (MrNV) Gangnonngiw et al. (2020). Extra small virus XSV Senapin et al. (2012).
2.4. Histopathology
For histological analysis, samples were prepared following the procedures outlined by Bell and Lightner (1998). Briefly, the samples were fixed in Davidson’s AFA for at least 24 h or 72 h in the case of broodstock before processing for routine histological analysis of 5 µm thick tissue sections stained with hematoxylin and eosin (H&E). In addition, methyl green-pyronin-modified stain was also used to distinguish DNA and RNA. From each juvenile to broodstock shrimp sample, 2 to 4 paraffin blocks were prepared. For postlarvae (PL), approximately 1,500 animals were taken from each tank, and more than half (i.e., approximately 750+) were fixed in Davidson’s AFA for 24 h and embedded in paraffin blocks, each containing approximately 35 to 50 PLs.
3. Results
This section may be divided by subheadings. This should provide a concise and precise description of the experimental results, their interpretation, and the experimental conclusions that can be drawn.
3.1. PCR Results
In region 1, as in the other 2 regions, WzSv8 was the most important pathogen detected. Similar to previous studies, both the UAZ and Biotech primers (Cruz Flores et al. 2023; Srisala et al. 2024) produced the highest prevalence (Table 1). In region 4, WzSV8 was present in 100% of the samples tested. The average prevalence of WzSV8 in all 3 regions was 82%. WzSv8 was detected in the PL, farm animal, and broodstock of P. vannamei. It was found in wild P. vannamei, P. stylirostris and P. monodon.
The hepanhamaparvovirus (DHPV) was most common in regions 1 and 4. Region 3 was detected in only one sample of PL. DHPV was most commonly found in farm animals, followed by broodstock. In contrast to WzSV8, DHPV was never detected in wild stocks (Table 2). In the first study (Intriago et al., 2024), we included Macrobrachium Bidnavirus (MrBdv) because of its histological similarity to HDPV lesions. However, it was never detected. The most effective primers for DHPV were those reported by Phromjai et al. (2002) and Umesha et al. (2006).
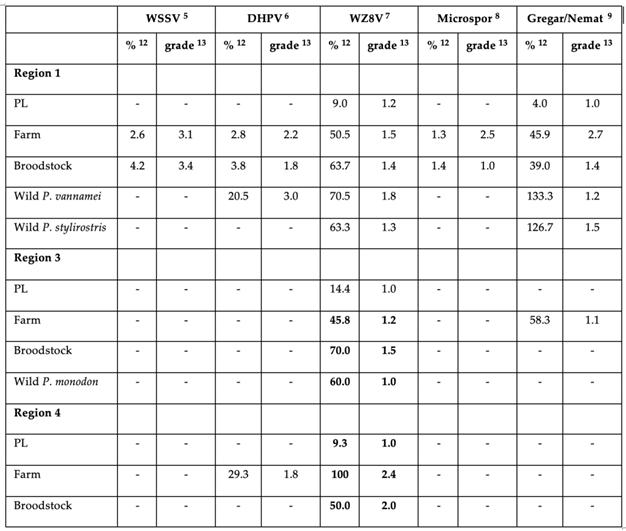
The most common PCR screening tools for intracellular bacteria in shrimp are Rickettsia-like organisms (RLOs) (Nunan et al. 2003) and the necrotizing hepatopancreatitis bacterium NHPB (Aranguren et al. 2010) (Table 3). We compared these two methods with the Rickettsia genus-specific primers Rp877p/Rp1258n (Potts et al. 2020). In general, only RLO was detected, and the Rickettsia genus-specific primers Rp877p/Rp1258n were most prevalent. Figure 1 shows that between intracellular bacteria, RLO was more important than NHPB and that Rickettsia genus-specific primers became more prevalent during the second part of the study.
We tested three methods to detect Spiroplasma: Nunan et al. (2004), Bastian et al. (2004), and Ding et al. (2007). The highest prevalence was obtained using S. penaei primers (Nunan et al. (2004), averaging 20% prevalence in the 3 regions, followed by S. mirum primers (Ding et al. 2007). The average prevalence was 9% in the 3 regions. The prevalence of Spiroplasma ribosomal DNA in Creutzfeldt‒Jakob disease (CJD) was low when primers were used (Bastian et al. 2004), and Spiroplasma ribosomal DNA was present only in region 1. In general, this pathogen was detected mainly in farm animals, with a very low prevalence (1/15) in PL and two cases in broodstock (2/37) (Table 4). No positive bacteria were detected in the wild animals. There was no detection in region 4 at any stage.
The presence of non-EHP microsporidia (Table 5) was determined using three methods. The first method, developed by Pasharawipas and Flegel (1994), was designed to detect Agmasoma penaei in P. merguensis and P. monodon. This method yielded the highest prevalence, averaging 85% across all three regions. Although the lowest concentration was observed in PL, positive results were obtained for all stages, including farm animal, broodstock, and wild samples. We also tested a more recent method (Sokolova et al. 2015) designed to be selective for A. penaei in Litopenaeus setiferus from the Gulf of Mexico. However, this method showed the lowest prevalence, averaging 21% across the three regions. Among farm animals, PL, broodstock, and wild animals had very low or nonexistent prevalence, while the highest prevalence (80%) was found in region 4. The third method utilized universal primers widely used for microsporidians (Weiss and Vossbrinck, 1999) and provided intermediate results, with an average prevalence of 47% across the three regions. The lowest prevalence was observed in the PL samples.
When testing IHHNV (Table 6), it is necessary to distinguish viruses from endogenous viral elements (eve). For this reason, we used all four PCR primers (Nunan et al. 2000, Tang et al. 2000, Tang and Lightner, 2006, Tang et al. 2007a). All these viruses must be present for a functional virus to replicate. However, if some but not all primers amplify, this pattern is a clear indication that the PCR primers are measuring an eve and not the complete virus.
The primers with the highest and lowest prevalence in descending order were 309 F/R, 392 F/R, 389 F/R and 77012 F/77353R, averaging 45%, 40%, 39% and 24%, respectively, in the 3 regions. These results revealed average virus and eve prevalence rates of 23% and 26%, respectively, in the 3 regions. Interestingly, P. monodon from region 3 had zero prevalence of IHHNV virus and 50% prevalence of eve, but when primers for an exclusive sequence for eve in monodon MG831F/R primers were used [19,21], all animal samples were positive. Except for one sample (1/3) of PL in region 3, all farm animals and broodstock were negative.
Based on a previous study by Intriago et al. (2024), we tested different sets of primers for the detection of Enterocytozoon hepatopenaei (EHP). Table 7 summarizes the results obtained from six different primers used in our tests. Generally, most existing PCR detection methods target the EHP small subunit ribosomal RNA (SSU rRNA) gene (SSU-PCR). However, our results clearly indicate that these methods often produce false positives due to the cross-reactivity of SSU-PCR primers with DNA from closely related microsporidia. For now, we consider the method targeting the spore wall protein (SWP) gene of EHP, as described by Jaroenlak et al. (2016), to be the most reliable. This method minimizes false positives and provides a more accurate detection of EHP.
Th white spot syndrome virus (WSSV) was detected in only regions 1 and 3. In region 1, its prevalence among PL, farm animals and broodstock was 20%, 45% and 10%, respectively. It was not present in wild P. vannamei but was detected in wild P. stylirostris, with a 13% prevalence. In region 3, it was present in PL and broodstock, with 33 and 90% prevalence, respectively. It was not detected in wild P. monodon. Pathogens such as CMNV, TSV, and DIV1.
The following pathogens were never detected in any region: MrBdv, PvNV, IMNV, YHV, MrNV and XSV. A comparison of the most prevalent pathogens in this study with those found during the first survey revealed that WzSV8 had a high prevalence (> 80%) during the year of the study and that the prevalence of almost all pathogens increased in the last six months, with special attention given to EHP (Table 7, Figure 2).
3.2. Histopathology Results
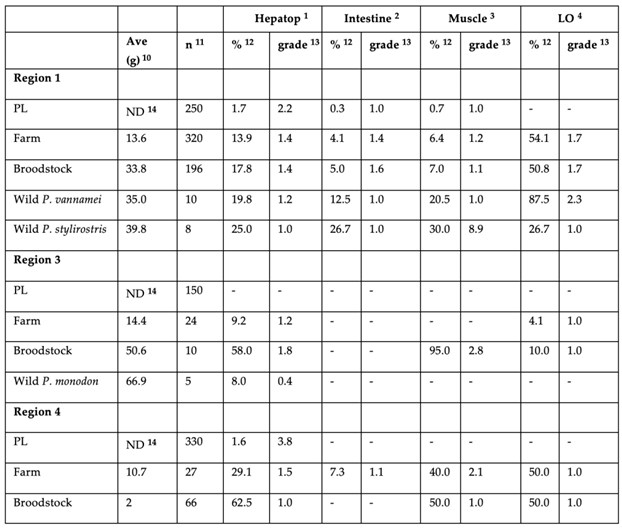
Approximately 150 samples were analyzed for histopathology in this second part of the survey. In general, coinfections that consisted of WzSV8, DHPV, chronic midgut inflammation and tubule distension/epithelial atrophy consistent with Pir A/B toxicity or other bacteria were the most common pathologies in all regions (Table 8).
WzSV8 viral inclusions (VINs) are basophilic inside vacuoles within the HP tubule cell cytoplasm and are mostly circular in cross section, while those of DHPV VIN are in the nuclei, mostly magenta, ovoid and separated from marginated chromatin by an unstained space. On occasion, especially with small VINs, a crescent-shaped nucleolus can sometimes be seen, pushed aside by the expanding viral inclusion. Hepatopancreas infection with both DHPV and WzSV8 is not uncommon (Figure 3). Histological staining using methyl green-pyronin-modified stain, which is used to distinguish DNA and RNA, was used, and it is tempting to suggest that both DHPV and WzSV8 are not exclusive to the hepatopancreas but also infect the ceca and intestine (Figure 3). WzSV8 with lighter double inclusions (LDI) (Srisala et al. 2023) can be considered pathognomonic for WzSV8 because of the unique morphology of additional (usually smaller) eosinophilic inclusions adjacent to single, circular basophilic inclusions in vacuoles. In some specimens that were rounded, sloughed WzSV8-infected cells were noted in the lumen of the tubules. Interestingly, WzSV8 VIN, including both basophilic and LDI VIN, was also found frequently in the ovaries of some specimens and was previously reported (Intriago et al. 2024) at both the developing and mature stages.
WzSV8 VINs were detected in PL, farm animals, broodstock and wild animals in all 3 regions. Its prevalence ranged from 9% in larvae in regions 1 and 4 to 100% in farm animals in region 4 (Table 8). DHPV VIN were never found in PL, and they were not found either in any sample of region 3. Region 1 included farm animals, broodstock and wild P. vannamei but not P. stylirostris.
After WzSV8, the next most important disorder observed in the hepatopancreas was chronic midgut inflammation and tubule distension/epithelial atrophy, consistent with Pir A/B or different bacterial toxicity (Table 8 and Figure 4). In general, lesions in the hepatopancreas were observed in all sizes from the PL to the broodstock; however, it is easier to note from Table 8 that the larger the size of the lesion was. Interferingly, these lesions were found in all species of wild Penaeus sampled in regions 1 and 3. Baculovirus penaei (BP), also known as singly enveloped nucleopolyhedron virus (PvSNPV), was occasionally detected in the hepatopancreas of broodstock from 3 animals from 196 in region 1 and 5 animals from 10 in region 3 (Figure 4).
In addition, inflammation of intestinal walls with hemocyte infiltration (hemocytic enteritis) was commonly found in all sizes in region 1 and in farm animals in region 4. In region 1, this lesion was always found together with muscle necrosis and the presence of a pack of bacteria with hemocyte infiltration. Melanized reactions in the cuticle of the exoskeleton were sometimes part of this bacterial infection (Figure 5).
Lymphoid organ spheroids (LOSs) were never present in PL and wild P. monodon but could be found in farms, broodstock and in wild P. vannamei and P. stylirostris (Table 8 and Figure 6). In contrast to region 3, regions 1 and 4 had the highest prevalence. Interestingly, LOS was also found in wild P. vannamei and P. stylirostris.
Histopathology of the PLs (regions 1 and 4) revealed sloughing cells and slight hemocyte infiltration with a clear bacterial infection in the hepatopancreatic tubules (Figure 7). WzSV8 VIN was also detected (Figure 7E, F).
Lesions caused by WSSV were reported only in region 1, farm and broodstock. The prevalence was low, but the lesion grade was high (>3) (Table 8). WSSV VIN was detected in the stomach, gill, hematopoietic tissue, antennal gland, epidermis and stomach (Figure 8A). IHHNV was observed in 1 sample from farm animals out of 64 in region 1. Cowdry type A inclusions, which are typical of IHHNV, were observed in the rostrum, pleopod and nervous tissue (Figure 8B).
Clusters of microsporidian spores were mainly observed in hepatopancreatic cells within striated muscle sarcomeres antennal glands (Figure 9). Histopathological lesions caused by microsporidia were found only in region 1 (Table 8).
Gregarines and nematodes of all sizes, including wild P. vannamei and P. stylirostris, were found in region 1. It was found only in farm animals in region 3 and was absent in animals in region 4. Larval nematodes (Figure 10A, B) were commonly found encysted in the wall of the foregut near the junction with the midgut. Gregarine trophozoites were within the lumen of the anterior midgut cecum and midgut intestine, while gametocytes were within the lumen of the posterior midgut cecum (Figure 10C to F).
4. Discussion
The present study details the second part of a one-year study of histopathological lesions and identifies agents using PCR in P. vannamei samples collected in hatcheries, farms and maturation units from three distinct regions in Latin America. Additionally, samples from wild P. vannamei, P. stylirostris, and P. monodon were also included. This study compares different sets of primers targeting various agents to understand potential geographical variations and their specificity. An important aspect to note is the variation in coinfection levels among the three regions, which is linked to the culture system or the health status of the broodstock/source of the postlarvae.
WzSV8 emerged as the most prevalent pathogen detected across all sampled regions by both PCR and histology, irrespective of animal size or environment, including wild penaeid samples from both the previous (Intriago et al., 2024) and current studies. WzSV8 VINs were rarely found in isolation; they were often accompanied by midgut lesions presumed to be caused by bacteria, along with a lower prevalence of DHPV, WSSV, or BP. The presence of WzSV8 in both farmed and wild animals suggests that it is endemic to the region and has a global distribution.
In this study, the midgut histopathology findings were indicative of potential bacterial infections such as septic hepatopancreatic necrosis (SHPN), chronic AHPND, and/or RLO. However, due to the lack of in situ hybridization capability, we were unable to directly link these names to specific infections or digestive system alterations observed. Consequently, we could not confirm whether the pathogens identified in the lesions corresponded to those detected by PCR in samples from different shrimp of the same source. Like previous findings reported by Intriago et al. (2024), the high prevalence of PCR samples positive for Vibrio, RLB, and AHPND, coupled with histopathological lesions in the hepatopancreas and intestine (with bacteria), strongly suggests a potential association between these bacterial pathogens and the observed lesions. These findings further support the notion that Vibrio species and associated or bacterial diseases are predominant in the region (Intriago et al., 2024).
In Region 1, situated in the Pacific Ocean, there was minimal biosecurity and no rigorous selection process; broodstock were chosen from farms without considering their health status. In contrast, animals from region 3 are from an open farm in the Atlantic Ocean that lacks biosecurity measures but uses specific pathogen-free (SPF) broodstock and larvae. Similarly, Region 4 animals are sourced from a superintensive farm with lined ponds in the Pacific Ocean, utilizing well water and employing SPF broodstock and larvae.
Histopathological lesions in the PLs (postlarvae) from regions 1 and 4 were primarily of bacterial origin. The hepatopancreatic tubules exhibited sloughing cells and slight hemocyte infiltration, indicative of a clear bacterial infection. WzSV8 VIN was also observed (Figure 7). High and sudden mortalities of P. vannamei PL were reported in Latin American hatcheries (Intriago et al., 2023). These mortalities were attributed to a Vibrio species carrying VpPirAB genes. Histopathology revealed massive sloughing and detachment of hepatopancreocytes. This condition was tentatively termed post larval acute hepatopancreatic necrosis disease (PL-AHPND) to distinguish it from other pathologies affecting post larval shrimp (Intriago et al., 2023).
It is intriguing that pathogens such as Spiroplasma and EHP were detected by PCR but were not histopathologically detected. Specifically, in the case of Spiroplasma, there is a notable possibility of the presence of S. mirum, especially in farm animals in region 3 (Table 4). For EHP, it is worth noting the increase in prevalence from the first survey (Intriago et al., 2024) to the present study (Figure 2). Although there are still key factors to be determined, EHP, in conjunction with opportunistic bacteria and Propionigenium and unknown stress factors, could be the primary cause of a condition termed white feces, which poses a significant threat to P. vannamei culture (Munkongwongsiri et al., 2022).
In penaeid species, the lymphoid organ plays an important role in immune defense and is possibly the major phagocytic organ (Rusaini and Owens, 2010). It is also a major site for viral degradation within lymphoid organ spheroids (LOSs) (Rusaini and Owens, 2010). LOSs were found in all regions, all sizes and in wild P. vannamei and P. stylirostris, except for PL and P. monodon (Table 8). The extensive LOS formation observed in a previous report (Intriago et al. 2024) might be related to the tolerance of penaeid prawns to bacterial and viral infection.
PCR analysis revealed that 26% of the IHHNV was associated with EVE, while 23% might have arisen from infectious IHHNV (Table 6). It is important to note that only one sample of 69 (from farm animals from region 1) resulted in pathognomonic Cowdry type A inclusion (CAI) lesions (Figure 8 D) (albeit in different specimens from the same source). It is important to realize that when testing samples using multiple PCR primer sets of equal sensitivity and covering different regions of a virus genome, all the pairs must give the expected amplicons. Failure to do so is a strong indication that the primer sets yielding amplicons arose from the EVE and not from the complete virus genomes (Brock et al. 2013, 2022). IHHNV has been shown to be integrated into host genomic DNA and is not associated with histological lesions typical of IHHNV infection. A study by Taengchaiyaphum et al. (2022) revealed that a separate scrambled cluster of genome fragments from a currently existing type of IHHNV also occurred in the genome of the same shrimp specimen and that false positive PCR results could be obtained using methods for IHHNV detection recommended by the World Organization for Animal Health (WOAH). The existence of EVE arising from IHHNV, and other viruses reveals that caution is needed, especially when reporting a virus from a new location for the first time (Saksmerprome et al. 2011, Alday-Sanz et al. 2018).
More than 20 species of Microsporidia belonging to 17 genera have been reported from a variety of decapod species belonging to the families Penaeidae and Caridea (Sokolova et al. 2015). The two most abundant non-EHP microsporidian parasites of penaeids are Agmasoma penaei and Perezia nelsoni, which have completely different life cycles, morphologies, and tissue tropisms. Pasharawipas and Flegel (1994) and Pasharawipas et al. (1994) designed a specific DNA probe to identify the microsporidian Agmasoma as the parasite infecting muscles in both P. merguensis and P. monodon in Thailand. Later, Sokolova et al. (2016) suggested that muscle infections previously attributed to A. penaei might have been due to overlooked P. nelson in dual infections with A. penaei; they believe that these infections are confined to the gonads, while P. nelsoni infections are confined to the muscles. In the present study, an 85% average PCR prevalence of microsporidia was revealed in the three regions. However, according to histopathology, less than 2% of the microsporidia were present. It was found not only in muscles but also in the hepatopancreas and was restricted to farm animals and broodstock from region 1. More studies are needed to understand the dynamics and importance of this group.
A high prevalence of gregarines and nematodes was found, especially in regions 1 and 3, during the whole study. In region 1, they were found in all sizes, including wild P. vannamei and P. stylirostris, at very high concentrations (100%, Table 8). In region 4, they were absent, and their absence might be associated with the liner ponds. In the wild, Penaeids act as intermediate and final hosts for a wide variety of parasites, and they typically infect shrimp through the ingestion of either free-living stages or infected planktonic hosts (Dall et al. 1990; Marcogliese, 2005). Furthermore, the presence of gregarines, microsporidia, trematodes, cestodes, and nematodes in wild penaeids has been well documented over the past six decades. (Kruse 1959, Iversen and Van Meter, 1964, Feigenbaum, 1975). This highlights the potential role of parasites in introducing and spreading pathogens within the ecosystem, even in seemingly healthy individuals.
Throughout the year, it was observed that animals, despite appearing healthy, often carried multiple pathogens, including viruses, bacteria, and metazoan parasites. Generally, a pathogenic agent must be present for disease to occur. However, the presence of that pathogen alone is not always sufficient to cause disease (Armstrong, 1993). A variety of factors can influence whether exposure to an agent will result in disease. For example, stocking density and physical and chemical factors, among others, can cause a host to become immunocompromised and cause disease (Méthot and Alizon, 2014, Díaz-Muñoz, 2017).
In addition, it is common to attribute a viral disease to infection by a single agent. However, multiple infections are commonly found in shrimp culture and may have more serious consequences than infections caused by a single pathogen (Bonnichon et al., 2006). Crustaceans collected from farm and wild habitats can host a taxonomically diverse array of DNA and RNA viruses (Jang et al. 2014). Thus, health outcomes may be influenced by contributions from more than a single agent, which is often not considered in diagnostic laboratories (Kumar et al. 2018, Bateman and Stentiford, 2017). On the other hand, coinfections can also lead to genetic exchange between pathogens, giving rise to recombinant viruses that can impact viral evolution, disease dynamics, and ultimately the fate of the host (Kumar et al., 2018; Bateman and Stentiford, 2017). Coinfections can also play a pivotal role in either reducing or augmenting disease severity. When individual cells are coinfected, one virus typically influences the replication of the other, a phenomenon known as viral interference. Such interactions have been reported between IHHNV and WSSV (Melena et al. 2006), as well as between TSV and YHV (Aranguren et al. 2012; Flegel, 2020).
Mutualistic viruses can help the host by supplying new genes or through epigenetic changes in the host genome with beneficial results (Goic and Saleh, 2012). This tolerance to the pathogen does not mean resistance, so shrimp can still become infected and may develop the disease if there is a trigger (Goic and Saleh, 2012).
We emphasized the critical nature of selecting the correct set of PCR primers specific to the target agents, which can vary based on geographical location. The inconsistency observed in several primers may stem from the genetic diversity of pathogens influenced by host distribution, environmental factors, and viral adaptations to different environmental conditions across geographical areas (Safeena et al. 2012).
Finally, this study highlights the importance of the following:
Geographic Distribution and Variability of Pathogen Prevalence: Understanding how pathogen prevalence varies across different regions is crucial for effective disease management and control in shrimp populations. The critical nature of selecting the correct set of PCR primers specific to the target agents can vary based on geographical location. Existing PCR detection methods can yield false positive results due to primer cross-reactivity with closely related organisms, as well as false positives arising from environmental DNA that includes the PCR target sequence.
Author Contributions
P.I. conceived the study and directed the writing, editing and interpretation of the results. J. G. performed the molecular analysis. B.M. and M.V. performed the histological analysis and contributed to the histopathological examination. Y.C. sampling and processing.
Funding
This research received no external funding.
Acknowledgments
We would like to thank Nicole Cercado and Milena Burgos for their assistance with histological staining and processing, as well as for their review, and Andrés Medina , and Alejandra Montenegro for his contributions to the molecular work.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Alday-Sanz, V; Brock, J; Flegel, T.W; McIntosh, R; Bondad-Reantaso, M; Salazar, M; Subasinghe, R, 2018. Facts, truths and myths about SPF shrimp in Aquaculture. Reviews in Aquaculture. 12: 76-84. [CrossRef]
- Armstrong D (1993) History of opportunistic infection in the immunocompromised host. Clin Infect Dis 1993; 17:S318-21; [CrossRef] [PubMed]
- Aranguren LF, Tang KFJ, Lightner DV (2010) Quantification of the bacterial agent of necrotizing hepatopancreatitis (NHP-B) by real-time PCR and comparison of survival and NHP load of two shrimp populations. Aquaculture. 307: 187-192. [CrossRef]
- Aranguren LF, Tang KFJ, Lightner DV (2012) Protection from yellow head virus (YHV) infection in Penaeus vannamei preinfected with Taura syndrome virus (TSV). Dis Aquat Organ. 98:185-192.
- Bastian FO, Dash S, Garry RF (2004) Linking chronic wasting disease to scrapie by comparison of Spiroplasma mirum ribosomal DNA sequences. Exp Mol Pathol. 77:49–56.
- Bateman KS, Stentiford GD (2017) A taxonomic review of viruses infecting crustaceans with an emphasis on wild hosts. J Invertebr Pathol. 147:86-110. [CrossRef]
- Bell TA, Lightner DV (1998) A handbook of normal penaeid shrimp histology. The World Aquaculture Society, Baton Rouge, LA.
- Bonnichon V, Lightner DV, Bonami JR (2006) Viral interference between infectious hypodermal and hematopoietic necrosis virus and white spot syndrome virus in Litopenaeus vannamei. Dis Aquat Organ. 72:179-184.
- Brock, J. A; Antonio, N; Argue, B, 2013. Genomic IHHN-related DNA sequences in black tiger shrimp. Global Aquaculture Advocate. Sep; 16:1-8.
- Brock, J. A; Intriago, P; Medina, A, 2022. OIE should eliminate IHHNV as a reportable farmed shrimp pathogen. Responsible Seafood Advocate. Dec 15;12(8):1-8.
- Cruz-Flores, R; Andrade, T.P; Mai, H.N; Alenton, R.R.R; Dhar, A.K. 2022. Identification of a Novel Solinvivirus with Nuclear Localization Associated with Mass Mortalities in Cultured Whiteleg Shrimp (Penaeus vannamei). Viruses. 14:2220. [CrossRef] [PubMed]
- Dall W, Hill BJ, Rothlisberg PC, Staples DJ (1990) Parasites. In: Dall W, Hill BJ, Rothlisberg PC, Staples DJ (eds) The biology of the Penaeidae. Advance in Marine Biology. 27. Chapter 12. Academic Press, London, pp 251–275.
- Dangtip S, Sirikharin R, Sanguanrut P, Thitamadee S, Sritunyalucksana K, Taengchaiyaphum S, Mavichak R, Proespraiwong P, Flegel TW (2015) AP4 method for two-tube nested PCR detection of AHPND isolates of Vibrio parahaemolyticus. Aquaculture Reports. 2: 158-162. [CrossRef]
- Díaz-Muñoz SL (2017) Viral coinfection is shaped by host ecology and virus–virus interactions across diverse microbial taxa and environments. Virus evolution. 3:vex011. [CrossRef] [PubMed]
- Ding Z, Bi K, Wu T, Gu W, Wang W, Chen J (2007) A simple PCR method for the detection of pathogenic spiroplasmas in crustaceans and environmental samples. Aquaculture. 272:49-54. [CrossRef]
- Feigenbaum DL (1975) Parasites of the Commercial Shrimp Penaeus vannamei Boone and Penaeus brasiliensis Latreille. Bulletin of Marine Science. 25: 491-514.
- Flegel TW (2020) Research progress on viral accommodation 2009 to 2019. Dev Comp Immunol. 112:103771. [CrossRef] [PubMed]
- Gangnonngiw W, Bunnontae M, Phiwsaiya K, Senapin S, Dhar AK (2020) In experimental challenge with infectious clones of Macrobrachium rosenbergii nodavirus (MrNV) and extra small virus (XSV), MrNV alone can cause mortality in freshwater prawn (Macrobrachium rosenbergii). Virology. 540:30–37. [CrossRef]
- Gangnonngiw W, Bunnontae M, Kayansamruaj P, Senapin S, Srisala J, Flegel TW, Wongprasert (2023) A novel ssDNA Bidnavirus in the giant freshwater prawn Macrobrachium rosenbergii. Aquaculture. 568:739340. [CrossRef]
- Goic B, Saleh MC (2012) Living with the enemy: viral persistent infections from a friendly viewpoint. Curr Opin Microbiol. 15:531–537. [CrossRef] [PubMed]
- Intriago P, Medina A, Espinoza J, Enriquez X, Arteaga K, Aranguren LF, Shinn AP, Romero X (2023) Acute mortality of Penaeus vannamei larvae in farm hatcheries associated with the presence of Vibrio sp. carrying the VpPirAB toxin genes. Aquacult Int 31: 3363-338210.1007/s10499-023-01129-0.
- Intriago, P; Medina, A; Cercado, N; Arteaga, K; Montenegro, A; Burgos, M; Espinoza, J; Brock, J.A; McIntosh, R; Flegel, T, 2024. Passive surveillance for shrimp pathogens in Penaeus vannamei submitted from 3 Regions of Latin America. Aquaculture Reports. 36: 102092. [CrossRef]
- Iversen ES, Van Meter NN (1964) A Record of Microsporidian, Thelohania duorara, parasitizing the shrimp, Penaeus Brasiliensis. Bulletin of Marine Science. 14.: 549-553.
- Jang, I. K; Qiao, G; Kim, S.K, 2014. Effect of multiple infections with white spot syndrome virus and Vibrio anguillarum on Pacific white shrimp Litopenaeus vannamei (L.): mortality and viral replication. J Fish Dis. 37:911-920. [CrossRef] [PubMed]
- Jaroenlak P, Sanguanrut P, Williams BAP, Stentiford GD, Flegel TW, Sritunyalucksana K, Itsathitphaisarn O (2016) A nested PCR assay to avoid false positive detection of the microsporidian Enterocytozoon hepatopenaei (EHP) in environmental samples in shrimp farms. PLoS ONE. 11. [CrossRef] [PubMed]
- Kruse DN (1959) Parasites of the commercial shrimps, Penaeus aztecus Ives, P. duorarum Burkenroad and P. setiferus (Linnaeus). Tulane Studies In Zoology. 7: 123-144. [CrossRef]
- Kumar N, Sharma S, Barua S, Tripathi BN, Rouse BT (2018) Virological and immunological outcomes of coinfections. Clin Microbiol Rev. 31:e00111-17. [CrossRef] [PubMed]
- Lanza (2022). Personal communication.
- Lo CF, Ho CH, Peng SE, Chen CH, Hsu HC, Chiu YL, Chang CF, Liu KF, Su MS, Wang CH, Kou GH (1996) White Spot Syndrome Baculovirus (WSBV) Detected in Cultured and Captured Shrimp, Crabs and Other Arthropods. Dis Aquat Organ. 27:215–225. [CrossRef]
- Marcogliese DJ (2005) Parasites of the superorganism: are they indicators of ecosystem health? Int. J. Parasitol. 35, 705–716. [CrossRef]
- Melena J, Bayot B, Betancourt I, Amano Y, Panchana F, Alday V, Calderón J, Stern S, Roch P, Bonami JR (2006) Preexposure to infectious hypodermal and hematopoietic necrosis virus or to inactivated white spot syndrome virus (WSSV) confers protection against WSSV in Penaeus vannamei (Boone) postlarvae. J Fish Dis. 29:589–600. [CrossRef]
- Méthot PO, Alizon S (2014) What is a pathogen? Toward a process view of host- parasite interactions. Virulence 5, 775–785. [CrossRef]
- Mohr PG, Moody NJG, Hoad J, Williams LM, Bowater RO, Cummins DM, Cowley JA, Crane M StJ (2015) New yellow head virus genotype (YHV7) in giant tiger shrimp Penaeus monodon indigenous to northern Australia. Dis Aquat Org. 115: 263-268.
- Munkongwongsiri N, Prachumwat A, Eamsaard W, Lertsiri K, Flegel TW, Stentiford GD, Sritunyalucksana K (2022) Propionigenium and Vibrio species identified as possible component causes of shrimp white feces syndrome (WFS) associated with the microsporidian Enterocytozoon hepatopenaei. J Invertebr Pathol. 192:107784. [CrossRef]
- Navarro SA, Tang KFJ, Lightner DV (2009) An improved Taura syndrome virus (TSV) RT‒PCR using newly designed primers. Aquaculture. 293:290–292. [CrossRef]
- Nunan LM, Poulos BT, Lightner DV (1998) Reverse transcription polymerase chain reaction (RT‒PCR) used for the detection of Taura Syndrome Virus (TSV) in experimentally infected shrimp. Dis Aquat Organ. 34:87–91. [CrossRef]
- Nunan LM, Poulos BT, Lightner DV (2000) Use of polymerase chain reaction (PCR) for the detection of infectious hypodermal and hematopoietic necrosis virus (IHHNV) in penaeid shrimp. Mar Biotechnol. 2:319–328. [CrossRef]
- Nunan LM, Poulos B, Redman R, Groumellec ML, Lightner DV (2003) Molecular detection methods developed for a systemic rickettsia-like bacterium (RLB) in Penaeus monodon (Decapoda: Crustacea). Dis Aquat Organ. 53:15–23. [CrossRef]
- Nunan LM, Pantoja CR, Salazar M, Aranguren F, Lightner DV (2004) Characterization and molecular methods for detection of a novel spiroplasma pathogenic to Penaeus vannamei. Dis Aquat Organ. 62:255–264. [CrossRef]
- Pantoja CR, Lightner DV (2000) A nondestructive method based on the polymerase chain reaction for detection of hepatopancreatic parvovirus of penaeid shrimp. Dis Aquat Organ. 39:177-182.
- Pasharawipas, T; Flegel, T.W, 1994. A specific DNA probe to identify the intermediate host of a common microsporidian parasite of Penaeus merguensis and P. monodon. Asian Fisheries Sci. 7:157–167.
- Pasharawipas T, Flegel TW, Chaiyaroj S, Mongkolsuk S, Sirisinha S (1994) Comparison of Amplified RNA gene sequences from microsporidian parasites (Agmasoma or Thelohania) in Penaeus merguiensis and P. monodon. Asian Fisheries Sci. 7:169-178.
- Phromjai J, Boonsaeng V, Withyachumnarnkul B, Flegel TW (2002) Detection of hepatopancreatic parvovirus in Thai shrimp Penaeus monodon by in situ hybridization, dot blot hybridization and PCR amplification. Dis Aquat Organ. 51:227–232. [CrossRef]
- Qiu L, Chen MM, Wan XY, Li C, Zhang QL, Wang RY, Cheng DY, Dong X, Yang B, Wang XH, Xiang JH, Huang J (2017) Characterization of a new member of Iridoviridae, Shrimp hemocyte iridescent virus (SHIV), found in white leg shrimp (Litopenaeus vannamei). Sci Rep. 7:11834. [CrossRef]
- Rusaini, Owens L (2010) Insight into the lymphoid organ of penaeid prawns: a review. Fish Shellfish Immunol. 29:367-377. [CrossRef]
- Potts R, Molina I, Sheele JM, Pietri JE (2020) Molecular detection of Rickettsia infection in field-collected bed bugs. New Microbes New Infect. 34:1-6.
- Poulos BT, Lightner DV (2006) Detection of infectious myonecrosis virus (IMNV) of penaeid shrimp by reverse-transcriptase polymerase chain reaction (RT‒PCR). Dis Aquat Organ. 73:69-72.
- Safeena MP, Rai P, Karunasagar I (2012) Molecular Biology and Epidemiology of Hepatopancreatic parvovirus of Penaeid Shrimp. Indian J Virol. 23:191-202.
- Saksmerprome, V; Jitrakorn, S; Chayaburakul, K; Laiphrom, S; Boonsua, K; Flegel, T. W, 2011. Additional random, single to multiple genome fragments of Penaeus stylirostris densovirus in the giant tiger shrimp genome have implications for viral disease diagnosis. Virus Research. 160: 180–190. [CrossRef]
- Senapin S, Jaengsanong C, Phiwsaiya K, Prasertsri S, Laisutisan K, Chuchird N, Limsuwan C, Flegel TW (2012) Infections of MrNV (Macrobrachium rosenbergii nodavirus) in cultivated whiteleg shrimp Penaeus vannamei in Asia. Aquaculture. 338-341:41-46. [CrossRef]
- Sokolova, Y; Pelin, A; Hawke, J; Corradi, N, 2015. Morphology and phylogeny of Agmasoma penaei (Microsporidia) from the type of host, Litopenaeus setiferus, and the type locality, Louisiana, USA. Int J Parasitol. 45:1-16.
- Sokolova Y, Hawke JP (2016) Perezia nelsoni (Microsporidia) in Agmasoma penaei infected Atlantic white shrimp Litopenaeus setiferus (Penaeidae, Decapoda) and phylogenetic analysis of Perezia spp. Complex. Protistology. 10:67–78.
- Srisala J, Thaiue D, Sanguanrut P, Aldama-Cano DJ, Flegel TW, Sritunyalucksana K (2021) Potential universal PCR method to detect decapod hepanhamaparvovirus (DHPV) in crustaceans. Aquaculture. 541:736782. [CrossRef]
- Srisala J, Thaiue D, Saguanrut P, Taengchaiyaphum S, Flegel TW, Sritunyalucksana K (2023) Wenzhou shrimp virus 8 (WzSV8) detection by unique inclusions in shrimp hepatopancreatic E-cells and by RT‒PCR. Aquaculture. 572:739483. [CrossRef]
- Taengchaiyaphum S, Wongkhaluang P, Sittikankaew K, Karoonuthaisiri N, Flegel TW, Sritunyalucksana K (2022) Shrimp genome sequence contains independent clusters of ancient and current Endogenous Viral Elements (EVE) of the parvovirus IHHNV. BMC Genomics. 23:565. [CrossRef]
- Tang KFJ, Durand SV, White BL, Redman RM, Pantoja CR, Lightner DV (2000) Postlarvae and juveniles of a selected line of Penaeus stylirostris are resistant to infectious hypodermal and hematopoietic necrosis virus infection. Aquaculture. 190: 203-210.
- Tang KFJ, Lightner DV (2006) Infectious hypodermal and hematopoietic necrosis virus (IHHNV) in the genome of the black tiger prawn Penaeus monodon from Africa and Australia. Virus Res. 118:185–191.
- Tang KFJ, Navarro SA, Lightner DV (2007a) A PCR assay for discriminating between infectious hypodermal and hematopoietic necrosis virus (IHHNV) and the virus-related sequences in the genome of Penaeus monodon. Dis Aquat Organ. 74:165–170.
- Tang KFJ, Pantoja CR, Redman RM, Lightner DV (2007b) Development of in situ hybridization and RT‒PCR assay for the detection of a nodavirus (PvNV) that causes muscle necrosis in Penaeus vannamei. Dis Aquat Organ. 75:183-190. [CrossRef]
- Tang KFJ, Pantoja CR, Redman RM, Han JE, Tran LH, Lightner DV (2015) Development of in situ hybridization and PCR assays for the detection of Enterocytozoon hepatopenaei (EHP), a microsporidian parasite infecting penaeid shrimp. J Invertebr Pathol. 130:37–41. [CrossRef]
- Tang KFJ, Aranguren LF, Piamsomboon P, Han JE, Maskaykina IY, Schmidt MM (2017) Detection of the microsporidian Enterocytozoon hepatopenaei (EHP) and Taura syndrome virus in Penaeus vannamei cultured in Venezuela. Aquaculture. 480:17–21.
- Tangprasittipap A, Srisala J, Chouwdee S, Somboon M, Chuchird N, Limsuwan C, Srisuvan T, Flegel TW, Sritunyalucksana K (2013) The microsporidian Enterocytozoon hepatopenaei is not the cause of white feces syndrome in white leg shrimp Penaeus (Litopenaeus) vannamei. BMC Vet Res. 9:139. [CrossRef]
- Thompson JR, Randa MA, Marcelino LA, Tomita-Mitchell A, Lim E, Polz MF (2004) Diversity and dynamics of a North Atlantic coastal Vibrio community. Appl Environ Microbiol. 70:4103-4110.
- Tourtip S, Wongtripop S, Stentiford GD, Bateman KS, Sriurairatana S, Chavadej J, Sritunyalucksana K, Withyachumnarnkul B (2009) Enterocytozoon hepatopenaei sp. nov. (Microsporidia: Enterocytozoonidae), a parasite of the black tiger shrimp Penaeus monodon (Decapoda: Penaeidae): fine structure and phylogenetic relationships. J Invertebr Pathol. 102:21-29. [CrossRef]
- Umesha KR, Kennedy B, Dass M, Manja Naik B, Venugopal MN, Karunasagar I, Karunasagar I (2006) High prevalence of dual and triple viral infections in black tiger shrimp ponds in India. Aquaculture. 258:91–96.
- Weiss LM, Vossbrinck CR (1999) Molecular biology, molecular phylogeny, and molecular diagnostic approaches to the microsporidia. In The microsporidia and microsporidiosis (pp. 129-171). ch4. [CrossRef]
- Zhang Q, Xu TT, Wan Liu, Wang LX, Dong X, Yang B, Huang J (2017) Prevalence and distribution of covert mortality nodavirus (CMNV) in cultured crustacean. Virus Research. [CrossRef]
Figure 1.
Comparison of the prevalence between methods for detecting intracellular bacteria (Nunan et al. 2003, Aranguren et al. 2010, Potts et al. 2020) found during a one-year survey. The data were obtained from Oct 22-Apr 23 (Intriago et al. 2024). 1 Nunan et al. (2003) 2 Potts et al. (2020) 3 Aranguren et al. (2010).
Figure 1.
Comparison of the prevalence between methods for detecting intracellular bacteria (Nunan et al. 2003, Aranguren et al. 2010, Potts et al. 2020) found during a one-year survey. The data were obtained from Oct 22-Apr 23 (Intriago et al. 2024). 1 Nunan et al. (2003) 2 Potts et al. (2020) 3 Aranguren et al. (2010).
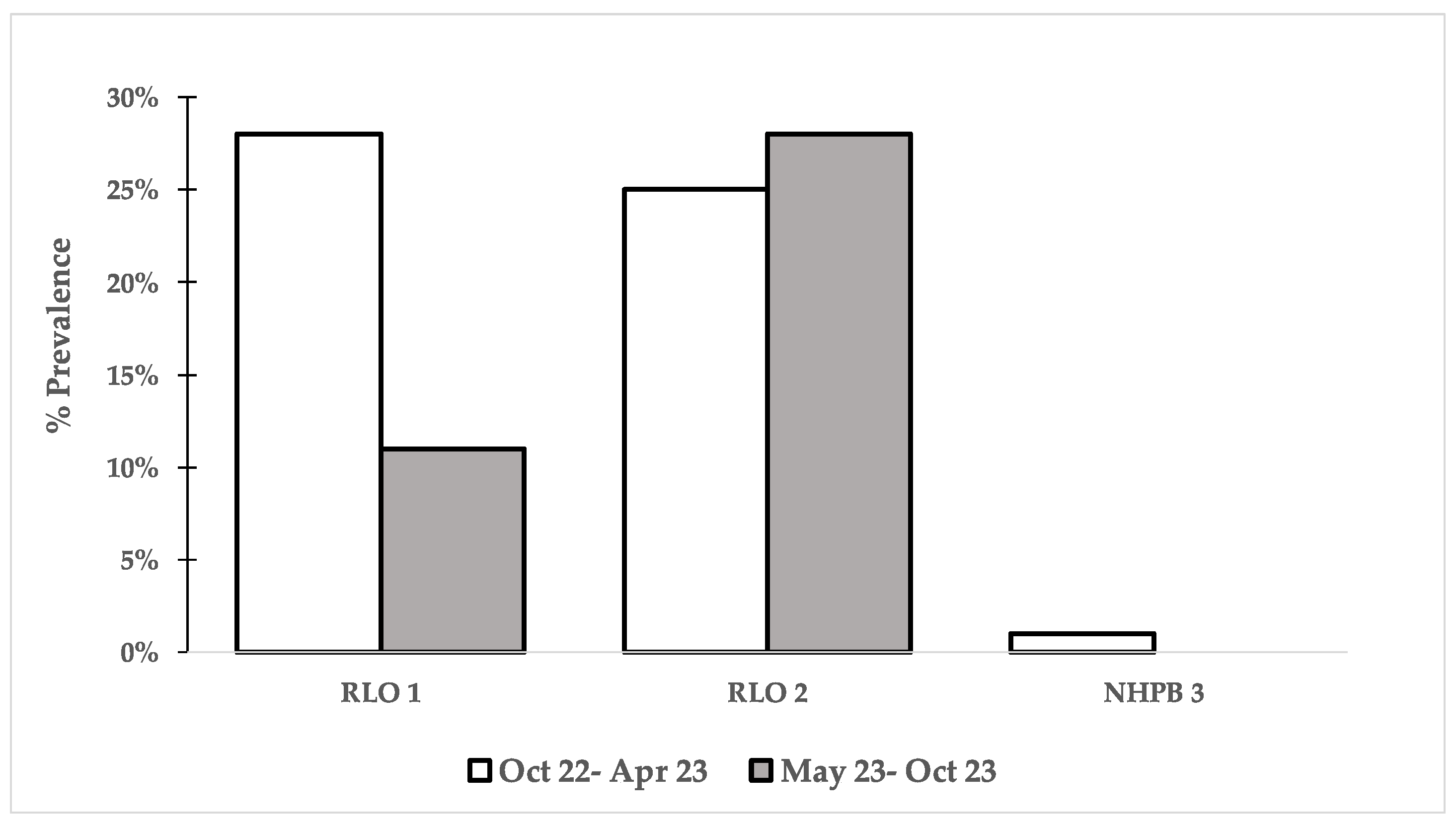
Figure 2.
Comparison of PCR incidence (averaging the 3 regions) between the 6 most important pathogens found during a one-year survey. Data from Oct 22-Apr 23 were obtained from Intriago et al. (2024).
Figure 2.
Comparison of PCR incidence (averaging the 3 regions) between the 6 most important pathogens found during a one-year survey. Data from Oct 22-Apr 23 were obtained from Intriago et al. (2024).
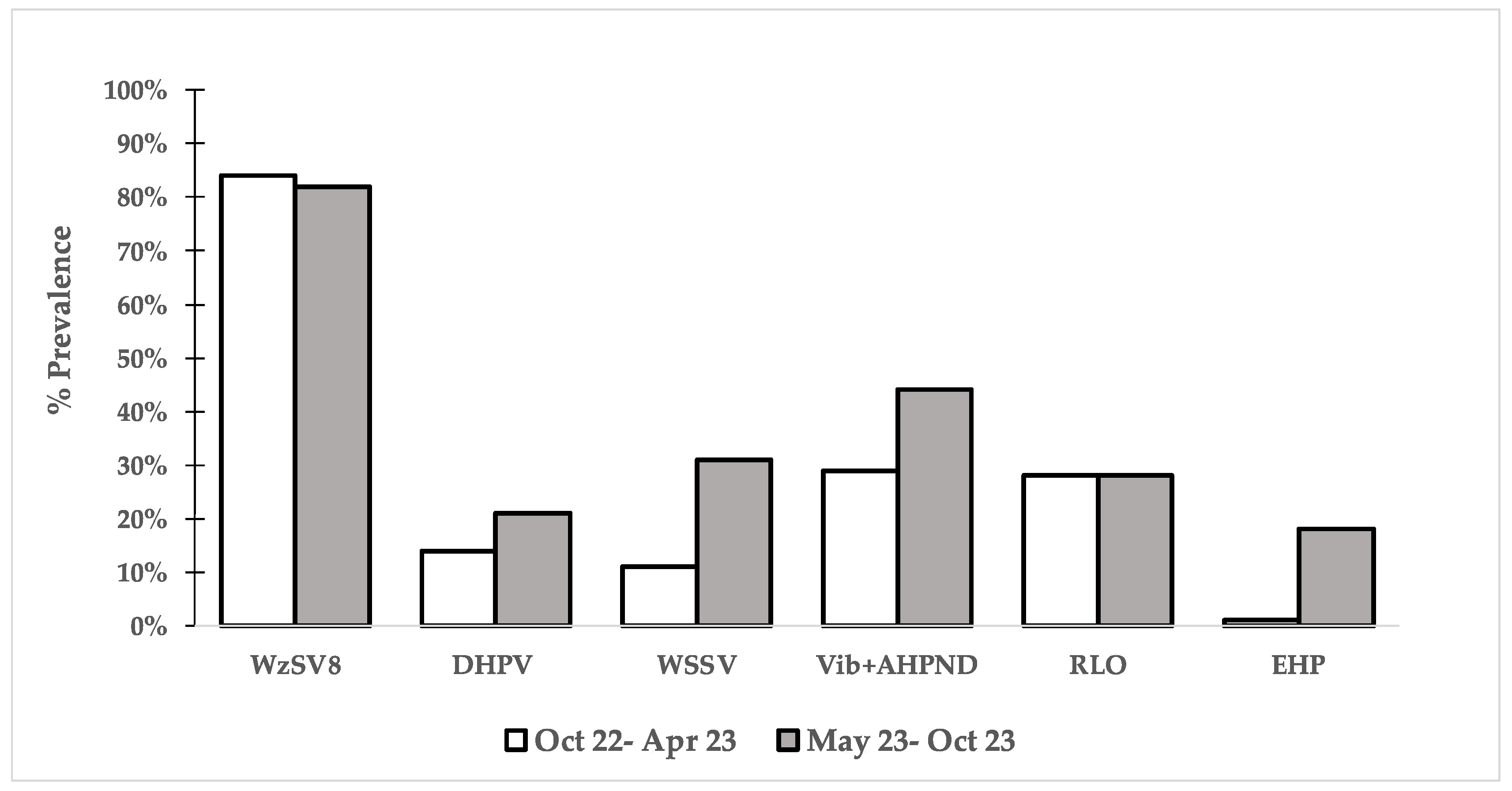
Figure 3.
HPV/DHPV (arrow) inclusion bodies in the hepatopancreatic cells (A, 40X, B, 100X). Anterior caecum cells infected with HPV/DHPV (arrow) (B, 40X). H&E Stain. Anterior caecum cells infected with WzSV8 (arrow) and HPV/DHPV (arrowhead) (C, 40X). Methyl Green - Pyronin modified Stain to distinguish DNA and RNA. Intestine cells infected with HPV/DHPV (arrow) (D, 100X). Methyl Green - Pyronin modified Stain to distinguish DNA and RNA. Ceca infected with WzSV8 (arrow) (E, 100X). Methyl Green - Pyronin modified Stain to distinguish DNA and RNA. Hepatopancreatic cells with inclusion bodies of WZV-8, (DLI) (F, 100X). H&E Stain.
Figure 3.
HPV/DHPV (arrow) inclusion bodies in the hepatopancreatic cells (A, 40X, B, 100X). Anterior caecum cells infected with HPV/DHPV (arrow) (B, 40X). H&E Stain. Anterior caecum cells infected with WzSV8 (arrow) and HPV/DHPV (arrowhead) (C, 40X). Methyl Green - Pyronin modified Stain to distinguish DNA and RNA. Intestine cells infected with HPV/DHPV (arrow) (D, 100X). Methyl Green - Pyronin modified Stain to distinguish DNA and RNA. Ceca infected with WzSV8 (arrow) (E, 100X). Methyl Green - Pyronin modified Stain to distinguish DNA and RNA. Hepatopancreatic cells with inclusion bodies of WZV-8, (DLI) (F, 100X). H&E Stain.
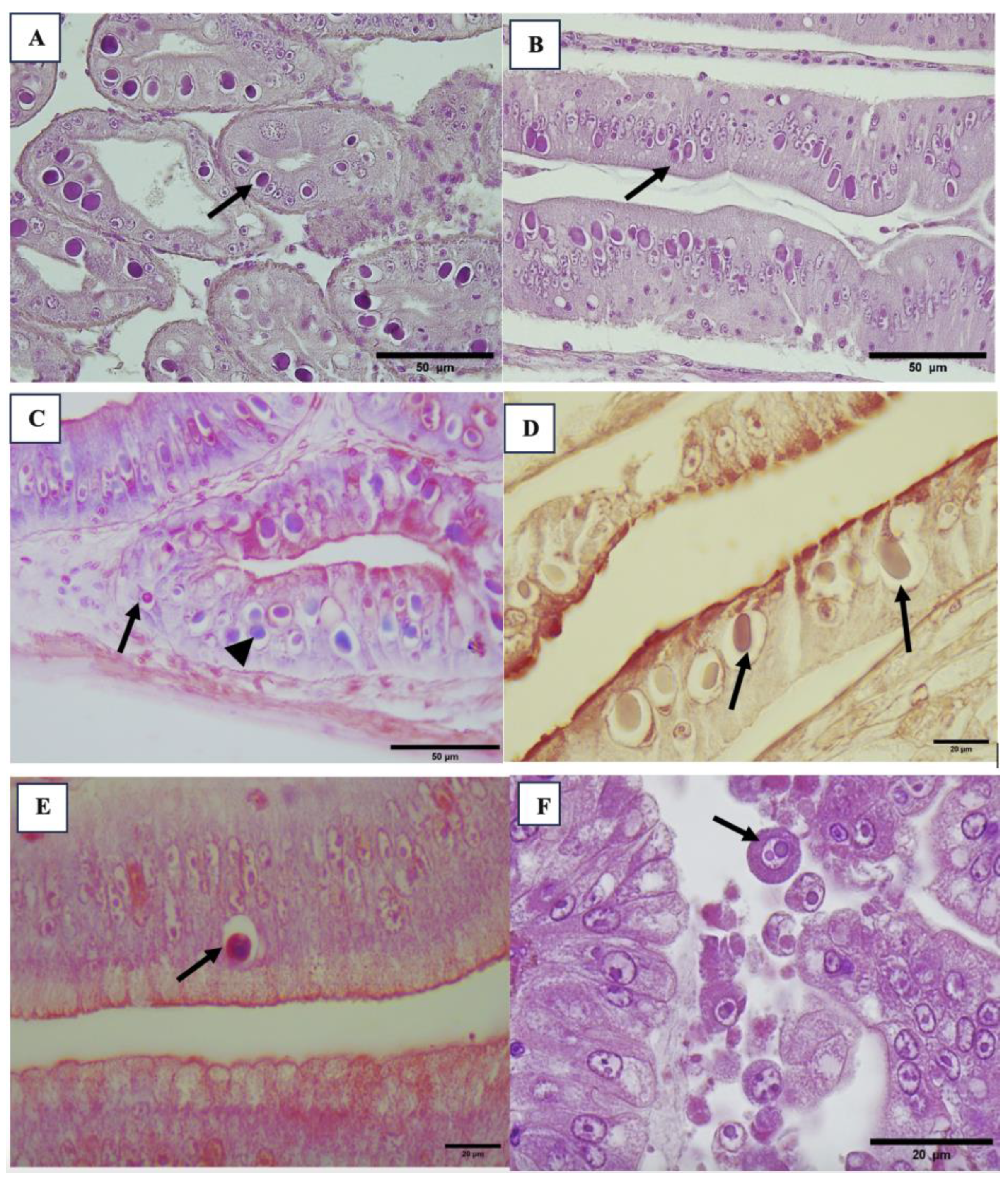
Figure 4.
Histological samples of hepatopancreas with presence of hemocytes, melanized tubules and necrotic tubules (arrow), intracellular bacteria (arrowhead) (A, B, C, D, 40X). Hepatopancreatic cells infected with occlusion body of BP- PvSNPV (arrowhead) and free tetrahedral body in the lumen (arrowhead) (F, 40X). Tetrahedral BP-body of PvSNPV in intestine lumen (M, 10X. N, 40X). H&E Stain.
Figure 4.
Histological samples of hepatopancreas with presence of hemocytes, melanized tubules and necrotic tubules (arrow), intracellular bacteria (arrowhead) (A, B, C, D, 40X). Hepatopancreatic cells infected with occlusion body of BP- PvSNPV (arrowhead) and free tetrahedral body in the lumen (arrowhead) (F, 40X). Tetrahedral BP-body of PvSNPV in intestine lumen (M, 10X. N, 40X). H&E Stain.
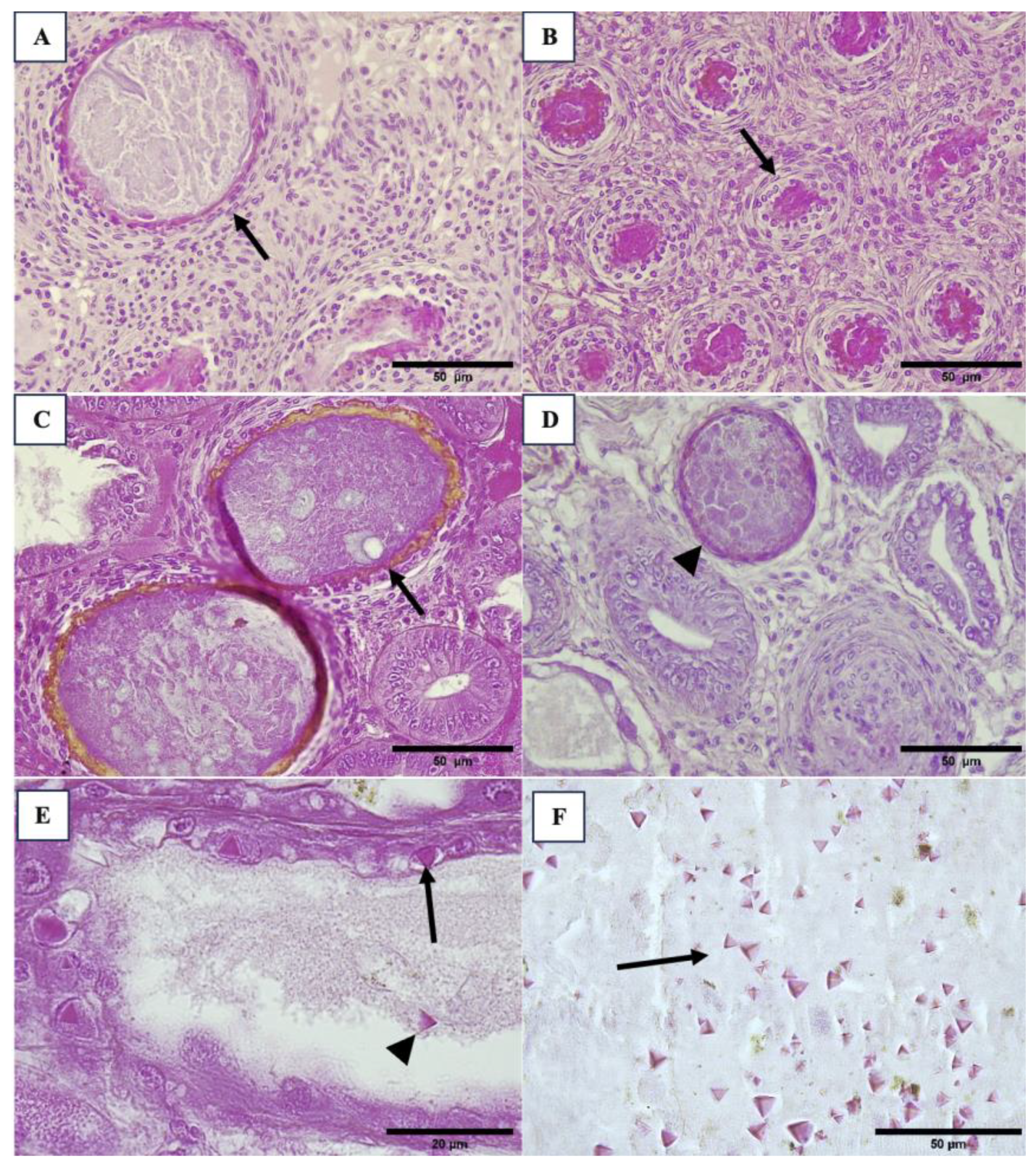
Figure 5.
Cross section views intestinal epithelium with enteritis, observing the inflammation of intestine walls with hemocyte infiltration (arrow) (A, 40X). Melanized reaction in cuticle of exoskeleton (arrow), infiltrated reaction in epidermis (arrowhead) (B, 40X, G, 10X) and muscle necrosis (thick arrow). Muscle necrosis and presence of pack of bacteria with hemocyte infiltration (arrowhead) (D, 100X). H&E Stain.
Figure 5.
Cross section views intestinal epithelium with enteritis, observing the inflammation of intestine walls with hemocyte infiltration (arrow) (A, 40X). Melanized reaction in cuticle of exoskeleton (arrow), infiltrated reaction in epidermis (arrowhead) (B, 40X, G, 10X) and muscle necrosis (thick arrow). Muscle necrosis and presence of pack of bacteria with hemocyte infiltration (arrowhead) (D, 100X). H&E Stain.
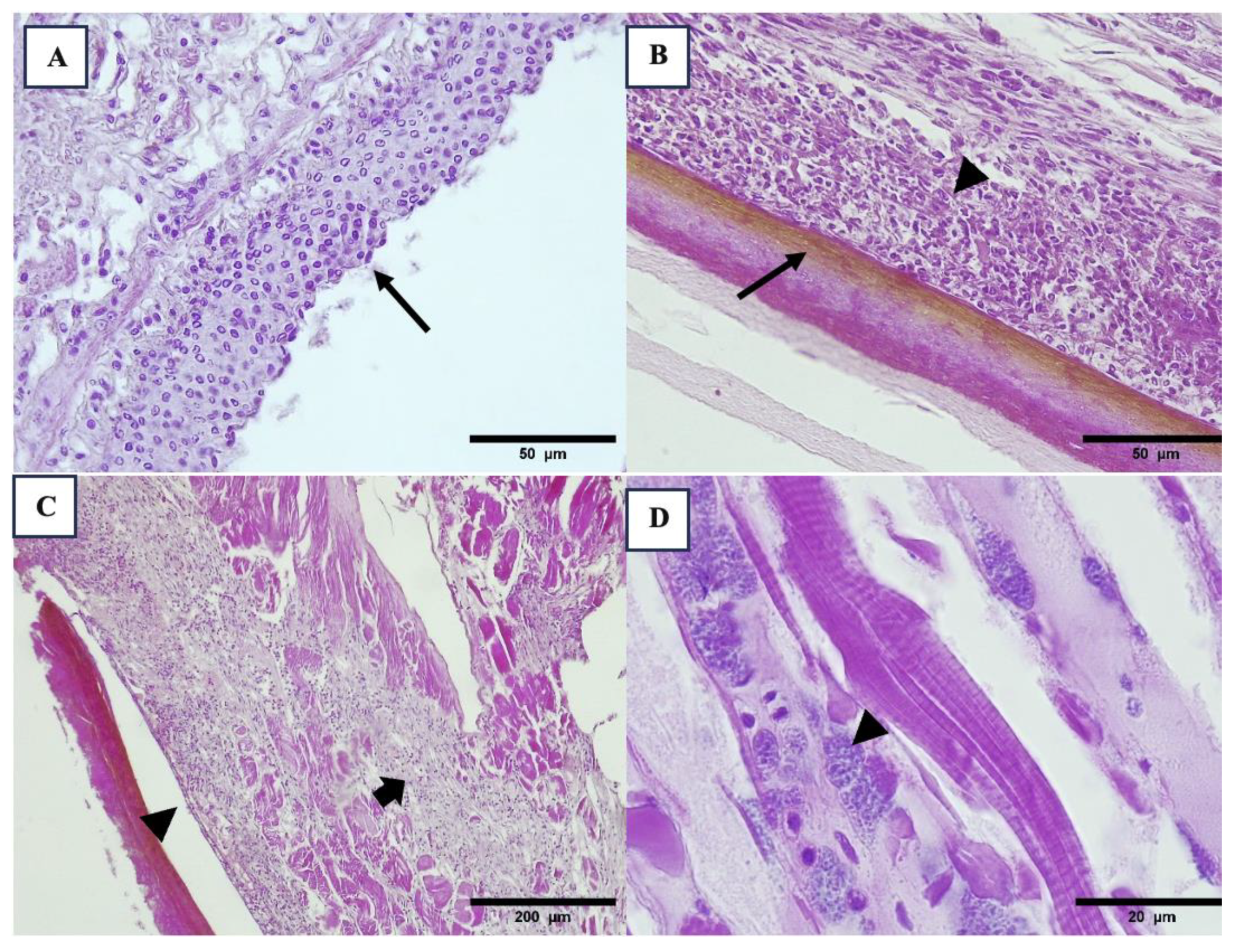
Figure 6.
Presence of LOS in lymphoid organ, observing the different cellular reaction in LOS, pycnotic nuclei (arrow), vacuoles (arrowhead) and RNA-like virus (thick arrow) (A, 40X). Lymphoid organ with melanized reaction (arrow) and LOS (arrowhead) (B, 40X). H&E Stain.
Figure 6.
Presence of LOS in lymphoid organ, observing the different cellular reaction in LOS, pycnotic nuclei (arrow), vacuoles (arrowhead) and RNA-like virus (thick arrow) (A, 40X). Lymphoid organ with melanized reaction (arrow) and LOS (arrowhead) (B, 40X). H&E Stain.
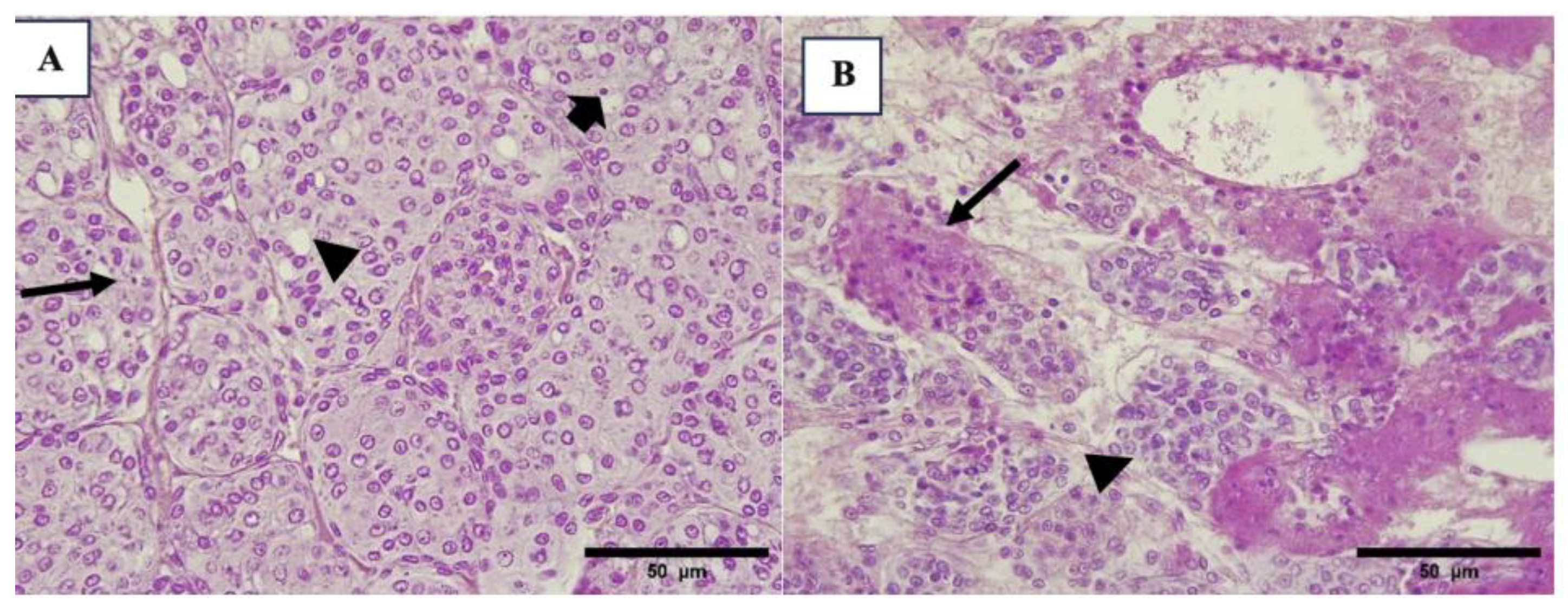
Figure 7.
Hepatopancreas of P. vannamei larvae with sloughing cells and slight hemocyte infiltration (arrow) (A, 10X. B, 40X) and bacterial infection in hepatopancreatic tubules (arrowhead) (C, 10X. D, 40X,). Hepatopancreatic cell and sloughing cells of hepatopancreas with WZV-8 inclusion body (arrow) (K, L, 100X). H&E Stain. .
Figure 7.
Hepatopancreas of P. vannamei larvae with sloughing cells and slight hemocyte infiltration (arrow) (A, 10X. B, 40X) and bacterial infection in hepatopancreatic tubules (arrowhead) (C, 10X. D, 40X,). Hepatopancreatic cell and sloughing cells of hepatopancreas with WZV-8 inclusion body (arrow) (K, L, 100X). H&E Stain. .
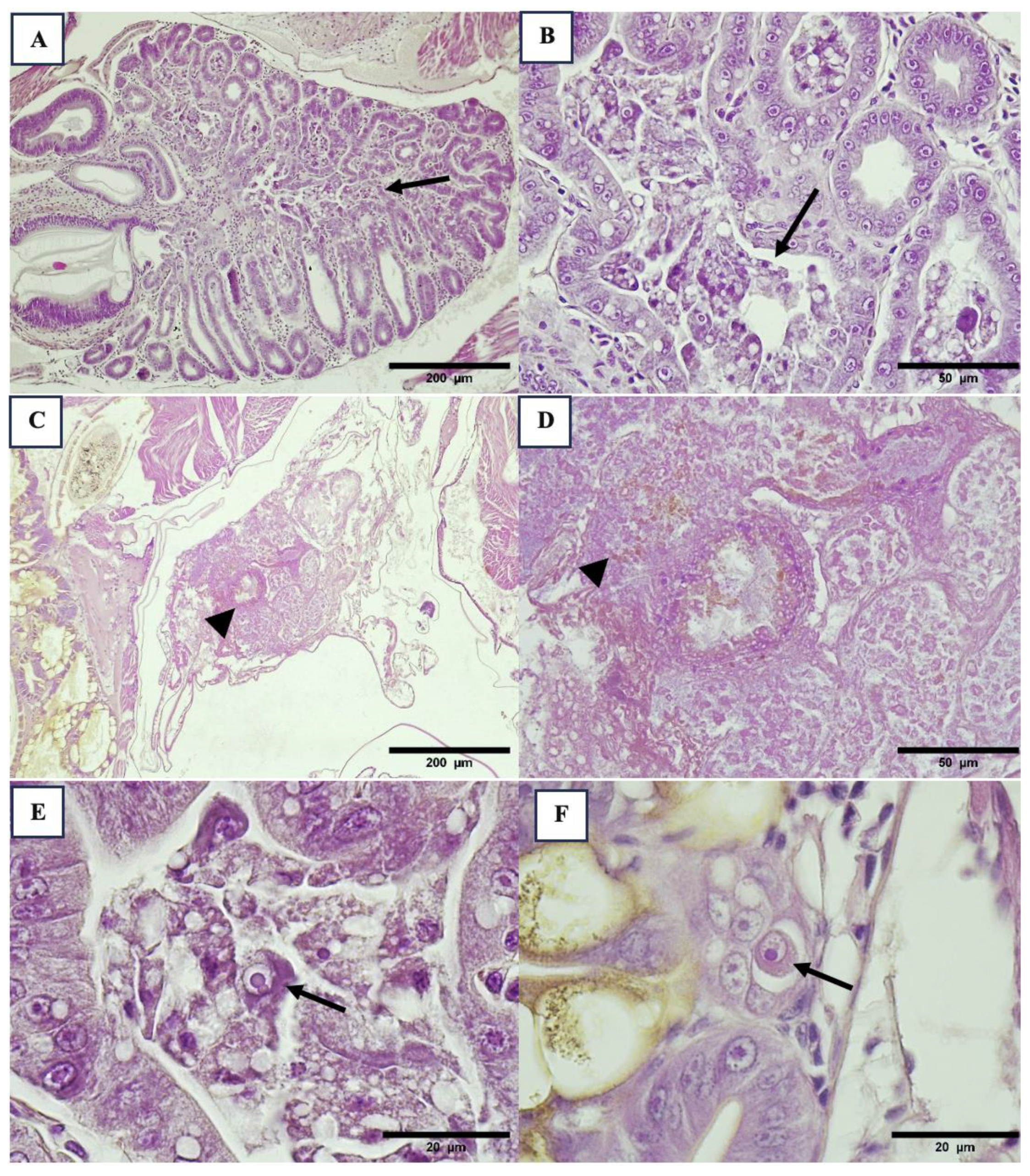
Figure 8.
Histological sections of epithelial of stomach with WSSV (arrowhead) (A, 40X). Histological section of nerve cordon with nervous cell presented inclusion body Cowdry type A typical of IHHNV (B, 40X). H&E Stain.
Figure 8.
Histological sections of epithelial of stomach with WSSV (arrowhead) (A, 40X). Histological section of nerve cordon with nervous cell presented inclusion body Cowdry type A typical of IHHNV (B, 40X). H&E Stain.
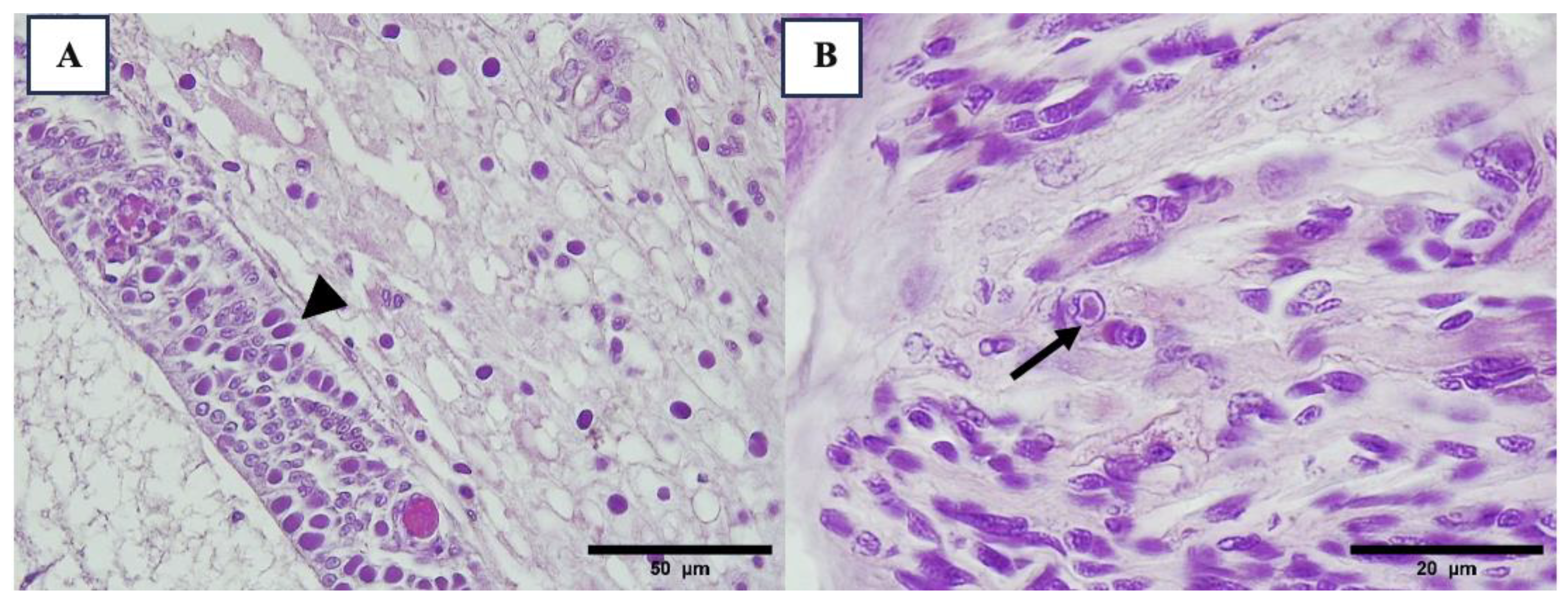
Figure 9.
Infection by microsporidian (arrow) in hepatopancreatic cells (A, 40X). Microsporidia packed was identified in antennal gland (C) and connective tissue of stomach (arrow) (B, C, 100X) both tissues have a slight hemocyte infiltration. Shrimp nephrocomplex with sign of damage in the antennal gland, observing hemocytes infiltration and tubules within melanized reaction and melanized encapsulations (arrow) (D, 40X).H&E Stain.
Figure 9.
Infection by microsporidian (arrow) in hepatopancreatic cells (A, 40X). Microsporidia packed was identified in antennal gland (C) and connective tissue of stomach (arrow) (B, C, 100X) both tissues have a slight hemocyte infiltration. Shrimp nephrocomplex with sign of damage in the antennal gland, observing hemocytes infiltration and tubules within melanized reaction and melanized encapsulations (arrow) (D, 40X).H&E Stain.
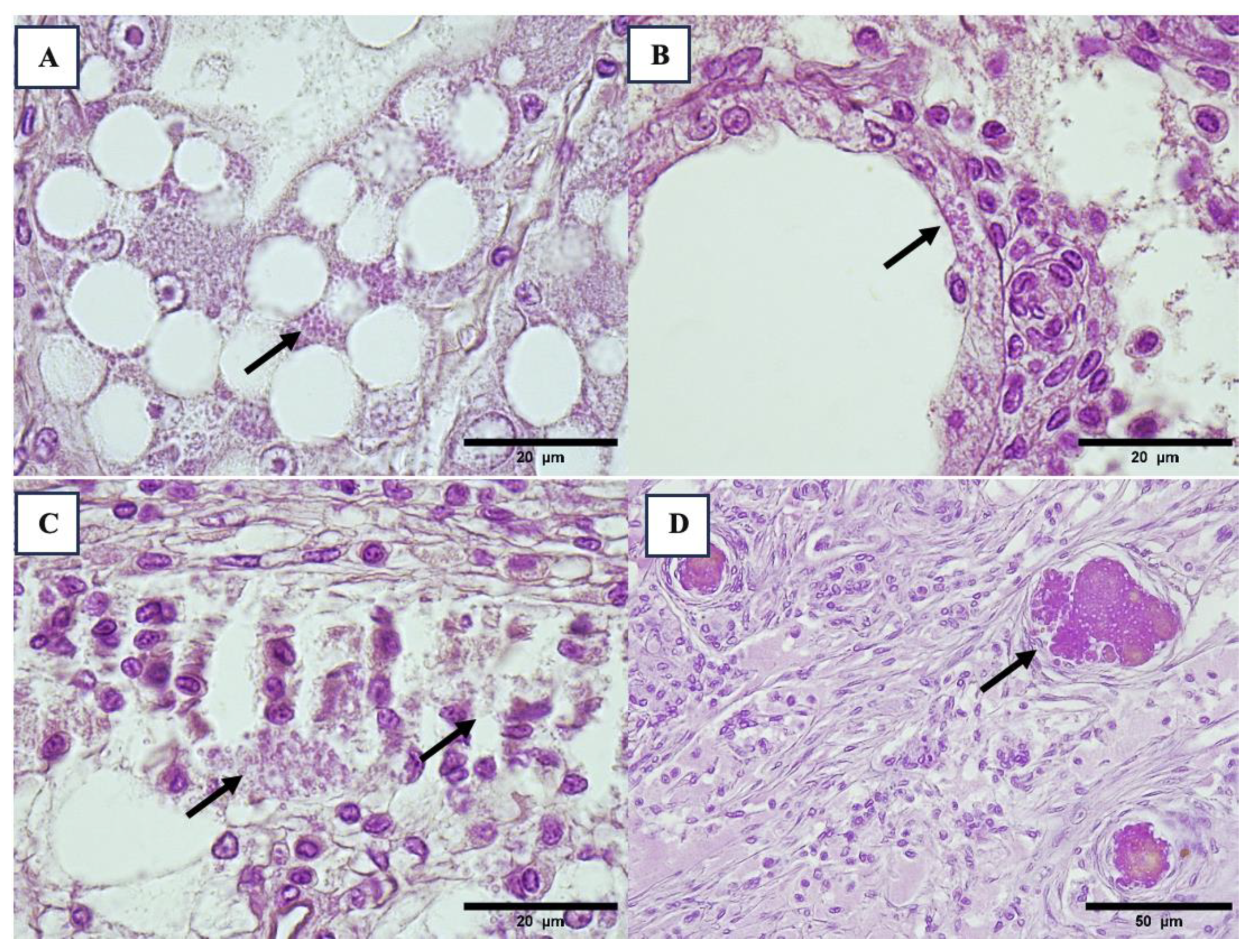
Figure 10.
Histological sections of anterior digestive part with presence of nematodes cyst encysted in connective tissue of stomach or anterior caecum and surrounded by hemocytes (arrow) (A, 10X. B, 40X). Presence of gregarines cyst in tegumental gland of posterior intestine folds (C, 10X). Trophozoite stage of gregarines in intestine lumen (D, 40X). Presence of ciliate protozoans in the gills (with hemocyte infiltration or melanized reaction (arrow) (E, F, 40X).H&E Stain. .
Figure 10.
Histological sections of anterior digestive part with presence of nematodes cyst encysted in connective tissue of stomach or anterior caecum and surrounded by hemocytes (arrow) (A, 10X. B, 40X). Presence of gregarines cyst in tegumental gland of posterior intestine folds (C, 10X). Trophozoite stage of gregarines in intestine lumen (D, 40X). Presence of ciliate protozoans in the gills (with hemocyte infiltration or melanized reaction (arrow) (E, F, 40X).H&E Stain. .
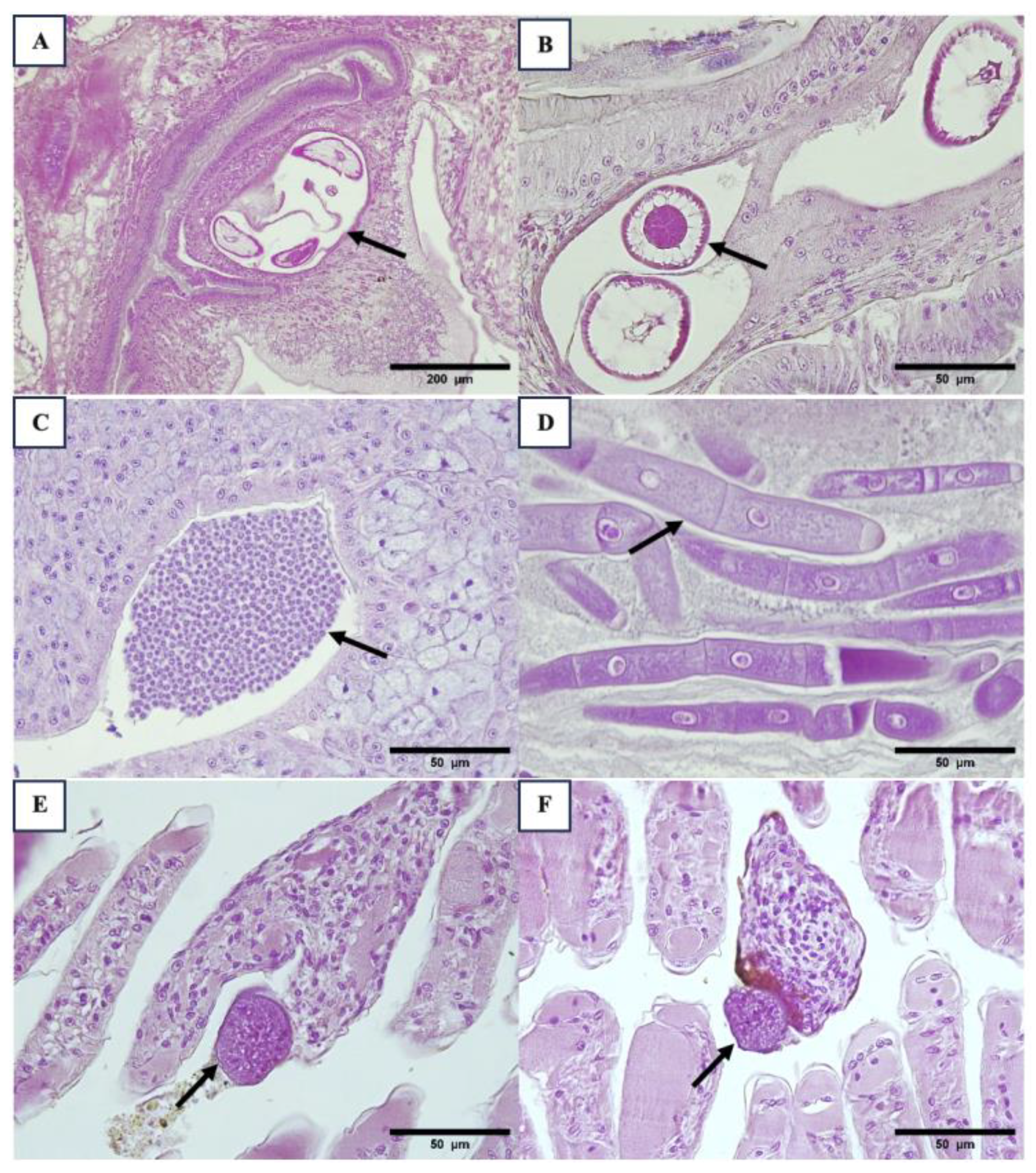

Table 1.
Comparison of prevalence using 3 different sets of primers for WzSV8 in P. vannamei, P. stylirostris and P. monodon in samples from different regions of Latin America.
Table 1.
Comparison of prevalence using 3 different sets of primers for WzSV8 in P. vannamei, P. stylirostris and P. monodon in samples from different regions of Latin America.
| WzSV8 1 504F/R 170F/R 540 bp 170 bp |
WzSV8 2 428F/R 168F/R 428 bp 168 bp |
3136F/3268 R 133 bp3 (PvSV) |
|
|---|---|---|---|
| Region #1 | |||
| PL | 3/5 (60%) | 5/5 (100%) | 4/5 (80%) |
| Farm | 52/58 (90%) | 57/58 (98%) | 55/58 (95%) |
| Broodstock | 23/29 (79%) | 26/29 (90%) | 25/29 (86%) |
| Wild P. vannamei | 3/10 (30%) | 0/10 (0%) | 2/10 (20%) |
| Wild P. stylirostris | 3/8 (38%) | 7/8 (88%) | 8/8 (100%) |
| Region #3 | |||
| PL | 3/3 (100%) | 2/3 (67%) | 2/3 (67%) |
| Farm | 5/8 (63%) | 8/8 (100%) | 8/8 (100%) |
| Broodstock | 0/10 (0%) | 3/10 (30%) | 1/10 (10%) |
| Wild P. monodon | 0/4 (0%) | 1/4 (25%) | 3/4 (75%) |
| Region #4 | |||
| PL | 6/7 (86%) | 7/7 (100%) | 7/7 (100%) |
| Farm | 4/5 (80%) | 5/5 (100%) | 5/5 (100%) |
| Broodstock | 1/1 (100%) | 1/1 (100%) | 1/1 (100%) |
| Average** | 103/148 (70%) | 122/148 (82%) | 121/148 (82%) |
**Average pool of all samples in all regions. 1 Lanza (2022). Personal communication 2 Srisala et al. (2023) 3 Cruz-Flores et al. (2022).
Table 2.
Comparison of sensitivity using 4 different sets of primers for Hepanhamaparvovirus (DHPV) and 1 for Macrobrachium Bidnavirus (MrBdv) in P. vannamei, P. stylirostris and P. monodon in samples from different regions of Latin America.
Table 2.
Comparison of sensitivity using 4 different sets of primers for Hepanhamaparvovirus (DHPV) and 1 for Macrobrachium Bidnavirus (MrBdv) in P. vannamei, P. stylirostris and P. monodon in samples from different regions of Latin America.
| H441F/R 1 HPVnF/R |
H441F/R 2 HPVnF/R |
1120F/R 592 bp 3 |
DHPV-U 1538 F DHPV-U 1887R DHPV-U 1622F 350 bp 256 bp4 |
MrBdv L/R 392 bp 5 |
|
|---|---|---|---|---|---|
| Region #1 | |||||
| PL | 0/5 (0%) | 0/5 (0%) | 0/5 (0%) | 0/5 (0%)) | 0/5 (0%) |
| Farm | 23/58 (40%) | 24/58 (41%) | 4/58 (7%) | 8/58 (14%) | 0/58 (0%) |
| Broodstock | 4/26 (15%) | 4/26 (15%) | 0/26 (0%) | 0/26 (0%) | 0/26 (0%) |
| Wild P. vannamei | 0/10 (0%) | 0/10 (0%) | 0/10 (0%) | 0/10 (0%) | 0/10 (0%) |
| Wild P. stylirostris | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) |
| Region #3 | |||||
| PL | 1/3 (33%) | 0/3 (0%) | 0/3 (0%) | 0/3 (0%) | 0/3 (0%) |
| Farm | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) |
| Broodstock | 0/10 (0%) | 0/10 (0%) | 0/10 (0%) | 0/10 (0%) | 0/10 (0%) |
| Wild P. monodon | 0/4 (0%) | 0/4 (0%) | 0/4 (0%) | 0/4 (0%) | 0/4 (0%) |
| Region #4 | |||||
| PL | 0/7 (0%) | 0/7 (0%) | 0/7 (0%) | 0/7 (0%) | 0/7 (0%) |
| Farm | 2/5 (40%) | 1/5 (20%) | 0/5 (0%) | 0/5 (0%) | 0/5 (0%) |
| Broodstock | 1/1 (100%) | 0/1 (0%) | 0/1 (0%) | 0/1 (0%) | 0/1 (0%) |
| Average** | 31/145 (21%) | 29/145 (20%) | 4/145 (3%) | 8/145 (6%) | 0/145 (0%) |
**Average pool of all samples in all regions. 1 Phromjai et al. (2002) 2 Umesha et al. (2006) 3 Pantoja and Lightner (2000) 4 Srisala et al. (2021) 5 Gangnonngiw et al. (2023).
Table 3.
Comparison of prevalence using 3 different primers for RLB/NHPB in P. vannamei, P. stylirostris and P. monodon in samples from different regions of Latin America.
Table 3.
Comparison of prevalence using 3 different primers for RLB/NHPB in P. vannamei, P. stylirostris and P. monodon in samples from different regions of Latin America.
| Bact F/R 1500 bp Rik F/R532 bp 1 |
Rp877p Rp1258 n 382 bp2 |
NHPF2 NHPR2 379 bp3 |
|
|---|---|---|---|
| Region #1 | |||
| PL | 0/5 (0%) | 1/5 (20%) | 0/5 (0%) |
| Farm | 1/58 (2%) | 17/58 (29%) | 0/66 (0%) |
| Broodstock | 17/41 (41%) | 21/41 (51%) | 0/26 (0%) |
| Wild P. vannamei | 0/10 (0%) | 0/10 (0%) | 0/10 (0%) |
| Wild P. stylirostris | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) |
| Region #3 | |||
| PL | 0/3 (0%) | 0/3 (0%) | 0/3 (0%) |
| Farm | 0/8 (0%) | 5/8 (63%) | 0/8 (0%) |
| Broodstock | 0/10 (0%) | 0/10 (0%) | 0/10 (0%) |
| Wild P. monodon | 0/4 (0%) | 0/4 (0%) | 0/4 (0%) |
| Region #4 | |||
| PL | 0/7 (0%) | 1/7 (0%) | 0/7 (0%) |
| Farm | 0/5 (0%) | 0/5 (0%) | 0/5 (0%) |
| Broodstock | 0/1 (0%) | 0/1 (0%) | 0/1 (0%) |
| Average** | 18/160 (11%) | 45/160 (28%) | 0/153 (0%) |
**Average pool of all samples in all regions. 1 Nunan et al. (2003) 2 Potts et al. (2020) 3 Aranguren et al. (2010).
Table 4.
Comparison of sensitivity using 3 different sets of primers for Spiroplasma in P. vannamei, P. stylirostris and P. monodon in samples from different regions of Latin America.
Table 4.
Comparison of sensitivity using 3 different sets of primers for Spiroplasma in P. vannamei, P. stylirostris and P. monodon in samples from different regions of Latin America.
| CSF/R 269 bp1 | F28/R5 271 bp2 | Pri-1/2 1200 bp3 | |
|---|---|---|---|
| Region #1 | |||
| PL | 1/5 (20%) | 0/5 (0%) | 0/5 (0%) |
| Farm | 28/62 (45%) | 1/62 (2%) | 6/62 (10%) |
| Broodstock | 0/26 (0%) | 2/26 (8%) | 0/26 (0%) |
| Wild P. vannamei | 0/10 (0%) | 0/10 (0%) | 0/10 (0%) |
| Wild P. stylirostris | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) |
| Region #3 | |||
| PL | 0/3 (0%) | 0/3 (0%) | 0/3 (0%) |
| Farm | 1/8 (13%) | 0/8 (0%) | 8/8 (100%) |
| Broodstock | 0/10 (0%) | 0/10 (0%) | 0/10 (0%) |
| Wild P. monodon | 0/4 (0%) | 0/4 (0%) | 0/4 (0%) |
| Region #4 | |||
| PL | 0/7 (0%) | 0/7 (0%) | 0/7 (0%) |
| Farm | 0/5 (0%) | 0/5 (0%) | 0/5 (0%) |
| Broodstock | 0/1 (0%) | 0/1 (0%) | 0/1 (0%) |
| Average** | 30/149 (20%) | 3/149 (2%) | 14/149 (9%) |
**Average pool of all samples in all regions. 1 Nunan et al. (2004) 2 Bastian et al. (2004) 3 Ding et al. (2007).
Table 5.
Comparison of the prevalence of Microsporidia in P. vannamei, P. stylirostris and P. monodon using 3 different primers in samples from different regions of Latin America.
Table 5.
Comparison of the prevalence of Microsporidia in P. vannamei, P. stylirostris and P. monodon using 3 different primers in samples from different regions of Latin America.
| TS1/TS2 790 bp 1 |
18f/1492r 1200 bp 2 |
Ss2 18f3/ ss1492r 3 |
|
|---|---|---|---|
| Region #1 | |||
| PL | 3/5 (60%) | 0/5 (0%) | 0/5 (0%) |
| Farm | 46/57 (81%) | 16/57 (28%) | 31/57 (54%) |
| Broodstock | 20/26 (77%) | 6/26 (23%) | 8/26 (31%) |
| Wild P. vannamei | 9/10 (90%) | 2/10 (20%) | 2/10 (20%) |
| Wild P. stylirostris | 7/8 (88%) | 0/8 (0%) | 6/8 (75%) |
| Region #3 | |||
| PL | 1/3 (33%) | 0/3 (0%) | 0/3 (0%) |
| Farm | 8/8 (100%) | 2/8 (25%) | 4/8 (50%) |
| Broodstock | 8/10 (80%) | 0/10 (0%) | 6/10 (60%) |
| Wild P. monodon | 3/4 (75%) | 0/4 (0%) | 3/4 (75%) |
| Region #4 | |||
| PL | 1/7 (14%) | 0/7 (0%) | 1/7 (14%) |
| Farm | 5/5 (100%) | 4/5 (80%) | 5/5 (100%) |
| Broodstock | 1/1 (100%) | 0/1 (0%) | 1/1 (100%) |
| Average** | 112/144 (85%) | 30/144 (21%) | 67/144 (47%) |
**Average pool of all samples in all regions. 1 Pasharawipas and Flegel (1994) 2 Sokolova et al. (2015) 3 Weiss and Vossbrinck (1999).
Table 6.
Comparison of prevalence using 4 different primers for IHHNV in P. vannamei, P. stylirostris and P. monodon in samples from different regions of Latin America.
Table 6.
Comparison of prevalence using 4 different primers for IHHNV in P. vannamei, P. stylirostris and P. monodon in samples from different regions of Latin America.
| 309 F/R 309 bp 1 |
392 F/R 392 bp 2 |
389 F/R 389 bp 3 |
77012F/77353R (356 bp) 4 |
IHHNV*** | eve*** | |
|---|---|---|---|---|---|---|
| Region #1 | ||||||
| PL | 3/5 (60%) | 3/5 (60%) | 3/5 (60%) | 0/5 (0%) | 0/5 (0%) | 3/5 (60%) |
| Farm | 44/58 (76%) | 38/58 (66%) | 37/58 (64%) | 26/58 (45%) | 26/58 (45%) | 19/58 (33%) |
| Broodstock | 6/26 (23%) | 8/26 (31%) | 6/26 (21%) | 4/26 (15%) | 4/26 (15%) | 4/26 (15%) |
| Wild P. vannamei | 8/10 (80%) | 7/10 (70%) | 5/10 (50%) | 3/10 (30%) | 3/10 (30%) | 5/10 (50%) |
| Wild P. stylirostris | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) |
| Region #3 | ||||||
| PL | 0/3 (0%) | 0/3 (0%) | 1/3 (33%) | 0/3 (0%) | 0/3 (0%) | 1/3 (33%) |
| Farm | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) |
| Broodstock | 0/10 (0%) | 0/10 (0%) | 0/10 (0%) | 0/10 (0%) | 0/10 (0%) | 0/10 (0%) |
| Wild P. monodon | 0/4 (0%) | 1/4 (25%) | 2/4 (50%) | 1/4 (25%) | 0/4 (0%) | 2/4 (50%) |
| Region #4 | ||||||
| PL | 3/7 (43%) | 0/7 (0%) | 1/7 (14%) | 0/7 (0%) | 0/7 (0%) | 3/7 (43%) |
| Farm | 1/5 (20%) | 1/5 (20%) | 1/5 (20%) | 1/5 (20%) | 1/5 (20%) | 0/5 (0%) |
| Broodstock | 0/1 (0%) | 0/1 (0%) | 0/1 (0%) | 0/1 (0%) | 0/1 (0%) | 0/1 (0%) |
| Average** | 65/145 (45%) | 58/145 (40%) | 56/145 (39%) | 35/145 (24%) | 34/145 (23%) | 37/145 (26%) |
*All wild P. monodon were positive MG831F/R primers [19,21] indicating the presence of eve. **Average pool of all samples in all regions. *** When tested using multiple PCR primers, all primers must be present for a functional virus to replicate. However, if some but not all primers amplify, this pattern is a clear indication that the PCR primers are measuring an eve (endogenous viral element) and not the complete virus. 1 Tang et al. (2007a) 2 Tang et al. (2000) 3 Tang and Lightner (2006) 4 [22] Nunan et al. (2000).
Table 7.
Comparison of prevalence using 6 different primers for EHP in P. vannamei, P. stylirostris and P. monodon in samples from different regions of Latin America.
Table 7.
Comparison of prevalence using 6 different primers for EHP in P. vannamei, P. stylirostris and P. monodon in samples from different regions of Latin America.
| EHP 510 bp 1 | EHP900-1000 bp 2 | VE-EHP-SWP-365 bp 3 |
EHP B-tubulin 262 bp 3 |
SSU ENF779 F1 SSU ENF779 R1 SSU ENF176 F1 SSU ENF176 R1 779 bp 176 bp 4 |
SWP_1F SWP_1R SWP_2F SWP_2R 514 bp 148 bp 5 |
|
|---|---|---|---|---|---|---|
| Region #1 | ||||||
| PL | 0/5 (0%) | 0/5 (0%) | 0/5 (0%) | 0/5 (0%) | 0/5 (0%) | 0/5 (0%) |
| Farm | 20/72 (28%) | 10/72 (14%) | 1/72 (1%) | 3/72 (4%) | 30/72 (42%) | 19/72 (26%) |
| Broodstock | 8/29 (28%) | 1/29 (3%) | 0/29 (0%) | 5/29 (17%) | 8/29 (28%) | 10/29 (34%) |
| Wild P. vannamei | 0/10 (0%) | 0/10 (0%) | 0/10 (0%) | 0/10 (0%) | 0/10 (0%) | 0/10 (0%) |
| Wild P. stylirostris | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) | 1/8 (13%) | 0/8 (0%) |
| Region #3 | ||||||
| PL | 0/3 (0%) | 0/3 (0%) | 0/3 (0%) | 0/3 (0%) | 0/3 (0%) | 0/3 (0%) |
| Farm | 0/8 (0%) | 5/8 (0%) | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) |
| Broodstock | 0/10 (0%) | 0/10 (0%) | 0/10 (0%) | 0/10 (0%) | 0/10 (0%) | 0/10 (0%) |
| Wild P. monodon | 0/4 (0%) | 0/4 (0%) | 0/4 (0%) | 0/4 (0%) | 0/4 (0%) | 0/4 (0%) |
| Region #4 | ||||||
| PL | 0/7 (0%) | 0/7 (0%) | 0/7 (0%) | 0/7 (0%) | 1/7 (14%) | 0/7 (0%) |
| Farm | 0/5 (0%) | 0/5 (0%) | 0/5 (0%) | 0/5 (0%) | 5/5 (100%) | 0/5 (0%) |
| Broodstock | 0/1 (0%) | 0/1 (0%) | 0/1 (0%) | 0/1 (0%) | 1/1 (100%) | 0/1 (0%) |
| Average** | 28/162 (17%) | 16/162 (10%) | 1/162 (1%) | 8/162 (5%) | 46/162 (28%) | 29/162 (18%) |
**Average pool of all samples in all regions. 1 Tang et al. (2015) 2 Tourtip et al. (2009) 3 Tang et al. (2017) 4 Tangprasittipap et al. (2013) 5 Jaroenlak et al. (2016).
Table 8.
Histological average of major lesions in the 3 regions.
 |
 |
1. The average hepatopancreas abnormalities (cell sloughing, hemocytic – melanized and necrotic tubules) were recorded. atrophied/destroyed tubules, hemocytic enteritis) 2. Intestinal epithelium with enteritis: observing the inflammation of intestinal walls with hemocyte infiltration. 3. Muscle necrosis, the presence of bacterial packs, and hemocyte infiltration. 4. Lymphoid organ spheroids or any pathology. 5. White spot syndrome virus lesions. 6. Hepanhamaparvovirus (DHPV) VIN. 7. Wenzhou shrimp virus 8 (WzSV8) VIN. 8. Infection by microsporidians in the hepatopancreas, antennal gland, and connective tissue of the stomach. 9. Sum of gregarines and nematodes in the intestine, hepatopancreas, etc. 10. Average weight (g). 11. Total number of animals analyzed. 12. Percentage of prevalence. 13. The average severity was adopted from Lightner (1996) and simplified as follows: 0= no lesions, 1=lesions or infection present in <25% of area or organ or tissue sections, 2=lesions or infection present in 25-50% of area or organ or tissue sections, 3=lesions or infection present in 50-75% of area or organ or tissue sections, and 4=lesions or infection present in >75% of area or organ or tissue sections [42]. 14. ND Not determined.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



