
Submitted:
03 December 2024
Posted:
06 December 2024
You are already at the latest version
Abstract
Aquaporins (AQPs) are membrane proteins facilitating water and other small solutes to be transported across cell membranes. They are crucial in maintaining cellular homeostasis by regulating water permeability in various tissues. Moreover, they regulate cell migration, signaling pathways, inflammation, tumor growth, and metastasis. In critically ill patients, such as trauma, sepsis, and patients with acute respiratory distress syndrome (ARDS), which are frequently encountered in intensive care units (ICUs), water transport regulation is crucial for maintaining homeostasis, as dysregulation can lead to edema or dehydration, with the latter also implicating hemodynamic compromise. Indeed, AQPs are involved in fluid transport in various organs, including the lungs, kidneys, and brain, where their dysfunction can exacerbate conditions like ARDS, acute kidney injury (AKI), or cerebral edema. In this review, we discuss the implication of AQPs in the clinical entities frequently encountered in ICUs, such as systemic inflammation and sepsis, ARDS, acute kidney injury (AKI), and brain edema due to different types of primary brain injury from a clinical perspective. Current and possible future therapeutic implications are also considered.
Keywords:
AQP
; critically ill
; sepsis
; ARDS
; AKI
; ABI
1. Introduction
1.1. Aquaporin Localization and Role in Physiology and Disease
Aquaporins (AQPs) are specialized water channel proteins crucial for maintaining cellular homeostasis by regulating water and other small solutes’ transport across cell membranes. Their discovery revolutionized our understanding of how water moves through biological systems. In 1992, Peter Agre and his team identified the first aquaporin, which was later named aquaporin-1 (AQP1), while studying Rh blood group antigens. They noticed an unexplained protein in red blood cell membranes and kidney tissues that facilitated rapid water transport. By isolating and cloning this protein, they confirmed its function as a water channel [1,2,3,4]. This groundbreaking work earned Agre the Nobel Prize in Chemistry in 2003. Aquaporins are now recognized to be essential for processes such as kidney filtration, glandular secretion, and maintaining water balance in cells.
The family of aquaporins consists of at least 13 different isoforms in humans (AQP0–AQP12), each with unique tissue distribution, distinct physiological roles, and regulatory mechanisms. Table 1 summarizes the findings thus far on the distinct tissue distributions and physiological roles of human AQPs [5,6,7,8].
The key physiological functions across AQPs include water homeostasis, exocrine secretion, energy metabolism, neuroprotection, osmoregulation, and detoxification. The importance of AQPs in physiology extends to various pathophysiological processes, such as cell migration, signaling pathways, inflammation, tumor growth, metastasis, and neurological disorders, highlighting their multifunctional nature in maintaining cellular integrity and responding to physiological challenges [5,7,9,10].
In critically ill patients, such as sepsis, patients with acute respiratory distress syndrome (ARDS), acute kidney injury (AKI), and acute brain injury (ABI), which are frequently encountered in intensive care units (ICUs) [11,12], water transport regulation is crucial for maintaining homeostasis, as dysregulation can lead to edema or dehydration, with the latter also implicating hemodynamic compromise [13,14,15,16]. Indeed, AQPs are involved in fluid transport in various organs, including the lungs, kidneys, and brain, where their dysfunction can exacerbate conditions like ARDS, AKI, or cerebral edema [17,18,19,20,21,22,23,24,25,26,27,28,29,30,31]. Additionally, altered AQP expression and function can affect patient outcomes by contributing to vascular permeability and interstitial edema [32,33,34]. Moreover, AQP implication in cell migration and the inflammatory response during critical illness can influence patient response in sepsis [35,36]. Finally, in trauma cases, AQPs can contribute to the body’s attempt to maintain fluid balance and restore homeostasis [31,37,38,39,40,41,42,43,44,45,46,47]. Understanding the roles of AQPs in these contexts can aid clinicians in managing fluid therapy more effectively and minimizing complications associated with organ dysfunction.
In the present review, we discuss the implication of AQPs in the clinical entities frequently encountered in ICUs, such as systemic inflammation and sepsis, ARDS, AKI, and brain edema due to different types of primary brain injury. The role of AQPs is viewed from a clinical perspective. Current and possible future therapeutic implications are included in the discussion.
1.2. Aquaporin Structure and Function
The structure of AQPs is characterized by six transmembrane alpha-helices (H1–H6) that form a central pore (Figure 1). These helices are connected by five loops extending into the extracellular and cytoplasmic spaces. This arrangement creates an hourglass-shaped channel, lined with hydrophilic amino acids, allowing the passage of water molecules through the AQP monomer while excluding ions and larger solutes [48,49].
The selectivity of AQPs for water is primarily due to the size and charge of the channel, which is finely tuned to permit the rapid transit of water molecules in response to osmotic gradients [48]. The water transport mechanism through AQPs is facilitated by a unique property known as “single-file diffusion.” Water molecules enter the AQP channel, and are aligned in a single file, enabling rapid movement through the narrow pore [50]. The channel’s structure also contains specific residues playing critical roles in maintaining selectivity and function. For instance, the presence of an asparagine-proline-alanine (NPA) motif, found in the second and fifth loops, is crucial for forming hydrogen bonds with water molecules, thus ensuring the proper conduction of water. Another structural element of the AQP channel important for water selectivity is the “aromatic/arginine (ar/R) selectivity filter”, in which four amino acids, histidine 180, cysteine 189, phenylalanine 56, and arginine 195 (the numbers of these amino acids may differ in different isoforms) come together in the pore and form the part of the pore with the smallest diameter, excluding sodium, potassium, and hydrogen [48]. (Figure 1).
Although each AQP monomer can independently transport water, the channel’s quaternary structure forms a tetramer [51,52,53]. The tetrameric arrangement also forms a central, primarily hydrophobic channel, whose specific function remains unclear (Figure 1). Some believe it doesn’t act as a pore, whereas others have data showing that it acts as an ion channel that is gated with the loops of the AQP protein being involved in determining when the channel is open or closed [48,54].
AQPs primarily facilitate water transport, but many of them also transport other small solutes like glycerol, nitrate, urea, ammonia, hydrogen peroxide (H₂O₂), carbon dioxide (CO₂), oxygen (O₂), arsenic, antimony, and more, with roles in both physiological functions and in mediating toxicity [6,55,56,57,58,59,60,61,62,63,64]. Human AQPs are, therefore, further classified into orthodox AQPs, aquaglyceroporins, and super aquaporins, with each group facilitating the transport of distinct types of molecules [5]. AQPs 1, 3, 6, 7, 8, 9, and 10 facilitate ammonia transport [65,66]. Aquaglyceroporins, including AQP 3, 6, 7, 9, and 10 also transport urea, potentially linking it to energy metabolism [66]. AQPs 3, 7, 9, and 10 can transport antimonite [58,67,68]. AQPs 0, 1, 4, 5, and 6 are implicated in CO₂ transport, with recent evidence showing that AQP1 also facilitates the transport of O₂ and nitric oxide (NO) [69,70].
1.3. Aquaporin Regulation
An important aspect of AQP function affecting their role in pathophysiological processes is their regulation. AQPs are regulated at transcriptional, post-transcriptional, translational, and post-translational levels. These regulatory mechanisms ensure that AQPs respond appropriately to maintain water balance in different tissues and under varying physiological conditions.
At the transcriptional level, hormones such as vasopressin (antidiuretic hormone, ADH) can upregulate AQP expression, especially AQP2 in the kidneys [72,73]. Changes in osmotic gradients are another major factor affecting AQP regulation. Hyperosmolar or hypoosmolar conditions can influence the expression of AQPs like AQP1 and AQP4 in the brain and kidneys [5,74]. Cytokines and inflammatory mediators can modulate AQP gene transcription in response to injury or infection [75,76]. Finally, hypoxic conditions can upregulate or downregulate AQPs, such as AQP1 and AQP4, depending on the tissue and context [77,78,79].
Post-translational modifications include alternative splicing and microRNAs (miRNAs). Different splicing patterns can affect the production of functional AQP isoforms [80], whereas miRNAs can bind to AQP mRNA and suppress their translation, impacting protein levels [81].
Phosphorylation, ubiquitination, and methylation at the post-translational level also play a crucial role in regulating AQPs. For example, phosphorylation at serine residues in the AQP2 protein is essential for its trafficking to the plasma membrane. The dephosphorylation of AQP2 by phosphatases can reverse the effects, promoting the removal of AQP2 from the membrane. Phosphorylation not only controls the localization and membrane abundance of AQPs but can also modulate their water permeability directly [82,83,84,85,86,87,88]. Ubiquitination marks AQPs for degradation, affecting their turnover and cellular abundance, while methylation and/or acetylation can alter protein function or interactions, although this is less studied for AQPs [89].
There are other regulatory mechanisms also. AQPs can be stored in intracellular vesicles and trafficked to the plasma membrane in response to ADH stimulation [90] or can be inserted and removed from the plasma membrane through endocytosis and exocytosis, respectively [91]. AQPs often interact with other proteins, such as anchoring proteins or scaffolding molecules, to stabilize their localization and function [71,92]. Changes in osmotic gradients or mechanical forces can directly influence the gating of AQPs, altering their water permeability [5,74]. Finally, pathological conditions can dysregulate AQP expression, as described in detail below.
These regulatory mechanisms allow AQPs to respond dynamically to changes in the cellular and extracellular environment, maintaining homeostasis and adapting to physiological and pathological conditions.
2. Aquaporins in Critical Illness
2.1. The Role of Aquaporins in ICU Patients with Sepsis
2.1.1. Sepsis
The Third International Consensus (Sepsis-3) currently defines sepsis as “organ dysfunction caused by a dysregulated host response to infection.” In the most severe form of sepsis, septic shock, there is profound circulatory and cellular/metabolic dysfunction. Persistent hypotension despite fluid resuscitation, along with elevated lactate levels, are hallmarks. Multiorgan failure can occur as a result of prolonged hypoperfusion and metabolic disturbances, with death being the likely outcome if untreated. Its pathophysiology includes an exaggerated inflammatory response to an infection leading to endothelium dysfunction, and increased vascular permeability, triggering coagulation abnormalities including microthrombi formation and decreased fibrinolysis [93]. Despite great progress in research and clinical management made in the last three decades, sepsis mortality is still high and, therefore, novel regulatory mechanisms and therapeutic strategies are still needed.
2.1.2. Aquaporin Expression in Human Immune Cells
Aquaporins (AQPs) are expressed in various human immune cells, including lymphocytes, macrophages, neutrophils, and dendritic cells, where they play crucial roles in immune functions such as migration, proliferation, and response to inflammation (Figure 2). AQP1 is expressed in neutrophils, monocytes and lymphocytes, contributing to water transport and cell volume regulation during migration and differentiation [75,94,95,96]. AQP3 is expressed in leukocytes, including lymphocytes, monocytes, macrophages, and dendritic cells, facilitating chemokine-driven migration and supporting energy metabolism through glycerol transport [75,94,95,96,97,98,99]. AQP5 is expressed in some immune cells, particularly lymphocytes, neutrophils, and dendritic cells [94,100]. AQP5 plays a role in facilitating water transport, which is critical for maintaining cell shape and motility during migration. In immune context, AQP5 has been implicated in the regulation of lymphocyte activation and migration, although its precise mechanisms in immune responses remain less well-characterized compared to AQP3 or AQP9 [94,100]. AQP9 is found most prominently in leukocytes, aids in respiratory bursts and cell motility during pathogen defense [101]. Immune cells in which AQP9 seems to have a role are neutrophils, macrophages, monocytes, lymphocytes, and dendritic cells [33,75,96,97,102,103,104]. Other AQPs detected at low concentrations in immune cells are AQP6, AQP7, AQP8, AQP11, and AQP12, expressed in monocytes, macrophages, and dendritic cells [75,96,97].
2.1.3. Aquaporins and Sepsis
Aquaporins are emerging as a novel area of research in sepsis, given their roles in regulating fluid balance/edema, inflammation/immune response, and vascular permeability. Their effects seem to be cell and/or tissue and aquaporin-specific. A significant number of experimental studies are available unraveling the underlying pathophysiological mechanisms attributed to aquaporins during sepsis. Clinical studies have shown that the expression of AQPs is often altered during sepsis. In particular, studies have highlighted changes in AQP levels in the lungs, kidneys, and other organs during sepsis, correlating with disease severity and outcomes. This makes AQPs potential biomarkers for sepsis severity or targets for intervention aimed at improving organ function during septic states. However, clinical applications are still in early stages, and further investigations are required to fully understand their impact on sepsis outcomes.
Detailed analysis of clinical studies related to aquaporins and critical sepsis are presented below. Investigations have focused on the expression of AQP1, AQP3, AQP5, and AQP9 in blood cells. A possible explanation is that these aquaporins are related to immune cell regulation. Most studies have shown that the expression is increased, and, of importance, a number of them raise the possibility that AQP expression is linked to high mortality.
AQP9 is expressed in immune system cells and most prominently in leukocytes. Ishibashi et al. identified the AQP9 cDNA in human leukocytes by polymerase chain reaction. They showed that AQP9 is expressed at the highest levels in polymorphonuclear cells (PMNs), followed by liver, lung, and spleen, suggesting that AQP9 may play a role in the immunological function of leukocytes [102]. The ability of neutrophils to sense and move to sites of infection is essential for defense against pathogens. Matsushima et al. showed that the expression of AQP9 in PMNs is increased in patients with systemic inflammatory response syndrome (SIRS) compared to healthy volunteers. Moreover, they showed that PMNs exhibited morphological and functional changes [103]. Of note, AQP9 is permeable to water and other small molecules such as glycerol, urea, and certain metabolic by-products like lactate. Loitto et al. reported that AQP9 plays an essential role in water influx during the activation of PMNs. They proved that the water influx through AQP9 contributes to cell motility and shape changes in PMNs, which are essential for transmigration or chemotaxis of PMNs to the inflammatory site [106]. Taken together, these suggest that AQP9 may play a significant role in morphological and functional changes in PMNs during SIRS. In contrast, AQP9 expression was not altered during sepsis, and low levels were associated with increased survival [107].
Additionally, Thon et al. also measured AQP3 and showed that AQP3 mRNA expression in whole blood samples from septic patients increased over the course of sepsis, correlated with the lymphocyte count, and most importantly high AQP3 expression was associated with increased survival. Thus, the roles of AQP3 and AQP9 differ in the pathophysiology of sepsis and may be useful biomarkers for sepsis. Furthermore, modulation of their expression might offer a therapeutic option. This study also found that AQP3 correlated with IL-8 levels, suggesting that AQP3 might affect cytokine signaling pathways [107]. AQP3 is expressed in human leukocytes, monocytes, lymphocytes, and dendritic cells [94,95,96,97]. Moreover, AQP3 appears to be important for T-cell function [98,99]. AQP3 seems to regulate the inflammasome, which in turn stimulates cytokine production [96]. Taken together, these suggest that AQP3 has a key role in immune responses. Finally, in a gene expression analysis of 5 critically ill patients who developed sepsis and septic shock, AQP3 was found downregulated [95]. The AQP3 polymorphism (rs17553719) exhibited an association with 30-day survival in patients with sepsis. Patients with the homozygote CC genotype had lower 30-day survival rate compared to the CT and the TT genotypes, while patients with the CC genotype had higher AQP3 mRNA at study admission compared to the other genotypes [108].
The leukocyte AQP1 expression levels were measured in critically ill patients on ICU admission, upon diagnosis of sepsis, and at the occurrence of septic shock. AQP1 was induced in the leukocytes of patients with ICU-acquired sepsis and exhibited higher expression in septic shock. Furthermore, neutrophil AQP1 expression was induced by LPS in vitro [95]. AQP1 was the first mammalian AQP reported to be observed in erythrocytes and renal tubules and was originally named channel-forming integral membrane protein of 28 kDa (CHIP28) [1]. Subsequent studies have demonstrated that it is physiologically distributed in the choroidal plexus, corneal endothelium, pain-processing C-fibers of the spinal cord, and all vascular endothelial cells except in the central nervous system (CNS) [8]. Moreover, AQP1 plays a role in cell migration and angiogenesis in cancer [109]. In the context of sepsis, AQP1 overexpression in leukocytes may increase their cell membrane permeability, increasing their volume and, hence, migration. In addition, in experimental studies, AQP1 emerges as a critical scaffold for a plasma membrane-associated multiprotein complex important for cytoskeleton build-up, adhesion, and motility [110].
A growing number of studies have aimed to investigate the interplay between long non-coding RNAs (lnRNAs) and micro RNAs (miRNAs) in the regulation of aquaporins, and especially their potential role as therapeutic targets in sepsis [34,81]. Non-coding RNAs are non-protein coding RNAs that regulate gene expression on an epigenetic, transcriptional, and post-transcriptional level. The most studied classes of non-coding RNAs are lncRNAs and miRNAs. lncRNAs lack any protein-coding capacity and consist of more than 200 nucleotides, while miRNAs are short non-coding RNAs of 18–24 nucleotides [111,112]. The recent identification of lnRNAs and miRNAs as regulators of aquaporin expression in several conditions has instigated research into the potential role of these molecules in the setting of sepsis.
In one study, septic patients had lower serum lncRNA H19 and AQP1 mRNA levels and higher miR-874 levels compared to healthy controls, showing a negative relationship in septic patients. Based on their results, lncRNA H19 functioned as an AQP1 competitive endogenous RNA (ceRNA) in regulating miR-874, which directly interacted with AQP1 [113]. Similarly, a subsequent study showed that the serum levels of lnRNA H19 were decreased in septic patients compared to healthy controls [114]. The results of these studies propose that lnRNA H19 could be used as a biomarker of early sepsis diagnosis.
The lncRNA cancer susceptibility candidate 2 (CASC2) has been shown to regulate AQP1 expression via the miR-144-3p/AQP1 axis [115]. In a study including 184 septic patients, circulating lncRNA CASC2 levels were found to be decreased compared to healthy controls, while lncRNA CASC2 was associated with disease severity and multi-organ injuries and could be used as a prognostic biomarker [116].
In 2008, upon sequencing of the AQP5 promoter, a novel functional 1364A/C polymorphism was identified and was associated with decreased AQP5 expression. The presence of the C allele was associated with lower mRNA and protein AQP5 expression [117]. Subsequently, the same group studied 154 patients with sepsis and demonstrated that the 1364A/C polymorphism was associated with higher 30-day survival. Thus, they concluded that decreased AQP5 gene expression seemed to enhance survival by reducing the inflammatory burden during sepsis [118].
Epigenetic regulation via DNA methylation also contributes to a differential AQP5 expression in sepsis. More specifically, DNA methylation of the AQP5 promoter results in decreased reporter gene transcription, while demethylation induces AQP5 expression [119]. Rump et al. performed DNA methylation analysis within the AQP5 promoter in 135 septic patients on blood samples collected within the first 24-hrs after sepsis diagnosis. The methylation analysis revealed that CpG methylation at the AQP5 promoter position “nt-937” was independently associated with an increased risk of death within 30 days. Moreover, non-survivor septic patients had higher AQP5 mRNA expression levels in the blood cells compared to survivors, and, of importance, increased expression was linked to increased risk of death. They concluded that DNA methylation within the AQP5 promoter could act as a possible mechanism for differential AQP5 expression [89].
In a later study, the same group associated the presence of the C-allele of the AQP5-1364A/C polymorphism with a lower gene expression in septic patients, accompanied by a higher methylation level of the AQP5 promoter. Moreover, there was a differential AQP5 promoter methylation in different types of immune cells of septic patients, suggesting that AQP5 promoter methylation could drive genotype-dependent expression [120].
Zheng et al. aimed to investigate the relationship between miR-34b-5p, which has been associated with sepsis-induced organ injury, and AQP2. In the sera of septic patients with acute kidney injury miR-34b-5p expression levels were found elevated compared to healthy controls. This study reported miR-34b-5p as a novel biomarker in septic AKI patients, which could regulate sepsis-induced renal injury by inhibiting AQP2 [121].
2.1.4. Proposed Mechanisms of Aquaporin Involvement in Sepsis
Experimental studies have aided in unraveling the underlying pathophysiological mechanisms attributed to aquaporins during sepsis.
i) Increased Vascular Permeability and Edema
During sepsis, inflammation can disrupt the endothelial barrier, causing fluids to leak from blood vessels into tissues, resulting in edema. AQPs, particularly AQP1 (found in vascular endothelium) and AQP4 (found in the brain and lung), may contribute to this permeability by facilitating water movement, which exacerbates tissue edema in organs like the lungs (ARDS) and brain (cerebral edema) [122,123,124,125,126].
Moreover, studies have shown that AQPs can be upregulated in response to inflammatory mediators such as cytokines, which are elevated during sepsis [75,76,127]. This increase can lead to more water transport across membranes, contributing to the excessive tissue swelling seen in septic patients.
ii) Contribution to Organ Dysfunction
Aquaporins in the kidneys (notably AQP2 in the collecting duct) are critical for regulating water reabsorption [128]. Sepsis-induced dysregulation of AQP2 can impair water reabsorption, potentially leading to fluid imbalances and worsening renal function [121,129,130,131,132,133].
Aquaporins are also highly expressed in the brain and lung and have been implicated in cerebral and pulmonary edema during sepsis [34,134]. This renders them potential therapeutic targets to reduce brain swelling and improve neurological outcomes in severe sepsis cases. The involvement of aquaporins in brain and lung injury in critically ill patients is discussed further below.
iii) Inflammatory Modulation
Some aquaporins, like AQP1, may play roles in modulating cytokine production and other inflammatory processes. AQP-mediated changes in cell volume and migration can impact immune cell functions, influencing the systemic inflammatory response characteristic of sepsis [95,109,110].
iv) Barrier Integrity
In sepsis, aquaporins can influence the tight junctions of cells, which help maintain barrier integrity [135]. Dysregulation in aquaporin expression can thus contribute to the breakdown of barriers in various organs, including the intestines, lungs, and kidneys, leading to further complications in septic patients.
2.1.5. Aquaporins as Therapeutic Targets in Sepsis
Targeting specific AQPs has been proposed as a potential therapeutic strategy in sepsis management to reduce edema and improve organ function. For instance, AQP4 inhibitors have been proposed as potential therapeutic targets to alleviate cerebral edema [136,137]. However, the clinical application of AQP modulation in sepsis requires further research.
2.1.6. Conclusion
Aquaporins play an important role in the pathophysiology of sepsis by regulating water movement and potentially influencing inflammation. They are implicated in the fluid imbalances and edema formation that are prominent in sepsis-related organ dysfunction, particularly in the brain, lungs, and kidneys. Understanding and potentially modulating AQP activity could offer new avenues for therapeutic intervention in sepsis management, although more research is needed to clarify their exact roles and therapeutic viability.
2.2. The Role of Aquaporins in ICU Patients with Clinical Acute Lung Inflammation - Acute Respiratory Distress Syndrome (ARDS)
2.2.1. Clinical Acute Lung Inflammation - Acute Respiratory Distress Syndrome (ARDS)
Based on the revised Berlin Definition, acute respiratory distress syndrome (ARDS) is a heterogeneous syndrome characterized by acute onset, distinctive radiographic findings, and increased mortality. It constitutes a severe lung condition characterized by widespread inflammation, increased vascular permeability, and accumulation of fluid in the alveoli, which leads to impaired gas exchange and respiratory failure [138]. Several direct and indirect lung injury etiologies increase the risk of ARDS development. Sepsis, ischemic periods followed by reperfusion, aspiration, toxic inhalation, and radiation are the principal inciting events leading to ARDS [139].
The lung facilitates oxygen and carbon dioxide exchange between air and blood. A liquid layer directly exposed to the gaseous compartment covers the airways and the respiratory zone forming the air-liquid interface. The pulmonary epithelium separates this lining fluid from the extracellular compartment. Maintaining the fluid volume is essential for gas exchange, achieved by balancing fluid transport across the epithelium. While osmosis mainly drives airway fluid transport, filtration may contribute to alveolar fluid accumulation under pathological conditions.
In critically ill patients, particularly those with ARDS, the balance between fluid entry and clearance is disrupted due to increased vascular permeability, impaired alveolar clearance, oxidative stress, and the inflammatory environment of ARDS [140,141,142] (Figure 3). More specifically, inflammatory mediators such as cytokines (TNF-α, IL-1β) increase capillary permeability, leading to fluid leakage into the alveoli. When the process of removing fluid from the alveolar space is disrupted (e.g., through downregulation in response to inflammatory signals), pulmonary edema worsens.
AQPs are integral membrane proteins that act as water channels, facilitating water transport across cell membranes in response to osmotic gradients. This function is crucial for maintaining fluid balance between the alveolar and interstitial spaces and the capillaries. Disruption in alveolar fluid clearance, due to altered functional expression of respiratory AQPs, underscores their pathophysiological importance in respiratory disease associated with pulmonary edema [124,143]. In ARDS and lung injury in critically ill patients, AQPs are crucial for regulating lung water balance, directly impacting the development and resolution of pulmonary edema. Excessive pulmonary edema formation impairs gas exchange, worsens respiratory failure, and has been associated with adverse outcomes in ARDS patients [144].
2.2.2. Overview of Aquaporins in the Lung
AQP1, AQP3, AQP4, and AQP5 are the main aquaporins expressed in the lung. AQP1 is present in the capillary endothelial cells and plays a role in the control of water movement across the endothelial barrier. AQP3 and AQP4 are found in the airway epithelium and alveolar epithelial cells. AQP5 is expressed in the alveolar epithelial cells type I and is primarily responsible for water transport across the alveolar barrier (Figure 3). The specific localization of AQPs to both endothelial and epithelial cells in alveolar tissue suggests a role for these proteins in fluid movement between the airspace, interstitial, and capillary compartments. Indeed, data from AQP knock-out mice provide evidence for the specific involvement of AQP1 in capillary–airspace fluid movement [122,145]. The first data on the involvement of the AQPs in lung water transport came from experimental models using AQP1 and AQP4 knock-out mice [122,145]. AQP1 null mice had reduced water permeability between airspace and capillary compartments. The results obtained from these models showed that osmotically driven water transport across lung microvessels occurs transcellularly via AQP1 water channels and that the microvascular endothelium is a significant barrier for airspace-capillary osmotic water transport. The authors concluded that AQP1 facilitates hydrostatically driven lung edema but is not required for active near-isosmolar absorption of alveolar fluid. A year later, using AQP5 knock-out mice, Ma and coworkers demonstrated that AQP5 is responsible for most of the water transport occurring across the apical membrane of type I alveolar epithelial cells. Alveolar fluid clearance was unhindered in AQP5-null mice indicating that high alveolar water permeability is not required for active, near-isosmolar fluid transport [146]. The same group used mice deficient in each of the three principal lung aquaporins, AQP1, AQP4, and AQP5, to test the hypothesis that aquaporins are important in neonatal lung fluid balance, adult lung fluid clearance, and formation of lung edema following acute lung injury [147]. They found that despite aquaporins’ role in osmotically driven lung water transport, they are not required for the physiological clearance of lung water in the neonatal or adult lung, or the accumulation of extravascular lung water in the injured lung [147]. These findings support the expected function of AQPs in active fluid absorption and secretion, but they suggest that the requirement for AQP-facilitated water transport depends on the rate of water transport [124,148].
2.2.3. Aquaporins and ARDS
The functional characterization of AQPs, in addition to their tissue- and cell-specific distribution has garnered considerable scientific interest, particularly in understanding their role in pathological conditions, including ARDS and lung injury in critically ill patients [124]. Dysregulation of AQPs has been linked to the pathophysiology of ARDS. During lung injury and inflammation, changes in aquaporin expression and function can either worsen or improve the severity of fluid accumulation. Moreover, oxidative stress and the inflammatory environment of ARDS can directly impact aquaporin function [76,149].
To the best of our knowledge, the first study to report enhanced AQP1 expression levels in ARDS patients was performed by Lai et al. [150]. The study focused on post-mortem lung tissue samples from ARDS patients. Tissue samples were collected within 48-hrs from the time of death and immediately after the autopsy. Prior to ARDS development, patients were healthy, while the causes of ARDS included trauma, sepsis, stroke complicated by multi-organ failure, and burns. AQP1 was found to be constitutively expressed in the alveolar endothelium. Compared to lung tissue samples from age- and sex-matched non-smoking patients who had died of non-pulmonary diseases in the ICU, AQP1 expression was found to be mildly elevated (33%) in the alveolar capillary endothelium of ARDS patients. Based on these findings, they suggested that AQP1 upregulation could be, to a minor extent, a potential contributing factor of alveolar flooding in ARDS.
In a genomic expression analysis of 5 polytrauma, initially non-septic patients who were admitted to the ICU, 2 of whom developed sepsis and 3 septic shock and ARDS, many genes were found to be upregulated. To validate these genes, Vassiliou et al. corroborated their results with published expression profiling studies using animal models of LPS-induced and aseptic lung injury. This analysis revealed three genes that were commonly dysregulated in both human patients with severe sepsis and ARDS and in experimental models, among them AQP1 [95].
A study performed on mechanically ventilated critically ill COVID-19 patients with respiratory failure due to viral pneumonia, showed that serum AQP1 levels were higher in the patient group compared to the control group of 45 healthy individuals with negative PCR tests for SARS-CoV-2 and no chronic disease [151].
The AQP5 promoter 1364A/C polymorphism has also been studied in the context of ARDS. DNA was extracted from whole blood of 136 ARDS patients, and samples were genotyped for the 1364A/C SNP. The results indicated that the patients carrying the AQP5 AA genotype showed decreased recovery from AKI and increased mortality. Patients carrying the AQP5 AC genotype presented with significantly higher AKI recovery rates on day 30 compared to AA carriers. Hence, decreased AQP5 expression seems to be associated with increased recovery from AKI in ARDS patients [152]. Using the same pool of 136 patients with ARDS induced by either bacterial pneumonia or primary extrapulmonary sepsis with a secondary bacterial pneumonia, Rahmel et al. also investigated whether the AQP5 1364A/C SNP is associated with pulmonary inflammation and survival in ARDS. Based on their results, the presence of the AA genotype of the AQP5 1364A/C polymorphism was associated with aggravated pulmonary inflammation, as indicated by the elevated bronchoalveolar lavage protein and leukocyte concentrations, while ARDS patients with the C-allele of the AQP5 1364A/C promoter polymorphism had attenuated pulmonary inflammation and higher 30-day survival. However, the study was unable to define the mechanisms in which the AA and AC/CC genotypes contribute to mortality in ARDS since it omitted quantitative histochemistry and pulmonary biopsies [27].
Xie et al. analyzed the differentially expressed genes in the mRNA expression profile dataset GSE32707, which included mRNA expression data from whole-blood samples of 33 ARDS patients and 34 healthy control individuals. While the initial results of the analysis showed increased AQP9 mRNA expression in ARDS patients, a subsequent validation analysis in a different group of ARDS patients and healthy individuals did not confirm these findings [153].
The involvement of lncRNA-5657 in sepsis-induced ARDS has also been investigated. lncRNA-5657 expression was measured in the bronchoalveolar lavage fluid (BALF) cells of 15 patients with sepsis-induced ARDS. Compared to non-septic and non-ARDS patients, lncRNA-5657 BALF expression was found significantly elevated in sepsis-induced ARDS patients. Moreover, sepsis-induced ARDS patients also had higher levels of TNF-α and IL-1β. Based on the above findings and in vitro and in vivo data, they suggested that lncRNA-5657 was involved in the development of sepsis-induced ARDS [154]. While lncRNA-5657 has not been demonstrated to regulate aquaporin expression in experimental sepsis-induced ARDS, a connection between lncRNA-5657 and AQP4 has been identified in sepsis-associated encephalopathy (SAE) [155,156].
Data from an in vivo LPS-induced ALI model supported that miR-126-5p had a protective role, maintaining the expression of AQP1 [157]. Recently, Mao et al. investigated the miR-126-5p expression within 24 h of ICU admission in the plasma of 120 septic patients, 60 of whom had ARDS. Based on their results, miR-126-5p plasma levels were reduced in the septic patients with ARDS compared to the septic patients without ARDS and the healthy controls, while miR-126-5p could differentiate between sepsis patients with and without ARDS. Moreover, miR-126-5p expression negatively correlated with various systemic inflammatory markers, while positively correlated with immune function indicators, including immunoglobulins and T-cells in patients with sepsis-induced ARDS [158].
2.2.4. Proposed Mechanisms of Aquaporin Involvement in ARDS
Data from experimental studies have provided insight into how AQPs might be involved in ARDS.
i) Role in Pulmonary Edema Formation
In ARDS, the integrity of the alveolar-capillary barrier is disrupted due to inflammation, leading to increased permeability and leakage of fluid into the alveolar space (pulmonary edema). Aquaporins, particularly AQP1 (in endothelial cells) and AQP5 (in alveolar epithelial cells), are involved in the water movement across the alveolar-capillary barrier. Under physiological conditions, they usually facilitate the management of this fluid by transporting it out of the alveolar space into the interstitium or capillaries. Hence, increased water transport by AQPs could contribute to fluid accumulation in the alveoli if the inflammation and barrier dysfunction are uncontrolled, exacerbating pulmonary edema. On the other hand, when the expression or function of these AQPs is disrupted, as is often observed in ARDS, the lung’s ability to clear fluid is compromised, leading to worsened pulmonary edema. This effect has been observed in lung injury models using lipopolysaccharide (LPS) exposure, which suppresses AQP1 expression and exacerbates the fluid imbalance within the lungs [29,157,159,160,161,162,163,164,165,166]. Interventions that counteract AQP1 downregulation have shown the potential in mitigating these effects by improving fluid regulation and reducing inflammatory cytokine levels [167,168].
ii) Role in Pulmonary Edema Clearance
Conversely, efficient clearance of excess fluid from the alveolar space is critical for the resolution of ARDS. AQPs also play a role in clearing excess fluid from the alveoli. Alveolar epithelial cells use AQP5 to help reabsorb water from the alveolar space, transferring it into the interstitial and vascular compartments. Impairment or downregulation of AQPs, especially AQP5, has been observed in experimental lung injury models, resulting in impaired alveolar fluid clearance and contributing to respiratory failure. AQP5 expression has been shown to decrease in alveolar epithelial cells during experimental inflammatory conditions, reducing the ability to clear alveolar fluid [29,159,160,161,162,163,164,166,169,170,171,172].
Impairment in alveolar fluid clearance due to altered functional expression of respiratory AQPs highlights their pathophysiological significance in pulmonary edema-associated respiratory illness.
iii) Role in the Inflammatory Response
AQPs have been implicated in modulating inflammation in the lungs. Studies suggest that the expression of certain AQPs is altered in response to pro-inflammatory cytokines (e.g., TNF-α, IL-1β), which are abundant in ARDS [75,76]. Changes in AQP expression may affect not only water transport but also cellular processes like migration of inflammatory cells and maintenance of the alveolar epithelial barrier, influencing the overall course of ARDS [148].
iv) Oxidative Stress and Injury
Oxidative stress, a hallmark of ARDS, can impact AQP function. Reactive oxygen species (ROS) generated during inflammation and lung injury can modify AQPs, leading to changes in their permeability and function [76]. This may further contribute to edema and impaired fluid clearance.
In summary, aquaporins, especially AQP1 and AQP5, are involved in the pathophysiology of ARDS by regulating fluid transport and maintaining the balance between edema formation and clearance. Their dysregulation due to inflammation or injury can exacerbate alveolar fluid accumulation and contribute to respiratory distress.
2.2.5. Aquaporins as Therapeutic Targets in ARDS
Due to their role in water transport and edema formation, AQPs have been proposed as potential therapeutic targets for managing lung injury in ARDS. The modulation of AQP function by enhancing the activity or expression of AQPs could improve alveolar fluid clearance and reduce pulmonary edema. Repurposed FDA-approved drugs like Niclosamide look promising, as they have been shown to increase AQP5 abundance and reduce its degradation, which could help to counteract water transport impairments that often exacerbate lung injury. Moreover, high-throughput screenings of drug libraries have identified compounds that promote AQP5 stability, suggesting a potential therapeutic avenue for enhancing AQP5 expression in lung injury management [173]. Approaches to increase AQP5 levels or activity have been investigated in animal models but have yet to translate into clinical therapies [171,172].
On the other hand, where overactive AQPs contribute to excess water influx into the alveoli, aquaporin inhibitors could be used [149]. Therefore, AQP inhibitors could be explored to reduce water movement into the lung during the early stages of injury. This approach is still experimental and requires further research to assess its safety and efficacy.
A recent review by Lotsios et al. extensively refers to these experimental therapeutic approaches [34].
Finally, gene therapy targeting AQPs to restore their normal function or the development of small molecules that modulate the activity of aquaporins could represent future treatment options for ARDS. These therapies would aim to correct the fluid imbalances, a hallmark of the disease.
AQP-based therapies are still experimental, but modulation of these channels may offer new strategies for treating ARDS and other forms of lung injury in critically ill patients. Understanding the precise role of aquaporins in ARDS remains an active area of research. Their importance in lung water regulation suggests that they could be key to developing novel therapeutic interventions for this devastating condition.
2.2.6. Conclusion
Aquaporins, particularly AQP1 and AQP5, are critical in regulating water transport in the lungs. Their dysregulation in ARDS leads to impaired fluid clearance, exacerbating pulmonary edema and worsening respiratory failure. AQPs contribute significantly to the development and resolution of pulmonary edema (excess fluid in the lungs), which impairs gas exchange and worsens respiratory failure in critically ill patients. A reduction in the expression or function of specific aquaporins, such as AQP1 or AQP5, can impair fluid clearance from the alveoli, leading to ongoing pulmonary edema. Conversely, overexpression or increased activity of certain aquaporins may cause fluid imbalance by allowing excessive fluid to enter the lung tissue or alveolar space.
Although research on targeting aquaporins as a therapeutic strategy is still in its early stages, they hold promise for future interventions aimed at improving fluid management in ARDS and other critical lung injuries.
2.3. The Role of Aquaporins in ICU Patients with Acute Kidney Injury
2.3.1. Acute Kidney Injury (AKI)
Acute kidney injury (AKI) impacts 30–60% of critically ill patients, contributing significantly to acute morbidity and mortality [174]. Emerging evidence highlights that the consequences of AKI extend beyond the immediate phase, increasing risks for chronic kidney disease progression, cardiovascular complications, recurrent AKI episodes, and long-term mortality [175]. The Kidney Disease: Improving Global Outcome (KDIGO) workgroup proposed a consensus definition and staging system for clinical practice (the KDIGO definition) that relies on the increase of serum creatinine (of at least ≥ 0.3 mg/dL within the first 48 h, or ≥ 1.5 times the baseline value within 7 days) and/or the presence of oliguria (< 0.5 ml/Kg/h for at least 6 h), both surrogate markers of GFR [176].
2.3.2. Localization and Physiology of AQPs in the Kidney
AQP1 is a highly selective water channel found in the apical and basolateral plasma membranes of the proximal tubule, the descending thin limbs of Henle, and the descending vasa recta, where it facilitates water reabsorption [177]. Mice lacking AQP1 exhibit polyuria, underscoring its essential role in creating hypertonic conditions [178,179].
AQP2 is a crucial channel protein for regulating urine concentration, located on the apical membrane of principal cells in the collecting duct [180]. Its function in water reabsorption is largely regulated by arginine vasopressin, which increases intracellular cyclic adenosine monophosphate production and promotes serine phosphorylation to facilitate AQP2 trafficking to the plasma membrane [181,182,183]. Normal expression of AQP2 on the apical plasma membrane is essential for renal urine concentration and overall body water balance. AQP2 gene deletion or mutation leads to severe water imbalance and can result in nephrogenic diabetes insipidus [24].
AQP3 facilitates the transport of glycerol and hydrogen peroxide across the cell membrane, influencing intracellular signaling pathways that impact key cellular functions, including proliferation, apoptosis, and migration [184]. AQP7, expressed in the brush border of the S3 segment of the proximal tubule, plays a significant role in metabolism by regulating glycerol transport. While impaired AQP7 expression minimally affects water permeability in proximal tubules, it is linked to serious metabolic disorders, such as obesity and insulin resistance [185]. AQP8 enables bidirectional water and hydrogen peroxide transport across biological membranes [186]. AQP11 is uniquely located in the endoplasmic reticulum membrane of the proximal tubular cells. Interestingly, AQP11 knockout mice develop uremia due to polycystic renal disease [187].
Figure 4 depicts the localization and function of the AQP members in the kidney.
2.3.3. Aquaporins and AKI
Urine AQP2 levels increase in several clinical conditions, such as the syndrome of inappropriate secretion of antidiuretic hormone [188], cirrhosis [189], pregnancy [190], and diabetic nephropathy [191]. On the contrary, histological analysis revealed loss of AQP2 expression in the collecting ducts in patients with elevated bilirubin and cholemic nephropathy (CN), and consequently, the loss of AQP2 in such patients might be the result of toxic effects of cholestasis and in part be responsible for the impairment of renal function [192].
Given the potential diagnostic role for AKI, Chan et al. [193] aimed at determining whether urine AQP2 can predict AKI in patients with acute decompensated heart failure. They conducted a prospective, observational study in a coronary care unit, including 189 patients. AKI was diagnosed in 69 (36.5%) patients, and the median urine AQP2 levels were 61.5 ng/mL and 30.9 ng/mL, respectively, in the AKI and non-AKI groups (p<0.001). Urine AQP2 remained significantly associated with the risk of AKI despite adjustment for other covariates. Furthermore, the area under the receiver operating characteristic curve of urine AQP2 for AKI diagnosis demonstrated an acceptable value (0.795).
In humans, an altered AQP5 expression is linked with a common single nucleotide polymorphism (SNP; -1364A/C; rs3759129) in the AQP5 gene promoter [117]. Substitution of cytosine for adenosine at position -1364 is associated with decreased AQP5 expression [117] and has a significant impact on survival in patients suffering from sepsis [118]. Another study assessed the hypothesis that the AQP5 -1364A/C promoter polymorphism is associated with the duration and recovery of AKI in patients with ARDS. One hundred thirty-six patients with ARDS [79 males (58%), 57 females (42%), mean age: 43.7 years)] were prospectively included. ARDS developed in 110 cases (81%) by bacterial pneumonia and 26 cases (19%) by primary extrapulmonary sepsis with secondary bacterial pneumonia leading to ARDS. The ARDS patients were assigned to two groups (AA genotype vs. AC/CC genotype) depending on the -1364A/C polymorphism in the AQP5 gene promoter. The primary end point was AKI presence on day 30 of ICU stay. Homozygous AA genotypes (57%) showed an increased prevalence of AKI compared to AC/CC genotypes (24%), p=0.001. Furthermore, the AA genotype proved to be a strong, independent risk factor for predicting AKI persistence after adjustment for confounders (odds-ratio: 3.35; 95%-confidence interval: 1.2–9.0; p=0.017) [152].
2.3.4. Proposed Mechanisms of Aquaporin Involvement in AKI
Experimental studies have shed light on how AQPs may play a role in AKI.
i) Water Reabsorption and Urine Concentration
AQP1, AQP2, AQP3, and AQP4 are predominantly expressed in the kidneys and play crucial roles in water reabsorption, urine concentration, and maintaining water-electrolyte balance, as described earlier. AQP1, which is highly expressed in the proximal tubule and descending limb of the loop of Henle, supports passive water reabsorption. In cases of AKI, AQP1 dysfunction or downregulation can disrupt this process, leading to impaired water reabsorption, electrolyte imbalances, and fluid retention [177]. Regarding AQP2, studies show that its expression is downregulated in lipopolysaccharide (LPS)-induced AKI, contributing to impaired urinary concentration during sepsis [194]. Notably, downregulation of AQP2 and sepsis-induced AKI progression were alleviated by reduced activation of nuclear factor kappa B (NF-κB) signaling pathways [195].
ii) Inflammation and Oxidative Stress
In rats with LPS-induced AKI, AQP1 expression is significantly upregulated in the kidneys, making it a valuable biomarker for diagnosing septic AKI [196]. Additionally, higher levels of miR-144-3p have been linked to decreased AQP1 expression, potentially affecting kidney function during LPS-triggered systemic inflammation [197]. Interestingly, AQP1 also has a protective function in LPS-induced AKI by promoting M2 macrophage polarization [198]. Another study suggests that AQP1 helps protect against AKI by modulating the inflammatory response, reducing apoptosis, and attenuating fibrosis through downregulation of P53 in septic AKI or LPS-induced HK-2 cells [199]. Furthermore, AQP8 facilitates hydrogen peroxide (a reactive oxygen species) transport, and its dysregulation can heighten oxidative stress in renal tubular cells, leading to cell damage, mitochondrial dysfunction, and apoptosis during AKI [186].
iii) Tubular Cell Apoptosis and Necrosis
Lei et al. demonstrated that AQP3 knockdown exacerbates kidney injury by inhibiting mitogen-activated protein kinase (MAPK) signaling and increasing apoptosis in ischemia/reperfusion (I/R) injury in mice [181]. Reduced AQP3 levels may negatively affect renal cell viability after I/R injury, primarily through apoptosis induction. Presumably, increased permeability of AQP-mediated water channels in response to injury can lead to cell swelling or shrinkage, which triggers programmed cell death pathways. In severe AKI cases, excessive water influx and subsequent cell lysis can result in necrosis, intensifying inflammation, and kidney damage [200].
iv) Renal ischemia/reperfusion-induced AKI
Renal I/R injury is a common cause of AKI, and research has shown a strong association between AQP expression and I/R-induced AKI. For instance, alterations in urine AQP2 levels have been reported in animal models of I/R-induced AKI [201]. Notably, erythropoietin treatment prevented the downregulation of AQPs and sodium transporters in renal I/R injury, and this may play a critical role in improving I/R-induced urinary concentrating defects and impairment of tubular sodium reabsorption [202].
V) Edema Formation
Histological signs of necrosis are typically more common in proximal tubules than in distal tubules. This discrepancy may be due to the relatively high hydraulic conductivity of proximal tubule membranes, which promotes swelling secondary to sodium influx. AQP1, the primary water channel in the proximal tubule, plays a significant role in water reabsorption and may thus contribute to cell swelling [200].
2.3.5. Aquaporins as Therapeutic Targets in AKI
As discussed above, AQPs are vital water channel proteins involved in kidney function, making them promising therapeutic targets in AKI, by addressing fluid imbalance and cellular damage [133]. Modulation of AQPs, such as restoring AQP1 and AQP3 levels or function or stabilizing AQP2 to enhance water reabsorption, could prove beneficial in the treatment of loss-of-function aquaporin diseases such as nephrogenic diabetes insipidus (NDI), among others [203,204]. Indeed, AQP1, AQP2, AQP3, and AQP4-null mice exhibit diuresis, so restoration of these AQPs shows potential to treat NDI, while their inhibitors are predicted to be novel diuretics [24].
Gene therapy approaches could also regulate AQP expression to support tissue recovery or mitigate inflammation. Pharmacological agents, such as AQP-targeting small molecules, osmotic agents, and vasopressin analogs, offer potential for modulating AQP activity. Anti-inflammatory and antioxidant therapies, like steroids and N-acetylcysteine, could stabilize AQPs and reduce oxidative stress [24].
AQP1 and AQP2 levels in the urine (uAQP1 and uAQP2) have been shown to reflect their respective renal expression levels. Urine levels decrease significantly in animal models of AKI, including gentamicin [205] and I/R-induced AKI rats [201], while studies have reported that AQP5 levels are significantly upregulated in the collecting ducts of patients with diabetic nephropathy (DN) [191,206,207], suggesting that urine levels of these AQPs might be novel noninvasive biomarkers to diagnose DN. It has also been suggested that monitoring AQP levels as biomarkers might guide treatment and track AKI progression [208].
Despite these promising avenues, challenges remain in ensuring specificity, safety, and effective delivery systems, necessitating further research to translate these strategies into clinical practice.
2.3.6. Conclusions
AQPs are expressed in the kidneys and play crucial roles in water reabsorption, urine concentration, and maintaining water-electrolyte balance. Preclinical studies hint at the crucial roles AQPs might play in the development and evolution of AKI. However, there is a lack of clinical studies on their role in the pathogenesis as well as treatment of AKI syndromes in humans, especially in the ICU setting.
2.4. The Role of Aquaporins in ICU Patients with Acute Brain Injury
2.4.1. Acute Brain Injury (ABI)
Acute brain injury (ABI), such as traumatic brain injury (TBI), subarachnoid hemorrhage (SAH), acute ischemic stroke (AIS), and intracerebral hemorrhage (ICH), are prevalent in intensive care units (ICUs) due to their severe and life-threatening nature. These conditions often lead to significant neurological deficits and systemic complications, including elevated intracranial pressure, cerebral edema, and impaired cerebral perfusion, necessitating comprehensive monitoring and intervention [209]. Effective management of ABI patients in the ICU requires a multidisciplinary approach that incorporates advanced neuroimaging techniques, hemodynamic support, and strategies aimed at preventing secondary brain injury. Despite progress in critical care practices, patient outcomes remain varied, highlighting the ongoing need for research into the underlying mechanisms and optimal therapeutic approaches for this vulnerable group. Prompt detection and targeted treatment of complications are essential for enhancing survival rates and functional recovery in ABI patients [210].
2.4.2. Localization and Physiology of Aquaporins in the Brain
Under physiological conditions, AQPs are strategically expressed in the brain to maintain water homeostasis and facilitate cerebrospinal fluid (CSF) dynamics [211]. AQP1 is found in the choroid plexus, where it plays a key role in CSF production by enabling water transport across epithelial cells. AQP1-knockout mice exhibit impaired CSF production due to the lack of water transport in the choroid plexus [211]. AQP2 is expressed in astrocytes [212]. AQP4, the most abundant AQP in the brain, is highly expressed in astrocyte end-feet surrounding blood vessels, the glia limitans, and ependymal cells, where it regulates water exchange between the brain parenchyma and blood. AQP4 is also essential for the glymphatic system, which facilitates waste clearance during sleep. AQP4-null mice demonstrate significant alterations in water homeostasis and astrocytic function [9,123,213,214]. In models of cytotoxic edema (e.g., ischemic stroke), these mice show reduced brain swelling, suggesting a protective effect against water influx into the brain parenchyma [213,215,216,217]. Conversely, in cases of vasogenic edema (e.g., trauma or tumors), AQP4-null mice have impaired water clearance, exacerbating fluid accumulation [218,219]. AQP9 is expressed in astrocytes and some neurons, particularly in metabolically active regions, allowing the transport of small solutes like glycerol and lactate to support energy metabolism [220]. Together, these AQPs contribute to the brain’s water balance, metabolic support, and waste clearance under normal physiological conditions [80,215].
2.4.3. Aquaporins and ABI
AQP4, the most abundant aquaporin in the central nervous system, is particularly involved in the regulation of brain edema and the response to traumatic brain injury (TBI), including subdural hematomas (SDH) and acute ischemic stroke (AIS). Genetic variations in AQP4 have been linked to clinical outcomes in TBI. A study by Dardiotis et al. identified several single nucleotide polymorphisms (SNPs) in the AQP4 gene that were significantly associated with functional outcomes six months post-TBI. The SNPs rs3763043 and rs3875089 were not associated with the initial severity of the injury but were correlated with patient recovery, emphasizing AQP4’s role in influencing edema and neural repair processes [221].
Elevated levels of AQP4-expressing microparticles in the bloodstream further highlighted the significance of this aquaporin in TBI. A study demonstrated that patients with severe TBI had significantly higher concentrations of AQP4-positive microparticles in arterial and cerebrovenous blood compared to healthy controls. These findings suggest that AQP4-derived microparticles could serve as biomarkers for the extent of brain injury and systemic involvement [222].
Significantly elevated AQP4 levels were reported in the CSF of patients with severe TBI compared to healthy individuals [223]. Notably, AQP4 levels tended to increase further in patients whose intracranial pressure (ICP) was successfully managed, indicating its role in the metabolic response to brain water regulation.
In AIS, AQP4 is implicated in early neurological deterioration (END) due to its association with oxidative stress and blood-brain barrier (BBB) disruption. Elevated systolic blood pressure (SBP) within 24-hrs of thrombolysis was directly associated with increased AQP4 levels, along with markers of oxidative stress such as malondialdehyde (MDA) and matrix metalloproteinase-9 (MMP-9). These findings suggest that AQP4 contributes to BBB dysfunction and water accumulation, exacerbating ischemic damage and clinical deterioration [224].
The pathology of sepsis-associated encephalopathy (SAE) is linked to astrocyte inflammation, which is associated with AQP4. In patients with SAE, AQP4 protein levels were found to be elevated in peripheral blood. The authors, using a cecal ligation and perforation (CLP) mouse model, suggested that learning and memory impairments in SAE were mitigated by AQP4 knockout. This effect was achieved by activating astrocytic autophagy, reducing neuroinflammation, and ultimately providing neuroprotection [156].
Finally, in patients with acute and chronic subdural hematomas, plasma levels of AQPs, including AQP2 and AQP4, have shown potential as biomarkers of disease severity. AQP2 plasma concentrations strongly correlated with chronic hematoma volume and midline shift, highlighting its significance in chronic disease processes. However, no significant correlations were observed for AQP1, AQP4, and AQP9 in acute subdural hematomas, suggesting distinct roles of aquaporins in different phases and types of brain injury [225].
Human studies in populations not treated in an ICU setting provide further evidence on the importance of AQP function in ABI. Elevated AQP4 levels in both serum and brain tissue have been demonstrated, with higher levels observed in cases of more severe injury [226]. AQP4 levels can remain elevated post-injury, indicating a prolonged response to trauma [227,228]. In addition to AQP4, AQP1 may also contribute to edema formation following TBI [229]. Postmortem studies reinforce the role of AQP4 in AIS-related edema, revealing its upregulation in astrocytes at the ischemic boundary [230,231].
2.4.4. Proposed Mechanisms of Aquaporin Involvement in ABI
Human and animal studies have identified an increasing number of pathophysiological mechanisms for aquaporin involvement in ABI. These are depicted in Figure 5.
i) Cerebral Edema Formation
AQP4 facilitates water influx into the brain parenchyma, contributing to edema after ischemia, hemorrhage, or trauma. Its upregulation in reactive astrocytes around lesion sites intensifies water transport and swelling [232,233]. In white matter, AQP4 expression increases significantly, resulting in greater swelling compared to cortical areas [231]. AQP1 has been shown to increase in astrocytes under pathological conditions, such as multiple sclerosis and cerebral infarction. This suggests that AQP1, alongside AQP4, may contribute to the astrocytic response to brain injury, possibly influencing glial scar formation and edema regulation. Notably, AQP1’s response to inflammatory cytokines differs from that of AQP4, suggesting distinct roles in CNS injury repair [234]. Comparisons of AQP4 and AQP1 expression in ischemic areas reveal that AQP4 is more prevalent, particularly in the cortex and scarred regions, and has a stronger association with basement membranes, highlighting its dominant role in edema-related water transport [235].
In SAH, AQP4 expression in astrocytes increases, especially around blood vessels, and appears to aid in water distribution across the brain’s perivascular spaces. Following SAH, blood enters the brain’s paravascular spaces, leading to vasospasm, neuroinflammation, and microvascular dysfunction. AQP4 is central in modulating water movement in these spaces, and its absence exacerbates the neurological deficits and neuroinflammation caused by SAH, as demonstrated by studies using AQP4 knockout models. This suggests that while AQP4 facilitates fluid movement, it may also play a protective role in managing inflammation post-SAH [236].
AQP1 also plays a role in SAH, particularly in edema formation around the hemorrhage site. Both AQP1 and AQP4 expression is markedly upregulated in human brain tissue following SAH, particularly on astrocytic processes, suggesting that AQP1 and AQP4 contribute to the brain’s reactive response in regulating edema [237]. Interestingly, in SAH cases, the usual polarization of AQP4 on astrocytic end-feet is lost, implying that AQP4 may be redistributed to better manage edema during acute brain injuries like SAH. However, the precise functions of AQP1 in water management post-SAH remain less understood and require further exploration [237].
ii) Blood-Brain Barrier Disruption
Elevated blood pressure post-stroke enhances oxidative stress and AQP4-mediated BBB permeability, exacerbating edema and early neurological deterioration [224]. Several studies highlight the critical role of AQP4 in response to injury or toxic exposure. Wang et al. examined cases of methamphetamine intoxication and found increased AQP4 expression alongside elevated MMP-9 levels and reduced claudin-5 (CLDN5) expression. This suggests that methamphetamine use disrupts BBB integrity by modifying tight junctions and matrix composition, while increased AQP4 expression may serve as a compensatory mechanism to remove excess water from the brain’s extracellular space, potentially mitigating edema [60,134]. In ICH, miR-27a-3p loss deregulates AQP11 in endothelial cells, impairing BBB integrity and increasing edema [238].
iii) Inflammatory Signaling Pathways
Aquaporins, particularly AQP4, also play a significant role in the pathophysiology of SAE, characterized by vasogenic cerebral edema and cognitive impairment. During SAE, AQP4 is upregulated in response to cerebral inflammation driven by neutrophil infiltration, which exacerbates edema [239,240,241]. AQP4 also interacts with toll-like receptor 4 (TLR4) pathways, promoting microglial activation and cytokine release in TBI and ICH [212,242]. Moreover, AQP2 expression in astrocytes drives microglial activation toward a pro-inflammatory phenotype, aggravating neuroinflammation [212].
iv) Metabolic and Ionic Homeostasis
AQP9 contributes to lactate transport, supporting metabolic adaptation under ischemic conditions [243]. AQP4 collaborates with potassium channels like Kir4.1, balancing ionic and water transport in injured brain regions [244].
v) Genetic Susceptibility
SNPs in the AQP4 gene, such as rs9951307 (stroke) and rs1058427 (ICH), correlate with severe edema and poor outcomes, highlighting a genetic basis for edema vulnerability [245,246]. Other AQP4 polymorphisms have also been associated with altered ICH risk and onset age, although their significance is uncertain after correction for multiple comparisons, suggesting a complex genetic influence on AQP4 expression and cerebral edema vulnerability [247].
2.4.5. Aquaporins as Therapeutic Targets in ABI
Aquaporins, particularly AQP4, present promising therapeutic targets in the treatment of ABI. AQP4 inhibitors such as AER-270 and acetazolamide have demonstrated significant potential in reducing cerebral edema and improving outcomes in animal models of ABI. AER-270, a small-molecule inhibitor of AQP4-mediated water permeability, has been shown to reduce brain swelling in models of both TBI and stroke, highlighting its therapeutic potential [248].
Acetazolamide has also been investigated for its role in reducing edema in the early stages of stroke. While it has shown promise in reducing swelling, its effect on long-term recovery remains limited, indicating the need for further optimization [249].
MicroRNA-145 (miR-145) has been explored as a potential therapeutic agent for ABI. In animal models of ischemic stroke, miR-145 overexpression has been shown to downregulate AQP4, providing neuroprotection by reducing astrocytic injury and edema formation. This approach suggests that targeting the post-transcriptional regulation of AQP4 could be a novel therapeutic strategy to mitigate brain injury [250].
In the context of ICH, AQP2 expression has been linked to increased inflammation and edema. Targeting AQP2, particularly through the modulation of inflammatory pathways like TLR4/NFκB, could reduce neuroinflammation and edema, improving recovery outcomes [212]. Additionally, miR-27a-3p mimics, which target AQP11 in endothelial cells, have shown promise in enhancing BBB integrity and reducing edema after ICH [238].
Experimental models of sepsis underscore AQP4’s involvement in cognitive decline, as AQP4 deletion mitigates memory deficits, reduces neuroinflammation, and downregulates pro-inflammatory cytokines [156]. Increased AQP4 in endotoxemia-induced encephalopathy further implicates its role in SAE-related cognitive dysfunction. Collectively, these findings position AQP4 as a critical mediator in SAE pathogenesis, presenting it as a potential therapeutic target to alleviate brain edema and cognitive impairment in sepsis [251]. Moreover, research into spinal cord injury models has demonstrated that selective inhibition of AQP4 reduces edema and improves recovery outcomes, further supporting the idea that AQP4 modulation could benefit a wide range of CNS injuries [252]. These suggests that AQP4 inhibitors may have broad therapeutic potential beyond ABI, offering a strategy for managing brain and spinal cord injuries. In SAE and other inflammatory brain injuries, corticosteroids such as dexamethasone can reduce AQP4 expression by modulating inflammatory pathways. This reduction may help alleviate cerebral edema and improve cognitive function. However, the use of corticosteroids in such conditions remains controversial due to potential side effects [253].
Beyond water transport, AQP4 interacts with glutamate transporter-1 (GLT-1), the primary astrocytic glutamate buffer. In ischemic brain tissue, AQP4 and GLT-1 have distinct spatial distributions, with AQP4 concentrated around blood vessels and GLT-1 near neurons. This coordinated distribution suggests a role for astrocytes in both water and neurotransmitter regulation, with potential therapeutic implications for targeting AQP4 and GLT-1 to enhance water and glutamate buffering in ischemic regions [254].
A connection between lncRNA-5657 and AQP4 has been identified in SAE. Inhibiting lncRNA-5657 reduced neuronal degeneration and inflammatory markers, including AQP4, suggesting neuroprotection against septic brain injury [155].
Circulating AQP4 has also shown promise as a biomarker. Baseline serum AQP4 levels in AIS patients undergoing thrombolysis inversely correlated with neurological severity and infarct size, with higher levels associated with better recovery outcomes, suggesting its potential as a prognostic biomarker for recovery [255]. In sepsis-induced delirium (SID), increased astrocytic AQP4 expression, possibly detectable through exosomes, may serve as a biomarker for SID [256].
2.4.5. Conclusions
Aquaporins are crucial in maintaining CNS water homeostasis. Their expression and regulation have been implicated in a variety of neurological and neurosurgical diseases, such as TBI, AIS, SAH, and ICH.
The dual function of AQPs in both injury propagation and recovery highlights their potential as biomarkers for disease severity and therapeutic targets in managing brain injuries and other CNS disorders. Furthermore, the identification of genetic polymorphisms and their links to clinical outcomes underscores the importance of AQPs as potential biomarkers. Ongoing research continues to explore their therapeutic potential, offering hope for improved diagnostic precision and targeted interventions. The intricate and dynamic roles of AQPs highlight their clinical significance in neuroinflammatory and vascular processes and the need for further studies to fully elucidate their therapeutic implications.
Table 2 lists all the studies on aquaporins in critically ill patients with injuries involving the immune system, lung, kidney, and brain.
3. Conclusions
Aquaporins (AQPs) play critical roles in maintaining water homeostasis, regulating cellular function, and supporting tissue integrity, particularly in the context of critical illnesses. In conditions such as sepsis, ARDS, AKI, and ABI, AQPs contribute to fluid balance, inflammation, and cellular stress responses. The regulation of AQPs, either through gene expression changes or post-translational modifications, offers potential therapeutic targets for mitigating the effects of fluid imbalance, brain edema, and inflammation in critical care settings. Understanding the precise mechanisms of AQP involvement in these diseases and their potential role as biomarkers could lead to novel strategies for improving patient outcomes in severe, acute conditions.
Acknowledgments
The authors would like to thank Ms. Dimitra Patsatzi for her significant assistance in adapting Figures 2 and 3.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Denker, B.M.; Smith, B.L.; Kuhajda, F.P.; Agre, P. Identification, purification, and partial characterization of a novel Mr 28,000 integral membrane protein from erythrocytes and renal tubules. J Biol Chem 1988, 263, 15634-15642. [CrossRef]
- Preston, G.M.; Agre, P. Isolation of the cDNA for erythrocyte integral membrane protein of 28 kilodaltons: member of an ancient channel family. Proc Natl Acad Sci U S A 1991, 88, 11110-11114. https://doi.org/10.1073/pnas.88.24.11110.
- Preston, G.M.; Carroll, T.P.; Guggino, W.B.; Agre, P. Appearance of water channels in Xenopus oocytes expressing red cell CHIP28 protein. Science 1992, 256, 385-387. [CrossRef]
- Agre, P.; Preston, G.M.; Smith, B.L.; Jung, J.S.; Raina, S.; Moon, C.; Guggino, W.B.; Nielsen, S. Aquaporin CHIP: the archetypal molecular water channel. Am J Physiol 1993, 265, F463-476. [CrossRef]
- Azad, A.K.; Raihan, T.; Ahmed, J.; Hakim, A.; Emon, T.H.; Chowdhury, P.A. Human Aquaporins: Functional Diversity and Potential Roles in Infectious and Non-infectious Diseases. Front Genet 2021, 12, 654865. [CrossRef]
- Kreida, S.; Tornroth-Horsefield, S. Structural insights into aquaporin selectivity and regulation. Curr Opin Struct Biol 2015, 33, 126-134. [CrossRef]
- Verkman, A.S. Physiological importance of aquaporin water channels. Ann Med 2002, 34, 192-200.
- Verkman, A.S.; Anderson, M.O.; Papadopoulos, M.C. Aquaporins: important but elusive drug targets. Nat Rev Drug Discov 2014, 13, 259-277. [CrossRef]
- Verkman, A.S. Mammalian aquaporins: diverse physiological roles and potential clinical significance. Expert Rev Mol Med 2008, 10, e13. [CrossRef]
- Smith, I.M.; Stroka, K.M. The multifaceted role of aquaporins in physiological cell migration. Am J Physiol Cell Physiol 2023, 325, C208-c223. [CrossRef]
- Porter, D.; Johnston, A.M.; Henning, J. Medical conditions requiring intensive care. J R Army Med Corps 2009, 155, 141-146. [CrossRef]
- Joynt, G.M.; Tat, W.W. Ten diseases you need to know if you want to be a critical care specialist in Hong Kong. Intensive Care Med 2014, 40, 1144-1146. [CrossRef]
- Adler, S.M.; Verbalis, J.G. Disorders of body water homeostasis in critical illness. Endocrinol Metab Clin North Am 2006, 35, 873-894, xi. [CrossRef]
- Bouchard, J.E.; Mehta, R.L. Fluid balance issues in the critically ill patient. Contrib Nephrol 2010, 164, 69-78. [CrossRef]
- Bollaert, P.E.; Monnier, A.; Schneider, F.; Argaud, L.; Badie, J.; Charpentier, C.; Meziani, F.; Bemer, M.; Quenot, J.P.; Buzzi, M.; et al. Fluid balance control in critically ill patients: results from POINCARE-2 stepped wedge cluster-randomized trial. Crit Care 2023, 27, 66. [CrossRef]
- Robayo-Amortegui, H.; Quintero-Altare, A.; Florez-Navas, C.; Serna-Palacios, I.; Súarez-Saavedra, A.; Buitrago-Bernal, R.; Casallas-Barrera, J.O. Fluid dynamics of life: exploring the physiology and importance of water in the critical illness. Front Med (Lausanne) 2024, 11, 1368502. [CrossRef]
- Sartori, C.; Matthay, M.A.; Scherrer, U. Transepithelial sodium and water transport in the lung. Major player and novel therapeutic target in pulmonary edema. Adv Exp Med Biol 2001, 502, 315-338. [CrossRef]
- Zemans, R.L.; Matthay, M.A. Bench-to-bedside review: the role of the alveolar epithelium in the resolution of pulmonary edema in acute lung injury. Crit Care 2004, 8, 469-477. [CrossRef]
- Matthay, M.A.; Robriquet, L.; Fang, X. Alveolar epithelium: role in lung fluid balance and acute lung injury. Proc Am Thorac Soc 2005, 2, 206-213. [CrossRef]
- Cash, A.; Theus, M.H. Mechanisms of Blood-Brain Barrier Dysfunction in Traumatic Brain Injury. Int J Mol Sci 2020, 21. [CrossRef]
- Dadgostar, E.; Rahimi, S.; Nikmanzar, S.; Nazemi, S.; Naderi Taheri, M.; Alibolandi, Z.; Aschner, M.; Mirzaei, H.; Tamtaji, O.R. Aquaporin 4 in Traumatic Brain Injury: From Molecular Pathways to Therapeutic Target. Neurochem Res 2022, 47, 860-871. [CrossRef]
- Dutta, A.; Das, M. Deciphering the role of aquaporins in metabolic diseases: A mini review. Am J Med Sci 2022, 364, 148-162. [CrossRef]
- Gu, Y.; Zhou, C.; Piao, Z.; Yuan, H.; Jiang, H.; Wei, H.; Zhou, Y.; Nan, G.; Ji, X. Cerebral edema after ischemic stroke: Pathophysiology and underlying mechanisms. Front Neurosci 2022, 16, 988283. [CrossRef]
- He, J.; Yang, B. Aquaporins in Renal Diseases. Int J Mol Sci 2019, 20. [CrossRef]
- Kuebler, W.M. Acute Respiratory Distress Syndrome: Biomarkers, Mechanisms, and Water Channels. Anesthesiology 2019, 130, 364-366. [CrossRef]
- Ohmura, K.; Tomita, H.; Hara, A. Peritumoral Edema in Gliomas: A Review of Mechanisms and Management. Biomedicines 2023, 11. [CrossRef]
- Rahmel, T.; Rump, K.; Peters, J.; Adamzik, M. Aquaporin 5 -1364A/C Promoter Polymorphism Is Associated with Pulmonary Inflammation and Survival in Acute Respiratory Distress Syndrome. Anesthesiology 2019, 130, 404-413. [CrossRef]
- Szczygielski, J.; Kopańska, M.; Wysocka, A.; Oertel, J. Cerebral Microcirculation, Perivascular Unit, and Glymphatic System: Role of Aquaporin-4 as the Gatekeeper for Water Homeostasis. Front Neurol 2021, 12, 767470. [CrossRef]
- Vassiliou, A.G.; Manitsopoulos, N.; Kardara, M.; Maniatis, N.A.; Orfanos, S.E.; Kotanidou, A. Differential Expression of Aquaporins in Experimental Models of Acute Lung Injury. In vivo 2017, 31, 885-894. [CrossRef]
- Zhou, Z.; Zhan, J.; Cai, Q.; Xu, F.; Chai, R.; Lam, K.; Luan, Z.; Zhou, G.; Tsang, S.; Kipp, M.; et al. The Water Transport System in Astrocytes-Aquaporins. Cells 2022, 11. [CrossRef]
- Czyżewski, W.; Litak, J.; Sobstyl, J.; Mandat, T.; Torres, K.; Staśkiewicz, G. Aquaporins: Gatekeepers of Fluid Dynamics in Traumatic Brain Injury. Int J Mol Sci 2024, 25. [CrossRef]
- Messerer, D.A.C.; Schmidt, H.; Frick, M.; Huber-Lang, M. Ion and Water Transport in Neutrophil Granulocytes and Its Impairment during Sepsis. Int J Mol Sci 2021, 22. [CrossRef]
- da Silva, I.V.; Garra, S.; Calamita, G.; Soveral, G. The Multifaceted Role of Aquaporin-9 in Health and Its Potential as a Clinical Biomarker. Biomolecules 2022, 12. [CrossRef]
- Lotsios, N.S.; Keskinidou, C.; Dimopoulou, I.; Kotanidou, A.; Orfanos, S.E.; Vassiliou, A.G. Aquaporin Expression and Regulation in Clinical and Experimental Sepsis. Int J Mol Sci 2023, 25. [CrossRef]
- Kannan, A.; Panneerselvam, A.; Mariajoseph-Antony, L.F.; Loganathan, C.; Prahalathan, C. Role of Aquaporins in Spermatogenesis and Testicular Steroidogenesis. J Membr Biol 2020, 253, 109-114. [CrossRef]
- Tricarico, P.M.; Mentino, D.; De Marco, A.; Del Vecchio, C.; Garra, S.; Cazzato, G.; Foti, C.; Crovella, S.; Calamita, G. Aquaporins Are One of the Critical Factors in the Disruption of the Skin Barrier in Inflammatory Skin Diseases. Int J Mol Sci 2022, 23. [CrossRef]
- Robertson, G.L. Regulation of arginine vasopressin in the syndrome of inappropriate antidiuresis. Am J Med 2006, 119, S36-42. [CrossRef]
- Chesney, R.W. The role of the kidney in protecting the brain against cerebral edema and neuronal cell swelling. J Pediatr 2008, 152, 4-6. [CrossRef]
- Pasantes-Morales, H.; Cruz-Rangel, S. Brain volume regulation: osmolytes and aquaporin perspectives. Neuroscience 2010, 168, 871-884. [CrossRef]
- Nesic, O.; Guest, J.D.; Zivadinovic, D.; Narayana, P.A.; Herrera, J.J.; Grill, R.J.; Mokkapati, V.U.; Gelman, B.B.; Lee, J. Aquaporins in spinal cord injury: the janus face of aquaporin 4. Neuroscience 2010, 168, 1019-1035. [CrossRef]
- Lukaszewicz, A.C.; Soyer, B.; Payen, D. Water, water, everywhere: sodium and water balance and the injured brain. Curr Opin Anaesthesiol 2011, 24, 138-143. [CrossRef]
- Verbalis, J.G. Acquired forms of central diabetes insipidus: Mechanisms of disease. Best Pract Res Clin Endocrinol Metab 2020, 34, 101449. [CrossRef]
- Michinaga, S.; Koyama, Y. Pathophysiological Responses and Roles of Astrocytes in Traumatic Brain Injury. Int J Mol Sci 2021, 22. [CrossRef]
- Pan, Q.L.; Lin, F.X.; Liu, N.; Chen, R.C. The role of aquaporin 4 (AQP4) in spinal cord injury. Biomed Pharmacother 2022, 145, 112384. [CrossRef]
- Mavroudis, I.; Jabeen, S.; Balmus, I.M.; Ciobica, A.; Burlui, V.; Romila, L.; Iordache, A. Exploring the Potential of Exosomal Biomarkers in Mild Traumatic Brain Injury and Post-Concussion Syndrome: A Systematic Review. J Pers Med 2023, 14. [CrossRef]
- Garcia, T.A.; Jonak, C.R.; Binder, D.K. The Role of Aquaporins in Spinal Cord Injury. Cells 2023, 12. [CrossRef]
- Alfieri, L.; Montana, A.; Frisoni, P.; D’Errico, S.; Neri, M. Application of Aquaporins as Markers in Forensic Pathology: A Systematic Review of the Literature. Int J Mol Sci 2024, 25. [CrossRef]
- Agre, P. The aquaporin water channels. Proc Am Thorac Soc 2006, 3, 5-13. [CrossRef]
- Murata, K.; Mitsuoka, K.; Hirai, T.; Walz, T.; Agre, P.; Heymann, J.B.; Engel, A.; Fujiyoshi, Y. Structural determinants of water permeation through aquaporin-1. Nature 2000, 407, 599-605. [CrossRef]
- Horner, A.; Zocher, F.; Preiner, J.; Ollinger, N.; Siligan, C.; Akimov, S.A.; Pohl, P. The mobility of single-file water molecules is governed by the number of H-bonds they may form with channel-lining residues. Sci Adv 2015, 1, e1400083. [CrossRef]
- Heymann, J.B.; Agre, P.; Engel, A. Progress on the structure and function of aquaporin 1. J Struct Biol 1998, 121, 191-206. [CrossRef]
- Strand, L.; Moe, S.E.; Solbu, T.T.; Vaadal, M.; Holen, T. Roles of aquaporin-4 isoforms and amino acids in square array assembly. Biochemistry 2009, 48, 5785-5793. [CrossRef]
- Ozu, M.; Galizia, L.; Acuña, C.; Amodeo, G. Aquaporins: More Than Functional Monomers in a Tetrameric Arrangement. Cells 2018, 7. [CrossRef]
- Gonen, T.; Walz, T. The structure of aquaporins. Q Rev Biophys 2006, 39, 361-396. [CrossRef]
- Liu, Z.; Shen, J.; Carbrey, J.M.; Mukhopadhyay, R.; Agre, P.; Rosen, B.P. Arsenite transport by mammalian aquaglyceroporins AQP7 and AQP9. Proc Natl Acad Sci U S A 2002, 99, 6053-6058. [CrossRef]
- Carbrey, J.M.; Agre, P. Discovery of the aquaporins and development of the field. Handb Exp Pharmacol 2009, 3-28. [CrossRef]
- Bienert, G.P.; Bienert, M.D.; Jahn, T.P.; Boutry, M.; Chaumont, F. Solanaceae XIPs are plasma membrane aquaporins that facilitate the transport of many uncharged substrates. Plant J 2011, 66, 306-317. [CrossRef]
- Mukhopadhyay, R.; Bhattacharjee, H.; Rosen, B.P. Aquaglyceroporins: generalized metalloid channels. Biochim Biophys Acta 2014, 1840, 1583-1591. [CrossRef]
- Perez Di Giorgio, J.; Soto, G.; Alleva, K.; Jozefkowicz, C.; Amodeo, G.; Muschietti, J.P.; Ayub, N.D. Prediction of aquaporin function by integrating evolutionary and functional analyses. J Membr Biol 2014, 247, 107-125. [CrossRef]
- Wang, Q.; Ishikawa, T.; Michiue, T.; Zhu, B.L.; Guan, D.W.; Maeda, H. Molecular pathology of brain matrix metalloproteases, claudin5, and aquaporins in forensic autopsy cases with special regard to methamphetamine intoxication. Int J Legal Med 2014, 128, 469-474. [CrossRef]
- Finn, R.N.; Chauvigné, F.; Stavang, J.A.; Belles, X.; Cerdà, J. Insect glycerol transporters evolved by functional co-option and gene replacement. Nat Commun 2015, 6, 7814. [CrossRef]
- Garneau, A.P.; Carpentier, G.A.; Marcoux, A.A.; Frenette-Cotton, R.; Simard, C.F.; Rémus-Borel, W.; Caron, L.; Jacob-Wagner, M.; Noël, M.; Powell, J.J.; et al. Aquaporins Mediate Silicon Transport in Humans. PLoS One 2015, 10, e0136149. [CrossRef]
- Byrt, C.S.; Zhao, M.; Kourghi, M.; Bose, J.; Henderson, S.W.; Qiu, J.; Gilliham, M.; Schultz, C.; Schwarz, M.; Ramesh, S.A.; et al. Non-selective cation channel activity of aquaporin AtPIP2;1 regulated by Ca(2+) and pH. Plant Cell Environ 2017, 40, 802-815. [CrossRef]
- Huang, B.; Jin, L.; Zhang, L.; Cui, X.; Zhang, Z.; Lu, Y.; Yu, L.; Ma, T.; Zhang, H. Aquaporin-8 transports hydrogen peroxide to regulate granulosa cell autophagy. Front Cell Dev Biol 2022, 10, 897666. [CrossRef]
- Finn, R.N.; Cerdà, J. Evolution and functional diversity of aquaporins. Biol Bull 2015, 229, 6-23. [CrossRef]
- Litman, T.; Søgaard, R.; Zeuthen, T. Ammonia and urea permeability of mammalian aquaporins. Handb Exp Pharmacol 2009, 327-358. [CrossRef]
- Bhattacharjee, H.; Carbrey, J.; Rosen, B.P.; Mukhopadhyay, R. Drug uptake and pharmacological modulation of drug sensitivity in leukemia by AQP9. Biochem Biophys Res Commun 2004, 322, 836-841. [CrossRef]
- Rosen, B.P.; Tamás, M.J. Arsenic transport in prokaryotes and eukaryotic microbes. Adv Exp Med Biol 2010, 679, 47-55. [CrossRef]
- Herrera, M.; Hong, N.J.; Garvin, J.L. Aquaporin-1 transports NO across cell membranes. Hypertension 2006, 48, 157-164. [CrossRef]
- Zwiazek, J.J.; Xu, H.; Tan, X.; Navarro-Ródenas, A.; Morte, A. Significance of oxygen transport through aquaporins. Sci Rep 2017, 7, 40411. [CrossRef]
- Roche, J.V.; Törnroth-Horsefield, S. Aquaporin Protein-Protein Interactions. Int J Mol Sci 2017, 18. [CrossRef]
- Marples, D.; Knepper, M.A.; Christensen, E.I.; Nielsen, S. Redistribution of aquaporin-2 water channels induced by vasopressin in rat kidney inner medullary collecting duct. Am J Physiol 1995, 269, C655-664. [CrossRef]
- Nielsen, S.; Chou, C.L.; Marples, D.; Christensen, E.I.; Kishore, B.K.; Knepper, M.A. Vasopressin increases water permeability of kidney collecting duct by inducing translocation of aquaporin-CD water channels to plasma membrane. Proc Natl Acad Sci U S A 1995, 92, 1013-1017. [CrossRef]
- Chaumont, F.; Tyerman, S.D. Aquaporins: highly regulated channels controlling plant water relations. Plant Physiol 2014, 164, 1600-1618. [CrossRef]
- da Silva, I.V.; Cardoso, C.; Martínez-Banaclocha, H.; Casini, A.; Pelegrín, P.; Soveral, G. Aquaporin-3 is involved in NLRP3-inflammasome activation contributing to the setting of inflammatory response. Cell Mol Life Sci 2021, 78, 3073-3085. [CrossRef]
- da Silva, I.V.; Mlinarić, M.; Lourenço, A.R.; Pérez-Garcia, O.; Čipak Gašparović, A.; Soveral, G. Peroxiporins and Oxidative Stress: Promising Targets to Tackle Inflammation and Cancer. Int J Mol Sci 2024, 25. [CrossRef]
- Schuoler, C.; Haider, T.J.; Leuenberger, C.; Vogel, J.; Ostergaard, L.; Kwapiszewska, G.; Kohler, M.; Gassmann, M.; Huber, L.C.; Brock, M. Aquaporin 1 controls the functional phenotype of pulmonary smooth muscle cells in hypoxia-induced pulmonary hypertension. Basic Res Cardiol 2017, 112, 30. [CrossRef]
- Wang, C.; Yan, M.; Jiang, H.; Wang, Q.; He, S.; Chen, J.; Wang, C. Mechanism of aquaporin 4 (AQP 4) up-regulation in rat cerebral edema under hypobaric hypoxia and the preventative effect of puerarin. Life Sci 2018, 193, 270-281. [CrossRef]
- Huo, Z.; Lomora, M.; Kym, U.; Palivan, C.; Holland-Cunz, S.G.; Gros, S.J. AQP1 Is Up-Regulated by Hypoxia and Leads to Increased Cell Water Permeability, Motility, and Migration in Neuroblastoma. Front Cell Dev Biol 2021, 9, 605272. [CrossRef]
- Papadopoulos, M.C.; Verkman, A.S. Aquaporin water channels in the nervous system. Nat Rev Neurosci 2013, 14, 265-277. [CrossRef]
- Gomes, A.; da Silva, I.V.; Rodrigues, C.M.P.; Castro, R.E.; Soveral, G. The Emerging Role of microRNAs in Aquaporin Regulation. Front Chem 2018, 6, 238. [CrossRef]
- Fushimi, K.; Sasaki, S.; Marumo, F. Phosphorylation of serine 256 is required for cAMP-dependent regulatory exocytosis of the aquaporin-2 water channel. J Biol Chem 1997, 272, 14800-14804. [CrossRef]
- Leitch, V.; Agre, P.; King, L.S. Altered ubiquitination and stability of aquaporin-1 in hypertonic stress. Proc Natl Acad Sci U S A 2001, 98, 2894-2898. [CrossRef]
- van Balkom, B.W.; van Raak, M.; Breton, S.; Pastor-Soler, N.; Bouley, R.; van der Sluijs, P.; Brown, D.; Deen, P.M. Hypertonicity is involved in redirecting the aquaporin-2 water channel into the basolateral, instead of the apical, plasma membrane of renal epithelial cells. J Biol Chem 2003, 278, 1101-1107. [CrossRef]
- Kamsteeg, E.J.; Hendriks, G.; Boone, M.; Konings, I.B.; Oorschot, V.; van der Sluijs, P.; Klumperman, J.; Deen, P.M. Short-chain ubiquitination mediates the regulated endocytosis of the aquaporin-2 water channel. Proc Natl Acad Sci U S A 2006, 103, 18344-18349. [CrossRef]
- Hoffert, J.D.; Nielsen, J.; Yu, M.J.; Pisitkun, T.; Schleicher, S.M.; Nielsen, S.; Knepper, M.A. Dynamics of aquaporin-2 serine-261 phosphorylation in response to short-term vasopressin treatment in collecting duct. Am J Physiol Renal Physiol 2007, 292, F691-700. [CrossRef]
- Lu, H.J.; Matsuzaki, T.; Bouley, R.; Hasler, U.; Qin, Q.H.; Brown, D. The phosphorylation state of serine 256 is dominant over that of serine 261 in the regulation of AQP2 trafficking in renal epithelial cells. Am J Physiol Renal Physiol 2008, 295, F290-294. [CrossRef]
- Tamma, G.; Robben, J.H.; Trimpert, C.; Boone, M.; Deen, P.M. Regulation of AQP2 localization by S256 and S261 phosphorylation and ubiquitination. Am J Physiol Cell Physiol 2011, 300, C636-646. [CrossRef]
- Rump, K.; Unterberg, M.; Dahlke, A.; Nowak, H.; Koos, B.; Bergmann, L.; Siffert, W.; Schafer, S.T.; Peters, J.; Adamzik, M.; et al. DNA methylation of a NF-kappaB binding site in the aquaporin 5 promoter impacts on mortality in sepsis. Sci Rep 2019, 9, 18511. [CrossRef]
- Wilson, J.L.; Miranda, C.A.; Knepper, M.A. Vasopressin and the regulation of aquaporin-2. Clin Exp Nephrol 2013, 17, 751-764. [CrossRef]
- Markou, A.; Unger, L.; Abir-Awan, M.; Saadallah, A.; Halsey, A.; Balklava, Z.; Conner, M.; Tornroth-Horsefield, S.; Greenhill, S.D.; Conner, A.; et al. Molecular mechanisms governing aquaporin relocalisation. Biochim Biophys Acta Biomembr 2022, 1864, 183853. [CrossRef]
- Kreida, S.; Roche, J.V.; Olsson, C.; Linse, S.; Törnroth-Horsefield, S. Protein-protein interactions in AQP regulation - biophysical characterization of AQP0-CaM and AQP2-LIP5 complex formation. Faraday Discuss 2018, 209, 35-54. [CrossRef]
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.D.; Coopersmith, C.M.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 2016, 315, 801-810. [CrossRef]
- Moon, C.; Rousseau, R.; Soria, J.C.; Hoque, M.O.; Lee, J.; Jang, S.J.; Trink, B.; Sidransky, D.; Mao, L. Aquaporin expression in human lymphocytes and dendritic cells. Am J Hematol 2004, 75, 128-133. [CrossRef]
- Vassiliou, A.G.; Maniatis, N.A.; Orfanos, S.E.; Mastora, Z.; Jahaj, E.; Paparountas, T.; Armaganidis, A.; Roussos, C.; Aidinis, V.; Kotanidou, A. Induced expression and functional effects of aquaporin-1 in human leukocytes in sepsis. Crit Care 2013, 17, R199. [CrossRef]
- da Silva, I.V.; Soveral, G. Aquaporins in Immune Cells and Inflammation: New Targets for Drug Development. Int J Mol Sci 2021, 22. [CrossRef]
- de Baey, A.; Lanzavecchia, A. The role of aquaporins in dendritic cell macropinocytosis. J Exp Med 2000, 191, 743-748. [CrossRef]
- Hara-Chikuma, M.; Chikuma, S.; Sugiyama, Y.; Kabashima, K.; Verkman, A.S.; Inoue, S.; Miyachi, Y. Chemokine-dependent T cell migration requires aquaporin-3-mediated hydrogen peroxide uptake. J Exp Med 2012, 209, 1743-1752. [CrossRef]
- Rump, K.; Rahmel, T.; Rustige, A.M.; Unterberg, M.; Nowak, H.; Koos, B.; Schenker, P.; Viebahn, R.; Adamzik, M.; Bergmann, L. The Aquaporin3 Promoter Polymorphism -1431 A/G is Associated with Acute Graft Rejection and Cytomegalovirus Infection in Kidney Recipients Due to Altered Immune Cell Migration. Cells 2020, 9. [CrossRef]
- Rump, K.; Unterberg, M.; Bergmann, L.; Bankfalvi, A.; Menon, A.; Schäfer, S.; Scherag, A.; Bazzi, Z.; Siffert, W.; Peters, J.; et al. AQP5-1364A/C polymorphism and the AQP5 expression influence sepsis survival and immune cell migration: a prospective laboratory and patient study. J Transl Med 2016, 14, 321. [CrossRef]
- Talwar, S.; Munson, P.J.; Barb, J.; Fiuza, C.; Cintron, A.P.; Logun, C.; Tropea, M.; Khan, S.; Reda, D.; Shelhamer, J.H.; et al. Gene expression profiles of peripheral blood leukocytes after endotoxin challenge in humans. Physiol Genomics 2006, 25, 203-215. [CrossRef]
- Ishibashi, K.; Kuwahara, M.; Gu, Y.; Tanaka, Y.; Marumo, F.; Sasaki, S. Cloning and functional expression of a new aquaporin (AQP9) abundantly expressed in the peripheral leukocytes permeable to water and urea, but not to glycerol. Biochem Biophys Res Commun 1998, 244, 268-274. [CrossRef]
- Matsushima, A.; Ogura, H.; Koh, T.; Shimazu, T.; Sugimoto, H. Enhanced expression of aquaporin 9 in activated polymorphonuclear leukocytes in patients with systemic inflammatory response syndrome. Shock 2014, 42, 322-326. [CrossRef]
- Holm, A.; Karlsson, T.; Vikström, E. Pseudomonas aeruginosa lasI/rhlI quorum sensing genes promote phagocytosis and aquaporin 9 redistribution to the leading and trailing regions in macrophages. Front Microbiol 2015, 6, 915. [CrossRef]
- Vassiliou, A.G., Orfanos, Stylianos, E., Kotanidou, A. Clinical Assays in Sepsis: Prognosis, Diagnosis, Outcomes, and the Genetic Basis of Sepsis. In Sepsis, Vijay, K., Ed.; IntechOpen: Rijeka, 2017; p. Ch. 6.
- Loitto, V.M.; Forslund, T.; Sundqvist, T.; Magnusson, K.E.; Gustafsson, M. Neutrophil leukocyte motility requires directed water influx. J Leukoc Biol 2002, 71, 212-222.
- Thon, P.; Rahmel, T.; Ziehe, D.; Palmowski, L.; Marko, B.; Nowak, H.; Wolf, A.; Witowski, A.; Orlowski, J.; Ellger, B.; et al. AQP3 and AQP9-Contrary Players in Sepsis? Int J Mol Sci 2024, 25. [CrossRef]
- Ziehe, D.; Marko, B.; Thon, P.; Rahmel, T.; Palmowski, L.; Nowak, H.; von Busch, A.; Wolf, A.; Witowski, A.; Vonheder, J.; et al. The Aquaporin 3 Polymorphism (rs17553719) Is Associated with Sepsis Survival and Correlated with IL-33 Secretion. Int J Mol Sci 2024, 25. [CrossRef]
- Benga, G. The first discovered water channel protein, later called aquaporin 1: molecular characteristics, functions and medical implications. Mol Aspects Med 2012, 33, 518-534. [CrossRef]
- Monzani, E.; Bazzotti, R.; Perego, C.; La Porta, C.A. AQP1 is not only a water channel: it contributes to cell migration through Lin7/beta-catenin. PLoS One 2009, 4, e6167. [CrossRef]
- Mercer, T.R.; Dinger, M.E.; Mattick, J.S. Long non-coding RNAs: insights into functions. Nat Rev Genet 2009, 10, 155-159. [CrossRef]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat Rev Mol Cell Biol 2014, 15, 509-524. [CrossRef]
- Fang, Y.; Hu, J.; Wang, Z.; Zong, H.; Zhang, L.; Zhang, R.; Sun, L. LncRNA H19 functions as an Aquaporin 1 competitive endogenous RNA to regulate microRNA-874 expression in LPS sepsis. Biomed Pharmacother 2018, 105, 1183-1191. [CrossRef]
- Zhou, Y.; Sun, L.; Zhu, M.; Cheng, H. Effects and early diagnostic value of lncRNA H19 on sepsis-induced acute lung injury. Exp Ther Med 2022, 23, 279. [CrossRef]
- Li, H.; Shi, H.; Gao, M.; Ma, N.; Sun, R. Long non-coding RNA CASC2 improved acute lung injury by regulating miR-144-3p/AQP1 axis to reduce lung epithelial cell apoptosis. Cell Biosci 2018, 8, 15. [CrossRef]
- Wang, R.; Zhao, J.; Wei, Q.; Wang, H.; Zhao, C.; Hu, C.; Han, Y.; Hui, Z.; Yang, L.; Dai, Q.; et al. Potential of circulating lncRNA CASC2 as a biomarker in reflecting the inflammatory cytokines, multi-organ dysfunction, disease severity, and mortality in sepsis patients. J Clin Lab Anal 2022, 36, e24569. [CrossRef]
- Adamzik, M.; Frey, U.H.; Bitzer, K.; Jakob, H.; Baba, H.A.; Schmieder, R.E.; Schneider, M.P.; Heusch, G.; Peters, J.; Siffert, W. A novel-1364A/C aquaporin 5 gene promoter polymorphism influences the responses to salt loading of the renin-angiotensin-aldosterone system and of blood pressure in young healthy men. Basic Res Cardiol 2008, 103, 598-610. [CrossRef]
- Adamzik, M.; Frey, U.H.; Möhlenkamp, S.; Scherag, A.; Waydhas, C.; Marggraf, G.; Dammann, M.; Steinmann, J.; Siffert, W.; Peters, J. Aquaporin 5 Gene Promoter −1364A/C Polymorphism Associated with 30-day Survival in Severe Sepsis. Anesthesiology 2011, 114, 912-917. [CrossRef]
- Nomura, J.; Hisatsune, A.; Miyata, T.; Isohama, Y. The role of CpG methylation in cell type-specific expression of the aquaporin-5 gene. Biochem Biophys Res Commun 2007, 353, 1017-1022. [CrossRef]
- Rump, K.; Spellenberg, T.; von Busch, A.; Wolf, A.; Ziehe, D.; Thon, P.; Rahmel, T.; Adamzik, M.; Koos, B.; Unterberg, M. AQP5-1364A/C Polymorphism Affects AQP5 Promoter Methylation. Int J Mol Sci 2022, 23. [CrossRef]
- Zheng, C.; Wu, D.; Shi, S.; Wang, L. miR-34b-5p promotes renal cell inflammation and apoptosis by inhibiting aquaporin-2 in sepsis-induced acute kidney injury. Ren Fail 2021, 43, 291-301. [CrossRef]
- Verkman, A.S. Role of aquaporin water channels in kidney and lung. Am J Med Sci 1998, 316, 310-320. [CrossRef]
- Verkman, A.S. More than just water channels: unexpected cellular roles of aquaporins. Journal of Cell Science 2005, 118, 3225-3232. [CrossRef]
- Verkman, A.S. Role of aquaporins in lung liquid physiology. Respir Physiol Neurobiol 2007, 159, 324-330. [CrossRef]
- Loreto, C.; Reggio, E. Aquaporin and vascular diseases. Curr Neuropharmacol 2010, 8, 105-111. [CrossRef]
- González-Marrero, I.; Hernández-Abad, L.G.; González-Gómez, M.; Soto-Viera, M.; Carmona-Calero, E.M.; Castañeyra-Ruiz, L.; Castañeyra-Perdomo, A. Altered Expression of AQP1 and AQP4 in Brain Barriers and Cerebrospinal Fluid May Affect Cerebral Water Balance during Chronic Hypertension. Int J Mol Sci 2022, 23. [CrossRef]
- Sisto, M.; Ribatti, D.; Lisi, S. Aquaporin water channels: New perspectives on the potential role in inflammation. Adv Protein Chem Struct Biol 2019, 116, 311-345. [CrossRef]
- Kwon, T.H.; Frøkiær, J.; Nielsen, S. Regulation of aquaporin-2 in the kidney: A molecular mechanism of body-water homeostasis. Kidney Res Clin Pract 2013, 32, 96-102. [CrossRef]
- Grinevich, V.; Knepper, M.A.; Verbalis, J.; Reyes, I.; Aguilera, G. Acute endotoxemia in rats induces down-regulation of V2 vasopressin receptors and aquaporin-2 content in the kidney medulla. Kidney Int 2004, 65, 54-62. [CrossRef]
- Wang, W.; Li, C.; Summer, S.N.; Falk, S.; Wang, W.; Ljubanovic, D.; Schrier, R.W. Role of AQP1 in endotoxemia-induced acute kidney injury. Am J Physiol Renal Physiol 2008, 294, F1473-1480. [CrossRef]
- Olesen, E.T.; de Seigneux, S.; Wang, G.; Lutken, S.C.; Frokiaer, J.; Kwon, T.H.; Nielsen, S. Rapid and segmental specific dysregulation of AQP2, S256-pAQP2 and renal sodium transporters in rats with LPS-induced endotoxaemia. Nephrol Dial Transplant 2009, 24, 2338-2349. [CrossRef]
- Rodrigues, C.E.; Sanches, T.R.; Volpini, R.A.; Shimizu, M.H.; Kuriki, P.S.; Camara, N.O.; Seguro, A.C.; Andrade, L. Effects of continuous erythropoietin receptor activator in sepsis-induced acute kidney injury and multi-organ dysfunction. PLoS One 2012, 7, e29893. [CrossRef]
- Ozden, E.S.; Asci, H.; Buyukbayram, H.I.; Sevuk, M.A.; Imeci, O.B.; Dogan, H.K.; Ozmen, O. Dexpanthenol protects against lipopolysaccharide-induced acute kidney injury by restoring aquaporin-2 levels via regulation of the silent information regulator 1 signaling pathway. Korean J Anesthesiol 2023, 76, 501-509. [CrossRef]
- Rump, K.; Adamzik, M. Aquaporins in sepsis- an update. Front Immunol 2024, 15, 1495206. [CrossRef]
- Kwong, R.W.; Kumai, Y.; Perry, S.F. The role of aquaporin and tight junction proteins in the regulation of water movement in larval zebrafish (Danio rerio). PLoS One 2013, 8, e70764. [CrossRef]
- Tang, G.; Yang, G.Y. Aquaporin-4: A Potential Therapeutic Target for Cerebral Edema. Int J Mol Sci 2016, 17. [CrossRef]
- Abo El Gheit, R.E.; Atef, M.M.; Badawi, G.A.; Elwan, W.M.; Alshenawy, H.A.; Emam, M.N. Role of serine protease inhibitor, ulinastatin, in rat model of hepatic encephalopathy: aquaporin 4 molecular targeting and therapeutic implication. J Physiol Biochem 2020, 76, 573-586. [CrossRef]
- Force*, T.A.D.T. Acute Respiratory Distress Syndrome: The Berlin Definition. JAMA 2012, 307, 2526-2533. [CrossRef]
- Bernard, G.R.; Artigas, A.; Brigham, K.L.; Carlet, J.; Falke, K.; Hudson, L.; Lamy, M.; Legall, J.R.; Morris, A.; Spragg, R. The American-European Consensus Conference on ARDS. Definitions, mechanisms, relevant outcomes, and clinical trial coordination. Am J Respir Crit Care Med 1994, 149, 818-824. [CrossRef]
- Huppert, L.A.; Matthay, M.A.; Ware, L.B. Pathogenesis of Acute Respiratory Distress Syndrome. Semin Respir Crit Care Med 2019, 40, 31-39. [CrossRef]
- Vassiliou, A.G.; Kotanidou, A.; Dimopoulou, I.; Orfanos, S.E. Endothelial Damage in Acute Respiratory Distress Syndrome. Int J Mol Sci 2020, 21. [CrossRef]
- Keskinidou, C.; Vassiliou, A.G.; Dimopoulou, I.; Kotanidou, A.; Orfanos, S.E. Mechanistic Understanding of Lung Inflammation: Recent Advances and Emerging Techniques. J Inflamm Res 2022, 15, 3501-3546. [CrossRef]
- Wittekindt, O.H.; Dietl, P. Aquaporins in the lung. Pflugers Arch 2019, 471, 519-532. [CrossRef]
- Matthay, M.A.; Folkesson, H.G.; Clerici, C. Lung epithelial fluid transport and the resolution of pulmonary edema. Physiological reviews 2002, 82, 569-600. [CrossRef]
- Bai, C.; Fukuda, N.; Song, Y.; Ma, T.; Matthay, M.A.; Verkman, A.S. Lung fluid transport in aquaporin-1 and aquaporin-4 knockout mice. J Clin Invest 1999, 103, 555-561. [CrossRef]
- Ma, T.; Fukuda, N.; Song, Y.; Matthay, M.A.; Verkman, A.S. Lung fluid transport in aquaporin-5 knockout mice. J Clin Invest 2000, 105, 93-100. [CrossRef]
- Song, Y.; Fukuda, N.; Bai, C.; Ma, T.; Matthay, M.A.; Verkman, A.S. Role of aquaporins in alveolar fluid clearance in neonatal and adult lung, and in oedema formation following acute lung injury: studies in transgenic aquaporin null mice. J Physiol 2000, 525 Pt 3, 771-779. [CrossRef]
- Verkman, A.S. Knock-out models reveal new aquaporin functions. Handb Exp Pharmacol 2009, 359-381. [CrossRef]
- Meli, R.; Pirozzi, C.; Pelagalli, A. New Perspectives on the Potential Role of Aquaporins (AQPs) in the Physiology of Inflammation. Front Physiol 2018, 9, 101. [CrossRef]
- Lai, K.N.; Leung, J.C.; Metz, C.N.; Lai, F.M.; Bucala, R.; Lan, H.Y. Role for macrophage migration inhibitory factor in acute respiratory distress syndrome. J Pathol 2003, 199, 496-508. [CrossRef]
- Bayraktar, N.; Bayraktar, M.; Ozturk, A.; Ibrahim, B. Evaluation of the Relationship Between Aquaporin-1, Hepcidin, Zinc, Copper, and İron Levels and Oxidative Stress in the Serum of Critically Ill Patients with COVID-19. Biol Trace Elem Res 2022, 200, 5013-5021. [CrossRef]
- Rahmel, T.; Nowak, H.; Rump, K.; Siffert, W.; Peters, J.; Adamzik, M. The aquaporin 5 -1364A/C promoter polymorphism impacts on resolution of acute kidney injury in pneumonia evoked ARDS. PLoS One 2018, 13, e0208582. [CrossRef]
- Xie, Y.; Luo, J.; Hu, W.; Ye, C.; Ren, P.; Wang, Y.; Li, X. Whole Transcriptomic Analysis of Key Genes and Signaling Pathways in Endogenous ARDS. Dis Markers 2022, 2022, 1614208. [CrossRef]
- Liu, F.; Hu, S.; Zhao, N.; Shao, Q.; Li, Y.; Jiang, R.; Chen, J.; Peng, W.; Qian, K. LncRNA-5657 silencing alleviates sepsis-induced lung injury by suppressing the expression of spinster homology protein 2. Int Immunopharmacol 2020, 88, 106875. [CrossRef]
- Zhan, Y.A.; Qiu, X.L.; Wang, X.Z.; Zhao, N.; Qian, K.J. Reducing LncRNA-5657 expression inhibits the brain inflammatory reaction in septic rats. Neural Regen Res 2021, 16, 1288-1293. [CrossRef]
- Zhu, D.D.; Huang, Y.L.; Guo, S.Y.; Li, N.; Yang, X.W.; Sui, A.R.; Wu, Q.; Zhang, Y.; Kong, Y.; Li, Q.F.; et al. AQP4 Aggravates Cognitive Impairment in Sepsis-Associated Encephalopathy through Inhibiting Na(v) 1.6-Mediated Astrocyte Autophagy. Adv Sci (Weinh) 2023, 10, e2205862. [CrossRef]
- Tang, R.; Pei, L.; Bai, T.; Wang, J. Down-regulation of microRNA-126-5p contributes to overexpression of VEGFA in lipopolysaccharide-induced acute lung injury. Biotechnol Lett 2016, 38, 1277-1284. [CrossRef]
- Mao, X.; Wu, Y.; Xu, W. miR-126-5p expression in the plasma of patients with sepsis-induced acute lung injury and its correlation with inflammation and immune function. Clin Respir J 2023, 17, 629-637. [CrossRef]
- Li, B.; Chen, D.; Wang, G.; Dong, C.; Wang, X.; Bai, C. Expression of AQP-1, AQP-3, AQP-4 and AQP-5 in pulmonary tissues of mice with endotoxin-induced acute lung injury. Academic Journal of Second Military Medical University 2008, 29, 131-135.
- Rump, K.; Brendt, P.; Frey, U.H.; Schäfer, S.T.; Siffert, W.; Peters, J.; Adamzik, M. Aquaporin 1 and 5 expression evoked by the β2 adrenoreceptor agonist terbutaline and lipopolysaccharide in mice and in the human monocytic cell line THP-1 is differentially regulated. Shock 2013, 40, 430-436. [CrossRef]
- Hasan, B.; Li, F.-s.; Siyit, A.; Tuyghun, E.; Luo, J.-h.; Upur, H.; Ablimit, A. Expression of aquaporins in the lungs of mice with acute injury caused by LPS treatment. Respiratory Physiology & Neurobiology 2014, 200, 40-45.
- Hong-Min, F.; Chun-Rong, H.; Rui, Z.; Li-Na, S.; Ya-Jun, W.; Li, L. CGRP 8-37 enhances lipopolysaccharide-induced acute lung injury and regulating aquaporin 1 and 5 expressions in rats. J Physiol Biochem 2016, 73, 381-386. [CrossRef]
- Tao, B.; Liu, L.; Wang, N.; Wang, W.; Jiang, J.; Zhang, J. Effects of hydrogen-rich saline on aquaporin 1, 5 in septic rat lungs. J Surg Res 2016, 202, 291-298. [CrossRef]
- Li, B.; Liu, C.; Tang, K.; Dong, X.; Xue, L.; Su, G.; Zhang, W.; Jin, Y. Aquaporin-1 attenuates macrophage-mediated inflammatory responses by inhibiting p38 mitogen-activated protein kinase activation in lipopolysaccharide-induced acute kidney injury. Inflamm Res 2019, 68, 1035-1047. [CrossRef]
- Wang, X.; Zhou, X.; Xia, X.; Zhang, Y. Estradiol attenuates LPS-induced acute lung injury via induction of aquaporins AQP1 and AQP5. European Journal of Inflammation 2021, 19. [CrossRef]
- Wang, X.; Zhang, Y.; Zhou, X.; Xia, X.; Teng, W.; Sheng, L.; Ding, J. Soy isoflavone reduces LPS-induced acute lung injury via increasing aquaporin 1 and aquaporin 5 in rats. Open Life Sci 2023, 18, 20220560. [CrossRef]
- Nieman, G.F.; Gatto, L.A.; Andrews, P.; Satalin, J.; Camporota, L.; Daxon, B.; Blair, S.J.; Al-Khalisy, H.; Madden, M.; Kollisch-Singule, M.; et al. Prevention and treatment of acute lung injury with time-controlled adaptive ventilation: physiologically informed modification of airway pressure release ventilation. Ann Intensive Care 2020, 10, 3. [CrossRef]
- Yamashita, A.; Ito, Y.; Osada, M.; Matsuda, H.; Hosono, K.; Tsujikawa, K.; Okamoto, H.; Amano, H. RAMP1 Signaling Mitigates Acute Lung Injury by Distinctively Regulating Alveolar and Monocyte-Derived Macrophages. Int J Mol Sci 2024, 25. [CrossRef]
- Zhang, Y.; Chen, M.; Zhang, Y.; Peng, P.; Li, J.; Xin, X. miR-96 and miR-330 overexpressed and targeted AQP5 in lipopolysaccharide-induced rat lung damage of disseminated intravascular coagulation. Blood Coagul Fibrinolysis 2014, 25, 731-737. [CrossRef]
- Xu, J.; Yang, L.; Dong, L. Tanshinol upregulates the expression of aquaporin 5 in lung tissue of rats with sepsis. Oncol Lett 2018, 16, 3290-3296. [CrossRef]
- Ba, F.; Zhou, X.; Zhang, Y.; Wu, C.; Xu, S.; Wu, L.; Li, J.; Yin, Y.; Gu, X. Lipoxin A4 ameliorates alveolar fluid clearance disturbance in lipopolysaccharide-induced lung injury via aquaporin 5 and MAPK signaling pathway. J Thorac Dis 2019, 11, 3599-3608. [CrossRef]
- Wang, J.J.; Kong, H.; Xu, J.; Wang, Y.L.; Wang, H.; Xie, W.P. Fasudil alleviates LPS-induced lung injury by restoring aquaporin 5 expression and inhibiting inflammation in lungs. J Biomed Res 2019, 33, 156-163. [CrossRef]
- Villandre, J.; White, V.; Lear, T.B.; Chen, Y.; Tuncer, F.; Vaiz, E.; Tuncer, B.; Lockwood, K.; Camarco, D.; Liu, Y.; et al. A Repurposed Drug Screen for Compounds Regulating Aquaporin 5 Stability in Lung Epithelial Cells. Front Pharmacol 2022, 13, 828643. [CrossRef]
- Hoste, E.A.J.; Kellum, J.A.; Selby, N.M.; Zarbock, A.; Palevsky, P.M.; Bagshaw, S.M.; Goldstein, S.L.; Cerdá, J.; Chawla, L.S. Global epidemiology and outcomes of acute kidney injury. Nat Rev Nephrol 2018, 14, 607-625. [CrossRef]
- James, M.T.; Bhatt, M.; Pannu, N.; Tonelli, M. Long-term outcomes of acute kidney injury and strategies for improved care. Nat Rev Nephrol 2020, 16, 193-205. [CrossRef]
- Pickkers, P.; Darmon, M.; Hoste, E.; Joannidis, M.; Legrand, M.; Ostermann, M.; Prowle, J.R.; Schneider, A.; Schetz, M. Acute kidney injury in the critically ill: an updated review on pathophysiology and management. Intensive Care Med 2021, 47, 835-850. [CrossRef]
- Chou, C.L.; Knepper, M.A.; Hoek, A.N.; Brown, D.; Yang, B.; Ma, T.; Verkman, A.S. Reduced water permeability and altered ultrastructure in thin descending limb of Henle in aquaporin-1 null mice. J Clin Invest 1999, 103, 491-496. [CrossRef]
- Yang, B.; Ma, T.; Verkman, A.S. Erythrocyte water permeability and renal function in double knockout mice lacking aquaporin-1 and aquaporin-3. J Biol Chem 2001, 276, 624-628. [CrossRef]
- Verkman, A.S.; Yang, B. Aquaporin gene delivery to kidney. Kidney Int 2002, 61, S120-124. [CrossRef]
- Ren, H.; Yang, B.; Molina, P.A.; Sands, J.M.; Klein, J.D. NSAIDs Alter Phosphorylated Forms of AQP2 in the Inner Medullary Tip. PLoS One 2015, 10, e0141714. [CrossRef]
- Lei, L.; Huang, M.; Su, L.; Xie, D.; Mamuya, F.A.; Ham, O.; Tsuji, K.; Paunescu, T.G.; Yang, B.; Lu, H.A.J. Manganese promotes intracellular accumulation of AQP2 via modulating F-actin polymerization and reduces urinary concentration in mice. Am J Physiol Renal Physiol 2018, 314, F306-F316. [CrossRef]
- Ren, H.; Yang, B.; Ruiz, J.A.; Efe, O.; Ilori, T.O.; Sands, J.M.; Klein, J.D. Phosphatase inhibition increases AQP2 accumulation in the rat IMCD apical plasma membrane. Am J Physiol Renal Physiol 2016, 311, F1189-F1197. [CrossRef]
- Kavanagh, C.; Uy, N.S. Nephrogenic Diabetes Insipidus. Pediatr Clin North Am 2019, 66, 227-234. [CrossRef]
- Hara-Chikuma, M.; Satooka, H.; Watanabe, S.; Honda, T.; Miyachi, Y.; Watanabe, T.; Verkman, A.S. Aquaporin-3-mediated hydrogen peroxide transport is required for NF-κB signalling in keratinocytes and development of psoriasis. Nat Commun 2015, 6, 7454. [CrossRef]
- Rodríguez, A.; Catalán, V.; Gómez-Ambrosi, J.; Frühbeck, G. Aquaglyceroporins serve as metabolic gateways in adiposity and insulin resistance control. Cell Cycle 2011, 10, 1548-1556. [CrossRef]
- Medraño-Fernandez, I.; Bestetti, S.; Bertolotti, M.; Bienert, G.P.; Bottino, C.; Laforenza, U.; Rubartelli, A.; Sitia, R. Stress Regulates Aquaporin-8 Permeability to Impact Cell Growth and Survival. Antioxid Redox Signal 2016, 24, 1031-1044. [CrossRef]
- Tanaka, Y.; Watari, M.; Saito, T.; Morishita, Y.; Ishibashi, K. Enhanced Autophagy in Polycystic Kidneys of AQP11 Null Mice. Int J Mol Sci 2016, 17. [CrossRef]
- Ishikawa, S.; Saito, T.; Fukagawa, A.; Higashiyama, M.; Nakamura, T.; Kusaka, I.; Nagasaka, S.; Honda, K.; Saito, T. Close association of urinary excretion of aquaporin-2 with appropriate and inappropriate arginine vasopressin-dependent antidiuresis in hyponatremia in elderly subjects. J Clin Endocrinol Metab 2001, 86, 1665-1671. [CrossRef]
- Ivarsen, P.; Frokiaer, J.; Aagaard, N.K.; Hansen, E.F.; Bendtsen, F.; Nielsen, S.; Vilstrup, H. Increased urinary excretion of aquaporin 2 in patients with liver cirrhosis. Gut 2003, 52, 1194-1199. [CrossRef]
- Buemi, M.; D’Anna, R.; Di Pasquale, G.; Floccari, F.; Ruello, A.; Aloisi, C.; Leonardi, I.; Frisina, N.; Corica, F. Urinary excretion of aquaporin-2 water channel during pregnancy. Cell Physiol Biochem 2001, 11, 203-208. [CrossRef]
- Rossi, L.; Nicoletti, M.C.; Carmosino, M.; Mastrofrancesco, L.; Di Franco, A.; Indrio, F.; Lella, R.; Laviola, L.; Giorgino, F.; Svelto, M.; et al. Urinary Excretion of Kidney Aquaporins as Possible Diagnostic Biomarker of Diabetic Nephropathy. J Diabetes Res 2017, 2017, 4360357. [CrossRef]
- Bräsen, J.H.; Mederacke, Y.S.; Schmitz, J.; Diahovets, K.; Khalifa, A.; Hartleben, B.; Person, F.; Wiech, T.; Steenbergen, E.; Großhennig, A.; et al. Cholemic Nephropathy Causes Acute Kidney Injury and Is Accompanied by Loss of Aquaporin 2 in Collecting Ducts. Hepatology 2019, 69, 2107-2119. [CrossRef]
- Chan, M.J.; Chen, Y.C.; Fan, P.C.; Lee, C.C.; Kou, G.; Chang, C.H. Predictive Value of Urinary Aquaporin 2 for Acute Kidney Injury in Patients with Acute Decompensated Heart Failure. Biomedicines 2022, 10. [CrossRef]
- Suh, S.H.; Lee, K.E.; Kim, I.J.; Kim, O.; Kim, C.S.; Choi, J.S.; Choi, H.I.; Bae, E.H.; Ma, S.K.; Lee, J.U.; et al. Alpha-lipoic acid attenuates lipopolysaccharide-induced kidney injury. Clin Exp Nephrol 2015, 19, 82-91. [CrossRef]
- Höcherl, K.; Schmidt, C.; Kurt, B.; Bucher, M. Inhibition of NF-kappaB ameliorates sepsis-induced downregulation of aquaporin-2/V2 receptor expression and acute renal failure in vivo. Am J Physiol Renal Physiol 2010, 298, F196-204. [CrossRef]
- Lv, W.; Xue, L.; Liang, L.; Liu, D.; Li, C.; Liao, J.; Jin, Y. Endotoxin induced acute kidney injury modulates expression of AQP1, P53 and P21 in rat kidney, heart, lung and small intestine. PLoS One 2023, 18, e0288507. [CrossRef]
- Tod, P.; Róka, B.; Kaucsár, T.; Szatmári, K.; Vizovišek, M.; Vidmar, R.; Fonovič, M.; Szénási, G.; Hamar, P. Time-Dependent miRNA Profile during Septic Acute Kidney Injury in Mice. Int J Mol Sci 2020, 21. [CrossRef]
- Liu, C.; Li, B.; Tang, K.; Dong, X.; Xue, L.; Su, G.; Jin, Y. Aquaporin 1 alleviates acute kidney injury via PI3K-mediated macrophage M2 polarization. Inflamm Res 2020, 69, 509-521. [CrossRef]
- Lv, W.; Liao, J.; Li, C.; Liu, D.; Luo, X.; Diao, R.; Wang, Y.; Jin, Y. Aquaporin 1 is renoprotective in septic acute kidney injury by attenuating inflammation, apoptosis and fibrosis through inhibition of P53 expression. Front Immunol 2024, 15, 1443108. [CrossRef]
- Basile, D.P.; Anderson, M.D.; Sutton, T.A. Pathophysiology of acute kidney injury. Compr Physiol 2012, 2, 1303-1353. [CrossRef]
- Asvapromtada, S.; Sonoda, H.; Kinouchi, M.; Oshikawa, S.; Takahashi, S.; Hoshino, Y.; Sinlapadeelerdkul, T.; Yokota-Ikeda, N.; Matsuzaki, T.; Ikeda, M. Characterization of urinary exosomal release of aquaporin-1 and -2 after renal ischemia-reperfusion in rats. Am J Physiol Renal Physiol 2018, 314, F584-f601. [CrossRef]
- Gong, H.; Wang, W.; Kwon, T.H.; Jonassen, T.; Li, C.; Ring, T.; Froki, A.J.; Nielsen, S. EPO and alpha-MSH prevent ischemia/reperfusion-induced down-regulation of AQPs and sodium transporters in rat kidney. Kidney Int 2004, 66, 683-695. [CrossRef]
- Vukićević, T.; Schulz, M.; Faust, D.; Klussmann, E. The Trafficking of the Water Channel Aquaporin-2 in Renal Principal Cells-a Potential Target for Pharmacological Intervention in Cardiovascular Diseases. Front Pharmacol 2016, 7, 23. [CrossRef]
- Soveral, G.; Casini, A. Aquaporin modulators: a patent review (2010-2015). Expert Opin Ther Pat 2017, 27, 49-62. [CrossRef]
- Abdeen, A.; Sonoda, H.; El-Shawarby, R.; Takahashi, S.; Ikeda, M. Urinary excretion pattern of exosomal aquaporin-2 in rats that received gentamicin. Am J Physiol Renal Physiol 2014, 307, F1227-1237. [CrossRef]
- Wu, H.; Chen, L.; Zhang, X.; Zhou, Q.; Li, J.M.; Berger, S.; Borok, Z.; Zhou, B.; Xiao, Z.; Yin, H.; et al. Aqp5 is a new transcriptional target of Dot1a and a regulator of Aqp2. PLoS One 2013, 8, e53342. [CrossRef]
- Lu, Y.; Chen, L.; Zhao, B.; Xiao, Z.; Meng, T.; Zhou, Q.; Zhang, W. Urine AQP5 is a potential novel biomarker of diabetic nephropathy. J Diabetes Complications 2016, 30, 819-825. [CrossRef]
- Su, W.; Cao, R.; Zhang, X.Y.; Guan, Y. Aquaporins in the kidney: physiology and pathophysiology. Am J Physiol Renal Physiol 2020, 318, F193-f203. [CrossRef]
- Newcombe, V.; Muehlschlegel, S.; Sonneville, R. Neurological diseases in intensive care. Intensive Care Med 2023, 49, 987-990. [CrossRef]
- Robba, C.; Zanier, E.R.; Lopez Soto, C.; Park, S.; Sonneville, R.; Helbolk, R.; Sarwal, A.; Newcombe, V.F.J.; van der Jagt, M.; Gunst, J.; et al. Mastering the brain in critical conditions: an update. Intensive Care Med Exp 2024, 12, 1. [CrossRef]
- Papadopoulos, M.C.; Krishna, S.; Verkman, A.S. Aquaporin water channels and brain edema. Mt Sinai J Med 2002, 69, 242-248.
- Deng, S.; Chen, X.; Lei, Q.; Lu, W. AQP2 Promotes Astrocyte Activation by Modulating the TLR4/NFκB-p65 Pathway Following Intracerebral Hemorrhage. Front Immunol 2022, 13, 847360. [CrossRef]
- Papadopoulos, M.C.; Verkman, A.S. Aquaporin-4 and brain edema. Pediatr Nephrol 2007, 22, 778-784. [CrossRef]
- Szu, J.I.; Binder, D.K. The Role of Astrocytic Aquaporin-4 in Synaptic Plasticity and Learning and Memory. Front Integr Neurosci 2016, 10, 8. [CrossRef]
- Manley, G.T.; Fujimura, M.; Ma, T.; Noshita, N.; Filiz, F.; Bollen, A.W.; Chan, P.; Verkman, A.S. Aquaporin-4 deletion in mice reduces brain edema after acute water intoxication and ischemic stroke. Nat Med 2000, 6, 159-163. [CrossRef]
- Manley, G.T.; Binder, D.K.; Papadopoulos, M.C.; Verkman, A.S. New insights into water transport and edema in the central nervous system from phenotype analysis of aquaporin-4 null mice. Neuroscience 2004, 129, 983-991. [CrossRef]
- Yao, X.; Uchida, K.; Papadopoulos, M.C.; Zador, Z.; Manley, G.T.; Verkman, A.S. Mildly Reduced Brain Swelling and Improved Neurological Outcome in Aquaporin-4 Knockout Mice following Controlled Cortical Impact Brain Injury. J Neurotrauma 2015, 32, 1458-1464. [CrossRef]
- Papadopoulos, M.C.; Manley, G.T.; Krishna, S.; Verkman, A.S. Aquaporin-4 facilitates reabsorption of excess fluid in vasogenic brain edema. Faseb j 2004, 18, 1291-1293. [CrossRef]
- Papadopoulos, M.C.; Saadoun, S.; Binder, D.K.; Manley, G.T.; Krishna, S.; Verkman, A.S. Molecular mechanisms of brain tumor edema. Neuroscience 2004, 129, 1011-1020. [CrossRef]
- Mori, S.; Kurimoto, T.; Miki, A.; Maeda, H.; Kusuhara, S.; Nakamura, M. Aqp9 Gene Deletion Enhances Retinal Ganglion Cell (RGC) Death and Dysfunction Induced by Optic Nerve Crush: Evidence that Aquaporin 9 Acts as an Astrocyte-to-Neuron Lactate Shuttle in Concert with Monocarboxylate Transporters To Support RGC Function and Survival. Mol Neurobiol 2020, 57, 4530-4548. [CrossRef]
- Dardiotis, E.; Paterakis, K.; Tsivgoulis, G.; Tsintou, M.; Hadjigeorgiou, G.F.; Dardioti, M.; Grigoriadis, S.; Simeonidou, C.; Komnos, A.; Kapsalaki, E.; et al. AQP4 tag single nucleotide polymorphisms in patients with traumatic brain injury. J Neurotrauma 2014, 31, 1920-1926. [CrossRef]
- Nekludov, M.; Bellander, B.M.; Gryth, D.; Wallen, H.; Mobarrez, F. Brain-Derived Microparticles in Patients with Severe Isolated TBI. Brain Inj 2017, 31, 1856-1862. [CrossRef]
- Lo Pizzo, M.; Schiera, G.; Di Liegro, I.; Di Liegro, C.M.; Pál, J.; Czeiter, E.; Sulyok, E.; Dóczi, T. Aquaporin-4 distribution in control and stressed astrocytes in culture and in the cerebrospinal fluid of patients with traumatic brain injuries. Neurol Sci 2013, 34, 1309-1314. [CrossRef]
- He, Y.; Yang, Q.; Liu, H.; Jiang, L.; Liu, Q.; Lian, W.; Huang, J. Effect of blood pressure on early neurological deterioration of acute ischemic stroke patients with intravenous rt-PA thrombolysis may be mediated through oxidative stress induced blood-brain barrier disruption and AQP4 upregulation. J Stroke Cerebrovasc Dis 2020, 29, 104997. [CrossRef]
- Czyżewski, W.; Korulczyk, J.; Szymoniuk, M.; Sakwa, L.; Litak, J.; Ziemianek, D.; Czyżewska, E.; Mazurek, M.; Kowalczyk, M.; Turek, G.; et al. Aquaporin 2 in Cerebral Edema: Potential Prognostic Marker in Craniocerebral Injuries. Int J Mol Sci 2024, 25. [CrossRef]
- Yang, Z.H.; Yin, X.J.; Fu, G.Y. The correlation between CT findings of diffuse axonal injury and the expression of neuronal aquaporin in patients with craniocerebral injury. Eur Rev Med Pharmacol Sci 2022, 26, 6871-6878. [CrossRef]
- Hu, H.; Yao, H.T.; Zhang, W.P.; Zhang, L.; Ding, W.; Zhang, S.H.; Chen, Z.; Wei, E.Q. Increased expression of aquaporin-4 in human traumatic brain injury and brain tumors. J Zhejiang Univ Sci B 2005, 6, 33-37. [CrossRef]
- Braun, M.; Sevao, M.; Keil, S.A.; Gino, E.; Wang, M.X.; Lee, J.; Haveliwala, M.A.; Klein, E.; Agarwal, S.; Pedersen, T.; et al. Macroscopic changes in aquaporin-4 underlie blast traumatic brain injury-related impairment in glymphatic function. Brain 2024, 147, 2214-2229. [CrossRef]
- Suzuki, R.; Okuda, M.; Asai, J.; Nagashima, G.; Itokawa, H.; Matsunaga, A.; Fujimoto, T.; Suzuki, T. Astrocytes co-express aquaporin-1, -4, and vascular endothelial growth factor in brain edema tissue associated with brain contusion. Acta Neurochir Suppl 2006, 96, 398-401. [CrossRef]
- Aoki, K.; Uchihara, T.; Tsuchiya, K.; Nakamura, A.; Ikeda, K.; Wakayama, Y. Enhanced expression of aquaporin 4 in human brain with infarction. Acta Neuropathol 2003, 106, 121-124. [CrossRef]
- Stokum, J.A.; Mehta, R.I.; Ivanova, S.; Yu, E.; Gerzanich, V.; Simard, J.M. Heterogeneity of aquaporin-4 localization and expression after focal cerebral ischemia underlies differences in white versus grey matter swelling. Acta Neuropathol Commun 2015, 3, 61. [CrossRef]
- Wu, H.; Zhao, R.; Qi, J.; Cong, Y.; Wang, D.; Liu, T.; Gu, Y.; Ban, X.; Huang, Q. The expression and the role of protease nexin-1 on brain edema after intracerebral hemorrhage. J Neurol Sci 2008, 270, 172-183. [CrossRef]
- Reeves, C.; Pradim-Jardim, A.; Sisodiya, S.M.; Thom, M.; Liu, J.Y.W. Spatiotemporal dynamics of PDGFRβ expression in pericytes and glial scar formation in penetrating brain injuries in adults. Neuropathol Appl Neurobiol 2019, 45, 609-627. [CrossRef]
- Satoh, J.; Tabunoki, H.; Yamamura, T.; Arima, K.; Konno, H. Human astrocytes express aquaporin-1 and aquaporin-4 in vitro and in vivo. Neuropathology 2007, 27, 245-256. [CrossRef]
- Roşu, G.C.; Pirici, I.; Istrate-Ofiţeru, A.M.; Iovan, L.; Tudorică, V.; Mogoantă, L.; Gîlceavă, I.C.; Pirici, D. Expression patterns of aquaporins 1 and 4 in stroke. Rom J Morphol Embryol 2019, 60, 823-830.
- Luo, C.; Yao, X.; Li, J.; He, B.; Liu, Q.; Ren, H.; Liang, F.; Li, M.; Lin, H.; Peng, J.; et al. Paravascular pathways contribute to vasculitis and neuroinflammation after subarachnoid hemorrhage independently of glymphatic control. Cell Death Dis 2016, 7, e2160. [CrossRef]
- Badaut, J.; Brunet, J.F.; Grollimund, L.; Hamou, M.F.; Magistretti, P.J.; Villemure, J.G.; Regli, L. Aquaporin 1 and aquaporin 4 expression in human brain after subarachnoid hemorrhage and in peritumoral tissue. Acta Neurochir Suppl 2003, 86, 495-498. [CrossRef]
- Xi, T.; Jin, F.; Zhu, Y.; Wang, J.; Tang, L.; Wang, Y.; Liebeskind, D.S.; Scalzo, F.; He, Z. miR-27a-3p protects against blood-brain barrier disruption and brain injury after intracerebral hemorrhage by targeting endothelial aquaporin-11. J Biol Chem 2018, 293, 20041-20050. [CrossRef]
- Davies, D.C. Blood-brain barrier breakdown in septic encephalopathy and brain tumours. J Anat 2002, 200, 639-646. [CrossRef]
- Tong, D.M.; Zhou, Y.T.; Wang, G.S.; Chen, X.D.; Yang, T.H. Early prediction and outcome of septic encephalopathy in acute stroke patients with nosocomial coma. J Clin Med Res 2015, 7, 534-539. [CrossRef]
- Tauber, S.C.; Eiffert, H.; Brück, W.; Nau, R. Septic encephalopathy and septic encephalitis. Expert Rev Anti Infect Ther 2017, 15, 121-132. [CrossRef]
- Laird, M.D.; Shields, J.S.; Sukumari-Ramesh, S.; Kimbler, D.E.; Fessler, R.D.; Shakir, B.; Youssef, P.; Yanasak, N.; Vender, J.R.; Dhandapani, K.M. High mobility group box protein-1 promotes cerebral edema after traumatic brain injury via activation of toll-like receptor 4. Glia 2014, 62, 26-38. [CrossRef]
- Tu, Q.; Yan, L.; Wang, C.; Han, A.; Qin, Y.; Cui, L.; Xiang, Q. Associations between Aquaglyceroporin Gene Polymorphisms and Risk of Stroke among Patients with Hypertension. Biomed Res Int 2020, 2020, 9358290. [CrossRef]
- Saadoun, S.; Papadopoulos, M.C.; Krishna, S. Water transport becomes uncoupled from K+ siphoning in brain contusion, bacterial meningitis, and brain tumours: immunohistochemical case review. J Clin Pathol 2003, 56, 972-975. [CrossRef]
- Kleffner, I.; Bungeroth, M.; Schiffbauer, H.; Schäbitz, W.R.; Ringelstein, E.B.; Kuhlenbäumer, G. The role of aquaporin-4 polymorphisms in the development of brain edema after middle cerebral artery occlusion. Stroke 2008, 39, 1333-1335. [CrossRef]
- Appelboom, G.; Bruce, S.; Duren, A.; Piazza, M.; Monahan, A.; Christophe, B.; Zoller, S.; LoPresti, M.; Connolly, E.S. Aquaporin-4 gene variant independently associated with oedema after intracerebral haemorrhage. Neurol Res 2015, 37, 657-661. [CrossRef]
- Dardiotis, E.; Siokas, V.; Marogianni, C.; Aloizou, A.M.; Sokratous, M.; Paterakis, K.; Dardioti, M.; Grigoriadis, S.; Brotis, A.; Kapsalaki, E.; et al. AQP4 tag SNPs in patients with intracerebral hemorrhage in Greek and Polish population. Neurosci Lett 2019, 696, 156-161. [CrossRef]
- Farr, G.W.; Hall, C.H.; Farr, S.M.; Wade, R.; Detzel, J.M.; Adams, A.G.; Buch, J.M.; Beahm, D.L.; Flask, C.A.; Xu, K.; et al. Functionalized Phenylbenzamides Inhibit Aquaporin-4 Reducing Cerebral Edema and Improving Outcome in Two Models of CNS Injury. Neuroscience 2019, 404, 484-498. [CrossRef]
- Liu, M.; Li, H.; Fan, L.; Yan, W.; Yan, Y.F. Treatment Effects of Acetazolamide on Ischemic Stroke: A Meta-Analysis and Systematic Review. World Neurosurg 2024, 185, e750-e757. [CrossRef]
- Zheng, L.; Cheng, W.; Wang, X.; Yang, Z.; Zhou, X.; Pan, C. Overexpression of MicroRNA-145 Ameliorates Astrocyte Injury by Targeting Aquaporin 4 in Cerebral Ischemic Stroke. Biomed Res Int 2017, 2017, 9530951. [CrossRef]
- Kuwahara-Otani, S.; Maeda, S.; Tanaka, K.; Hayakawa, T.; Seki, M. Systemic administration of lipopolysaccharide increases the expression of aquaporin-4 in the rat anterior pituitary gland. J Vet Med Sci 2013, 75, 1081-1084. [CrossRef]
- Masterman, E.; Ahmed, Z. Experimental Treatments for Oedema in Spinal Cord Injury: A Systematic Review and Meta-Analysis. Cells 2021, 10. [CrossRef]
- Du, Y.; Meng, Y.; Lv, X.; Guo, L.; Wang, X.; Su, Z.; Li, L.; Li, N.; Zhao, S.; Zhao, L.; et al. Dexamethasone attenuates LPS-induced changes in expression of urea transporter and aquaporin proteins, ameliorating brain endotoxemia in mice. Int J Clin Exp Pathol 2014, 7, 8443-8452.
- Mogoanta, L.; Ciurea, M.; Pirici, I.; Margaritescu, C.; Simionescu, C.; Ion, D.A.; Pirici, D. Different dynamics of aquaporin 4 and glutamate transporter-1 distribution in the perineuronal and perivascular compartments during ischemic stroke. Brain Pathol 2014, 24, 475-493. [CrossRef]
- Ramiro, L.; Simats, A.; Penalba, A.; Garcia-Tornel, A.; Rovira, A.; Mancha, F.; Bustamante, A.; Montaner, J. Circulating Aquaporin-4 as A biomarker of early neurological improvement in stroke patients: A pilot study. Neurosci Lett 2020, 714, 134580. [CrossRef]
- Sfera, A.; Price, A.I.; Gradini, R.; Cummings, M.; Osorio, C. Proteomic and epigenomic markers of sepsis-induced delirium (SID). Front Mol Biosci 2015, 2, 59. [CrossRef]
Figure 1.
a) side monomer view parallel to the membrane (the two NPA motifs are shown in yellow), b) top view of the AQP tetramer from the extracellular side (water molecules in the pore are shown as red spheres), c) the AQP monomer with the six transmembrane alpha-helices (1–6) connected by five loops extending into the extracellular and cytoplasmic spaces (A-E), and d) illustration of the hourglass-shaped channel, depicting the two selectivity filters (ar/R and NPA), allowing the passage of water molecules and other solutes, as shown, while excluding ions and larger solutes. AQP, aquaporin; ar/R, aromatic/arginine; NPA, asparagine-proline-alanine. Image adapted from [33,71].
Figure 1.
a) side monomer view parallel to the membrane (the two NPA motifs are shown in yellow), b) top view of the AQP tetramer from the extracellular side (water molecules in the pore are shown as red spheres), c) the AQP monomer with the six transmembrane alpha-helices (1–6) connected by five loops extending into the extracellular and cytoplasmic spaces (A-E), and d) illustration of the hourglass-shaped channel, depicting the two selectivity filters (ar/R and NPA), allowing the passage of water molecules and other solutes, as shown, while excluding ions and larger solutes. AQP, aquaporin; ar/R, aromatic/arginine; NPA, asparagine-proline-alanine. Image adapted from [33,71].
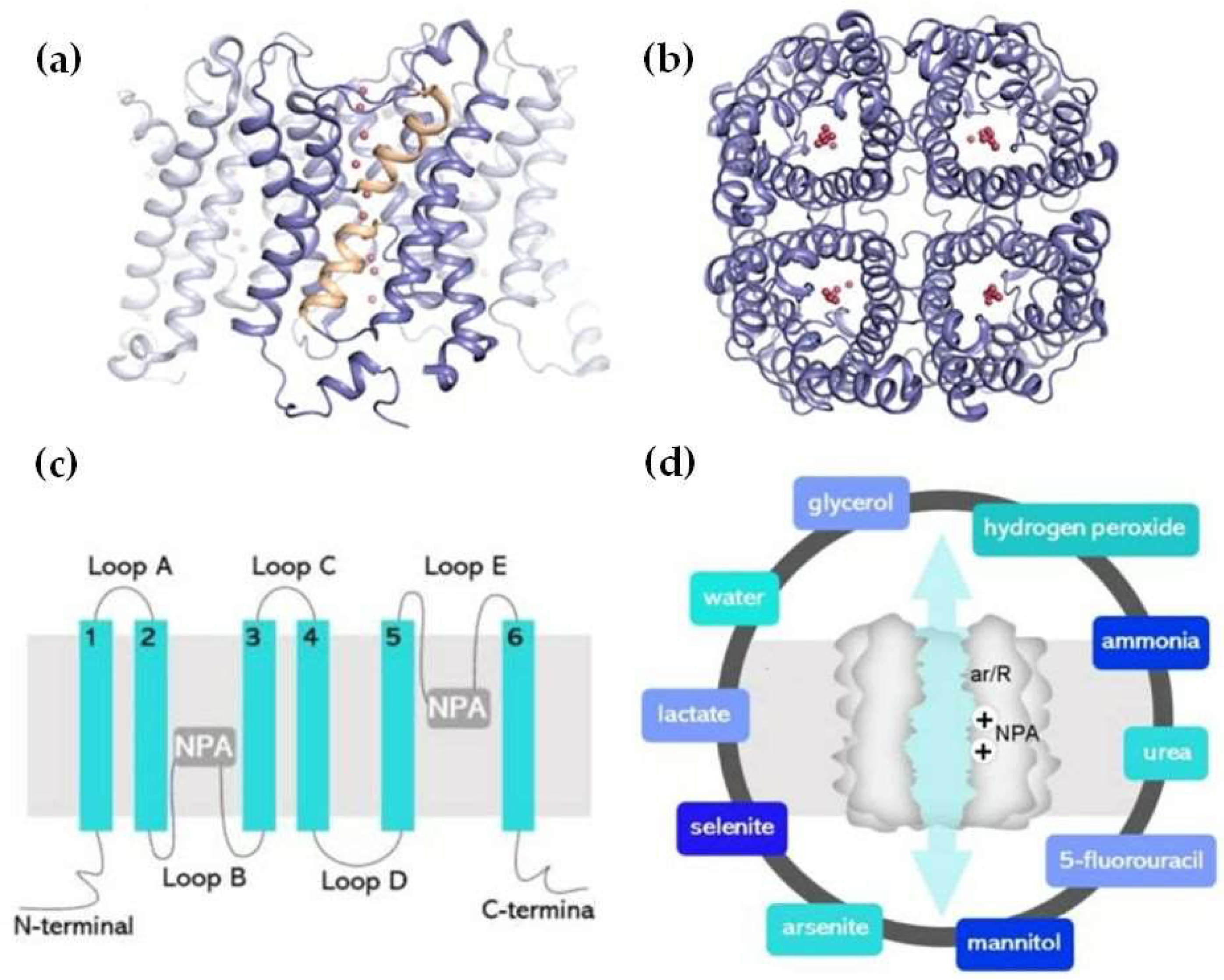
Figure 2.
Aquaporin expression in human immune cells. AQP1 is expressed in neutrophils, monocytes, and lymphocytes, contributing to water transport and cell volume regulation during migration and differentiation. AQP3 is expressed in macrophages, lymphocytes, and dendritic cells, facilitating chemokine-driven migration and supporting energy metabolism through glycerol transport. AQP5 is expressed in some immune cells, particularly lymphocytes, neutrophils, and dendritic cells. AQP5 plays a role in facilitating water transport, which is critical for maintaining cell shape and motility during migration. AQP9 is most prominently expressed in leukocytes, aiding in respiratory bursts and cell motility during pathogen defense. It is expressed in neutrophils, macrophages, monocytes, lymphocytes, and dendritic cells. Other AQPs detected at low concentrations in immune cells are AQP6, AQP7, AQP8, AQP11, and AQP12, expressed in monocytes, macrophages, and dendritic cells. Image adapted from [105].
Figure 2.
Aquaporin expression in human immune cells. AQP1 is expressed in neutrophils, monocytes, and lymphocytes, contributing to water transport and cell volume regulation during migration and differentiation. AQP3 is expressed in macrophages, lymphocytes, and dendritic cells, facilitating chemokine-driven migration and supporting energy metabolism through glycerol transport. AQP5 is expressed in some immune cells, particularly lymphocytes, neutrophils, and dendritic cells. AQP5 plays a role in facilitating water transport, which is critical for maintaining cell shape and motility during migration. AQP9 is most prominently expressed in leukocytes, aiding in respiratory bursts and cell motility during pathogen defense. It is expressed in neutrophils, macrophages, monocytes, lymphocytes, and dendritic cells. Other AQPs detected at low concentrations in immune cells are AQP6, AQP7, AQP8, AQP11, and AQP12, expressed in monocytes, macrophages, and dendritic cells. Image adapted from [105].
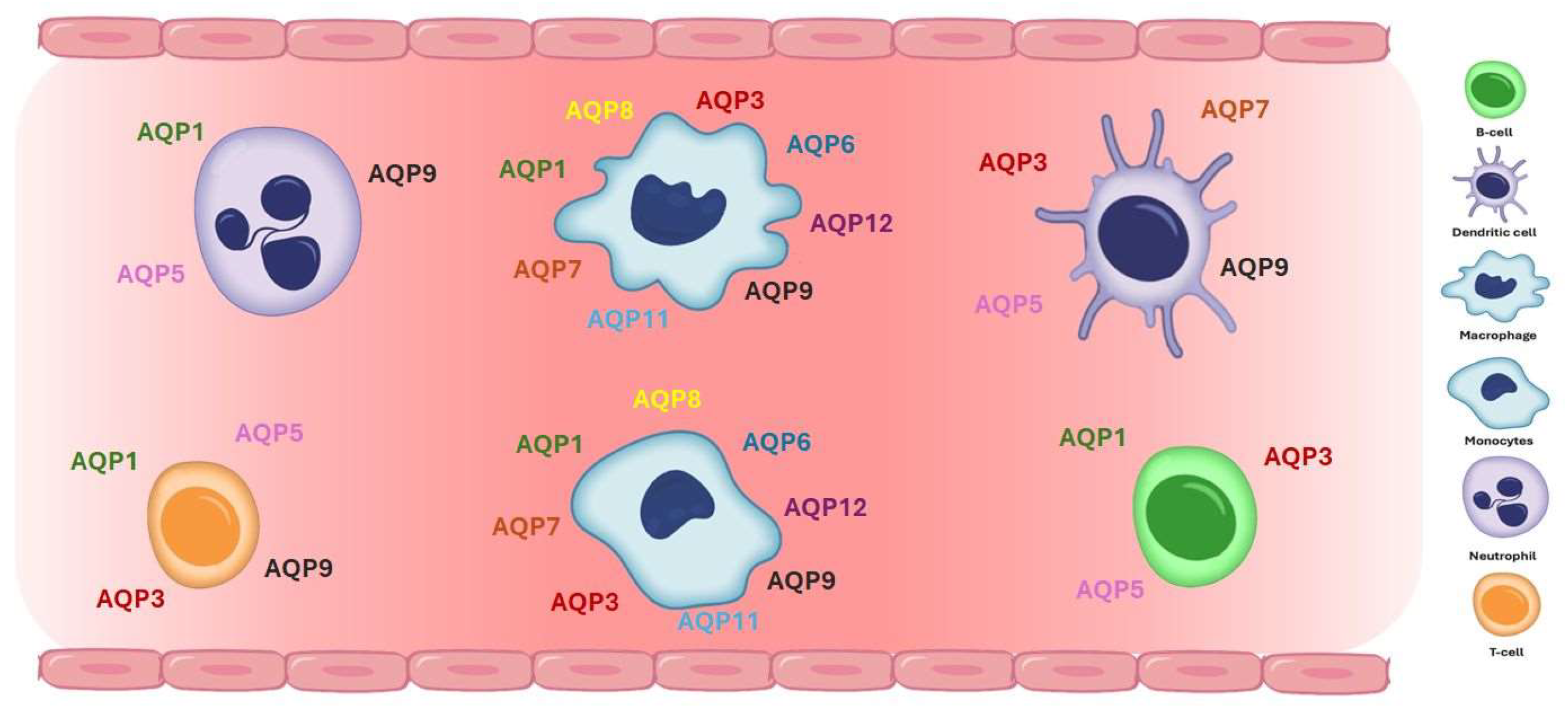
Figure 3.
Aquaporins in the human lung. The lung facilitates oxygen and carbon dioxide exchange between air and blood. Maintaining the fluid volume is essential for gas exchange, achieved by balancing fluid transport across the epithelium. Aquaporins (AQPs) are integral membrane proteins that act as water channels, facilitating water transport across cell membranes in response to osmotic gradients. Under physiological conditions, aquaporins usually facilitate the management of fluid by transporting it out of the alveolar space into the interstitium or capillaries. Τhe main aquaporins expressed in the lung are AQP1, AQP3, AQP4, and AQP5. AQP1 is present in the capillary endothelial cells and controls water movement across the endothelial barrier. AQP3 and AQP4 are found in the airway epithelium and alveolar epithelial cells. AQP5 is expressed in the alveolar epithelial cells type I and is primarily responsible for water transport across the alveolar barrier. Acute respiratory distress syndrome (ARDS) constitutes a severe lung condition characterized by the widespread release of pro- and anti-inflammatory cytokines, oxidative stress (production of reactive oxygen species-ROS), increased vascular permeability, and accumulation of fluid in the alveoli, which leads to impaired gas exchange and respiratory failure. In ARDS increased water transport by AQPs could contribute to fluid accumulation in the alveoli if the inflammation and barrier dysfunction are uncontrolled, exacerbating pulmonary edema. Disruption of the expression or function of these AQPs, which is often observed in ARDS, compromises the lung’s ability to clear fluid, leading to worsened pulmonary edema. Adapted from [141].
Figure 3.
Aquaporins in the human lung. The lung facilitates oxygen and carbon dioxide exchange between air and blood. Maintaining the fluid volume is essential for gas exchange, achieved by balancing fluid transport across the epithelium. Aquaporins (AQPs) are integral membrane proteins that act as water channels, facilitating water transport across cell membranes in response to osmotic gradients. Under physiological conditions, aquaporins usually facilitate the management of fluid by transporting it out of the alveolar space into the interstitium or capillaries. Τhe main aquaporins expressed in the lung are AQP1, AQP3, AQP4, and AQP5. AQP1 is present in the capillary endothelial cells and controls water movement across the endothelial barrier. AQP3 and AQP4 are found in the airway epithelium and alveolar epithelial cells. AQP5 is expressed in the alveolar epithelial cells type I and is primarily responsible for water transport across the alveolar barrier. Acute respiratory distress syndrome (ARDS) constitutes a severe lung condition characterized by the widespread release of pro- and anti-inflammatory cytokines, oxidative stress (production of reactive oxygen species-ROS), increased vascular permeability, and accumulation of fluid in the alveoli, which leads to impaired gas exchange and respiratory failure. In ARDS increased water transport by AQPs could contribute to fluid accumulation in the alveoli if the inflammation and barrier dysfunction are uncontrolled, exacerbating pulmonary edema. Disruption of the expression or function of these AQPs, which is often observed in ARDS, compromises the lung’s ability to clear fluid, leading to worsened pulmonary edema. Adapted from [141].
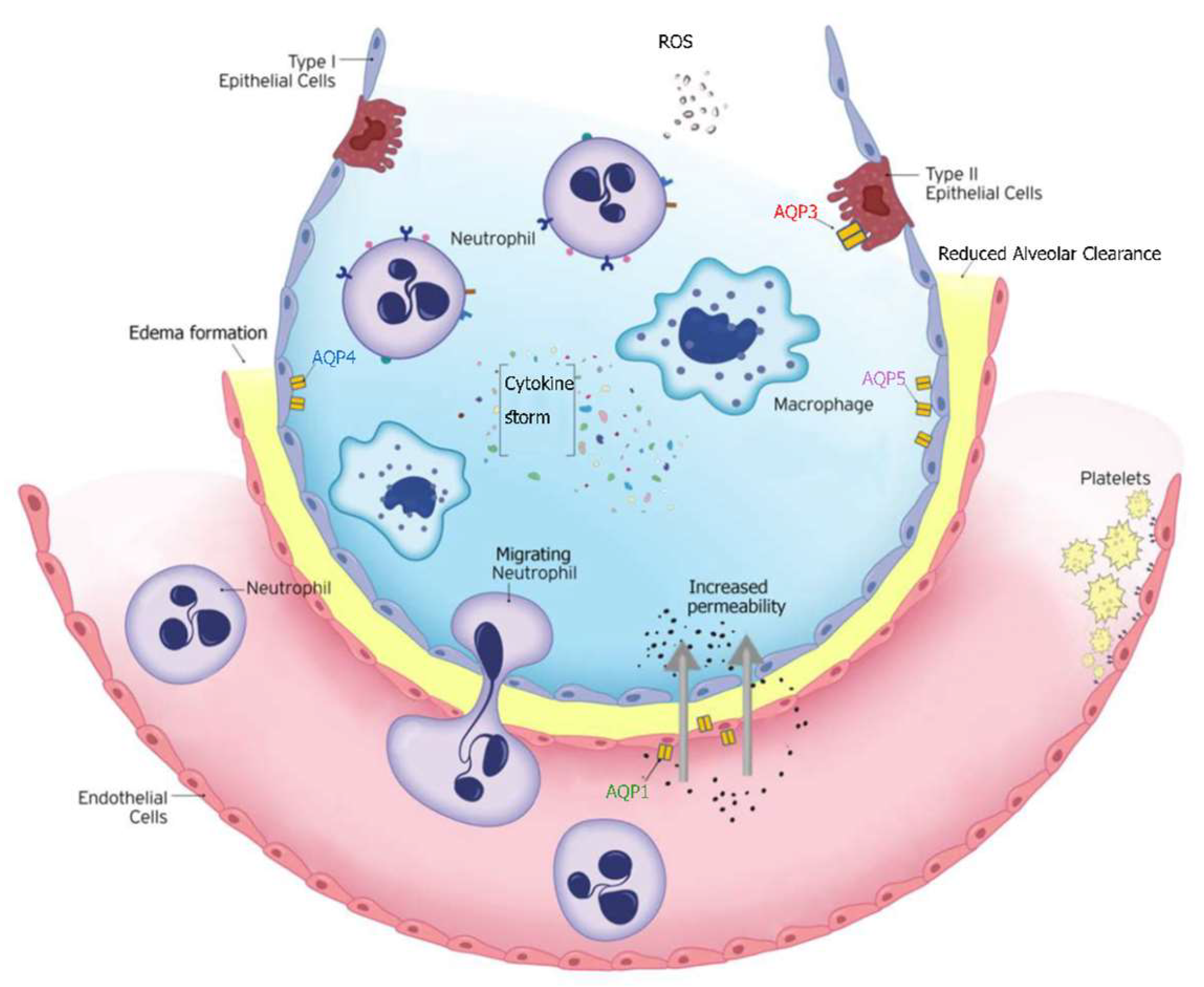
Figure 4.
Aquaporins in the human kidney. Aquaporins (AQPs) play a critical role in kidney function by regulating water transport across membranes, crucial for urine concentration and maintaining fluid balance. AQP1 facilitates water reabsorption in the proximal tubules and descending limb of the loop of Henle, while AQP2 is key in the collecting ducts, regulated by antidiuretic hormone (ADH) to concentrate urine. AQPs 3 and 4 support water exit from the collecting duct cells to the bloodstream. AQP7 is expressed in the brush border of the S3 segment of the proximal tubule and plays a significant role in metabolism by regulating glycerol transport. AQP8 enables bidirectional water and hydrogen peroxide transport across biological membranes, while AQP11 is uniquely located in the endoplasmic reticulum membrane of proximal tubular cells. Dysfunction of AQPs can lead to conditions like nephrogenic diabetes insipidus or fluid imbalances. Image reproduced from [24].
Figure 4.
Aquaporins in the human kidney. Aquaporins (AQPs) play a critical role in kidney function by regulating water transport across membranes, crucial for urine concentration and maintaining fluid balance. AQP1 facilitates water reabsorption in the proximal tubules and descending limb of the loop of Henle, while AQP2 is key in the collecting ducts, regulated by antidiuretic hormone (ADH) to concentrate urine. AQPs 3 and 4 support water exit from the collecting duct cells to the bloodstream. AQP7 is expressed in the brush border of the S3 segment of the proximal tubule and plays a significant role in metabolism by regulating glycerol transport. AQP8 enables bidirectional water and hydrogen peroxide transport across biological membranes, while AQP11 is uniquely located in the endoplasmic reticulum membrane of proximal tubular cells. Dysfunction of AQPs can lead to conditions like nephrogenic diabetes insipidus or fluid imbalances. Image reproduced from [24].
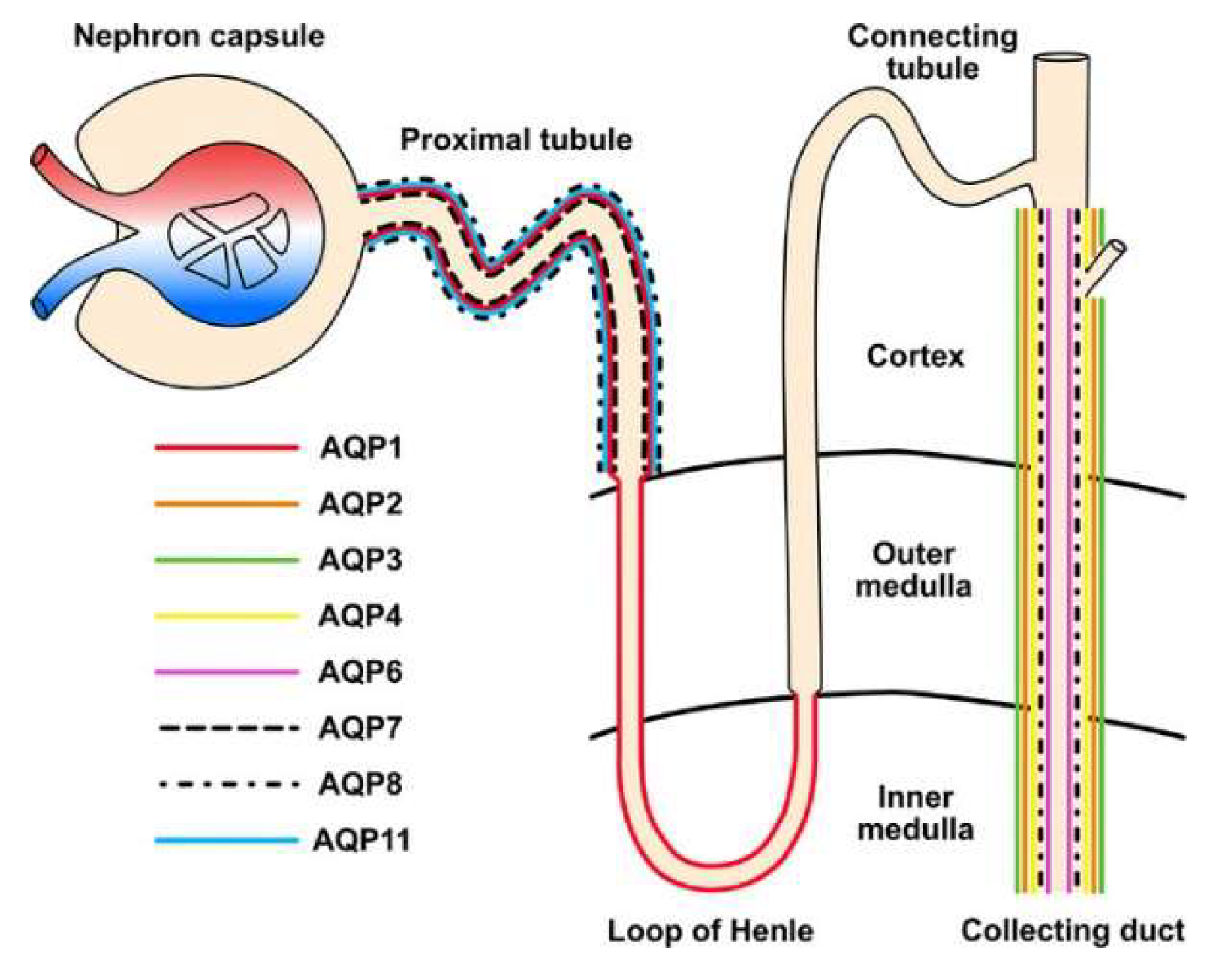
Figure 5.
Main pathophysiological roles of aquaporins in acute brain injury. Aquaporins (AQPs) are pivotal in the pathophysiology of acute brain injury, and their role extends across key processes such as cerebral edema formation, blood-brain barrier regulation, and inflammatory response modulation. AQP4 has emerged as a central player, influencing water homeostasis and edema resolution, while other aquaporins, such as AQP1, AQP2, and AQP9, contribute to diverse pathophysiological mechanisms, including immune cell migration and metabolic adaptation. Aspects of this figure were adapted with permission from Servier Medical Art library, available under Creative Commons license.
Figure 5.
Main pathophysiological roles of aquaporins in acute brain injury. Aquaporins (AQPs) are pivotal in the pathophysiology of acute brain injury, and their role extends across key processes such as cerebral edema formation, blood-brain barrier regulation, and inflammatory response modulation. AQP4 has emerged as a central player, influencing water homeostasis and edema resolution, while other aquaporins, such as AQP1, AQP2, and AQP9, contribute to diverse pathophysiological mechanisms, including immune cell migration and metabolic adaptation. Aspects of this figure were adapted with permission from Servier Medical Art library, available under Creative Commons license.
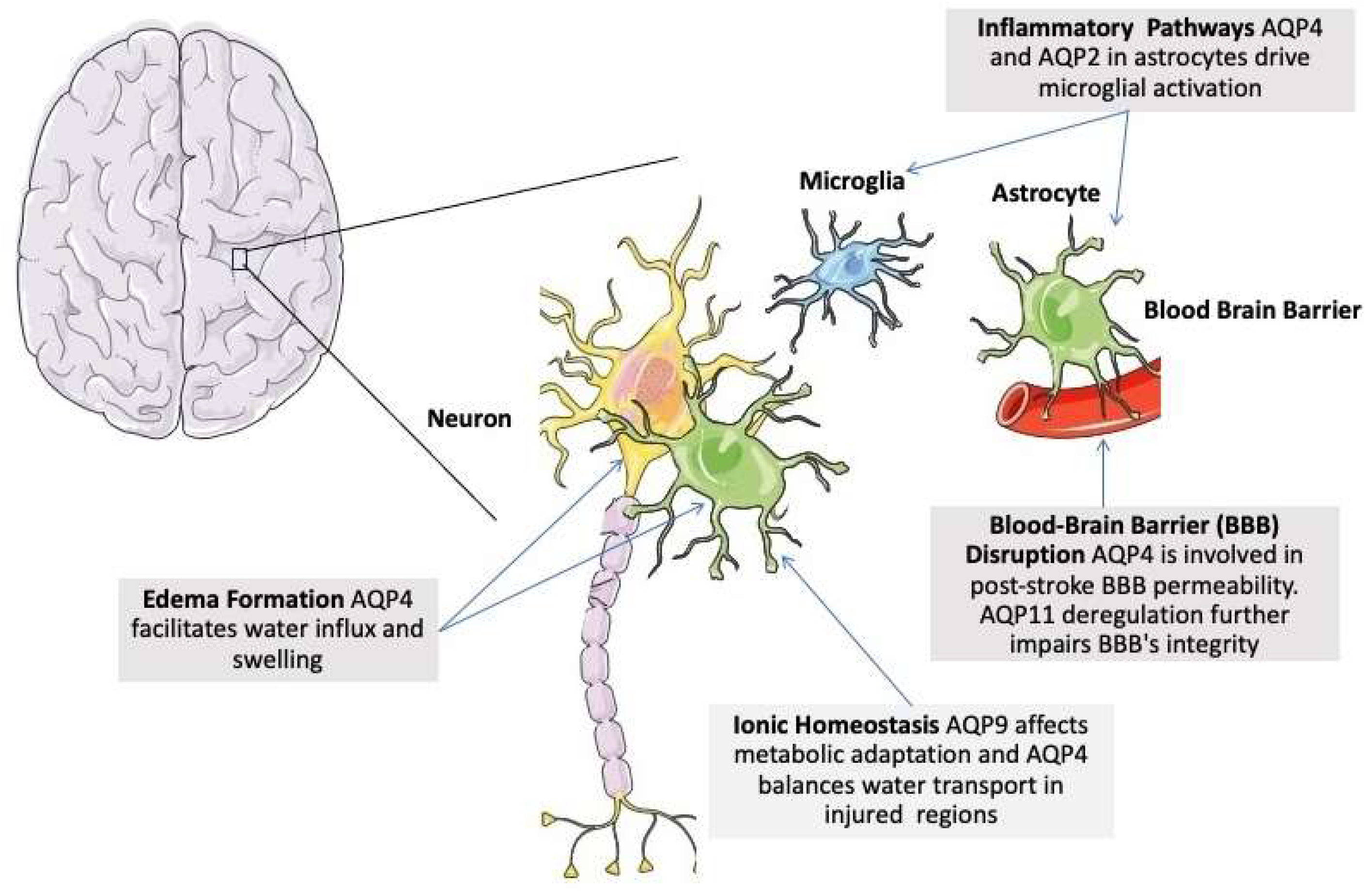

Table 1.
Aquaporins: Localization and role in physiology and disease.
| Aquaporin | Localization | Function | Role in Critical Illness |
|---|---|---|---|
| AQP0 | Eye lens, skin, male reproductive system | Acts as both a water channel and a structural protein in the lens. Maintains lens transparency and hydration | Impairment may contribute to cataracts and other ocular complications in critically ill patients |
| AQP1 | Kidneys, eye, brain, heart, lung, liver, skeletal muscle, blood cells, and various glands | Facilitates rapid water transport for maintaining plasma volume, cerebrospinal fluid (CSF) production, and microvascular fluid exchange | Altered expression affects fluid retention, edema, and organ function in critical illness |
| AQP2 | Kidneys (collecting ducts), ear, stomach, intestines, reproductive systems | Regulated by vasopressin; essential in water reabsorption, to concentrate urine and maintain body water balance | Dysregulation linked to electrolyte imbalances and water retention complications |
| AQP3 | Kidney, skin, immune cells, gastrointestinal tract (salivary and pancreatic secretion), lung, spinal cord | Transports water, glycerol, and small solutes, contributing to hydration, skin elasticity, and energy metabolism | |
| AQP4 | Brain, spinal cord, lung, kidney, stomach, skeletal muscle | Regulates osmotic balance in the central nervous system, critical in brain edema, supports glymphatic system function | Key role in cerebral edema management in brain injury and neurological illness |
| AQP5 | Epithelial tissues (salivary glands, airways), lung, immune cells, pancreas, skin | Facilitates fluid secretion, essential for saliva production and lung function | Altered levels associated with respiratory distress and secretion deficits |
| AQP6 | Kidney (intercalated cells), ear, female reproductive system | Transports water and anions, aiding acid-base balance | Potentially impacts acid-base imbalances and renal dysfunction in critical illness |
| AQP7 | Adipose tissue, kidney, gastrointestinal tract, heart | Transports glycerol and water, facilitating energy balance by releasing glycerol from adipose tissue for gluconeogenesis | Implicated in metabolic dysregulation and fluid imbalances under critical conditions |
| AQP8 | Liver, pancreas, kidney, gastrointestinal tract | Supports cellular osmoregulation, ammonia detoxification, particularly in the liver, and bile secretion | Dysfunction associated with liver and pancreatic issues in critical illness |
| AQP9 | Liver, immune cells, heart, spinal cord, spleen | Transports water, glycerol, urea; role in metabolic processes and immune cell function | Dysregulation affects immunity, metabolic balance, and organ perfusion |
| AQP10 | Intestine, ear, heart | Transports water and glycerol, contributing to intestinal absorption and lipid metabolism | Linked to fluid absorption issues and nutrient transport under stress |
| AQP11 | Kidney, heart, gastrointestinal tract, reproductive systems | Associated with renal development and function | Deficiency may contribute to renal and cardiovascular complications |
| AQP12 | Pancreas, female reproductive system | Involved in digestive fluid secretion and pancreatic enzyme secretion regulation | Potential role in pancreatic insufficiency and digestive issues in critical care |
Table 2.
Studies on aquaporins in critically ill patients with injuries involving the immune system, lung, kidney, and brain.
Table 2.
Studies on aquaporins in critically ill patients with injuries involving the immune system, lung, kidney, and brain.
| Aquaporin | Disease/condition | Findings | References |
| Sepsis | Higher mRNA expression in leukocytes from septic patients | [95] | |
| Sepsis | Total peripheral blood mRNA decreased in sepsis | [113] | |
| AQP1 | ARDS | Lung tissue mRNA expression mildly elevated in ARDS patients | [150] |
| ARDS | Total blood RNA expression elevated in ARDS | [95] | |
| COVID-19 respiratory failure | Higher serum levels in the patient group | [151] | |
| AKI | Urine AQP2 levels were higher in the AKI group; independently associated with AKI; AUC for AKI diagnosis= 0.795 | [193] | |
| AQP2 | TBI/Acute and chronic SDH | AQP2 linked to hematoma volume and midline shift in chronic SDH | [225] |
| Sepsis | Whole blood mRNA expression increased over the duration of sepsis and correlated with survival | [107] | |
| AQP3 | Sepsis | CC carriers of the rs17553719 SNP exhibited lower 30-day survival and higher admission mRNA expression | [108] |
| TBI | CSF AQP4 levels increased post-TBI | [223] | |
| TBI | Specific AQP4 SNPs linked to patient outcome, with no association with initial injury severity | [221] | |
| AQP4 | TBI | Elevated microparticles with AQP4 expression in blood; higher levels in TBI patients vs. controls, with progressive changes post-injury | [222] |
| AIS | High SBP after thrombolysis correlates with oxidative stress-induced BBB disruption and AQP4 upregulation, linked to early neurological decline | [224] | |
| SAE | AQP4 protein upregulation in blood samples | [156] | |
| Acute and chronic SDH | No significant findings for AQP4 in acute SDH | [225] | |
| Sepsis | 1364A/C SNP associated with lower AQP5 expression and higher survival | [118] | |
| Sepsis | AQP5 promoter DNA methylation associated with higher expression and increased mortality risk | [89] | |
| Sepsis | Healthy volunteer neutrophils carrying the A allele of the 1364A/C SNP show increased migration | [100] | |
| AQP5 | Sepsis | The C-allele in sepsis is accompanied by a higher methylation level of the AQP5 promoter | [120] |
| ARDS | Patients with AA genotype of the 1364A/C SNP showed increased mortality | [152] | |
| ARDS | The presence of the AA genotype of the 1364A/C SNP was associated with aggravated pulmonary inflammation, while the carriers of the C allele had higher 30-day survival | [27] | |
| AKI | AA genotype of the AQP5-1364A/C promoter polymorphism showed an increased prevalence of AKI compared to the AC/CC genotype and was independently associated with persistent AKI | [152] | |
| Sepsis | Increased mRNA expression in PMNs of SIRS patients | [103] | |
| Sepsis | Whole blood mRNA expression unaltered during sepsis and low levels associated with increased survival | [107] | |
| AQP9 | ARDS | The initial results showed increased whole blood mRNA expression in ARDS patients. However, subsequent validation analysis in a different group of ARDS patients did not confirm these findings | [153] |
| TBI/Acute and chronic SDH | No significant findings for AQP9 in acute SDH | [225] |
AIS, acute ischemic stroke; AKI, acute kidney injury; AQP, aquaporin; ARDS, acute respiratory distress syndrome; AUC, area under the curve; BBB, blood-brain barrier; CSF, cerebrospinal fluid; PMNs, polymorphonuclear neutrophils; RF, respiratory failure; SBP, systolic blood pressure; SDH, subdural hematoma; SIRS, systemic inflammatory response syndrome; SNPs, single nucleotide polymorphisms; TBI, traumatic brain injury.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



