
Submitted:
10 December 2024
Posted:
11 December 2024
You are already at the latest version
Abstract
The seasonal and interannual temporal variation of the structure, richness, diversity and similarity of fish larval assemblages associated to an Eastern Tropical Pacific coral reef system, was assessed for the period 2017 to 2019. A total of 4779 fish larvae were captured, of which 4556 (95% of the total) were identified, belonging to 88 species and 46 families, and increasing the number of larvae of fish species reported to 162 taxa. Based on the analysis of similarity between oceanographic seasons and between years, six significant groups were established (SIMPROF, p < 0.05), each group made up of the nine sampling stations belonging to each study period (ANOSIM, R = 0.764). C. mysticetus, D. pacificus, Anchoa sp., Anisotremus sp., B. bathymaster, O. saurus, Caranx sp., Seriola sp., Gobiidae sp., Microgobius sp., and S. evermanni were the species that contributed to the differentiation of the groups. Species richness distribution during the sampling periods was variable and did not show a clear temporal or annual trend. However, the fish larvae assamblages were ordered according with the cold and warm seasons, suggesting that intra-annual variation has a strong influence on the temporal dynamics of fish larval assemblages in ETP coral reef systems.
Keywords:
Eastern Tropical Pacific
; Coral Reef
; community structure
; diversity
; Marine Ecology
; Colombia
1. Introduction
The Eastern Tropical Pacific (ETP) has unique environmental characteristics for marine life. Specifically, the abiotic conditions in the region were modulated by the combined effects of seasonal and regional fluctuations in local environmental and oceanographic factors across a range of spatial and temporal scales [1,2,3] The Colombian Pacific Ocean (CPO) is located in the easternmost sector of the ETP, and there are different types of marine-coastal ecosystems, including rocky shores, sandy beaches, estuaries, mangrove forests, and coral reefs, which support a wide variety of fish with a wide range of life history strategies [4,5].
Coral reefs are highly biodiverse ecosystems and host the richest communities of fish species on the planet [6,7,8]. In general, fish have a life cycle with two different phases: the first pelagic larval phase, which allows them to disperse, and the second demersal or pelagic adult phase [9,10]. Fish larvae are a phase of great interest in marine ecology research because of the replenishment of populations, the connectivity or exchange of individuals between them, the understanding of fish dispersal patterns, and in general, a wide range of ecological processes of these populations depend largely on the understanding of the dynamics of the larval stages [11,12]. Therefore, studies on variations in the abundance and structure of fish larval assemblages are necessary to understand the dynamics of the fish populations present in coral reefs.
In coastal marine environments, variations in environmental conditions at different time scales can modulate the attributes of fish larval assemblages [13], such that during an annual cycle, which incorporates different hydrographic conditions, changes in their composition and abundance have been reported [14,15,16,17,18,19]. In general, the spatiotemporal variation of physical factors generally contributes the most to the biological responses of marine communities. For example, fluctuations in circulation, water column stability, turbidity, salinity, oxygen, and temperature can determine the viability of a particular species and consequently modulate the composition of a set or assemblage [20,21,22,23].
To date, research efforts on reef fishes in the ETP have focused on both the adult and larval stages [15,24,25,26,27,28,29,30]; however, in the CPO, efforts have focused mainly on adults [31,32,33,34,35,36,37]. In this region of the ETP, only two studies have analyzed larval fish assemblages, the first of which was carried out by Calle-Bonilla et al. [38], who evaluated the role of surface circulation in the movement of fish larvae associated with coral reef environments on Gorgona Island during two contrasting oceanographic periods, and the other one carried out by Ramírez et al. [39], who evaluated the temporal variability of the assembly of fish larvae associated with a coral reef formation on Gorgona Island on a daily (day-night) and monthly scale.
In this study, we assessed the response of ichthyoplankton assemblages associated with well-developed coral reef formation into intra-annual and inter-annual variability of hydrographic conditions. Specifically, we addressed the following questions: (1) Does ichthyoplankton abundance and community structure differ among the intra-annual contrasting oceanographic periods described for this region? (2) Is the structure of the fish larval assemblage similar on an inter-annual scale? (3) What are the main environmental factors affecting the abundance and community structure of fish larvae in this coral reef formation?.
2. Materials and Methods
The Colombian Pacific Ocean is an integral part of the eastern region of the ETP. In this region, the most developed coral reefs are located on the eastern side of Gorgona Island (2°58'N - 78°11'W) (Figure 1), on the continental shelf of Colombia's southern Pacific region, 36 km from the mainland [40]. This area, declared National Natural Park since 1984, has an average air temperature of 27°C (varying between 24°C and 30°C) and annual rainfall ranges between 4000 mm and 8000 mm distributed in two rainy periods, the most intense between April and November, and a second less intense period between December and March [41]. Two contrasting oceanographic periods have been described for this locality, the first from May to December with low surface salinity values and a deep thermocline (c.a. 47 m) and the second from January to April with high surface salinity and a shallow thermocline (c.a. 7.5 m), with a semi-diurnal tide that can reach a maximum record of 5.7 m [42].
Gorgona Island has a great variety of coastal habitats [43], among which the coral formations of La Azufrada, Playa Blanca, El Muelle, La Ventana, and Farallones stand out as habitats for diverse marine fauna [21,26,44,45,46]. All these coral formations have a low impact from anthropogenic activities, and the La Azufrada coral reef is considered one of the largest and well-developed coral reefs in the ETP [46]. The coral reef is 780 m long and 80-180 m wide, and it is formed mainly by coral species of the Pocilloporidae family, although there are also coral species from Psammocoridae, Agariciidae and Poritidae families represented by genera such as Psammocora, Pavona, Porites, and Gardineroseris [45,46]
Since 2007, the pelagic environment of the La Azufrada coral reef has been monitored twice a year as part of the protected area's oceanographic monitoring program, which consists of a sampling grid of nine equidistant stations over the reef (Figure 1). For this study, only samples collected during March and September 2017, 2018, and 2019 were used during the cold and warm oceanographic periods, which have been described for this location of ETP [38,42,47,48,49,50].
Discrete water samples were collected at a depth of 1 m using a Niskin bottle, and temperature, salinity, chlorophyll-a, and dissolved oxygen were recorded using a YSIpro® multiparameter probe and water column transparency using a Secchi disk. Fish larvae were obtained by day and night surface trawls of 5 min duration, using a bongo net of 30 cm diameter and 180 cm length, equipped with 300 and 500 μm pore meshes, with Hydrobios® flowmeters calibrated to quantify the volume of filtered water. Samples were fixed in a formalin solution (formalin-seawater) at a final concentration of 4% for transport to the Oceanographic Sciences Laboratory of the Universidad del Valle. All samples were reviewed to separate fish larvae from each zooplankton sample, and taxonomic identification was carried out to the closest possible species level, following the guidelines [51,52]. The identified species were deposited in the Zoology Teaching Collection of the Universidad del Valle.
To quantify the zooplankton biomass, half of the sample collected with the 300 μm mesh was concentrated on cellulose filters that were previously dried and weighed. The samples were dried at 60°C for 24 h and weighed on an analytical balance with a precision of 0.0001 g, determining the dry biomass by weight difference.
To reduce the effect of day and night variation in the structuring of the data matrix of fish larvae in the water column of the La Azufrada coral reef, the day and night records made during each sampling campaign were taken together. In addition, the information obtained with the 300 μm and 500 μm nets was consolidated, assuming that they were complementary sources of taxonomic information [53].
Temperature, salinity, dissolved oxygen, chlorophyll-a, zooplankton biomass, and fish larval abundance were compared between periods and years of study (summing the cold and warm periods for each year) using a non-parametric Kruskall Wallis test (KW) with Bonferroni correction and a post-hoc Tukey rank test. The representativeness of the sampling effort was established from the construction of rarefaction curves for the study period and the expected richness was defined from Clench's non-linear estimator, which predicts that the probability of finding a new species will increase (up to a maximum) as the sampling effort increases, that is, the probability of adding new species eventually decreases, but field work increases it [54]. The community attribute indices Shannon and Simpson were used to describe the diversity of the assemblages, and a comparison was made between periods by resampling 1000 replicates using the bootstrap technique.
The structure of the assemblage was described based on the Bray Curtis similarity index, defined from the abundance matrix of species with a frequency of occurrence equal to or greater than 5%. Transformation was performed using the log (x+1) function to reduce the abundance effect of dominant species. The average clustering algorithm was used to construct a similarity dendrogram, and the similarity profile routine (SIMPROF) was used to establish the significance of the clusters. Moreover, the contribution of species within the clusters generated in the similarity analysis was established using the similarity percentages (SIMPER) routine in the PRIMER v6.0® software. Finally, to determine whether there were differences in fish larval assemblage structure between study periods, a one-way abundance matrix similarity analysis (ANOSIM) was used to evaluate the differences in the structure of fish larval assemblages between study periods (totally different assemblages when R values are between 0.75 and 1) [55,56].
Finally, the relationship between the established assemblage structure and local environmental conditions (temperature, salinity, chlorophyll-a, and dissolved oxygen) was explored using a canonical correspondence analysis (CCA) carried out with the free ecological software Past® v3.12. CCA is a multivariate association analysis that maximizes the degree of correlation between fish larval abundance and physicochemical parameters.
3. Results
3.1. Fish Larval Assemblage Composition and Structure
A total of 4779 fish larvae were captured, of which 4556 (95% of the total), belonging to 88 species and 46 families, were identified during the six study periods. In our study, 41 species were first recorded in the La Azufrada coral reef, increasing the number of larvae of fish species associated with the coral formations of this locality to 162 taxa (Table 1, Table A1).
3.2. Ambient Conditions
Mean sea surface temperature ranged between 28.4 ± 0.13 °C (March 2017), 28.8 ± 0.13 °C (September 2017), 27.4 ± 0.24 °C (March 2018), 27.7 ± 0.09 °C (September 2018), 25.8 ± 0.29 °C (March 2019) and 27.5 ± 0.13 °C (September 2019), no significant differences were found for SST between oceanographic periods (KW, P>0.05). Mean sea surface salinity ranged from 29.6 ± 0.03 (March 2017), 25.2 ± 0.45 (September 2017), 26.7 ± 0.18 (March 2018), 26.1 ± 1.14 (September 2018), 30.7 ± 0.44 (March 2019) and 30.3 ± 0.13 (September 2019), there were significant differences between March and September 2017 (KW, P>0.05). The maximum temperature and salinity values were recorded in September 2017 and March 2019, respectively (Table 4).
3.3. Relationship Between Fish Larval Assemblages and Environmental Variables
The first two ordination axes of the canonical correspondence analysis (CCA) explained 67% of the total variance of the fish larval assemblage present in the water column associated with the La Azufrada coral reef. Salinity, chlorophyll-a, zooplankton biomass, pH and dissolved oxygen were inversely related to axis 1, whereas chlorophyll-a, temperature and pH were inversely related to axis 2 (Table 5, Figure 5).
The Bregmaceros bathymaster, Ophichthus zophochir, Bathycongrus macrurus, Antennarius sanguineus, Gobiesox sp., Lythrypnus sp., Achirus sp., and Coryphaena hippurus were directly related to chlorophyll-a and pH, whereas Anchoa sp., Anisotremus sp., Auxis sp., and Opistognathus panamensis were directly correlated with temperature. Finally, most of the species identified in the study periods and years evaluated were directly correlated with salinity, dissolved oxygen and zooplankton biomass, such as Cetengraulis mysticetus, Gobiidae sp., Oligoplites saurus, Caranx sp., Chromis sp., and Lampanyctus parvicauda (Figure 5).
% (Clench's nonlinear estimator) and the species richness and diversity varied significantly among the study periods and years, being higher in March 2018 and September 2019 than in the rest of the periods and years evaluated (Table 2).
Larval density (ind/100 m3) between the study periods did not show significant differences, except between March and September 2018. Zooplankton biomass (g/100 m3) was significantly higher in March 2019 than in the other periods evaluated (Figure 2). A non-significant positive correlation was established between larval density and zooplankton biomass (R= 0.14, p<0.05; Figure 3) in the study area, suggesting that zooplankton biomass did not have a dominant effect on larval density.
Based on the analysis of similarity between periods and years, six significant groups (I, II, III, IV, V, and VI) were established (SIMPROF, p < 0.05), with dissimilarity percentages between groups above than 65% (Figure 4). Each established group consisted of nine sampling stations in each study period. Group I (March-2018) was established due to the contribution of C. mysticetus, D. pacificus, Anchoa sp., and Anisotremus sp. (SIMPER, 57.36% similarity), which showed a tendency to cluster with those of group II. Group II (March-2017) was formed by the contribution of species such as: Anchoa sp., C. mysticetus, B. bathymaster, and O. saurus (SIMPER, 72% similarity). Group III (March-2019) was formed owing to the contribution of species such as: C. mysticetus, Caranx sp. Seriola sp., and Anisotremus sp. (SIMPER, 70% similarity) (Figure 4).
Group IV (September-2018) was formed due to the contribution of B. bathymaster, Gobiidae sp. 6, Anisotremus sp., and Gobiidae sp (SIMPER, 62.65% similarity). On the other hand, group V (September-2017) was established due to the contribution of species such as: Anisotremus sp., C. mysticetus, Microgobius sp., and S. evermanni (SIMPER, 58.72% of similarity) and group VI (September 2019) was formed due to the contribution of C. mysticetus, Anisotremus sp., Gobiidae sp. and Caranx sp. (SIMPER, 49.0% similarity). Finally, the ANOSIM similarity analysis established a significance level of 0.1% for the periods, with an overall R of 0.764, indicating that there was a difference between assemblages from all periods (Table 3).
4. Discussion
In the water column of the La Azufrada coral reef, there were abundant larvae of fish species that are typical of different marine environments. This characteristic is frequently reported in scientific literature, where species have been recorded from oceanic areas, sandy bottoms, and even mangrove environments in rocky or coral reef areas [9,58,59]. Planktonic organisms, such as fish larvae, are modulated by physical processes of transport (such as advection), behavioral (vertical migrations), and life history characteristics, and they tend to exhibit a high dispersal capacity [60]. This means that they can be present in environments that are atypical for adults. Furthermore, on the reef coral structures can function as originators of accumulation areas by mechanically disrupting currents, waves, and tidal flows [61,62,63].
The species richness distribution during the sampling periods was variable and did not show a clear temporal or annual trend. However, there were significant differences in larval density between the periods (March and September 2018), which coincided with a temporal order consistent with the climatic and oceanographic periods of the area (cold and warm seasons). Similarly, it was observed that there is a higher total larval density in the months of March in all the years evaluated, this may be due to the fact that in general there was a greater availability of zooplankton biomass in the months of March, finding significant differences between March and September of the years 2017 and 2018, thus favoring the fish species to synchronize their spawning periods for these dates. Furthermore, although the cluster analysis suggested that each period evaluated was a separate group from the others, in a general way, a tendency was observed for the groups to be ordered concordantly with the cold and warm seasons. Similarly, the analysis of similarity (ANOSIM), where the overall R determined that all groups (periods evaluated) were different, but there was a tendency for the fish larval assemblages from September to be more like each other.
Over the last few decades, marine fish larval assemblages have been described to present high spatial and temporal dynamics, where variations in composition and abundance occur on time scales ranging from hours to annual seasons [64]. In general terms, this temporal variability would modulate the simultaneous effect of multiple hydrographic and environmental factors that may come to determine larval survival [15,65,66,67], so it could come to determine the distribution, abundance and, in general, the structure of fish larval assemblages [68,69,70,71,72].
Another determining factor in the distribution, abundance and structure of fish larval assemblages is food availability, which is largely responsible for their survival and growth in the early stages of fish development [73]. Generally, fish larval density is closely related to zooplankton biomass and correlates with an increase in local food availability [74,75,76]. This coincides with the results obtained in this study, in which most fish larval species found on the coral reef were directly correlated with zooplankton biomass. Moreover, it has been documented that marine surface circulation patterns have a strong effect on the spatial distribution of fish larvae [77,78,79], giving rise to retention or advective processes that can decrease or increase the abundance of fish larvae in a locality [80,81]. During September, on the eastern side of Gorgona Island, the instantaneous surface circulation pattern is towards the sea, favoring the drifting movement of fish larvae, while during February the trend is towards the coast, which increases retention of larvae above the La Azufrada coral reef [38]. In our study, a trend towards a higher larval density in March compared to September was evident in all years evaluated. This result would be associated with the formation during the first months of the year of a cyclonic gyre in the vicinity of the la Azufrada coral reef [38], which may be favoring the retention of fish larvae and directly increasing the larval density in the cold period of the year.
The structural complexity of coral reefs is considered one of the most important factors in the organization of fish assemblages [82]. These habitats provide refuge areas against predators for many fish species and generate a food source consisting mainly of benthic invertebrates and macroalgae [83,84,85,86]. They also constitute natural sites where fish larvae can find the protection and food necessary for their rapid growth [87]. However, although the La Azufrada coral reef may serve a function as a nursery area for different fish species, most of these species probably exhibit life history strategies that reduce dependence on coral environments, and are likely to exploiting different habitat types during vulnerable life stages [15], this is reflected in the few species of fish larvae found in this study that in their adult stage are related to coral reefs (Antennarius sanguineus, Pseudogramma thaumasium, Oligoplites saurus, and Chaetodipterus zonatus), in addition to the fact that all were found exclusively in the month of March with the exception of Oligoplites saurus.
Another aspect to consider is the seasonal fluctuations in larval production, which are strongly associated with the reproductive events of the species and seem to be synchronized with the requirements of the larvae to increase recruitment success, defining the spatiotemporal pattern of distribution [65,88,89,90]. Thus, species such as C. mysticetus, B. bathymaster, Caranx sp., and Anisotremus sp. reproduce several times a year. Species collected only during a local oceanographic period, could be larvae of species that present only one reproductive event per year, such as Dormitator sp., and Menticirrhus sp. that were recorded in the month of March and Microgobius sp., and Stelifer sp. that were present in September.
5. Conclusions
The composition and structure of the fish larval assemblage in all evaluated periods were different and specific for each period. However, a tendency was observed for them to group together or be more similar between the climatic and oceanographic periods described for the study area (cold and warm seasons). This suggests that the intra-annual variation has a greater effect on the fish larval assemblages, where it is evident that the cold period (March) presents better conditions for the fish species to carry out their spawning season; and subsequently there is a higher larval density in the la Azufrada coral reef.
The results of this study increase the available knowledge on the assemblage of fish larvae associated with the ETP coral reef and how it varies during the year and between the years, showing that intra-annual variation, and in particular the cold season, have a greater effect on the assemblage of fish larvae associated whit the La Azufrada coral reef. Some species were frequent in the years and seasons evaluated and others in part of these, probably reflecting different spawning strategies or ontogenetic changes in distribution. These differences resulted in temporal variations in the species assemblage. However, considering the method implemented to capture fish larvae in our study (zooplankton trawls), it is necessary to implement complementary capture methods, such as light traps, to obtain information on more advanced stages of development and thus obtain information on population recruitment processes, which allows a complete overview of the temporal dynamics of the different fish species that are related to the coral reefs of the Eastern Tropical Pacific.
Supplementary Materials
Table A1 can be downloaded as supplementary materials.
Author Contributions
JJGZ: Methodology, Validation, Formal analysis, Investigation, writing original draft, Writing review & editing, Data curation, Visualization. DFCR: Conceptualization, Methodology, Formal analysis, Supervision. AGL: Conceptualization, Methodology, Investigation, Writing review & editing, Supervision, Funding acquisition.
Funding
This research was funded by the Ministry of Environment and Sustainable Development of the Republic of Colombia (National Natural Parks - PNN) and Universidad del Valle (Grupo de Investigación en Ciencias Oceanográficas), associated to Gorgona National Park Pelagic Environment monitoring program.
Data Availability Statement
Data are contained within the article and Supplementary Materials
Acknowledgments
The authors express their gratitude to members of the research group in oceanographic sciences of the Universidad del Valle and the officials of the Gorgona National Natural Park for all their support during the sampling efforts. Ricardo Saldierna y Gerardo Aceves from the CICIMAR-IPN Ichthyoplankton laboratory confirmed the taxonomic identification of the fish larvae collected during our study. All the biological material was deposited in the collection of zoological practices of the Universidad del Valle.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Amador, J.A.; Alfaro, E.J.; Lizano, O.G.; Magaña, V.O. Atmospheric forcing of the eastern tropical Pacific: a review. Prog. Oceanogr. 2006, 69, 101–142. [Google Scholar] [CrossRef]
- Fiedler, P.C.; Talley, L.D. Hydrography of the eastern tropical Pacific: a review. Progress. Oceanogr. 2006, 69, 143–160. [Google Scholar] [CrossRef]
- Fiedler, P.C.; Lavín, M.F. Oceanographic Conditions of the Eastern Tropical Pacific. In Coral Reefs of the Eastern Tropical Pacific. Coral Reefs of the World; Glynn, P.; Manzello, D.; Enochs, I., Eds., vol 8.; Springer, Dordrecht, 2017; pp. 59-83. [CrossRef]
- Eslava, J.A. (1993). Climatología. En P. Leyva (Ed.), Colombia Pacífico. Tomo I; Fondo para la protección del medio ambiente, FEN, Santafé de Bogotá, Colombia, 1993; pp. 136–147.
- Velandia, M.C.; Scheel, M.; Puentes, C.; Durán, D.; Osorio, P.; Delgado, P.; Obando, N.; Prieto, A.; Díaz, J.M. 2019.
- Sale, P.F. Connectivity, recruitment variation, and the structure of reef fish communities. Integr Comp Biol. [CrossRef]
- Cortés, J.; Enochs, I.C.; Sibaja-Cordero, J.; Hernández, L.; Alvarado, J.J.; Breedy, O. ;... Zapata, F.A. Marine biodiversity of Eastern Tropical Pacific coral reefs. In Coral reefs of the eastern tropical Pacific; Glynn et al., Eds.; Springer, Dordrecht, 2017; pp. 203-250. [CrossRef]
- Álvarez-Barco, J.A.; Rodríguez-Quintal, J.G.; Rodríguez, C.T.; Narciso-Fejure, S.E. Moluscos asociados al arrecife coralino de Isla Larga, Parque Nacional San Esteban, Estado Carabobo, Venezuela. Mem. Fund. La Salle Cien. Nat. Fundación 2019, 77. [Google Scholar]
- Brogan, M.W. Distribution and retention of larval fishes near reefs in the Gulf of California. Mar. Ecol. Prog. Ser. 1994, 115, 1–1. [Google Scholar] [CrossRef]
- Hamner, W.M; Largier, J.L. Oceanography of the planktonic stages of aggregation spawning reef fishes. In Reef fish spawning aggregations: biology, research and management; Sadovy de Mitcheson, Y.; Colin, P.L., Eds.; Springer, Dordrecht, 2012; pp. 159-190. [CrossRef]
- Cowen, R.K.; Paris, C.B.; Srinivasan, A. Scaling of connectivity in marine populations. Sci. 2006, 311, 522–527. [Google Scholar] [CrossRef]
- Lefevre, C.D.; Bellwood, D.R. Disturbance and recolonisation by small reef fishes: the role of local movement versus recruitment. Mar. Ecol. Prog. Ser. 2015, 537, 205–215. [Google Scholar] [CrossRef]
- Kingsford, M.J. Diel patterns of abundance of presettlement reef fishes and pelagic larvae on a coral reef. Mar. Biol. 2001, 138, 853–867. [Google Scholar] [CrossRef]
- Flores-Coto, C.; Gutiérrez, R.M.; González-Félix, M.; Sanvicente-Añorve, L.; García, F.Z. Annual Variation of Ichthyoplancton Assemblages in Neritic Waters of the Southern Gulf of Mexico. Caribb. J. Sci. 2000, 36, 233–243. [Google Scholar]
- Leis, J.M.; McCormick, M.I. The biology, behavior and ecology of the pelagic, larval stage of coral-reef fishes. In Sale, P.F. (Ed.). Coral reef fishes: New insights into their ecology. Academic Press, San Diego, 2022; pp. 171-199. [CrossRef]
- Jiménez-Rosenberg, S.P.A.; Saldierna-Martínez, R.J.; Aceves-Medina, G.; Cota–Gómez, V.M. (2007) Fish larvae in Bahía Sebastián Vizcaíno and the adjacent oceanic region, Baja California, México. Check. list. 2007, 3, 204–223. [Google Scholar] [CrossRef]
- León-Chávez, C.A.; Beier, E.; Sánchez-Velasco, L.; Barton, E.D.; Godínez, V.M. Role of circulation scales and water mass distributions on larval fish habitats in the Eastern Tropical Pacific off Mexico. J. Geophys. Res. Oceans. 2015, 120, 3987–4002. [Google Scholar] [CrossRef]
- Ribeiro, F.; Hale, E.; Hilton, E.J.; Clardy, T.R. : Deary, A.L.; Targett, T.E.; Olney, J.E. Composition and temporal patterns of larval fish communities in Chesapeake and Delaware Bays, USA. Mar. Ecol. Progr. Ser. 2015, 527, 167–180. [Google Scholar] [CrossRef]
- Gallego-Zerrato, J.J.; Córdoba-Rojas, D.F.; Giraldo, A. Composición taxonómica de las larvas de peces en el arrecife coralino de La Azufrada, Pacífico colombiano, entre 2017 a 2019. Bol. Cient. Mus. Hist. 2023, 27, 245–261. [Google Scholar] [CrossRef]
- Blaxter, J.H.S. The effect of temperature on larval fishes. Neth. J. Zool. 1991, 42, 336–357. [Google Scholar] [CrossRef]
- Lambert, Y.; Yaragina, N.; Kraus, G.; Marteinssdottir, G.; Wright, P.J. Using environmental and biological indices as proxies for egg and larval production of marine fish. J. Northwest Atl. Fish. Sci. 2003, 33, 115–159. [Google Scholar] [CrossRef]
- Perry, R.I.; Cury, P.; Brander, K.; Jennings, S.; Möllmann, C.; Planque, B. Sensitivity of marine systems to climate and fishing: concepts, issues and management responses. J. Mar. Syst. 2010, 79, 427–435. [Google Scholar] [CrossRef]
- Villamizar, N.; Blanco-Vives, B.; Migaud, H.; Davie, A.; Carboni, S.; Sanchez-Vazquez, F.J. Effects of light during early larval development of some aquacultured teleosts: A review. Aquac. 2011, 315, 86–94. [Google Scholar] [CrossRef]
- Leis, J.M. Larval fish assemblages near Indo-Pacific coral reefs. Bull. Mar. Sci. 1993, 53, 362–392. [Google Scholar]
- Victor, B.C.; Wellington, G.M. Endemism and the pelagic larval duration of reef fishes in the eastern Pacific Ocean. Mar. Ecol. Prog. 2000, 205, 241–248. [Google Scholar] [CrossRef]
- Leis, J.M. Pacific coral-reef fishes: the implications of behaviour and ecology of larvae for biodiversity and conservation, and a reassessment of the open population paradigm. Environ. Biol. Fishes. 2002, 65, 199–208. [Google Scholar] [CrossRef]
- Cortés, J.; et al. Marine Biodiversity of Eastern Tropical Pacific Coral Reefs. In Coral Reefs of the Eastern Tropical Pacific. Coral Reefs of the World; Glynn, P.; Manzello, D.; Enochs, I., Eds., vol 8.; Springer, Dordrecht, 2017. Pp. [CrossRef]
- Edgar, G.J.; Banks, S.A.; Bessudo, S.; Cortés, J.; Guzmán, H.M.; Henderson, S.; Martinez, C.; Rivera, F.; Soler, G.; Ruiz, D.; Zapata, F.A. Variation in reef fish and invertebrate communities with level of protection from fishing across the Eastern Tropical Pacific seascape. Glob. Ecol. Biogeogr. 2011, 20, 730–743. [Google Scholar] [CrossRef]
- Dubuc, A.; Quimbayo, J.P.; Alvarado, J.J.; Araya-Arce, T.; Arriaga, A.; Ayala-Bocos, A.; Julio Casas-Maldonado, J.; Chasqui, L.; Cortés, J.; Cupul-Magaña, A.; Olivier, D.; Olán-González, M.; González-Leiva, A.; López-Pérez, A.; Reyes-Bonilla, H.; Smith, F.; Rivera, F.; Rodríguez-Zaragoza, F.A.; Rodríguez-Villalobos, J.C.; Segovia, J.; Zapata, F.A.; Bejarano, S. Patterns of reef fish taxonomic and functional diversity in the Eastern Tropical Pacific. Ecography. 2023, 2023. [Google Scholar] [CrossRef]
- Eisele, M.H.; Madrigal-Mora, S.; Espinoza, M. Drivers of reef fish assemblages in an upwelling region from the Eastern Tropical Pacific Ocean. J. Fish Biol. 2021, 98, 1074–1090. [Google Scholar] [CrossRef] [PubMed]
- Rubio, E.A.; Gutiérrez, B.; Franke, R. Peces de la isla de Gorgona. Universidad del Valle, Cali, Colombia, 1987; pp. 323.
- Mora, C.; Ospina, A. Experimental effect of cold, la nina temperatures on the survival of reef fishes from Gorgona island (eastern Pacific Ocean). Mar. Biol. 2002, 141, 789–793. [Google Scholar] [CrossRef]
- Ospina, A.F.; Mora, C. Effect of body size on reef fish tolerance to extreme low and high temperatures. Environ. Biol. Fishes. 2004, 70, 339–343. [Google Scholar] [CrossRef]
- Palacios, M.M.; Zapata, F.A. Fish community structure on coral habitats with contrasting architecture in the Tropical Eastern Pacific. Rev. Biol. Trop. 2014, 62, 343–357. [Google Scholar] [CrossRef]
- Palacios, M.M.; Muñoz, C.G.; Zapata, F.A. Fish corallivory on a pocilloporid reef and experimental coral responses to predation. Coral Reefs. 2014, 33, 625–636. [Google Scholar] [CrossRef]
- Alzate, A.; Zapata, F.A.; Giraldo, A. A comparison of visual and collection-based methods for assessing community structure of coral reef fishes in the Tropical Eastern Pacific. Rev. Biol. Trop. 2014, 62, 359–371. [Google Scholar] [CrossRef]
- Quimbayo, J.P.; Zapata, F.A. Cleaning interactions by gobies on a tropical eastern Pacific coral reef. J. Fish. Biol. 2018, 92, 1110–1125. [Google Scholar] [CrossRef] [PubMed]
- Calle-Bonilla, I.C.; López, A.G.; Cuéllar-Chacón, A. Spaciotemporal variation in the coral fish larvae assembly of Gorgona Island, Colombian Pacific. Bol. Invest. Mar. Cost. 2017, 46, 55–72. [Google Scholar] [CrossRef]
- Ramírez-Martínez, G.A.; Giraldo, A.; Rivera-Gómez, M.; Aceves-Medina, G. Daily and monthly ichthyoplankton assemblages of La Azufrada coral reef, Gorgona Island, Eastern Tropical Pacific. Reg. Stud. Mar. Sci. 2022, 52. [Google Scholar] [CrossRef]
- Rojas, P.; Zapata, L.A. Peces demersales del Parque Nacional Natural Gorgona y su área de influencia, Pacífico colombiano. Bio. Col. 2006, 7, 211–244. [Google Scholar]
- Blanco, J.F. “The hydroclimatology of Gorgona Island: seasonal and ENSO–related patterns”. Act. Biol. 2009, 91, 111–121. [Google Scholar] [CrossRef]
- Giraldo, A.; Rodríguez-Rubio, E.; Zapata, F. Condiciones oceanográficas en isla Gorgona, Pacífico oriental tropical de Colombia. Lat. Am. J. Aquat. 2008, 36, 121–128. [Google Scholar] [CrossRef]
- Giraldo, A.; Díaz-Granados, M.C.; Gutiérrez-Landázuri, C.F. Isla Gorgona, enclave estratégico para los esfuerzos de conservación en el Pacífico Oriental Tropical. Rev. biol. Trop. 2014, 62, 1–12. [Google Scholar] [CrossRef]
- Zapata, F.A.; Morales, Y.A. Spatial and temporal patterns of fish diversity in a coral reef at Gorgona Island, Colombia. In Proceedings of the 8th international Coral Reef symposium, Vol. 1.; Ciudad de, Panamá, Panamá; 1997; pp. 1029–1034. [Google Scholar]
- Zapata, F.A.; Vargas-Ángel, B. Corals and coral reefs of the Pacific coast of Colombia. En: Coral reefs of Latin America Elsevier Science; Cortes, J. Eds.; Amsterdam, Holanda, 2003; pp. 419-447. [CrossRef]
- Glynn, P.W.; Von Prahl, H.; Guhl, F. Coral reefs of Gorgona Island, Colombia, with special reference to corallivores and their influence on community structure and reef development. An. Inst. Invest. Mar. Punta. BetõÂn. 1982, 12, 185–214. [Google Scholar] [CrossRef]
- Valencia, B.; Lavaniegos, B.; Giraldo, A.; Rodríguez-Rubio, E. Temporal and spatial variation of hyperiid amphipod assemblages in response to hydrographic processes in the Panama Bight, eastern tropical Pacific. Deep-Sea Res. I: Oceanogr. Res. 2013, 73, 46–61. [Google Scholar] [CrossRef]
- Rivera-Gómez, M.; Giraldo, A.; Lavaniegos, B. Structure of euphausiid assemblages in the Eastern Tropical Pacific off Colombia during El Niño, La Niña and neutral conditions. J. Exp. Mar. Biol. Ecol. 2019, 516, 1–15. [Google Scholar] [CrossRef]
- Valencia, B.; Giraldo, A. Structure of hyperiid amphipod assemblages on Isla Gorgona, eastern tropical Pacific off Colombia. J. Mar. Biol. Assoc. 2012, 92. [Google Scholar] [CrossRef]
- Giraldo, A.; Valencia, B.; Acevedo, J.D.; Rivera, M. Fitoplancton y zooplancton en el área marina protegida de Isla Gorgona, Colombia, y su relación con variables oceanográficas en estaciones lluviosa y seca. Rev. biol. Trop. [CrossRef]
- Moser, H.G. The early stages of fishes in the California current region. California cooperative oceanic fisheries investigations, Atlas No. 33. Nat. Mar. Fish. Serv. SFSC. La Jolla, Calif. Allen Press, Inc., Lawrence, Kans, 1996; 18. 9358. [Google Scholar]
- Beltrán-León, B.S.; Ríos, R. Estadios tempranos de peces del Pacífico colombiano. Instituto Nacional de Pesca y Acuicultura INPA-Buenaventura, Colombia, 2000; pp. 727. 9: ISBN, 9589. [Google Scholar]
- Boeing, W.J.; Duffy-Anderson, J.T. Ichthyoplankton dynamics and biodiversity in the Gulf of Alaska: responses to environmental change. Ecol. Indic. 2008, 8, 292–302. [Google Scholar] [CrossRef]
- Soberón, M.J.; Llorente, B.J. The use of species accumulation functions for the prediction of species richness. Conserv. Biol. 1993, 7, 480–488. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation. 2nd Edition, PRIMER-E, Ltd. 2001. [Google Scholar]
- Schiller, C. Monitoring river health initiative technical report number 36. 2003. www.deh.gov.au/index.html. 12/05/07.
- Escarria, E.; Beltrán, B.; Giraldo, A.; Zapata, F. Ichthyoplankton in the National Natural Park Isla Gorgona (Pacific Ocean of Colombia) during September 2005. Bol. Investig. Mar. 2007, 35, 127–133. [Google Scholar] [CrossRef]
- Leis, J.M.; Goldman, B. A preliminary distributional study of fish larvae near a ribbon coral reef in the Great Barrier Reef. Coral reefs. 1984, 2, 197–203. [Google Scholar] [CrossRef]
- Sponaugle, S.; Fortuna, J.; Grorud, K.; Lee, T. Dynamics of larval fish assemblages over a shallow coral reef in the Florida Keys. Mar. Biol. 2003, 143, 175–189. [Google Scholar] [CrossRef]
- Leis, J.M. Are larvae of demersal fishes plankton or nekton? Adv. Mar. Biol. 2006, 51, 57–141. [Google Scholar] [CrossRef]
- Hamner, W.M.; Wolanski, E. Hydrodynamic forcing functions and biological processes on coral reefs: a status review. In Proc. 6th Int. Coral Reef Symp, 1988; Vol 1; pp. 103-113.
- Irisson, J.O.; LeVan, A.; De Lara, M.; Planes, S. Strategies and trajectories of coral reef fish larvae optimizing self-recruitment. J. Theor. Bio. 2004, 227, 205–218. [Google Scholar] [CrossRef]
- Leahy, S.M.; Russ, G.R.; Abesamis, R.A. Pelagic larval duration and settlement size of a reef fish are spatially consistent, but post-settlement growth varies at the reef scale. Coral Reefs. 2015, 34, 1283–1296. [Google Scholar] [CrossRef]
- Leis, J.M. The pelagic stage of reef fishes: the larval biology of coral reef fishes. In: The ecology of fishes on coral reefs; Sale, P.F.; Eds; University of Hawaii, Honolulu, 1991; pp. 183-230. [CrossRef]
- Robertson, D.R. The role of adult biology in the timing of spawning of tropical reef fishes. En: The Ecology of Fishes on Coral Reefs; Sale, P.F; Eds.; Academic Press, San Diego, USA, 1991; pp. 356–386. [CrossRef]
- Lo, W.T.; Hsieh, H.Y.; Wu, L.J.; Jian, H.B.; Liu, D.C.; Su, W.C. Comparison of larval fish assemblages between during and after northeasterly monsoon in the waters around Taiwan, western North Pacific. J. Plankton. Res. 2010, 32, 1079–1095. [Google Scholar] [CrossRef]
- Sánchez-Velasco, L.; Beier, E.; Godínez, V.M.; Barton, E.D.; Santamaría-del-Angel, E.; Jiménez-Rosemberg, S.P.A.; Marinone, S.G. Hydrographic and fish larvae distribution during the “Godzilla El Niño 2015–2016” in the northern end of the shallow oxygen minimum zone of the Eastern Tropical Pacific Ocean. J. Geophys. Res. Oceans. 2017, 122, 2156–2170. [Google Scholar] [CrossRef]
- Franco-Gordo, C.; Godínez-Domínguez, E.; Suárez-Morales, E. Larval fish assemblages in waters off the central Pacific coast of Mexico. J. Plankton. Res. 2002, 24, 775–784. [Google Scholar] [CrossRef]
- Funes-Rodríguez; R. ; Flores-Coto, C.; Esquivel-Herrera, A.; Fernández-Alamo, M.A.; Gracia-Gásca, A. Larval fish community structure along the west coast of Baja California during and after the El Niño event (1983). Bull. Mar. Sci. 2002, 70, 41–54. [Google Scholar]
- Aceves-Medina, G.; Jiménez-Rosenberg, S.P.A.; Hinojosa-Medina, A.; Funes-Rodríguez, R.; Saldierna, R.J.; Lluch-Belda, D. . Watson, W. Fish larvae from the Gulf of California. Sci. Mar. 2003, 67, 1–11. [Google Scholar] [CrossRef]
- Aceves-Medina, G.; Saldierna-Martínez, R.J.; Hinojosa-Medina, A.; Jiménez-Rosenberg, S.P.; Hernández-Rivas, M.E.; Morales-Ávila, R. Vertical structure of larval fish assemblages during diel cycles in summer and winter in the southern part of Bahía de La Paz, México. Estuar. Coas. Shelf. Sci. 2008, 76, 889–901. [Google Scholar] [CrossRef]
- Hsieh, H.Y.; Lo, W.T.; Chen, H.H.; Meng, P.J. Larval fish assemblages and hydrographic characteristics in the coastal waters of southwestern Taiwan during non-and post-typhoon summers. Zool. Stud. 2016, 55, 1–18. [Google Scholar] [CrossRef]
- Somarakis, S.; Drakopoulos, P.; Filippou, V. Distribution and abundance of larval fish in the northern Aegean Sea—eastern Mediterranean—in relation to early summer oceanographic conditions. J. Plankton. Res. 2002, 24, 339–358. [Google Scholar] [CrossRef]
- Kaunda-Arara, B.; Mwaluma, J.M.; Locham, G.A.; Øresland, V.; Osore, M.K. Temporal variability in fish larval supply to Malindi Marine Park, coastal Kenya. Aquat. Conserv: Mar. Freshw. Ecosyst. 2009, 19, S10–S18. [Google Scholar] [CrossRef]
- Sponaugle, S.; Llopiz, J.K.; Havel, L.N.; Rankin, T.L. Spatial variation in larval growth and gut fullness in a coral reef fish. Mar. Ecol. Prog. Ser. 2009, 383, 239–249. [Google Scholar] [CrossRef]
- Leis, J.M. Ontogeny of behaviour in larvae of marine demersal fishes. Ichthyol. Res. 2010, 7, 325–342. [Google Scholar] [CrossRef]
- Cowen, R.K.; Castro, L.R. Relation of coral reef fish larval distributions to island scale circulation around Barbados, West Indies. Bull. Mar. Sci. 1994, 54, 228–244. [Google Scholar]
- Reiss, C.S.; Panteleev, G.; Taggart, C.T.; Sheng, J.; DeYoung, B. Observations on larval fish transport and retention on the Scotian Shelf in relation to geostrophic circulation. Fish. Oceanogr. 2000, 9, 195–213. [Google Scholar] [CrossRef]
- Feng, M.; Slawinski, D.; Beckley, L.E.; Keesing, J.K. Retention and dispersal of shelf waters influenced by interactions of ocean boundary current and coastal geography. Mar. Freshw. Res. 2010, 61, 1259–1267. [Google Scholar] [CrossRef]
- Sánchez-Velasco, L.; Beier, E.; Avalos-García, C.; Lavín, M.F. Larval fish assemblages and geostrophic circulation in Bahía de La Paz and the surrounding southwestern region of the Gulf of California. J. Plankton. Res. 2006, 28, 1081–1098. [Google Scholar] [CrossRef]
- Olivar, M.P.; Emelianov, M.; Villate, F.; Uriarte, I.; Maynou, F.; Alvarez, I.; Morote, E. The role of oceanographic conditions and plankton availability in larval fish assemblages off the Catalan coast (NW Mediterranean). Fish. Oceanogr. 2010, 19, 209–229. [Google Scholar] [CrossRef]
- Beck, M.W. Comparison of the measurement and effects of habitat structure on gastropods in rocky intertidal and mangrove habitats. Mar. Ecol. Prog. Ser. 1998, 169, 165–178. [Google Scholar] [CrossRef]
- Moberg, F.; Folke, C. Ecological goods and services of coral reef ecosystems. Ecol. Econ. 1999, 29, 215–233. [Google Scholar] [CrossRef]
- Nagelkerken, I.; Dorenbosch, M.; Verberk, W.C.E.P.; De La Morinière, E.C.; van Der Velde, G. Importance of shallow-water biotopes of a Caribbean Bay for juvenile coral reef fishes: patterns in biotope association, community structure and spatial distribution. Mar. Ecol. Prog. Ser. 2000, 202, 175–192. [Google Scholar] [CrossRef]
- Bellwood, D.R.; Wainwright, P.C. (2002) The history and biogeography of fishes on coral reefs. In: Coral reef fishes: dynamics and diversity in a complex ecosystem: Sale, P.F., Eds.; Academic Press, 2002; pp. 5-32. [CrossRef]
- Mumby, P.J.; Broad, K.; Brumbaugh, D.R.; Dahlgren, C.P.; Harborne, A.R.; Hastings, A. . Sanchirico, J.N. Coral reef habitats as surrogates of species, ecological functions, and ecosystem services. Conserv. Biol. 2008, 22, 941–951. [Google Scholar] [CrossRef] [PubMed]
- McConnell, R.; Lowe-McConnell, R.H. Ecological studies in tropical fish communities. Cambridge University Press, Cambridge, 1987; pp. 382. [CrossRef]
- Johannes, R.E. Traditional marine conservation methods in Oceania and their demise. Annu. Rev. Ecol. Evol. Syst. 1978, 9, 349–364. [Google Scholar] [CrossRef]
- Pineda, J.; Porri, F.; Starczak, V.; Blythe, J. Causes of decoupling between larval supply and settlement and consequences for understanding recruitment and population connectivity. J. Exp. Mar. Biol. Ecol. 2010, 392, 9–21. [Google Scholar] [CrossRef]
- China, V.; Holzman, R. Hydrodynamic starvation in first-feeding larval fishes. Proc. Natl. Acad. Sci. 2014, 111, 8083–8088. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Geographical location of Gorgona Island and sampling stations on La Azufrada reef. The area of the Azufrada reef plain and slope and the position of La Azufrada on Gorgona Island and the position of Gorgona Island in the Eastern Tropical Pacific are highlighted. Source: Own.
Figure 1.
Geographical location of Gorgona Island and sampling stations on La Azufrada reef. The area of the Azufrada reef plain and slope and the position of La Azufrada on Gorgona Island and the position of Gorgona Island in the Eastern Tropical Pacific are highlighted. Source: Own.
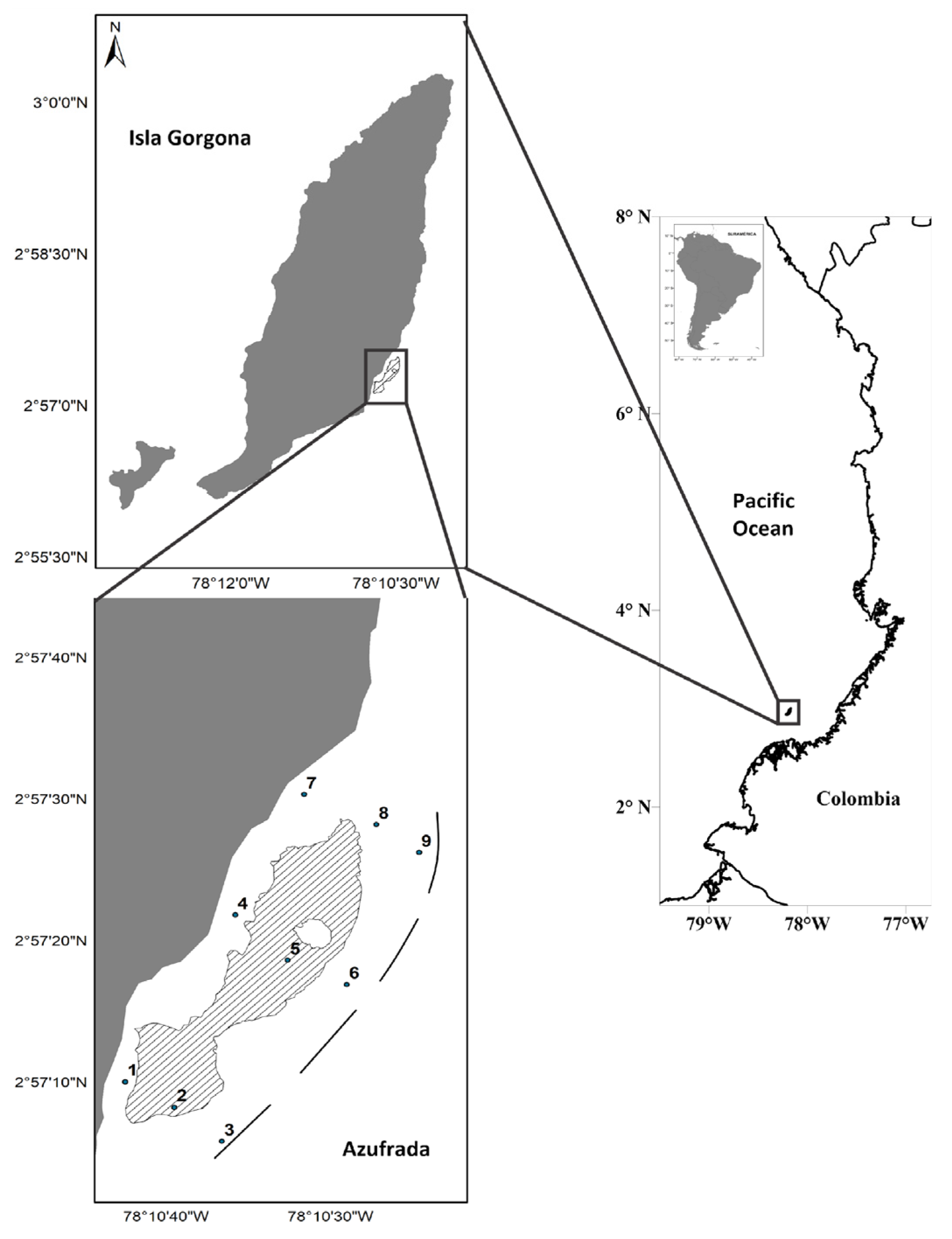
Figure 2.
Zooplankton biomass and fish larval density present in the water column associated with the La Azufrada coral reef during March and September 2017 to 2019. The y-axis is in logarithmic scale.
Figure 2.
Zooplankton biomass and fish larval density present in the water column associated with the La Azufrada coral reef during March and September 2017 to 2019. The y-axis is in logarithmic scale.
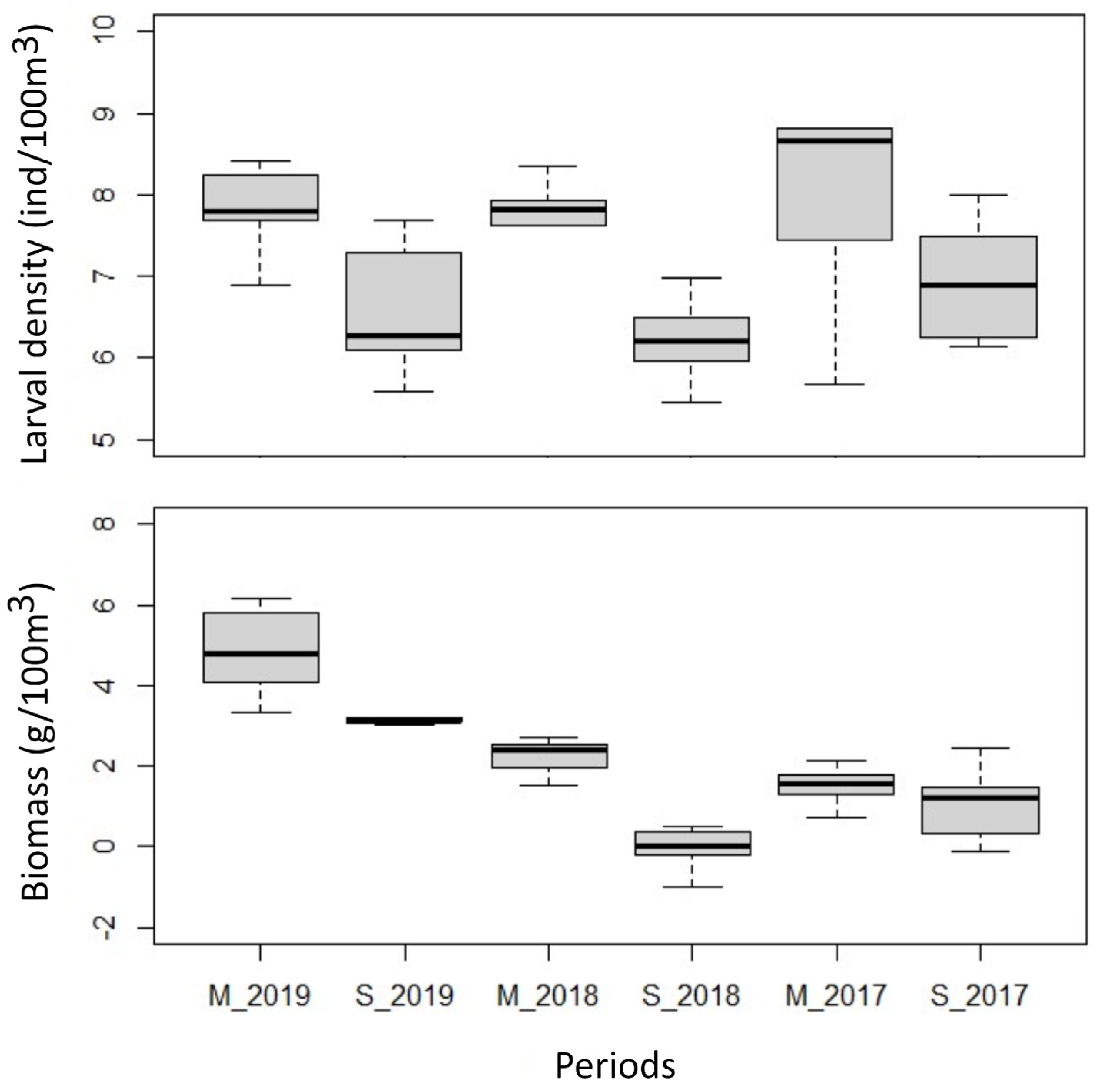
Figure 3.
Spearman correlation showing the relationship between zooplankton biomass and larval density in the study area. The two axes are in logarithmic scale.
Figure 3.
Spearman correlation showing the relationship between zooplankton biomass and larval density in the study area. The two axes are in logarithmic scale.
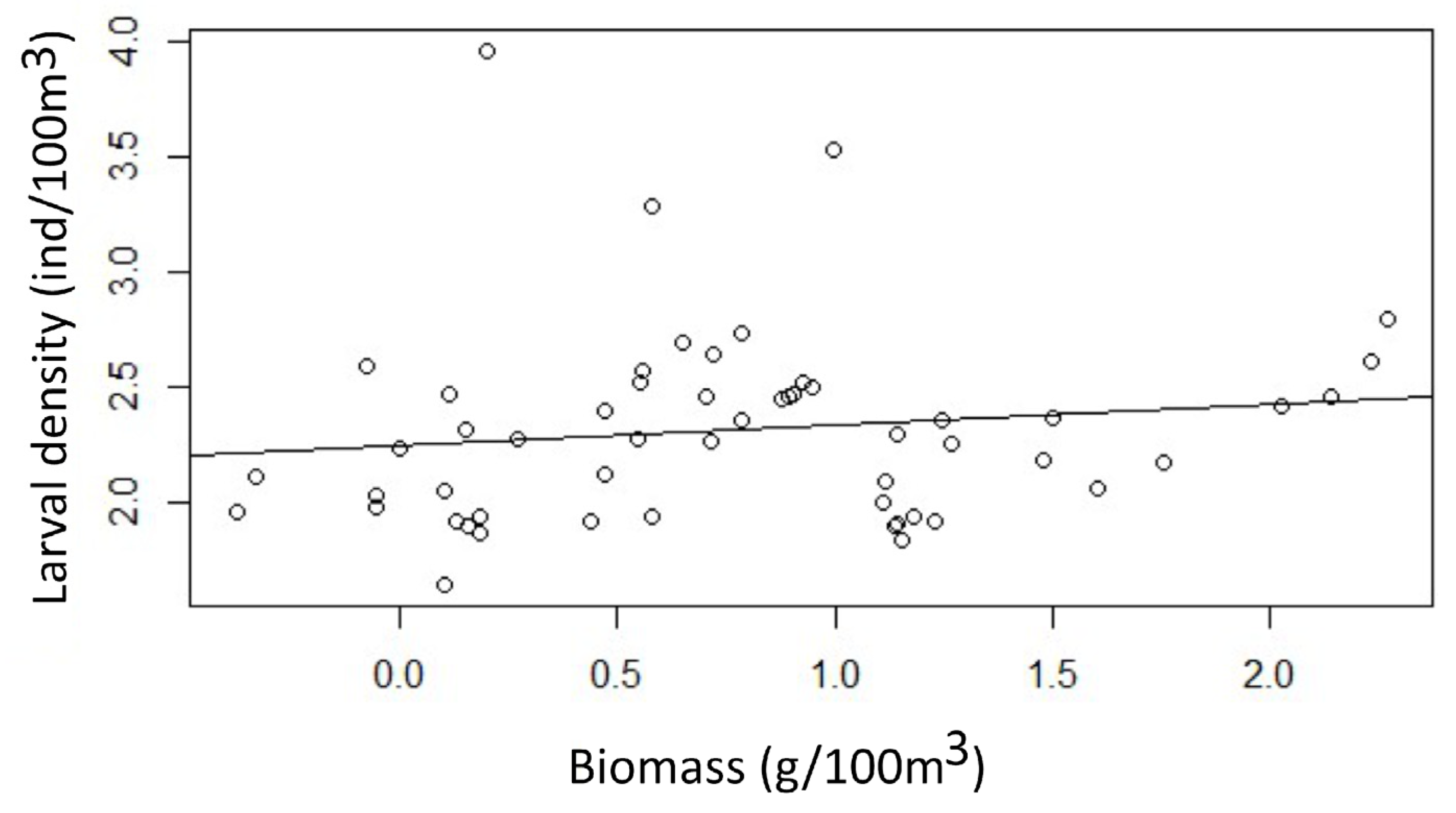
Figure 4.
Similarity dendrogram of the fish larval assemblage present in the water column associated with La Azufrada reef in the March-September periods between 2017 to 2019.
Figure 4.
Similarity dendrogram of the fish larval assemblage present in the water column associated with La Azufrada reef in the March-September periods between 2017 to 2019.
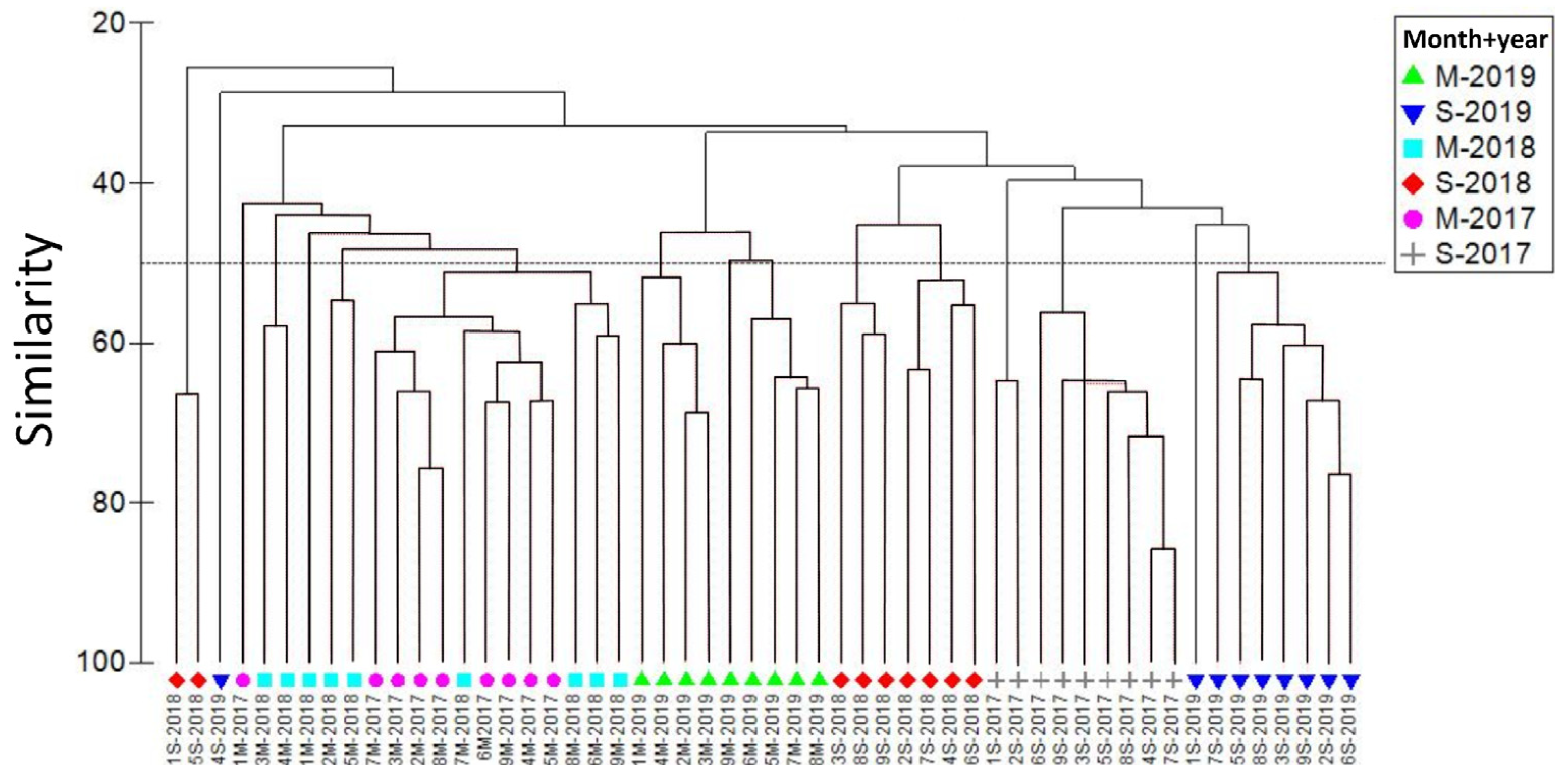
Figure 5.
Canonical correspondence analysis using the abundance of species found with frequency > 5%. The first two canonical axes explain 67% of the total variance.
Figure 5.
Canonical correspondence analysis using the abundance of species found with frequency > 5%. The first two canonical axes explain 67% of the total variance.
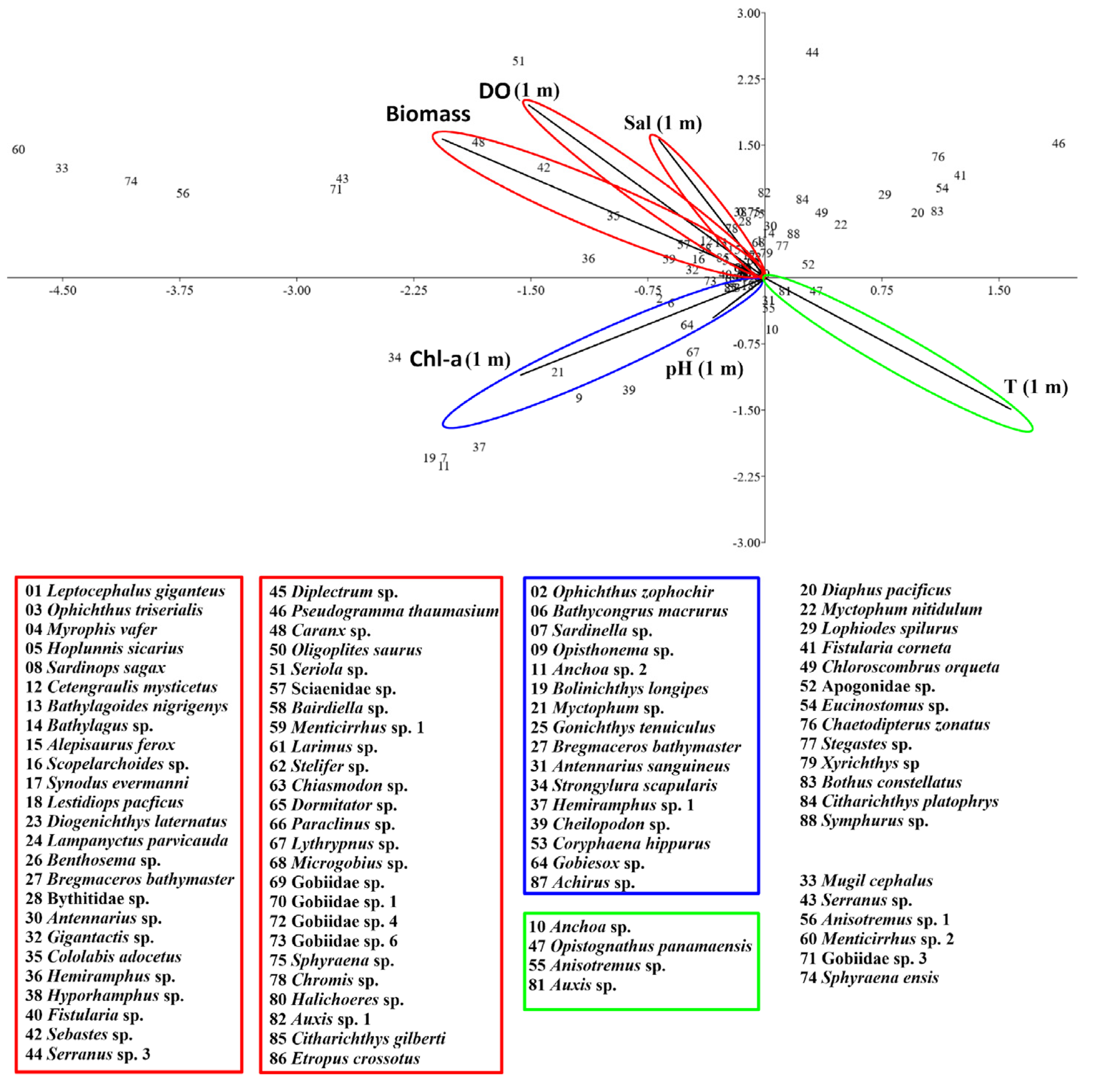

Table 1.
Taxa of fish larvae reported for Gorgona Island and the Azufrada coral reef.
| Authors | Taxa reported | New records | Accumulated taxa |
|---|---|---|---|
| Escarria et al. [57] | 35 | 35 | 35 |
| Calle-Bonilla et al [38] | 29 | 29 | 64 |
| Ramírez et al. [39] | 87 | 57 | 121 |
| This work | 88 | 41 | 462 |
Table 2.
Diversity and richness indices by period evaluated.
| Periods | Richness (d) | Diversity H' (nats/ind) |
|---|---|---|
| March_2017 | 1.64 | 2.25 |
| September_2017 | 1.80 | 2.90 |
| March_2018 | 2.23 | 4.38 |
| September_2018 | 1.71 | 2.99 |
| March_2019 | 1.98 | 2.34 |
| September_2019 | 2.59 | 4.20 |
Table 3.
One-way ANOSIM, with sampling periods (month + year).
| Groups | Statistic R | Significance level P (%) |
|---|---|---|
| M-2019, S-2019 | 0.752 | 0.1 |
| M-2019, M-2018 | 0.921 | 0.1 |
| M-2019, S-2018 | 0.859 | 0.1 |
| M-2019, M-2017 | 0.903 | 0.1 |
| M-2019, S-2017 | 0.791 | 0.1 |
| S-2019, M-2018 | 0.837 | 0.1 |
| S-2019, S-2018 | 0.682 | 0.1 |
| S-2019, M-2017 | 0.962 | 0.1 |
| S-2019, S-2017 | 0.601 | 0.1 |
| M-2018, S-2018 | 0.757 | 0.1 |
| M-2018, M-2017 | 0.287 | 0.1 |
| M-2018, S-2017 | 0.849 | 0.1 |
| S-2018, M-2017 | 0.856 | 0.1 |
| S-2018, S-2017 | 0.576 | 0.1 |
| M-2017, S-2017 | 0.896 | 0.1 |
Table 4.
Physicochemical parameters recorded in the water column associated with La Azufrada coral reef during March-September between 2017-2019. n: number of stations where physicochemical parameters were recorded, μ: mean, S.D.: standard deviation, Min: minimum value recorded and Max: maximum value recorded.
Table 4.
Physicochemical parameters recorded in the water column associated with La Azufrada coral reef during March-September between 2017-2019. n: number of stations where physicochemical parameters were recorded, μ: mean, S.D.: standard deviation, Min: minimum value recorded and Max: maximum value recorded.
| Parameters | Statistics | M-2017 | S-2017 | M-2018 | S-2018 | M-2019 | S-2019 |
|---|---|---|---|---|---|---|---|
| T (°C) | µ | 28.4 | 28.8 | 27.4 | 27.7 | 25.8 | 27.5 |
| S.D. | 0.13 | 0.13 | 0.24 | 0.09 | 0.29 | 0.13 | |
| Min | 28.3 | 28.7 | 27.2 | 27.5 | 25.4 | 27.4 | |
| Max | 28.7 | 29.1 | 27.8 | 27.8 | 26.2 | 27.8 | |
| S (PSU) | µ | 29.6 | 25.2 | 26.7 | 26.1 | 30.7 | 30.3 |
| S.D. | 0.03 | 0.45 | 0.18 | 1.14 | 0.44 | 0.13 | |
| Min | 29.6 | 24.2 | 26.5 | 23.1 | 30.1 | 30.0 | |
| Max | 29.7 | 25.7 | 27.0 | 26.7 | 31.4 | 30.4 | |
| OD (ml/L) | µ | 5.6 | 6.7 | 5.9 | 4.9 | 8.9 | 6.7 |
| S.D. | 0.20 | 0.50 | 0.80 | 0.46 | 0.24 | 0.63 | |
| Min | 5.2 | 6.1 | 5.3 | 4.1 | 8.6 | 6.0 | |
| Max | 5.8 | 7.5 | 7.4 | 5.5 | 9.3 | 7.9 |
Table 5.
Correlation values of each variable with respect to the first two axes in the canonical correspondence analysis. The first two ordination axes explain 67% of the total variance.
Table 5.
Correlation values of each variable with respect to the first two axes in the canonical correspondence analysis. The first two ordination axes explain 67% of the total variance.
| Axis 1 | Axis 2 | |
|---|---|---|
| Variance explained | 36.27 | 31.42 |
| T (1 m) | 0.45 | -0.42 |
| UPS (1 m) | -0.19 | 0.44 |
| DO (1 m) | -0.43 | 0.56 |
| pH-1 m | -0.10 | -0.13 |
| Chl-a (1 m) | -0.45 | -0.32 |
| Biomass | -0.59 | 0.45 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



