
Submitted:
03 December 2024
Posted:
03 December 2024
You are already at the latest version
Abstract
Weaning in piglets presents significant physiological and immunological challenges, including gut dysbiosis and increased susceptibility to post-weaning diarrhoea (PWD). Abrupt dietary, environmental, and social changes during this period disrupt the intestinal barrier and microbiota, often necessitating antimicrobial use. Sustainable dietary strategies are critical to addressing these issues while reducing reliance on antimicrobials. Reducing dietary crude protein mitigates the availability of undigested proteins for pathogenic bacteria, lowering harmful byproducts like ammonia and branched-chain fatty acids, which exacerbate dysbiosis. Organic acid supplementation improves gastric acidification, nutrient absorption, and microbial balance, while also serving as an energy-efficient alternative to traditional grain preservation methods. Increasing intestinal butyrate, a key short-chain fatty acid with anti-inflammatory and gut-protective properties, is particularly promising. Butyrate strengthens intestinal barrier integrity by upregulating tight junction proteins, reduces inflammation by modulating cytokine responses, and promotes anaerobic microbial stability. Exogenous butyrate supplementation via salts provides immediate benefits, while endogenous stimulation through prebiotics (e.g., resistant starch) and probiotics promotes sustained butyrate production. These interventions selectively enhance butyrate-producing bacteria such as Roseburia and Faecalibacterium prausnitzii, further stabilising the gut microbiota. Integrating these strategies can enhance gut integrity, microbial resilience, and immune responses in weaned piglets. Their combination offers a sustainable, antimicrobial-free approach to improving health and productivity in modern pig production systems.
Keywords:
post-weaning
; microbiota
; gut health
; organic acids
; grain
; crude protein
; intestinal dysfunction
; antimicrobial resistance
; butyrate
; swine
1. Introduction
Weaning represents a critical developmental phase for piglets, characterised by abrupt dietary, environmental, and social changes that pose significant physiological and immunological challenges. The transition from sow’s milk to solid feed disrupts the gut environment, often leading to impaired digestion, gut dysbiosis, and post-weaning diarrhoea (PWD), a major health and economic challenge in swine production [1,2]. During the suckling phase, piglets rely on lactose as their primary energy source, with its fermentation producing lactic acid that compensates for underdeveloped hydrochloric acid (HCl) secretion. However, the shift to solid feed reduces lactic acid production, leaving gastric pH elevated (approximately 5.0), which is insufficient for optimal pepsin activation and protein digestion [3]. Consequently, undigested proteins reach the distal colon, where they are fermented by nitrogen-utilising bacteria, generating harmful by-products that disrupt gut homeostasis, increase inflammation, and compromise intestinal barrier function [4,5,6].
Traditionally, pharmacological levels of zinc oxide have been widely used in piglet diets to address weaning-related challenges, improving growth performance and reducing the incidence of PWD. However, growing concerns regarding its environmental impact, specifically the accumulation of zinc in soil and water ecosystems, and its role in promoting antimicrobial resistance have led to regulatory bans, including its prohibition in the European Union as of 2022 (Commission Implementing Decision, 2017).
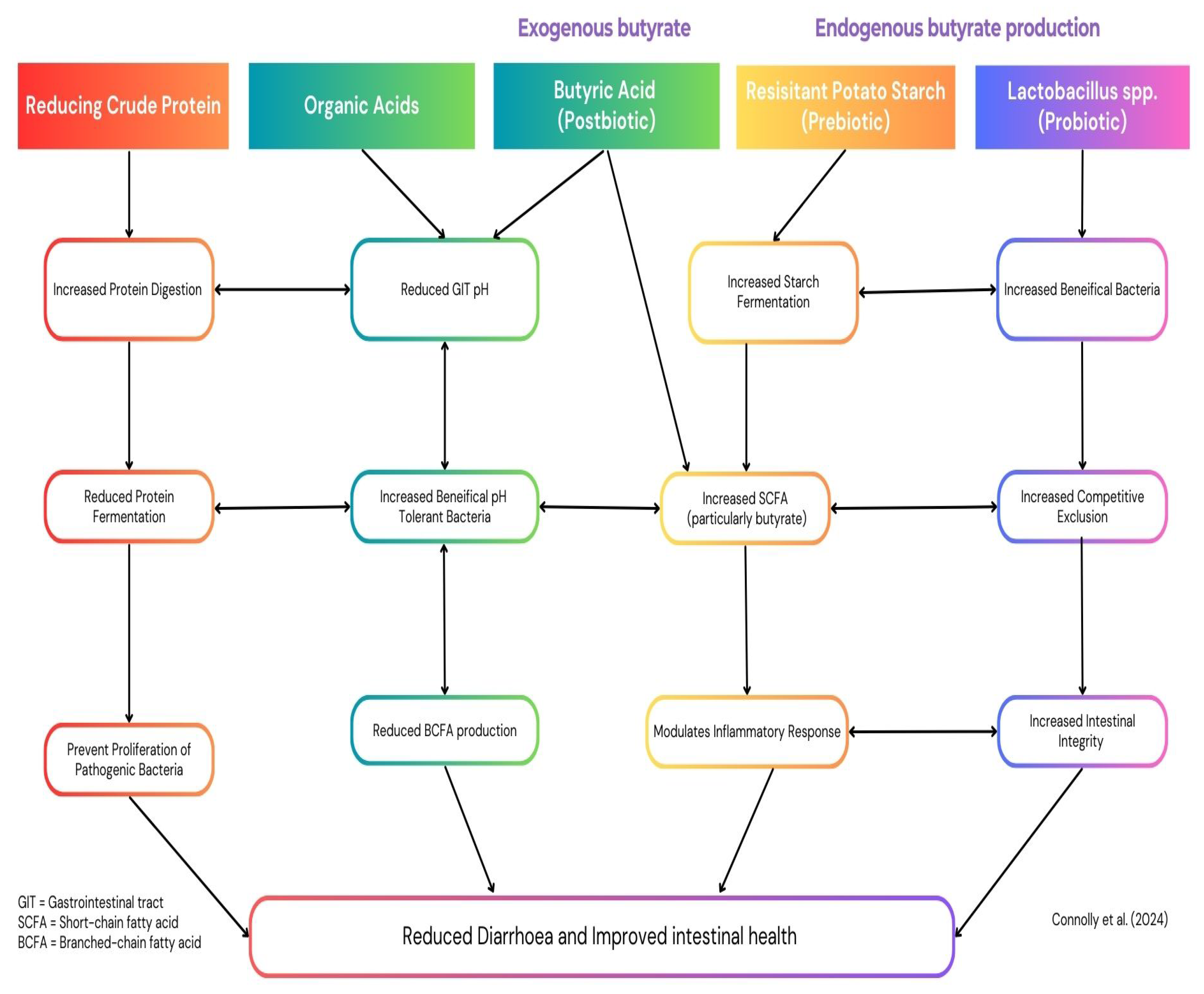
This review evaluates alternative nutritional strategies that address the physiological and microbial challenges associated with weaning, focusing on reducing dietary crude protein (CP), supplementing organic acids, and enhancing intestinal butyrate levels. Reducing CP limits undigested substrates that promote the growth of pathogenic bacteria, while organic acids lower gastric pH, enhance nutrient absorption, and inhibit pathogens. Butyrate, a short-chain fatty acid (SCFA) with potent anti-inflammatory and gut-protective properties, can be supplied directly or promoted endogenously via prebiotics and probiotics, supporting gut integrity and microbial stability. While each of these strategies shows considerable promise when applied individually, their synergistic potential offers a more effective and sustainable solution for mitigating the adverse effects of weaning, as illustrated in Figure 1. This review explores these strategies in detail, proposing a framework for their integration into modern pig production systems to enhance resilience, growth performance, and sustainability.
2. Impact of Standard Crude Protein Concentrations in Weaner Pig Diets: Effects on Growth, Gut Health and Immune Function
Weaning is a critical developmental phase in piglets, marked by abrupt dietary changes that exert significant physiological and immunological stress on the gastrointestinal tract. To support rapid growth and maximise feed efficiency during this transition, weaner pig diets are often formulated with high CP levels of 20–23% in pre-starter feeds and 18–20% in starter feeds, incorporating high-quality protein sources such as whey protein, soya bean concentrate, potato protein, and fish meal [7,8]. These diets are designed to meet the piglet’s high amino acid demands, particularly lysine, while compensating for their limited feed intake [9,10]. However, while high-CP diets are essential for supporting growth, they also present considerable challenges, including heightened risk of PWD, impaired gut health, and suboptimal post-weaning performance [11].
During weaning, stress and reduced feed consumption compromise digestive and immune functions. The immature gut undergoes morphological changes, including villous atrophy, crypt hyperplasia, and reduced activity of brush-border enzymes such as sucrase, lactase, and lipase [12,13]. These changes impair the piglet’s ability to digest and absorb nutrients efficiently. High dietary CP exacerbates this issue by delivering excess undigested protein to the distal intestine, reducing overall digestive efficiency and contributing to gastrointestinal disturbances [14]. In the colon, undigested protein undergoes microbial fermentation, raising colonic pH and promoting the proliferation of pathogenic bacteria such as Clostridium and Bacteroides [15]. Protein fermentation produces harmful by-products, including ammonia, phenols, and amines, which disrupt gut homeostasis, induce inflammation, and exacerbate PWD [5,16,17]. Additionally, osmotic effects associated with protein fermentation further aggravate diarrhoea by drawing excess water into the intestinal lumen [15]. High-CP diets also alter the gut microbiota by reducing bacterial diversity and enriching pro-inflammatory species such as Fusobacterium [18]. These changes increase coliform counts in the colon and faeces while decreasing the beneficial Lactobacillus-to-coliform ratio, compounding dysbiosis and intestinal dysfunction [19].
Compounding these microbiological and physiological challenges, is the piglets' underdeveloped immune system, which is further stressed by weaning. Elevated levels of pro-inflammatory cytokines and increased intestinal permeability are common responses to weaning stress [20,21]. High-CP diets can intensify these effects by promoting allergic reactions and inflammatory responses, further exacerbating gastrointestinal disturbances and PWD [22]. Effective protein digestion, facilitated by sufficient digestive enzyme activity and bile salt production, is essential for breaking down dietary proteins into non-immunogenic fragments and mitigating these effects [23].
2.1. The Effects of Low Crude Protein in Weaner Pig Diets : Diarrhoea, Gut Health, Immune Function and Growth
Managing CP levels plays a pivotal role in enhancing intestinal health and reducing the incidence of PWD in piglets. Extensive research has demonstrated that lowering CP levels in piglet diets effectively limits the amount of undigested protein reaching the gastrointestinal tract (GIT). This reduces protein fermentation, thereby minimising the production of harmful by-products such as ammonia, amines, and polyamines [19], [24,25,26], which damage colonic epithelial cells and decrease villous height in the small intestine, significantly impairing nutrient absorption [16,27]. Furthermore, elevated ammonia levels disrupt the oxidative metabolism of SCFAs, reducing energy availability in the large intestine and compromising its overall functionality [5]. Hence, lowering dietary CP levels is an essential strategy for improving gut health and optimising post-weaning performance in piglets.
Numerous studies have highlighted the benefits of low CP diets supplemented with amino acids in weaned piglets which have been highlighted in Figure 1. For example García et al., [28] demonstrated reduced diarrhoea scores and increased SCFA production in piglets fed a 16% CP diet supplemented with amino acids and probiotics compared to a 20% CP diet. Similarly, Tian et al., [29] observed that a 14% CP diet with essential amino acid supplementation enhanced pancreatic enzyme expression compared to a 17% CP diet and a 20% CP diet, while Opapeju et al., [30] reported reduced ammonia concentrations and shorter crypt depths in pigs fed a 19% CP diet compared to those offered a 21% CP diet. However, reducing CP levels to 17% or lower may impair growth performance by decreasing villous height and reducing pancreatic function, which can compromise nutrient absorption [31,32]. [24,25,26]
Additionally, low-CP diets have been shown to promote beneficial microbial populations, such as butyrate-producing bacteria like Roseburia and Eubacterium rectale, which support gut health and reduce inflammation [5]. Marchetti et al., [33] observed that piglets on a 15.5% CP diet had higher abundances of anti-inflammatory bacteria (Succinivibrionaceae), fibre-degrading bacteria (Fibrobacteraceae), and Lactobacillus, indicating a more stable and beneficial microbiome. However, excessively low CP levels may reduce key butyrate-producing bacteria such as Faecalibacterium prausnitzii and lower SCFA butyrate production, potentially compromising gut health [34,35]. These results suggest that microbial responses to CP levels can vary depending on factors such as sanitation, genetics, social stress, or the protein source used in diets.
Low-CP diets also have implications for immune modulation. High-CP diets have been shown to suppress the expression of the MCT1 gene, which is crucial for mucus production and gut barrier function. Conversely, low-CP diets stimulate MCT1 expression, which is enhanced by butyrate and helps control intestinal inflammation [5]. Zhang et al,. [36] found that piglets on a low-CP diet exhibited reduced expression of pro-inflammatory cytokines in the colon, correlating with lower levels of ammonia and E. coli. Additionally, Fan et al., [37] reported that moderate reductions in CP levels increased the expression of tight junction proteins, strengthening gut integrity. However, certain low-CP diets have been associated with increased inflammation, particularly in the presence of soya antigens or synthetic amino acids. Rattigan et al., [38] observed elevated pro-inflammatory markers in piglets on a 18% low-CP diet, likely due to heightened sensitivity to dietary antigens. Furthermore, synthetic amino acids may lack the ability to bind to immune cells, unlike natural proteins, potentially failing to stimulate regulatory immune responses and exacerbating inflammation [39]. Variability in feed ingredient quality, feeding patterns, and environmental conditions complicates the consistent implementation of low-CP diets [27].
A significant challenge of low-CP diets is maintaining growth performance. When CP levels are reduced, careful dietary formulation is required to ensure amino acid requirements are met [27]. It is recommended to supplement with the first four limiting amino acids or branched-chain amino acids when CP levels are lowered below 3%, and with dietary nitrogen or non-essential amino acids (NEAAs) when CP levels are lowered below 6% [27]. Lowering CP levels can limit the availability of nitrogen for synthesising NEAAs, which are essential for protein synthesis and growth. However variations in feed ingredients, feeding patterns and environmental conditions can make it difficult to achieve consistent results [27]. This variability is a major limitation in the widespread adoption of low-CP diets. Supporting this, studies report mixed results. For example, Nyachoti et al,. [40] and Deng et al,. [31] observed reduced growth performance when CP levels were lowered to 17% and 12.7%, respectively. Conversely, Rattigan et al,. [34] and Fang et al,. [14] found that reducing CP levels to 18% maintained growth performance, with further reductions improving feed efficiency.
Balancing CP levels in piglet diets remains critical for optimising growth, gut health, and immune function while minimising PWD. Although reducing CP levels promotes microbiome stability and reduces inflammation, care must be taken to avoid compromising growth and digestive performance. Fine-tuning CP levels with precise amino acid supplementation is crucial for developing optimal nutritional strategies for weaned piglets. Reducing CP levels offers benefits such as reduced nitrogen excretion, improved gut health, and decreased risk of PWD. However, achieving these benefits while maintaining growth performance remains challenging due to the need for balanced nitrogen and amino acid availability.
3. The Role of Organic Acids in Weaner Pig Diets
The inclusion of organic acids in weaning piglet diets has proven to be an effective strategy for improving growth performance, stimulating feed intake, enhancing gut health, and bolstering resilience during the critical post-weaning period, with their mechanisms outlined in Figure 1. Organic acids can be classified into three primary categories: SCFAs, medium-chain fatty acids (MCFAs), and tricarboxylic acids (TCAs). These acids are added to diets either individually or as blends, in free forms or as salts [41]. As naturally occurring cellular metabolites, organic acids have low toxicity and are well-tolerated by animals [42]. Short-chain fatty acids, such as acetic, propionic, and butyric acids (≤5 carbons), are particularly important for maintaining intestinal morphology and barrier integrity [43]. Medium-chain fatty acids (7–12 carbons) act as efficient energy sources [44], while TCAs, intermediates of the Krebs cycle, positively influence gut barrier function, intestinal microbiome balance, and structural integrity [43].
Piglets exhibit a natural preference for sour tastes, making organic acids like tartaric and citric acids appealing dietary supplements [42]. The addition of 1.6% lactic acid to piglet diets significantly increases average daily feed intake (ADFI), comparable to diets containing antibiotics such as Lincospectin (lincomycin and spectinomycin) [45]. Organic acid supplementation generally enhances ADFI compared to non-supplemented diets, although excessive concentrations of acids like acetic, propionic, or formic acids can cause discomfort, reducing feed intake [46]. This highlights that the efficacy of organic acids depends on factors such as acid type, inclusion levels, and interactions with other dietary components.
One of the major physiological challenges during weaning is the increase in gastric pH, which compromises the stomach's role as a pathogen barrier [47]. Before weaning, bacteria such as Streptococcus suis constitute less than 4% of the stomach’s microbiome, while Lactobacillus represents approximately 14% [48]. Post-weaning, the proportion of S. suis increases, while Lactobacillus levels decrease, increasing the pathogenic load throughout the GIT. High gastric pH not only facilitates bacterial overgrowth but also impairs protein digestion and the absorption of nutrients like calcium, iron, and vitamin B12 [49]. Organic acids, with their low acid-binding capacities, help reduce dietary and gastric pH, improving digestion and suppressing pathogen growth in post-weaned piglets [50]. However, research on the effects of organic acids on gastric pH shows mixed results. While Hansen et al,. [51] observed that lactic and formic acids reduced stomach pH in weaned piglets, Roth et al,. [52] and Li et al,. [53] found no significant impact of certain organic acids or blends on gastric pH. These variations suggest that the benefits of organic acids extend beyond pH reduction and include mechanisms such as improved nutrient digestibility. For instance, organic acid supplementation has been shown to enhance pancreatic enzyme production and increase the digestibility of amino acids like methionine, lysine, and leucine [54]. By lowering pH, organic acids can also shorten feed transit time, allowing for more efficient nutrient absorption and better utilisation of minerals like phosphorus, zinc, and calcium [3,55].
The effects of organic acids on intestinal morphology are similarly variable. Some studies report no changes in gut structure with organic acid supplementation [56,57], while others demonstrate improvements in villous height-to-crypt depth ratios, indicating enhanced nutrient absorption [58,59].
Beyond their digestive benefits, organic acids also modulate the microbiome. They create an acidic environment that is unfavourable for pathogens like E. coli and Clostridia while supporting beneficial bacteria such as Lactobacillus and SCFA-producing bacteria [3,60]. For example, Pluske et al,. [61] found that piglets supplemented with organic acids, MCFAs, and phenolic compounds exhibited an increased abundance of beneficial bacterial families, including Lachnospiraceae and Lactobacillaceae. These bacteria are associated with anti-inflammatory effects, improved immune function, and tight junction integrity [62]. However, the efficacy of organic acids as antimicrobials varies with their chain length, saturation, and inclusion levels [42].
Despite their benefits, the inappropriate use of organic acids can lead to unintended effects. For instance, high levels of potassium diformate (1.8%) can reduce beneficial lactic acid bacteria in faeces, emphasising the need for precise formulation [63]. Overall, incorporating organic acids into piglet diets offers significant potential for improving gut health, nutrient utilisation, and microbiome balance, but their application must be carefully tailored to optimise benefits while minimising drawbacks.
3.1. The Potential of Organic Acid-Preserved Grain in Weaner Diets : Enhancing Grain Quality, Gut Health, Growth and Sustainability
Grain preservation is a fundamental aspect of animal feed production, as it ensures the maintenance of feed quality while preventing spoilage. Traditional grain drying methods, though widely adopted, are unsustainably energy-intensive and often lead to uneven moisture distribution, which fosters mould growth and increases the risk of mycotoxin contamination [64,65]. This is particularly detrimental in piglet nutrition, as mycotoxins are known to impair growth, compromise immune function, and negatively affect overall health, thereby reducing productivity [66].
Organic acids have emerged as a versatile solution in feed preservation, offering a more energy-efficient alternative to traditional drying processes. Preserving grain using organic acid is an uncomplicated process. Initially, the grain is emptied into a screw conveyor, where the acid is applied at the desired concentration. The grain and acid are then mixed thoroughly before storage in a corrosive-resistant storage facility. By lowering grain pH and inhibiting fungal proliferation, organic acids such as propionic acid effectively prevent mould growth and mycotoxin formation, preserving feed quality and extending shelf life [69]. Moreover, organic acid-preserved grains have demonstrated superior nutritional and performance benefits compared to heat-dried grains. Supplementing organic acid-preserved grains exhibited higher digestible and metabolisable energy values, contributing to improved average daily gain (ADG) and ADFI in pigs [70]. Similarly, piglets fed organic acid-preserved grain exhibited greater growth performance compared to those receiving the same organic acid blend as a dietary additive [71], emphasising the added nutritional value of preserved grains.
Beyond nutritional benefits, organic acid-preserved grains positively impact piglet health. Maher et al,. [67] observed enhanced growth performance, improved nutrient digestibility, and increased proliferation of beneficial gut microbiota, such as Faecalibacterium, in piglets consuming organic acid-preserved grains. These findings suggest that organic acid preservation not only improves feed quality but also enhances post-weaning gastrointestinal health and resilience.
The adoption of organic acid-preserved grains has the potential to promote sustainable swine production by reducing microbial contamination in feed, improving nutritional value, and directly supporting gut health through pathogen inhibition and nutrient optimisation. However, further research is warranted to evaluate the long-term effects of organic acid-preserved grains on intestinal health, growth performance, and interactions with other dietary strategies. Such investigations will provide a deeper understanding of their role in enhancing piglet performance and advancing sustainable livestock production practices.
4. The Protective Effects of Butyrate
Gut dysbiosis during the weaning phase disrupts the proliferation of butyrate-producing bacteria, significantly impairing the intestinal health and resilience of weaned piglets. This imbalance is characterised by an overgrowth of facultative anaerobic pathogens such as E. coli and Salmonella and a corresponding decline in beneficial butyrate-producing bacteria like Roseburia and Faecalibacterium prausnitzii [57], [72,73,74]. Butyrate, a key SCFA, plays a critical role in maintaining intestinal homeostasis, promoting gut barrier integrity, regulating immune responses, and providing energy to colonocytes. Enhancing butyrate levels in weaned pigs through dietary strategies is a promising solution to mitigate post-weaning challenges such as PWD, inflammation, and compromised growth performance. Butyrate provides up to 70% of the energy requirements of colonocytes, making it vital for maintaining the structural integrity and regenerative capacity of the intestinal lining [75,76,77]. Colonocytes absorb butyrate through transporters such as SLC16A1 (MCT1) and SLC5A8 (SMCT1), which facilitate its rapid oxidation to generate ATP. This energy supports the maintenance of tight junctions, crucial for preventing pathogen translocation and maintaining intestinal barrier function. In addition to its role as an energy source, butyrate has profound immunomodulatory effects. It suppresses pro-inflammatory cytokines such as IL-6, CXCL8,TNF-α, and IFN-γ, while upregulating anti-inflammatory markers like IL-10 and TGF-β [78,79]. These actions reduce intestinal inflammation, support immune tolerance, and promote resilience against weaning-associated stress. Butyrate also preserves an anaerobic gut environment, which is critical for maintaining a healthy microbiota. Anaerobic bacteria dominate the microbiota under homeostatic conditions, effectively suppressing facultative anaerobes, including pathogens like E. coli and Salmonella. However, during dysbiosis, facultative anaerobes proliferate, further disrupting the microbial balance and outcompeting butyrate producers [73]. By fostering an anaerobic environment, butyrate indirectly supports beneficial microbes and strengthens intestinal defenses against infections.
4.1. The Effects of Supplementing Exogenous Butyrate on Post-Weaned Pig Growth and Gut Health
Strategies to enhance butyrate levels include exogenous supplementation with butyrate salts and promoting endogenous butyrate production through microbial fermentation. Exogenous supplementation typically uses sodium, potassium, magnesium, or calcium butyrate, which are less odorous and more practical for animal feed than pure butyric acid [80]. Exogenous butyrate supplementation has exhibited varying impacts on the performance of weaned piglets, with its proposed mechanism of action detailed in Figure 1. Piva et al,. [81] found that sodium butyrate at 0.8 g/kg increased ADG by 20% and feed intake by 16% during the first 15 days post-weaning. Similarly, Feng et al,. [82] showed that 2,000 mg/kg sodium butyrate improved gut barrier integrity by increasing the expression of tight junction proteins such as occludin, claudin-3, and ZO-1, reducing intestinal permeability and diarrhoea. However, not all studies report consistent benefits. Biagi et al [81] observed no significant effects of sodium butyrate on growth performance or mucosal morphology, though they noted increased caecal pH and ammonia concentrations. Jang et al,. [84] reported that 500–1,000 ppm of coated sodium butyrate reduced feed intake, emphasising the need for optimising dosages and delivery methods.
4.2. Promoting Endogenous Butyrate Production in Weaned Pigs via Prebiotic and Probiotic Supplementation: Effects on Growth, Gut Health and the Gut Microbiome
Endogenous butyrate production relies on the fermentation of dietary fibres and prebiotics, with resistant starch (RS) being particularly effective. Resistant starch, especially RS2 (e.g., raw potato starch), resists enzymatic digestion in the small intestine and reaches the colon intact, where it undergoes fermentation by butyrate-producing bacteria such as Roseburia and Faecalibacterium prausnitzii [85]. This fermentation process promotes butyrate synthesis while reducing harmful byproducts like branched-chain fatty acids and ammonia, which are associated with proteolytic fermentation [17,86]. Studies demonstrate that low to moderate RS supplementation (0.5%–5%) optimises butyrate production, supports beneficial microbial populations, and improves overall gut health [87,88]. Additionally, RS enhances gut barrier function by increasing MUC2 expression, a critical mucin protein, and IgA secretion, strengthening mucosal defenses [89]. Excessive RS supplementation (≥7%), however, can disrupt microbial balance, reduce butyrate synthesis, and exacerbate diarrhoea [90,91]. This emphasises the importance of precise inclusion to achieve optimal benefits without adverse effects.
Probiotics enhance the effectiveness of resistant starch (RS) in promoting butyrate production by creating a gut environment conducive to fermentation, as illustrated in Figure 1. Strains such as Lactobacillus plantarum and Lactobacillus reuteri produce lactate and acetate, which act as precursors for butyrate synthesis via cross-feeding with butyrate-producing bacteria [92]. For instance, supplementation with Lactobacillus plantarum has been shown to increase colonic butyrate concentrations and promote the growth of beneficial species like Roseburia spp. and Faecalibacterium prausnitzii [93]. Probiotics also strengthen gut barrier integrity by upregulating tight junction proteins, reducing intestinal permeability, and alleviating damage caused by weaning stress [94,95]. Their immunomodulatory effects, including increased IL-10 and TGF-β1 production, further enhance the gut environment for sustained butyrate synthesis [78].
The combination of RS and probiotics presents a synergistic strategy for improving gut health. RS provides fermentable substrates for butyrate production, while probiotics stabilise the gut microbiota and facilitate cross-feeding interactions. For example, co-supplementation with RS and Lactobacillus plantarum has been shown to elevate SCFA levels, fortify the gut barrier, and reduce intestinal inflammation during weaning stress [93]. However, balancing RS and probiotics is crucial, as excessive supplementation can disrupt microbial communities and reduce butyrate synthesis [90,96].
Boosting butyrate production through RS and probiotics offers a sustainable, long-lasting solution for improving gut health, immune function, and growth performance in weaned piglets. This approach effectively addresses challenges such as PWD and intestinal inflammation while supporting piglet development and overall resilience.
5. Exploring the Synergistic Effects of Combining these Dietary Strategies for Optimal Health and Growth in Weaner Pigs
A review of the literature indicates that strategies such as reducing dietary CP levels, incorporating organic acids, and enhancing intestinal butyrate levels—either through exogenous supplementation or by promoting endogenous production with pre- and probiotics are promising approaches to mitigate the physiological challenges associated with weaning in piglets. These strategies show significant potential for improving growth performance and intestinal health without relying on antimicrobials. However, the inconsistencies and variabilities in their outcomes present challenges for consistent application in pig production systems. Despite the complementary effects of these strategies, there remains a notable gap in the literature exploring their combined implementation, which could reduce individual limitations and enhance overall efficacy.
For instance, the use of organic acid-preserved grain may provide additional benefits compared to traditional organic acid supplementation by improving grain quality while directly influencing gastrointestinal health and function. Furthermore, integrating organic acid-preserved grain into low-CP diets could mitigate the potential negative effects of reduced CP levels on growth performance by improving protein digestive efficiency, as demonstrated in broiler studies [97]. Similarly, incorporating butyrate supplementation into low-CP diets could bolster intestinal fermentative capacity and compensate for the reduced butyrate production often observed in pigs consuming low-CP diets [35]. A combination of exogenous butyrate supplementation with endogenous butyrate-promoting additives, such as RS and Lactobacillus spp., presents an innovative opportunity. This approach could not only enhance initial intestinal butyrate levels post-weaning to counteract the negative effects of weaning but also prime the intestine for sustained, lifelong butyrate production.
The interconnected mechanisms of these dietary strategies suggest that their combined application could improve post-weaning growth performance and intestinal health while addressing the limitations associated with each individual approach. The potential synergistic effects of these strategies, as depicted in Figure 1, highlight the importance of further research to identify the most effective combinations for optimising gut health and growth performance in weaned piglets. Identifying the optimal integration of these approaches could significantly enhance post-weaning pig production, providing a robust and sustainable alternative to conventional antimicrobial use.
6. Conclusions
In conclusion, lowering dietary CP levels effectively reduces the availability of undigested substrates in the gut, limiting the proliferation of pathogenic bacteria and mitigating PWD. This approach improves gastrointestinal health and promotes a more stable gut microbiome. The incorporation of organic acids further supports gut health by lowering gastric pH, inhibiting harmful bacteria, enhancing nutrient digestibility, and fostering the growth of beneficial microbial populations. Strategies to increase gut butyrate levels, whether through direct supplementation or by stimulating endogenous production, provide critical benefits, including anti-inflammatory effects, improved intestinal integrity, and enhanced immune function, which collectively strengthen the piglet’s resilience during the post-weaning period. Integrating these approaches offers a sustainable and multifaceted strategy to improve weaning outcomes by optimising nutrient utilisation, enhancing growth performance, and promoting a balanced intestinal environment while reducing reliance on antimicrobials. These interventions have the potential to address the physiological and microbial challenges of weaning and support lifelong gut health.
However, the effectiveness of these strategies can be influenced by various factors, including dietary composition, production environments, and individual piglet variability. Further research is needed to refine their application, optimise dosage and timing, and explore interactions between strategies to maximise their synergistic potential. Advancing these nutritional approaches is essential for improving piglet health and welfare, supporting productivity, and promoting sustainability in swine production systems, ultimately contributing to a more resilient and innovative livestock industry.
Author Contributions
Conceptualization, K.R.C, J.V.O and T.S; writing—original draft preparation, K.R.C.; writing—review and editing, K.R.C., J.V.O and T.S; supervision, J.V.O. and T.S; funding acquisition, J.V.O and T.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Department of Agriculture, Food and the Marine (DAFM), grant number 2019R518.
Institutional Review Board Statement
Not Applicable.
Informed Consent Statement
Not Applicable.
Data Availability Statement
Not Applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Gilbert, M.S.; Ijssennagger, N.; Kies, A.K.; Van Mil, S.W.C. Protein fermentation in the gut; implications for intestinal dysfunction in humans, pigs, and poultry. American Journal of Physiology-Gastrointestinal and Liver Physiology 2018, 315, G159–G170. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.M.; Opapeju, F.O.; Pluske, J.R.; Kim, J.C.; Hampson, D.J.; Nyachoti, C.M. Gastrointestinal health and function in weaned pigs: a review of feeding strategies to control post-weaning diarrhoea without using in-feed antimicrobial compounds: Feeding strategies without using in-feed antibiotics. Journal of Animal Physiology and Animal Nutrition 2013, 97, 207–237. [Google Scholar] [CrossRef]
- Suiryanrayna, M.V.A.N.; Ramana, J.V. A review of the effects of dietary organic acids fed to swine. J Animal Sci Biotechnol 2015, 6, 45. [Google Scholar] [CrossRef] [PubMed]
- O’Doherty, J.V.; Bouwhuis, M.A.; Sweeney, T. Novel marine polysaccharides and maternal nutrition to stimulate gut health and performance in post-weaned pigs. Anim. Prod. Sci. 2017, 57, 2376. [Google Scholar] [CrossRef]
- Pieper, R.; Tudela, C.V.; Taciak, M.; Bindelle, J.; Pérez, J.F.; Zentek, J. Health relevance of intestinal protein fermentation in young pigs. Anim. Health. Res. Rev. 2016, 17, 137–147. [Google Scholar] [CrossRef]
- Zhang, H.; Wielen, N.V.D.; Hee, B.V.D.; Wang, J.; Hendriks, W.; Gilbert, M. Impact of Fermentable Protein, by Feeding High Protein Diets, on Microbial Composition, Microbial Catabolic Activity, Gut Health and beyond in Pigs. Microorganisms 2020, 8, 1735. [Google Scholar] [CrossRef]
- Collins, C.L.; et al. Post-weaning and whole-of-life performance of pigs is determined by live weight at weaning and the complexity of the diet fed after weaning. Animal Nutrition 2017, 3, no. 4, pp. 372–379. [Google Scholar] [CrossRef]
- Veldkamp, T.; Vernooij, A.G. Use of insect products in pig diets. JIFF, 2021; 5. [Google Scholar] [CrossRef]
- Gloaguen, M.; Le Floc’h, N.; Corrent, E.; Primot, Y.; van Milgen, J. The use of free amino acids allows formulating very low crude protein diets for piglets1. Journal of Animal Science 2014, 92, 637–644. [Google Scholar] [CrossRef]
- NRC, Nutritional Requirements of Swine. Washington DC: National Academcic Press: 2012.
- Wensley, M.R.; et al. Maintaining continuity of nutrient intake after weaning. II. Review of post-weaning strategies. Translational Animal Science, 2021; 1. [Google Scholar] [CrossRef]
- Lallès, J.-P.; et al. Gut function and dysfunction in young pigs: physiology. Anim. Res. 2004, 53, 301–316. [Google Scholar] [CrossRef]
- Pluske, J.R.; Hampson, D.J.; Williams, I.H. Factors influencing the structure and function of the small intestine in the weaned pig: a review. Livestock Production Science 1997, 51, 215–236. [Google Scholar] [CrossRef]
- Fang, L.H.; et al. Effects of dietary energy and crude protein levels on growth performance, blood profiles, and nutrient digestibility in weaning pigs. Asian-Australas J Anim Sci. [CrossRef]
- Batson, K.L.; et al. Effects of feeding diets containing low crude protein and coarse wheat bran as alternatives to zinc oxide in nursery pig diets. Journal of Animal Science 2021, 99, skab090. [Google Scholar] [CrossRef]
- Williams, B.A.; Verstegen, M.W.A.; Tamminga, S. Fermentation in the large intestine of single-stomached animals and its relationship to animal health. NRR 2001, 14, 207. [Google Scholar] [CrossRef]
- Windey, K.; De Preter, V.; Verbeke, K. Relevance of protein fermentation to gut health. Mol. Nutr. Food Res. 2012; 1. [Google Scholar] [CrossRef]
- Gao, J.; et al. Protein Level and Infantile Diarrhea in a Postweaning Piglet Model. Mediators of Inflammation 2020, 2020, 1–15. [Google Scholar] [CrossRef]
- Wellock, I.J.; Fortomaris, P.D.; Houdijk, J.G.M.; Kyriazakis, I. The effect of dietary protein supply on the performance and risk of post-weaning enteric disorders in newly weaned pigs. Anim. Sci. 2006, 82, 327–335. [Google Scholar] [CrossRef]
- Bauer, E.; Metzler-Zebeli, B.U.; Verstegen, M.W.A.; Mosenthin, R. Intestinal gene expression in pigs: effects of reduced feed intake during weaning and potential impact of dietary components. Nutr. Res. Rev. 2011, 24, 155–175. [Google Scholar] [CrossRef]
- McCracken, B.A.; Spurlock, M.E.; Roos, M.A.; Zuckermann, F.A.; Gaskins, H.R. Weaning Anorexia May Contribute to Local Inflammation in the Piglet Small Intestine. The Journal of Nutrition 1999, 129, no. 3, pp. 613–619. [Google Scholar] [CrossRef]
- Xia, J.; et al. Research progress on diarrhoea and its mechanism in weaned piglets fed a high-protein diet. Animal Physiology Nutrition 2022, 106, 1277–1287. [Google Scholar] [CrossRef]
- Mayer, L. Mucosal immunity. Pediatrics 2003, 1595–1600. [Google Scholar] [CrossRef]
- Heo, J.M.; Kim, J.C.; Hansen, C.F.; Mullan, B.P.; Hampson, D.J.; Pluske, J.R. Feeding a diet with decreased protein content reduces indices of protein fermentation and the incidence of postweaning diarrhea in weaned pigs challenged with an enterotoxigenic strain of Escherichia coli1. Journal of Animal Science 2009, 87, 2833–2843. [Google Scholar] [CrossRef]
- Kim, J.C.; Heo, J.M.; Mullan, B.P.; Pluske, J.R. Efficacy of a reduced protein diet on clinical expression of post-weaning diarrhoea and life-time performance after experimental challenge with an enterotoxigenic strain of Escherichia coli. Animal Feed Science and Technology 2011, 170, 222–230. [Google Scholar] [CrossRef]
- Yue, L.Y.; Qiao, S.Y. Effects of low-protein diets supplemented with crystalline amino acids on performance and intestinal development in piglets over the first 2 weeks after weaning. Livestock Science 2008, 115, 144–152. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, J.; Wang, G.; Cai, S.; Zeng, X.; Qiao, S. Advances in low-protein diets for swine. J Animal Sci Biotechnol 2018, 9, 60. [Google Scholar] [CrossRef]
- García, K.E.; de Souza, T.C.R.; Landín, G.M.; Barreyro, A.A.; Santos, M.G.B.; Soto, J.G.G. Microbial Fermentation Patterns, Diarrhea Incidence, and Performance in Weaned Piglets Fed a Low Protein Diet Supplemented with Probiotics. FNS 2014, 5, 1776–1786. [Google Scholar] [CrossRef]
- Tian, Z.; et al. Influence of low protein diets on gene expression of digestive enzymes and hormone secretion in the gastrointestinal tract of young weaned piglets. J. Zhejiang Univ. Sci. B 2016, 17, 742–751. [Google Scholar] [CrossRef]
- Opapeju, F.O.; Rademacher, M.; Blank, G.; Nyachoti, C.M. Effect of low-protein amino acid-supplemented diets on the growth performance, gut morphology, organ weights and digesta characteristics of weaned pigs. Animal 2008, 2, 1457–1464. [Google Scholar] [CrossRef]
- Deng, D.; et al. Impaired translation initiation activation and reduced protein synthesis in weaned piglets fed a low-protein diet. The Journal of Nutritional Biochemistry 2009, 20, 544–552. [Google Scholar] [CrossRef]
- Yu, D.; Zhu, W.; Hang, S. Effects of low-protein diet on the intestinal morphology, digestive enzyme activity, blood urea nitrogen, and gut microbiota and metabolites in weaned pigs. Archives of Animal Nutrition, 2019; 4. [Google Scholar] [CrossRef]
- Marchetti, R.; et al. Protein Content in the Diet Influences Growth and Diarrhea in Weaning Piglets. Animals, 7952; 5. [Google Scholar] [CrossRef]
- Rattigan, R.; Sweeney, T.; Vigors, S.; Rajauria, G.; O’Doherty, J.V. Effects of reducing dietary crude protein concentration and supplementation with laminarin or zinc oxide on the faecal scores and colonic microbiota in newly weaned pigs. J Anim Physiol Anim Nutr 2020, 104, no. 5, pp. 1471–1483. [Google Scholar] [CrossRef]
- Wang, Y.; et al. Dietary crude protein time-dependently modulates the bacterial community and metabolites and changes dietary nutrient efficiency in growing pigs. Animal Nutrition 2024, 17, 1–10. [Google Scholar] [CrossRef]
- Zhang, C.; Yu, M.; Yang, Y.; Mu, C.; Su, Y.; Zhu, W. Differential effect of early antibiotic intervention on bacterial fermentation patterns and mucosal gene expression in the colon of pigs under diets with different protein levels. Appl Microbiol Biotechnol 2017, 101, 2493–2505. [Google Scholar] [CrossRef]
- Fan, P.; Liu, P.; Song, P.; Chen, X.; Ma, X. Moderate dietary protein restriction alters the composition of gut microbiota and improves ileal barrier function in adult pig model. Sci Rep 2017, 7, 43412. [Google Scholar] [CrossRef]
- Rattigan, R.; Sweeney, T.; Maher, S.; Ryan, M.T.; Thornton, K.; O’Doherty, J.V. Effects of reducing dietary crude protein concentration and supplementation with either laminarin or zinc oxide on the growth performance and intestinal health of newly weaned pigs. Animal Feed Science and Technology 2020, 270, 114693. [Google Scholar] [CrossRef]
- Moreira, T.G.; et al. Dietary protein modulates intestinal dendritic cells to establish mucosal homeostasis. Mucosal Immunology 2024, S1933021924000606. [Google Scholar] [CrossRef]
- Nyachoti, C.M.; Omogbenigun, F.O.; Rademacher, M.; Blank, G. Performance responses and indicators of gastrointestinal health in early-weaned pigs fed low-protein amino acid-supplemented diets1. Journal of Animal Science 2006, 84, 125–134. [Google Scholar] [CrossRef]
- Ferronato, G.; Prandini, A. Dietary Supplementation of Inorganic, Organic, and Fatty Acids in Pig: A Review. Animals 2020, 10, 1740. [Google Scholar] [CrossRef]
- Nowak, P.; Zaworska-Zakrzewska, A.; Frankiewicz, A.; Kasprowicz-Potocka, M. The effects and mechanisms of acids on the health of piglets and weaners – a review. Annals of Animal Science 2021, 21, 433–455. [Google Scholar] [CrossRef]
- Tugnoli; Giovagnoni; Piva; Grilli, “From Acidifiers to Intestinal Health Enhancers: How Organic Acids Can Improve Growth Efficiency of Pigs. Animals 2020, 10, 134. [CrossRef]
- Schönfeld, P.; Wojtczak, L. Short- and medium-chain fatty acids in energy metabolism: the cellular perspective. Journal of Lipid Research 2016, 57, 943–954. [Google Scholar] [CrossRef]
- Tsiloyiannis, V.K.; Kyriakis, S.C.; Vlemmas, J.; Sarris, K. The effect of organic acids on the control of porcine post-weaning diarrhoea. Research in Veterinary Science 2001, 70, 287–293. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Chang, R.B.; Allgood, S.D.; Silver, W.L.; Liman, E.R. A TRPA1-dependent mechanism for the pungent sensation of weak acids. Journal of General Physiology 2011, 137, 493–505. [Google Scholar] [CrossRef]
- Hutchens, W.M.; et al. The effects of pharmacological levels of zinc, diet acidification, and dietary crude protein on growth performance in nursery pigs. Journal of Animal Science 2021, 99, skab259. [Google Scholar] [CrossRef]
- Su, Y.; Yao, W.; Perez-Gutierrez, O.N.; Smidt, H.; Zhu, W.-Y. Changes in abundance of Lactobacillus spp. and Streptococcus suis in the stomach, jejunum and ileum of piglets after weaning: Changes in the gastrointestinal microbiota of the weaning piglet. FEMS Microbiology Ecology 2008, 66, 546–555. [Google Scholar] [CrossRef]
- Schubert, M.L.; Peura, D.A. Control of Gastric Acid Secretion in Health and Disease. Gastroenterology 2008, 134, 1842–1860. [Google Scholar] [CrossRef]
- Wang, L.F.; Bergstrom, J.R.; Hahn, J.D.; Young, M.G.; Zijlstra, R.T. Acid-binding capacity of feed in swine nutrition. Animal Feed Science and Technology 2023, 295, 115519. [Google Scholar] [CrossRef]
- Hansen, C.F.; Riis, A.L.; Bresson, S.; Højbjerg, O.; Jensen, B.B. Feeding organic acids enhances the barrier function against pathogenic bacteria of the piglet stomach. Livestock Science 2007, 108, 206–209. [Google Scholar] [CrossRef]
- Roth, F.; Kirchgessner, M. Organic acids as feed additives for young pigs:Nutritional and gastrointestinal effects. J. Anim. Feed Sci. 1998, 7, 25–33. [Google Scholar] [CrossRef]
- Li, Z.; Yi, G.; Yin, J.; Sun, P.; Li, D.; Knight, C. Effects of Organic Acids on Growth Performance, Gastrointestinal pH, Intestinal Microbial Populations and Immune Responses of Weaned Pigs. Asian Australas. J. Anim. Sci 2008, 21, 252–261. [Google Scholar] [CrossRef]
- Kuang, Y.; et al. Effects of dietary combinations of organic acids and medium chain fatty acids as a replacement of zinc oxide on growth, digestibility and immunity of weaned pigs. Animal Feed Science and Technology 2015, 208, 145–157. [Google Scholar] [CrossRef]
- Mroz, Z.; Jongbloed, A.W.; Partanen, K.H.; Vreman, K.; Kemme, P.A.; Kogut, J. The effects of calcium benzoate in diets with or without organic acids on dietary buffering capacity, apparent digestibility, retention of nutrients, and manure characteristics in swine. Journal of Animal Science 2000, 78, 2622. [Google Scholar] [CrossRef]
- Ferrara, F.; Tedin, L.; Pieper, R.; Meyer, W.; Zentek, J. Influence of medium-chain fatty acids and short-chain organic acids on jejunal morphology and intra-epithelial immune cells in weaned piglets. J Anim Physiol Anim Nutr 2017, 101, 531–540. [Google Scholar] [CrossRef]
- Li, S.; et al. Supplementation with organic acids showing different effects on growth performance, gut morphology and microbiota of weaned pigs fed with highly or less digestible diets. Journal of Animal Science 2018. [CrossRef]
- Long, S.F.; et al. Mixed organic acids as antibiotic substitutes improve performance, serum immunity, intestinal morphology and microbiota for weaned piglets. Animal Feed Science and Technology 2018, 235, 23–32. [Google Scholar] [CrossRef]
- Zeng, X.; et al. Dietary butyrate, lauric acid and stearic acid improve gut morphology and epithelial cell turnover in weaned piglets. Animal Nutrition 2022, 11, 276–282. [Google Scholar] [CrossRef]
- Namkung, H.; Gong, M.L.J.; Yu, H.; Cottrill, M.; de Lange, C.F.M. Impact of feeding blends of organic acids and herbal extracts on growth performance, gut microbiota and digestive function in newly weaned pigs. Can. J. Anim. Sci. 2004, 84, 697–704. [Google Scholar] [CrossRef]
- Pluske, J.R.; Turpin, D.L.; Sahibzada, S.; Pineda, L.; Han, Y.; Collins, A. Impacts of feeding organic acid-based feed additives on diarrhea, performance, and fecal microbiome characteristics of pigs after weaning challenged with an enterotoxigenic strain of Escherichia coli. Translational Animal Science 2021, 5, txab212. [Google Scholar] [CrossRef]
- Diao, H.; Jiao, A.R.; Yu, B.; Mao, X.B.; Chen, D.W. Gastric infusion of short-chain fatty acids can improve intestinal barrier function in weaned piglets. Genes Nutr 2019, 14, 4. [Google Scholar] [CrossRef]
- Canibe, N.; Steien, S.H.; Overland, M.; Jensen, B.B. Effect of K-diformate in starter diets on acidity, microbiota, and the amount of organic acids in the digestive tract of piglets, and on gastric alterations. Journal of Animal Science 2001, 79, 2123. [Google Scholar] [CrossRef]
- Magan, N.; Aldred, D. Post-harvest control strategies: Minimizing mycotoxins in the food chain. International Journal of Food Microbiology 2007, 119, 131–139. [Google Scholar] [CrossRef]
- Melin, P.; Sundh, I.; Håkansson, S.; Schnürer, J. Biological preservation of plant derived animal feed with antifungal microorganisms: safety and formulation aspects. Biotechnol Lett 2007, 29, 1147–1154. [Google Scholar] [CrossRef]
- Pierron, A.; Alassane-Kpembi, I.; Oswald, I.P. Impact of mycotoxin on immune response and consequences for pig health. Animal Nutrition, 6820. [Google Scholar] [CrossRef]
- Maher, S.; et al. Organic acid preservation of cereal grains improves grain quality, growth performance, and intestinal health of post-weaned pigs. Animal Feed Science and Technology 2024, 116078. [Google Scholar] [CrossRef]
- Jokiniemi, T.; Jaakkola, S.; Turunen, M.; Ahokas, J. Energy consumption in different grain preservation methods. Agronomy Research 2014, 12, 81–94. [Google Scholar]
- Jokiniemi, H.T.; Ahokas, J.M. Drying process optimisation in a mixed-flow batch grain dryer. Biosystems Engineering 2014, 121, 209–220. [Google Scholar] [CrossRef]
- Xu, X.; et al. A comparison of the nutritional value of organic-acid preserved corn and heat-dried corn for pigs. Animal Feed Science and Technology 2016, 214, 95–103. [Google Scholar] [CrossRef]
- Connolly, R.; Sweeney, T.; Maher, S. Organic acid and salt treatment of cereal at harvest improves growth performance in the post weaned pig. Animal-science proceedings 2022, 13, 204. [Google Scholar] [CrossRef]
- Dou, S.; et al. Characterisation of Early-Life Fecal Microbiota in Susceptible and Healthy Pigs to Post-Weaning Diarrhoea. PLoS ONE 2017, 12, e0169851. [Google Scholar] [CrossRef]
- Jha, R.; Fouhse, J.M.; Tiwari, U.P.; Li, L.; Willing, B.P. Dietary Fiber and Intestinal Health of Monogastric Animals. Front. Vet. Sci. 2019, 6, 48. [Google Scholar] [CrossRef]
- Nowland, T.L.; Kirkwood, R.N.; Pluske, J.R. Review: Can early-life establishment of the piglet intestinal microbiota influence production outcomes? Animal 2021, 100368. [Google Scholar] [CrossRef]
- Hamer, H.M.; Jonkers, D.; Venema, K.; Vanhoutvin, S.; Troost, F.J.; Brummer, R.-J. Review article: the role of butyrate on colonic function: REVIEW: ROLE OF BUTYRATE ON COLONIC FUNCTION. Alimentary Pharmacology & Therapeutics 2007, 27, 104–119. [Google Scholar] [CrossRef]
- Leonel, A.J.; Alvarez-Leite, J.I. Butyrate: implications for intestinal function. Current Opinion in Clinical Nutrition and Metabolic Care 2012, 15, 474–479. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, C.; Jiang, Q.; Yin, Y. Butyrate in Energy Metabolism: There Is Still More to Learn. Trends in Endocrinology & Metabolism 2021, 32, 159–169. [Google Scholar] [CrossRef]
- Geng, T.; et al. Probiotics Lactobacillus rhamnosus GG ATCC53103 and Lactobacillus plantarum JL01 induce cytokine alterations by the production of TCDA, DHA, and succinic and palmitic acids, and enhance immunity of weaned piglets. Research in Veterinary Science 2021, 137, 56–67. [Google Scholar] [CrossRef]
- Siddiqui, M.T.; Cresci, G.A. The Immunomodulatory Functions of Butyrate. JIR 2021, 14, 6025–6041. [Google Scholar] [CrossRef] [PubMed]
- Guilloteau, P.; Martin, L.; Eeckhaut, V.; Ducatelle, R.; Zabielski, R.; Van Immerseel, F. From the gut to the peripheral tissues: the multiple effects of butyrate. Nutr. Res. Rev. 2010, 23, 366–384. [Google Scholar] [CrossRef] [PubMed]
- Piva, A.; Morlacchini, M.; Casadei, G.; Gatta, P.P.; Biagi, G.; Prandini, A. Sodium butyrate improves growth performance of weaned piglets during the first period after weaning. Italian Journal of Animal Science 2002, 1, 35–41. [Google Scholar] [CrossRef]
- Feng, W.; et al. Sodium Butyrate Attenuates Diarrhea in Weaned Piglets and Promotes Tight Junction Protein Expression in Colon in a GPR109A-Dependent Manner. Cell Physiol Biochem 2018, 47, 1617–1629. [Google Scholar] [CrossRef]
- Biagi, G.; Piva, A.; Moschini, M.; Vezzali, E.; Roth, F.X. Performance, intestinal microflora, and wall morphology of weanling pigs fed sodium butyrate1. Journal of Animal Science 2007, 85, 1184–1191. [Google Scholar] [CrossRef]
- Jang, Y.D.; Lindemann, M.D.; Monegue, H.J.; Monegue, J.S. The effect of coated sodium butyrate supplementation in sow and nursery diets on lactation performance and nursery pig growth performance. Livestock Science 2017, 195, 13–20. [Google Scholar] [CrossRef]
- Nugent, A.P. Health properties of resistant starch. Nutrition Bulletin 2005, 30, 27–54. [Google Scholar] [CrossRef]
- Desai, M.S.; et al. A Dietary Fiber-Deprived Gut Microbiota Degrades the Colonic Mucus Barrier and Enhances Pathogen Susceptibility. Cell 2016, 167, 1339–1353e21. [Google Scholar] [CrossRef]
- Heo, J.M.; Agyekum, A.K.; Yin, Y.L.; Rideout, T.C.; Nyachoti, C.M. Feeding a diet containing resistant potato starch influences gastrointestinal tract traits and growth performance of weaned pigs1. Journal of Animal Science 2014, 92, 3906–3913. [Google Scholar] [CrossRef]
- Yi, S.-W.; et al. Raw potato starch diet supplement in weaned pigs could reduce Salmonella Typhimurium infection by altering microbiome composition and improving immune status. Front. Vet. Sci. 2023, 10, 1183400. [Google Scholar] [CrossRef]
- Trachsel, J.; Briggs, C.; Gabler, N.K.; Allen, H.K.; Loving, C.L. Dietary Resistant Potato Starch Alters Intestinal Microbial Communities and Their Metabolites, and Markers of Immune Regulation and Barrier Function in Swine. Front. Immunol. 2019, 10, 1381. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, S.K.; Nyachoti, C.M.; Krause, D.O. Raw potato starch in weaned pig diets and its influence on postweaning scours and the molecular microbial ecology of the digestive tract1. Journal of Animal Science 2009, 87, 984–993. [Google Scholar] [CrossRef] [PubMed]
- Yi, S.-W.; et al. Effect of feeding raw potato starch on the composition dynamics of the piglet intestinal microbiome. Anim Biosci 2022, 35, 1698–1710. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Qian, K.; Wang, C.; Wu, Y. Roles of Probiotic Lactobacilli Inclusion in Helping Piglets Establish Healthy Intestinal Inter-environment for Pathogen Defense. Probiotics & Antimicro. Prot. 2018, 10, 243–250. [Google Scholar] [CrossRef]
- Pieper, R.; et al. Effect of Lactobacillus plantarum on intestinal microbial community composition and response to enterotoxigenic Escherichia coli challenge in weaning piglets. Livestock Science 2010, 133, 98–100. [Google Scholar] [CrossRef]
- Su, W.; Gong, T.; Jiang, Z.; Lu, Z.; Wang, Y. The Role of Probiotics in Alleviating Postweaning Diarrhea in Piglets From the Perspective of Intestinal Barriers. Front. Cell. Infect. Microbiol. 2022, 12, 883107. [Google Scholar] [CrossRef]
- Sultana, R.; McBain, A.J.; O’Neill, C.A. Strain-Dependent Augmentation of Tight-Junction Barrier Function in Human Primary Epidermal Keratinocytes by Lactobacillus and Bifidobacterium Lysates. Appl Environ Microbiol 2013, 79, 4887–4894. [Google Scholar] [CrossRef]
- Li, X.-Q.; et al. Risks Associated with High-Dose Lactobacillus rhamnosus in an Escherichia coli Model of Piglet Diarrhoea: Intestinal Microbiota and Immune Imbalances. PLoS ONE 2012, 7, e40666. [Google Scholar] [CrossRef]
- Dehghani, N. INTERACTIVE IMPACTS OF DIETARY ORGANIC ACIDS AND CRUDE PROTEIN LEVELS ON PERFORMANCE AND GUT MORPHOLOGY OF BROILER CHICKENS. Book of Abstracts 2012.
Figure 1.
The potential synergistic effects of reduced dietary crude protein, organic acid and butyrate on post-weaned pig growth and health.
Figure 1.
The potential synergistic effects of reduced dietary crude protein, organic acid and butyrate on post-weaned pig growth and health.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



