
Submitted:
07 December 2024
Posted:
09 December 2024
You are already at the latest version
Abstract
Breast cancer (BC) is the most common cancer among women worldwide, therefore, the efforts of many scientists are aimed at finding effective biomarkers of this disease. It is known that exosomes are nanosized extracellular vesicles (EVs), which are released from various cell types, including cancer cells. Exosomes are directly involved in governing physiological and pathological processes of an organism by horizontal transfer of functional molecules (proteins, microRNA, etc.) from producing to receiving cells. Since the diagnosis and treatment of BC were improved substantially with exosomes, in this study, we isolated breast carcinoma cells‐derived exosomes, primary endoteliocytes-derived exosomes and blood exosomes from BC patients at 1st stage of disease and investigated their proteomic profile. Exosomes were isolated from the samples by ultrafiltration and ultracentrifugation, followed by mass spectrometric and bioinformatics analysis of the data. Exosomal nature of vesicles has been verified using transmission electron microscopy and flow cytometry. Exosome proteins secreted by MCF-7 and BT-474 cells were found to form 4 clusters, three of which enhanced the malignant potential of cancer cells, and one cluster coincided with a cluster of HUVEC-derived exosome proteins. Despite the different ensembles of proteins in exosomes from the MCF-7 and BT-474 lines, these portions of these proteins are involved in similar biological pathways. Comparison analysis reveals more BC-associated proteins were found in the exosomal content from the blood of BCPs than in the content of conditioned medium of cells mimicking the corresponding cancer subtype (89% and 82% for luminal A BC and MCF-7 cells and 86% and 75% for triple-positive BC and BT-474 cells, respectively). Thus, tumor-associated proteins should be sought not in exosomes secreted by cell lines, but in the composition of blood exosomes of cancer patients, while the contribution of endotheliocyte exosomes to the total pool of blood exosomes can be neglected.
Keywords:
exosomes
; extracellular vesicles
; MALDI-TOF mass spectrometry
; bioinformatics analysis
; HUVEC
; MCF-7
; BT-474
; breast cancer patients
; plasma
; blood
1. Introduction
In recent decades, there has been a steady increase in the number of new cases of malignant neoplasms, which is primarily due to increased life expectancy, as well as changes in lifestyle and exposure to unfavorable environmental factors. Today oncology ranks 2nd in mortality, conceding only to cardiovascular diseases. In 2022, breast cancer (BC) accounted for 23.8% of all new cancer cases in women worldwide. In particular, there were 2.3 million new cases of BC and 666,000 deaths from BC worldwide in 2022 [1]. The incidence of BC continues to increase despite the success of mammographic screening (depending on the country, from 14.2% (Republic of Moldova) to 47.3% (Czech Republic) of patients are diagnosed at stage I) [2]. Obviously, BC early diagnosis will increase the effectiveness of anti-tumor therapy and the survival rate of tumor patients. Thus, there is an urgent need to identify more effective and non-invasive surrogate markers that can guide not only early diagnosis, but also the selection of therapeutic strategies for individual patients and accurate assessment of prognosis [3]. Furthermore, BC is characterized by considerable tissue heterogeneity, showing distinct clinical and biological features, which make tumors respond differently to treatments and adverse in their management [4]. Today, molecular profiles have been largely explored, providing a well-established classification of BCs into five well-settled subtypes: Luminal A, Luminal B Her2+, Luminal B Her2-, Basal-like, and human epidermal growth factor receptor 2 (Her2)-enriched [2,5]. These molecular subtypes of BC are established by tumor biopsy, which can cause displacement of tumor cells, promoting metastasis and various pathological changes in breast tissue [6]. In addition, biopsy has an inaccuracy in determining the BC subtype, and also does not pose to track patients follow up [4]. In this regard, a promising direction in molecular oncology has been the search for new tumor markers in the composition of extracellular vesicles (EVs) by liquid biopsy. Among EVs, exosomes, small vesicles 30-150 nm in diameter, with a lipid bilayer membrane and tetraspanins CD9, CD63 and CD81 on their crown, are prominent [7,8]. The advantage of exosomes is the sorting of biologically active molecules (proteins, different types of RNA) at the maturation stage of these vesicles [9]. Recently, exosomes from breast cancer cell lines have been shown to be a rich source of breast cancer-related proteins and surface biomarkers, and can be used for diagnosis and prognosis of the disease [10,11]. However, when analyzing the content of exosomes circulating in the blood of cancer patients, it is necessary to take into account that in addition to tumor exosomes, exosomes of endotheliocytes and other non-tumor cells, as well as cells of the tumor microenvironment, are present in the blood.
To assess the diagnostic potential of blood exosomes, we performed differential analysis of exosomal proteomes from primary endotheliocytes, from two cell lines mimicking luminal A and luminal Her2-positive BC and from the blood of patients with these BC subtypes.
2. Materials and Methods
2.1. Isolation and Cultivation of Human Umbilical Vein Endothelial Cells (HUVEC)
HUVEC were obtained from three donors. Each vein washed sequentially with 50 mL phosphate-buffered saline (PBS) and 20 mL collagenase IV buffer (0.1% collagenase IV in buffer containing 1.5 mM HEPES, 14 mM NaCl, 0.4 mM KCl, 0.12 mM CaCl2, 0.04 mM MgSO4, and 0.76 mM D-glucose, pH 7.4) [12]. The vein was incubated with 0.1% collagenase IV solution at 37°C for 15 min to release endothelial cells. The collagenase solution containing detached cells was collected and combined with an additional PBS wash of the vein. The pooled solution was centrifuged at 800 g for 10 minutes, and the cell pellet was resuspended in IMDM (Gibco, Aucland, New Zealand) supplemented with 10% Fetal Bovine Serum (FBS) (Thermo Fisher Scientific) and Penicillin-Streptomycin (100 μg/ml) (Thermo Fisher Scientific). HUVEC were cultured in CO2-incubator (5% CO2) at 37°C, adherent cells were washed the next day with fresh IMDM to remove residual blood cells. HUVECs were cultured to 70–80% confluence, cells from the first passage were used for exosome isolation. For dissociation, cells were treated with 0.1% collagenase IV.
2.2. Cancer Cell Lines Cultivation
MCF-7 (ATCC® HTB-22™) and BT-474 (ATCC® HTB-20™) breast cancer cell lines were cultured in Dulbecco's Modified Eagle Medium (DMEM) (Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% FBS (Gibco, USA) and Penicillin-Streptomycin (100 μg/ml) (Thermo Fisher Scientific) in a CO₂ incubator (5% CO₂) at 37°C up to 70-80% confluence. Cells were subcultured with the solution of 0.25% trypsin in PBS, 5 mM EDTA.
2.3. Isolation of Exosomes from Conditioned Medium
All cells are negative for mycoplasma infection as confirmed by PCR analysis of 16S mycoplasma ribosomal gene [13].
FBS was centrifuged at 100,000g for 2 h at 4°C to remove bovine exosomes. The supernatant was collected and used to prepare bovine exosome-depleted medium. Three days prior to cell harvesting, the culture medium was replaced with a depleted medium containing IMDM for HUVEC or DMEM for MCF-7 and BT-474, a mixture of antibiotics, and 10% FBS devoid of bovine exosomes.
For isolation of exosomes, after 72 h conditioned medium was collected and subjected to two successive centrifugations at 300g and 15,000g for 10 min and 20 min, respectively to remove dead cells and cellular debris. To eliminate large EVs, the supernatant was filtered through 100 nm pore-size filters (Minisart high flow, 16553-K, Sartorius, Goettingen, Germany). Exosomes were isolated from the pre-cleared conditioned medium by centrifugation at 100,000g for 90 min at 100,000 g at 4°C. The pellets were suspended in 12 ml of PBS and again centrifuged for 90 min at 100,000 g at 4°C. The washing stage was repeated two times. Then, supernatant was removed, and exosomes were resuspended in 300 μl of PBS, aliquoted, frozen in liquid nitrogen, and stored at -80°C.
2.4. Ethics Statement
The study protocol was approved by the Ethics Committee of the Institute of Chemical Biology and Fundamental Medicine. Written informed consent was obtained from every female. Human samples were obtained according to the principles expressed in the Declaration of Helsinki. Blood samples from untreated BC patients with from luminal A (n = 5 age range 56–61 years, median age 61) and triple-positive (n = 8, age range 44–69 years, median age 61) subtypes [2,5] (Table 1) were obtained from Novosibirsk Regional Oncology Dispensary.
2.5. Exosome Isolation from Blood
Venous blood (9 mL) was collected by venipuncture in K3EDTA spray-coated vacutainers (Improvacuter, Guangzhou, China), and processed within one hour. To isolate total blood exosomes by ultrafiltration and differential ultracentrifugation, the previously described protocol was used [14]. The pellet containing blood exosomes was re-suspended in 300 μl of PBS.
2.6. Characterization of Exosomes
Morphology and membrane integrity of the isolated exosomes was assessed by transmission electron microscopy (TEM) as described previously [15]. The initial volumes of exosomes using the TEM were 15 μl.
To evaluate the protein concentration of exosomes, a NanoOrange Protein Quantitation kit (NanoOrange Protein Quantitation Kit, Molecular Probes, USA) was used as described previously [16].
The presence of CD9, CD63 and CD 81 tetraspanins in the exosomal crown was confirmed by flow cytometry as described previously [16]. The MFI of stained exosomes was analyzed and compared to the isotype control (BD bioscience, Heidelberg, Germany). Flow cytometry was performed on the Cytoflex (Becman Coulter, BioBay, China), using CytExpert 2.0 Software.
2.7. Mass Spectrometry Analysis
For the identification of exosomal proteins by MALDI-TOF mass-spectrometry, proteins were separated using SDS-disk electrophoresis, than fragments of polyacrylamide gel containing the studied proteins were washed from SDS and subjected to trypsinolysis as described previously [14]. Peptide fragments of proteins were extracted from the gel, concentrated, and desalted on C18 ZipTips microcolumns (Milipore, Burlington, MA, USA). Next, 5 μl of a saturated solution of α-Cyano-4-Hydroxycinnamic Acid (HCCA) in 70% acetonitrile was added and then spotted onto the MTP 384 ground steel target plate. After crystallization, the target was loaded into the mass spectrometer to obtain the protein molecular weight spectrum. The acquisition and registration of mass spectra was carried out on a time-of-flight tandem mass spectrometer MALDI-TOF/TOF spectrometer Ultraflex III (Bruker Daltonics, Bremen, Germany). To ensure the reliability and reproducibility of the results, all analyses were performed in five biological replicates for each sample.
Spectra were acquired using the following parameters: shots – 150, laser frequency – 66.7 Hz, laser attenuator offset – 85%, laser attenuator range – 21%, laser attenuator set – 5_ularge, laser focus offset – 0%, laser focus range – 100%, and laser focus value – 4%. The instrument was pre-calibrated for a mass range of 500-3800 kDa. The obtained spectra were converted into mass values using flexAnalysis software. Protein identification was performed by searching for appropriate candidates in the annotated NCBI and SwissProt databases using the Mascot program (Matrix Science Ltd., London, UK, www.matrixscience.com/search_form_select.html accessed on 20 October 2024) with the following search parameters: species – Homo sapiens, error tolerance – ±300 ppm, maximum number of missed cleavages – 2, fixed modifications – Propionamide (C), variable modifications – Oxidation (M), Phospho (ST); identification reliability not lower than 95%.
2.8. Bioinformatics Analysis
Data from the SwissProt database were translated into the UniProt database for further analysis using the Retrieve/ID mapping platform (https://www.uniprot.org). Functional enrichment analysis of exosomal proteomes according to Gene Ontologies was conducted using the STRING software (https://www.string-db.org/). Cellular localization, molecular functions, and involvement in biological processes were determined using FunRich 3.1.3 software based on the Gene Ontologies GO Component, GO Function, and GO Process. Involvement in biological pathways was assessed using the Reactome service (https://reactome.org/). For primary data processing and graph generation, Python libraries such as Pandas, Numpy, Matplotlib, and Seaborn were utilized.
3. Results
3.1. Characterization of Isolated Exosomes
To characterize exosomes isolated from conditioned medium and BCP blood, TEM and flow cytometry were used. TEM revealed the presence of clearly structured cup-shaped vesicles (40-95 nm) of low electron density with preserved membrane in all samples; the portion of vesicles with the size smaller than 30 nm, was no more than 13% (Figure 1). We then confirmed that the exosomes isolated from all sources contained the typical exosome protein markers, CD9 and CD63 tetraspanins. For this purpose, exosomes adsorbed onto aldehyde/sulfate latex beads using anti-CD9 antibodies were stained with FITC-labeled antibodies to CD63 tetraspanin. It was shown that CD9 and CD63 tetraspanins were comparably represented in all vesicles (Figure 1).
Collectively, obtained data reveal that the sEVs isolated from conditioned medium and blood have all properties of exosomes [20].
3.2. Annotation of Identified Proteins from HUVEC-Derived Exosomes
Since in addition to tumor exosomes, exosomes of normal cells, including those of endothelial origin, circulate in the blood of cancer patients, the first stage of the work involved the identification and characterization of exosome proteins secreted by primary endothelial cells from three donors. A total of 38 proteins were identified with high confidence (P < 0.05) using MALDI-TOF mass-spectrometry in the HUVEC-derived exosomes (Supplemental Table S1), of these, 29% (11 proteins) were common for three groups (Figure 2A). Main part (30/38) of identified exosomal proteins from HUVEC were previously discovered in other studies, using mass-spectrometry and annotated in the Vesiclepedia database [21].
Thus, about 21% of exosomal proteins are identified in our study for the first time as a part of extracellular vesicles; previously, they were not annotated in this database (Figure 2B).
The protein-protein interaction was mainly concentrated in the relevance among ALB, CD9, CD63, CD81, ADAM10, MMP9, GTF2F1, SPEK1, of which 6 major exosomal proteins form a cluster (Figure 3).
Predominantly eight proteins (ALB, CD9, CD63, CD81, MMP9, GTF2F1, RPS3A, PSMD10) were found to be significantly (P<0.05) involved in biological pathways (Table 2). In particular, the common proteomic profiles of endothelial exosomes are enriched with terms related to biological processes such as “transport of organic anions”, “bile acid and bile salt metabolism”, “metabolism of mRNA”, “adaptive immune system”, “pausing and recovery of elongation”, “integrin signaling”, etc.
3.3. Annotation of Protein Cargo from BC-Derived Exosomes
Using MALDI-TOF mass-spectrometry, 61 and 65 proteins were identified with high reliability (p < 0.05) in the exosomes from BC cell line - BT-474 and MCF-7, respectively (Supplementary Tables S2 and S3). Of these, only 5% (6 proteins) were common to both groups (Supplementary Tables S2 and S3, Figure 4A). It should be noted that the same proteins (CD9, CD24, CD63, CD81, MMP9, ALB) were common to exosomes of cancer cells and primary cells. Approximately 36% of identified exosomal proteins from cancer cells were previously discovered in other studies, using mass spectrometry and annotated in the Vesiclepedia database (Figure 4B).
For cancer-derived exosomes the protein-protein interaction was mainly concentrated in the relevance among NDUFS4, COPS5, CNKN1B, H2ACB, H2AC14, H2AC7, PPIA, RBBP6, CSTF1, UPF3B, SRSF6, SRSF5, RBMX, PRPF38A, RPS20, KCNC4, RPS27I, RPL28, RPL14, SPCS2, VAMP8, CRX, CALML5, RALB, EDN1, EFNB2, CD9, CD63, CD81, CD86, ADAM10 and MMP9, of which 28 exosomal proteins form four clusters: in addition to the cluster of major proteins identified by molecular cargo analysis of primary endotheliocytes, three clusters were identified that enhance the malignant potential of MCF-7 and BT-474 cells (Figure 5).
Analysis of the protein cargo of exosomes secreted by MCF-7 showed the involvement of proteins in biological pathways such as “Membrane Trafficking”, “Clathrin derived vesicle budding”, “trans-Golgi Network Vesicle Budding”, “Post-Elongation Processing of Intron-Containing pre-mRNA”, “Cleavage of Growing Transcript in the Termination Region”, “Integrins in angiogenesis”, “Signaling by SCF-KIT”, “Notch signaling pathway”, “Signaling by EGFR”, etc. (Table 3). It should be noted that 8 of the 10 (80%) proteins from Table 3 were identified by protein-protein interaction analysis as proteins interacting with a large number of other proteins (Figure 5).
Similar bioinformatics analysis of proteins from BT-474 secreted exosomes revealed the involvement of in biological pathways such as “Nonsense Mediated Decay Enhanced by the Exon Junction Complex”, “Insulin Synthesis and Processing”, “Gene Expression”, “Peptide chain elongation”, “Metabolism of proteins”, “mRNA Splicing”, “Eukaryotic Translation Initiation”, “Transcription”, “CXCR4-mediated signaling events”, “Hypoxic and oxygen homeostasis regulation of HIF-1-alpha”, etc. (Table 4). As with exosomal proteins from MCF-7, in exosomes from BT-474, 13 of the 17 proteins (76%) presented in Table 4 were also identified by protein-protein interaction analysis as proteins that interact with a large number of other proteins (Figure 5). It was also shown that despite the different ensembles of proteins within exosomes from MCF-7 and BT-474 lines, these exosomal proteins are involved in many of the same biological pathways such as “Post-Elongation Processing of Intron-Containing pre-mRNA”, “mRNA 3'-end processing”, “Cleavage of Growing Transcript in the Termination Region”, “Post-Elongation Processing of the Transcript”, “RNA Polymerase II Transcription Termination” and “Receptor-ligand binding initiates the second proteolytic cleavage of Notch receptor” (Table 3 and Table 4).
3.4. Comparative Proteomic Analysis of Exosomes in the Blood of BCPs with Luminal A and Triple-Positive Subtypes
Using MALDI-TOF mass spectrometry with high confidence (p < 0.05) were identified 74 and 110 proteins in exosomes from blood of untreated BCPs with luminal A (n = 5, Table 1) and triple-positive (n = 8, Table 1) subtypes, respectively (Supplementary Tables S4 and S5). It should be noted that 8 out of 41 universal exosomal proteins were detected in half of the samples. The analysis of the identified exosomal proteins by Vesiclepedia database reveals that among 89 proteins that have been previously found to be associated with vesicles, a total of 47 and 74 proteins were identified in the luminal A BC and triple-positive BC exosomes, respectively. Thus, 38% of proteins identified in exosomes from blood of BCPs in this study were not previously annotated in the Vesiclepedia database (Figure 6).
To determine whether the exosomal proteins we identified were found in breast neoplasms, the list of proteins was analyzed, using FunRich 3.13 software to search publicly available database for proteins in BC—The Human Protein Atlas (HPA). It was shown that 66 BC-associated proteins from blood exosomes of BCPs with luminal A subtype and 53 - from MCF-7-derived exosomes were annotated in the HPA database (Figure 7A).
Similar results were obtained for protein cargo of exosomes from blood of triple-positive BCPs and from BT-474 conditioned medium (Figure 7B). Thus, more BC-associated proteins were found in the composition of exosomes from the blood of BCPs than in the exosomes of the composition of conditioned medium of cells mimicking the corresponding cancer subtype (89% and 82% for luminal A BC and MCF-7 cells and 86% and 75% for triple-positive BC and BT-474 cells, respectively).
4. Discussion
Tumor exosomes able to transport biological active molecules (RNA, proteins, and metabolites) in recipient cells have been recognized fundamental mediators of cell-to-cell communication in cancer, including BC. Since the molecular cargo of exosomes reflects the composition of the parent cell, in the field of molecular diagnostics, great expectations are associated with the identification of tumor-specific signatures in the composition of exosomes for the development of a method for the diagnosis of malignant neoplasms by liquid biopsy [22,23]. One of the undoubted advantages of exosomal proteome study is the possibility of removing ballast proteins of blood plasma and increasing the concentration of tumor-specific proteins, including membrane proteins. It should be noted that the membrane of vesicles protects their contents from the action of proteases and nucleases, and vesicle preparations are stable and can be stored for a long time in laboratory conditions [24].
The search for tumor markers in exosomes is complicated by the high individual variability of the exosome proteome even in the group of healthy donors. At comparing the proteomic profiles of blood plasma exosomes of fifteen clinically healthy donors, only 9 out of 109 identified exosomal proteins were present in all samples [25]. Our profiling of the proteome of HUVEC-derived exosomes partially confirms these data: only 29% of proteins within primary endotheliocytes were common to the three human umbilical vein donors.
Avoiding the problems of low reproducibility between different samples, as well as solving the problem of the amount of protein for mass spectrometry, most researchers work with cell culture exosomes, which is reflected in the Vesiclepedia database. As a result, the authors provide data on thousands of proteins in the composition of exosomes obtained from conditioned media of various cell lines. In particular, Altelaar's group successfully propitiated exosome proteomes from cell lines mimicking triple-negative (BT-549, Hs578T, LM2, MDAMB-231), HER-2 positive (HCC1419, HCC1954, JIMT1, SKBR-3) and luminal A (MCF-7) breast cancer subtypes [26], identifying 4648 proteins. However, the authors do not take into account since the size of exosomes is less than 150 nm, vesicle cannot contain more than 100 proteins. Thus, there is no point in obtaining too large quantities of exosomes and describing proteins that can be detected in blood exosomes with very low probability.
Modern BC diagnostic methods (mammography, breast ultrasound, and MRI) can detect neoplasms larger than 0.5 cm; at the same time, these tests are often ambiguous and have documented drawbacks [3]. For simplicity of calculation, we assume that only the surface cells of the tumor secrete exosomes to the external space. Then the surface area of a 5 mm diameter sphere-shaped tumor is 78.5 mm2, and taking into account the average diameter of a breast carcinoma cell of 20 µm, a quarter of a million cells will be located on the outer surface of the tumor. Taking into account the above calculations, in the current work, for the analysis of exosome proteins by mass spectrometry, the proteins of exosomes secreted by 500,000 MCF-7 or BT-474 cells were applied to a gel and separated. The obtained data on the proteomic profile of exosomes from conditioned media of breast carcinoma lines correspond to the number of exosomes secreted by the tumor at stage T1. Comparative analysis of proteins of secreted by primary endotheliocytes exosomes and breast carcinoma cells exosomes showed that it is possible to neglect the contribution of the endotheliocyte exosome proteome when searching for tumor markers in the composition of blood exosomes.
To confirm that EVs are ideal diagnostic tools, Hoshino’s group analyzed proteomes of EVs from different sources. It was shown that EV proteins from human plasma overlapped best with human serum-derived EVs (r2 = 0.92), followed by human bone marrow (r2 = 0.65) and lymphatic fluid EVs (r2 = 0.64), and correlated least with human cell line (r2 = 0.15) and tissue explant -derived EVs (r2 = 0.24), suggesting the complexity of plasma and lymph EV proteomes may drive the divergence from tissue EV proteomes [27]. Our study also revealed an extremely low coincidence of exosome proteins secreted by MCF-7 cells and blood exosomes of patients with luminal A subtype of BC, as well as exosome proteins secreted by BT-474 cells and blood exosomes of patients with triple-positive subtype of BC. However, when comparing protein profiles of blood exosomes of patients and tissue proteins, a significant coincidence of proteins was found. It should be noted, that breast tumors are characterized by significant variability in cellular composition, as well as histological, expression, and genotypic heterogeneity. In particular, intratumoral morphologic heterogeneity of invasive breast carcinoma of nonspecific type, the most common histologic form of breast cancer (incidence rate up to 80%), has been described [28]. As a consequence, exosomes secreted by tumor cells have a more diverse composition than exosomes originating from MCF-7 or BT-474 cells. In addition, in the body, besides cancer cells, exosomes are secreted by other cells, in particular cells of the tumor microenvironment [29]. Thus, exosomal content in blood of BCPs at T1 stage reflect the cancer-associated changes occurring not only in the developing primary tumor, but also the tumor microenvironment. Despite previous studies searching for protein biomarkers within exosomes secreted by breast carcinoma cell lines [11,30,31], there is no consensus on exosomal tumor markers due to limited EV proteomic datasets from human samples and appropriate controls for data analysis and interpretation.
Taken together, our results support the idea that tumor-associated proteins should be sought not in exosomes secreted by cell lines, but in the composition of blood exosomes of cancer patients, while the contribution of endotheliocyte exosomes to the total pool of blood exosomes can be neglected.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, Y.T. and S.T.; methodology, A.S., L.Y., O.T., A.G., N.Y.; formal analysis, A.S.; investigation, A.S., L.Y., K.P., O.T., A.G., N.Y.; resources, Y.T.; data curation, S.T.; writing—original draft preparation, A.S., L.Y. and S.T.; writing—review and editing, Y.T. and S.T.; project administration, L.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by a grant from the Russian Science Foundation, N23-25-00462, https://rscf.ru/project/23-25-00462/, accessed on 25 January 2023.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and was approved by the Local Ethics Committee of the Institute of Chemical Biology and Fundamental Medicine (the protocol N3 from 2 September 2024).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef] [PubMed]
- Pu, Q.; Gao, H. The Role of the Tumor Microenvironment in Triple-Positive Breast Cancer Progression and Therapeutic Resistance. Cancers 2023, 15, 5493. [Google Scholar] [CrossRef] [PubMed]
- Shefer, A.; Yalovaya, A.; Tamkovich, S. Exosomes in breast cancer: Involvement in tumor dissemination and prospects for liquid biopsy. Int. J. Mol. Sci. 2022, 23, 8845. [Google Scholar] [CrossRef] [PubMed]
- Bandini, E.; Rossi, T.; Scarpi, E.; Gallerani, G.; Vannini, I.; Salvi, S.; Azzali, I.; Melloni, M.; Salucci, S.; Battistelli, M.; Serra, P.; Maltoni, R.; Cho, W.C.; Fabbri, F. Early Detection and Investigation of Extracellular Vesicles Biomarkers in Breast Cancer. Front. Mol. Biosci. 2021, 8, 732900. [Google Scholar] [CrossRef]
- Johnson, K.S.; Conant, E.F.; Soo, M.S. Molecular Subtypes of Breast Cancer: A Review for Breast Radiologists. J. Breast Imaging 2021, 3, 12–24. [Google Scholar] [CrossRef]
- Bilous, M. Breast core needle biopsy: Issues and controversies. Mod Pathol. 2010, 23 (Suppl. S2), S36–S45. [Google Scholar] [CrossRef]
- Lee, Y.; Ni, J.; Wasinger, V.C.; Graham, P.; Li, Y. Comparison Study of Small Extracellular Vesicle Isolation Methods for Profiling Protein Biomarkers in Breast Cancer Liquid Biopsies. Int. J. Mol. Sci. 2023, 24, 15462. [Google Scholar] [CrossRef]
- Kugeratski, F.G.; Hodge, K.; Lilla, S.; McAndrews, K.M.; Zhou, X.; Hwang, R.F.; Zanivan, S.; Kalluri, R. Quantitative proteomics identifies the core proteome of exosomes with syntenin-1 as the highest abundant protein and a putative universal biomarker. Nat. Cell Biol. 2021, 23, 631–641. [Google Scholar] [CrossRef]
- Neagu, A.-N.; Whitham, D.; Bruno, P.; Morrissiey, H.; Darie, C.A.; Darie, C.C. Omics-Based Investigations of Breast Cancer. Molecules 2023, 28, 4768. [Google Scholar] [CrossRef]
- Risha, Y.; Minic, Z.; Ghobadloo, S.M.; Berezovski, M.V. The proteomic analysis of breast cell line exosomes reveals disease patterns and potential biomarkers. Sci. Rep. 2020, 10, 13572. [Google Scholar] [CrossRef]
- Gangoda, L.; Liem, M.; Ang, C.S.; Keerthikumar, S.; Adda, C.G.; Parker, B.S.; Mathivanan, S. Proteomic Profiling of Exosomes Secreted by Breast Cancer Cells with Varying Metastatic Potential. Proteomics 2017, 17. [Google Scholar] [CrossRef] [PubMed]
- Stepanova, A.O.; Laktionov, P.P.; Cherepanova, A.V.; Chernonosova, V.S.; Shevelev, G.Y.; Zaporozhchenko, I.A.; Karaskov, A.M.; Laktionov, P.P. General Study and Gene Expression Profiling of Endotheliocytes Cultivated on Electrospun Materials. Materials 2019, 12, 4082. [Google Scholar] [CrossRef] [PubMed]
- Cherepanova, A.V.; Bushuev, A.V.; Kharkova, M.V.; Vlassov, V.V.; Laktionov, P.P. DNA inhibits dsRNA-induced secretion of pro-inflammatory cytokines by gingival fibroblasts. Immunobiology 2013, 218, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Tutanov, O.; Orlova, E.; Proskura, K.; Grigor’eva, A.; Yunusova, N.; Tsentalovich, Y.; Alexandrova, A.; Tamkovich, S. Proteomic Analysis of Blood Exosomes from Healthy Females and Breast Cancer Patients Reveals an Association between Different Exosomal Bioactivity on Non-tumorigenic Epithelial Cell and Breast Cancer Cell Migration in Vitro. Biomolecules 2020, 10, 495. [Google Scholar] [CrossRef] [PubMed]
- Tamkovich, S.; Tutanov, O.; Efimenko, A.; Grigor'eva, A.; Ryabchikova, E.; Kirushina, N.; Vlassov, V.; Tkachuk, V.; Laktionov, P. Blood Circulating Exosomes Contain Distinguishable Fractions of Free and Cell-Surface-Associated Vesicles. Curr. Mol. Med. 2019, 19, 273–285. [Google Scholar] [CrossRef]
- Tamkovich, S.N.; Yunusova, N.V.; Tugutova, E.; Somov, A.K.; Proskura, K.V.; Kolomiets, L.A.; Stakheeva, M.N.; Grigor’eva, A.E.; Laktionov, P.P.; Kondakova, I.V. Protease cargo in circulating exosomes of breast cancer and ovarian cancer patients. Asian Pac. J. Cancer Prev. 2019, 20, 255–262. [Google Scholar] [CrossRef]
- Aebersold, R.; Mann, M. Mass spectrometry-based proteomics. Nature 2003, 422, 198–207. [Google Scholar] [CrossRef]
- Domon, B.; Aebersold, R. Mass Spectrometry and Protein Analysis. Science 2006, 312, 212–217. [Google Scholar] [CrossRef]
- Suckau, D.; Resemann, A.; Schuerenberg, M.; Hufnagel, P.; Franzen, J.; Holle, A. A novel MALDI LIFT-TOF/TOF mass spectrometer for proteomics. Anal. Bioanal. Chem. 2003, 376, 952–965. [Google Scholar] [CrossRef]
- Batista, I.A.; Machado, J.C.; Melo, S.A. Advances in exosomes utilization for clinical applications in cancer. Trends Cancer. 2024, 10, 947–968. [Google Scholar] [CrossRef]
- Vesiclepedia database. www.microvesicles.org (accessed on 30 November 2024).
- Bates, M.; Mohamed, B.M.; Lewis, F.; O'Toole, S.; O'Leary, J.J. Biomarkers in high grade serous ovarian cancer. Biochim. Biophys. Acta Rev. Cancer. 2024, 1879, 189224. [Google Scholar] [CrossRef] [PubMed]
- Bao, H.; Min, L.; Bu, F.; Wang, S.; Meng, J. Recent advances of liquid biopsy: Interdisciplinary strategies toward clinical decision-making. Interdisciplinary Medicine 2023, 1, e20230021. [Google Scholar] [CrossRef]
- Zhang, Y.; Bi, J.; Huang, J.; Tang, Y.; Du, S.; Li, P. Exosome: A Review of Its Classification, Isolation Techniques, Storage, Diagnostic and Targeted Therapy Applications. Int. J. Nanomedicine 2020, 15, 6917–6934. [Google Scholar] [CrossRef] [PubMed]
- Bastos-Amador, P.; Royo, F.; Gonzalez, E.; Conde-Vancells, J.; Palomo-Diez, L.; Borras, F.E.; Falcon-Perez, J.M. Proteomic analysis of microvesicles from plasma of healthy donors reveals high individual variability. J. Proteomics 2012, 75, 3574–3584. [Google Scholar] [CrossRef]
- Rontogianni, S.; Synadaki, E.; Li, B.; Liefaard, M.C.; Lips, E.H.; Wesseling, J.; Wu, W.; Altelaar, M. Proteomic profiling of extracellular vesicles allows for human breast cancer subtyping. Commun. Biol. 2019, 2, 325. [Google Scholar] [CrossRef]
- Hoshino, A.; Kim, H.S.; Bojmar, L.; Gyan, K.E.; Cioffi, M.; Hernandez, J.; Zambirinis, C.P.; Rodrigues, G.; Molina, H.; Heissel, S.; Mark, M.T.; Steiner, L.; Benito-Martin, A.; Lucotti, S.; Di Giannatale, A.; Offer, K.; Nakajima, M.; Williams, C.; Nogués, L.; Pelissier Vatter, F.A.; Hashimoto, A.; Davies, A.E.; Freitas, D.; Kenific, C.M.; Ararso, Y.; Buehring, W.; Lauritzen, P.; Ogitani, Y.; Sugiura, K.; Takahashi, N.; Alečković, M.; Bailey, K.A.; Jolissant, J.S.; Wang, H.; Harris, A.; Schaeffer, L.M.; García-Santos, G.; Posner, Z.; Balachandran, V.P.; Khakoo, Y.; Raju, G.P.; Scherz, A.; Sagi, I.; Scherz-Shouval, R.; Yarden, Y.; Oren, M.; Malladi, M.; Petriccione, M.; De Braganca, K.C.; Donzelli, M.; Fischer, C.; Vitolano, S.; Wright, G.P.; Ganshaw, L.; Marrano, M.; Ahmed, A.; DeStefano, J.; Danzer, E.; Roehrl, M.H.A.; Lacayo, N.J.; Vincent, T.C.; Weiser, M.R.; Brady, M.S.; Meyers, P.A.; Wexler, L.H.; Ambati, S.R.; Chou, A.J.; Slotkin, E.K.; Modak, S.; Roberts, S.S.; Basu, E.M.; Diolaiti, D.; Krantz, B.A.; Cardoso, F.; Simpson, A.L.; Berger, M.; Rudin, C.M.; Simeone, D.M.; Jain, M.; Ghajar, C.M.; Batra, S.K.; Stanger, B.Z.; Bui, J.; Brown, K.A.; Rajasekhar, V.K.; Healey, J.H.; de Sousa, M.; Kramer, K.; Sheth, S.; Baisch, J.; Pascual, V.; Heaton, T.E.; La Quaglia, M.P.; Pisapia, D.J.; Schwartz, R.; Zhang, H.; Liu, Y.; Shukla, A.; Blavier, L.; DeClerck, Y.A.; LaBarge, M.; Bissell, M.J.; Caffrey, T.C.; Grandgenett, P.M.; Hollingsworth, M.A.; Bromberg, J.; Costa-Silva, B.; Peinado, H.; Kang, Y.; Garcia, B.A.; O'Reilly, E.M.; Kelsen, D.; Trippett, T.M.; Jones, D.R.; Matei, I.R.; Jarnagin, W.R.; Lyden, D. Extracellular Vesicle and Particle Biomarkers Define Multiple Human Cancers. Cell 2020, 182, 1044–1061.e18. [Google Scholar] [CrossRef]
- Gerashchenko, T.S.; Zavyalova, M.V.; Denisov, E.V.; Krakhmal, N.V.; Pautova, D.N.; Litviakov, N.V.; Vtorushin, S.V.; Cherdyntseva, N.V.; Perelmuter, V.M. Intratumoral Morphological Heterogeneity of Breast Cancer as an Indicator of the Metastatic Potential and Tumor Chemosensitivity. Acta Naturae 2017, 9, 56–67. [Google Scholar] [CrossRef]
- Eguchi, T.; Sheta, M.; Fujii, M.; Calderwood, S.K. Cancer extracellular vesicles, tumoroid models, and tumor microenvironment. Semin. Cancer Biol. 2022, 86 Pt 1, 112–126. [Google Scholar] [CrossRef]
- Kugeratski, F.G.; Hodge, K.; Lilla, S.; McAndrews, K.M.; Zhou, X.; Hwang, R.F.; Zanivan, S.; Kalluri, R. Quantitative proteomics identifies the core proteome of exosomes with syntenin-1 as the highest abundant protein and a putative universal biomarker. Nat Cell Biol. 2021, 23, 631–641. [Google Scholar] [CrossRef]
- Gelsomino, L.; Barone, I.; Caruso, A.; Giordano, F.; Brindisi, M.; Morello, G.; Accattatis, F.M.; Panza, S.; Cappello, A.R.; Bonofiglio, D.; et al. Proteomic Profiling of Extracellular Vesicles Released by Leptin-Treated Breast Cancer Cells: A Potential Role in Cancer Metabolism. Int. J. Mol. Sci. 2022, 23, 12941. [Google Scholar] [CrossRef]
Figure 1.
Exosomes characterization by TEM and flow cytometry analysis. At total view of exosomes preparation and expression of CD63 on CD9-positive vesicles obtained from: (a) conditioned medium from HUVEC; (b) conditioned medium from MCF-7; (c) conditioned medium from BT-474; (d) blood from patients with luminal A BC; (e) blood from patients with triple positive BC, scale bars correspond to 100 nm. Electron microscopy, negative staining by phosphotungstate acid. For flow cytometry mean MFI are shown. Negative control (f) (latex beads labeled anti-CD9 with anti CD63 FITC antibody).
Figure 1.
Exosomes characterization by TEM and flow cytometry analysis. At total view of exosomes preparation and expression of CD63 on CD9-positive vesicles obtained from: (a) conditioned medium from HUVEC; (b) conditioned medium from MCF-7; (c) conditioned medium from BT-474; (d) blood from patients with luminal A BC; (e) blood from patients with triple positive BC, scale bars correspond to 100 nm. Electron microscopy, negative staining by phosphotungstate acid. For flow cytometry mean MFI are shown. Negative control (f) (latex beads labeled anti-CD9 with anti CD63 FITC antibody).
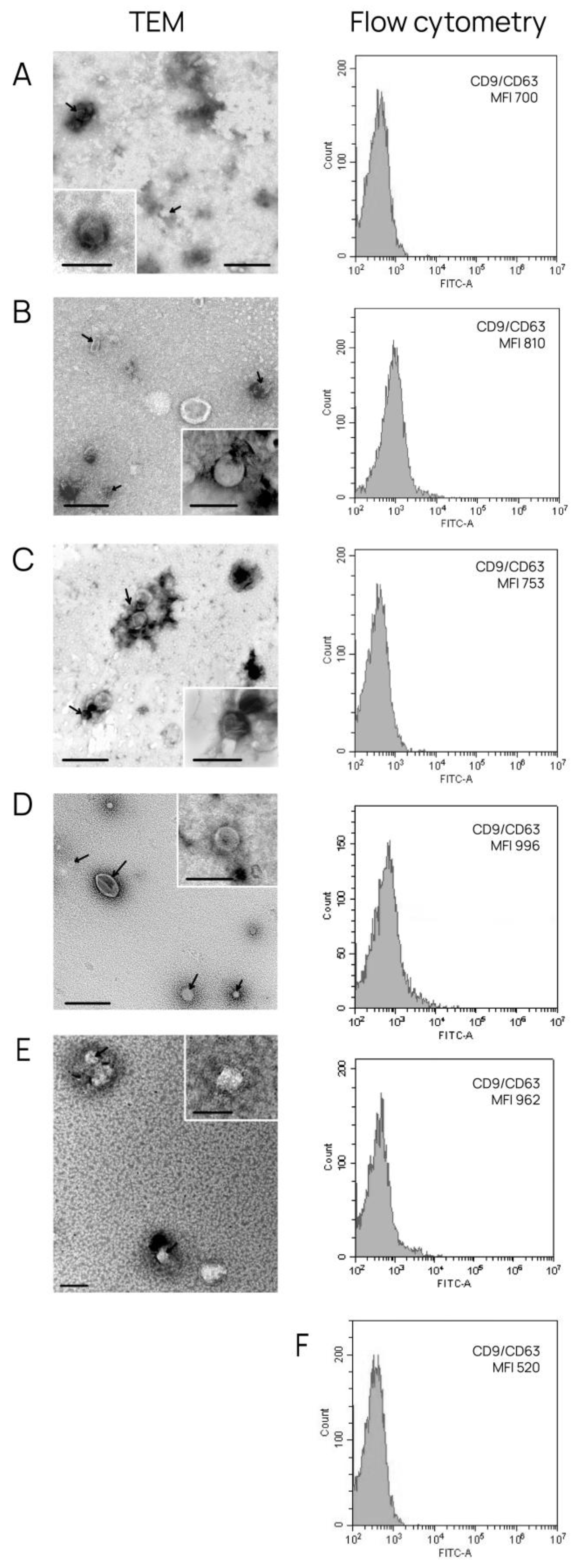
Figure 2.
Venn–Euler diagrams of proteins in exosomes from HUVEC from three donors (A) and with Vesiclepedia databases (B).
Figure 2.
Venn–Euler diagrams of proteins in exosomes from HUVEC from three donors (A) and with Vesiclepedia databases (B).
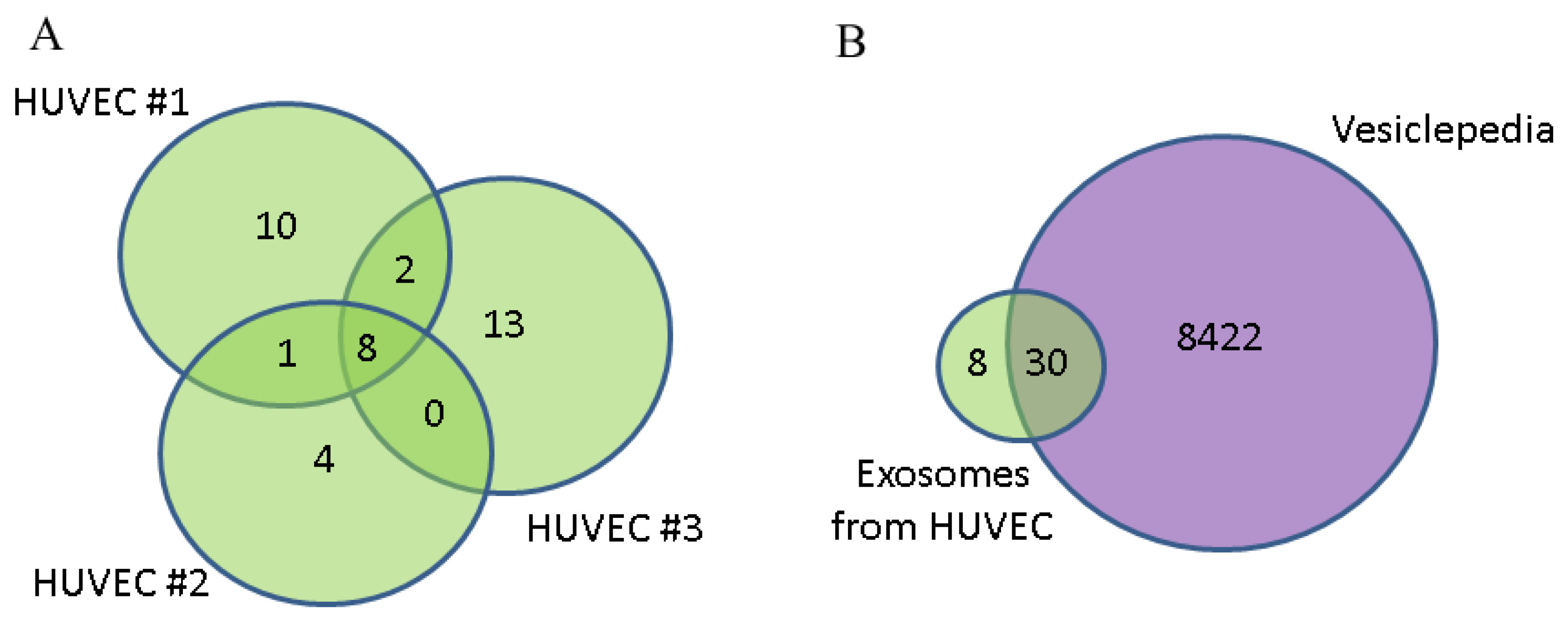
Figure 3.
Protein–protein interaction network of 31 proteins from HUVEC-derived exosomes. PPI networks plotted with STRING (http://string-db.org/, accessed on 30 November 2024) with following settings—minimum interaction score: high confidence (0.400); active interaction sources: textmining, experiments, databases, co-expression, neighborhood, gene fusion, co-occurrence.
Figure 3.
Protein–protein interaction network of 31 proteins from HUVEC-derived exosomes. PPI networks plotted with STRING (http://string-db.org/, accessed on 30 November 2024) with following settings—minimum interaction score: high confidence (0.400); active interaction sources: textmining, experiments, databases, co-expression, neighborhood, gene fusion, co-occurrence.
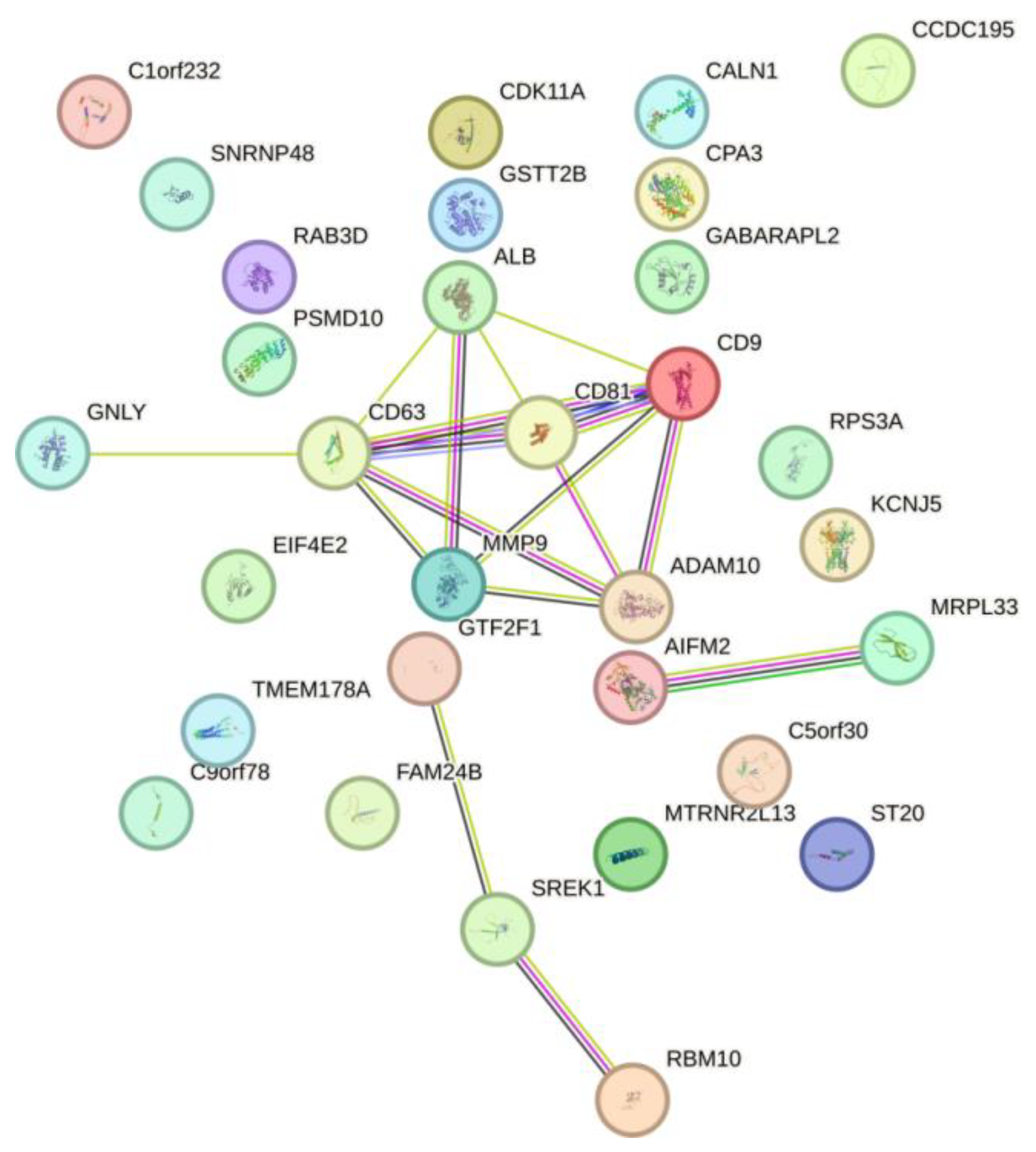
Figure 4.
Venn–Euler diagrams of proteins in exosomes from HUVEC, BT-474 and MCF-7 (A) and with Vesiclepedia databases (B).
Figure 4.
Venn–Euler diagrams of proteins in exosomes from HUVEC, BT-474 and MCF-7 (A) and with Vesiclepedia databases (B).
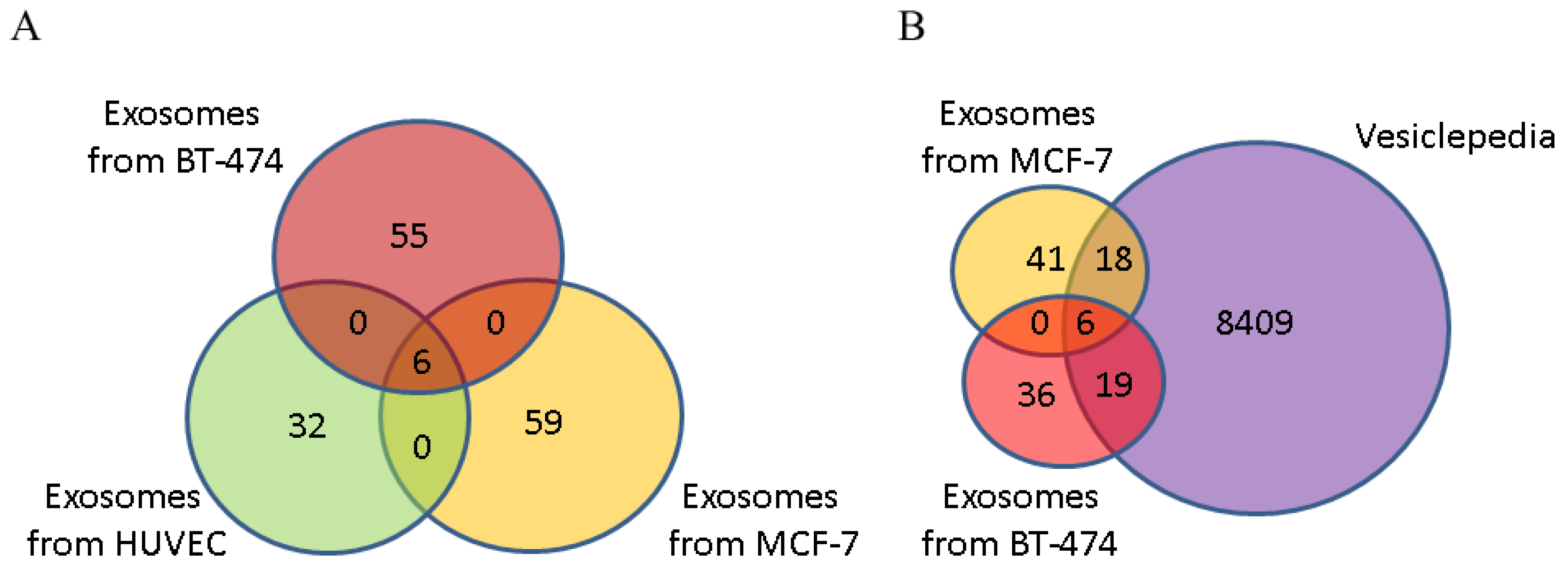
Figure 5.
Protein–protein interaction network of 110 proteins from cancer cells-derived exosomes. PPI networks plotted with STRING (http://string-db.org/, accessed on 30 November 2024) with following settings—minimum interaction score: high confidence (0.400); active interaction sources: textmining, experiments, databases, co-expression, neighborhood, gene fusion, co-occurrence.
Figure 5.
Protein–protein interaction network of 110 proteins from cancer cells-derived exosomes. PPI networks plotted with STRING (http://string-db.org/, accessed on 30 November 2024) with following settings—minimum interaction score: high confidence (0.400); active interaction sources: textmining, experiments, databases, co-expression, neighborhood, gene fusion, co-occurrence.
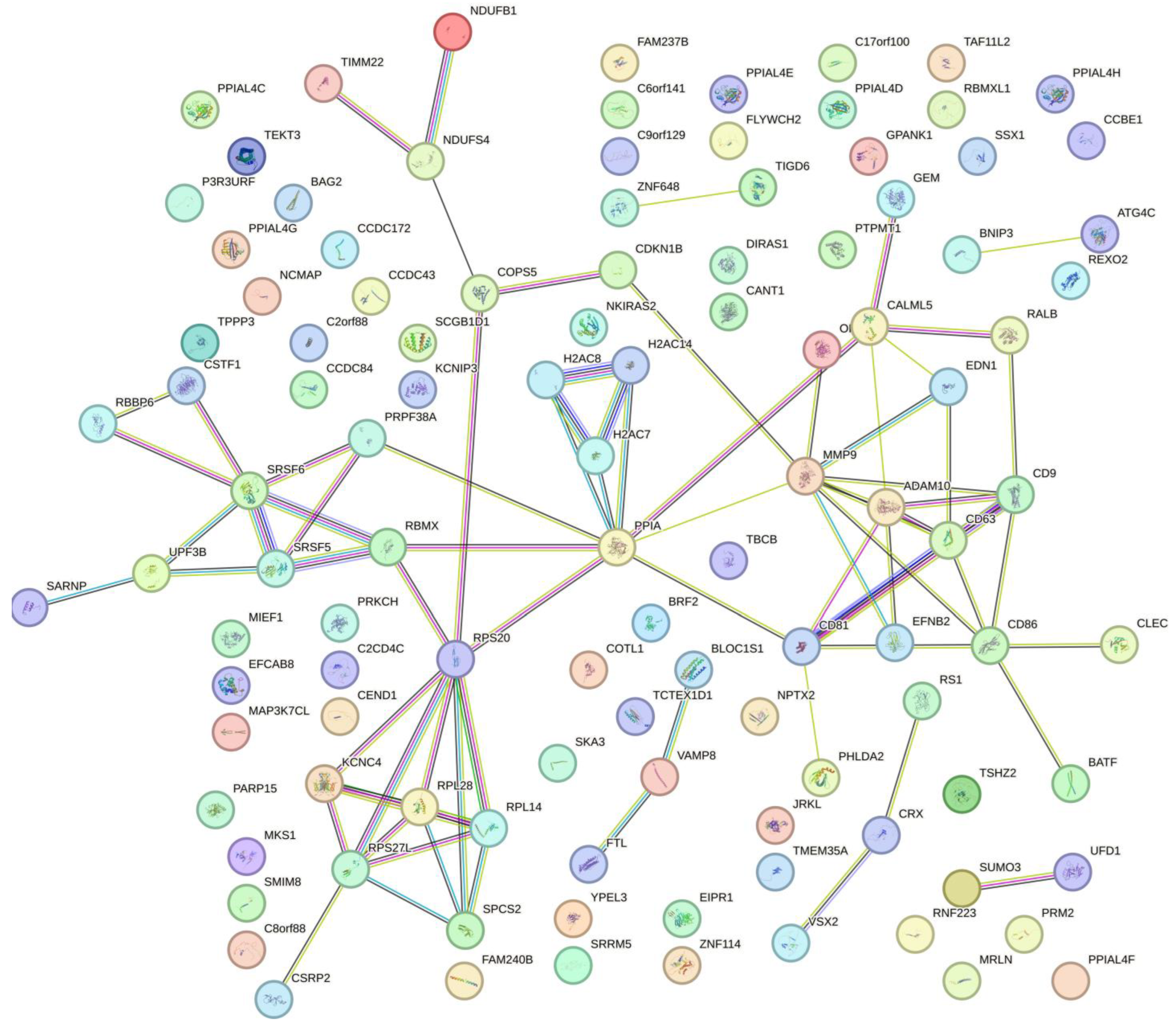
Figure 6.
Venn–Euler diagrams of proteins in exosomes from blood of BCPs with luminal A and triple-positive subtypes with Vesiclepedia databases.
Figure 6.
Venn–Euler diagrams of proteins in exosomes from blood of BCPs with luminal A and triple-positive subtypes with Vesiclepedia databases.
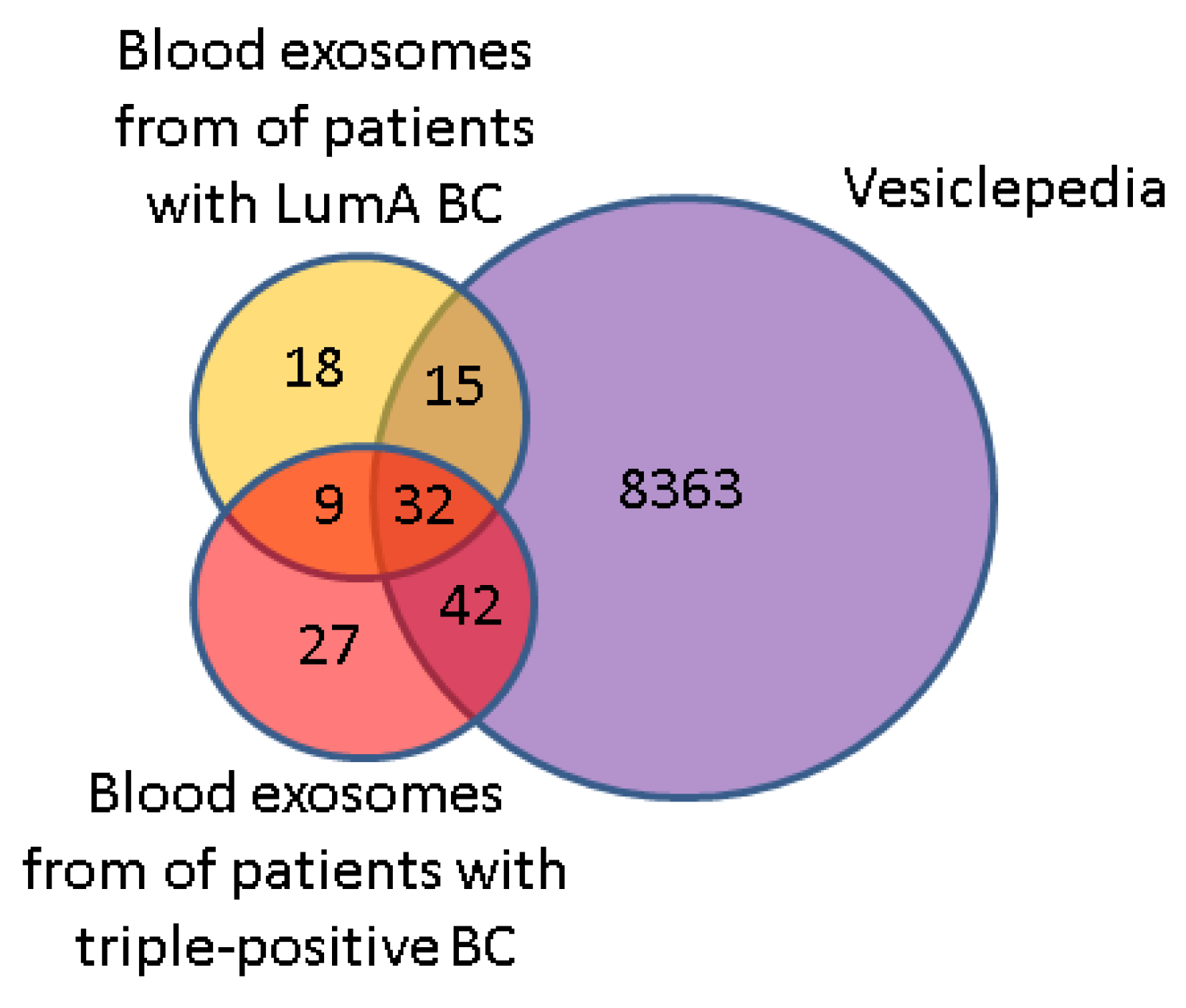
Figure 7.
Venn–Euler diagram of exosomal proteins from cancer cells and BCP blood with luminal A and triple-positive subtypes in dbHPA, composed using QuickGO 2.0 and FunRich 3.13 software.
Figure 7.
Venn–Euler diagram of exosomal proteins from cancer cells and BCP blood with luminal A and triple-positive subtypes in dbHPA, composed using QuickGO 2.0 and FunRich 3.13 software.
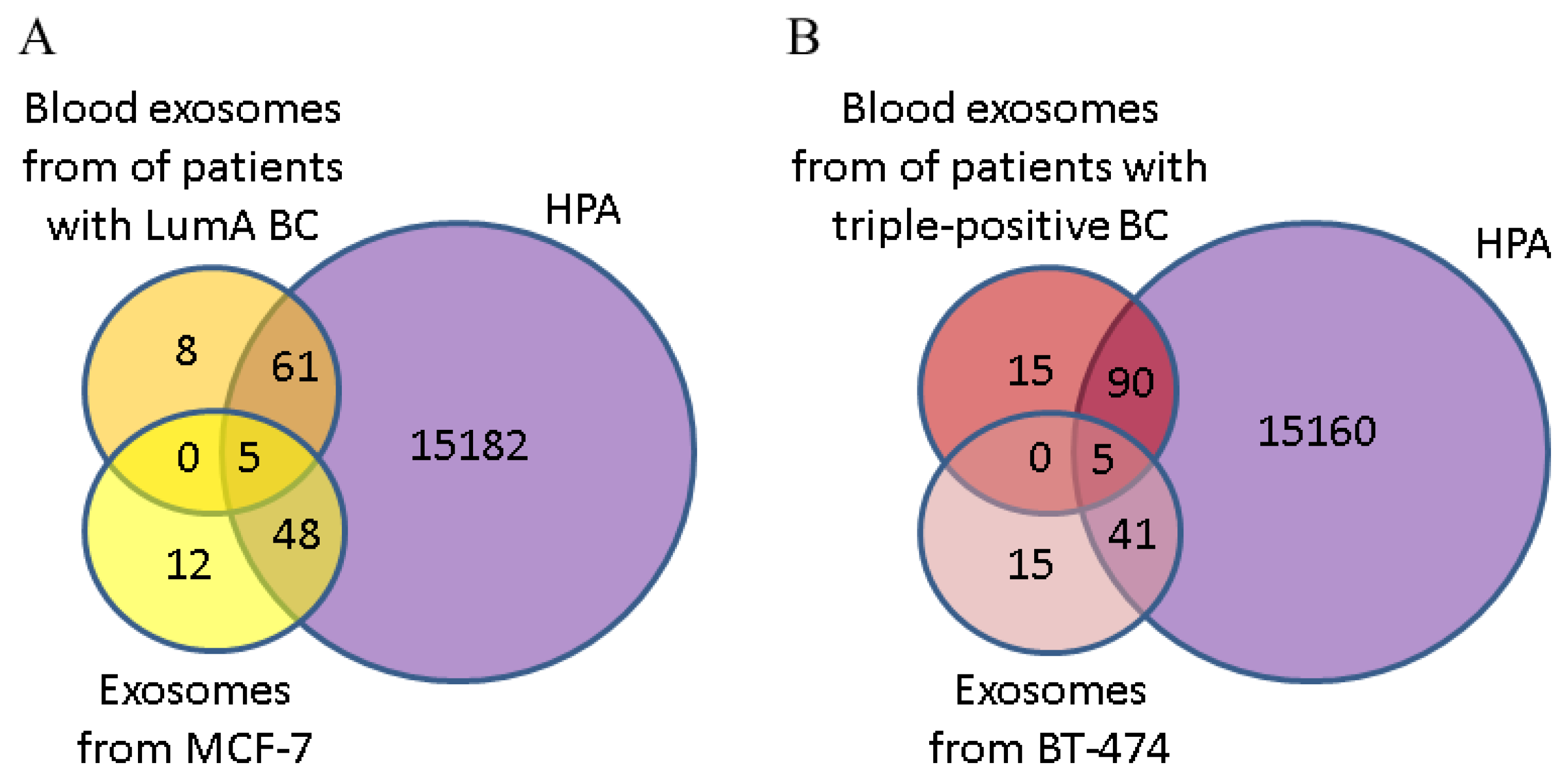

Table 1.
Clinical characteristics of untreated BCPs.
| Subtype | Hormonal status | HER2/neo status | Age | T | N | M | Ki-67 | G | Infiltrative Ductal Carcinoma |
|---|---|---|---|---|---|---|---|---|---|
| Luminal An = 5 | ER+PR+ | Negative | 61 | 1 | 0 | 0 | 10-15% | 2 | Yes |
| 61 | 1 | 0 | 0 | 12-14% | 2 | Yes | |||
| 56 | 2 | 0 | 0 | 5-10% | 2 | Yes | |||
| 59 | 2 | 0 | 0 | 5-10% | 2 | Yes | |||
| 61 | 2 | 1 | 0 | 5% | 2 | Yes | |||
| Triple-positiven = 8 | ER+PR+ | Positive | 52 | 1 | 0 | 0 | 10% | 2 | Yes |
| 62 | 1 | 0 | 0 | 10-15% | 2 | Yes | |||
| 66 | 1 | 0 | 0 | 15% | 2 | Yes | |||
| 68 | 1 | 0 | 0 | 5-10% | 2 | Yes | |||
| 69 | 1 | 0 | 0 | 5-10% | 2 | Yes | |||
| 44 | 2 | 0 | 0 | 5% | 2 | Yes | |||
| 61 | 2 | x* | 0 | 15-17% | 2 | Yes | |||
| 67 | 2 | 0 | 0 | 20-25% | 2 | Yes |
* - no information available.
Table 2.
Analysis of the biological pathways in which the HUVEC-derived exosomal protein is involved.
Table 2.
Analysis of the biological pathways in which the HUVEC-derived exosomal protein is involved.
| Protein name | Biological pathways | P-value |
| ALB | Transport of organic anions | 0,0127 |
| ALB | Recycling of bile acids and salts | 0,0139 |
| ALB | HDL-mediated lipid transport | 0,0189 |
| GTF2F1 | RNA Pol II CTD phosphorylation and interaction with CE | 0,0326 |
| RPS3A, PSMD10 | Metabolism of mRNA | 0,0333 |
| ALB | Bile acid and bile salt metabolism | 0,0338 |
| ALB | Lipoprotein metabolism | 0,0338 |
| CD81, PSMD10 | Adaptive Immune System | 0,0341 |
| GTF2F1 | mRNA Capping | 0,0351 |
| CD63 | Platelet degranulation | 0,0351 |
| MMP9 | Osteopontin-mediated events | 0,0363 |
| ALB | Transport of vitamins, nucleosides, and related molecules | 0,0375 |
| GTF2F1 | Pausing and recovery of elongation | 0,0388 |
| GTF2F1 | Elongation arrest and recovery | 0,0388 |
| GTF2F1 | Formation of the Early Elongation Complex | 0,0400 |
| CD81 | Alpha4 beta1 integrin signaling events | 0,0412 |
| CD9 | a6b1 and a6b4 Integrin signaling | 0,0437 |
| RPS3A, PSMD10 | Metabolism of RNA | 0,0466 |
| GTF2F1 | RNA Polymerase II Transcription Pre-Initiation And Promoter Opening | 0,0486 |
Table 3.
Analysis of the biological pathways in which the MCF-7-derived exosomal protein is involved *.
Table 3.
Analysis of the biological pathways in which the MCF-7-derived exosomal protein is involved *.
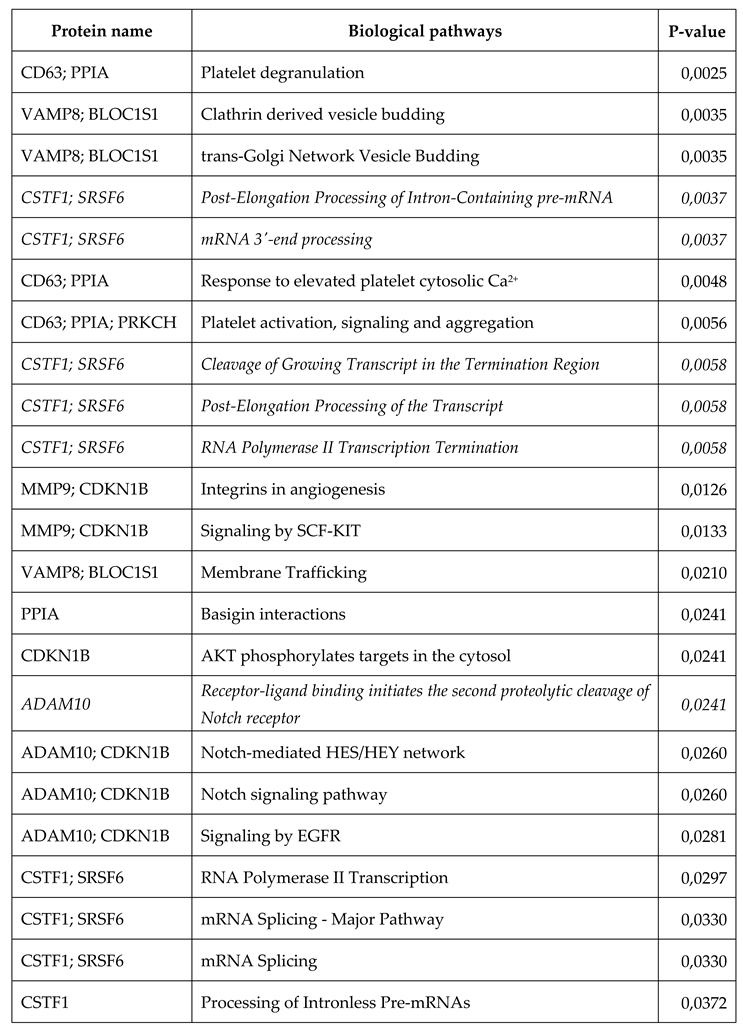 |
* biological pathways universal to exosome proteins secreted by cancer cells are highlighted in italics.
Table 4.
Analysis of the biological pathways in which the BT-474-derived exosomal protein is involved *.
Table 4.
Analysis of the biological pathways in which the BT-474-derived exosomal protein is involved *.
| Protein name | Biological pathways | P-value |
| RPS20; RPL28; UPF3B; RPL14 | Nonsense Mediated Decay Enhanced by the Exon Junction Complex | 0,0008 |
| RPS20; RPL28; SPCS2; RPL14 | Insulin Synthesis and Processing | 0,0019 |
| RBMX; SRSF5; RPS20; RPL28; UPF3B; RPL14 | Gene Expression | 0,0037 |
| RPS20; RPL28; RPL14 | Peptide chain elongation | 0,0045 |
| RPS20; RPL28; RPL14 | Eukaryotic Translation Termination | 0,0045 |
| RPS20; RPL28; RPL14 | Eukaryotic Translation Elongation | 0,0049 |
| RPS20; RPL28; RPL14 | Formation of a pool of free 40S subunits | 0,0062 |
| RPS20; RPL28; RPL14 | Regulation of gene expression in beta cells | 0,0074 |
| RPS20; RPL28; RPL14 | L13a-mediated translational silencing of Ceruloplasmin expression | 0,0082 |
| RPS20; RPL28; RPL14 | GTP hydrolysis and joining of the 60S ribosomal subunit | 0,0084 |
| SRSF5; UPF3B | mRNA 3'-end processing | 0,0085 |
| SRSF5; UPF3B | Post-Elongation Processing of Intron-Containing pre-mRNA | 0,0085 |
| RBMX; SRSF5; UPF3B | mRNA Splicing | 0,0094 |
| RPS20; RPL28; RPL14 | Regulation of beta-cell development | 0,0096 |
| RPS20; RPL28; RPL14 | Eukaryotic Translation Initiation | 0,0101 |
| RPS20; RPL28; RPL14 | Cap-dependent Translation Initiation | 0,0101 |
| RPS20; RPL28; RPL14 | Translation | 0,0119 |
| SRSF5; UPF3B | Post-Elongation Processing of the Transcript | 0,0134 |
| SRSF5; UPF3B | Cleavage of Growing Transcript in the Termination Region | 0,0134 |
| SRSF5; UPF3B | RNA Polymerase II Transcription Termination | 0,0134 |
| RPS20; RPL28; UPF3B; RPL14 | Metabolism of mRNA | 0,0147 |
| SRSF5; UPF3B | Transport of Mature mRNA derived from an Intron-Containing Transcript | 0,0185 |
| RBMX; SRSF5; UPF3B | Processing of Capped Intron-Containing Pre-mRNA | 0,0186 |
| TBCB; RPS20; RPL28; RPL14 | Metabolism of proteins | 0,0206 |
| RBMX; SRSF5; UPF3B | mRNA Processing | 0,0260 |
| RPS20; RPL28; UPF3B; RPL14 | Metabolism of RNA | 0,0268 |
| SRSF5; BRF2; UPF3B | Transcription | 0,0359 |
| ADAM10 | Receptor-ligand binding initiates the second proteolytic cleavage of Notch receptor | 0,0366 |
| NDUFS4; NDUFB1 | Respiratory electron transport | 0,0388 |
| RBMX; SRSF5; UPF3B | Formation and Maturation of mRNA Transcript | 0,0396 |
| MMP9; CD86; RALB | CXCR4-mediated signaling events | 0,0424 |
| BNIP3; EDN1 | Hypoxic and oxygen homeostasis regulation of HIF-1-alpha | 0,0426 |
| TBCB | Post-chaperonin tubulin folding pathway | 0,0485 |
* Biological pathways universal to exosome proteins secreted by cancer cells are highlighted in italics.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



