
Submitted:
02 December 2024
Posted:
05 December 2024
You are already at the latest version
Abstract
Colorectal cancer (CRC) is a leading cause of cancer-related deaths, largely due to metastasis, particularly to the liver, and the limited understanding of the molecular mechanisms underlying this process. In this study, we focused on investigating the role of long non-coding RNAs (lncRNAs) as key regulatory factors in CRC progression and metastasis to liver tissues. Using high-throughput sequencing and microarray approaches, we analyzed gene expression profiles from two independent lncRNA datasets to identify potential players involved in liver metastasis.
Our findings revealed five lncRNAs—PROX1-AS1, SOX9-AS1, LINC01594, LINC01555, and APOA1-AS—previously known for their roles in CRC progression, now identified as being involved in the liver metastatic process. Additionally, 20 other lncRNAs, including VCAN-AS1, SYP-AS1, SMIM2-IT1, NCOA7-AS1, and LINC01449, were also identified as potential contributors to CRC liver metastasis. Notably, two lncRNAs—SATB2-AS1 and LINC01116—emerged as common candidates across both datasets, suggesting their significant role in promoting CRC metastasis to the liver. These two lncRNAs hold promise as molecular targets for therapeutic and diagnostic development.
In conclusion, our study uncovers a novel layer of regulatory mechanisms involving lncRNAs in CRC liver metastasis. These findings advance our understanding of the molecular behaviors that drive CRC progression and offer new avenues for targeted therapeutic strategies and diagnostic tools, particularly for liver metastasis in CRC.
Keywords:
Colorectal cancer (CRC)
; Liver metastasis
; Long non-coding RNAs (lncRNAs)
; Gene expression profiling
; Therapeutic targets
1. Introduction
Colorectal cancer (CRC) is one of the most prevalent malignancies globally and a leading cause of cancer-related deaths. Despite advancements in treatment strategies, the prognosis for patients with advanced or metastatic CRC remains poor. A key challenge in combating CRC is the insufficient understanding of the molecular mechanisms that drive tumor initiation, progression, and metastasis, particularly at the genetic and regulatory levels. This lack of knowledge limits the discovery of novel therapeutic targets and impedes the development of effective diagnostic tools, making CRC an often-untreatable condition in its advanced stages [1,2].
In this study, we aim to fill this critical knowledge gap by investigating the molecular behaviors and pathways that contribute to CRC progression, with a particular focus on metastasis to the liver—one of the most common and life-threatening metastatic sites for CRC. Our primary goal is to identify potential therapeutic and diagnostic targets by delving into the molecular underpinnings of CRC at a high resolution. To achieve this, we employed cutting-edge high-throughput sequencing technologies, coupled with microarray-based gene expression profiling, to systematically analyze the vast landscape of genes implicated in CRC [3-5].
Given the complexity of cancer biology and the sheer volume of gene expression data generated, we utilized systems biology approaches to streamline our analysis. By integrating and analyzing thousands of gene expression profiles, we aimed to uncover key molecular players—particularly those involved in regulatory networks that influence CRC metastasis. A major focus of our research is on the role of regulatory factors, especially long non-coding RNAs (lncRNAs), which have emerged as important regulators of gene expression and cancer progression. Through upstream regulatory analysis, we sought to gain deeper insights into the function of lncRNAs and their potential as novel therapeutic or diagnostic targets in CRC [6-10].
By shedding light on these critical molecular mechanisms, our study seeks to contribute to a better understanding of CRC at the molecular level, potentially paving the way for the development of more effective, targeted interventions for CRC patients. Our findings may offer new avenues for therapeutic strategies aimed at halting CRC progression and improving patient outcomes, particularly in cases of liver metastasis.
2. Materials and Methods:
2.1. Expression Profile Analysis
In this study, we utilized the GSE148918 dataset from the Gene Expression Omnibus (GEO) to identify differentially expressed genes (DEGs). The analysis was conducted using R programming with packages such as Limma and DESeq2. A threshold of ±1 for log fold change and a p-value ≤ 0.05 were set as the criteria for significant gene expression changes (Figure 1A). To validate these findings, we employed another dataset (GSE174602), obtained through microarray techniques. This dataset was also analyzed in R, using the same thresholds as in the initial analysis [11-13].
2.2. Upstream Analysis
Identifying DEGs helps to understand which genes are altered during the progression of metastatic colorectal cancer to liver tissue, but it is crucial to determine the factors regulating these genes. Several regulatory elements, such as transcription factors (TFs), microRNAs, and protein kinases, play pivotal roles in this process. While many of these regulators have been extensively studied in cancer, others, such as long non-coding RNAs (lncRNAs), remain less understood, particularly in metastatic cancers. In this study, we used the lncHUB_lncRNA_Co-Expression database from the EnrichR web tool (https://maayanlab.cloud/Enrichr/) (Fig.1E). This database revealed lncRNAs that are co-expressed with our candidate genes, providing insights into their potential regulatory roles [14].
2.3. Protein Network Construction
This section aims to study protein interactions following transcriptional changes. We utilized the STRING database to source protein-protein interaction data, and Cytoscape software was employed to visualize and analyze the network. The protein network was constructed using the default settings of the STRING-db package within Cytoscape (Fig.1C). Given the complexity of the network, we applied the CentiScape plug-in to calculate betweenness centrality (Fig.1D), which measures the importance of each node within the network. Nodes with the highest betweenness scores were identified as hub genes, likely playing significant roles in the metastatic progression from primary colorectal cancer to liver metastasis [15-18].
2.4. DE-LncRNAs
The results from the upstream analysis were verified by comparing them with the expression profiles obtained in the current study. Specifically, the lncRNA database results were cross-referenced with our identified DEGs (Fig.1G). To further strengthen these findings, a second dataset of lncRNAs derived from microarray techniques was identified and re-analyzed. The outcomes from this second dataset were then compared to the DEGs found in the current study, reinforcing the robustness of our results (Fig.1H).
3. Results
3.1. Differential Expression Genes
We analyzed 12 samples from the GSE148918 dataset, which includes 8 Organoid_CRC samples and 4 Organoid_Liver samples (Supplementary Figure 1). The dataset contains 64,099 genes, comprising both mRNA and LncRNA (Supplementary Table 1). Of these, only 1,921 genes met the significance criteria, with a fold change of less than -1 or more than 1, and a p-value of ≤ 0.05. Among the 1,921 genes, 1,118 were overexpressed in liver metastatic samples, while 805 genes were downregulated (Figure 2) (Supplementary Table 2).
To validate our LncRNA results, we used an additional dataset, GSE147602, which consists of 20 samples: 10 CRC samples and 10 liver metastatic samples. Following Principal Component Analysis (PCA), 4 noisy CRC samples and 1 noisy liver sample were removed, leaving 15 samples for further analysis (Supplementary Figure 2). These samples contained 38,213 genes each (Supplementary Table 3). After filtration, we identified 2,963 significant genes. Of these, 1,468 showed overexpression, while 1,495 were downregulated in liver metastatic samples (Supplementary Figure 3) (Supplementary Table 4).
3.2. Construction of PPI Network
A total of 1,921 genes were used to construct the protein-protein interaction (PPI) network. The STRING database identified 1,607 proteins, with 159 proteins found to have no connections. This indicates that while these nodes were identified in the database, their interactions with other genes in our dataset were not found. As a result, these unconnected nodes were excluded from further analysis. The final network consisted of 1,448 nodes and 8,748 edges (Figure 3).
Given the complexity of the network, we needed a method to evaluate the interactions of each node and determine which nodes are more critical. To achieve this, we used the betweenness centrality criterion. The top 10 nodes, including FN1, JUN, and SOX2, were identified as hub genes, playing a significant role in our network. These genes are particularly important in the later stages of liver metastasis compared to the earlier stages in colon tissue (Table 1).
3.3. Upstream analysis
After identifying the alterations, the next important question was to understand how these alterations occurred. To address this, we performed upstream analysis within the molecular biology framework. Using 1,921 DEGs, we queried an lncRNA database to identify potential interactions, finding approximately 400 lncRNAs that met the significance threshold of a p-value ≤ 0.05. When comparing these with the 1,921 DEGs, only 23 lncRNAs were identified as differentially expressed lncRNAs (DE-lncRNAs). We also mapped their range of activity by constructing an interaction network (Fig.1I).
Additionally, we utilized other databases to validate the influence of these lncRNAs on the 1,921 DEGs. From the second dataset, 2,963 DEGs were identified, and when compared to the ~400 lncRNAs, only 16 lncRNAs were found to be DE-lncRNAs. Interestingly, both datasets revealed two common lncRNAs, LINC01116 and SATB2-AS1, which exhibited significant alterations in expression between primary and liver metastatic tumor samples. LINC01116 showed a 5-fold increase in expression, while SATB2-AS1 was upregulated by 7-fold (Table 2) (Figure 4).
4. Discussion
Colorectal cancer is one of the major causes of mortality in the Iranian population. A key approach to addressing this issue is halting the progression of the disease. To achieve this, a comprehensive understanding of the molecular players involved in the progression is essential, with metastasis being the primary mechanism of concern. This study aims to address this problem by investigating the molecular mechanisms at play, particularly through upstream analysis focusing on lncRNA regulation, using high-throughput techniques such as RNA-seq and microarray datasets.
LINC00365 has been reported to function as a tumor suppressor by inhibiting HIF-1α-mediated glucose metabolism reprogramming in breast cancer, as demonstrated by Liu et al. in 2023 [19]. Additionally, Taghehchian et al. (2022) found LINC00365 to be overexpressed in gastric cancers, influencing the epithelial-mesenchymal transition (EMT) mechanism [20]. Zhu et al. (2020) further identified LINC00365 as a promoter of colorectal cancer progression [21]. Collectively, these studies highlight the role of LINC00365 across various cancer types, with its overexpression being linked to colorectal cancer progression.
However, in the current study, LINC00365 was found to be downregulated in liver tissue, where it displayed 26 interactions (Table 2), the highest number of interactions among the lncRNAs identified in the PPI network. Based on these findings, LINC00365 appears to play a barrier role in preventing metastasis to liver tumors.
In addition, LINC00278 has previously been reported to play a role in metastatic liver tumors in colorectal cancer (CRC) [22,23]. In our study, LINC00278 was overexpressed and exhibited 22 interactions in the PPI network. Similarly, LINC01559 was also overexpressed in liver metastatic tumors, with 21 regulatory interactions in the protein network (Table 2). A previous study reported that LINC01559 suppresses CRC progression by regulating miR-106b [24], and it has also been linked to triple-negative breast cancer, as shown by Yang et al. in 2022 [25]. Therefore, LINC01559 is proposed as a candidate lncRNA in liver metastatic cancers.
Furthermore, Yao et al. (2024) reported that LINC01133 could inhibit the progression of CRC to metastatic liver by regulating miR-186 [26]. In our study, LINC01133 was identified as a DE-lncRNA and was downregulated in metastatic liver tumors (Table 2), which aligns with its role in inhibiting metastasis. This downregulation supports the findings of our methodology, highlighting LINC01133’s inhibitory role in liver metastasis from CRC.
Furthermore, Yao et al. (2024) reported that LINC01133 could inhibit the progression of CRC to metastatic liver by regulating miR-186 [26] . In our study, LINC01133 was identified as a DE-lncRNA and was downregulated in metastatic liver tumors, which aligns with its role in inhibiting metastasis. This downregulation supports the findings of our methodology, highlighting LINC01133’s inhibitory role in liver metastasis from CRC.
LINC02826 has been implicated in hepatocellular carcinoma [27], and in our study, it was highly downregulated during the metastatic process to the liver in CRC samples, with 21 interactions in the PPI network (Table 2). These observations suggest that LINC02826 may be involved in the metastatic process, given its significant alteration from primary to metastatic liver tumors and its interactions. Therefore, LINC02826 is proposed as a candidate lncRNA for further study in CRC progression to liver metastasis.
Moreover, the relationship between CLCA4-AS1 and head and neck squamous carcinoma was reported by Liao in 2024 [28]. In our study, CLCA4-AS1 was identified as a DE-lncRNA, downregulated in liver metastatic tumors, with 21 connections in the PPI network (Table 2), suggesting its regulatory role in the metastatic process of CRC to the liver.
LINC00462, one of our DE-lncRNAs, may also be related to the progression network, as it displayed 20 interactions and was highly downregulated (Table 2). Previous studies by Zhang et al. in 2021 identified LINC00462 as a biomarker for liver cancers [29]. Therefore, we propose, for the first time, that LINC00462 may be involved in the progression of CRC to liver metastasis.
Moghadami et al. previously reported VLDLR-AS1 as a biomarker for CRC due to its downregulation in healthy samples [30]. In our study, VLDLR-AS1 was overexpressed in metastatic tumors compared to primary CRC tumors, with 19 interactions in the progression network (Table 2), suggesting its potential role in the metastatic progression to the liver and its value as a therapeutic target.
Additionally, PROX1-AS1 was identified as a DE-lncRNA in our study, with 19 interactions in the progression network and overexpression in liver metastatic tumors compared to primary CRC tumors (Table 2). Liu et al. previously reported in January 2023 that PROX1-AS1 plays a role in CRC progression mechanisms [31]. Our study extends this by demonstrating its role in CRC progression to liver metastasis. This finding suggests that PROX1-AS1 could be a potential molecular therapy target to reduce CRC progression to liver tissue.
The lncRNA PRICKLE2-DT was found to be highly overexpressed (5-fold change) in liver metastatic tumors, with 19 interactions within the progression network (Table 2). This suggests that PRICKLE2-DT could play a novel role in the progression of CRC metastasis to liver tissue, marking its potential importance for the first time in this context.
LINC00887 was previously reported to be involved in liver metastasis progression by Liao et al. in September 2024 [32]. This supports our methodology, which identified LINC00887 as a DE-lncRNA influencing the PPI network through 18 interactions (Table 2), further validating its potential as a key player in metastatic progression. This finding encourages attention to other candidate lncRNAs discovered in this study.
EVX1-AS was reported as a molecular driver in colon cancer progression by Gao in 2021 [33], and our study confirmed its role as a DE-lncRNA in CRC liver metastasis with 18 connections in the current PPI network analysis (Table 2). Similarly, LINC02418, previously identified in 2020 by Tian et al. as associated with CRC progression [34], was also found to be a DE-lncRNA in this study with 17 interactions, highlighting its relevance to the metastatic process in liver tissue. Additionally, LINC00957 was linked to CRC progression in 2019 by Zhang et al. [35], further supporting its potential role in this context.
SOX9-AS1, known for its role in regulating SOX8 and promoting proliferation in hepatocellular cancer cells (HCC) via miR-5590-3p, was reported by Moghadam et al. in 2021 [36]. In our study, SOX9-AS1 was identified as a DE-lncRNA driving progression from CRC to liver metastasis through 16 interactions in the PPI network (Table 2).
LINC01594 was reported as a CRC metastatic biomarker by Liu et al. in 2023 [37]. In our analysis, LINC01594 was downregulated in liver metastatic tumors, with 16 interactions in the metastatic PPI network (Table 2), making it a promising target for suppressing metastasis or serving as a diagnostic biomarker for liver involvement.
Three lncRNAs—LHFPL3-AS2, NEUROG2-AS1, and ADAMTS19-AS1—were identified as DE-lncRNAs involved in CRC progression from primary to liver metastatic tumors, interacting with the PPI network through 17, 17, and 18 connections, respectively (Table 2).
Our analysis also identified 16 DE-lncRNAs from the second dataset, with 14 being unique. Among these, LINC00607 was found to be downregulated in liver metastatic tumors, with 16 interactions in the progression network (Table 2). Its role in tumor progression was previously reported by Dong in hepatocellular carcinoma cells in 2023 [38]. Similarly, LINC00114 was downregulated in metastatic CRC liver tissue, with 18 interactions in the PPI network (Table 2), aligning with Qin et al.'s 2022 findings that LINC00114 plays a role in liver tumor progression via EZH2 [39].
Additionally, LINC01555 and APOA1-AS were identified as DE-lncRNAs in our study, with 18 and 17 interactions in the CRC progression network (Table 2), respectively. Both have been suggested as potential therapeutic targets to prevent metastasis from primary CRC tumors based on previous studies in 2022 and 2024 [40,41].
Among the 10 other DE-lncRNAs identified in our analysis—such as VCAN-AS1 and LINC00626 (upregulated), and SYP-AS1, SMIM2-IT1, NCOA7-AS1, LINC01449, LINC00974, LINC00440, LINC00379, and ERVH-1 (downregulated)—no previous reports link them directly to CRC liver metastasis (Table 2). Given their significant differential expression and their direct interactions within the progression network, these lncRNAs may play roles in CRC metastasis to the liver. Further experimental validation, such as through RT-PCR, is recommended to confirm their involvement.
SATB2-AS1, a known suppressor of metastasis in CRC and other cancers, was found to be downregulated in our study, with 25 direct connections in the progression network. Although SATB2-AS1 typically inhibits metastasis, its minimal downregulation (-1 logFC) in this study suggests it may play a nuanced role in the early stages of CRC metastasis to the liver. Further investigation using experimental approaches is warranted to clarify SATB2-AS1’s exact function.
We identified two shared lncRNAs that influenced the PPI network during the transition from primary colon tumors to metastatic liver tumors. One of them, SATB2-AS1, is a known suppressor of metastasis in colorectal cancer, where its overexpression has been associated with reduced tumor aggression and metastatic potential [42,43]. Additionally, in hepatocellular carcinoma, SATB2-AS1 was reported by Huang in 2023 to play a developmental role via the miR3678-3b pathway [44]. In our study, SATB2-AS1 was one of the two shared DE-lncRNAs, interacting with 25 direct connections in the progression network (Table 2), linking primary CRC tumors to liver metastases. Interestingly, SATB2-AS1 was downregulated in our analysis (Table 2), which might suggest its involvement in the metastatic process, possibly indicating a shift in its regulatory function as the tumor progresses from the primary site to the liver. The slight downregulation (-1 logFC) hints at a nuanced role in this transition. These findings suggest the need for further assessment of SATB2-AS1's exact role in the progression of colorectal cancer to liver metastasis, with future studies utilizing laboratory techniques like RT-PCR to better understand its function.
Long intergenic non-protein coding RNA 1116 (LINC01116) is a long non-coding RNA (lncRNA), a type of RNA over 200 nucleotides in length that is not translated into proteins. Aberrant expression of LINC01116 has been associated with several cancers, including lung cancer, gastric cancer, colorectal cancer (CRC), glioma, and osteosarcoma. It plays a critical role in promoting cell proliferation, invasion, migration, and apoptosis [45]. Previous studies have shown that inhibiting LINC01116 can reduce tumor progression in CRCs [46]. Furthermore, LINC01116's role in hepatocellular carcinoma (HCC) was highlighted by Kun Wang in 2023 [47]. In our study, LINC01116 was found to be overexpressed and involved in 16 interactions within the progression network from CRC to liver metastasis (Table 2). This positions LINC01116 as a strong candidate for further investigation into its role in metastatic CRC to liver tumors. Future experimental studies should focus on inhibiting its expression and monitoring its effects on tumor progression in liver metastasis.
In summary, our study provides a comprehensive analysis of lncRNAs involved in the progression of colorectal cancer (CRC) to liver metastasis. By integrating data from multiple datasets, we identified several differentially expressed lncRNAs (DE-lncRNAs) with significant roles in the metastatic process, such as LINC01116 and SATB2-AS1. These lncRNAs not only interact within key protein-protein interaction (PPI) networks but also present potential as diagnostic biomarkers and therapeutic targets. The results suggest that these lncRNAs play vital roles in regulating CRC progression, making them promising candidates for future research aimed at disrupting the metastatic process.
5. Conclusion
In this study, we aimed to deepen our understanding of the regulatory factors, particularly long non-coding RNAs (lncRNAs), and their roles in the progression and metastasis of colorectal cancer (CRC) to liver tissues. Our analysis identified five lncRNAs—PROX1-AS1, SOX9-AS1, LINC01594, LINC01555, and APOA1-AS—which were previously known for their roles in CRC progression but, for the first time, were shown to affect the liver metastatic process. Additionally, from two lncRNA expression datasets, we identified 20 novel lncRNAs (VCAN-AS1, SYP-AS1, SMIM2-IT1, NCOA7-AS1, LINC01449, LINC00974, LINC00626, LINC00440, LINC00379, ERVH-1, VLDLR-AS1, PRICKLE2-DT, ADAMTS19-AS1, NEUROG2-AS1, LHFPL3-AS2, LINC00365, LINC02826, LINC01559, CLCA4-AS1, LINC02563, and LINC00462) that may play a role in CRC progression and liver metastasis.
Moreover, two lncRNAs—SATB2-AS1 and LINC01116—emerged as the only common candidates across both datasets, suggesting a potential role in the metastatic process from primary CRC tumors to liver tissues. This makes them promising targets for molecular therapy and diagnostic applications.
In summary, our findings highlight a new dimension of regulatory mechanisms driven by lncRNAs in the metastatic network of CRC to liver tissues. Several novel molecular players involved in this progression process from primary CRC to liver metastasis were uncovered, contributing to our understanding of molecular behaviors. These insights pave the way for designing new therapeutic or diagnostic strategies focused on CRC progression, particularly in liver metastasis.
Author Contributions
Conceptualization, A.S. and A.P.; methodology, A.S.; validation, M.K. and A.S.; formal analysis, M.K.; investigation, M.K. and A.S.; resources, A.P. and A.S.; data curation, M.K. and A.S.; writing—original draft preparation, M.K. and A.S.; writing—review and editing, A.S.; visualization, M.K. and A.S.; supervision, A.P. and A.S.; project administration, A.S.; funding acquisition, A.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no funding
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
The authors of the present study are grateful for the assistance of the Arak University and Omicsdade Institute in improving the forthcoming research.
Confilicts of Interest: The authors declare no competing interests.
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Van Cutsem, E.; Cervantes, A.; Adam, R.; Sobrero, A.; Van Krieken, J.H.; Aderka, D.; Aranda Aguilar, E.; Bardelli, A.; Benson, A.; Bodoky, G.; et al. ESMO consensus guidelines for the management of patients with metastatic colorectal cancer. Ann Oncol 2016, 27, 1386–1422. [Google Scholar] [CrossRef] [PubMed]
- Manfredi, S.; Lepage, C.; Hatem, C.; Coatmeur, O.; Faivre, J.; Bouvier, A.M. Epidemiology and management of liver metastases from colorectal cancer. Ann Surg 2006, 244, 254–259. [Google Scholar] [CrossRef]
- Mardis, E.R. Next-generation sequencing platforms. Annu Rev Anal Chem (Palo Alto Calif) 2013, 6, 287–303. [Google Scholar] [CrossRef]
- Lockhart, D.J.; Dong, H.; Byrne, M.C.; Follettie, M.T.; Gallo, M.V.; Chee, M.S.; Mittmann, M.; Wang, C.; Kobayashi, M.; Horton, H.; et al. Expression monitoring by hybridization to high-density oligonucleotide arrays. Nat Biotechnol 1996, 14, 1675–1680. [Google Scholar] [CrossRef]
- Peng, W.X.; Koirala, P.; Mo, Y.Y. LncRNA-mediated regulation of cell signaling in cancer. Oncogene 2017, 36, 5661–5667. [Google Scholar] [CrossRef]
- Schmitt, A.M.; Chang, H.Y. Long Noncoding RNAs in Cancer Pathways. Cancer Cell 2016, 29, 452–463. [Google Scholar] [CrossRef]
- Kitano, H. Systems biology: a brief overview. Science 2002, 295, 1662–1664. [Google Scholar] [CrossRef]
- Hozhabri, H.; Ghasemi Dehkohneh, R.S.; Razavi, S.M.; Razavi, S.M.; Salarian, F.; Rasouli, A.; Azami, J.; Ghasemi Shiran, M.; Kardan, Z.; Farrokhzad, N.; et al. Comparative analysis of protein-protein interaction networks in metastatic breast cancer. PLoS One 2022, 17, e0260584. [Google Scholar] [CrossRef]
- Rasti, A.; Abazari, O.; Dayati, P.; Kardan, Z.; Salari, A.; Khalili, M.; Motlagh, F.M.; Modarressi, M.H. Identification of Potential Key Genes Linked to Gender Differences in Bladder Cancer Based on Gene Expression Omnibus (GEO) Database. Adv Biomed Res 2023, 12, 157. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic acids research 2015, 43, e47–e47. [Google Scholar] [CrossRef] [PubMed]
- Love, M.; Anders, S.; Huber, W. Differential analysis of count data–the DESeq2 package. Genome Biol 2014, 15, 10–1186. [Google Scholar]
- Huang, P.; Deng, W.; Bao, H.; Lin, Z.; Liu, M.; Wu, J.; Zhou, X.; Qiao, M.; Yang, Y.; Cai, H.; et al. SOX4 facilitates PGR protein stability and FOXO1 expression conducive for human endometrial decidualization. Elife 2022, 11. [Google Scholar] [CrossRef]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma'ayan, A. Enrichr: interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinformatics 2013, 14, 128. [Google Scholar] [CrossRef]
- Mering, C.v.; Huynen, M.; Jaeggi, D.; Schmidt, S.; Bork, P.; Snel, B. STRING: a database of predicted functional associations between proteins. Nucleic acids research 2003, 31, 258–261. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome research 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P. The STRING database in 2021: customizable protein–protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic acids research 2021, 49, D605–D612. [Google Scholar] [CrossRef]
- Scardoni, G.; Petterlini, M.; Laudanna, C. Analyzing biological network parameters with CentiScaPe. Bioinformatics 2009, 25, 2857–2859. [Google Scholar] [CrossRef]
- Liu, B.; Qu, X.; Wang, J.; Xu, L.; Zhang, L.; Xu, B.; Su, J.; Bian, X. LINC00365 functions as a tumor suppressor by inhibiting HIF-1α-mediated glucose metabolism reprogramming in breast cancer. Experimental Cell Research 2023, 425, 113514. [Google Scholar] [CrossRef]
- Taghehchian, N.; Farshchian, M.; Mahmoudian, R.A.; Asoodeh, A.; Abbaszadegan, M.R. The expression of long non-coding RNA LINC01389, LINC00365, RP11-138J23. 1, and RP11-354K4. 2 in gastric cancer and their impacts on EMT. Molecular and Cellular Probes 2022, 66, 101869. [Google Scholar] [CrossRef]
- Zhu, Y.; Bian, Y.; Zhang, Q.; Hu, J.; Li, L.; Yang, M.; Qian, H.; Yu, L.; Liu, B.; Qian, X. LINC00365 promotes colorectal cancer cell progression through the Wnt/β-catenin signaling pathway. Journal of Cellular Biochemistry 2020, 121, 1260–1272. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Du, X.; Chen, M.; Han, L.; Sun, J. Novel insight into the functions of N 6-methyladenosine modified lncRNAs in cancers. International journal of oncology 2022, 61, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Man, C.-F.; He, R.; He, L.; Huang, J.-B.; Xiang, S.-Y.; Dai, Z.; Wang, X.-Y.; Fan, Y. The interaction between N6-methyladenosine modification and non-coding RNAs in gastrointestinal tract cancers. Frontiers in Oncology 2022, 11, 784127. [Google Scholar] [CrossRef]
- Shi, K.; Yang, S.; Chen, C.; Shao, B.; Guo, Y.; Wu, X.; Zhao, L.; Yang, X.; Zhang, Q.; Yuan, W. RNA methylation-mediated LINC01559 suppresses colorectal cancer progression by regulating the miR-106b-5p/PTEN axis. International journal of biological sciences 2022, 18, 3048. [Google Scholar] [CrossRef]
- Yang, X.; Yang, Y.; Qian, X.; Xu, X.; Lv, P. Long non-coding RNA LINC01559 serves as a competing endogenous RNA accelerating triple-negative breast cancer progression. biomedical journal 2022, 45, 512–521. [Google Scholar] [CrossRef]
- Yao, Y.; Zhang, F.; Liu, F.; Xia, D. Propofol-induced LINC01133 inhibits the progression of colorectal cancer via miR-186-5p/NR3C2 axis. Environmental Toxicology 2024, 39, 2265–2284. [Google Scholar] [CrossRef]
- Zhang, P.; Chen, L.; Wu, S.; Ye, B.; Chen, C.; Shi, L. Construction of a metabolism-related long non-coding RNAs-based risk score model of hepatocellular carcinoma for prognosis and personalized treatment prediction. Pathology and Oncology Research 2022, 28, 1610066. [Google Scholar] [CrossRef]
- Liao, H.; He, B. Predictive value of cuproptosis and disulfidptosis-related lncRNA in head and neck squamous cell carcinoma prognosis and treatment. Heliyon 2024. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, R.; Sun, L.; Hu, X. An lncRNA model for predicting the prognosis of hepatocellular carcinoma patients and ceRNA mechanism. Frontiers in Molecular Biosciences 2021, 8, 749313. [Google Scholar] [CrossRef]
- Moqadami, A.; Ahmadi, A.; Khalaj-Kondori, M. lncRNA VLDLR-AS1 Gene Expression in Colorectal Cancer in Patients from East Azerbaijan Province, Iran. Jentashapir J Cell Mol Biol 2023, 14, e141522. [Google Scholar] [CrossRef]
- Liu, J.; Zhan, W.; Chen, G.; Yan, S.; Chen, W.; Li, R. SP1-induced PROX1-AS1 contributes to tumor progression by regulating miR-326/FBXL20 axis in colorectal cancer. Cellular Signalling 2023, 101, 110503. [Google Scholar] [CrossRef] [PubMed]
- Liao, M.; Zheng, W.; Wang, Y.; Li, M.; Sun, X.; Liu, N.; Yao, J.; Dong, F.; Wang, Q.; Ma, Y.; et al. LINC00887 promotes GCN5-dependent H3K27cr level and CRC metastasis via recruitment of YEATS2 and enhancing ETS1 expression. Cell Death & Disease 2024, 15, 711. [Google Scholar] [CrossRef]
- Gao, M.; Guo, Y.; Xiao, Y.; Shang, X. Comprehensive analyses of correlation and survival reveal informative lncRNA prognostic signatures in colon cancer. World journal of surgical oncology 2021, 19, 1–15. [Google Scholar] [CrossRef]
- Tian, J.; Cui, P.; Li, Y.; Yao, X.; Wu, X.; Wang, Z.; Li, C. LINC02418 promotes colon cancer progression by suppressing apoptosis via interaction with miR-34b-5p/BCL2 axis. Cancer Cell International 2020, 20, 460. [Google Scholar] [CrossRef]
- Zhang, L.H.; Li, L.H.; Zhang, P.F.; Cai, Y.F.; Hua, D. LINC00957 Acted as Prognostic Marker Was Associated With Fluorouracil Resistance in Human Colorectal Cancer. Frontiers in Oncology 2019, 9. [Google Scholar] [CrossRef]
- Khashkhashi Moghadam, S.; Bakhshinejad, B.; Khalafizadeh, A.; Mahmud Hussen, B.; Babashah, S. Non-coding RNA-associated competitive endogenous RNA regulatory networks: Novel diagnostic and therapeutic opportunities for hepatocellular carcinoma. J Cell Mol Med 2022, 26, 287–305. [Google Scholar] [CrossRef]
- Liu, B.; Song, A.; Gui, P.; Wang, J.; Pan, Y.; Li, C.; Li, S.; Zhang, Y.; Jiang, T.; Xu, Y.; et al. Long noncoding RNA LINC01594 inhibits the CELF6-mediated splicing of oncogenic CD44 variants to promote colorectal cancer metastasis. Cell Death & Disease 2023, 14, 427. [Google Scholar] [CrossRef]
- Dong, S.; Wang, W.; Liao, Z.; Fan, Y.; Wang, Q.; Zhang, L. MYC-activated LINC00607 promotes hepatocellular carcinoma progression by regulating the miR-584-3p/ROCK1 axis. The Journal of Gene Medicine 2023, 25, e3477. [Google Scholar] [CrossRef]
- Qin, J.; Li, Y.; Li, Z.; Qin, X.; Zhou, X.; Zhang, H.; Li, S. LINC00114 stimulates growth and glycolysis of esophageal cancer cells by recruiting EZH2 to enhance H3K27me3 of DLC1. Clinical epigenetics 2022, 14, 51. [Google Scholar] [CrossRef]
- Feng, Z.; Liu, Z.; Peng, K.; Wu, W. A Prognostic Model Based on Nine DNA Methylation-Driven Genes Predicts Overall Survival for Colorectal Cancer. Frontiers in Genetics 2022, 12. [Google Scholar] [CrossRef]
- Rasool, M.; Karim, S.; Haque, A.; Alharthi, M.; Chaudhary, A.G.; Natesan Pushparaj, P. Deciphering gene expression signatures in liver metastasized colorectal cancer in stage IV colorectal cancer patients. Journal of King Saud University - Science 2024, 36, 103415. [Google Scholar] [CrossRef]
- Xu, M.; Xu, X.; Pan, B.; Chen, X.; Lin, K.; Zeng, K.; Liu, X.; Xu, T.; Sun, L.; Qin, J. LncRNA SATB2-AS1 inhibits tumor metastasis and affects the tumor immune cell microenvironment in colorectal cancer by regulating SATB2. Molecular cancer 2019, 18, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-Q.; Jiang, D.-M.; Hu, S.-S.; Zhao, L.; Wang, L.; Yang, M.-H.; Ai, M.-L.; Jiang, H.-J.; Han, Y.; Ding, Y.-Q. SATB2-AS1 suppresses colorectal carcinoma aggressiveness by inhibiting SATB2-dependent Snail transcription and epithelial–mesenchymal transition. Cancer research 2019, 79, 3542–3556. [Google Scholar] [CrossRef]
- Huang, J.; Yang, Y.; Zhao, F.; Zhang, Z.; Deng, J.; Lu, W.; Jiang, X. LncRNA SATB2-AS1 overexpression represses the development of hepatocellular carcinoma through regulating the miR-3678-3p/GRIM-19 axis. Cancer Cell International 2023, 23, 82. [Google Scholar] [CrossRef]
- Xu, Y.; Yu, X.; Zhang, M.; Zheng, Q.; Sun, Z.; He, Y.; Guo, W. Promising advances in LINC01116 related to cancer. Frontiers in Cell and Developmental Biology 2021, 9, 736927. [Google Scholar] [CrossRef]
- Ghasemian, A.; Omear, H.A.; Mansoori, Y.; Mansouri, P.; Deng, X.; Darbeheshti, F.; Zarenezhad, E.; Kohansal, M.; Pezeshki, B.; Wang, Z.; et al. Long non-coding RNAs and JAK/STAT signaling pathway regulation in colorectal cancer development. Frontiers in Genetics 2023, 14. [Google Scholar] [CrossRef]
- Wang, X.-K.; Zhang, X.-D.; Luo, K.; Yu, L.; Huang, S.; Liu, Z.-Y.; Li, R.-F. Comprehensive analysis of candidate signatures of long non-coding RNA LINC01116 and related protein-coding genes in patients with hepatocellular carcinoma. BMC Gastroenterology 2023, 23, 216. [Google Scholar] [CrossRef]
Figure 1.
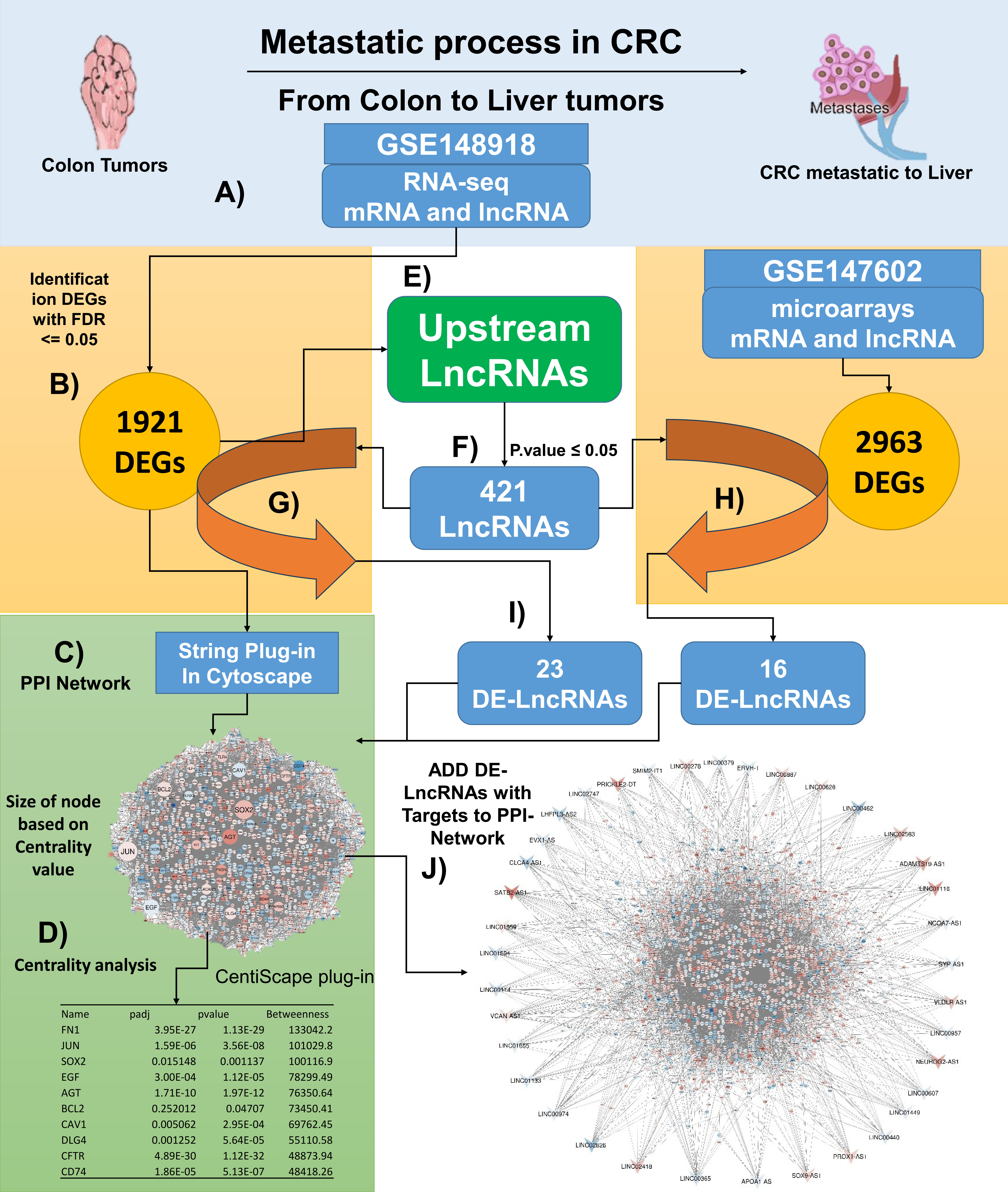
Overview of Analytical Workflow Leading to Current Study Results. This figure summarizes the steps taken in the study to analyze and interpret the data. A) The comparison between metastatic liver tumors and primary CRC tumors was performed using bulk RNA-seq techniques. B) Re-analysis of the data identified 1921 significantly differentially expressed genes (DEGs). C) These DEGs were input into Cytoscape software, using the STRING database to construct the protein-protein interaction (PPI) network. D) Centrality analysis was performed using the CentiScape plugin to determine the most influential genes within the PPI network, identifying key hub genes. E) Upstream analysis was conducted to predict which regulatory lncRNAs might regulate the identified DEGs. F) This upstream analysis highlighted 421 lncRNAs as potential regulators of the DEGs. G) The expression of these 421 lncRNAs was cross-referenced with the DEGs to filter relevant lncRNAs. H) To further refine this, an additional microarray dataset was re-analyzed and compared with the 421 candidate lncRNAs. I) From this analysis, 23 lncRNAs were identified as DE-lncRNAs in the first dataset, and 16 DE-lncRNAs were found in the second dataset. J) Integration of both DE-lncRNAs and their target DEGs was performed to map them to the PPI network.
Figure 1.
Overview of Analytical Workflow Leading to Current Study Results. This figure summarizes the steps taken in the study to analyze and interpret the data. A) The comparison between metastatic liver tumors and primary CRC tumors was performed using bulk RNA-seq techniques. B) Re-analysis of the data identified 1921 significantly differentially expressed genes (DEGs). C) These DEGs were input into Cytoscape software, using the STRING database to construct the protein-protein interaction (PPI) network. D) Centrality analysis was performed using the CentiScape plugin to determine the most influential genes within the PPI network, identifying key hub genes. E) Upstream analysis was conducted to predict which regulatory lncRNAs might regulate the identified DEGs. F) This upstream analysis highlighted 421 lncRNAs as potential regulators of the DEGs. G) The expression of these 421 lncRNAs was cross-referenced with the DEGs to filter relevant lncRNAs. H) To further refine this, an additional microarray dataset was re-analyzed and compared with the 421 candidate lncRNAs. I) From this analysis, 23 lncRNAs were identified as DE-lncRNAs in the first dataset, and 16 DE-lncRNAs were found in the second dataset. J) Integration of both DE-lncRNAs and their target DEGs was performed to map them to the PPI network.
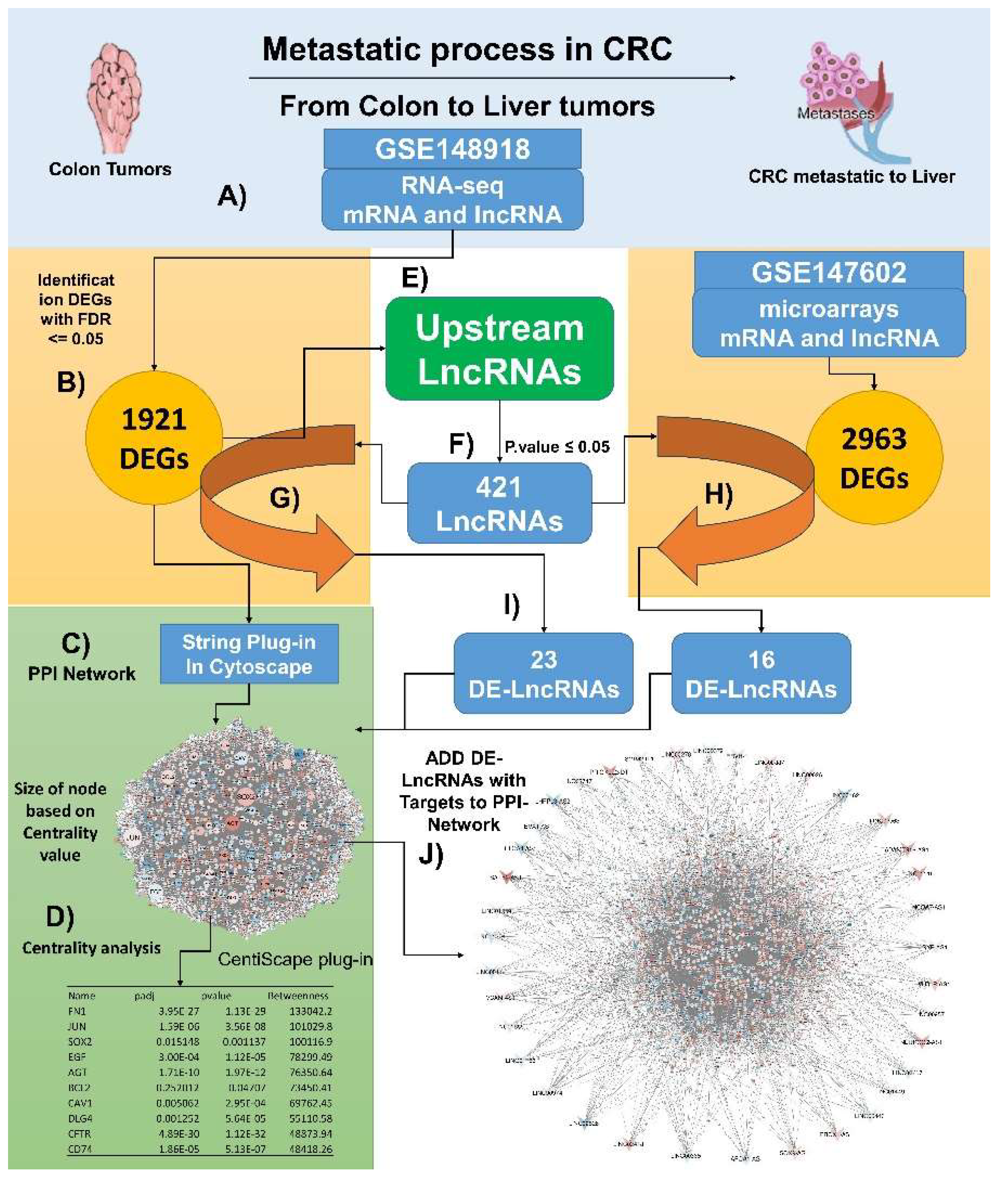
Figure 2.
Dispersion of Expressed Genes from Primary CRC Tumors to Liver Metastatic Tumors. This figure illustrates the distribution of gene expression changes between primary colorectal cancer (CRC) tumors and liver metastatic tumors. Genes shown in red are overexpressed, and those in blue are downregulated. Black nodes represent non-significant genes. Significant genes are defined by a p-value ≤ 0.05, with upregulated genes having a log2 fold change (Log2FC) ≥ +1 and downregulated genes having a Log2FC ≤ -1.
Figure 2.
Dispersion of Expressed Genes from Primary CRC Tumors to Liver Metastatic Tumors. This figure illustrates the distribution of gene expression changes between primary colorectal cancer (CRC) tumors and liver metastatic tumors. Genes shown in red are overexpressed, and those in blue are downregulated. Black nodes represent non-significant genes. Significant genes are defined by a p-value ≤ 0.05, with upregulated genes having a log2 fold change (Log2FC) ≥ +1 and downregulated genes having a Log2FC ≤ -1.
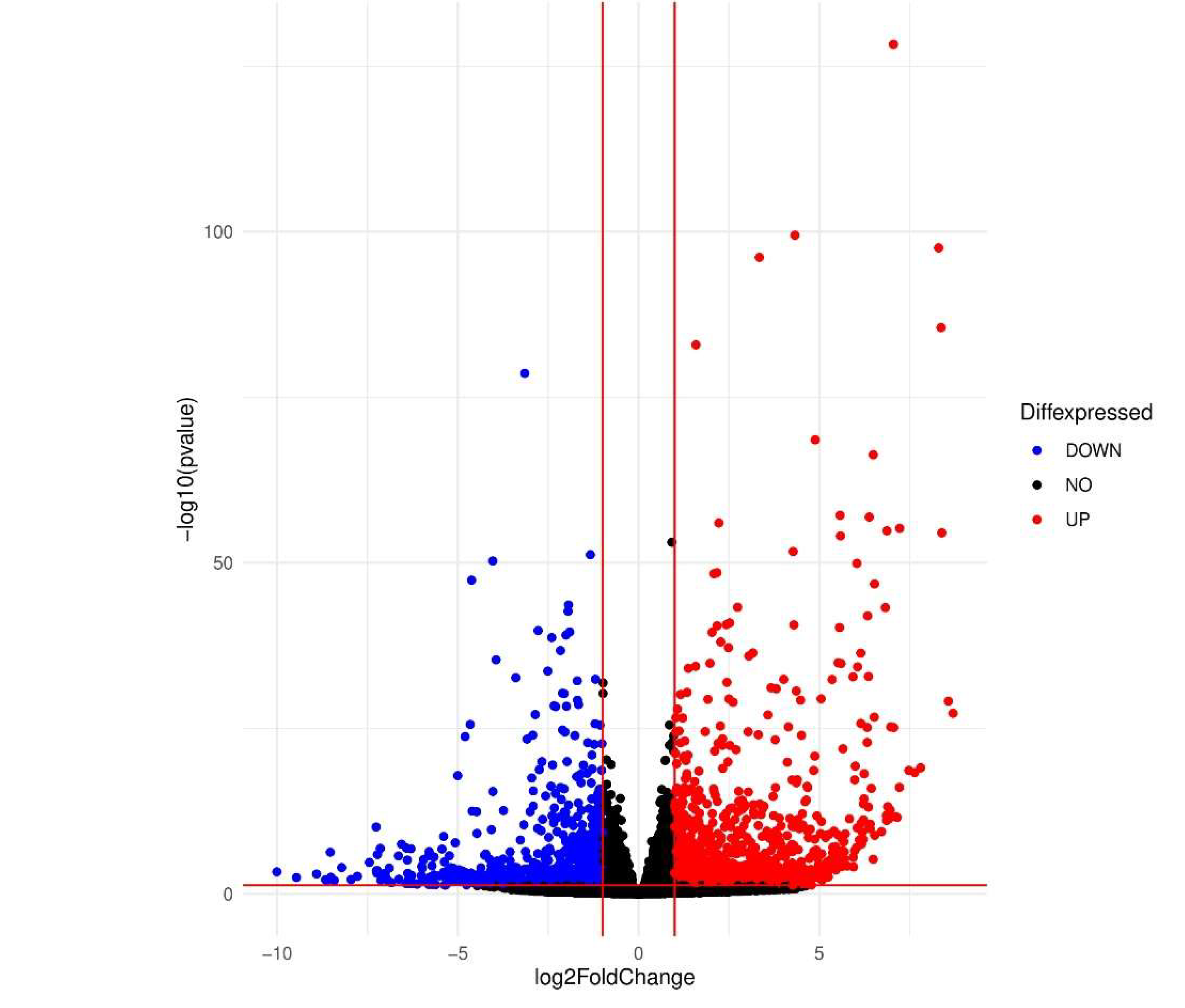
Figure 3.
Illustration of the Protein-Protein Interaction (PPI) Network. This figure depicts the PPI network consisting of 1,448 nodes and 8,748 edges. Nodes in red represent upregulated genes, while nodes in blue represent downregulated genes. The size of the nodes and their labels are proportional to their centrality scores, reflecting their significance within the network.
Figure 3.
Illustration of the Protein-Protein Interaction (PPI) Network. This figure depicts the PPI network consisting of 1,448 nodes and 8,748 edges. Nodes in red represent upregulated genes, while nodes in blue represent downregulated genes. The size of the nodes and their labels are proportional to their centrality scores, reflecting their significance within the network.
Figure 4.
The integration of 37 differentially expressed lncRNAs (DE-lncRNAs) into the PPI network, representing 1,523 nodes and 9,450 edges. Red nodes indicate overexpressed genes, while blue nodes indicate down-expressed genes. DE-lncRNAs are represented by V-shaped nodes.
Figure 4.
The integration of 37 differentially expressed lncRNAs (DE-lncRNAs) into the PPI network, representing 1,523 nodes and 9,450 edges. Red nodes indicate overexpressed genes, while blue nodes indicate down-expressed genes. DE-lncRNAs are represented by V-shaped nodes.
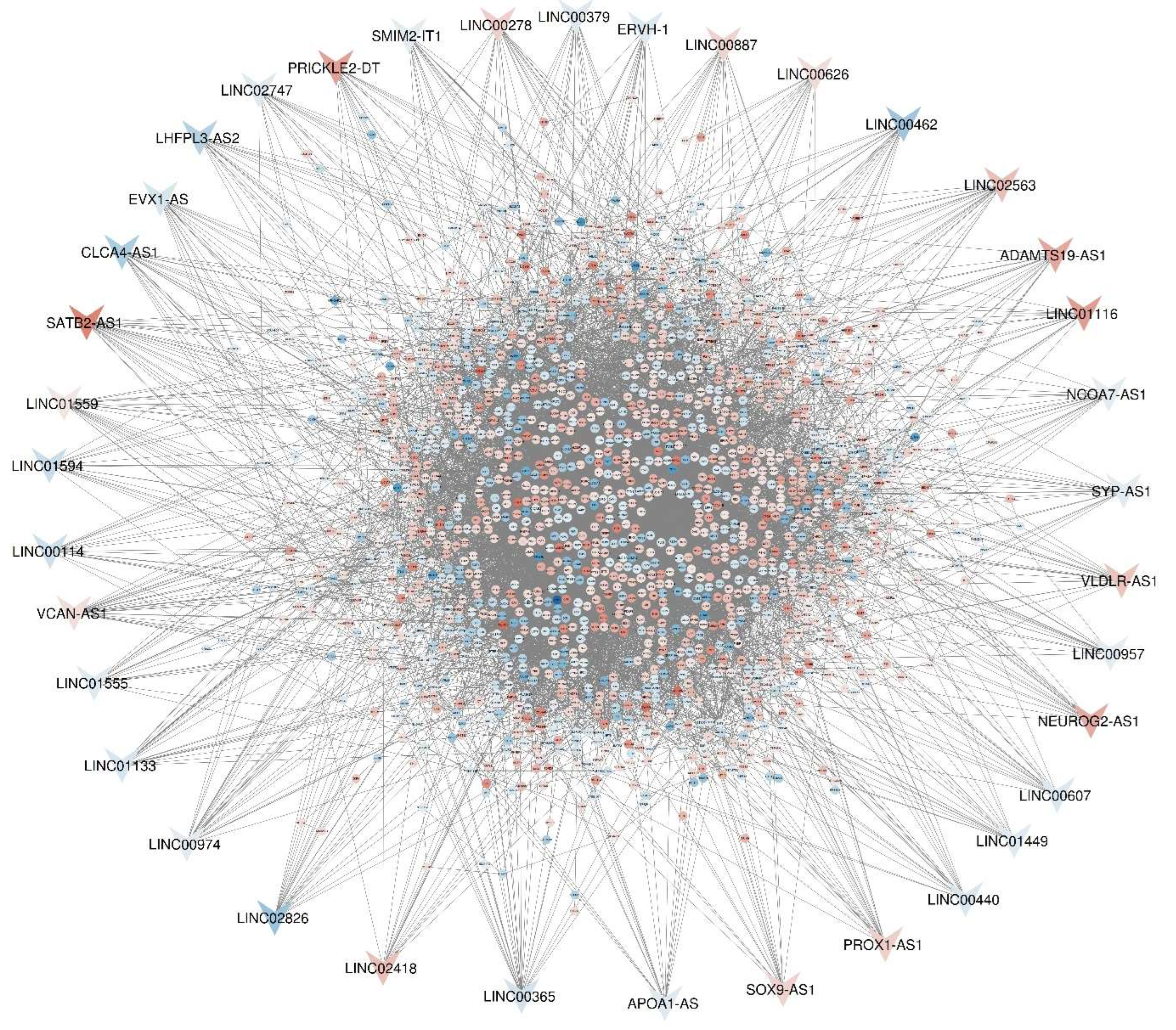

Table 1.
Top 10 Hub Genes in the PPI Network. This table presents the top 10 hub genes identified in the PPI network. The first column lists the hub gene names, followed by the adjusted p-values in the second column, p-values in the third column, and betweenness scores in the fourth column. FN1, with the highest betweenness score, emerges as the most significant hub gene, indicating its central role in interactions within the PPI network associated with colorectal cancer metastasis to liver tissue.
Table 1.
Top 10 Hub Genes in the PPI Network. This table presents the top 10 hub genes identified in the PPI network. The first column lists the hub gene names, followed by the adjusted p-values in the second column, p-values in the third column, and betweenness scores in the fourth column. FN1, with the highest betweenness score, emerges as the most significant hub gene, indicating its central role in interactions within the PPI network associated with colorectal cancer metastasis to liver tissue.
| Name | padj | Pvalue | Betweenness |
| FN1 | 3.95E-27 | 1.13E-29 | 133042.2 |
| JUN | 1.59E-06 | 3.56E-08 | 101029.8 |
| SOX2 | 0.015148 | 0.001137 | 100116.9 |
| EGF | 3.00E-04 | 1.12E-05 | 78299.49 |
| AGT | 1.71E-10 | 1.97E-12 | 76350.64 |
| BCL2 | 0.252012 | 0.04707 | 73450.41 |
| CAV1 | 0.005062 | 2.95E-04 | 69762.45 |
| DLG4 | 0.001252 | 5.64E-05 | 55110.58 |
| CFTR | 4.89E-30 | 1.12E-32 | 48873.94 |
| CD74 | 1.86E-05 | 5.13E-07 | 48418.26 |
Table 2.
The table represent indicated DE-lncRNAs from first and second datasets. The first columns indicate symbol names, the second columns express logFC criteria which indicate amount of differential expressed genes, the third and fourth columns representing P.value and adjusted.P.value respectively and the final column report DE-lncRNA targets from our DEGs.
Table 2.
The table represent indicated DE-lncRNAs from first and second datasets. The first columns indicate symbol names, the second columns express logFC criteria which indicate amount of differential expressed genes, the third and fourth columns representing P.value and adjusted.P.value respectively and the final column report DE-lncRNA targets from our DEGs.
| First Dataset | Second Dataset | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Symbol | LogFC | P.Value | Padj | Targets | Symbol | LogFC | P.Value | Padj | Targets |
| LINC00365 | -1.96228 | 0.01873 | 0.13292 | 26 | VCAN-AS1 | 1.6172 | 0.00036 | 0.00448 | 29 |
| SATB2-AS1 | 1.3E-52 | 7.22302 | 6.2E-56 | 25 | SATB2-AS1 | -1.4441 | 0.04048 | 0.12922 | 25 |
| LINC00278 | 2.477 | 0.0095 | 0.07997 | 22 | LINC00974 | -1.1489 | 0.00107 | 0.00973 | 24 |
| LINC02826 | -4.91475 | 0.00683 | 0.06268 | 21 | SYP-AS1 | -1.1536 | 7.57E-05 | 0.00148 | 19 |
| LINC01559 | 1.14904 | 5.6E-11 | 3.9E-09 | 21 | LINC00440 | -1.4691 | 0.0008 | 0.00789 | 19 |
| CLCA4-AS1 | -3.87085 | 0.02089 | 0.14368 | 21 | SMIM2-IT1 | -1.03 | 0.00295 | 0.02051 | 18 |
| LINC01133 | -1.71347 | 1.9E-18 | 3.2E-16 | 21 | LINC01555 | -1.3152 | 0.007 | 0.03816 | 18 |
| LINC02563 | 3.48567 | 0.00133 | 0.01714 | 20 | LINC01449 | -1.2126 | 2.05E-07 | 6.73E-05 | 18 |
| LINC00462 | -4.69382 | 0.01943 | 0.13672 | 20 | LINC00114 | -2.1923 | 0.00927 | 0.04644 | 18 |
| VLDLR-AS1 | 3.19298 | 0.01845 | 0.13161 | 19 | NCOA7-AS1 | -1.0249 | 0.0004 | 0.00479 | 17 |
| PRICKLE2-DT | 5.4572 | 0.00024 | 0.00433 | 19 | ERVH-1 | -1.3516 | 0.00025 | 0.00347 | 17 |
| PROX1-AS1 | 2.56183 | 3.6E-09 | 1.9E-07 | 19 | APOA1-AS | -1.248 | 7.72E-07 | 0.00011 | 17 |
| ADAMTS19-AS1 | 4.95854 | 4.1E-06 | 0.00012 | 18 | LINC01116 | 1.9556 | 0.00066 | 0.00683 | 16 |
| LINC00887 | 2.25373 | 0.0002 | 0.00369 | 18 | LINC00626 | 1.7331 | 0.0161 | 0.06867 | 16 |
| EVX1-AS | -1.64684 | 0.00097 | 0.01333 | 18 | LINC00607 | -1.411 | 0.00133 | 0.01147 | 16 |
| NEUROG2-AS1 | 4.80579 | 1.6E-06 | 5.2E-05 | 17 | LINC00379 | -1.3688 | 0.01837 | 0.07516 | 16 |
| LHFPL3-AS2 | -3.44886 | 0.01825 | 0.13062 | 17 | |||||
| LINC02418 | 3.48651 | 0.00144 | 0.01826 | 17 | |||||
| LINC00957 | -1.05286 | 0.01165 | 0.09339 | 17 | |||||
| SOX9-AS1 | 2.3242 | 3.7E-24 | 8.9E-22 | 16 | |||||
| LINC02747 | -1.30917 | 0.00271 | 0.03019 | 16 | |||||
| LINC01594 | -2.46856 | 0.01107 | 0.09008 | 16 | |||||
| LINC01116 | 6.3E-07 | 5.67987 | 1.3E-08 | 16 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



