
Submitted:
09 December 2024
Posted:
10 December 2024
You are already at the latest version
Abstract
RNA G-quadruplexes (rG4s) in 5'-UTRs represent complex regulatory elements capable of both inhibiting and activating mRNA translation through diverse mechanisms. This review analyzes the evolution of our understanding of 5'-UTR rG4-mediated translation regulation, from early discoveries of simple translation inhibitors to the current recognition of their multifaceted regulatory roles. We discuss canonical and non-canonical rG4 structures, their interactions with regulatory proteins including helicases and FMRP, and their function in both cap-dependent and IRES-mediated translation. Special attention is given to the synergistic effects between rG4s and upstream open reading frames (uORFs), stress-responsive translation regulation, and their role in repeat-associated non-AUG (RAN) translation linked to neurodegenerative diseases. We critically evaluate methodological challenges in the field, including limitations of current detection methods, reporter system artifacts, and the necessity to verify rG4 presence in endogenous transcripts. Recent technological advances, including genome editing and high-throughput sequencing approaches, have revealed that rG4 effects are more complex and context-dependent than initially thought. This review highlights the importance of developing more robust methodologies for studying rG4s at endogenous levels and carefully reevaluating previously identified targets, while emphasizing their potential as therapeutic targets in various diseases.
Keywords:
RNA G-quadruplex
; translation regulation
; IRES
; helicases
; FMRP
; 5’-UTR
1. Introduction
RNA molecules exhibit remarkable functional versatility in cellular regulation, orchestrating diverse processes including translational modulation and subcellular localization, thereby facilitating rapid cellular adaptation to environmental changes [1,2]. The application of advanced structural biology methodologies, in conjunction with high-throughput sequencing technologies, has revealed that this regulatory capacity is derived from RNA's exceptional structural plasticity and dynamic conformational landscapes [3]. The repertoire of structural motifs includes, but is not limited to, stem-loops, pseudoknots, triple helices, i-motifs and G-quadruplexes.
RNA G-quadruplex structures (rG4s) have been identified as a particularly significant regulatory element. These structures, preferentially formed in guanine-enriched sequences, are characterized by planar arrangements of four guanine residues (G-quartets) stabilized through Hoogsteen hydrogen bonding, with multiple G-quartets stacking to form the rG4 [4]. The structural integrity of these complexes is maintained through coordination with monovalent cations, particularly potassium, which occupy the central channel between stacked G-quartets. rG4s demonstrate enhanced thermodynamic stability and preferential parallel topology compared to their DNA counterparts due to conformational constraints and specific hydrogen bonding patterns imposed by the ribose 2'-hydroxyl moiety [5].
Although rG4s have been shown to exhibit robust stability under in cuvette conditions, the question of their intracellular formation remains unresolved. This necessitates the use of diverse analytical methodologies, including chemical probing, immunological detection and high-throughput sequencing approaches, in order to achieve comprehensive characterization [3]. Genome-wide methodologies including rG4-seq [6], utilizing reverse transcriptase stalling, and G4RP-seq [7], applying G4-specific ligand capture, suggest widespread rG4 formation, while DMS-seq [8] data suggested rG4s to be globally unfolded in eukaryotic cells. The formation of rG4s within cells was additionally supported by 19F NMR spectroscopy [9], immunofluorescence microscopy with a G4-specific antibody BG4 [10], and real-time visualization using rG4-specific fluorescent probe QUMA-1 [11]. These findings have led to the hypothesis that folded and unfolded rG4s exist in a dynamic equilibrium, subject to precise cellular regulation.
In 5'-UTR contexts, rG4s exhibit multiple regulatory functions, primarily modulating translation efficiency through interaction with scanning ribosomes [4,12]. Although initially regarded as mere impediments to scanning pre-initiation complexes, rG4s in 5'-UTRs are now acknowledged as sophisticated regulatory elements that orchestrate translation through intricate interactions with other regulatory elements, such as IRES and upstream open reading frames, and with rG4-binding proteins that modulate rG4 stability. These interactions facilitate dynamic modulation of translation in response to cellular signalling pathways and environmental cues. The latest research has led to significant updates in our understanding of rG4 structural diversity and biological significance. This review aims to examine the mechanisms of rG4-mediated translational regulation, evaluate methodological developments, and update the inventory of potential rG4-containing 5'-UTR regulatory elements affecting translation efficiency.
2. Translation-Inhibiting rG4s
2.1. Origins: First Identified Translation-Inhibiting rG4s
First translation-inhibiting rG4s in human mRNA were found in the 5'-UTRs of NRAS and Zic-1 transcripts. For NRAS, the rG4's influence on translation was demonstrated through in vitro translation assays using rabbit reticulocyte lysate [13]. The Zic-1 rG4 became the first target whose effect was demonstrated in cellulo using dual luciferase reporter assay (DLR) coupled with RT-qPCR to distinguish between translational and transcriptional effects [14].
The inhibitory effect of 5'-UTR rG4s on translation was initially attributed to steric hindrance created by the rG4 structure impeding RNA scanning by the preinitiation complex (PIC) from the cap to the start codon (Figure 1). Model systems using DLR assay established a correlation between rG4 thermodynamic stability and translation inhibition efficiency [15,16]. However, no consensus was reached regarding how rG4 positioning within the 5'-UTR affects the magnitude of repression. Investigation of rG4-binding proteins' influence on translation remained largely unexplored. A notable exception is Fragile X messenger ribonucleoprotein FMR1, which stabilizes rG4s and inhibits translation [17].
The discovery of regulatory rG4s in NRAS and Zic-1 5'-UTRs initiated a wave of reports between 2007-2012 identifying translation-inhibiting rG4s in the 5'-UTRs of biologically significant genes (Table 1). Most studies followed a common pipeline established in the pioneer publications: 1) bioinformatic identification of sequences in target mRNA 5'-UTRs with a potential to form rG4s; 2) investigation of corresponding oligonucleotide properties using CD and UV spectroscopy, along with reverse transcriptase (RT) stop, RNase T1 and in-line assays; 3) comparison of rG4-forming sequence versus mutant effects on translation efficiency using reporter systems, primarily through DLR assay supplemented by RT-qPCR, less frequently via in vitro translation or western blotting of transfected vector products. In some cases, researchers investigated the effects of G4 ligands on translation efficiency in reporter systems. For instance, G4 stabilizers 360A, Phen-DC3, and Phen-DC6 were shown to reduce translation efficiency by 15-90% in a concentration-dependent manner for TRF2 [18], while TMPyP4 attenuated translation inhibition by approximately 40% for metalloproteinase MT3-MMP [19].
However, most studies did not progress beyond reporter systems, and downstream effects of rG4-controlled translation on biological processes remained unexplored. Only for CCND3 and ADAM10 was the impact of rG4 mutation on protein overexpression levels and intracellular processes demonstrated. Flow cytometry and MTT assays revealed that CCND3 overexpression under wild-type (WT) rG4 control only marginally stimulated cell S-phase transition and division, whereas the vector with mutated rG4 significantly increased both S-phase cell proportion and division rate [21]. For ADAM10, vectors with mutated rG4 showed elevated levels of APPsα, the primary protein activity product, in the supernatant [24].
Even in these cases, the effect was not confirmed at the endogenous mRNA level. Moreover, for rG4 in the 5'-UTR of Bcl-2, a known anti-tumor target and apoptosis inhibitor, CRISPR-Cas9 deletion of the rG4-forming sequence from melanoma cell genome caused no changes in mRNA levels, protein levels, or response to apoptosis-activating compounds compared to unedited cells [23]. This led to the conclusion that at the endogenous mRNA level, rG4 presence does not significantly affect translation, and effects observed in the reporter assays may result from oversaturation of the putative rG4 helicase activity. This same issue is known in in vitro translation assays: for Bcl-2, no rG4 inhibitory effect was observed at 20 ng/μL plasmid concentration, but translation inhibition through rG4 became detectable (2-fold) at 320 ng/μL [22]. However, the study on Bcl-2 rG4 with CRISPR-Cas9 had limitations due to choosing an rG4 with relatively weak effects. In this study, DLR assays in HEK293 cells showed only a 20% restoration of translation efficiency upon deletion of the G4-forming sequence, and a comparable change might have been missed in subsequent CRISPR-Cas9 tests due to significant experimental error. While this work undermines the prospects of Bcl-2 rG4 as an anti-tumor therapy target, it raised important questions about the adequacy of primary DLR and in vitro translation methods for assessing rG4's influence on translation efficiency.
The mechanism of rG4-mediated translation inhibition remained largely unexplored, primarily reduced to the hypothesis of steric hindrance created by rG4s impeding scanning PIC. Except for rG4-resolving helicases and FMRP, only for MT3-MMP and ARPC2 mRNAs rG4s were protein partners later identified [28]. Among these were small ribosomal subunit proteins, generating a hypothesis about PIC scanning deceleration due to its proteins' affinity for 5'-UTR rG4s.
In many cases, researchers overlooked the critical step of verifying expression levels of mRNA containing rG4-forming sequences. It is well established that determining mRNA 5'-ends presents significant challenges, and mRNA sequence databases undergo constant refinement as methodology improves [29,30]. Furthermore, cell line-specific and condition-dependent variations in promoter activation and splicing patterns can alter target mRNA levels. Nevertheless, only for estrogen receptor 1 (ESR1) was the presence of rG4-forming sequences in cellular mRNAs confirmed by qRT-PCR, albeit with caveats regarding method sensitivity [20]. Surprisingly, for NRAS rG4, which served as a control in numerous studies, near-complete absence of mRNAs containing the cap-proximal rG4-forming sequence was demonstrated across 14 cell lines [31]. According to an updated NCBI annotation, NRAS mRNA contains a 131nt-long 5'-UTR, while the rG4 was previously reported 222 nt upstream of the translation start site (TSS). The conducted experiments, including qRT-PCR across 14 cell lines, rapid amplification of cDNA 5'-ends (5'-RACE) on HEK293T cells, and cap analysis of gene expression (CAGE)-Seq data confirmed that most NRAS mRNAs lack the rG4-forming sequence. In 2016, new rG4s were discovered in NRAS 5'-UTR using the rG4-specific ligand RGB-1; however, these rG4 sequences are also absent in the current annotation [32]. Thus, critical revision of other known rG4 targets is recommended for each tissue type/cell line, as may also be absent in cellular mRNAs.
Additionally, many 5'-UTR rG4-forming sequence identifications may have been erroneous or incomplete, as contemporary G4 predictors relied on canonical motifs, while many rG4s possess non-canonical structures with long loops, bulges, two G-tetrads, etc. (Figure 2) [33]. Therefore, investigation of most rG4 targets subsequently evolved toward: 1) searching for non-canonical rG4s; 2) identifying rG4s whose biological role was supported by genome editing and helicase inhibition experiments; 3) discovering new mechanisms of rG4-mediated translation regulation in 5'-UTRs.
2.2. Non-Canonical rG4s Inhibiting Translation
In previously discussed examples, rG4s were identified using bioinformatic methods employing the classical motif G3+-N1-7-G3+-N1-7-G3+-N1-7-G3+ (Figure 2A). However, DHX-36, a G4-unwinding helicase, was shown to predominantly target non-canonical rG4s containing two tetrads and bulges in the 5'-UTRs of transcripts of myoblast cells [34]. Identifying such non-canonical rG4s remained methodologically challenging. In 2010, it was demonstrated that rG4s with loop lengths up to 30 nucleotides could fold in cuvette (Figure 2B) [35]. J.P. Perreault’s group was among the first to address this, conducting bioinformatic searches in 5'-UTR databases for rG4-forming motifs with two single-nucleotide loops and one long central loop up to 90 nucleotides. They successfully identified rG4s inhibiting translation of therapeutically significant HIRA, TOM1L2, and APC mRNAs by 90%, 75%, and 40% respectively, according to DLR assay data [36]. Analysis via in-line probing methodology revealed that these rG4 structures possessed loop regions spanning from 11 to 32 nucleotides, whereby the assay provided detailed insights into the specific nucleotide contributions to structural assembly.
Since the study revealed that G4s with loops longer than 30 nucleotides do not form regulatory sequences in cells (although they fold in cuvette), subsequent work employed a modified motif G3-N1-G3-N1-20-G3-N1-G3 [36,37]. This approach, combined with the classical motif, identified three additional rG4 targets involved in intestinal cancer development and inhibiting cap-dependent translation by 2-4 fold: BAG-1, MAPK3, and CASP8AP2. Analysis of in-line cleavage patterns revealed that a subset of G-rich sequences demonstrated the capacity to form rG4 structures featuring loops that exceed the conventional 7 nt length. Thus, this approach significantly expanded the repertoire of known rG4 regulators and identified several targets for anti-tumor therapy.
Non-canonical rG4s were also discovered during investigation of the effects of G4-stabilizer RGB-1 on oncogene mRNA translation efficiency [38]. To identify targets whose translation, but not transcription, depends on RGB-1, western blotting and qRT-PCR data were compared before and after RGB-1 cell treatment. This approach identified two targets, Nectin-4 and CapG. Nectin-4 regulates Ca2+-independent cellular adhesion and cancer proliferation [39]. CapG enhances cell motility in nasopharyngeal carcinoma by promoting MLC2 phosphorylation, regulates NF-κB signaling in acute myeloid leukemia, and accelerates proliferation and invasion in diffuse large B-cell lymphoma through the PI3K/AKT pathway, highlighting its multifaceted role in cancer progression [40,41,42,43]. Although canonical rG4 motifs were present in 5'-UTRs of both mRNAs', reverse transcriptase stop assay (RT-stop) revealed that rG4 formation in both cases ignores the fourth adjacent G-tract, instead incorporating a G-tract located more than seven nucleotides away. Moreover, for CapG, the equilibrium between canonical/non-canonical rG4s was shown to depend on potassium concentration. At near-physiological potassium concentration (100 mM), a non-canonical rG4 forms that can inhibit translation by 50%. At lower potassium concentration (25 mM), rG4 formation results in only 20% translation inhibition. The authors suggested this regulating mechanism of the translation efficiency in response to changes in potassium concentrations might have biological significance.
Besides RT-stop and in-line assays, which identify residues forming rG4s, DLR test can track translation inhibition efficiency depending on sequence mutations. This approach helped to establish the structural ensemble of highly polymorphic rG4s in the 5'-UTR of mRNA that codes VWF protein, an antihemophilic factor carrier and platelet-vessel wall mediator in blood coagulation crucial for hemostasis, whose deficiency leads to von Willebrand disease [44,45]. Its 5'-UTR contains three GGG tracts, flanked by several GG tracts and isolated Gs at both 5'- and 3'-ends. When G-to-A mutations were introduced into the G-rich sequence at the 5'-end, the inhibitory effect on translation increased compared to WT rG4, suggesting formation of a more homogeneous and stable structure by the remaining guanines and indicating minimal contribution of 5'-terminal guanines to the major inhibitory structure. Conversely, mutation of most guanines at 3'-end attenuated the inhibitory effect, likely due to rG4 structure destabilization. Analysis revealed that G38 and G45 likely participate in the formation of most ensemble structures, while the intervening G residue between them can vary (although G42 is predominantly involved), with bulges forming in all cases (Figure 2C). While this system clearly exceeds the resolution of employed methods and requires confirmation by NMR or X-ray crystallography, this study raises questions about expanding bioinformatic rG4 search motifs to include bulge-containing patterns. Thus, the approach significantly expanded the repertoire of known rG4 translation inhibitors and identified several anti-tumor therapy targets.
2.3. Involvement of rG4-Binding Proteins in Translation Inhibition
2.3.1. Investigation of Inhibitory rG4s in Context of Their Helicase Interactions
As the field developed, it became possible to study rG4's influence on translation in the context of rG4-binding protein interactions. DHX36 (also known as RHAU, G4R1) is of particular interest as a helicase with high rG4 affinity and predominantly cytoplasmic localization, capable of unwinding rG4s and thereby regulating translation (Figure 3) [46]. DHX36 plays a special role during active cell proliferation, including embryonic development and injury recovery, consequently DHX36 expression decreases during embryonic and stem cell differentiation [7,34,47]. In myoblasts, DHX36 targets, most of which contain rG4-forming motifs, were shown to have the highest density in 5'-UTRs, and the helicase knockdown primarily decreased translation efficiency of mRNAs containing 5'-UTR rG4s [34]. Thus, DHX36 can enhance translation efficiency of rG4-containing mRNAs in rapidly proliferating cells, and its knockdown/overexpression can serve as a tool for identifying rG4 targets, including non-canonical ones.
Mice with conditional DHX36 knockout in myogenic cells showed smaller fibers, limb muscle weakness, and significantly reduced regeneration efficiency after injury, attributed to decreased rate of cell proliferation [34]. Search for downstream targets of DHX36 knockout whose translation increases without mRNA level changes in myoblasts using cross-linking and immunoprecipitation (CLIP)-seq and polysome profiling coupled with RNA-seq identified mRNA encoding G protein subunit alpha i2 (GNAI2). This protein participates in hormonal regulation of adenylate cyclase and regulates cell proliferation, muscle size, and regeneration. The 5'-UTR of GNAI2 mRNA contains a G-rich conservative sequence whose rG4-forming ability was confirmed by probing with fluorogenic dyes NMM and ThT along with RT-stop assay. RNA pull-down revealed the 5'-UTR association with DHX36, and DHX36 antibody effectively retrieved GNAI2 mRNA via RNA immunoprecipitation (RIP). GNAI2 protein levels were significantly reduced without mRNA level changes in DHX36-knockout stem cells and myoblasts, with effects enhanced by the treatment with rG4-stabilizer cPDS, alongside decreased proliferation rate. Insertion of full-length 5'-UTR and G4-forming sequences into a plasmid vector upstream of EGFP-coding sequence downregulated translation compared to non-G4-forming mutants according to western blotting data. The inhibitory effect of the rG4s was more pronounced in DHX36 knockout cells. Moreover, DHX36 plasmid overexpression increased GNAI2 protein levels and rescued SC proliferation by 30.8%. Therefore, GNA2I rG4s can be considered a promising target with potential for improving injured muscle recovery efficiency.
DHX36 deletion also disrupts heart development in mouse embryos, leading to death at day E7.5 due to cardiomyocyte proliferation impairment [47]. RNA microarray analysis, northern blotting, qRT-PCR, and whole-mount in situ hybridization (WISH) revealed increased levels of Nkx2-5 mRNA. However, protein levels, according to western blotting analysis and immunofluorescence (IF) staining, decreased with DHX36 knockout. Investigation of this phenomenon using RIP and RNA pull-down assay showed that DHX36 could form complexes with both 5'- and 3'-UTR of Nkx2-5 mRNA. When binding to 3'-UTR, DHX36 accelerates mRNA degradation. The 5'-UTR of Nkx2-5 mRNA contains a highly conserved G-rich sequence capable of forming rG4. Nkx2-5 5'-UTR sequence, when inserted into a GFP expression vector, inhibited its translation. In the presence of DHX36, the GFP levels significantly increased, indicating improved translation efficiency due to unfolding of the inhibitory rG4. Inserting this 5’-UTR with the WT rG4 and corresponding 6G-A mutant into a Nkx2-5 expression vector increased translation efficiency according to western blotting data. A DLR assay showed that DHX36 knockdown reduced translation efficiency of Nkx2-5 5’-UTR containing mRNA by 49%. An addition of siDHX36 also decreased the proportion of the mRNA in the polysomal fraction. Notably, Nkx2-5 rG4 contains five G-tracts and must include a 15-nucleotide loop in any assembly, indicating its non-canonical structure.
Based on rG4-seq data, an rG4 target was found in the 5'-UTR of ADAR1 mRNA [6]. ADAR protein is an adenosine deaminase acting on RNA, proceeding with adenosine to inosine substitution and triggering translation frameshift and codon substitution in the open reading frame region, affecting alternative splicing, translation efficacy, miRNA processing, and RNA stability in UTRs [48,49]. RT-stop and selective hydroxyl acylation analyzed by lithium-ion mediated primer extension (SHALiPE) assays were used to precisely determine nucleotides participating in structure formation [50]. In the presence of DHX36, the rG4-forming oligoribonucleotide showed greater tendency to form duplexes with complementary trap than intramolecular rG4, according to native electrophoresis data. The ability to form rG4 in cellulo was supported by confocal microscopy: upon ADAR 5’UTR RNA transfection, cell fixation and permeabilization, colocalization was observed between FAM-adar and ISCH-adar oligonucleotide probes complementary to rG4 flanking sequences and fluorescently labeled with FAM and G4-specific light-up moiety respectively. When repeating the experiment in DHX36-knockout cells, the rG4 light-up/FAM signal ratio increased 2-fold compared to intact cells, indicating increased stable rG4 proportion. According to DLR assay, the rG4 inserted in 5'-UTR of the Renilla luciferase gene reduced translation efficiency by 2.7-fold or 4.3-fold, respectively, compared to rG4 mutant. In the former case, treatment with the G4 stabilizer PDS led to an additional 3.0-fold decrease in translation efficiency. When conducting the test under DHX36 knockdown and knockout conditions, the translation efficiency of mRNA bearing WT rG4 was decreased additionally by 1.6 and 2.9-fold compared to intact DHX36 levels. The rG4's influence on translation was also confirmed by western blotting using vectors containing ADAR1 5'-UTR and coding sequence (CDS), with translation efficiency decreasing in DHX36 knockout cells. Thus, the ADAR1 rG4 study demonstrated DHX36 level influence on stable rG4 proportion.
The 5'-UTR mRNA of the TWIK-related acid-sensitive K+ channel (Task3) was also shown to have a regulatory rG4 [51]. Task3 dysfunction and knockdown leads to impaired neuronal migration of cerebral cortical neurons in developing murine brain and maternally imprinted intellectual disability Birk-Barel mental retardation. The repeat (GGN)13, due to its length and polymorphic nature, represents a complex object for investigation, including control selection. Therefore, the authors proposed using GC-scrambled or complementary sequences as controls, along with comparing CD spectra before and after DMS treatment. RIP with G4-binding BG4 antibody confirmed intracellular Task3 rG4 formation. Western-blotting of 3xFLAG-tagged products of reporter constructs possessing WT or mutated regions upstream of Task-M1 CDS revealed the translation inhibition for WT construct by 3-fold compared to the reporter containing a GC-scrambled sequence of identical length. Following cellular transfection with a construct encoding N-terminally FLAG-tagged full-length DHX36, FLAG immunoprecipitation experiments validated the molecular interaction between DHX36 and Task3 mRNA. DHX36 overexpression increased translation efficiency of the Task3-M1 coding reporter construct by 2-fold and endogenous Task3 expression by 1.25-fold, increasing cell hyperpolarization in the second case. Thus, Task3 5'-UTR rG4, which mediates Task3 production affecting cell hyperpolarization and whose existence in endogenous mRNA was confirmed by 5'-RACE , represents an attractive target for developing therapeutic agents against Birk-Barel mental retardation.
Recently, an rG4 was identified in PBX1 mRNA 5'-UTR [52]. PBX1 is a transcription factor whose overexpression is linked to pre-B-cell acute lymphoblastic leukemia and melanoma development. Using a GFP-expressing reporter construct, PBX1 rG4 mutation was shown to improve translation efficiency by approximately 20%. Furthermore, G4 ligands PDS and TMPyP4 further inhibited translation to 25% of WT level. Additionally, helicase DHX9 was demonstrated to regulate PBX1 mRNA translation by resolving the rG4 structure. PBX1-rG4 edited cell lines showed no dependence on rG4-stabilizing compounds, and immunohistochemistry of normal and tumor tissues revealed that DHX9 absence corresponded to PBX1 absence. Notably, DHX9 knockdown in melanoma reduced tumor progression.
In hepatitis B virus (HBV)-related hepatocellular carcinoma context, RNA helicase DDX5 was known to be downregulated during HBV replication, correlating with poor prognosis. Recent research demonstrated DDX5 facilitates STAT1 transcription factor translation; specifically, siRNA-mediated DDX5 knockdown reduced STAT1 protein levels without affecting STAT1 mRNA levels or splicing. Transcriptome-wide rG4-seq revealed that the 5'-UTR of STAT1 mRNA exhibits significant potential for rG4 formation [6]. Three G-rich sequences potentially capable of forming rG4 were discovered in STAT1 5'-UTR mRNA, with the cap-proximal sequence's influence on translation thoroughly investigated [53]. According to DLR results, the mutation of the cap-proximal rG4 increased translation efficiency by more than 2-fold. Simultaneously, both the treatment with rG4-stabilizer (Phen-DC3 or PDS) and DDX5 knockdown decreased translation efficiency in the reporter system without significantly affecting the rG4-mutated vector. This indicated rG4's inhibitory effect on STAT1 mRNA translation, whose degree depends on DDX5 helicase activity. To obtain data about effects on endogenous mRNA levels, CRISPR-Cas9 was used to delete the rG4-forming sequence from the STAT1 gene. Cells with biallelic deletion showed significant and reproducible increase in STAT1 protein levels compared to unedited cells, while STAT1 mRNA levels remained unchanged. DDX5 knockdown and treatment with Phen-DC3 and PDS had no effect on STAT1 protein levels in biallelic cells but reduced them in WT cells. DDX5 and STAT1 mRNA rG4 interaction was also confirmed by RIP, RNA pull-down, and EMSA methods. The study also showed that HBV replication reduces DDX5 levels and STAT1 levels, clinically significant for interferon therapy. Besides therapeutic significance, this case provided the first example of rG4 whose influence at endogenous mRNA level was confirmed by genome editing methods, and drew attention to helicase DDX5, which was less known than DHX36 and DHX9.
Thus, helicases DHX36, DHX9, and DDX5 significantly regulate the degree of rG4-mediated translation inhibition. This is important from two perspectives. First, it opens therapeutic horizons for controlling rG4-regulated mRNA translation through helicase activity and, conversely, for increasing translation efficiency by G4-specific disruptors [54,55,56] under helicase deficiency, for example, during HBV infection. Second, helicase interaction-based methods enable more effective rG4 studies, including non-canonical and in cellulo forms.
2.3.2. FMRP: rG4-Binding Protein Inhibiting Translation
The FMR1 locus encodes Fragile X Mental Retardation Protein (FMRP), a neuronally-enriched RNA-binding protein whose dysfunction through compromised transcription or translation leads to neurodevelopmental disorders, including Fragile X Syndrome (FXS) and Fragile X-associated tremor/ataxia syndrome (FXTAS) [57,58]. The pathogenesis of these conditions is attributed to dysregulation of FMRP-dependent gene expression networks, specifically perturbations in target mRNA translation and subcellular localization. FMRP's molecular architecture includes three distinct RNA-binding modules: RGG, KH1, and KH2 domains. The RGG domain exhibits particular specificity for rG4-forming sequences [59]. Biochemical analyses demonstrate that FMRP-rG4 interactions result in translational repression, as evidenced by luciferase-based reporter systems [60]. The mechanism appears to involve both rG4 stabilization and interference with ribosomal scanning. Additionally, FMRP mediates cap-dependent translational inhibition through CYFIP1 recruitment/stabilization, which modulates eIF4E activity. While FMRP predominantly associates with rG4 elements within coding sequences and 3'-UTRs - including an autoregulatory interaction within its own coding region - at least three characterized mRNAs are regulated through 5'-UTR rG4 interactions.
A notable example is MAP1B (microtubule-associated protein 1B), whose expression FMRP downregulates during developmental synaptogenesis. In FMR1-deficient conditions or knockout models, MAP1B overexpression leads to aberrant microtubule stabilization and impaired dendritic spine development [40]. Structural and biochemical characterization of the MAP1B 5'-UTR revealed a G-rich sequence capable of forming rG4, confirmed through CD and NMR spectroscopy [61]. The RGG domain of FMRP exhibits nanomolar affinity for this structure, primarily through hydrophobic interactions. Notably, the FMRP:mRNA stoichiometry influences rG4 stability: low ratios promote structure formation while higher ratios induce destabilization, suggesting a potential activity-dependent translational control mechanism during neuronal stimulation (Figure 4).
Studies have demonstrated that FMRP functions as a translational repressor of protein phosphatase 2A catalytic subunit (PP2Ac) mRNA [62]. In FMRP-deficient conditions, elevated PP2Ac expression disrupts cytoskeletal dynamics, specifically affecting dendritic spine morphogenesis in FXS patients. Biochemical analyses have revealed that FMRP exhibits nanomolar binding affinity for the PP2Ac mRNA 5'-UTR, which harbors four distinct rG4s - two exhibiting high stability and two with lower stability, as determined by RT stop assays. Detailed structural characterization of one particular rG4, positioned four nucleotides upstream of the translation initiation site, was accomplished through CD spectroscopy and RNase T1 footprinting, enabling precise identification of nucleotides involved in rG4 formation [63]. Functional analyses using dual luciferase reporter assays demonstrated this rG4's capacity to suppress translation by 70% relative to its mutant form.
Further investigations identified an additional rG4-forming sequence at position 163 within the 333-nucleotide 5'-UTR of SMNDC1 mRNA [64]. SMNDC1, a splicing regulatory factor expressed in muscle tissue and neural structures, whose elevated expression correlates with apoptotic activation, was identified as a potential FMRP target through PAR-CLIP analysis. Biophysical characterization using CD spectroscopy, NMR analysis, and UV melting studies confirmed the formation of a physiologically stable three-tetrad rG4. Electrophoretic mobility shift assays demonstrated high-affinity, selective binding of FMRP's RGG domain to this rG4, corroborated by RNA pull-down experiments. While direct evidence for rG4-mediated translational regulation of SMNDC1 by FMRP remains to be established, the observed interactions suggest a potential regulatory mechanism.
Additionally, while a rG4-forming sequence was identified in hASH1 mRNA 5'-UTR, comprehensive analysis using multiple experimental approaches revealed that a U10 uridine tract, rather than the rG4, serves as the primary regulatory element [65].
The accumulated experimental evidence indicates that rG4s exhibit regulatory capacity in translational control not only through creating structural impediments to scanning PIC but also serving as recognition elements for regulatory trans-acting factors, particularly exemplified by FMRP-mediated translation regulation. However, comprehensive mechanistic investigations, employing methodological approaches similar to those utilized in hASH1 mRNA characterization, are required to establish detailed molecular mechanisms underlying rG4-dependent FMRP-mediated translational control in the identified regulatory systems.
3. rG4s Inhibiting Translation Through Interaction with uORF
3.1. Synergistic Translation Inhibition by rG4s and Upstream Open Reading Frames
The inhibitory effect of rG4s on translation has been studied repeatedly at the transcriptome-wide level. A recent study focused on the relationship between upstream open reading frames (uORFs) and rG4s in inhibiting main reading frame translation [66].
Transcriptome-wide ribosomal profiling of cells treated with a translation elongation inhibitor (cycloheximide) was coupled with matched transcriptional mRNA sequencing and calculation of length-corrected minimum free energies of folded RNA secondary structures in 5'-UTRs to observe the relationship between translation efficiency and secondary structure stability. Among mRNAs with rG4-regulated translation, 80S ribosome presence in 5'-UTR was observed more frequently than average, while less frequently in CDS, which indicated more active uORF translation. The distribution of ribosomal footprints was non-random: 1) more actively translated uORFs corresponded to more stable rG4s; 2) rG4s were positioned downstream of uORF start codons in a way that generated queues of periodically scanning ribosomes. rG4s located downstream of uORF start codons force PICs to remain longer in the uORF start codon region, thereby increasing the probability of uORF translation initiation while inhibiting main reading frame translation (Figure 5).
In the same study [66], the role of rG4-unwinding helicases in regulating translation of mRNAs containing uORF and downstream rG4 was investigated. Polysome profiling coupled with proteomics mass spectrometry was employed. Data showed DHX9 and DHX36 helicases to concentrate in the polysomal fraction, so their mRNA binding corresponds to active translation. Ribosome profiling was subsequently repeated on cells with DHX9 and DHX36 knockdown. This revealed an additional increase and a decrease in uORF and main reading frame translation efficiency, respectively. Such mRNAs also contained more stable rG4s. Translation efficiency changes correlated for both helicases, indicating a shared target pool. Analysis of helicase-dependent translation revealed significant enrichment of genes encoding of transcription factors (e.g., STAT6 or FOXM1), epigenetic regulators (e.g., SUZ12, MLL1, or MLL5), and kinases (e.g., MAPK3, MAP2K1, or CDC42BPB). Individual-nucleotide resolution UV crosslinking and immunoprecipitation (iCLIP) method was used to map DHX9 binding sites to determine whether DHX9 binds mRNAs directly via rG4s. The distribution of ribosomes on transcripts with DHX9 iCLIP peaks in 5'-UTRs showed enrichment of ribosome-protected fragments upstream of rG4-containing DHX9 binding sites in 5'-UTRs. When DHX9 was depleted, ribosome occupancy in the 5'-UTR increased, while it decreased in the downstream coding sequence. These findings indicate that DHX9 regulates ribosome distribution and translation efficiency by directly binding to its rG4 substrate.
To further verify the roles of DHX9/DHX36 and their targets, GFP expression vectors with 5’-UTRs containing either an rG4, a short rG4-containing uORF, or a mutated rG4/alternative translation initiation site were constructed, using DDX23 5'-UTR as a template. The rG4 from 5′-UTR of DDX23 mRNA is characterized to be bound in cells by DHX9 and control its translation efficiency in a DHX36- and DHX9-dependent manner. Comparing GFP expression with an rG4 versus a mutated rG4 revealed minimal impact from the rG4. Further tests showed that an alternative translation initiation site moderately influences expression. Interestingly, comparing the expression of the reporter gene with a 5'-UTR containing an alternative translation initiation site and a downstream rG4 to one where only the rG4 was mutated revealed that the rG4 enhances the uORF's repressive effect. Furthermore, the depletion of DHX36 or DHX9 resulted in a reduction in GFP expression in the context of an rG4-containing uORF, whereas no effect was observed in the analogue with a mutated rG4. The data demonstrated that the repressive effect of the uORF was DHX36- and DHX9-dependent, and observed only in the presence of rG4. Thus, this study discovered and showed synergy between uORF and downstream rG4 in inhibiting main reading frame translation.
3.2. FMR1 mRNA and Repeat-Associated Non-AUG Translation
Secondary structures can also cause translation from non-AUG start codons and inhibit cap-dependent translation of the main open reading frame [67]. This is especially characteristic of microsatellite repeats associated with the development of neurodegenerative disease and the synthesis of aggregation-prone toxic peptides and is called repeat-associated non-AUG (RAN) translation (Figure 6) [68]. A proposed mechanism for amyotrophic lateral sclerosis (ALS) and frontotemporal dementia (FTD) is the RAN translation of the r(G4C2)n hexanucleotide repeat in C9Orf72 pre-mRNA, causing accumulation of toxic polyPR/polyGR dipeptide repeats [68,69]. An additional example is FXTAS, where affected individuals possess 55-200 repeats of the r(GGC)n sequence in FMR1 mRNA 5'-UTR, leading to production of toxic polyG, polyR, and polyA, as well as chimeric peptides [49,50]. Importantly, when 200-2000 repeats accumulate, transcript production halts due to methylation of CpG islands in this GC-rich region, resulting in FXS, which is also associated with intellectual disability from FMR1 protein deficiency.
The inhibition of translation from the main open reading frame and initiation of non-canonical RAN translation likely result from the formation of a highly structured RNA region due to repeat expansion [68]. The PIC assembles on the cap structure and scans the 5'-UTR for a start codon, but secondary structures encountered during this process impede PIC movement, reducing translation efficiency. This slowdown increases chances of near-cognate non-AUG codons being recognized as start codons, thereby triggering abnormal repeat translation [68]. Specifically, the mechanism is enhanced by displacement of eIF1 protein from PIC by eIF5, which is crucial for accurate start codon recognition and maintaining PIC's open scanning conformation [67]. Additionally, these structures can induce a frameshift in the repeat region after non-canonical translation begins, resulting in chimeric peptides that are more toxic and aggregation-prone compared to their homopeptide equivalents [70]. RAN translation also leads to sequestration of ribosomes and translation factors, necessitating specific corrective enzymes for their release.
The r(GGC)n and r(G4C2)n repeats can form both hairpins and rG4s, but there is no consensus in the literature about which secondary structure serves as a more relevant biological target for regulating translation in ALS, FTD, and FXTAS [71,72,73,74]. Proteins such as hnRNP A2 and CBF-A, which can unwind rG4s, have been shown to alleviate the inhibitory effect of FMR1's structured 5'-UTR on main open reading frame translation, though these rG4-unwinding proteins lack structural specificity [72]. The rG4-destabilizing ligand TMPyP4 and lessens translation inhibition [74]. Replacing of the r(GGC)n motif with r(GGN)n still allows polyG production but inhibits hairpin formation without significantly affecting rG4 assembly, resulting in notable reduction in peptides' toxicity and ribosome trapping [69,70]. This suggests hairpin structures contribute significantly to the negative effects of RAN translation. As far as r(G4C2)n from C9Orf72 is concerned, support for the hairpin theory arises from several observations: 1) the hairpin form is more kinetically favored and becomes more preferable as repeat length increases; 2) RAN translation is less efficient in samples folded in potassium-rich buffer favoring rG4 stabilization; 3) impact of G4-ligands on RAN translation is less pronounced than that of hairpin-interacting agents [71]. Furthermore, the inherent complexity of these repetitive sequences complicates determination of their precise structure and whether that structure is consistent throughout the sequence, making it challenging to fully comprehend potential implications of their three-dimensional conformation and dynamic variations. Ultimately, while establishing secondary structures of GC-rich repeats remains uncertain, this interplay between canonical and RAN translation exemplifies secondary structures' regulatory role, offering promising avenues for therapeutic applications in neurodegenerative diseases.
4. rG4s Enhancing Translation
4.1. rG4s Activating Translation as IRES Components
Until now, the role of rG4s in regulation of cap-dependent translation has been discussed. However, rG4s can also mediate cap-independent translation, acting as elements of internal ribosome entry sites (IRES) (Figure 7) [2,4]. The spatial structure of IRES allows recruitment of the 40S ribosomal subunit, an incomplete set of translation initiation factors, and IRES trans-acting factors (ITAFs) for translation initiation without the need for cap recognition [75]. The function of some smaller IRES elements, although lacking rG4, in eukaryotic cells has been described as mediating interactions with 18S rRNA in a Shine-Dalgarno-like manner, which is actually known in prokaryotes [76]. Subsequent investigation demonstrated that rG4s are able to directly interact with the 40S ribosomal subunit in the absence of other protein factors [77]. Most frequently, rG4s in the IRES region simultaneously inhibit cap-dependent translation and activate cap-independent translation, the engagement of which becomes necessary under stress conditions during the repression of canonical translation [28,37,78,79].
The first rG4 as IRES component that enhances cap-independent translation efficiency was found in FGF2 mRNA transcript [80]. FGF-2 protein is involved in cell growth, migration, and differentiation, influencing various systems such as cardiovascular and central nervous systems, and bone physiology; its dysfunction is linked to diseases like cardiovascular disorders, osteoporosis, and neurodegenerative conditions [81]. The IRES that controls translation from four out of five start codons is localized in the first 176 nucleotides of FGF2 mRNA 5'-UTR and contains two hairpins and a rG4 between them. Interestingly, rG4 represents an antiparallel five-layered chair-type structure with unusual central single-nucleotide and two long loops. Since the main IRES element is the second hairpin's loop, rG4 rather plays a supporting role and cannot ensure alone efficient cap-independent translation, providing no more than 20% of IRES activity.
The second known rG4 translation activator was discovered in IRES-A of VEGF mRNA 5'-UTR. Despite VEGF rG4's widespread recognition, its role in translation regulation remains unclear. Human VEGF is a key physiological and pathological angiogenic growth factor. In 2010, a G-rich sequence capable of forming a wide array of two-layered rG4s was shown to exist in VEGF mRNA IRES-A [82]. The introduction of four G-U mutations that prevents rG4 formation almost completely eliminated IRES-A activity, suggesting rG4 was absolutely necessary for IRES-A-mediated cap-independent translation. Through various combinations of two G-U mutations limiting possible rG4 arrangements, translation efficiency could be either significantly decreased, or slightly increased. This led to the concept of a switchable rG4 motif that could provide fine-tuning of cap-independent translation efficiency through adopting different rG4 structures. Later, another research group showed that the 40S ribosomal subunit has high affinity for this G-rich motif, enabling IRES-A function; although whether the effect was structure- or sequence-specific was not specified [77]. While four G-U mutations did reduce translation efficiency, G-A and G-C mutations at the same positions, despite preventing rG4 formation, resulted in translation efficiency nearly identical to the wild-type (WT) sequence [83]. The insertion of a more stable three-layered rG4 from NRAS mRNA 5'-UTR and the treatment of the WT system with G4-stabilizing ligands decreased cap-independent translation efficiency. Moreover, exposure of cells to G4-stabilizing ligand Phen-DC6 decreased the ratio of polysomal to non-polysomal VEGF mRNA, further supporting rG4's inhibitory role in IRES-dependent VEGF mRNA translation. Importantly, G-U mutations introduced an AUG codon absent in WT mRNA, creating a small open reading frame that could explain decreased translation efficiency upon rG4 disruption [77,82,83]. However, mutants with two and four G-U mutations both contained this AUG codon, but only the second caused decreased translation efficiency, while the first had no effect [82]. This might indicate that neither rG4 disruption nor uORF appearance caused translation efficiency reduction with G-U mutations. In summary: 1) rG4 is not necessary for VEGF IRES-A activity; 2) rG4 inhibits VEGF translation when stabilized by G4-ligands and may represent a therapeutic target; 3) the reasons for the decrease in translation efficiency with four G-U mutations remains unknown, but its determination could illuminate IRES-A functioning. Later a new IRES-A activity regulation mechanism was discovered: A856 methylation, which could deactivate endogenous uORF start codon in IRES-A and recruit YTHDC2/eIF4GI complex to trigger cap-independent translation [84]. Interestingly, knockdown of METTL3 encoding N6-adenosine-methyltransferase 70 kDa subunit reduced the effect of G4-stabilizer 360A, and the treatment with 360A decreased METTL3 knockdown's impact, though both effects remained statistically significant. A connection between G-rich region mutations and A856 methylation might be worth investigating to establish the mechanism of VEGF cap-independent translation regulation through IRES-A.
BAG-1 mRNA translation regulation is particularly interesting. As mentioned earlier, BAG-1 5'-UTR rG4 mutation led to a 2.4-4.0-fold increase in cap-dependent translation efficiency depending on a cell line in DLR assays. However, BAG-1 translation regulation is quite complex: besides cap-proximal rG4, the 5'-end contains four start codons, including the first non-canonical CUG, as well as IRES before the third codon and uORF [78]. The first three start codons encode protein isoforms differing in N-terminal length and intracellular localization. DLR assay established that rG4 inhibits translation of all three protein isoforms and uORF, exerting general suppression on cap-dependent translation. The inhibitory effect intensified when cells were treated with G4 stabilizers cPDS and Phen-DC3. Despite proximity to cap structure, the effect is not related to disruption of cap-dependent synthesis or loss of affinity to translation initiation factor eIF4E. Meanwhile, cap-independent translation showed a 20% decrease in efficiency when rG4 was mutated. After unveiling the spatial structure of 5'-UTR with intact or mutated rG4 using SHAPE method, IRES with mutated rG4 was shown to adopt more rigid structure and remains in closed conformation, presumably preventing interaction with previously established ITAFs PTB-1 and PCBP1, thereby inhibiting translation. Thus, BAG-1 provides an excellent example of rG4's distal influence on IRES functionality, unlike most cases where rG4 is its constituent element.
Similarly, rG4 in the 5'-UTR of actin-related protein 2 (ARPC2) mRNA regulates translation. ARPC2 is part of the ARP2/3 protein complex, participating in actin cytoskeleton branching and facilitating cell migration. The rG4 inhibits cap-dependent translation by 40% according to DLR assay and can bind to ribosomal proteins within the 43S pre-initiation complex [28]. Complex formation with rG4, rather than steric blocking alone, might be responsible for translation inhibition. Nevertheless, ARPC2 mRNA can also be translated cap-independently due to IRES presence. The study established its structure, with rG4 positioned in a hairpin loop within a cruciform structure [79]. Mutational destabilization of the rG4 resulted in a 30% reduction in cap-independent translation efficiency. However, determining the exact cause of this effect proved challenging: it could be due to either IRES structure changes or inadvertent creation of new ITAF binding sites through mutations. Notably, ARPC2 cap-independent translation efficiency increases under stress conditions at high cell density, suggesting possible connection between rG4-mediated regulation and cellular stress.
There exists another example of rG4 participation in cap-independent translation with its increasing influence under stress conditions. NRF2 functions as a key transcription factor that induces antioxidant and detoxification gene expression, providing cytoprotection to multiple organ systems [85]. Its translation efficiency increases under oxidative stress conditions, as demonstrated during H2O2 treatment both at the endogenous protein level and in a bicistronic reporter system. A rG4-forming sequence was identified in the 5'-UTR and rG4 folding was confirmed by CD and NMR spectroscopy and DMS footprinting. The reporter system showed that the mutation of the rG4 eliminated response to H2O2 treatment, indicating its important regulatory role in oxidative stress response. Liquid chromatography-tandem mass spectrometry (LC-MS/MS)-based proteomics was employed to identified related protein factors and revealed the rG4-interacting protein, eukaryotic elongation factor 1 alpha (EF1a). An increase in translation efficiency during H2O2 treatment of the cells did not occur upon EF1a knockdown. Additionally, RIP and RNA pull-down methods showed that EF1a association with NRF2 rG4 increases during H2O2 treatment. Thus, the case represents an example of stress-activated rG4-mediated translation and, importantly, revealed one of the rG4-interacting proteins as a translation regulator.
There is a report on rG4 targets in alpha-synuclein (SNCA) mRNA 5'-UTR [86,87]. SNCA is a neuronal protein primarily located in presynaptic terminals, involved in modulating synaptic neurotransmission, and implicated in the pathogenesis of Parkinson's disease (PD) and other synucleinopathies. However, therapeutic targeting prospects for these rG4s are questionable. For cap-dependent translation, mutation of only one of three rG4 led to 36% increase in protein levels without changes in mRNA level, but its formation is unlikely based on sequence and was not confirmed in cuvette. Cap-independent translation efficiency decreased less than 15% when mutating all three rG4-forming sequences, though the effect remained statistically significant. These data question rG4 significance in SNCA mRNA 5'-UTR.
Thus, some mRNAs can be translated both through classical pathways and cap-independently under stress conditions and cap-dependent translation inhibition. rG4s in such mRNAs may serve dual roles, inhibiting cap-dependent translation while activating cap-independent one, supporting the idea of particular importance of rG4-mediated translation regulation under stress conditions. rG4s can either be part of the IRES structure or regulate its structure from a distance.
4.2. Other Translation-Enhancing rG4s
Previously, we discussed rG4s activating translation as IRES components. However, there are systems with rG4 translation activators functioning independently of IRES or lacking data on cap-independent translation. For instance, rG4 presence in FOXE3 mRNA 5'-UTR is known to enhance translation efficiency by 60% compared to rG4-mutant according to DLR data [27]. FOXE3 is a transcription factor characterized by a distinct forkhead domain and plays an important role in vertebrate lens formation.
rG4s in transforming growth factor-beta 2 (TGFβ2) mRNA 5'-UTR have been studied more thoroughly [88,89]. Two rG4s were discovered that jointly enhance translation efficiency with no evidence they are IRES elements. TGFβ2 is a multifunctional cytokine involved in various cell proliferation, differentiation, embryogenesis, migration, tissue repair, and immune response modulation processes. It is crucial for epicardial biology and heart development and linked to carcinogenesis, causing epithelial−mesenchymal transition, tumor invasiveness, and metastasis. The first rG4 is located 313 nt downstream of the cap structure [88]. Although incorporating the rG4-forming sequence alone into 5'-UTR of a reporter system showed a decrease in translation efficiency compared to the rG4-mutant, it enhances translation efficiency by up to 2-fold compared to the rG4-mutant in the full-length 5'-UTR context. Subsequently, a second rG4-forming sequence located 138 nt downstream of the cap structure was discovered [89]. According to DLR data, compared to the double mutant, wild type 5'-UTR sequence provides translation efficiency enhancement up to 224%, while mutation of either rG4 reduces activation level to approximately 130%. Interestingly, through mutations in rG4 loops, it was demonstrated that the G138–G155 regulatory element is active due to structure rather than sequence. The system of two translation-activating rG4s in TGFβ2 mRNA 5'-UTR is unique and deserves special attention due to the therapeutic potential of the target protein.
One more example of IRES-independent rG4 translation activator is the rG4 in cellular inhibitor of apoptosis protein 1 (cIAP1) mRNA 5'-UTR [90]. cIAP1 modulates various signal transduction pathways, especially TNFα-mediated signaling, to promote cell survival. In its longest version, the 5'-UTR is 1.7 kb long and contains several regulatory elements, including stress-activated IRES and uORF. In addition to these elements, a highly conserved G-rich sequence containing seven G-tracts was discovered near the cap structure. Notably, the search was conducted using G4RNA screener program, which utilizes machine learning results rather than specific rG4 motifs. In cellulo rG4 formation was supported by G4-RNA specific precipitation (G4RP) method. This involved stabilization of transiently formed rG4 structures by G4-stabilizer BRACO-19 followed by formaldehyde cross-linking before cell lysis and pull-down of rG4-forming RNA transcripts from the lysate using a biotin-linked biomimetic G4 ligand BioCyTASQ. Using luciferase reporter plasmids, rG4 mutation in the context of the entire 5'-UTR was shown to unexpectedly lead to dramatic decrease in translation efficiency, to a greater degree than typically observed for rG4s. When checking possible rG4 influence on distal IRES, IRES activity level under normal growth conditions proved insignificant and does not explain rG4's activating effect. Thus, rG4 in cIAP1 mRNA 5'-UTR represents a unique example of highly efficient and IRES-independent rG4 translation activator.
5. Discussion
One early article devoted to the investigation of rG4s' influence on mRNA 5'-UTR translation efficiency was titled "Predictable suppression of gene expression by 5'-UTR-based RNA quadruplexes"[15]. However, currently available data presents a much more complex picture of rG4-mediated translation regulation. These structures can both inhibit and activate translation. They can inhibit translation either through sterically hindering PIC scanning of 5'-UTR or by interacting with proteins like FMRP. While inhibiting downstream main reading frame translation, they can simultaneously activate translation from uORF and upstream near-cognate codons, thus acting as translation activators for these elements. In the case of uORFs, a synergistic inhibitory effect is observed. Interestingly, in prokaryotes, rG4 localization in coding sequences has been shown to enhance main reading frame translation efficiency, as with uORFs, through preventing leaky scanning and slowing PIC movement, which then better recognizes the start codon. This same secondary structure effect - enhancing and reducing translation efficiency of downstream and upstream codons respectively - has also been well studied in model systems with hairpins. This effect is yet be demonstrated for rG4s in eukaryotic coding sequences.
However, rG4s can activate translation: they can be integral parts of IRES, distally influence its structure, or even activate translation independently. In the latter case mechanisms are insufficiently studied, though in cIAP1's case it might be related to PIC assembly efficiency due to cap-proximal positioning. Moreover, helicases contribute significantly to regulating rG4-controlled translation.
Thus, translation of rG4-containing mRNAs can be finely tuned and tightly regulated for rapid cell response to changing environmental conditions.
However, the field of rG4 influence study faces numerous methodological challenges and artifacts. First is the challenge of bioinformatic identification of rG4-forming sequences. Initially, searches were conducted using the canonical motif G3+-N1-7-G3+-N1-7-G3+-N1-7-G3+ [91]. However, this approach proved insufficient: not all such sequences actually form rG4s (partly due to competition with other secondary structures), and simultaneously, some experimentally confirmed rG4s defy this motif, and contain long loops, bulges, or non-canonical tetrads [36,92,93,94,95,96]. Current search methods can be generally divided into two groups: those using hand-crafted rules and those employing machine learning [97]. Among the former, G4Catchall and G4Hunter are most effective, both allowing identification of a wide range of non-canonical motifs [93,97,98]. G4RNA screener, used in ADAR1 and BAG-1 studies, was one of the first machine learning-based methods [99]. Recently, the G4Boost algorithm was developed, surpassing known machine learning methods in accuracy and providing G4 thermodynamic stability assessment [100]. Thus, the toolkit for addressing this challenge has significantly improved.
The second methodological challenge derives from the usage of reporter systems as primary methods for confirming rG4s' influence on translation. Despite their widespread application and several undisputed advantages, such as rapid execution and ability to provide precise quantitative results, they have several inherent limitations. The main limitations are inadequate corresponding mRNA levels and increased cellular process load due to viral promoter usage [23,101,102]. This substantially alters the distribution and availability of transcription and translation enzymes in the cell. This alone causes cross-influence between test and normalization plasmid expression and reduces endogenous gene expression levels, including housekeeping genes [101,102]. Thus, the test inadequately models regulatory element effects during endogenous expression. Additionally, results strongly depend on experimental conditions: chosen promoter, transfection efficiency, and plasmid quantity, making data comparison challenging. Therefore, the application of developed robust methods to study rG4 effects at endogenous mRNA levels, such as genome editing and helicase knockdown, is necessary.
The third methodological challenge involves accurate sequencing of mRNA cap-proximal regions. It is hindered by the lack of TSS-specific signal sequence and presence of transcriptional noise [103,104]. Second, RNA-seq read lengths are typically shorter than required for quality mapping, and standard RNA-seq does not prevent the production of 5'-truncated sequences. To address this, methods were developed using special primers for 5'-end amplification (5'-RACE) or cap-containing mRNA pull-down (CAGE) [105]. The problem is that these methods were developed significantly later than many studies on rG4-regulating role in translation, and annotation and mRNA sequence refinement according to new data continues to this day. Moreover, transcription start sites and splicing can vary depending on cell line and condition, producing different mRNA sets. These challenges explain why the NRAS rG4, discovered in 2007 and becoming a classical example, was not detected in at least 14 cell lines in 2024 [31].
Contemporary rG4 research necessitates both the development of novel methodologies for evaluating endogenous rG4-mediated effects and the systematic reevaluation of previously identified rG4 targets using advanced analytical techniques, especially to confirm their presence in cellular mRNAs (Figure 8).
Author Contributions
Conceptualization, P.N.K; writing—original draft preparation, P.N.K; writing—review and editing, A.M.V and A.V.A; visualization, V.A.A.; supervision, V.A.K.; project administration, A.V.A.; funding acquisition, A.V.A. All authors have read and agreed to the published version of the manuscript.”
Funding
The study was supported by The Russian Scientific Foundation grant 24-15-00236 (https://rscf.ru/project/24-15-00236/) to A.V.A.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Tants, J.-N.; Schlundt, A. The Role of Structure in Regulatory RNA Elements. Biosci. Rep. 2024, 44, BSR20240139. [Google Scholar] [CrossRef] [PubMed]
- Leppek, K.; Das, R.; Barna, M. Functional 5′ UTR mRNA Structures in Eukaryotic Translation Regulation and How to Find Them. Nat. Rev. Mol. Cell Biol. 2018, 19, 158–174. [Google Scholar] [CrossRef]
- Bose, R.; Saleem, I.; Mustoe, A.M. Causes, Functions, and Therapeutic Possibilities of RNA Secondary Structure Ensembles and Alternative States. Cell Chem. Biol. 2024, 31, 17–35. [Google Scholar] [CrossRef]
- Varshney, D.; Spiegel, J.; Zyner, K.; Tannahill, D.; Balasubramanian, S. The Regulation and Functions of DNA and RNA G-Quadruplexes. Nat. Rev. Mol. Cell Biol. 2020, 21, 459–474. [Google Scholar] [CrossRef]
- Fay, M.M.; Lyons, S.M.; Ivanov, P. RNA G-Quadruplexes in Biology: Principles and Molecular Mechanisms. J. Mol. Biol. 2017, 429, 2127–2147. [Google Scholar] [CrossRef]
- Kwok, C.K.; Marsico, G.; Sahakyan, A.B.; Chambers, V.S.; Balasubramanian, S. rG4-Seq Reveals Widespread Formation of G-Quadruplex Structures in the Human Transcriptome. Nat. Methods 2016, 13, 841–844. [Google Scholar] [CrossRef]
- Yang, S.Y.; Lejault, P.; Chevrier, S.; Boidot, R.; Robertson, A.G.; Wong, J.M.Y.; Monchaud, D. Transcriptome-Wide Identification of Transient RNA G-Quadruplexes in Human Cells. Nat. Commun. 2018, 9, 4730. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.U.; Bartel, D.P. RNA G-Quadruplexes Are Globally Unfolded in Eukaryotic Cells and Depleted in Bacteria. Science 2016, 353, aaf5371. [Google Scholar] [CrossRef] [PubMed]
- Bao, H.-L.; Xu, Y. Investigation of Higher-Order RNA G-Quadruplex Structures in Vitro and in Living Cells by 19F NMR Spectroscopy. Nat. Protoc. 2018, 13, 652–665. [Google Scholar] [CrossRef] [PubMed]
- Miglietta, G.; Marinello, J.; Capranico, G. Immunofluorescence Microscopy of G-Quadruplexes and R-Loops. In Methods in Enzymology; Elsevier, 2024; Vol. 695, pp. 103–118 ISBN 978-0-443-21774-6.
- Chen, X.; Chen, S.; Dai, J.; Yuan, J.; Ou, T.; Huang, Z.; Tan, J. Tracking the Dynamic Folding and Unfolding of RNA G-Quadruplexes in Live Cells. Angew. Chem. Int. Ed. 2018, 57, 4702–4706. [Google Scholar] [CrossRef] [PubMed]
- Bugaut, A.; Balasubramanian, S. 5’-UTR RNA G-Quadruplexes: Translation Regulation and Targeting. Nucleic Acids Res. 2012, 40, 4727–4741. [Google Scholar] [CrossRef]
- Kumari, S.; Bugaut, A.; Huppert, J.L.; Balasubramanian, S. An RNA G-Quadruplex in the 5′ UTR of the NRAS Proto-Oncogene Modulates Translation. Nat. Chem. Biol. 2007, 3, 218–221. [Google Scholar] [CrossRef] [PubMed]
- Arora, A.; Dutkiewicz, M.; Scaria, V.; Hariharan, M.; Maiti, S.; Kurreck, J. Inhibition of Translation in Living Eukaryotic Cells by an RNA G-Quadruplex Motif. RNA 2008, 14, 1290–1296. [Google Scholar] [CrossRef] [PubMed]
- Halder, K.; Wieland, M.; Hartig, J.S. Predictable Suppression of Gene Expression by 5′-UTR-Based RNA Quadruplexes. Nucleic Acids Res. 2009, 37, 6811–6817. [Google Scholar] [CrossRef]
- Kumari, S.; Bugaut, A.; Balasubramanian, S. Position and Stability Are Determining Factors for Translation Repression by an RNA G-Quadruplex-Forming Sequence within the 5′ UTR of the NRAS Proto-Oncogene. Biochemistry 2008, 47, 12664–12669. [Google Scholar] [CrossRef] [PubMed]
- Edwards, M.; Joseph, S. The Fragile X Proteins Differentially Regulate Translation of Reporter mRNAs with G-Quadruplex Structures. J. Mol. Biol. 2022, 434, 167396. [Google Scholar] [CrossRef]
- Gomez, D.; Guédin, A.; Mergny, J.-L.; Salles, B.; Riou, J.-F.; Teulade-Fichou, M.-P.; Calsou, P. A G-Quadruplex Structure within the 5′-UTR of TRF2 mRNA Represses Translation in Human Cells. Nucleic Acids Res. 2010, 38, 7187–7198. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.J.; Wingate, K.L.; Silwal, J.; Leeper, T.C.; Basu, S. The Porphyrin TmPyP4 Unfolds the Extremely Stable G-Quadruplex in MT3-MMP mRNA and Alleviates Its Repressive Effect to Enhance Translation in Eukaryotic Cells. Nucleic Acids Res. 2012, 40, 4137–4145. [Google Scholar] [CrossRef] [PubMed]
- Balkwill, G.D.; Derecka, K.; Garner, T.P.; Hodgman, C.; Flint, A.P.F.; Searle, M.S. Repression of Translation of Human Estrogen Receptor α by G-Quadruplex Formation. Biochemistry 2009, 48, 11487–11495. [Google Scholar] [CrossRef] [PubMed]
- Weng, H.-Y.; Huang, H.-L.; Zhao, P.-P.; Zhou, H.; Qu, L.-H. Translational Repression of Cyclin D3 by a Stable G-Quadruplex in Its 5′ UTR: Implications for Cell Cycle Regulation. RNA Biol. 2012, 9, 1099–1109. [Google Scholar] [CrossRef]
- Shahid, R.; Bugaut, A.; Balasubramanian, S. The BCL-2 5′ Untranslated Region Contains an RNA G-Quadruplex-Forming Motif That Modulates Protein Expression. Biochemistry 2010, 49, 8300–8306. [Google Scholar] [CrossRef]
- Serikawa, T.; Eberle, J.; Kurreck, J. Effects of Genomic Disruption of a Guanine Quadruplex in the 5′ UTR of the Bcl-2 mRNA in Melanoma Cells. FEBS Lett. 2017, 591, 3649–3659. [Google Scholar] [CrossRef]
- Lammich, S.; Kamp, F.; Wagner, J.; Nuscher, B.; Zilow, S.; Ludwig, A.-K.; Willem, M.; Haass, C. Translational Repression of the Disintegrin and Metalloprotease ADAM10 by a Stable G-Quadruplex Secondary Structure in Its 5′-Untranslated Region. J. Biol. Chem. 2011, 286, 45063–45072. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.J.; Basu, S. An Unusually Stable G-Quadruplex within the 5′-UTR of the MT3 Matrix Metalloproteinase mRNA Represses Translation in Eukaryotic Cells. Biochemistry 2009, 48, 5313–5319. [Google Scholar] [CrossRef] [PubMed]
- Beaudoin, J.-D.; Perreault, J.-P. 5’-UTR G-Quadruplex Structures Acting as Translational Repressors. Nucleic Acids Res. 2010, 38, 7022–7036. [Google Scholar] [CrossRef]
- Agarwala, P.; Pandey, S.; Maiti, S. Role of G-Quadruplex Located at 5ʹ End of mRNAs. Biochim. Biophys. Acta BBA - Gen. Subj. 2014, 1840, 3503–3510. [Google Scholar] [CrossRef]
- Hacht, A.V.; Seifert, O.; Menger, M.; Schütze, T.; Arora, A.; Konthur, Z.; Neubauer, P.; Wagner, A.; Weise, C.; Kurreck, J. Identification and Characterization of RNA Guanine-Quadruplex Binding Proteins. Nucleic Acids Res. 2014, 42, 6630–6644. [Google Scholar] [CrossRef] [PubMed]
- Kawaji, H.; Lizio, M.; Itoh, M.; Kanamori-Katayama, M.; Kaiho, A.; Nishiyori-Sueki, H.; Shin, J.W.; Kojima-Ishiyama, M.; Kawano, M.; Murata, M.; et al. Comparison of CAGE and RNA-Seq Transcriptome Profiling Using Clonally Amplified and Single-Molecule next-Generation Sequencing. Genome Res. 2014, 24, 708–717. [Google Scholar] [CrossRef]
- Adamopoulos, P.G.; Tsiakanikas, P.; Stolidi, I.; Scorilas, A. A Versatile 5′ RACE-Seq Methodology for the Accurate Identification of the 5′ Termini of mRNAs. BMC Genomics 2022, 23, 163. [Google Scholar] [CrossRef] [PubMed]
- Balaratnam, S.; Torrey, Z.R.; Calabrese, D.R.; Banco, M.T.; Yazdani, K.; Liang, X.; Fullenkamp, C.R.; Seshadri, S.; Holewinski, R.J.; Andresson, T.; et al. Investigating the NRAS 5′ UTR as a Target for Small Molecules. Cell Chem. Biol. 2023, 30, 643–657.e8. [Google Scholar] [CrossRef]
- Katsuda, Y.; Sato, S.; Asano, L.; Morimura, Y.; Furuta, T.; Sugiyama, H.; Hagihara, M.; Uesugi, M. A Small Molecule That Represses Translation of G-Quadruplex-Containing mRNA. J. Am. Chem. Soc. 2016, 138, 9037–9040. [Google Scholar] [CrossRef]
- Lightfoot, H.L.; Hagen, T.; Tatum, N.J.; Hall, J. The Diverse Structural Landscape of Quadruplexes. FEBS Lett. 2019, 593, 2083–2102. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Yuan, J.; Xue, G.; Campanario, S.; Wang, D.; Wang, W.; Mou, X.; Liew, S.W.; Umar, M.I.; Isern, J.; et al. Translational Control by DHX36 Binding to 5′UTR G-Quadruplex Is Essential for Muscle Stem-Cell Regenerative Functions. Nat. Commun. 2021, 12, 5043. [Google Scholar] [CrossRef]
- Guédin, A.; Gros, J.; Alberti, P.; Mergny, J.-L. How Long Is Too Long? Effects of Loop Size on G-Quadruplex Stability. Nucleic Acids Res. 2010, 38, 7858–7868. [Google Scholar] [CrossRef] [PubMed]
- Jodoin, R.; Bauer, L.; Garant, J.-M.; Mahdi Laaref, A.; Phaneuf, F.; Perreault, J.-P. The Folding of 5′-UTR Human G-Quadruplexes Possessing a Long Central Loop. RNA 2014, 20, 1129–1141. [Google Scholar] [CrossRef] [PubMed]
- Jodoin, R.; Perreault, J.-P. G-Quadruplexes Formation in the 5’UTRs of mRNAs Associated with Colorectal Cancer Pathways. PLOS ONE 2018, 13, e0208363. [Google Scholar] [CrossRef]
- Katsuda, Y.; Sato, S.; Inoue, M.; Tsugawa, H.; Kamura, T.; Kida, T.; Matsumoto, R.; Asamitsu, S.; Shioda, N.; Shiroto, S.; et al. Small Molecule-Based Detection of Non-Canonical RNA G-Quadruplex Structures That Modulate Protein Translation. Nucleic Acids Res. 2022, 50, 8143–8153. [Google Scholar] [CrossRef]
- Chatterjee, S.; Sinha, S.; Kundu, C.N. Nectin Cell Adhesion Molecule-4 (NECTIN-4): A Potential Target for Cancer Therapy. Eur. J. Pharmacol. 2021, 911, 174516. [Google Scholar] [CrossRef]
- Silacci, P.; Mazzolai, L.; Gauci, C.; Stergiopulos, N.; Yin, H.L.; Hayoz, D. Gelsolin Superfamily Proteins: Key Regulators of Cellular Functions. Cell. Mol. Life Sci. 2004, 61, 2614–2623. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Zhang, X.; Liang, X.; Chen, Y.; Chen, Z.; Xiao, Z. CapG Promoted Nasopharyngeal Carcinoma Cell Motility Involving Rho Motility Pathway Independent of ROCK. World J. Surg. Oncol. 2022, 20, 347. [Google Scholar] [CrossRef]
- Wang, G.; Liu, H.; An, L.; Hou, S.; Zhang, Q. CAPG Facilitates Diffuse Large B-Cell Lymphoma Cell Progression through PI3K/AKT Signaling Pathway. Hum. Immunol. 2022, 83, 832–842. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Zhao, M.; Long, B.; Li, H. Super-Enhancer-Associated Gene CAPG Promotes AML Progression. Commun. Biol. 2023, 6, 622. [Google Scholar] [CrossRef]
- Agarwala, P.; Pal, G.; Pandey, S.; Maiti, S. Mutagenesis Reveals an Unusual Combination of Guanines in RNA G-Quadruplex Formation. ACS Omega 2017, 2, 4790–4799. [Google Scholar] [CrossRef] [PubMed]
- Fogarty, H.; Doherty, D.; O’Donnell, J.S. New Developments in von Willebrand Disease. Br. J. Haematol. 2020, 191, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Yao, J.; Yi, H.; Huang, X.; Zhao, W.; Yang, Z. To Unwind the Biological Knots: The DNA / RNA G-quadruplex Resolvase RHAU ( DHX36 ) in Development and Disease. Anim. Models Exp. Med. 2022, 5, 542–549. [Google Scholar] [CrossRef] [PubMed]
- Nie, J.; Jiang, M.; Zhang, X.; Tang, H.; Jin, H.; Huang, X.; Yuan, B.; Zhang, C.; Lai, J.C.; Nagamine, Y.; et al. Post-Transcriptional Regulation of Nkx2-5 by RHAU in Heart Development. Cell Rep. 2015, 13, 723–732. [Google Scholar] [CrossRef] [PubMed]
- Nishikura, K. Functions and Regulation of RNA Editing by ADAR Deaminases. Annu. Rev. Biochem. 2010, 79, 321–349. [Google Scholar] [CrossRef]
- Yitzhaki, S.; Huang, C.; Liu, W.; Lee, Y.; Gustafsson, Å.B.; Mentzer, R.M.; Gottlieb, R.A. Autophagy Is Required for Preconditioning by the Adenosine A1 Receptor-Selective Agonist CCPA. Basic Res. Cardiol. 2009, 104, 157–167. [Google Scholar] [CrossRef]
- Lyu, K.; Chen, S.; Chow, E.Y.; Zhao, H.; Yuan, J.; Cai, M.; Shi, J.; Chan, T.; Tan, J.; Kwok, C.K. An RNA G-Quadruplex Structure within the ADAR 5′UTR Interacts with DHX36 Helicase to Regulate Translation. Angew. Chem. Int. Ed. 2022, 61, e202203553. [Google Scholar] [CrossRef]
- Maltby, C.J.; Schofield, J.P.R.; Houghton, S.D.; O’Kelly, I.; Vargas-Caballero, M.; Deinhardt, K.; Coldwell, M.J. A 5′ UTR GGN Repeat Controls Localisation and Translation of a Potassium Leak Channel mRNA through G-Quadruplex Formation. Nucleic Acids Res. 2020, 48, 9822–9839. [Google Scholar] [CrossRef] [PubMed]
- Sui, Y.; Xu, Q.; Liu, D.; Liu, J. RNA Helicase DHX9 Enables PBX1 mRNA Translation by Unfolding RNA G-Quadruplex in Melanoma. EJC Skin Cancer 2024, 2, 100135. [Google Scholar] [CrossRef]
- Sun, J.; Wu, G.; Pastor, F.; Rahman, N.; Wang, W.-H.; Zhang, Z.; Merle, P.; Hui, L.; Salvetti, A.; Durantel, D.; et al. RNA Helicase DDX5 Enables STAT1 mRNA Translation and Interferon Signalling in Hepatitis B Virus Replicating Hepatocytes. Gut 2022, 71, 991–1005. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Qi, Q.; Xiong, W.; Shen, W.; Zhang, K.; Fan, R.; Zhang, Y.; Zhao, Y.; Xu, X.; Li, M.; et al. Unveiling a Potent Small Molecule Disruptor for RNA G-Quadruplexes Tougher Than DNA G-Quadruplex Disruption. ACS Chem. Biol. 2024, 19, 2032–2040. [Google Scholar] [CrossRef] [PubMed]
- Mitteaux, J.; Lejault, P.; Wojciechowski, F.; Joubert, A.; Boudon, J.; Desbois, N.; Gros, C.P.; Hudson, R.H.E.; Boulé, J.-B.; Granzhan, A.; et al. Identifying G-Quadruplex-DNA-Disrupting Small Molecules. J. Am. Chem. Soc. 2021, 143, 12567–12577. [Google Scholar] [CrossRef]
- Teng, X.; Hu, D.; Dai, Y.; Jing, H.; Hu, W.; Zhang, Q.; Zhang, N.; Li, J. Discovery of A G-Quadruplex Unwinder That Unleashes the Translation of G-Quadruplex-Containing mRNA without Inducing DNA Damage. Angew. Chem. Int. Ed. 2024, 63, e202407353. [Google Scholar] [CrossRef]
- Melko, M.; Bardoni, B. The Role of G-Quadruplex in RNA Metabolism: Involvement of FMRP and FMR2P. Biochimie 2010, 92, 919–926. [Google Scholar] [CrossRef]
- Song, J.; Perreault, J.-P.; Topisirovic, I.; Richard, S. RNA G-Quadruplexes and Their Potential Regulatory Roles in Translation. Translation 2016, 4, e1244031. [Google Scholar] [CrossRef]
- Vasilyev, N.; Polonskaia, A.; Darnell, J.C.; Darnell, R.B.; Patel, D.J.; Serganov, A. Crystal Structure Reveals Specific Recognition of a G-Quadruplex RNA by a β-Turn in the RGG Motif of FMRP. Proc. Natl. Acad. Sci. 2015, 112. [Google Scholar] [CrossRef] [PubMed]
- Schaeffer, C. The Fragile X Mental Retardation Protein Binds Specifically to Its mRNA via a Purine Quartet Motif. EMBO J. 2001, 20, 4803–4813. [Google Scholar] [CrossRef] [PubMed]
- Menon, L.; Mader, S.A.; Mihailescu, M.-R. Fragile X Mental Retardation Protein Interactions with the Microtubule Associated Protein 1B RNA. RNA 2008, 14, 1644–1655. [Google Scholar] [CrossRef] [PubMed]
- Castets, M.; Schaeffer, C.; Bechara, E.; Schenck, A.; Khandjian, E.W.; Luche, S.; Moine, H.; Rabilloud, T.; Mandel, J.-L.; Bardoni, B. FMRP Interferes with the Rac1 Pathway and Controls Actin Cytoskeleton Dynamics in Murine Fibroblasts. Hum. Mol. Genet. 2005, 14, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Pany, S.P.P.; Sapra, M.; Sharma, J.; Dhamodharan, V.; Patankar, S.; Pradeepkumar, P.I. Presence of Potential G-Quadruplex RNA-Forming Motifs at the 5′-UTR of PP2Acα mRNA Repress Translation. ChemBioChem 2019, 20, 2955–2960. [Google Scholar] [CrossRef]
- McAninch, D.S.; Heinaman, A.M.; Lang, C.N.; Moss, K.R.; Bassell, G.J.; Rita Mihailescu, M.; Evans, T.L. Fragile X Mental Retardation Protein Recognizes a G Quadruplex Structure within the Survival Motor Neuron Domain Containing 1 mRNA 5′-UTR. Mol. Biosyst. 2017, 13, 1448–1457. [Google Scholar] [CrossRef]
- Fähling, M.; Mrowka, R.; Steege, A.; Kirschner, K.M.; Benko, E.; Förstera, B.; Persson, P.B.; Thiele, B.J.; Meier, J.C.; Scholz, H. Translational Regulation of the Human Achaete-Scute Homologue-1 by Fragile X Mental Retardation Protein. J. Biol. Chem. 2009, 284, 4255–4266. [Google Scholar] [CrossRef]
- Murat, P.; Marsico, G.; Herdy, B.; Ghanbarian, A.; Portella, G.; Balasubramanian, S. RNA G-Quadruplexes at Upstream Open Reading Frames Cause DHX36- and DHX9-Dependent Translation of Human mRNAs. Genome Biol. 2018, 19, 229. [Google Scholar] [CrossRef] [PubMed]
- Singh, C.R.; Glineburg, M.R.; Moore, C.; Tani, N.; Jaiswal, R.; Zou, Y.; Aube, E.; Gillaspie, S.; Thornton, M.; Cecil, A.; et al. Human Oncoprotein 5MP Suppresses General and Repeat-Associated Non-AUG Translation via eIF3 by a Common Mechanism. Cell Rep. 2021, 36, 109376. [Google Scholar] [CrossRef] [PubMed]
- Reyes, C.J.F.; Asano, K. Between Order and Chaos: Understanding the Mechanism and Pathology of RAN Translation. Biol. Pharm. Bull. 2023, 46, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Tseng, Y.-J.; Krans, A.; Malik, I.; Deng, X.; Yildirim, E.; Ovunc, S.; Tank, E.M.H.; Jansen-West, K.; Kaufhold, R.; Gomez, N.B.; et al. Ribosomal Quality Control Factors Inhibit Repeat-Associated Non-AUG Translation from GC-Rich Repeats. Nucleic Acids Res. 2024, 52, 5928–5949. [Google Scholar] [CrossRef] [PubMed]
- Wright, S.E.; Rodriguez, C.M.; Monroe, J.; Xing, J.; Krans, A.; Flores, B.N.; Barsur, V.; Ivanova, M.I.; Koutmou, K.S.; Barmada, S.J.; et al. CGG Repeats Trigger Translational Frameshifts That Generate Aggregation-Prone Chimeric Proteins. Nucleic Acids Res. 2022, 50, 8674–8689. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-F.; Ursu, A.; Childs-Disney, J.L.; Guertler, R.; Yang, W.-Y.; Bernat, V.; Rzuczek, S.G.; Fuerst, R.; Zhang, Y.-J.; Gendron, T.F.; et al. The Hairpin Form of r(G4C2)Exp in c9ALS/FTD Is Repeat-Associated Non-ATG Translated and a Target for Bioactive Small Molecules. Cell Chem. Biol. 2019, 26, 179–190.e12. [Google Scholar] [CrossRef]
- Khateb, S.; Weisman-Shomer, P.; Hershco-Shani, I.; Ludwig, A.L.; Fry, M. The Tetraplex (CGG)n Destabilizing Proteins hnRNP A2 and CBF-A Enhance the in Vivo Translation of Fragile X Premutation mRNA. Nucleic Acids Res. 2007, 35, 5775–5788. [Google Scholar] [CrossRef]
- Zumwalt, M.; Ludwig, A.; Hagerman, P.J.; Dieckmann, T. Secondary Structure and Dynamics of the r(CGG) Repeat in the mRNA of the Fragile X Mental Retardation 1 (FMR1) Gene. RNA Biol. 2007, 4, 93–100. [Google Scholar] [CrossRef]
- Ofer, N.; Weisman-Shomer, P.; Shklover, J.; Fry, M. The Quadruplex r(CGG)n Destabilizing Cationic Porphyrin TMPyP4 Cooperates with hnRNPs to Increase the Translation Efficiency of Fragile X Premutation mRNA. Nucleic Acids Res. 2009, 37, 2712–2722. [Google Scholar] [CrossRef] [PubMed]
- Hoque, M.E.; Mahendran, T.; Basu, S. Reversal of G-Quadruplexes’ Role in Translation Control When Present in the Context of an IRES. Biomolecules 2022, 12, 314. [Google Scholar] [CrossRef] [PubMed]
- Meng, Z.; Jackson, N.L.; Shcherbakov, O.D.; Choi, H.; Blume, S.W. The Human IGF1R IRES Likely Operates through a Shine–Dalgarno-like Interaction with the G961 Loop (E-site) of the 18S rRNA and Is Kinetically Modulated by a Naturally Polymorphic polyU Loop. J. Cell. Biochem. 2010, 110, 531–544. [Google Scholar] [CrossRef]
- Bhattacharyya, D.; Diamond, P.; Basu, S. An Independently Folding RNA G-Quadruplex Domain Directly Recruits the 40S Ribosomal Subunit. Biochemistry 2015, 54, 1879–1885. [Google Scholar] [CrossRef]
- Jodoin, R.; Carrier, J.C.; Rivard, N.; Bisaillon, M.; Perreault, J.-P. G-Quadruplex Located in the 5′UTR of the BAG-1 mRNA Affects Both Its Cap-Dependent and Cap-Independent Translation through Global Secondary Structure Maintenance. Nucleic Acids Res. 2019, 47, 10247–10266. [Google Scholar] [CrossRef]
- Al-Zeer, M.A.; Dutkiewicz, M.; Von Hacht, A.; Kreuzmann, D.; Röhrs, V.; Kurreck, J. Alternatively Spliced Variants of the 5’-UTR of the ARPC2 mRNA Regulate Translation by an Internal Ribosome Entry Site (IRES) Harboring a Guanine-Quadruplex Motif. RNA Biol. 2019, 16, 1622–1632. [Google Scholar] [CrossRef] [PubMed]
- Bonnal, S.; Schaeffer, C.; Créancier, L.; Clamens, S.; Moine, H.; Prats, A.-C.; Vagner, S. A Single Internal Ribosome Entry Site Containing a G Quartet RNA Structure Drives Fibroblast Growth Factor 2 Gene Expression at Four Alternative Translation Initiation Codons. J. Biol. Chem. 2003, 278, 39330–39336. [Google Scholar] [CrossRef] [PubMed]
- Freiin Von Hövel, F.; Kefalakes, E.; Grothe, C. What Can We Learn from FGF-2 Isoform-Specific Mouse Mutants? Differential Insights into FGF-2 Physiology In Vivo. Int. J. Mol. Sci. 2020, 22, 390. [Google Scholar] [CrossRef]
- Morris, M.J.; Negishi, Y.; Pazsint, C.; Schonhoft, J.D.; Basu, S. An RNA G-Quadruplex Is Essential for Cap-Independent Translation Initiation in Human VEGF IRES. J. Am. Chem. Soc. 2010, 132, 17831–17839. [Google Scholar] [CrossRef] [PubMed]
- Cammas, A.; Dubrac, A.; Morel, B.; Lamaa, A.; Touriol, C.; Teulade-Fichou, M.-P.; Prats, H.; Millevoi, S. Stabilization of the G-Quadruplex at the VEGF IRES Represses Cap-Independent Translation. RNA Biol. 2015, 12, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhou, J.; Li, J.; Wang, Z.; Chen, Z.; Lv, Z.; Ge, L.; Xie, G.; Deng, G.; Rui, Y.; et al. N6 -Methyladenosine Promotes Translation of VEGFA to Accelerate Angiogenesis in Lung Cancer. Cancer Res. 2023, 83, 2208–2225. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.C.; Zhang, J.; Strom, J.; Yang, D.; Dinh, T.N.; Kappeler, K.; Chen, Q.M. G-Quadruplex in the NRF2 mRNA 5′ Untranslated Region Regulates De Novo NRF2 Protein Translation under Oxidative Stress. Mol. Cell. Biol. 2017, 37, e00122–16. [Google Scholar] [CrossRef]
- Cave, J.W.; Willis, D.E. G-quadruplex Regulation of Neural Gene Expression. FEBS J. 2022, 289, 3284–3303. [Google Scholar] [CrossRef]
- Koukouraki, P.; Doxakis, E. Constitutive Translation of Human α-Synuclein Is Mediated by the 5′-Untranslated Region. Open Biol. 2016, 6, 160022. [Google Scholar] [CrossRef]
- Agarwala, P.; Pandey, S.; Mapa, K.; Maiti, S. The G-Quadruplex Augments Translation in the 5′ Untranslated Region of Transforming Growth Factor Β2. Biochemistry 2013, 52, 1528–1538. [Google Scholar] [CrossRef] [PubMed]
- Agarwala, P.; Pandey, S.; Ekka, M.K.; Chakraborty, D.; Maiti, S. Combinatorial Role of Two G-Quadruplexes in 5′ UTR of Transforming Growth Factor Β2 (TGFβ2). Biochim. Biophys. Acta BBA - Gen. Subj. 2019, 1863, 129416. [Google Scholar] [CrossRef]
- Singha Roy, A.; Majumder, S.; Saha, P. Stable RNA G-Quadruplex in the 5′-UTR of Human cIAP1 mRNA Promotes Translation in an IRES-Independent Manner. Biochemistry 2024, acs.biochem.3c00521. [Google Scholar] [CrossRef]
- Huppert, J.L.; Balasubramanian, S. Prevalence of Quadruplexes in the Human Genome. Nucleic Acids Res. 2005, 33, 2908–2916. [Google Scholar] [CrossRef]
- Martadinata, H.; Phan, A.T. Formation of a Stacked Dimeric G-Quadruplex Containing Bulges by the 5′-Terminal Region of Human Telomerase RNA (hTERC). Biochemistry 2014, 53, 1595–1600. [Google Scholar] [CrossRef] [PubMed]
- Beaudoin, J.-D.; Jodoin, R.; Perreault, J.-P. New Scoring System to Identify RNA G-Quadruplex Folding. Nucleic Acids Res. 2014, 42, 1209–1223. [Google Scholar] [CrossRef]
- Liu, H.; Matsugami, A.; Katahira, M.; Uesugi, S. A Dimeric RNA Quadruplex Architecture Comprised of Two G:G(:A):G:G(:A) Hexads, G:G:G:G Tetrads and UUUU Loops. J. Mol. Biol. 2002, 322, 955–970. [Google Scholar] [CrossRef] [PubMed]
- Fratta, P.; Mizielinska, S.; Nicoll, A.J.; Zloh, M.; Fisher, E.M.C.; Parkinson, G.; Isaacs, A.M. C9orf72 Hexanucleotide Repeat Associated with Amyotrophic Lateral Sclerosis and Frontotemporal Dementia Forms RNA G-Quadruplexes. Sci. Rep. 2012, 2, 1016. [Google Scholar] [CrossRef] [PubMed]
- Bolduc, F.; Garant, J.-M.; Allard, F.; Perreault, J.-P. Irregular G-Quadruplexes Found in the Untranslated Regions of Human mRNAs Influence Translation. J. Biol. Chem. 2016, 291, 21751–21760. [Google Scholar] [CrossRef] [PubMed]
- Miskiewicz, J.; Sarzynska, J.; Szachniuk, M. How Bioinformatics Resources Work with G4 RNAs. Brief. Bioinform. 2021, 22, bbaa201. [Google Scholar] [CrossRef]
- Doluca, O. G4Catchall: A G-Quadruplex Prediction Approach Considering Atypical Features. J. Theor. Biol. 2019, 463, 92–98. [Google Scholar] [CrossRef]
- Garant, J.-M.; Perreault, J.-P.; Scott, M.S. Motif Independent Identification of Potential RNA G-Quadruplexes by G4RNA Screener. Bioinformatics 2017, 33, 3532–3537. [Google Scholar] [CrossRef] [PubMed]
- Cagirici, H.B.; Budak, H.; Sen, T.Z. G4Boost: A Machine Learning-Based Tool for Quadruplex Identification and Stability Prediction. BMC Bioinformatics 2022, 23, 240. [Google Scholar] [CrossRef] [PubMed]
- Di Blasi, R.; Marbiah, M.M.; Siciliano, V.; Polizzi, K.; Ceroni, F. A Call for Caution in Analysing Mammalian Co-Transfection Experiments and Implications of Resource Competition in Data Misinterpretation. Nat. Commun. 2021, 12, 2545. [Google Scholar] [CrossRef]
- Frei, T.; Cella, F.; Tedeschi, F.; Gutiérrez, J.; Stan, G.-B.; Khammash, M.; Siciliano, V. Characterization and Mitigation of Gene Expression Burden in Mammalian Cells. Nat. Commun. 2020, 11, 4641. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Shumate, A.; Pertea, G.; Varabyou, A.; Breitwieser, F.P.; Chang, Y.-C.; Madugundu, A.K.; Pandey, A.; Salzberg, S.L. CHESS: A New Human Gene Catalog Curated from Thousands of Large-Scale RNA Sequencing Experiments Reveals Extensive Transcriptional Noise. Genome Biol. 2018, 19, 208. [Google Scholar] [CrossRef]
- Batut, P.; Dobin, A.; Plessy, C.; Carninci, P.; Gingeras, T.R. High-Fidelity Promoter Profiling Reveals Widespread Alternative Promoter Usage and Transposon-Driven Developmental Gene Expression. Genome Res. 2013, 23, 169–180. [Google Scholar] [CrossRef]
- The RGASP Consortium; Steijger, T.; Abril, J.F.; Engström, P.G.; Kokocinski, F.; Hubbard, T.J.; Guigó, R.; Harrow, J.; Bertone, P. Assessment of Transcript Reconstruction Methods for RNA-Seq. Nat. Methods 2013, 10, 1177–1184. [Google Scholar] [CrossRef]
Figure 1.
rG4 inhibits cap-dependent translation through steric hindrance.
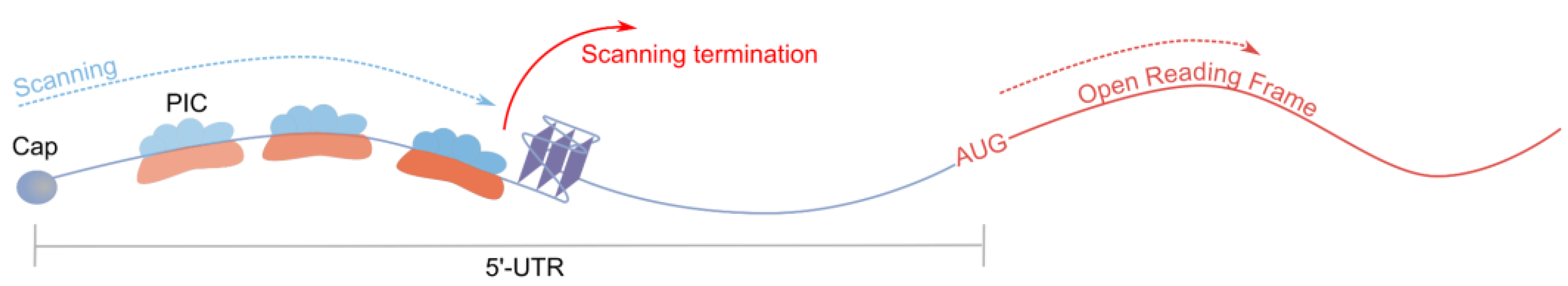
Figure 2.
Canonical (A) and non-canonical rG4s with long loop (B) and bulges (C).
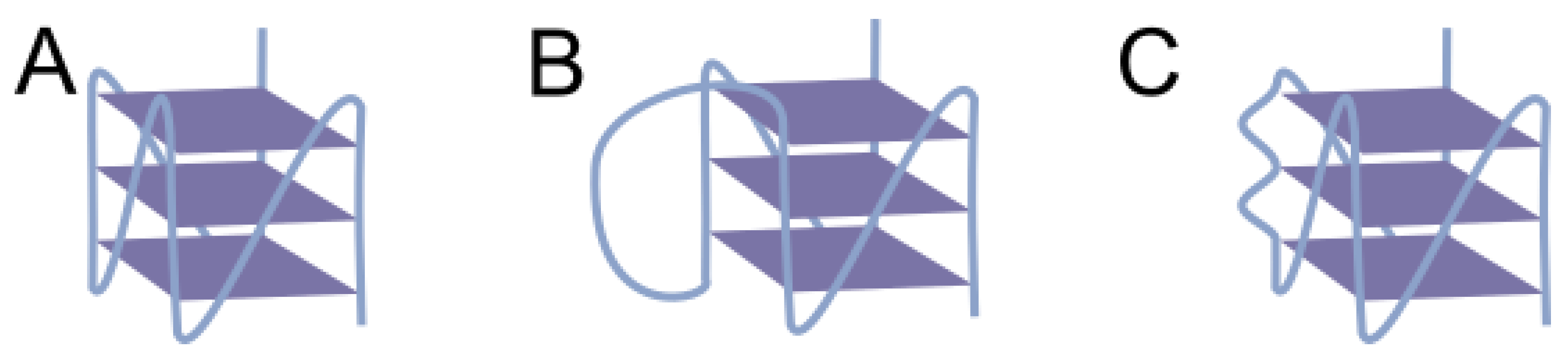
Figure 3.
A rG4-resolving helicase alleviates rG4-dependent translation inhibition.
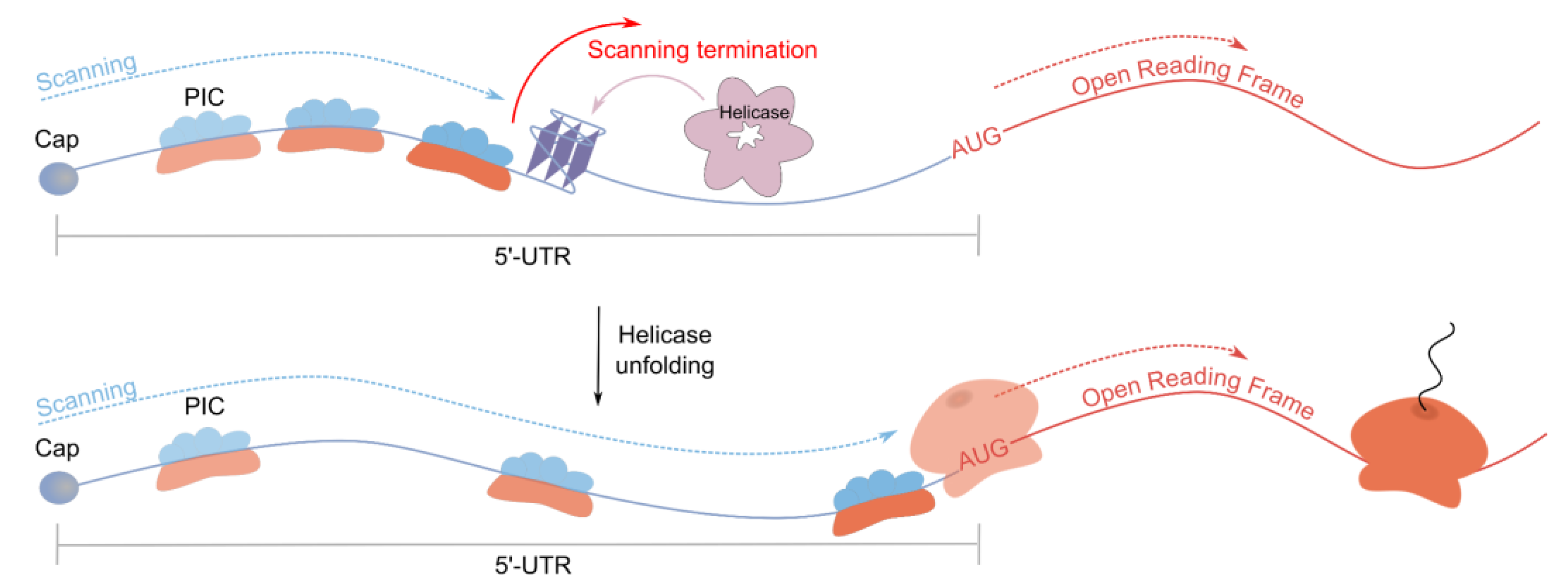
Figure 4.
FMRP regulates translation of MAP1B mRNA through rG4 in the 5’-UTR.
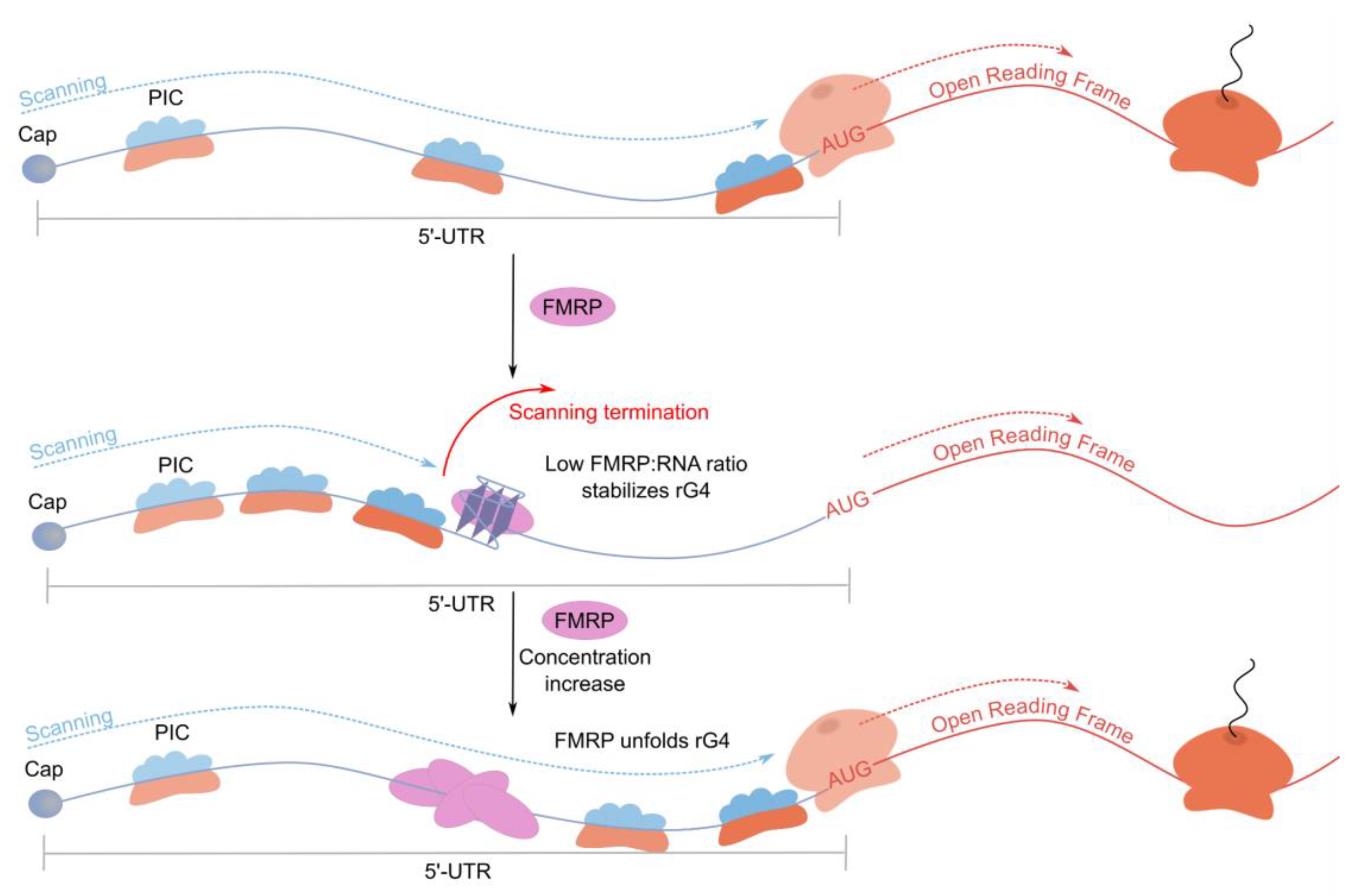
Figure 5.
rG4s and Upstream Open Reading Frames synergistically inhibit translation of the main ORF.
Figure 5.
rG4s and Upstream Open Reading Frames synergistically inhibit translation of the main ORF.
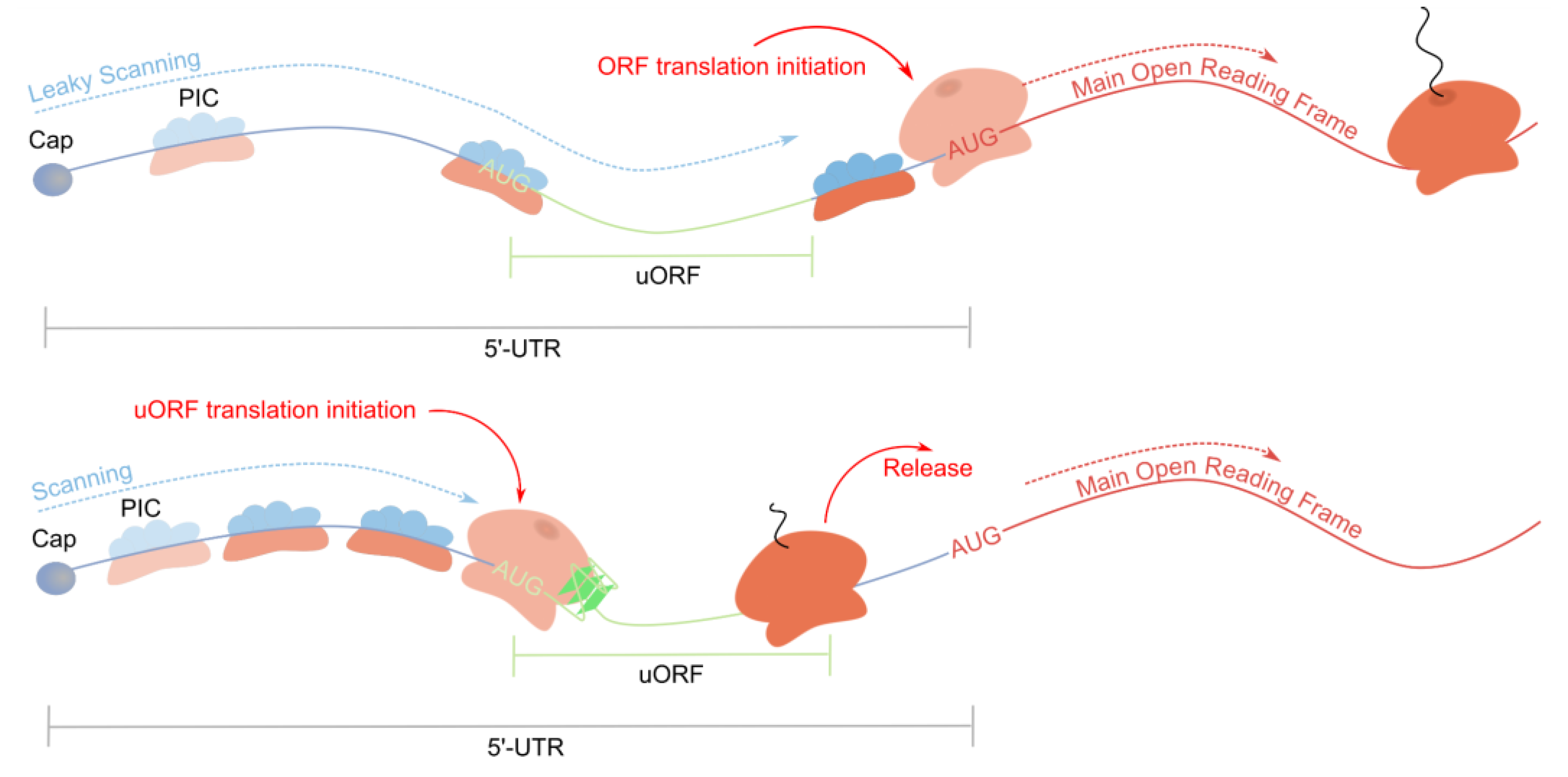
Figure 6.
rG4s promote repeat-associated non-AUG translation.
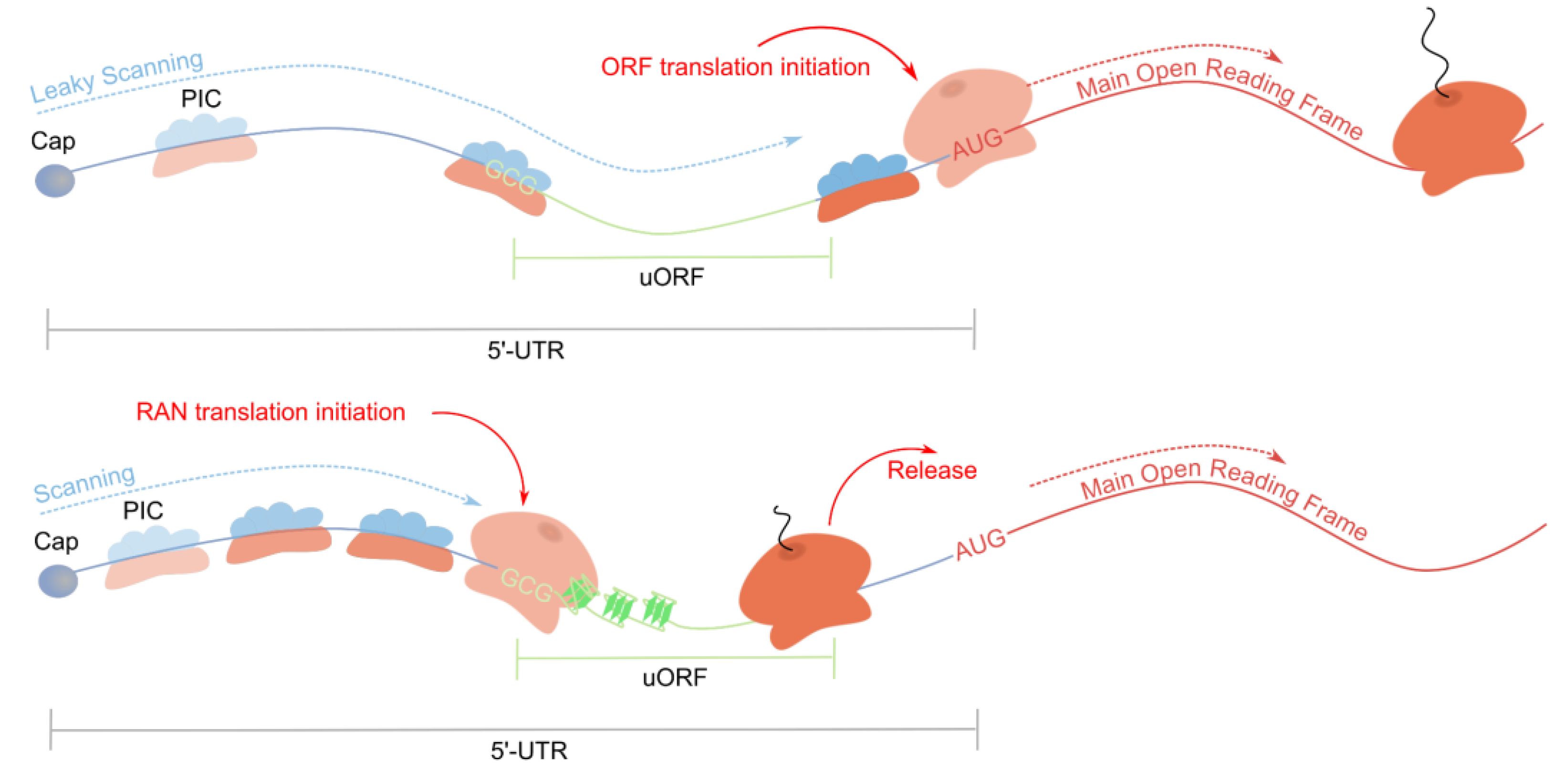
Figure 7.
rG4s as a part of IRES promote cap-independent translation.
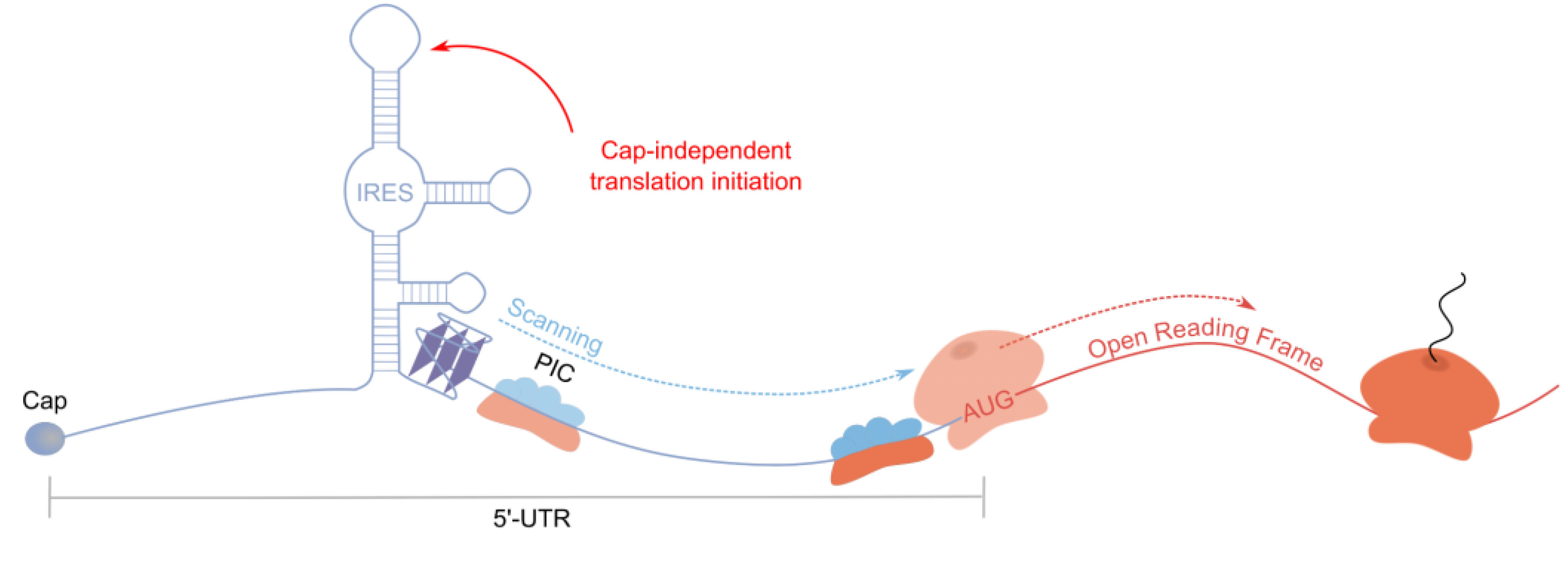
Figure 8.
Methods of investigation of rG4s in 5’-UTRs mRNAs as translation regulators.
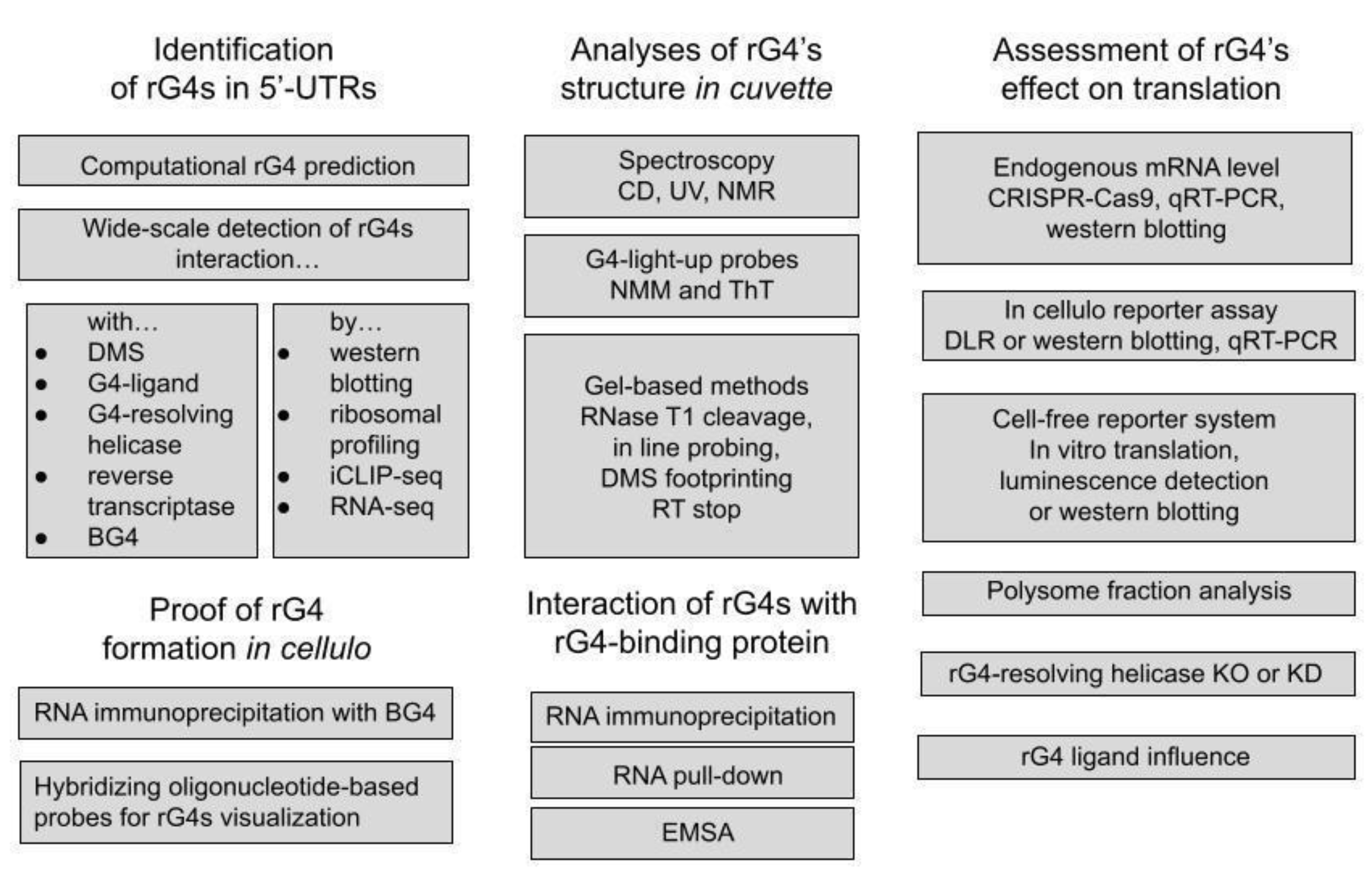

Table 1.
5'-UTR rG4s inhibiting translation.
| Target | Function | Methods used for verification of rG4s /theirs effect on translation | rG4’s effect on translation, fold change versus mutant | Ref. |
|---|---|---|---|---|
| NRAS | Membrane protein regulating cell proliferation and differentiation, associated with somatic rectal cancer, follicular thyroid cancer, juvenile myelomonocytic leukemia | UV-melting (Tm=63℃ at 1 mM KCl)/ in vitro translation | 3.6 1 | [13] |
| ZIC-1 | Transcription factor regulating cerebellar development, overexpressed in medulloblastoma | UV-melting (Tm=79℃ at 25 mM KCl)/DLR, western blotting | 52 | [14] |
| ESR1 | Transcription factor, regulating cell proliferation and differentiation, upregulated in estrogen receptor-positive breast cancers | CD-melting (Tm>85℃ at 100 mM KCl)/in vitro translation | 61 | [20] |
| TRF2 | Component of the telomere nucleoprotein shelterin complex, overexpressed in liver hepatocarcinomas, breast carcinomas and lung carcinomas | UV-melting (Tm=62℃ at 1 mM KCl)/ western blotting | 2.453 | [18] |
| CCND3 | Cyclin regulating the G0/G1 to S phase transition, overexpressed in myeloma, melanoma and mature B-cell malignancies | CD-melting (Tm=73℃ at 1 mM KCl), RNase T1/western blotting | 22 | [21] |
| BCL-2 | Integral outer mitochondrial membrane protein that blocks apoptosis and associated with follicular lymphoma and high-grade B-cell lymphoma | UV-melting (Tm=81℃ at 50 mM KCl)/ DLR, in vitro translation, western blotting | 3.51, 1.4-1.92 | [22,23] |
| ADAM10 | ɑ-Secretase of a disintegrin and metalloproteases family with anti-amyloidogenic activity | CD-melting (Tm=60℃ at 1 mM KCl)/ DLR, in vitro translation, western blotting | 31,2, 9-153 | [24] |
| MT3-MMP | Matrix metalloproteinase degrading extracellular matrix proteins, associated with invasiveness of gastric cancer, prostate cancer or renal carcinoma | CD-melting (Tm=72℃ at 1 mM KCl), RNase T1/ DLR | 2.22 | [25] |
| EBAG9 | Activator of apoptotic protease, associated with cervical adenocarcinoma and endometrial cancer | in-line probing/DLR | 1.82 | [26] |
| FZD2 | Transmembrane receptor coupled to the beta-catenin canonical signaling pathway, associated with omodysplasia, Robinow Syndrome and hepatocellular carcinoma | in-line probing/DLR | 2.52 | [26] |
| BARHL1 | Transcription factor involved in midbrain development/neuron migration, associted with triple-receptor negative breast cancer and medulloblastoma | in-line probing/DLR | 1.92 | [26] |
| NCAM2 | Membrane protein regulating cell adhesion/neuron adhesion, associated with epilepsy, familial temporal lobe and lissencephaly | in-line probing/DLR | 1.62 | [26] |
| THRA | Thyroid hormone receptor, associated with hypothyroidism, congenital, nongoitrous, and resistance to thyroid hormone | in-line probing/DLR | 1.62 | [26] |
| AASDHPPT | Phosphopantetheinyl transferase, associated with Pipecolic Acidemia and Lissencephaly | in-line probing/DLR | 2.22 | [26] |
| AKTIP | Protein interacting with protein kinase B (PKB)/Akt and modulating its activity by enhancing the phosphorylation of PKB regulatory sites | UV-melting (Tm=79℃ at 1 mM KCl)/DLR | 1.32 | [27] |
| CTSB | Lysosomal cysteine proteinase involved in the proteolytic processing of amyloid precursor protein (APP) | UV-melting (Tm=68℃ at 1 mM KCl)/DLR | 1.42 | [27] |
rG4’s effect on translation versus mutant was measured by means of 1 in vitro translation, 2 DLR, 3 western blotting.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



