Submitted:
02 December 2024
Posted:
03 December 2024
You are already at the latest version
Abstract
Herein, we report the presence of a plant paleocommunity dominated by ferns of the family Osmundaceae embedded in siliceous chert blocks from the only known Mesozoic, fossiliferous geothermal deposits, from the La Matilde Formation (Middle-Upper Jurassic), in the Deseado Massif, southern Patagonia, Argentina. A total of 13 blocks sampled in an area of approximately 250 m2, which includes multiple structurally preserved, monotypic, in-situ, rhizomatous stems of Osmundaceae, embedded within their original swampy substrate are documented. Additional Osmundaceae and other ferns, and conifers are present in chert levels adjacent to the previous blocks. This mimics similar monotypic associations of Osmundaceae in high-altitude lagoons in northeastern Argentina in the middle of the Paraná Forest, suggesting that the fern assemblage found occupied similar open spaces in the Jurassic, becoming preserved in analogous configurations. In addition, observation of the preserved fern tissues and surrounding substrate revealed a variety of interacting microorganisms, which are comparable to those that make up the microbiological communities inhabiting modern osmundaceous environments. This finding reveals a case of ecological stasis, where members of the same fern family separated by more than 150 million years formed similar exclusive groupings of individuals in microecosystems of comparable structure and general characteristics.
Keywords:
Osmunda
; wetlands
; Deseado Massif
; Patagonia
; ecological stasis
1. Introduction
The fossil record represents only a fraction of the total biodiversity that has existed through the geological time [1]. It is inherently biased and conditioned by the characteristics of the organisms and the depositary environments in which they were preserved, both before and after fossilization [1]. Notwithstanding, the fossil record provides qualitative and quantitative information for the reconstruction of the evolution of life and of its host environments through time at different hierarchical scales [1].
At the group level, the fossil record of plants is highly variable due to differences in taphonomic processes [2,3]. Some groups are well-represented temporally and spatially, whereas others lack known fossils [4,5]. These situations greatly influence the accuracy to trace their evolution and to reconstruct their paleoenvironments [4,5].
The preservation of organisms as fossils may occur in different forms and continues to be influenced by external factors up to the moment of discovery and collection [2,3]. Therefore, the mode and taphonomic history of preservation determine the quality of the information inferable from a given fossil [2,3]. Typically, only parts of an organism are preserved, and it is rare to find whole plants [2,3]. In particular, plants are mostly known in the geological record based on isolated parts and sometimes by a couple of organically connected organs [5]. Even in these situations preservation is somewhat incomplete when considering the internal structures of the plants (e.g., organs, cells, organelles). A few very rare examples of exceptional preservation of plants, having even some of their organelles preserved, are known [6]. These unique situations, such as in a lagerstatten, provide exceptional scenarios for the reconstruction of organisms, their structure, habits, interactions, and the paleoenvironments they inhabited [6]. In other instances, reconstruction of whole plants, including their structure, habit, and interactions has been possible by the combined study of isolated and organically connected organs [7].
Examples of exceptionally preserved organisms have extensively shown their more robust bearing as type cases for ecological and evolutionary studies, such as of some plant groups [8,9]. However, even in cases of exceptional preservation, broader ecological aspects of biological communities, such as spatial arrangement, physical structuring, and use of morphspace through time, are only possible to examine in uniquely preserved paleoenvironments. Another challenge in interpreting ancient plant communities is the taxonomic compositional differences between these ancient ecosystems and modern equivalents [3], which can complicate the reconstruction of entire ecosystems and limit the precise comparisons with modern environments.
The possibility of more precise comparisons with respect to modern environments is more likely for plant lineages with long fossil records [10]. Recognition of these unique paleontological situations in the fossil record are most unusual depending upon multiple factors, including the preservation of the organisms in life-position within their original substrate, and provide a unique opportunity to reconstruct the ecological history of biotic communities and their ecosystems through time [3]. In the geological record, examples of this can be detected from the conservation of community structure and morphological characteristics of taxa with modern representatives within their original paleoecosystems and denote certain cases of ecological stasis [11]. Ecological stasis refers to those moments in the paleontological record where biotic communities remain ecologically and morphologically stable through some periods of time [11]. In those cases, evolutionary processes are centered on the ecosystem organization resulting from the structuring of biological communities, which as such show a resilient tendency to face disturbances [11]. This is, selection via environmental processes on populations is at the same time constrained by their influence at the ecosystem scale, where its effect is less prominent, deriving in higher ecological stability. The achieved equilibrium should not be assumed as a steady state, but rather one where different expressions of analogous conformations for a given community replace each other through time, which are at the same time a reflection of its biological complexity [12]. However, stability moments are usually interrupted by periods where the opposite is the case, and populations and species are subject to disturbances that drive their change or evolution to eventually their extinction or to their modern forms [11].
The family Osmundaceae is a stereotypical case of conserved morphological and ecological characteristics, especially since the Mesozoic [5,10,13]. Its general body plan, habit, and even apparently its genetic configuration are perhaps the relatively most conserved ones among ferns [6,8,14]. Its fossil record is abundant and constant since the Permian and numerous morphospecies have been described in Patagonia and worldwide [5,15,16,17]. It is a monophyletic clade that represents a distinctive line of evolution, whose abundance apparently shrunk after the Mesozoic and gave rise to new forms during the Cenozoic, but which remained overall similar morphologically throughout [5,18]. The observed history of morphological and molecular stability for Osmundaceae based on its extensive fossil record is also apparent for its ecology [5]. Osmundaceous fossils have been described from wet, warm to temperate paleoenvironments, which is also how generally their modern habitats are, although variable with respect to their distribution and other environmental aspects [5,19]. However, the majority of the research papers carried out have focused on taxonomic studies of single organs and on phylogenetic analyses of selected fossil and extant species [14,20,21]. The study of whole fossil communities has been hindered by the lack of examples. As far as has been published, no examples of entirely preserved Osmundaceous communities, in-situ and in-life position are known from the fossil record. Herein, we describe a monotypic assemblage of ferns of the family Osmundaceae embedded in life within its original substrate from a Jurassic geothermal landscape in southern Patagonia, Argentina. This unique example provides information on the structure, composition, and network of interactions within a Jurassic swamp in Patagonia, which allows characterizing the paleoecosystem and evaluating its similarity to Osmunda-dominated, modern analogous ecosystems.
2. Results
2.1. Composition of the Main Chert Blocks (Includes Spatial Arrangement and Habit Within Individual Cherts and Relationships Among Chert Blocks)
Osmundaceous ferns in-situ preserved within 13 discrete chert blocks (up to 45 x 30 x 25 cm in size), holding up to 3 specimens together per block and around 21 individuals total occur exclusively distributed over an area of around 250 mts2 in the intact geothermal paleolandscape (Figure 1). Ferns are preserved embedded within the chert matrix, where mainly the rhizomatous stems together with their persistent leaf bases and associated roots attached to the original substrate persist in life position (Figure 2A-B; see S1-S3 in Supplementary material). These are accompanied by a few conifer remains, including a stump in life position with attached woods and other wood fragments, leafy branches, seeds, stems of Equisetum thermale, and numerous remains of other osmundaceae individuals that were scattered in the nearby area [23]. Within the mapped chert blocks occur from 1 to up to 3 ferns discernable, which vary in size (stem diameter 1.5 to 16 cm) and in their angle of orientation with respect to each other (Figure 2C–E). The angle of orientation within their respective substrates is variable and includes from almost horizontal to vertical individuals. Some larger stems of 7-14 cm of diameter and 10-15 petiole cycles have smaller, subordinate individuals attached laterally of 1.5-4 cm in diameter and 3-6 petiole cycles. Individual rhizomatous stems are characterized by having parenchymatous inner cortex and sclerotic outer zone clearly differentiated, stipulate petiole bases with a strong circular homogenous sclerotic ring surrounding the vascular bundle (Figure 2C–E). In addition, their vascular arrangement corresponds to a dictyoxylic siphonostele (Figure 2C–E). In particular, an ectophloic dictyostele with complete leaf gaps is observed and the leaf traces separate from the xylem cylinder with only one protoxylem cluster, whose division occurs in the outer cortex (Figure 2C–E). Further characteristics include one large mass of sclerenchyma near the ring plus several aligned masses in the stipular wings of the petiole bases (Figure 2C). These characteristics conform to the previously described morphotaxon Millerocaulis zamunerae from the same locality in the adjacencies of the assemblage described here [16].
The quantitative traits used to compare the fossils with the current specimens are summarized in Table 1. Figure 3 shows the Osmunda spectabilis populations from Misiones used for the comparison.
Finally, numerous dispersed sporangia, isolated or grouped, dehisced with none, a few, or completely filled with intact trilete or somewhat collapsed spores within, and also individual pinnae, were observed in the chert matrix (Figure 4A–D).
2.2. Associated Plants
In the nearby adjacencies of this main accumulation of fern-dominated chert blocks there are also further, identical, frequently fragmentary, osmundaceous fern fossils and a greater number of conifers [23]. These ferns are in general characterized by having a two-layered cortex, an ectophloic, dictyoxylic siphonostele with subexarch protoxylem, C-shaped leaf traces with a circular homogeneous sclerotic ring, an homogeneous parenchymatous pith, and a persistent mantle of winged petiole bases and diarch adventitious roots [23]. This combination of features place them within the Osmundoideae subfamily [16,23].
In addition, a couple of fern rhizomatous stems hold the presence of minuscule epiphyllous bryophytes. These have not been studied in detail, thus their systematic affinities are yet unknown.
Additionally, present in the chert matrix and associated with the tissues of the ferns preserved within the chert blocks there is a variety of fungi and other microorganism remains. Furthermore, the infected plant organs display a variety of decay and consumption patterns that variously affect their tissues, including modifications of the structure and appearance of the infected tissues, which sometimes also include the presence of coprolites.
In particular, in the interspaces between the fern petioles there are numerous, variously preserved, adventitious diarch roots, the best preserved of which show a variety of endobiotic, filamentous structures, which are distributed within the host cells (Figure 5A). The endobiotic filaments consist of hyphae that fill up some or most of the root’s cortex cells, occupying first the inner layers around the pericycle and from there extend outwards (Figure 5A,B). The filaments thin-out into highly ramified arbuscules that arise from wider trunk hyphae, and then profusely ramify into multiple tiny segments that occupy most of the space of the lumina of the colonized cells (Figure 5C). Some of the host roots are filled up with fungal material and may appear obscured by a “spongy” and opaque residue, whereas in other areas of the adventitious roots, cells are eroded or missing (Figure 5G). Included within the adventitious roots sometimes there are also further, broad, coenocytic hyphae, which are also obscured by the opaque deposits within the cortex of the roots, although they can be distinguished at different focal lengths, appearing somewhat coiled and occupying most of the lumina of the host cells (Figure 5H). Accompanying the hyphae are globose spores distributed also within the host root cells (Figure 5I). The globose structures are characterized by a uni- or bi-layered wall, of variable opacity, and by having a short, robust, subtending hypha (Figure 5J). In addition, attached to the outer surface of some of these degraded diarch roots via a single subtending hypha there are small, globose to ovoid, identical spores (Figure 5K).
The degraded tissues of the fern stems include filaments and globose structures of various shapes and sizes, which are variously arranged within the host cells or empty spaces created in the decayed fern stems. In particular, adventitious, diarch roots and especially petioles within the fern stems commonly appear degraded in various degrees, sometimes hollowed out and containing an uncountable number of globose, pyriform, and sometimes ameboid structures within the created spaces (Figure 6A,B). These structures are simple, variable with respect to their size (12 - 38 um), wall thickness (0.5 - 1.5 um), layering, and appearance, shape, and arrangement (Figure 6C). The smaller units are simpler and consist of rather spherical forms with no contents, nor further modifications observable on their surfaces (Figure 6C). Some of them occur as dispersed entities, individually discernable as float units in the inner space of the hollowed tissues, or they occupy the lumina of other globose structures and of healthy to degraded host cells, alone, in loose groups, or in tight clusters (Figure 6C–F). Larger forms (up to ~54 um) sometimes display 2-3 circular, annulate scars and/or papillae, flat, or arising from short necks, on their surfaces; a thicker wall, 1- to 2- or 3-layered; a basal apophysis or similar apparent rhizoids attached to the substrate; and a cytoplasm or lumen that in some cases appears granulated or as if divided into small globose to quadrangular parts (Figure 6G–K). Some of these, having psilate to faintly granulated surfaces appear somewhat collapsed and may contain smaller forms inside (Figure 7A,B). In addition, some thinner-walled, larger forms, having clear walls, appear as if they have one half or two quarters of their bodies missing, and cast profiles corresponding to a small plate-like bowl or a tote-like bag (Figure 7C). In other forms, parts missing of their bodies are shallower and correspond to smaller portions (Figure 7D). Some of these forms and other smaller ones with different characteristics sometimes are attached to the outer surface of similar or intact structures (Figure 7C–E). One to several of the smaller structures in other cases appear included within their lumina or cytoplasm of bigger forms (Figure 7C–E). Sometimes the inner globose structures appear also to have further papillae or protrusions that extend outwards and remain in contact with the wall of the bigger structures that contain them (Figure 7F).
In some of the degraded petiole and root tissues, and also associated with a decayed sporangium and unidentifiable plant debris, alone or along with the globose structures there are inter and intracellular filaments (~ 2 - 6 µm wide) , some of which are connecting adjacent cells to each other (Figure 8A–C). Some of the filaments are wider (9-13 µm) than others, and generally appear coenocytic, sometimes completely filling the lumen of some host cells (Figure 8D–E). Other filaments are more clearly septate, branched at right to acute angles and give rise to small, globose to quadrangular consecutive subunits (Figure 8F–I). Some other branched filaments observed sometimes give rise to one or two arms that narrow distally and may also be apically septate (Figure 8J,K). Some of these latter branched filaments are attached to the lumen of host cells via a wide circular base or appressorium and also appear regularly septate, forming quadrangular to cylindrical, unicellular segments (Figure 8L). Other filaments present in the lumina of some of the cells within the degraded tissues are completely opaque, of smaller size, and branch into secondary ones having a “rosary-like” pattern (Figure 8M).
The tissues occupied by the globose structures and filaments display a range of decay features consisting of broken, teared-apart, deformed cells, which sometimes have their lumen filled with opaque substances (Figure 9A). At greater magnification some cells appear to have their innermost cell wall layer detached from the remaining components and projected towards the center of their lumina (Figure 9B). Some other cells within similar decay areas have their middle lamellae differentially decayed and their cell wall layers appear deformed, and little to indistinguishable from each other (Figure 9C). In other areas, cells have all their wall components synchronously degraded, remaining mainly some parts of them, especially the middle-lamella at the corners of adjoining cells (Figure 9D). Further decay of all cell components is observed in even other areas, which creates cell-free areas where some globose structures and filaments are distributed (Figure 9E,F). In other petioles, decay spots have a “furry” or spongy texture (Figure 9G). Sometimes, these decay spots having a more regular shape appear concentrated in some zones of the decayed petioles, which also have other degraded or cell-free areas filled with globose structures (Figure 9H). A closer look at these decay spots shows a radiating, fusiform pattern, where cells appear progressively more decayed and eventually individually indistinguishable (Figure 9I–L).
In addition, within the matrix embedding the plant remains associated with various organic residues, filaments and globose structures, there are some degraded fern petioles whose inner tissues around the vascular trace and cortical area are bored and contain clusters of elliptical to cylindrical coprolites (Figure 10A,B). In addition, some small diarch roots are also almost completely degraded and filled with coprolites (Figure 10C). Fern petiole tissues display additional borings of different shapes, which are empty or filled with organic residues, along with some coprolites too (Figure 10D,E). Present in the spaces created by the bored tissues are globose structures and filaments, which are attached to the degraded tissues and sometimes also to the coprolites (Figure 10D–G). The coprolites present vary in size, shape, texture and apparent contents and are variously smudged, compressed and broken (Figure 10F–I). Some of those associated with the decayed vascular trace and inside the decayed cortical tissues are of rather homogeneous size (70 - 50 x 40 - 35 um), elliptical, approximately circular in cross section, and have a soft texture consisting of finely comminuted organic particles (Figure 10F,H). Some of the coprolites in the degraded cortical tissues of the petioles and diarch roots are bigger (180 - 140 x 80 - 90 um), somewhat quadrangular to cylindrical, form loose groups of fewer individuals, and have rough textures given by some more angular contents similar to plant remains (Figure 10G,I).
Further remains present in the degraded tissues, either in the empty spaces created within petioles and adventitious roots or encysted in the decayed tissues or their residues, correspond to more or less pyriform, to elliptical, to circular, to somewhat irregular and flatten structures, of variable size (115 - 70 um) (Figure 11A–F). In addition, these are characterized by complex n-layered, hyaline to opaque, psilate, reticulate, or ornamented walls (e.g.,short, broad, spines, or similar elements), sometimes having a broad, subtending hypha or apparent apertures (Figure 11A–F). Some of these structures have a central or so additional inclusion within their lumina (Figure 11E–G). In addition, smaller (30 - 65 um), thick-walled, opaque or hyaline, multi-celled, psilate, dispersed spores have been observed in the degraded tissues or within the lumina of host cells (Figure 11H–I).
In addition to the microorganisms and decay features of the fern stems, numerous chunks of wood preserved along with the latter in adjacent chert blocks or as isolated pieces show various biodeterioration patterns. These include general areas of decay interspersed along with xylem cells that appear sound, sometimes resulting in variable mottled patterns (Figure 12A,B). Included are spots where the tissues appear hyaline or less opaque than regular xylem cells, and irregular to circular areas where the tissues are missing, completely eroded away (Figure 12A,B). In the hyaline zones the cells are variously deformed, broken and torn apart, whereas the xylem in the borders of the completely degraded areas shows a progressive decay, appearing increasingly decayed and hyaline inwards (Figure 12C,D). Other areas show organic residues and opaque deposits distributed in irregular to discrete decay areas, sometimes arranged in somewhat parallel lines that include the ray cells and regular tracheids (Figure 12E–G). In these later zones the opaque deposits occupy the lumina of sound cells (Figure 12H). In addition, the opaque deposits sometimes are concentrated at the growth rings, around which the xylem appears partly to completely degraded (Figure 12I).
In some of the woods present there are also numerous traumatic resin ducts variable with respect to their size and appearance, which are placed immersed within the sound and degraded secondary xylem cells and aligned parallel to the growth rings (Figure 13A). The traumatic resin ducts are approximately ovoid to diamond shaped, have an ample lumen, empty or filled with an amorphous opaque content, and a ring of cylindrical to quadrangular, bulging, subsidiary cells, which can also look hyaline or somewhat opaque (Figure 13B,C). Continuous with the ring of cells, around the traumatic resin ducts there is further opaque material forming an amorphous mass, which may be organized in concentric lines in the adjacent xylem cells (Figure 13D). This mass of opaque substance extends further away into the surrounding xylem and is in direct contact with the degraded tissues, including some variously decayed areas and different growth rings, in turn connected to each other (Figure 13E–G). Together with the growth rings, the traumatic resin ducts form a tangential line of opaque deposits, around which different decay features are present (Figure 13G–H). Some zones around the latter have rather intact cells and others where they are variously decayed, including synchronous decay of all cell wall components or preferential decay of cellulose rich layers, resulting in their separation in individual parts (Figure 13I,J). In addition, in some of these traumatic resin ducts filled with massive amounts of an opaque substance there are clusters of globose structures embedded within it (Figure 13K). In other areas with more variable decayed xylem cells there are much smaller traumatic resin ducts consisting of a lumen the size of a single cell and surrounded by an incipient ring of subsidiary cells that look hyaline or variously opaque (Figure 13K–L). These sometimes appear clustered or closely together and an opaque substance appears also to have oozed out from a central nucleus and spread over the surrounding area (Figure 13 K–L).
2.3. Composition of the Main Chert Blocks (Include Spatial Arrangement and Habit Within Individual Cherts and Relationships Among Chert Blocks)
3. Discussion
We have described a unique fossil assemblage that preserves several biological and structural attributes of a paleoecosystem in a pristine configuration, which provides a very rare glimpse of Jurassic Patagonian landscapes. This complex case of preservation of an in situ and in life position community of osmundaceous ferns embedded together with their original substrate, the accompanying flora, and the associated microbiological communities there present allows for a multiple-way comparison to modern counterparts based on the identity of the ferns and other members of the greater plant community, trophic functional characterization of the microbes, and the general characteristics of the bearing-sediments. Overall, this consortium appears most closely comparable to similar osmundaceous fern communities present in modern lacustrine settings in terms of their structure, organization and composition. This suggests that despite geological and environmental changes that have happened since the Jurassic, the general characteristics of the ecosystem persisted through time and characterize similar modern osmundaceous ferns communities. This makes a call as to what attributes of the ecosystem can be postulated as having been key for the observed stability through time. In addition, understanding the characteristics of this ecosystem provides information about those features that have remained unchanged and which ones are comparatively different in terms of its composition, structure and ecological dynamics derived from biological interactions. In turn, this allows analyzing this example in terms of the concept of ecological stasis, which accounts for unique cases of morphological and ecological stability within flexible bounds through time [11,12,37].
Fossils described here consist of around 21 rhizomatous stems, which are variable with respect to their relative size and orientation within the siliceous matrix. In addition, some examples of dispersed empty sporangia, sometimes with collapsed spores, and pinnae are also present and these too occur embedded within the same bearing sediments. The variable shape and size displayed by the fern stocks suggests different developmental stages, whereas the presence of dispersed reproductive sexual structures is suggestive of generative and senescent life stages [38]. This arrangement is the result of taphonomic processes that started when the community was alive and, in turn, is an indication of the influence of the environment on the population dynamics [3,38,39]. Moreover, the fossil fern individuals appear tightly packed and sometimes appear bound together via lateral roots within the sediments, which probably represent clonal individuals. Current representatives of Osmundaceae reproduce vegetatively via the spread of lateral rhizomes that break apart into individual plants within the substrate, conforming to typical clonal populations (Figure 3E); this represents an adaptive advantage that favors horizontal spread within the occupied settings and, in turn, increases the probability of positive biotic interactions [40]. Although these are cases where slow growth rates were recorded, an age of 400 years has been calculated for populations of O. claytoniana growing through clones in humid and cold forests in western Virginia (USA) [41]. This highlights the importance of vegetative propagation for the maintenance of the species of this group [41].
Typical dynamics of the geothermal system could have influenced development of clonal growth organs in the jurassic osmundaceous community and, in particular, abundance of some (i.e., N) soil nutrients and scarcity of others (i.e., P), as have been suggested for other terrestrial settings, could have been most influential [42]. There is no evidence of nitrogen fixation for the geothermal soils where the osmundaceous community lived in the La Bajada locality during the Jurassic; although, based on the presence of potential nitrogen-fixing microorganisms in acidic, modern geothermal soils holding even higher temperatures than those in the later, a complete nitrogen cycle and, therefore, free availability of it is suggested [43]. Moreover, clonal development is suggested to represent a reproductive strategy that regulates the frequency of sexual vs asexual reproduction, through which influences the development of optimal life histories in unpredictable, disturbed ecosystems [44]. Formation of clones in the Jurassic landscape by the osmundaceae ferns would have provided them with a mechanism to cope with recurrent disturbance resulting from the geothermal system dynamics [45].
However, proportionally, the majority of the fossils found correspond to greater rootstocks that account for relatively more mature individuals. This could be a reflection of changing environmental conditions negatively affecting the osmundaceous community and of the consequent relative impoverishment of younger life stages leading to a senile community [38]. Within the greater paleolandscape further rhizomatous osmundaceous stems are found in an area exceeding 2 km around, which could imply different scenarios, including the possibility that additional swamp-like settings containing analogous communities were present. However, different to the specific assemblage described here, additional rhizomatous stems generally occur as dispersed units associated with geothermal wetlands where conifers and other ferns also abound [16,23]. Then, it also remains possible that migration of thermal fluids along the landscape triggered necessary environmental changes that positively influenced the osmundaceous fern community dynamics ending in the observed arrangements, which at the same time favored their preservation as fossils. In any case, external disturbances, such as via the effects of the migrating thermal waters, were suggested as having a direct influence on the structuring of osmundaceous communities [46].
Whether the direct influence of the geothermal system had a positive or negative effect on the consolidation of the osmundaceous community, some features of it could be suggested as having been a sign of a healthy ecosystem. This is, for example, the relative absence of trees within the nucleus of the osmundaceous paleocommunity, which is like what has been observed in modern assemblages, where open environments lacking plants producing shade are positively correlated with the development of generative life cycle stages and overall healthy populations of Osmunda regalis producing sexual structures in modern assemblages [38,39]. In these restricted settings of the Jurassic of Patagonia, several individuals of osmundaceous ferns, were distributed in an area of ~ 250 m2 in monotypic stands and got preserved in life position, where conifers, other plants, or their remains are absent or only sporadically observed. Additional plants present occur both, as larger decayed debris together with rhizome stocks and also larger intact forms, represented by trunks, stems, foliose branches, and reproductive structures of other ferns and conifers in the immediate surroundings of the pure osmundaceous assemblage [23]. In the greater paleo-geothermal settings of La Bajada locality in Patagonia, Osmundaceae also was widespread and is present in an area of 4 km2, along with an undetermined number of conifer species of the Araucariales [47,48,49] and other herbaceous ferns comparable to the Gleicheniaceae and Equisetaceae [16,23]. This shows that comparatively with modern settings, the taxonomic composition has varied. Whereas ferns and conifers made up the main plant groups during the Jurassic, modern ecosystems have a greater number of components, and, especially, of angiosperms, which, in fact, did not exist by when the fossil assemblage in Patagonia became preserved [5]. Also at an even greater scale, the Jurassic of Gondwana was dominated by conifers (i.e., Cupressaceae, Araucariaceae, Podocarpaceae and Cheirolepidiaceae [50], which formed forests of variable density, where herbaceous and shrubby vegetation belonging to different fern groups (e.g., Osmundaceae, Gleichenaceae, Dipteridaceae) and other gymnosperm groups (e.g., Cycadales and Bennettitales) thrived in understory, more or less open settings [50,51].
There are some examples today where ferns and conifers play a structuring role in the ecosystem and dominate some ecosystems over angiosperms. One such place corresponds to the wetlands or swamps of the Paraná Forest in Northeastern Argentina [52] (Figure 3A). They are small depressions that form more or less open vegetated lagoons at relatively higher elevations (~500 - 800 m.a.s.l.) on gently sloping land or paleochannels, which are generally inserted in the forest and feed by creeks of variable magnitude. They also are extremely important due their organic-rich substrates, which is manifested by the high degree of species richness and endemism. In these kinds of habitats, Osmundaceous ferns constitute persistent hydrophilous communities (Figure 3B) together with marsh plants and shrubs, surrounded by trees, including conifers of Araucariaceae (Araucaria angustifolia). These swamp settings are variable with respect to their more specific composition and some of them are formed by monotypic associations of osmundaceous ferns, or associations of osmundaceae with other ferns with a similar habit, such as Neoblechnum brasiliense (Desv.) Gasper & V.A.O. Dittrich (Blechnaceae) and Cyathea atrovirens (Langsd. & Fisch.) (Cyatheaceae) (AY observation). [38] pointed out the relevance of spring-fed swamps as the most apt settings to sustain viable osmundaceous communities. Moreover, presence of hydrophilous vegetation and low sedimentation rate typical of swamps are further characteristics of those communities. Most importantly, it has been postulated that independency and constancy of some environmental parameters that regulate species abundance are key factors for the stability of some modern osmundaceous communities, including acidic soil pH, water availability, reduced tree cover, which in turn was related to their distribution in higher relative elevations [38,39].
These modern, high elevation open swamps in Misiones, Argentina appear as the most closely comparable environments inhabited by Osmundaceae to those where the osmundaceous communities are presently preserved in the Jurassic geothermal landscapes of the Deseado Massif in Patagonia, Argentina. As in the modern swamps, the paleo-settings in Patagonia were dominated by monotypic stands of Osmundaceae, accompanied by ferns of other families and conifers. Evidence shows that the accompanying ferns and conifers were the second most important ecological groups of the ecosystem considering their relative abundance and distribution, inhabiting the fringes or the surroundings of the restricted environment within the greater landscape. This scheme of organization appears to have repeated itself in the geothermal landscape as a result of the influence of the migrating geothermal system, which might be analogous to the effect that the fluctuating inundation of the terrain, due to the natural dynamics of existing small scale fluvial systems in the Paraná Forest highlands, has on the development of modern osmundaceae-dominated, fern ecosystems in Misiones [52]. Further, low sedimentation rate, organic-rich substrates, but rather neutral pH, in a terrain that might have had swiftly undulating paleotopography seem to coincide with the characteristics of the Paraná Forest highland lagoons [22,45]. More specifically, the pH characterizing the osmundaceous settings likely varied between neutral to somewhat acidic in response to the dynamics of the migrating thermal waters, switching to the later during dryer periods and, back to the former, when inundation proceeded. No published data is available about that, but evidence of typical porous textures in the cherts preserving the osmundaceous ferns is consistent with this regime’s dynamics. In addition, preserved anatomical characteristics of the fossil Osmundaceae rhizomatous stems, which conform to the previously described morphospecies Millerocaulis zamunerae, are also generally like those of O. spectabilis presently adapted to wetlands, swamps, and similar environments [10,16,52]. The similarities include size and disposition of petioles and roots (Table 1), as well as the arrangement of sclerotic tissues, which form a continuous layer around them [10,16].
In addition, even though the Osmundaceae as a family is restricted to mild climates and wet biomes, distribution of morphotaxa within species varies with respect to elevation, mean annual temperature, temperature seasonality, and annual precipitation, and this in time is correlated with their morphological variation [13]. For example, morphotaxon ‘palustris’ of O. spectabilis characterizes high elevation swamps with high mean annual temperature and precipitation, such as in South America, including northern Argentina, whereas similar, stunted forms of O. regalis also are typical of high elevation swamps, but in Africa and regional islands, forms which have been suggested to be connected genetically [13]. There is not a direct measure of what the possible paleoelevation at the Patagonian geothermal settings was when the Osmundaceae monotypic assemblages were alive, but the abundance of araucarialean remains are suggestive of a hilly landscape. Moreover, field geological evidence of the La Bajada locality indicates that relative to other geofeatures of the landscape, including the presence of surrounding breccia complexes, the specific swamp-like settings that hold the pure osmundaceous communities occupied rather depressed areas within the landscape. Alternatively, comparisons in an environmental and climatic framework between the fossils described and the current Osmunda populations, are difficult to establish because the morphological characters used in extant analyses [13] are predominantly related to external morphology, principally of the fronds. Additional comparative anatomical studies within the genus are necessary to contribute to the discussion on paleobotanical assignments and the relationship of taxa to the environment.
Microbial Community
Evidence that members of the Osmundaceae paleocommunity engaged in mutualistic associations with glomeromycete fungi consist of individual diarch roots of variable size showing abundant arbuscules arising from narrowing hyphae distributed in the cortex. The roots show intracellular arbuscules in different states of development that arise from short fragments of trunk hyphae, which occupy most of the cellular lumen of the majority of the cortex plant cells, especially toward the center, around the pericycle. The presence of arbuscules in the small, diarch roots of the rhizomatous osmundaceous stems is the physical proof consistent with the formation of endomycorrhizae in plant-fungi mutualistic interactions [53]. Further evidence of endomycorrhizae forming fungi is the presence of intrarhizal and external spores attached to the epidermal surface of the same diarch roots, and occurrence of broad, intracellular, coenocytic hyphae [54]. The spores are similar to those produced by different families within the glomeromycetes [54,55]. Their specific features (i.e., globose shape, 2-layered, single subtending hypha) generally conform to those of the Glomeraceae and Paraglomeraceae [55].
Formation of endomycorrhizae characterizes most plants and are fundamental for the distribution and structuring of plant communities in different environments [56]. This suggests that these associations were also very important for the Osmundaceae community in the Jurassic geothermal system in the Deseado Massif. Mycorrhizal associations of this kind are especially important in low-nutrient availability environments, and by engaging in this mutualistic association the osmundaceous paleocommunity most likely gained a key ecological asset that favored its presence within the swamp settings in the geothermal landscape [57]. In modern settings, ferns, and especially old terrestrial lineages, such as Osmundaceae engage in equivalent mutualistic association with glomeromycete fungi [58,59]. The origin of plant-fungal endomycorrhizal association dates back to the Devonian, during land colonization, and it was suggested to have had a key role for the establishment of the first plants on Earth [4,60]. Glomeromycete fungi are not the only ones that engage in analogous mutualistic associations with ferns, also Mucoromycotina, and, perhaps ascomycetes and basidiomycetes too [61,62]. However, glomeromycetes have become obligate symbionts with vascular plants, including ferns and, as such, they have evolutively accompanied the Osmundaceae for over 150 ma (Middle-Late Jurassic) and developed together a relationship that has not yet been clearly studied, but which seems fundamental for their mutual existence, at least based on the current record.
The arbuscule-bearing diarch roots of the osmundaceous ferns from the Jurassic of Patagonia show numerous arbuscules in different states of preservation, which based on their known periodicity of formation, restricted spatial distribution, and overall rapid turnover, suggests that active mature plants engaged in the formation of the observed endomycorrhizae [61,63]. Moreover, the fungi distribution within the cortex of their host roots is identical to that observed in their modern relatives, specifically in Osmunda spectabilis from swamps in the Paraná Forest highlands in Northeastern Argentina (Figure 1a). However, a possibility remains that the observed infection actually represents non-functional mycorrhizal associations, or simply, a facultatively one, and, perhaps, a sign of a different kind of interaction [61,64,65]. Some of the adventitious diarch roots present in the fossil specimens appear variously degraded, sometimes containing an amorphous residue in the place of arbuscules and cortical cells, which could be indicating naturally senescent endomycorrhizae or a different kind of interaction.
Some factors conditioning the effects of mycorrhizae formation include light and nutrient availability, in cases rebalancing the relationship towards a unilateral one where the fungal partner ends up taking up carbohydrates from the plant host, without any clear benefit for the later [66,67]. In addition, the effects of mycorrhizae formation are variable depending upon the specific host plant colonized, its habit, state of development and environmental conditions [68,69]. Moreover, the effects of endomycorrhizae formation might not be instantaneous, but appear stronger at a later stage of development, such as in clonal offshoots from mother plants themselves, having a wider ecological impact at the community level and, in turn, in ecosystem functioning [69,70]. The recovered fossil assemblage is composed of plants variable with respect to size, shape and distribution, which supports an active osmundaceous community, where most likely at least some of the individuals included were metabolically active and harboring glomeromycete fungi to form endomycorrhizae. Moreover, no anatomical signs of plant defense mechanisms (e.g., modifications of the cell wall, presence of secondary metabolites) expected for bio- or necrotrophic fungi-plant interactions are observed in the fungi-colonized root cells [71].
The status of the potential interactions in which the fungi and the individual osmundaceous ferns engaged based on the observed association may lie in other aspects, such as in the developmental stage and tendency to engage in symbiosis of the plant partner, and, especially, in some characteristics of the environment, such as the exploitation of the nutritional quality of the substrate and the limiting features of its physical characteristics [62,72]. With respect to this, considering the influence of the geothermal waters as a factor directly conditioning colonization of the landscape via the frequent input of different amounts and types of minerals and heated water, restrictive of normal growth and development, potential metabolic and, in turn, ecological gains via the formation of endomycorrhizae appears logical for the osmundaceous ferns in the Jurassic paleocommunity. This is supported by the observed thermotolerance obtained by grasses via the formation of endomycorrhizae and other plant- endophytic fungal partnerships, which by doing so improved their metabolic rates and extended their distribution deep within the modern geothermal landscapes [73,74]. Then, formation of endomycorrhizae by the Jurassic osmundaceous community could have in turn positively influenced resource acquisition and allocation, promoted clone formation, greater areal distribution, and increased biomass growth in the geothermal system, and, in this way, provided a means to cope with the rapid environmental turnover typical of the dynamics of the geothermal system [45,69,75].
To what extent the influence of the geothermal setting affected the assembling of the osmundaceous community can only be predicted based on the observed association and, for this, it seems logical that tighter tolerance of the generated environmental conditions resulted from the formation of endomycorrhizae. Moreover, as it has been shown that preferential formation of endomycorrhizae is less likely in waterlogged soils and in aquatic settings in general, it can be suggested that the osmundaceous community likely occupied sufficiently drained soils, or at least some where this type of interaction was possible [62,76]. Indeed, modern examples of endomycorrhizae illustrated in this paper, formed by Osmunda spectabilis in the highland lagoons in the Paraná Forest correspond to small adventitious roots present normally at the water-air interphase and above (Figure 5D–F). The apparent preference for endomycorrhizae formation in Osmundaceae based on the current record supports the observed persistence of this kind of symbiosis in analogous modern similar settings [77]. As the family seems to have tracked down through time identical settings arguably having correspondent physico-chemical conditions, so have it accordingly maintained its symbiotic status, which likely compensated for its presence in swamp settings characterized by low-availability nutrient soils and oxygen, and periodically subject to the disturbing effects of geothermal and comparable fluvial settings [59,62,77]. In turn, this possible scenario suggests that the migrating geothermal waters and the consequent inundation of the communities adjacent to the system were key for their fossilization and entombment. This opens even the possibility that the endomycorrhizal plants were actually facultative ones, having sheltered latent fungal propagules within their roots, apt for developing a mutualistic relationship to cope with the periodically disturbing input of the geothermal waters on the soil conditions [78]. In this context, formation of endomycorrhizae and clonal growth organs by the Osmundaceous fern community in the Jurassic might have been key ecological assets for the observed resiliency of the ecosystem faced with various kinds of disturbances through time.
Other remarkably abundant microbial remains preserved within the osmundaceous fossil assemblage include morphologically and developmentally variable globose structures associated with the decayed fern stems, being especially abundant in decay spots resulting from hollowed petioles and roots. They occur in masses of individuals sometimes hardly discernible from each other and include various forms variable with respect to size, shape, contents, and ornamentation. Their morphology is rather simple and internal contents vary only a little too. Based on these features they can be grossly compared to different protists and fungi, including some testate amoeba, algae, peronosporomycetes, protozoa and chytridiomycetes and dispersed mitospores [79,80,81,82,83,84].
These fossils can be compared to some unicellular algae with globose to pyriform bodies (e.g., Chlamydomonas; Volvox), but which have two or more flagella that are not observed in the fossils from Patagonia [81]. Some of these algae are further comparable to the fossils based on the presence of papillae, rhizoids, and apophysis, although they are usually of bigger size, commonly display internal contents, especially chloroplasts, and rarely show an epibiotic habit on other individuals [81]. Some globose algae with life stages without flagella, such as Chlorella, Chlorococcum, Desmococcus, and Pleurococcus have a more regular size, are epibiotic and form biofilms of individuals joined by their lateral walls, or via cytoplasmic unions, and reproduce by binary fission, which are not features that have been observed in the fossils from Patagonia either [81]. Their habitats would fit well with that characterizing the Osmundaceous community, although they would rarely occupy hollow inner spaces where not much light would have arrived, such as it seems the case for the decayed fern roots and petioles [81]. To some extent the globose structures associated with the decayed osmundaceous ferns can also be compared to testate amoeba and protozoa, such as Arcella, Pelomyxa, Nassula and Porodon [82,85]. Especially so might be for those fossils for which a section of their bodies appears missing and have a granulated appearance, and papillae, rhizoids or an apophysis; however, those features, although rapidly comparable to apertures, ornamentation, and various appendages, such as uroids, pseudopodia, or lobopodia in testate amoeba and ciliate protozoa, they are morphologically different, usually symmetrically located, and more robust or rather inconspicuous [82,85,86]. In addition, the testate amoeba and protozoa are often of bigger size, thicker walled, comparatively opaque, and they usually have further modifications of their wall surface and interior that are not observed in the fossils from Patagonia [85,87]. A few of the fossils from Patagonia also appear amoeboid, having a body comparable to some other amoeba, such as Mayorella, but, from which they differ based on the lack of the same characteristics present in the former testate forms (e.g., pseudopodia, visible contents, such as organelles and further protist remains) [87]. In addition, the globose structures that are tightly packed as endobiotic inclusions in some of the fern host cells are comparable to cystosori of some typical plant-parasitic Plasmodiophorales (e.g. Membranosorus, Anisomyxa); however, other characteristics of members of this extant group are not observed for further comparisons [88]. The habitats where some of these protists live (freshwater settings, sometimes with marked regimes of flooding and drying seasons) are comparable to that inferred for the Osmundaceous community, which suggest that their may have been present there, but morphologically the fossils from Patagonia might be related to other groups of microorganisms [89]. For instance, some of the fossil globose structures, especially bigger examples containing a single or several additional propagules within their lumina are reminiscent of monogonial and polygonial, plerotic and aplerotic peronosporomycete (water molds, filamentous protists) oogonia, respectively [84]. However, oogonia could be ornamented, have a subtending hypha, and other attached hyphae remains from the reproduction process, which are not observed in the fossils [84]. Some unornamented oogonia, such as Pythium and Achlya are more similar to the fossil globose structures, although extant taxa have thicker, n-layered walls, which is different from the mostly uni-layered, simple wall of the fossil globose structures [90,91]. Fern host cells filled with small globose structures could also represent endobiotic oogonia produced by some peronospormycetes (e.g., Olpidium, Rozzela), but since there are no other directly associated remains relatable to the group (e.g., associated coenocytic hyphae, oospores), their origin within it seems unlikely [91]. Based on shape and size the globose structures are also reminiscent of dispersed extant and fossil mitosporic, fungal spores, although they differ in having more complex apertures, attachment structures, and habits [80,92].
Another group to which the fossil globose to pyriform structures may be related to are the Chytridiomycetes based on shape, size, habit, habitat, presence of incipient attachment structures comparable to rhizoids and apophyses of extant taxa, and openings, protrusions, or similar modifications of the wall reminiscent of discharge features also present in extant forms [79,93,94,95]. Size and shape wise, there are several zoosporangia of extant chytrids that are comparable to the fossils from Patagonia, such as different Rhizophydium and Chytriomyces [79,93]. Additional similar characteristics to zoosporangia of these taxa include the presence of papillae, pores, and tubes on the wall surface, through which zoospores are released, distribution in decaying plants in freshwater settings, and apparent host substrate, attaching tapering rhizoids [[79,93]. In addition, some globose structures with parts of their bodies missing look like dehisced Rhizophidium zoosporangia, whose deliquescent wall partly dissolves previous to zoospore release (e.g., R. megarrhizum) [79]. In other species (e.g., Rhizophydium vaucherii, R. patellarium), papillae or similar structures become detached and leave scars also comparable to those observed on the wall of the fossil globose structures [79]. These extant taxa are typical saprotrophs, present in different decaying organic remains in aquatic settings, and they can also occur as parasitic endobionts or epibionts on different organisms, including their own species, in a fashion comparable to some of the smaller fossil globose structures, epibiotic and endobiotic on bigger forms [79,93]. This mode of parasitism is unspecific and shows a tendency to rapidly gain access to suitable organic substrates, having several examples of chytrids parasitic on any organic remains, on the same species and on other taxa with which they share the same settings, such as Rhizophydium on Chytriomyces [79]. The extant parasites develop on the surface and inside the lumen of infected spores, pollen grains, zoosporangia, and similar propagules and, through a number of steps give rise to one or several individuals, which are reminiscent of the fossil globose structures with their lumina filled with a variable number of smaller forms [79,93,94,95]. Moreover, some of the fossil globose structures show their lumen granulated, sometimes as if it were parted into smaller fragments, which also look like some of the developmental stages of Rhizophydium, Chytriomyces, and other parasitic chytrids [79,93,95]. Other globose structures, with smaller individuals within their lumina, sometimes with discharge-like tubes, also look like some developmental stages of additional endobiotic chytrids that produce a single zoosporangium within a host zoosporangium, such as Rhizophydium globosum and R. sphaeroteca [79,93]. Also comparable to the development of encysted zoosporangia of necro and biotrophic chytridiomycetes are those fern host cells sometimes tightly packed with small globose structures [86]. In addition, a few of the globose structures have additional wall layers, appearing more robust and opaque, which are reminiscent of resting spores produced by the same and other extant chytrid genera [79,93].
The presence of putative zoosporangia associated to the fern rhizomatous stems is consistent with the diversity of chytrids present in modern aquatic settings [96]. Despite the apparent diversity present, morphologically, zoosporangia are simple structures having only a limited number of features available for their taxonomic assignment, which, especially in fossils, precludes precision for such a task, making them comparable to several extant forms, such as is for the numerous fossils associated with the decaying osmundaceous rhizomatous stems [79,93]. Moreover, several of the fossil zoosporangia show features consistent with different developmental states, which also supports affinities to different extant forms and, as such, an active chytrid community. Chytrids have been found as main decomposers of suspended particulate organic matter in freshwater settings, such as in swamps where different plants, including osmundaceous ferns are present [96]. Moreover, chytrid zoosporangia are present suspended in the chert matrix and also occur on and inside the tissues of petioles and roots of the osmundaceous stems and on each other, including from zoospores grouped inside zoosporangia to empty, dehisced forms, “floating” as variable senescent casts. This is an ecological characteristic of the chytrids, which guarantees completion of their life cycles after zoospore release and promotes their fundamental role as the main recyclers of particulate and dispersed organic compounds in aquatic settings [96].
The observed abundance of chytrids is consistent with acidic pHs and low salinity, and in turn with available organic remains and aerated substrates, suggesting shallow, freshwater wetlands or similar settings [97]. Alternatively, chytrid’s abundance might have depended more on micro-ecological conditions, such as on moist and organic-rich ones expected for the rhizomatous stems. The possible fossil chytrids are preferentially distributed in the inner spaces created as a result of the decay of the fern rhizomatous stem tissues, usually concealed within hollowed out petioles and adventitious roots. Reports of extant fungi associated with ferns describe examples of endomycorrhizae and saprotrophs, although chytridiomycetes associated with osmundaceous or similar complex rhizomatous stems have not been reported [98,99]. What has been reported are similar ecological strategies developed by other fungi and protists, which are preferentially distributed within inner, hollow spaces of decayed plant stems [100,101,102]. In these microhabitats, fungi and protists are buffered from external, rougher environmental conditions, and obtain an stable ecological niche that protects them from potential predators, and favors their distribution in a wider area [100,101,102]. The fungi in hollowed out plant organs in wetlands are drivers of organic matter breakdown and nutrient recycling, occupying a pivotal role essential for the dynamics of aquatic ecosystems [96]. In addition, ancient chytrid communities have been previously reported in similar hollowed out fern stems with root mantles, where they were associated also with organic matter breakdown, especially of fern tissues and other trapped plant litter, which could have aided ferns in nutrient acquisition [103]. In addition, apparent mutualistic fungi were also reported for the fern root mantles from these later examples, which is comparable also to the presence endomycorrhizae formed by glomeromycetes with adventitious roots distributed in the osmundaceous rhizomatous stems from Patagonia [103]. Additional examples of a comparable community of fungi, epiphytic plants and detritivorous microorganisms were described from early Jurassic and Eocene osmundaceous rhizomatous stems and their immediate surroundings [103,104]. This underscores the ecological significance as saprotrophs and parasites of the putative chytridiomycetes in the trophic structure of the environments characterizing the ecosystems where the Osmundaceae community thrived. It also supports previous indications that point out the importance of fern rhizomatous stems as particular ecological microniches sustaining diverse communities of meso and microorganisms through geological time. Moreover, recurrent association of the same type of organisms underscores the stability of the trophic structure and dynamics within the osmundaceous rhizome and suggests this as a feature that has fueled the observed resilience of this ecological microniche.
Furthermore, the decayed host fern stem tissues show numerous, morphologically variable filaments consistent with saprotrophic activity and other possible interactions. The filaments vary in size, ornamentation, shape, extension, septation, and opacity. Some of these are branched and show further modifications including apparent apical, consecutive, unicellular propagules, and some have a wider base through which they are attached to host cells. Based on these features, the filaments are like those produced by different mitosporic fungi, which correspond to asexually produced structures of ascomycetes, and less likely, basidiomycetes [80,106,107]. The morphological features of these are common in several fungal taxa, whose affinity is more clearly discernible based on sexual structures [108,109]. However, some of the observed branched filaments bear 1 or 2 distally narrowed and finely septate arms, which look like phialide or similar conidiophores produced by some ascomycete anamorphs, such as different Penicillium and Trichoderma species [110]. Species of these extant genera have different nutritional modes, such as saprotrophic on plant litter, which supports a similar role within the decaying osmundaceous remains [111]. They have also been detected as fern endophytes or opportunistic parasites, which become saprotrophic after host plant senescence and death, suggesting another possibility for the fossils [112,113]. Moreover, some of the filaments bearing phialide-like conidiophores are attached via a wide base to host cells, which are reminiscent of appressoria produced by some parasitic Penicillium [114]. There are also some filaments without morphological modifications that appear finely septate and bear what look like arthrospores, which are widespread among mitosporic fungi and are produced within host tissues under stressful conditions [109]. Additional, comparatively smaller, opaque, branched filaments present are also regularly septate and give rise to a thread of small units in a rosary-like pattern, which are morphologically comparable to conidiospores produced in aerial hyphae by some filamentous bacteria, such as Streptomyces, a widespread plant saprotroph [115,116]. Some of the endophytic filaments lack septa or these are too faint to be clearly discernible, and are wider, hyaline, and tightly fill in a contorted fashion the lumina of host cells. They are morphologically similar to those produced by some endomycorrhizal fungi within host cells [117]. This consortium of fungi and fungi-like propagules provides additional evidence of the trophic complexity of the microhabitat represented by the osmundaceous, fern rhizomatous stems. It shows that they possibly engaged in different kinds of interactions, but which certainly have a main role as saprotrophs, especially of fern tissues and other plant litter.
Formation of clonal growth organs by mother plants, such as in Osmundaceae, has been demonstrated to favor the development and spread via colonization of new propagules by mutualist and pathogenic fungi [118]. This strategy, where the amount and quality of plant litter is also affected by the production and turnover of extra plant organs via clones appears to positively influence saprotrophic fungal communities development as well, indirectly, as it increases the availability of edible organic resources [119]. This creates an advantageous ecological context for the fungi in general since it reduces the costs of finding new hosts, and is also of similar convenience for plant host partners in the case of mutualistic associations [119]. Such a mechanism of coherent and facilitated transmission of microorganism communities between plant generations has been suggested to represent a characteristic favoring resilience of plant communities faced to ecological disturbance [120]. Based on this, it has been suggested that host plants and fungal partners forming mycorrhizae have closely linked traits that likely favor their coexistence and association (Bitterbe et al., 2020). In the case of the Osmundaceous fern communities from the Jurassic of Patagonia, characterized also by the formation of clonal individuals, the analogous composition and deduced relationships between the plant hosts and the microorganisms partners can be postulated as another feature of the ecosystem that provided it with the capacity to effectively respond to disturbance, maintaining its trophic organization and dynamics through time (Vannier et al., 2018).
On the other hand, some of the fern tissues display a number of decay features, which were probably caused by the fungi and additional microorganisms and mesoorganisms present. Observed are teared-apart, broken, and deformed cells, which sometimes have their lumina filled with opaque substances. These are generally comparable to broken and deformed cells that result from fungal decay of wood and other woody tissues [121]. In particular, woody plants in aquatic settings, such as some ferns in swamps and wetlands are affected by soft-rot causing fungi, which carry out strategies adapted for decay under water saturated and low-oxygen conditions [121]. Decay by this group of fungi avoids lignin-rich cell wall components, preferentially degrading cellulose-based tissues, such as parenchymatous pith and cortex regions of fern stems, which remind of some of the fossil fern stems with decay areas present in the cortical region of the petioles and adventitious roots [121]. Moreover, soft-rot decay type preferentially decays the secondary wall components and creates a typical pattern that sometimes appears as pits of increasing size, ending in the detachment of the tertiary wall, which look like some of the generally eroded cells with their tertiary wall partly to completely detached from the remaining wall components, observed in cross section in decay areas interspersed with sound tissues [121]. These characterize areas or “decay spots” where, also sometimes no. or, only a few distorted cells and abundant filaments and globose structures are preserved. At lower magnification, some of the observed decay areas in the fern rhizomes have a spongy to furry texture and appear variously eroded, with cells deformed, broken and some with irregular opaque contents, comparable to that expected from soft-rot fungal decay and also from bacterial decomposition [122]. Moreover, at greater magnification the decayed tissues show cells with their walls generally thinned, sometimes from the lumen outwards, and others completely dissolved, or deformed and collapsed, having their primary wall and middle lamella partly broken or entirely degraded and separated from each other, which are features comparable to those produced by plant degrading fungi and bacteria [122]. In other fossil rhizomatous stems, individual cell wall components are indistinguishable from each other and only a pattern consisting of minute, radiating, translucid to somewhat opaque threads, or just an amorphous wall layer residue remains, which possibly represent some of the typical lignin and non-cellulosic byproducts of wood decay left by erosion bacteria that characteristically alters the texture of the degraded tissues [123,124]. Apparent bacterial degradation is consistent with decay under water saturated and low oxygen conditions [124]. Additional decayed roots and cortical zones, adjacent to more massive decay areas in the petioles, show cells with only their S2 and S3 cell wall components preserved, although deformed and separated from adjacent cells, which is consistent with preferential decay of lignin-rich units, such as by white-rot fungi [121]. Other cells show synchronous decay of all cell wall components, with cells remaining attached to each other only by remains of the middle-lamella at their corners, which is also how woody tissues degraded by white-rot fungi look like [121]. Some fossil fern cells show opaque contents within their lumina, which might correspond to metabolic remains from decay processes or ergastic compounds deposited as a means to deter decay spread [125]. White-rot decay is unique in its strategy because it preferentially degrades lignin-rich or synchronously all cell wall components, the S1 component, and the middle lamella, or these and the S2 and S3 too, minimizing competition for woody resources with other degraders, which thrive in well-aerated and moist media [121]. These characteristic decay features observed indicate an active and diverse community of saprotrophs adapted to various microenvironmental conditions that through different strategies variously altered the fern rhizomatous stems structure, which indirectly influenced each other and promoted further decay, analogous to how plant degrading bacteria and fungi synergistically predispose wood to fungal decay in some modern ecosystems [126]. This pivotal role in organic matter breakdown and nutrient cycling in swamp settings during the Jurassic carried out by some fungi and bacteria shows their engagement in various types of interactions with the community of Osmundaceous ferns, helping maintain a dynamic trophic equilibrium in the paleoecosystem [126].
In addition to possible protists, fungi and bacteria associated with the fern rhizomatous stems, evidence of mesoorganisms is based on the presence of degraded petioles and leaves, hollowed by irregular borings, some of which are filled with coprolites of 2 different kinds. Smaller coprolites are elliptical and of circular cross section, with somewhat rounded ends, formed of finely comminuted plant material, without distinguishable contents, of fine to rough textures, and are distributed in small clusters within irregularly bored leaves and petiole tissues, which are features that conform to those described for oribatid mite coprolites and feces of extant forms too [127,128,129]. Oribatid mites are typical Paleozoic and Mesozoic, scavenging, opportunistic, micro detritivores that irregularly bore and feed on live and dead plant organs and debris, including fern rhizomes, which suggests a similar ecological role in the decayed fern rhizomatous stems [130,131]. Additional, cylindrical to quadrangular, bigger coprolites with blunt endings and a rough texture resulting from the inclusion of apparent plant remains, present within the degraded tissues of a fern petiole, are closer to those of some coleoptera, especially some curculionids (Curculionidae) based on shape and contents [132,133,134]. These and other coleopteran groups typically attack and consume woody tissues of plants decayed by fungi, which is consistent with the mycoflora present in the decay fern rhizomatous stems [135]. Moreover, fungal decay along disintegration of the fern tissues by boring arthropods, such as coleopterans, possibly acted synergistically towards their decomposition, which is how these organisms interact in modern ecosystems [136]. They engage in variably dependent upon each other interactions, where wood-consuming coleopteran preferentially target substrates pre-conditioned by fungal decay, which in turn are positively benefited by being phonetically dispersed among potential hosts [137]. In terms of shape, they are comparable to the feces of some plant-consuming suspension-feeder, microinvertebrates from aquatic settings [138].
Additionally, widespread globose to flat spores of circular to somewhat irregular outline, encysted or trapped within the fern tissues or unidentifiable plant debris, characterized by complex walls formed by a variable number of layers, sometimes externally ornamented, and with a single or more apertures, reminiscent to peronosporomycete and algae sporangia are present [81,84]. Some of the encysted structures have a short extended neck or short subtending hypha, at least 4 wall layers, and possibly more, but which are otherwise difficult to distinguish from surrounding organic remains, These characteristics make these structures comparable to oogonia in different developmental states [81,84]. Moreover, an apparent central oospore is observed sometimes, further supporting their similarity to peronosporomycete oosporangia (Dick; 1990; 2001). In addition, some of them are reminiscent of previously found spiny oogonia of the Combresomycetales, which were suggested as saprotrophs and parasites [139]. Others, consisting of a flattened structure of circular outline and somewhat trapezoidal, blunt external ornamentation look like prasynophycean algae, resting spores, which were also previously found in the La Bajada locality [23]. Some, multicelled structures also present look like conidia or sclerotia produced by some anamorphic fungi, including some Helicomyces and similar helicoidal forms, which are found in freshwater settings as saprotrophs; others look like typical saprotrophic and parasitic mitosporic fungi, such as Hermatomyces [80,106,107].
In addition, embedded in the chert blocks or isolated within the area of the osmundaceous community there are some chunks of wood that show various features relatable to decay by fungi. Included are woods showing various mottled patterns consisting of irregular, completely or partially degraded areas of secondary xylem interspersed within sound tissues, which are typical of decay by white-rot fungi [121]. A closer view of the degraded areas show some where xylem cells are hyaline as a result of the decay of lignin-rich opaque components, which is consistent with differential decay also by white-rot fungi [121]. Xylem cells within these decay areas appear deformed, broken and torn apart, which is a reflection of loss of rigidity due to selective lignin decay by white-rot fungi [121]. Further decay, irregular and circular to ovoid, areas show no tissues preserved and, therefore, cells with all their cell components synchronously decayed, which is also consistent with white-rot fungi [121]. Sometimes decay is more discretely distributed over ray cells, which coincides with decay strategies followed by white-rot fungi degrading wood of extant trees [121]. Presence of wood with decayed areas having residual xylem cells embedded together with an opaque substance, looks like a necrotic zone, where deposition of ergastic compounds, such as secondary phenolic, resin products by modern plants used as fungal deterrents, occurs [140,141,142]. These putative ergastic deposits are sometimes more regularly oriented, following the axial, tangential and parallel orientation of growth rings, perhaps coinciding with latewood, thick-walled, fibers, or discrete lines in intercellular spaces between degraded xylem cells, which are reminiscent of the structured response to fungal invasion deployed by some modern trees, such as a reaction zone [125,143].
Another evidence of an active interaction responding to a triggering effect of biotic or abiotic origin is the formation of numerous axial traumatic resin ducts in the secondary xylem of some of the fossil woods present [144,145]. These are commonly produced in the secondary xylem of conifers of the family Pinaceae and sometimes also in the Taxodiaceae and Cupressaceae constitutively or as a result of different types of stimuli, such as invasion by pathogenic microorganisms and even arthropods [144,145,146,147]. The traumatic resin ducts are tangentially aligned with respect to the growth rings, forming discontinuous rows, and occur among wood with a mottle-rot pattern typical of white-rot fungi, which could have been the causal agents [148]. Indeed, an opaque substance massively spread over the decayed areas from the traumatic resin ducts, sometimes connecting with tracheids further aligned with the growth rings, forming one or more, continuous tangential barriers, which is consistent with it being deployed as a sealant against an invading agent [125,149]. Decay appears more massive on one of the sides delimited by the later resiniferous barriers emerging from traumatic resin ducts, which further supports their role as pathogen deterrents [125,149]. Within the degraded xylem, examples of variously deformed and broken cells, sometimes showing differential and selective decay by white-rot fungi are present [119]. Other xylem cells have signs of soft-rot decay, such as preferential decay of the S3 cell wall layer [119]. Among the degraded xylem cells also present are smaller, incipient traumatic resin ducts and rays filled with further opaque material, which is consistent with a dynamic response to pathogenic fungi [121,150,151]. These smaller traumatic resin ducts consist of a few cells and their presence also indicates metabolically active tissues in proximity to the cambial zone, where their formation involving chemical and anatomical evidence, likely responded to an ongoing external stimulus [125,145]. Moreover, globose structures of unknown affinity appear trapped inside some of the traumatic resin ducts and, these may correspond to fungal or protists propagules that might get caught in the mass of resin extruded by modern trees [151]. These fungally-attacked woods provide additional evidence of the complex network of interactions that characterized the ecosystem dominated by a community of osmundaceous ferns. It adds another component to the structure of the ecosystem, represented by typical decay and reaction processes between conifers and fungi present, which together with the pivotal ecological role of the ferns seems to have been key for its stability through time.
The associated microbiota described is a proof of the role as an ecological microniche that the sheathed rhizomatous stems of osmundaceae had, and gives a remarkable glimpse of the interactions that characterized the swamps settings where these plants inhabited during the Jurassic in Patagonia. This paper deals with functional groups and more precise taxonomic resolution is left for future papers. Despite, a community of fungi belonging to the glomeromycetes, and others of unknown affinity but more likely related to the chytridiomycetes and to the ascomycetes and, perhaps, to the basidiomycetes is present directly associated with the osmundaceous plants. They associate with each other in various fashions, occurring in different tissues, and having variable arrangements, which exemplify the intricacy and specificity of each association. Some of the interactions that can be rapidly deduced from the observed associations are also characteristics of the same communities and environments inhabited by extant Osmundaceae and other ferns [59,77,152,153]. These networks of interactions are some of the key features that control the dynamics and trophic structure of their respective ecosystems [71]. Their observation in the Osmundaceous community provides a point of reference that allows tracing the evolutionary paths followed by the parties involved in the different interactions present, and, therefore, provides a basis for hypotheses about their origin and historical ecological significance [154]. It adds to the existing body of information about biotic interactions between fungi and osmundaceous ferns in the geological record [104,105].
It has been previously shown that Osmundaceae represents in itself a conserved ecological microniche, which hosts a variety of saprotrophs, epiphytes, and detritivores within both, extant and fossil representatives [59,104,105,152,155,156]. Perhaps, this specialized ecological microniche and the accompanying biological microcommunity there formed accounts too for the observed resiliency of the macrocommunity formed along with the osmundaceous populations in swamp settings through time. This per se should not be thought of as something unexpectable, but as proof that demonstrates the ecological importance osmundaceous ferns have as hosts of diverse microbiological communities, both by providing a specific substrate, and also by forming part of a larger consortium of plants in swamp settings where various forms of fungi tend to inhabit. Reciprocally, this added complexity to the ecosystem constrains its structure and dynamics via a network of biological interactions, making it overall more stable, and likely to host ecologically consistent community members [71,157]. In other words, what the diversity of fungal taxa could be thought of as representing is a “buffer” of species that carry out analogous functions, which then can be “replaced” if any sort of ecological disturbance affects any of them in particular [158]. Moreover, in a pure hypothetical situation it can be assumed that what we see is a window of the fungal communities that accompanied the Osmundaceous ecosystem, which shared a nucleus of species that structured it and carry out functions that have remained through time and that accounts for the observed resilience of the system up to today, although, consisting of a different taxonomic composition.
4. Materials and Methods
4.1. Geological Context of the Studied Area
The fossil material was collected from outcrops of the La Matilde Formation at La Bajada locality in the Deseado Massif. The Deseado Massif is a geological province with an approximately 60000 km2 extension, located in the Santa Cruz Province of southern Patagonia, Argentina [22,23]. The geological history of the area is characterized by a bimodal rhyolitic and andesitic volcanic event that led to the formation of the Bahía Laura Complex, approximately 178–151 million years ago [24,25]. These rocks are part of the Chon Aike Silicic Large Igneous Province that extends from Argentinean Patagonia to Antarctica [26] and whose genesis is associated with the break-up of Gondwana and the opening of the South Atlantic Ocean [27,28].
The La Bajad locality is located in the western portion of the Deseado Massif Jurassic volcanic outcrops [23], and it is characterized by siliceous hot spring-related deposits hosted in reworked volcaniclastic sediments of the La Matilde Formation [23]. The rocks at La Bajada show the typical association of geothermal features with volcaniclastic fluvial or lacustrine reworked deposits and volcanic lava domes (lithological control proposed by [22]). A series of fossiliferous sedimentary chert beds outcrop irregularly in a 2.5 x 0.5 km area in close proximity to the La Bajada farmhouse [23]. These rocks vary from strongly silicified volcaniclastics to pure cherts deposited in distal, geothermally influenced marsh facies setting [23,29].
Preliminary analyses of chert samples from La Bajada hot-spring-chert deposits offered a first glimpse of an exceptionally preserved biota [23], composed of different groups of animals, plants, and a broad range of microorganisms [23,30,31]. Along the vascular plants, there are abundant vegetative and reproductive fern remains with different systematic affinities [23], including the rhizomatous stems assigned to the Osmundaceae species Millerocaulis zamunerae [16]; pinnules, sori and isolated sporangia of putative Gleicheniaceae, Cyatheaceae, and Dicksoniaceae [23]. Conifer vegetative and reproductive remains with araucarialean affinities are also common in these cherts, including wood fragments, roots, leafy twigs, and cones with pollen in situ [23,30,31]. Further plant remains include stems of Equisetum thermale, frequently fragmentary [23]. The microcommunities are characterized by different filamentous and colonial cyanobacteria; filamentous and unicellular algae; abundant oomycetes with combresomycetelean affinities; testate amoeba and ciliates; and very diverse and abundant fungi [23,30,31,32]. Different systematic and ecological groups of fungi are frequently associated with different plant remains, and are represented by abundant vegetative and reproductive remains [23], including epibiont chytrids, mycorrhizal glomeromycetes, epiphyllous ascomycetes, and a variety of ascomycetes and basidiomycetes [23,30,31,32]. Furthermore, remains of terrestrial and freshwater invertebrates are also present (e.g., appendages, mouthparts, abdomens and compound eyes of arthropods; ostracod and other crustaceans carapaces; gastropod shells; worms, rotifers, nematodes, and coprolites of various morphologies) [23].
4.2. Preparation and Imaging of Fossil Material
Several chert blocks were cut following standard techniques [33] and using two different Hillquist (Denver, USA) slab saw machines, one for larger rock samples and one for hand-size rock samples. Some blocks were cut in several sections, following the longitudinal axis of the plant individuals to obtain serial planes and ensuring the exposure of as much surface area as possible to observe the surrounding matrix. The chert blocks and their sections were observed using a Zeiss (Oberkochen, Germany) MC80DX and a Nikon (Tokyo, Japan) SMZ1000 stereoscope. Additionally, thin sections of some chert samples were prepared following standard methodology [34] and using a Buehler petrothin sectioning machine. Slides were observed using transmitted light microscopy. Images of the slides were obtained with a Leica DM 2500 transmitted light microscope with an incorporated, DFC 295 Leica camera. Images of the unmounted chert blocks and their sections were obtained using a Canon (Tokyo, Japan) camera with a Canon macro lens of 60 mm. Multiple images of the samples at several focal planes were taken to compose an image with greater depth of focus, following an image-stacking technique [35]. The images were stacked using auto-align and auto-blend functions in Adobe Photoshop 2023 (San José, USA). Composite images were created using both image-stacking and image-stitching techniques to visualize the entire specimens and their surrounding matrix (e.g., Figure 2).
The macrospecimens and their sections are housed at the Museo Regional Padre Molina of Río Gallegos (Santa Cruz Province, Argentina) under accession numbers MPM-Pb 16084-16109.
4.3. Surface 3D-Scanning of Fossils
Selected chert samples preserving one to several fern individuals were 3D scanned in high resolution using an Artec Space Spider 3D Scanner (Senningerberg, Luxembourg). Images were processed with the software Artec Studio 17 to obtain tridimensional models of the blocks in order to observe the superficial morphological features of the fern individuals, their distribution in the chert matrix, their relative position and distance to each other when more than one individual is preserved within the same block, and the distribution of other associated plant remains (e.g., conifer wood and twigs).
4.4. Surface 3D-Scanning of Fossils
In order to make comparisons with current Osmundaceae species that grow in Argentina, we studied populations of Osmunda spectabilis from Misiones province. The specimens were collected during field trips to the Cruce Caballero Provincial Park (-26.515012, -53.995383), herborized using traditional techniques and deposited in Museo Argentino de Ciencias Naturales “Bernardino Rivadavia” (BA herbarium).
In order to carry out anatomical comparisons, samples of petiole, rhizome and roots were fixed in a mixture of formalin, alcohol and acetic acid [36]. Histological preparations were made free-hand and mounted using gelatin-glycerin. In some cases, methyl blue was used for staining fungal structures in order to detect fungal associations.
Collected vouchers. ARGENTINA. Misiones: Dto. San Pedro, Parque Provincial Cruce Caballero, -26.515012 -53.995383, 22-V-2022, Yañez et al. 596 (BA).
5. Conclusions
It has been suggested that higher levels of organization of ecosystems impose a constraint over disturbance from lower hierarchical ones, resulting in greater stability due to gained resiliency of the system [37]. And, that such characteristic can explain paleoecological stability reflected by conserved morphology, composition, structure and dynamics of ecosystems through geological times [37]. This characteristic mechanism, preventive of directional selection, is a direct measure of the degree and nature of the interactions within the ecosystem, each adding complexity to it and a stronger basis for avoiding disrupting evolutionary change in multiple lineages [37]. Further factors, including historical, biogeographical, population size (law of large numbers), niche differentiation, and biotic interactions also lead to increased ecological constraint, pre-emptive of structural changes [11].
In this context, the Osmundaceae community from the Jurassic of Patagonia appears as a relictual type of analogous modern communities, in which selection processes through time did not alter its characteristic ecological organization. Its characteristic ecological structuring kept disruptive selection off from acting at a scale deeply affecting the osmundaceous ecosystems. In other words, its characteristic organization buffered and prevented it from deriving due to ecological disturbance. A proof of this is the analogous organization and composition of the fossil assemblage found in comparison with modern ones, especially those developed in the highlands of the Parana Forest, in northeastern Argentina [52]. In these type of systems, given the presence of a series of characteristics, described as those typical of spring swamps or highland lagoons, including restricted sedimentation ratios, constant water availability, neutral to acidic soils, abundant sun insolation due to the lack of canopy vegetation, and absence of major scale ecological disturbance, the arrangement of persistent Osmundaceous communities have found an optimal setting. In addition, some more specific features of the fossils found, such as the conserved morphology of the osmundaceous ferns stems and the types of interactions detected, including the formation of endomycorrhizae and the abundance of saprotrophic and parasitic chytridiomycetes within the tissues of individual plants, also are a reflection of the persistence and stability of the system through time.
Recurrent association through time to the same type of environments of Osmundaceous communities, together with other biotic groups, characteristic of these types of settings, is an example of how stabilizing selection due to the absence of major environmental change in a broader sense, works. The observed resiliency is reflected by the persistence of its structure and energy flux between different levels within it, where Millerocaulis zamunerae and fungal and protist taxa present had analogous ecological roles as those typical of modern highland, swamp ecosystems [16,71]. However, although at the ecological scale the ecosystem remained fully functional and structurally analogous through time, its composition at a lower selecting level (species) was subject to selection because also likely disrupting events occurred [13,52,157]. In other words, what this record shows is that even though the taxonomic composition of the osmundaceous paleocommunity changed through the period of time that separates it from modern analogous communities, at the ecosystem scale likely fluctuated within some ecological bounds, although without modifying its main structure and dynamics [12].
In turn, deciphering ecosystem scale ecological questions is possible when well-preserved, whole plant communities in life position and in-situ within their original substrates, such as the osmundaceous one from the Jurassic of Patagonia studied here, rather than single, isolated organs or organisms, are found [3]. The study of this paleocommunity is a demonstration of the concept of ecological stasis in an ample sense, where the equilibrium of the system means not its deviation from a single set of variables, but its stable fluctuation around a set of possibilities. It is a direct measure that shows that despite the range of disturbances that some communities can experience through time, their persistence in analogous configurations can be a characteristic of themselves [5,10,157]. Moreover, this finding shows that at the macroevolutionary scale the system has remained ecologically stable, having maintained its trophic structure and trophic links, represented by fossils belonging to the same macro-taxonomic groups, although of different specific composition [159]. This accounts for the maintenance of a dynamic equilibrium tending to stabilize the ecosystem based on the constant exchange and movement of matter within it [159]. In other words, the inferred complexity of the ecosystem based on the different functional groups found and the implied range of relationships with the vegetation present, especially with the osmundaceous ferns within the jurassic ecosystem, acted as an ecological resource that allowed variation of the system within some bounds, and which at the same time kept it stable through today.
Supplementary Materials
The original 3D-scanning files presented in the study as part of Supplementary Material are openly available in FigShare (10.6084/m9.figshare.27868176).
Author Contributions
Conceptualization, JLGM and GCN; Methodology, JLGM, GCN and AY; Software, GCN and IHE; Validation, IHE.; Formal Analysis, JLGM, GCN and AY; Investigation, JLGM, GCN, and AY; Resources, JLGM and IHE ; Data Curation, JLGM, GCN, AY, IHE and DG; Writing – Original Draft Preparation, JLGM; Writing – Review & Editing, JLGM, GCN, AY and IHE; Visualization, GCN and AY; Supervision, IHE; Project Administration, JLGM and IHE; Funding Acquisition, JLGM, GCN, AY and IHE.
Funding
This research was funded by PALSIRP Sepkoski Grant (GCN); APA-B&B Research Grant (GCN); PIP 202 & 173, PIP KE1 11220200100064CO (JLGM); MINCyT, PICT 2104-3496, PICT-2020 SERIEA 00416 (JLGM) and PIBAA 0062 (AY).
Data Availability Statement
In Argentina, fossils and fossil localities are considered part of the Cultural Heritage, thus they are protected by law (Ley Nacional 25 743, Ley de Protección del Patrimonio Arqueológico y Paleontológico). Additional legislation protects the fossils and fossil localities of Santa Cruz Province (Ley Provincial N.2472, Ley de Protección del Patrimonio Cultural). Thus, the GPS coordinates are associated with the specimens, housed at the Museo Regional Padre Molina of Río Gallegos (Santa Cruz Province, Argentina) and are available upon request to the Culture Bureau of Santa Cruz Province. See Materials and Methods for accession numbers.
Acknowledgments
We thank Federico Castía and Gonzalo J. Márquez for their assistance in the fieldwork on Osmunda spectabilis, and to the Ministry of Ecology and Renewable Natural Resources of Misiones Province for granting the necessary specimen collection permits. Our thanks also go to Ethel Denning (MEF) for scanning the chert samples. We are grateful to technicians S. Mirabelli (CRILAR) and Mariano Caffa (MEF) for their work on fossil preparation. Additionally, we thank the Culture Bureau of Santa Cruz Province for authorizing research in the geothermal deposits of the Deseado Massif, as well as the local community and Patagonian Gold workers for their logistical support. This work was funded by PalSIRP Sepkoski Grant (to GCN), APA-B&B Grant (to GCN), CONICET (RES. D. 2318, P. 202 & 173 to J.L.G.M.) and ANPCyT (PICT 2014-3496 to J.L.G.M. and D.G.).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jablonski, D.; Gould, S.J.; Raup, D.M. The Nature of the Fossil Record: A Biological Perspective. In Patterns and Processes in the History of Life; Raup, D.M., Jablonski, D., Eds.; Springer Berlin Heidelberg: Berlin, Heidelberg, 1986; pp. 7–22. ISBN 978-3-642-70833-6. [Google Scholar]
- Greenwood, D.R. The Taphonomy of Plant Macrofossils. In The Processes of Fossilization; Donovan, S.K., Ed.; Belhaven Press: London, 1991; pp. 141–169. [Google Scholar]
- Spicer, R.A. Plant Taphonomic Processes. In Taphonomy; Allison, P.A., Briggs, D.E.G., Eds.; Topics in Geobiology; Springer US: Boston, MA, 1991; ISBN 978-1-4899-5036-9. [Google Scholar]
- Taylor, T.N.; Krings, M.; Taylor, E.L. Fossil Fungi; Academic Press: London, 2015. [Google Scholar]
- Taylor, T.N.; Taylor, E.L.; Krings, M. Paleobotany: The Biology and Evolution of Fossil Plants, 2nd ed.; Academic Press: New York, 2009. [Google Scholar]
- Bomfleur, B.; McLoughlin, S.; Vajda, V. Fossilized Nuclei and Chromosomes Reveal 180 Million Years of Genomic Stasis in Royal Ferns. Science 2014, 343, 1376–1377. [Google Scholar] [CrossRef]
- Bomfleur, B.; Decombeix, A.L.; Escapa, I.H.; Schwendemann, A.B.; Axsmith, B. Whole-Plant Concept and Environment Reconstruction of a Telemachus Conifer (Voltziales) from the Triassic of Antarctica. Int. J. Plant Sci. 2013, 174, 425–444. [Google Scholar] [CrossRef]
- Phipps, C.J.; Taylor, T.N.; Taylor, E.L.; Cúneo, N.R.; Boucher, L.D.; Yao, X.L. Osmunda (Osmundaceae) from the Triassic of Antarctica: An Example of Evolutionary Stasis. Am. J. Bot. 888, 85, 1998.
- Schwendemann, A.B.; Taylor, T.N.; Taylor, E.L.; Krings, M. Organization, Anatomy, and Fungal Endophytes of a Triassic Conifer Embryo. Am. J. Bot. 2010, 97, 1873–1883. [Google Scholar] [CrossRef]
- Miller, C.N. Evolution of the Fern Family Osmundaceae Based on Anatomical Studies. Contrib. Mus. Paleontol. Univ. Mich. 1971, 23, 105–169. [Google Scholar]
- DiMichele, W.A.; Behrensmeyer, A.K.; Olszewski, T.D.; Labandeira, C.C.; Pandolfi, J.M.; Wing, S.L.; Bobe, R. Long-Term Stasis in Ecological Assemblages: Evidence from the Fossil Record. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 285–322. [Google Scholar] [CrossRef]
- Holling, C.S. Resilience and Stability of Ecological Systems. Annu. Rev. Ecol. Syst. 1973, 1–23. [Google Scholar] [CrossRef]
- Lehnert, M.; Monjau, T.; Rosche, C. Synopsis of Osmunda (Royal Ferns; Osmundaceae): Towards Reconciliation of Genetic and Biogeographic Patterns with Morphologic Variation. Bot. J. Linn. Soc. 2024, 205, 341–364. [Google Scholar] [CrossRef]
- Escapa, I.H.; Cúneo, N.R. Fertile Osmundaceae from the Early Jurassic of Patagonia, Argentina. Int. J. Plant Sci. 2012, 173, 54–66. [Google Scholar] [CrossRef]
- Tidwell, W.D.; Ash, S.R. A Review of Selected Triassic to Early Cretaceous Ferns. J. Plant Res. 1994, 107, 417–442. [Google Scholar] [CrossRef]
- Sagasti, A.J.; Massini, J.G.; Escapa, I.H.; Guido, D.M.; Channing, A. Millerocaulis Zamunerae Sp. Nov. (Osmundaceae) from Jurassic, Geothermally Influenced, Wetland Environments of Patagonia, Argentina. Alcheringa Australas. J. Palaeontol. 2016, 40, 456–474. [Google Scholar] [CrossRef]
- Cheng, Y.; Liu, F.; Yang, X.; Sun, T. Two New Species of Mesozoic Tree Ferns (Osmundaceae: Osmundacaulis) in Eurasia as Evidence of Long-Term Geographic Isolation. Geosci. Front. 2020, 11, 1875–1888. [Google Scholar] [CrossRef]
- Bomfleur, B.; Grimm, G.W.; McLoughlin, S. The Fossil Osmundales (Royal Ferns)—a Phylogenetic Network Analysis, Revised Taxonomy, and Evolutionary Classification of Anatomically Preserved Trunks and Rhizomes. PeerJ 2017, 5, e3433. [Google Scholar] [CrossRef] [PubMed]
- Tryon, R.M.; Tryon, A.F. Ferns and Allied Plants: With Special Reference to Tropical America; Springer: New York, NY, 1982; ISBN 978-1-4613-8164-8. [Google Scholar]
- Vera, E. A New Specimen of Millerocaulis (Osmundales: Osmundaceae) from the Cerro Negro Formation (Lower Cretaceous), Antarctica. Rev. Mus. Argent. Cienc. Nat. Nueva Ser. 2010, 12, 41–46. [Google Scholar] [CrossRef]
- Grimm, G.W.; Kapli, P.; Bomfleur, B.; McLoughlin, S.; Renner, S.S. Using More Than the Oldest Fossils: Dating Osmundaceae with Three Bayesian Clock Approaches. Syst. Biol. 2015, 64, 396–405. [Google Scholar] [CrossRef]
- Guido, D.M.; Campbell, K.A. Jurassic Hot Spring Deposits of the Deseado Massif (Patagonia, Argentina): Characteristics and Controls on Regional Distribution. J. Volcanol. Geotherm. Res. 2011, 203, 35–47. [Google Scholar] [CrossRef]
- García Massini, J.; Escapa, I.H.; Guido, D.M.; Channing, A. First Glimpse of the Silicified Hot Spring Biota from a New Jurassic Chert Deposit in the Deseado Massif, Patagonia, Argentina. Ameghiniana 2016, 53, 205–230. [Google Scholar] [CrossRef]
- Pankhurst, R.J.; Riley, T.R.; Fanning, C.M.; Kelley, S.P. Episodic Silicic Volcanism in Patagonia and the Antarctic Peninsula: Chronology of Magmatism Associated with the Break-up of Gondwana. J. Petrol. 2000, 41, 605–625. [Google Scholar] [CrossRef]
- Guido, D.M. Subdivisión litofacial e interpretación del volcanismo jurásico (Grupo Bahía Laura) en el este del Macizo del Deseado, provincia de Santa Cruz. Rev. Asoc. Geológica Argent. 2004, 59, 727–742. [Google Scholar]
- Pankhurst, R.J.; Leat, P.T.; Sruoga, P.; Rapela, C.W.; Márquez, M.; Storey, B.C.; Riley, T.R. The Chon Aike Province of Patagonia and Related Rocks in West Antarctica: A Silicic Large Igneous Province. J. Volcanol. Geotherm. Res. 1998, 81, 113–136. [Google Scholar] [CrossRef]
- Riley, T.R.; Leat, P.T.; Pankhurst, R.J.; Harris, C. Origins of Large Volume Rhyolitic Volcanism in the Antarctic Peninsula and Patagonia by Crustal Melting. J. Petrol. 2001, 42, 1043–1065. [Google Scholar] [CrossRef]
- Richardson, N.J.; Underhill, J.R. Controls on the Structural Architecture and Sedimentary Character of Syn-Rift Sequences, North Falkland Basin, South Atlantic. Mar. Pet. Geol. 2002, 19, 417–443. [Google Scholar] [CrossRef]
- Guido, D.M.; Campbell, K.A. A Large and Complete Jurassic Geothermal Field at Claudia, Deseado Massif, Santa Cruz, Argentina. J. Volcanol. Geotherm. Res. 2014, 275, 61–70. [Google Scholar] [CrossRef]
- Nunes, C.I.; García Massini, J.; Escapa, I.H.; Guido, D.M.; Campbell, K. Conifer Root Nodules Colonized by Arbuscular Mycorrhizal Fungi in Jurassic Geothermal Settings from Patagonia, Argentina. Int. J. Plant Sci. 2020, 181, 196–209. [Google Scholar] [CrossRef]
- Nunes, C.I.; García Massini, J.L.; Escapa, I.H.; Guido, D.M.; Campbell, K.A. Sooty Molds from the Jurassic of Patagonia, Argentina. Am. J. Bot. 2021, 108, 1464–1482. [Google Scholar] [CrossRef] [PubMed]
- Garcia Massini, J.L.; Guido, D.M.; Campbell, K.C.; Sagasti, A.J.; Krings, M. Filamentous Cyanobacteria and Associated Microorganisms, Structurally Preserved in a Late Jurassic Chert from Patagonia, Argentina. J. South Am. Earth Sci. 2021, 107, 103111. [Google Scholar] [CrossRef]
- Jones, T.P.; Rowe, N.P. Fossil Plants and Spores: Modern Techniques; Geological Society of London, 1999; ISBN 978-1-86239-035-5.
- Hass, H.; Rowe, N.P. Thin Section and Wafering. In Fossil Plant and Spores: Modern Techniques.; Jones, T.P., Rowe, N.P., Eds.; Geological Society of London: London, 1999; pp. 76–81. [Google Scholar]
- Bercovici, A.; Hadley, A.; Villanueva-Amadoz, U. Improving Depth of Field Resolution for Palynological Photomicrography. Palaeontol. Electron. 2009, 12, 0–12. [Google Scholar]
- Zarlavsky, G.E. Histología Vegetal: Técnicas Simples y Complejas; 1st ed.; Sociedad Argentina de Botánica: Buenos Aires, 2014. [Google Scholar]
- Morris, P.J.; Ivany, L.C.; Schopf, K.M.; Brett, C.E. The Challenge of Paleoecological Stasis: Reassessing Sources of Evolutionary Stability. Proc. Natl. Acad. Sci. USA 1995, 92, 11269–11273. [Google Scholar] [CrossRef]
- Landi, M.; Angiolini, C. Population Structure of Osmunda Regalis in Relation to Environment and Vegetation: An Example in the Mediterranean Area. Folia Geobot. 2011, 46, 49–68. [Google Scholar] [CrossRef]
- Landi, M.; Angiolini, C. Ecological Responses of Osmunda Regalis to Forest Canopy Cover and Grazing. Am. Fern J. 2010, 100, 137–147. [Google Scholar] [CrossRef]
- Klimešová, J.; Ottaviani, G.; Charles-Dominique, T.; Campetella, G.; Canullo, R.; Chelli, S.; Janovský, Z.; Lubbe, F.C.; Martínková, J.; Herben, T. Incorporating Clonality into the Plant Ecology Research Agenda. Trends Plant Sci. 2021, 26, 1236–1247. [Google Scholar] [CrossRef]
- Faust, A.; Petersen, R.L. Longevity of Interrupted Fern Colonies. Southeast. Nat. 2015, 14, 203–209. [Google Scholar] [CrossRef]
- Tsakalos, J.L.; Ottaviani, G.; Chelli, S.; Rea, A.; Elder, S.; Dobrowolski, M.P.; Mucina, L. Plant Clonality in a Soil-Impoverished Open Ecosystem: Insights from Southwest Australian Shrublands. Ann. Bot. 2022, 130, 981–990. [Google Scholar] [CrossRef] [PubMed]
- Burr, M.D.; Botero, L.M.; Young, M.J.; Inskeep, W.P.; McDermott, T.R. Observations Concerning Nitrogen Cycling in a Yellowstone Thermal Soil Environment. In Geothermal Biology and Geochemistry in Yellowstone National Park: Proceeding of the Thermal Biology Institute Workshop, Yellowstone National Park, W.Y.; Inskeep, W.P., Ed.; Montana State University Publications, 2005; pp. 171–182.
- Price, E.A.C.; Marshall, C. Clonal Plants and Environmental Heterogeneity – An Introduction to the Proceedings. Plant Ecol. 1999, 141, 3–7. [Google Scholar] [CrossRef]
- Guido, D.G.; Channing, A.; Campbell, K.A.; Zamuner, A. Jurassic Geothermal Landscapes and Fossil Ecosystems at San Agustín, Patagonia, Argentina. J. Geol. Soc. Lond. 2010, 167. [Google Scholar] [CrossRef]
- Aguraiuja, R.; Zobel, M.; Zobel, K.; Moora, M. Conservation of the Endemic Fern Lineage Diellia (Aspleniaceae) on the Hawaiian Islands: Can Population Structure Indicate Regional Dynamics and Endangering Factors? Folia Geobot. 2008, 43, 3–18. [Google Scholar] [CrossRef]
- Nunes, C.I.; Massini, J.L.G.; Escapa, I.H.; Guido, D.M.; Campbell, K. Conifer Root Nodules Colonized by Arbuscular Mycorrhizal Fungi in Jurassic Geothermal Settings from Patagonia, Argentina. Int. J. Plant Sci. 2020, 181, 196–209. [Google Scholar] [CrossRef]
- Nunes, C.I.; García Massini, J.L.; Escapa, I.H.; Guido, D.M.; Campbell, K.A. Sooty Molds from the Jurassic of Patagonia, Argentina. Am. J. Bot. 2021, 108, 1464–1482. [Google Scholar] [CrossRef]
- Escapa, I.H.; Garcia Massini, J.L.; Nunes, C.; Guido, D. Assembling a Jurassic Puzzle: Anatomically Preserved Conifer Remains in Hot Spring Deposits from Santa Cruz Province, Argentina.; Minesota, USA, 2018.
- Escapa, I.H.; Elgorriaga, A.; Nunes, C.; Scasso, R.; Cúneo, N.R. Megafloras Del Jurásico En La Cuenca de Cañadón Asfalto: Biomas En Transformación.; Puerto Madryn, 2022; pp. 878–901.
- Iglesias, A.; Artabe, A.E.; Morel, E.M. The Evolution of Patagonian Climate and Vegetation from the Mesozoic to the Present. Biol. J. Linn. Soc. 2011, 103, 409–422. [Google Scholar] [CrossRef]
- Regiones de humedales de la Argentina; Benzaquén, L. , Blanco, D.E., Bo, Kandus, P., Lingua, G., Minotti, P., Quintana, R., Eds.; 1st ed.; Ministerio de Ambiente y Desarrollo Sustentable, Fundación Humedales: Buenos Aires, Argentina, 2017; ISBN 978-987-29811-6-7. [Google Scholar]
- van der Heijden, M.G.A.; Martin, F.M.; Selosse, M.-A.; Sanders, I.R. Mycorrhizal Ecology and Evolution: The Past, the Present, and the Future. New Phytol. 2015, 205, 1406–1423. [Google Scholar] [CrossRef]
- Schüßler, A.; Walker, C. The Glomeromycota: A Species List with New Families and New Genera; Createspace Independent Pub: Gloucester, England, 2010. [Google Scholar]
- Morton, J.B.; Redecker, D. Two New Families of Glomales, Archaeosporaceae and Paraglomaceae, with Two New Genera Archaeospora and Paraglomus, Based on Concordant Molecular and Morphological Characters. Mycologia 2001, 93, 181–195. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis; Academic Press, 2010; ISBN 978-0-08-055934-6.
- Kessler, M.; Jonas, R.; Strasberg, D.; Lehnert, M. Mycorrhizal Colonizations of Ferns and Lycophytes on the Island of La Réunion in Relation to Nutrient Availability. Basic Appl. Ecol. 2010, 11, 329–336. [Google Scholar] [CrossRef]
- Wang, B.; Qiu, Y.-L. Phylogenetic Distribution and Evolution of Mycorrhizas in Land Plants. Mycorrhiza 2006, 16, 299–363. [Google Scholar] [CrossRef] [PubMed]
- Lehnert, M.; Krug, M.; Kessler, M. A Review of Symbiotic Fungal Endophytes in Lycophytes and Ferns – a Global Phylogenetic and Ecological Perspective. Symbiosis 2017, 71, 77–89. [Google Scholar] [CrossRef]
- Pirozynski, K.A.; Malloch, D.W. The Origin of Land Plants: A Matter of Mycotrophism. Biosystems 1975, 6, 153–164. [Google Scholar] [CrossRef]
- Brundrett, M.C. Coevolution of Roots and Mycorrhizas of Land Plants. New Phytol. 2002, 154, 275–304. [Google Scholar] [CrossRef]
- Lehnert, M.; Kessler, M. Review Mycorrhizal Relationships in Lycophytes and Ferns. Fern Gaz. 2016, 20, 101–116. [Google Scholar]
- Iqbal, S.H.; Yousaf, M.; Younus, M. A Field Survey of Mycorrhizal Associations in Ferns of Pakistan. New Phytol. 1981, 87, 69–79. [Google Scholar] [CrossRef]
- Brundrett, M.C. Mycorrhizal Associations and Other Means of Nutrition of Vascular Plants: Understanding the Global Diversity of Host Plants by Resolving Conflicting Information and Developing Reliable Means of Diagnosis. Plant Soil 2009, 320, 37–77. [Google Scholar] [CrossRef]
- Brundrett, M.C. Biological reviews. 2004, pp. 473–495.
- Wright, D.P.; Scholes, J.D.; Read, D.J. Effects of VA Mycorrhizal Colonization on Photosynthesis and Biomass Production of Trifolium Repens L. Plant Cell Environ. 1998, 21, 209–216. [Google Scholar] [CrossRef]
- Du, J.; Yu, F.-H.; Alpert, P.; Dong, M. Arbuscular Mycorrhizal Fungi Reduce Effects of Physiological Integration in Trifolium Repens. Ann. Bot. 2009, 104, 335–344. [Google Scholar] [CrossRef]
- Johnson, N.C.; Graham, J.-H.; Smith, F.A. Functioning of Mycorrhizal Associations along the Mutualism–Parasitism Continuum. New Phytol. 1997, 135, 575–585. [Google Scholar] [CrossRef]
- Dominiak, M.; Olejniczak, P.; Lembicz, M. Diversified Impact of Mycorrhizal Inoculation on Mother Plants and Daughter Ramets in the Clonally Spreading Plant Hieracium Pilosella L. (Asteraceae). Plant Ecol. 2019, 220, 757–763. [Google Scholar] [CrossRef]
- Bittebiere, A.-K.; Benot, M.-L.; Mony, C. Clonality as a Key but Overlooked Driver of Biotic Interactions in Plants. Perspect. Plant Ecol. Evol. Syst. 2020, 43, 125510. [Google Scholar] [CrossRef]
- Dighton, J. Fungi in Ecosystem Processes, 2nd ed.; CRC Press: Boca Raton, 2016; ISBN 978-1-315-37152-8. [Google Scholar]
- Huey, C.J.; Gopinath, S.C.B.; Uda, M.N.A.; Zulhaimi, H.I.; Jaafar, M.N.; Kasim, F.H.; Yaakub, A.R.W. Mycorrhiza: A Natural Resource Assists Plant Growth under Varied Soil Conditions. 3 Biotech 2020, 10, 204. [Google Scholar] [CrossRef] [PubMed]
- Redman, R.S.; Sheehan, K.B.; Stout, R.G.; Rodriguez, R.J.; Henson, J.M. Thermotolerance Generated by Plant/Fungal Symbiosis. Science 2002, 298, 1581–1581. [Google Scholar] [CrossRef]
- Redman, R.S.; Litvintseva, A.; Sheehan, K.B.; Henson, J.M.; Rodriguez, R.J. Fungi from Geothermal Soils in Yellowstone National Park | Applied and Environmental Microbiology. Appl. Environ. Microbiol. 1999, 65, 5193–5197. [Google Scholar] [CrossRef]
- Streitwolf-Engel, R.; Van der Heijden, M.G.A.; Wiemken, A.; Sanders, I.R. The Ecological Significance of Arbuscular Mycorrhizal Fungal Effects on Clonal Reproduction in Plants. Ecology 2001, 82, 2846–2859. [Google Scholar] [CrossRef]
- Lehnert, M.; Kottke, I.; Setaro, S.; Pazmiño, L.F.; Suárez, J.P.; Kessler, M. Mycorrhizal Associations in Ferns from Southern Ecuador. Am. Fern J. 2009, 99, 292–306. [Google Scholar] [CrossRef]
- Ogura-Tsujita, Y.; Hirayama, Y.; Sakoda, A.; Suzuki, A.; Ebihara, A.; Morita, N.; Imaichi, R. Arbuscular Mycorrhizal Colonization in Field-Collected Terrestrial Cordate Gametophytes of Pre-Polypod Leptosporangiate Ferns (Osmundaceae, Gleicheniaceae, Plagiogyriaceae, Cyatheaceae) | Mycorrhiza. Mycorrhiza 2016, 26, 87–97. [Google Scholar] [CrossRef]
- Channing, A.; Edwards, D. Yellowstone Hot Spring Environments and the Palaeo-Ecophysiology of Rhynie Chert Plants: Towards a Synthesis. Plant Ecol. Divers. 2009. [CrossRef]
- Sparrow, F.K. Aquatic Phycomycetes; 2d rev. ed.; University of Michigan Press: Ann Arbor, 1960. [Google Scholar]
- Barnett, H.L.; Hunter, B.B. Illustrated Genera of Imperfect Fungi, 4th ed.; APS Press: St. Paul, 1998. [Google Scholar]
- Prescott, G.W. How to Know Freshwater Algae. Pictured Key Nature Series., 3rd ed.; Wm. C. Brown Company Publishers: Dubuque, 1980. [Google Scholar]
- Patterson, R.T.; Baker, T.; Burbridge, S.M. Arcellaceans (Thecamoebians) as Proxies of Arsenic and Mercury Contamination in Northeastern Ontario Lakes. J. Foraminifer. Res. 1996, 26, 172–183. [Google Scholar] [CrossRef]
- Cavalier-Smith, T. A Revised Six-Kingdom System of Life. Biol. Rev. 1998, 73, 203–266. [Google Scholar] [CrossRef] [PubMed]
- Dick, M.W. Straminipilous Fungi: Systematics of the Peronosporomycetes Including Accounts of the Marine Straminipilous Protists, the Plasmodiophorids and Similar Organisms; Kluwer Academic Publishers: London, 2001. [Google Scholar]
- Lynn, D.H.; Small, E.B. Phylum Ciliophora. In The illustrated guide to the Protozoa; Lee, J.J., Leedale, G.F., Bradbury, P., Eds.; Society of Protozoologists, Allen Press Inc.: Lawrence, 2000; Volume 2, pp. 371–656. [Google Scholar]
- Mesterfield, R. Order Arcellinida. In The illustrated guide to the Protozoa; Lee, J.J., Leedale, G.F., Eds.; Society of Protozoologists, Allen Press Inc.: Lawrence, 2000; Volume 2, pp. 827–859. [Google Scholar]
- Patterson, D.J. Free-Living Freshwater Protozoa. A Color Guide.; CRC Press: Boca Raton, USA, 1992. [Google Scholar]
- Karling, J.S. The Plasmodiophorales, 1st ed.; The author: New York city, 1942; pp. 1–168. [Google Scholar]
- Heger, T., J.; Foissner, W. Ecology and Biodiversity of Testate Amoebae in Freshwater Habitats. Hydrobiologia 2009, 634, 115–124. [Google Scholar]
- Huann-Ju, H.; Chang, H.-S. Five Species of Pythium, Two Species of Pythiogeton New for Taiwan and Pythium Afertile. Bot. Bull. Acad. Sin. 1976, 17, 141–150. [Google Scholar]
- Dick, M.W. Morphology and Taxonomy of the Oomycetes, with Special Reference to Saprolegniaceae, Leptomitaceae and Pythiaceae. New Phytol. 1969, 68, 751–775. [Google Scholar] [CrossRef]
- Kalgutkar, R.M.; Jansonius, J. Synopsis of Fossil Fungal Spores, Mycelia and Fructifications; Palynologists, Contribution Series, Ed.; American Association of Stratigraphic: Dallas, 2000. [Google Scholar]
- Karling, J.S. Chytridiomycetarum Iconographia; Lubrecht & Cramer Ltd: Germany, 1977. [Google Scholar]
- Longcore, J.E. Chytridiomycete Taxonomy since 1960. Mycotaxon 1996, 60, 149–174. [Google Scholar]
- Longcore, J.E. Chytridiomycota. In Systematics and Evolution of Fungi; Springer, 1995.
- Gleason, F.H.; Scholz, B.; Jephcott, T.G.; van Ogtrop, F.F.; Henderson, L.; Lilje, O.; Kittelmann, S.; Macarthur, D.J. Key Ecological Roles for Zoosporic True Fungi in Aquatic Habitats. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef]
- DiLeo, K.; Donat, K.; Min-Venditti, A.; Dighton, J. A Correlation between Chytrid Abundance and Ecological Integrity in New Jersey Pine Barrens Waters. Fungal Ecol. 2010, 3, 295–301. [Google Scholar] [CrossRef]
- Rimington, W.R.; Duckett, J.G.; Field, K.J.; Bidartondo, M.I.; Pressel, S. The Distribution and Evolution of Fungal Symbioses in Ancient Lineages of Land Plants. Mycorrhiza 2020, 30, 23–49. [Google Scholar] [CrossRef]
- Rimington, W.R.; Pressel, S.; Duckett, J.G.; Bidartondo, M.I. Fungal Associations of Basal Vascular Plants: Reopening a Closed Book? New Phytol. 2015, 205, 1394–1398. [Google Scholar] [CrossRef]
- Lund, J.W.G. The Ecology of the Freshwater Algae Potamogeton in the English Lakes. J. Ecol. 1954, 42, 366–385. [Google Scholar]
- Fisher, P.J.; Petrini, O. Fungal Endophytes in Phragmites Australis. Mycol. Res. 1992, 96, 1–5. [Google Scholar]
- Channing, A.; Wujek, D.E. Preservation of Protists within Decaying Plants from Geothermally Influenced Wetlands of Yellowstone National Park, Wyoming, United States. PALAIOS 2010, 25, 347–355. [Google Scholar] [CrossRef]
- Krings, M.; Harper, C.J.; White, J.F.; Barthel, M.; Heinrichs, J.; Taylor, E.L.; Taylor, T.N. Fungi in a Psaronius Root Mantle from the Rotliegend (Asselian, Lower Permian/Cisuralian) of Thuringia, Germany. Rev. Palaeobot. Palynol. 2017, 239, 14–30. [Google Scholar] [CrossRef]
- McLoughlin, S.; Bomfleur, B. Biotic Interactions in an Exceptionally Well Preserved Osmundaceous Fern Rhizome from the Early Jurassic of Sweden. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016, 464, 86–96. [Google Scholar] [CrossRef]
- Bippus, A.C.; Escapa, I.H.; Wilf, P.; Tomescu, A.M.F. Fossil Fern Rhizomes as a Model System for Biotic Interactions across Geologic Time: Evidence from Patagonia; PeerJ Inc. 2019. [Google Scholar]
- Ellis, M.B. British Fungi, Part 2; Jarrold Publishing: Norwich, 1976. [Google Scholar]
- Ellis, M.B. Dematiaceous Hyphomycetes; Kew Botanical Garden: Surrey, England, 1971. [Google Scholar]
- Cannon, P.F.; Kirk, P.M. Fungal Families of the World; CAB International: Surrey, England, 2007. [Google Scholar]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W. Dictionary of the Fungi, 3rd ed.; CABI Europe: UK, 2008. [Google Scholar]
- Domsch, K.H.; Gams, W. Anderson Compendium of Soil Fungi, 2nd ed.; IHW-Verlag: Eching, 2007. [Google Scholar]
- Osono, T. Ecology of Ligninolytic Fungi Associated with Leaf Litter Decomposition. Ecol. Res. 2007, 22, 955–974. [Google Scholar] [CrossRef]
- Deshmukh, S.K.; Mishra, R.R. Fungi Associated with the Fronds of Pteris Vittata L. (Pteridaceae). Trop. Ecol. 29-33, 35, 1994.
- Kumar, D.S.; Hyde, K.D. Endophytic Fungal Assemblages in Fern Species: A Preliminary Study. Mycoscience 2004, 45, 334–338. [Google Scholar]
- Howard, R.J.; Ferrari, M.A. Role of the Fungus Penicillium in the Formation of Appressoria and Pathogenicity in Plants. Mycol. Res. 93, 345–353.
- Chandra, G.; Chatter, K.F. Microbiology Reviews. 2014, pp. 345–379.
- Chandra, G.; Chater, K. Developmental Biology of Streptomyces from the Perspective of 100 Actinobacterial Genome Sequences. FEMS Microbiol. Rev. 2014, 38, 345–379. [Google Scholar] [CrossRef]
- Giovannetti, M.; Sbrana, C.; Avio, L.; Citernesi, A.S.; Logi, C. Differential Hyphal Morphogenesis in Arbuscular Mycorrhizal Fungi during Pre-Infection Stages. New Phytol. 1993, 125, 587–593. [Google Scholar] [CrossRef]
- Vannier, N.; Bittebiere, A.-K.; Vandenkoornhuyse, P.; Mony, C. AM Fungi Patchiness and the Clonal Growth of Glechoma Hederacea in Heterogeneous Environments. Sci. Rep. 2016, 6, 37852. [Google Scholar] [CrossRef]
- Bittebiere, A.-K.; Benot, M.-L.; Mony, C. Clonality as a Key but Overlooked Driver of Biotic Interactions in Plants. Perspect. Plant Ecol. Evol. Syst. 2020, 43, 125510. [Google Scholar] [CrossRef]
- Vannier, N.; Mony, C.; Bittebiere, A.-K.; Michon-Coudouel, S.; Biget, M.; Vandenkoornhuyse, P. A Microorganisms’ Journey between Plant Generations. Microbiome 2018, 6, 79. [Google Scholar] [CrossRef] [PubMed]
- Schwarze, F.W.M.R. Wood Decay under the Microscope. Fungal Biol. Rev. 2007, 21, 133–170. [Google Scholar] [CrossRef]
- Singh, A.P.; Nilsson, N.; Daniel, D. Variable Resistance of Pinus Sylvestris Wood Components to Attack by Wood Degrading Bacteria. In Recent Advances in Wood Anatomy; Donaldson, L.A., Singh, A.P., Butterfield, B.G., Whitehouse, L., Eds.; New Zealand Forest Research Institute: Rotorua, New Zealand, 1996; pp. 408–416. [Google Scholar]
- Kim, Y.S.; Singh, A.P. Micromorphological Characteristics of Wood Biodegradation in Wet Environments: A Review. IAWA J. 2000, 135–155. [Google Scholar] [CrossRef]
- Singh, A.P.; Kim, Y.S. ; Singh Tripti Bacterial Degradation of Wood. In Secondary Xylem Biology; Kim, Y.S., Funada, R., Singh, A.P., Eds.; Elsevier Inc., 2016; pp. 169–190.
- Shigo, A.L.; Marx, H.G. Compartimentalization of Decay in Trees. Bull. US Dep. Agric. 1977, 405, 73. [Google Scholar]
- Clausen, A.C. International biodeterioration & biodegradation. 1996,.
- Labandeira, C.C.; Lucas, S.G.; Kirkland, J.I.; Estep, J.W. The Role of Insects in Late Jurassic to Middle Cretaceous Ecosystems. In Lower and Middle Cretaceous Terrestrial Ecosystems; New Mexico Museum of Natural History and Science Bulletin: Albuquerque, USA, 1998. [Google Scholar]
- Kellogg, D.W.; Taylor, E.L. Evidence of Oribatid Mite Detritivory in Antarctica during the Late Paleozoic and Mesozoic. J. Paleontol. 2004, 78, 1146–1153. [Google Scholar] [CrossRef]
- Haq, M.A. Potential of Oribatid Mites in Biodegradation and Mineralization for Enhancing Plant Productivity. Acarol. Stud. 2019, 1, 101–122. [Google Scholar]
- Tidwell, W.D.; Clifford, H.T. Three New Species of Millerocaulis (Osmundaceae) from Queensland, Australia. Aust. Syst. Bot. 1995, 8, 667–685. [Google Scholar] [CrossRef]
- D’Rozario, A.; Labandeira, C.; Guo, W.-Y.; Yao, Y.-F.; Li, C.-S. Spatiotemporal Extension of the Euramerican Psaronius Component Community to the Late Permian of Cathaysia: In Situ Coprolites in a P. Housuoensis Stem from Yunnan Province, Southwest China. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 306, 127–133. [Google Scholar] [CrossRef]
- Solomon, J.D. Frass Characteristics for Identifying Insect Borers (Lepidoptera: Cossidae and Sesiidae; Coleoptera: Cerambycidae) in Living Hardwoods. Can. Entomol. 1977, 109, 295–303. [Google Scholar] [CrossRef]
- Solomon, J.D. Guide to Insect Borers in North American Broadleaf Trees and Shrubs; United States Department of Agriculture Forest Service: New Orleans, USA, 1995. [Google Scholar]
- Greppi, C.D.; García Massini, J.L.; Pujana, R.R. Saproxylic Arthropod Borings in Nothofagoxylon Woods from the Miocene of Patagonia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2021, 571, 110369. [Google Scholar] [CrossRef]
- Havrylenko, D.; Winterhalter, J.J. Insectos Del Parque Nacional Nahuel Huapí; Administración General de Parques Nacionales y Turismo. 1949. [Google Scholar]
- Yee, M.; Grove, S.J.; Richardson, A.M.M.; Mohammed, C.L. Brown Rot in Inner Heartwood: Why Large Logs Support Characteristic Saproxylic Beetle Assemblages of Conservation Concern. In Insect biodiversity and dead wood: proceedings of a symposium for the22nd International Congress of Entomology; Grove, G.J., Simon, J., Hanula, J.L., Eds.; Department of Agriculture Forest Service, SouthernResearch Station: Asheville, NC, USA, 2006; pp. 42–53. [Google Scholar]
- Kukor, J.J.; Martin, M.M. Cellulose Digestion inMonochamus Marmorator Kby (Coleoptera: Cerambycidae): Role of Acquired Fungal Enzymes. J. Chem. Ecol. 1986, 12, 1057–1070. [Google Scholar] [CrossRef] [PubMed]
- Wotton, R.S.; Malmqvist, B.; Muotka, T.; Larsson, K. Fecal Pellets from a Dense Aggregation of Suspension-Feeders in a Stream: An Example of Ecosystem Engineering. Limnol. Oceanogr. 1998, 43, 719–725. [Google Scholar] [CrossRef]
- Schwendemann, A.B.; Taylor, T.N.; Taylor, E.L.; Krings, M.; Dotzler, N. Combresomyces Cornifer from the Triassic of Antarctica: Evolutionary Stasis in the Peronosporomycetes. Rev. Palaeobot. Palynol. 2009, 154, 1–5. [Google Scholar] [CrossRef]
- Christiansen, E.; Krokene, P.; Berryman, A.A.; Franceschi, V.R.; Krekling, T.; Lieutier, F.; Lönneborg, A.; Solheim, H. Mechanical Injury and Fungal Infection Induce Acquired Resistance in Norway Spruce. Tree Physiol. 1999, 19, 399–403. [Google Scholar] [CrossRef]
- Franceschi, V.R.; Krokene, P.; Christiansen, E.; Krekling, T. Anatomical and Chemical Defenses of Conifer Bark against Bark Beetles and Other Pests. New Phytol. 2005, 167, 353–376. [Google Scholar] [CrossRef]
- Krokene, P.; Nagy, N.E.; Krekling, T. Traumatic Resin Ducts and Polyphenolic Parenchyma Cells in Conifers. In Induced Plant Resistance to Herbivory; Schaller, A., Ed.; Springer Science, 2008; pp. 147–169.
- Shain, L. Dynamic Responses of Differentiated Sapwood to Injury and Infection. Phytopathology 1979, 69, 1143–1147. [Google Scholar] [CrossRef]
- Hudgins, J.W.; Christiansen, E.; Franceschi, V.R. Induction of Anatomically Based Defense Responses in Stems of Diverse Conifers by Methyl Jasmonate: A Phylogenetic Perspective. Tree Physiol. 2004, 24, 251–264. [Google Scholar] [CrossRef]
- Krokene, P.; Nagy, N.E.; Krekling, T. Traumatic Resin Ducts and Polyphenolic Parenchyma Cells in Conifers. In Induced plant resistance to herbivory; Schaller, A., Ed.; Springer: Netherlands, 2008. [Google Scholar]
- Esteban, L.G.; Guindeo, A.; Peraza, C.; de Palacios, P. La Madera y Su Anatomía; Fundacion Conde del Valle de Salazar, Mundi-prensa y AiTiM: Madrid, España, 2003. [Google Scholar]
- Rombola, C.F.; Greppi, C.D.; Pujana, R.R.; García Massini, J.L.; Bellosi, E.S.; Marenssi, S.A. Brachyoxylon Fossil Woods with Traumatic Resin Canals from the Upper Cretaceous Cerro Fortaleza Formation, Southern Patagonia (Santa Cruz Province, Argentina). Cretac. Res. 2022, 130, 105065. [Google Scholar] [CrossRef]
- Bonello, P.; Blodgett, J.T. Pinus Nigra–Sphaeropsis Sapinea as a Model Pathosystem to Investigate Local and Systemic Effects of Fungal Infection of Pines. Physiol. Mol. Plant Pathol. 2003, 63, 249–261. [Google Scholar] [CrossRef]
- Shortle, W.C. Mechanisms of Compartmentalization of Decay in Living Trees. Phytopathology 1979, 69, 1147–1151. [Google Scholar] [CrossRef]
- Shain, I. Dynamic Responses of Differentiated Sapwood to Injury and Infection. Phytopathology 1919, 69, 1143–1147. [Google Scholar] [CrossRef]
- Christiansen, E.; Franceschi, V.R.; Nagy, N.E.; Krekling, T.; Berryman, A.A.; Krokene, P.; Solheim, H. Traumatic Resin Duct Formation in Norway Spruce (Picea Abies (L.) Karst.) after Wounding or Infection with a Bark Beetle-Associated Blue-Stain Fungus, Ceratocystis Polonica. In Physiology and genetics of tree-phytophage interactions; Lieutier, F., Mattson, W.J., Wagner, M.R., Eds.; Les Colloques de I’INRA: Versailles, 1999; pp. 79–89. [Google Scholar]
- Lara-Pérez, L.A.; Zulueta-Rodríguez, R.; Andrade-Torres, A. Micorriza arbuscular, Mucoromycotina y hongos septados oscuros en helechos y licófitas con distribución en México: una revisión global. Rev. Biol. Trop. 2017, 65, 1062–1081. [Google Scholar] [CrossRef]
- Ohgushi, T. Herbivore-Induced Indirect Interaction Webs on Terrestrial Plants: The Importance of Non-Trophic, Indirect, and Facilitative Interactions. Entomol. Exp. Appl. 2008, 128, 217–229. [Google Scholar] [CrossRef]
- Brunner, F.S.; Deere, J.A.; Egas, M.; Eizaguirre, C.; Raeymaekers, J.A.M. The Diversity of Eco-Evolutionary Dynamics: Comparing the Feedbacks between Ecology and Evolution across Scales. Funct. Ecol. 2019, 33, 7–12. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, L.-D.; Liu, R.-J. Arbuscular Mycorrhizal Fungi Associated with Common Pteridophytes in Dujiangyan, Southwest China. Mycorrhiza 2004, 14, 25–30. [Google Scholar] [CrossRef]
- Braun, U.; Nakashima, C.; Crous, P.W. Cercosporoid Fungi (Mycosphaerellaceae) 1. Species on Other Fungi, Pteridophyta and Gymnospermae. IMA Fungus 2013, 4, 265–345. [Google Scholar] [CrossRef] [PubMed]
- Holling, C.S. Resilience and Stability of Ecological Systems. Annu. Rev. Ecol. Evol. Syst. 1973, 4, 1–23. [Google Scholar] [CrossRef]
- Yachi, S.; Loreau, M. Biodiversity and Ecosystem Productivity in a Fluctuating Environment: The Insurance Hypothesis. Proc. Natl. Acad. Sci. USA 1999, 96, 1463–1468. [Google Scholar] [CrossRef]
- Vislobokova, I.A. The Concept of Macroevolution in View of Modern Data. Paleontol. J. 2017, 51, 799–898. [Google Scholar] [CrossRef]
Figure 1.
Studied area and distribution of in situ plants at the La Bajada locality, in Santa Cruz Province, Argentina. (a) General view of the studied area at the La Bajada locality were the geothermal deposits outcrop. (b) Distribution and excavation of chert blocks bearing numerous osmundaceous ferns in life position. (c) Cluster of semi-burried chert blocks with in situ fern stems. (d) Detail of in situ fern stem embedded in the chert bed in transverse section.
Figure 1.
Studied area and distribution of in situ plants at the La Bajada locality, in Santa Cruz Province, Argentina. (a) General view of the studied area at the La Bajada locality were the geothermal deposits outcrop. (b) Distribution and excavation of chert blocks bearing numerous osmundaceous ferns in life position. (c) Cluster of semi-burried chert blocks with in situ fern stems. (d) Detail of in situ fern stem embedded in the chert bed in transverse section.

Figure 2.
Fossil Osmundaceae. (a) Cross section of rhizomes of three individuals (white arrows). (b) Longitudinal section of the rhizome. (c) Cross section of the rhizome showing internal petiole cycles and root mantle. White dotted circle indicates a lateral rhizome separating from the main plant for vegetative reproduction. Notice the central stem comprising the collapsed pith (p), vascular cylinder (vc), parenchymatous inner cortex (ic), and sclerotic outer cortex (oc); and the outer cycles of petiole bases (pb) characterized by stipular wings with sclerotic masses (sm), and a sclerotic ring (sr). (d) Detail of (c) showing petioles (black arrow) close to the central cylinder and superficial roots (white arrow). (e) Detail of (c) showing lateral rhizome (white dotted circle). Observe the formation of the leaf trace (black dotted ellipse), which separates from the xylem cylinder with only one protoxylem cluster, which divides in the outer cortex. Bar = 5 cm, (d-e) 1 cm.
Figure 2.
Fossil Osmundaceae. (a) Cross section of rhizomes of three individuals (white arrows). (b) Longitudinal section of the rhizome. (c) Cross section of the rhizome showing internal petiole cycles and root mantle. White dotted circle indicates a lateral rhizome separating from the main plant for vegetative reproduction. Notice the central stem comprising the collapsed pith (p), vascular cylinder (vc), parenchymatous inner cortex (ic), and sclerotic outer cortex (oc); and the outer cycles of petiole bases (pb) characterized by stipular wings with sclerotic masses (sm), and a sclerotic ring (sr). (d) Detail of (c) showing petioles (black arrow) close to the central cylinder and superficial roots (white arrow). (e) Detail of (c) showing lateral rhizome (white dotted circle). Observe the formation of the leaf trace (black dotted ellipse), which separates from the xylem cylinder with only one protoxylem cluster, which divides in the outer cortex. Bar = 5 cm, (d-e) 1 cm.

Figure 3.
Current Osmunda spectabilis in wetlands from Misiones province. (a) Wetland dominated by O. spectabilis and N. brasiliense. (b) General view of O. spectabilis plants. (c) Exposed underground rhizome showing the emergence of densely arranged petioles. (d) Cross section of the rhizome showing petioles (black arrow) close to the central cylinder and superficial roots (white arrow). (e) Cross section of the rhizome showing petioles (black arrow). A dotted circle indicates a lateral rhizome separating from the main plant for vegetative reproduction. (f) Cross section of roots. White arrow indicates colonization by arbuscular mycorrhizae. (g) Surface view of branched roots of different diameters. Bar = (c) 4 cm, (d, g) 5 cm, (e) 8 cm, (f) 150 µm.
Figure 3.
Current Osmunda spectabilis in wetlands from Misiones province. (a) Wetland dominated by O. spectabilis and N. brasiliense. (b) General view of O. spectabilis plants. (c) Exposed underground rhizome showing the emergence of densely arranged petioles. (d) Cross section of the rhizome showing petioles (black arrow) close to the central cylinder and superficial roots (white arrow). (e) Cross section of the rhizome showing petioles (black arrow). A dotted circle indicates a lateral rhizome separating from the main plant for vegetative reproduction. (f) Cross section of roots. White arrow indicates colonization by arbuscular mycorrhizae. (g) Surface view of branched roots of different diameters. Bar = (c) 4 cm, (d, g) 5 cm, (e) 8 cm, (f) 150 µm.

Figure 4.
Sporangia embedded in the chert matrix, associated with the rhizomatous stems. (a) General view of chert matrix containing roots belonging to the Osmundaceae rhizomatous stems, showing associated remains of the annulus of a single sporangium, composed of thick-walled cells. (b). General view of group of sporangia with variable preservation with in situ spores, preserved along the rhizomatous stems. (c) Close up of some of the sporangia, showing the presence of a short stalk (white arrowhead). (d) Close up of a sporangium containing numerous trilete spores. Bar = (a-b) 1 mm, (c) 70 µm, (d) 20 µm.
Figure 4.
Sporangia embedded in the chert matrix, associated with the rhizomatous stems. (a) General view of chert matrix containing roots belonging to the Osmundaceae rhizomatous stems, showing associated remains of the annulus of a single sporangium, composed of thick-walled cells. (b). General view of group of sporangia with variable preservation with in situ spores, preserved along the rhizomatous stems. (c) Close up of some of the sporangia, showing the presence of a short stalk (white arrowhead). (d) Close up of a sporangium containing numerous trilete spores. Bar = (a-b) 1 mm, (c) 70 µm, (d) 20 µm.
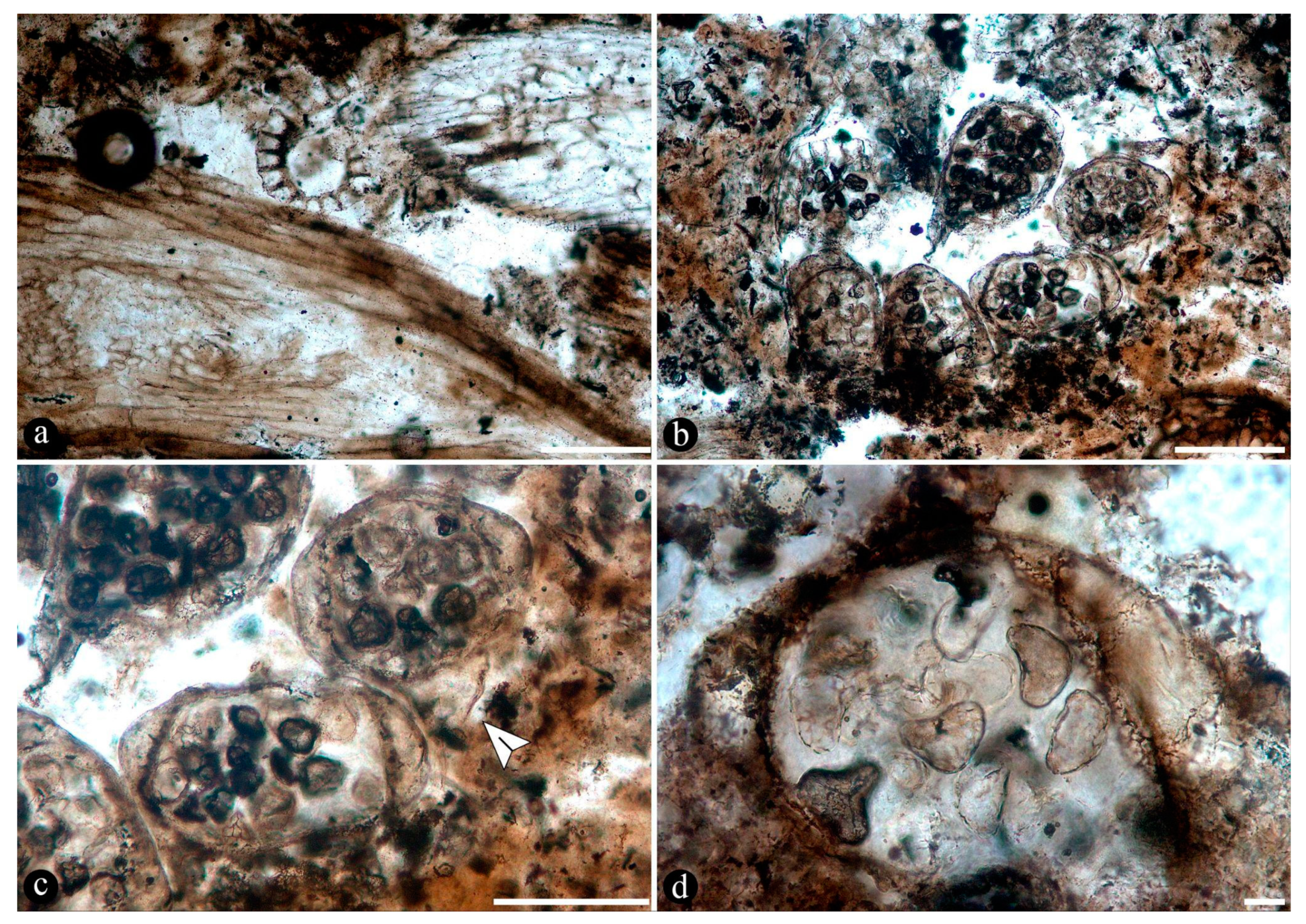
Figure 5.
Fungal interactions in the roots of fossil and extant Osmundaceae. (a) General view of silicified adventitious diarch root of rhizomatous stems of Osmundaceae preserved in a siliceous chert matrix, formed around an in-situ community in geothermal settings from the Jurassic of Patagonia, Argentina, filled with fungal material. (b) Close-up on cortical cells of fossil diarch root showing distribution of fungal material around the pericycle. (c) Close-up on a few cortical cells of fossil diarch root showing fungal material consisting of highly ramified arbuscules. Note arbuscules emerging from broader, trunk hypha (arrowhead). (d) General view of extant Osmunda spectabilis small diarch root filled with fungal material. (e) Close-up on cortical cells of O. spectabilis diarch root showing distribution of fungal material around the pericycle. (f) Close-up on a few cortical cells showing highly ramified arbuscules, in O. spectabilis small diarch root. (g) Smaller diarch root of fossil Osmundaceae, whose cortical cells are variously filled with fungal material and an amorphous, somewhat opaque residue. Also note, some broken cells and the resulting cell-free area to the upper right of the root. (h) Detail of fossil diarch root showing coenocytic hyphae within some cortical cells. (i) Detail of fossil diarch root showing globose, intracellular spore. (j) Close on cortex of fossil diarch root showing another, 2-layered, intracellular spore, subtended by a short hypha (arrow). (k) Identical globose spore in the surrounding silicified matrix, connected to the epidermis of a fossil small diarch root via a single hypha thread (arrow). Bar = (a) 200 µm, (b, f, i-k) 20 µm, (c, h) 10 µm, (d) 100 µm, (e) 65 µm, (g) 150 µm.
Figure 5.
Fungal interactions in the roots of fossil and extant Osmundaceae. (a) General view of silicified adventitious diarch root of rhizomatous stems of Osmundaceae preserved in a siliceous chert matrix, formed around an in-situ community in geothermal settings from the Jurassic of Patagonia, Argentina, filled with fungal material. (b) Close-up on cortical cells of fossil diarch root showing distribution of fungal material around the pericycle. (c) Close-up on a few cortical cells of fossil diarch root showing fungal material consisting of highly ramified arbuscules. Note arbuscules emerging from broader, trunk hypha (arrowhead). (d) General view of extant Osmunda spectabilis small diarch root filled with fungal material. (e) Close-up on cortical cells of O. spectabilis diarch root showing distribution of fungal material around the pericycle. (f) Close-up on a few cortical cells showing highly ramified arbuscules, in O. spectabilis small diarch root. (g) Smaller diarch root of fossil Osmundaceae, whose cortical cells are variously filled with fungal material and an amorphous, somewhat opaque residue. Also note, some broken cells and the resulting cell-free area to the upper right of the root. (h) Detail of fossil diarch root showing coenocytic hyphae within some cortical cells. (i) Detail of fossil diarch root showing globose, intracellular spore. (j) Close on cortex of fossil diarch root showing another, 2-layered, intracellular spore, subtended by a short hypha (arrow). (k) Identical globose spore in the surrounding silicified matrix, connected to the epidermis of a fossil small diarch root via a single hypha thread (arrow). Bar = (a) 200 µm, (b, f, i-k) 20 µm, (c, h) 10 µm, (d) 100 µm, (e) 65 µm, (g) 150 µm.
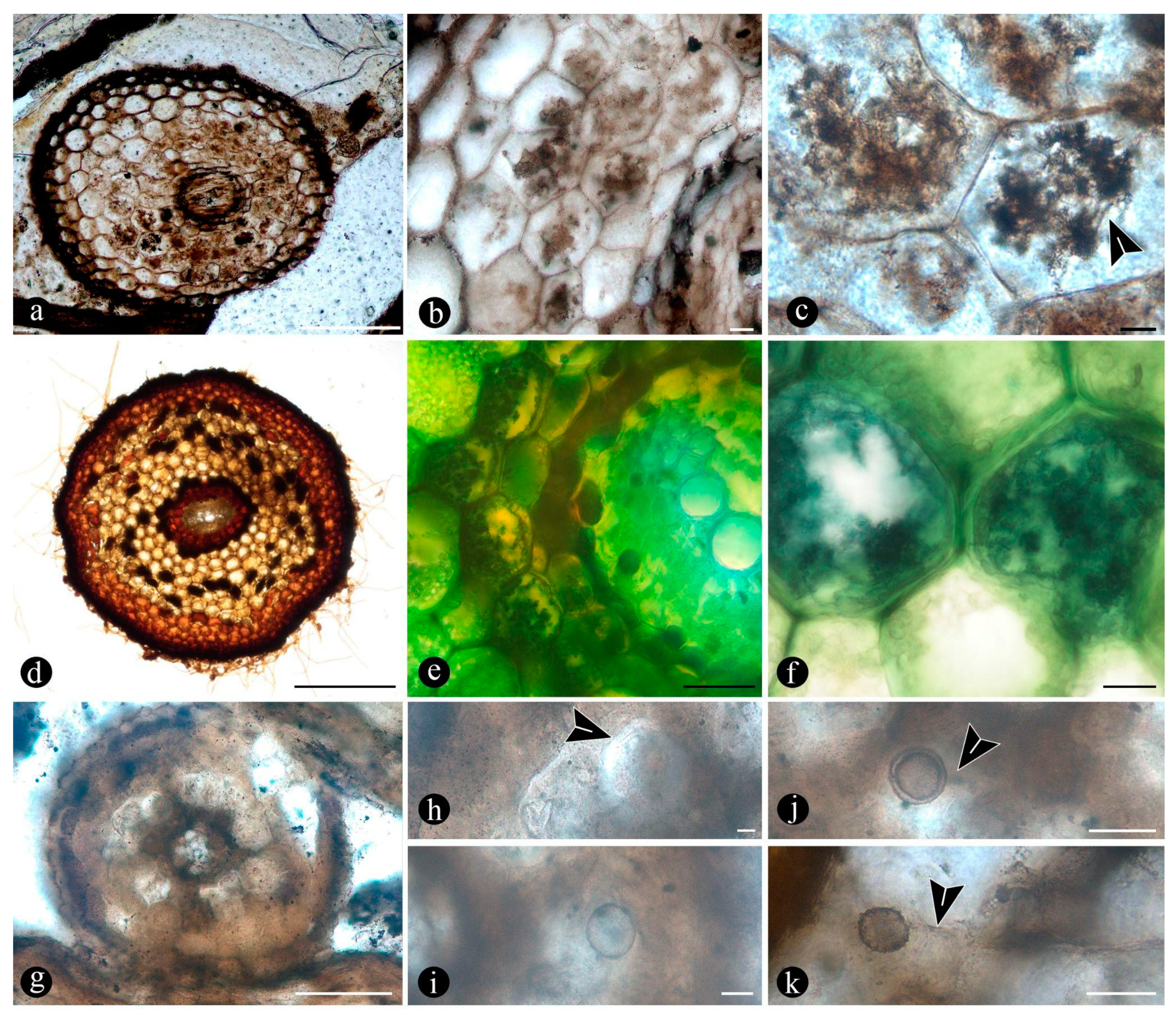
Figure 6.
(a) Silicified adventitious diarch root of Osmundaceae showing the complete decay of its central part, where numerous globose structures are present. (b) Decayed petiole of Osmundaceae with the area of the vascular trace hollowed out and filled by additional globose structures. (c) Close up on the decayed petiole area showing numerous, simple, spherical to pyriform globose structures. (d) Additional globose structures of variable size, texture, and general appearance, included in the decayed areas of a petiole of Osmundaceae. (e) Close up on additional globose structures present among the decayed fern petiole cortex, sometimes singly placed within individual cells. (f) Close up on other fern petiole cortex cells, showing some fully filled by small globose structures. (g) Detail of a bigger globose, to somewhat amoeboid structure displaying 3 symmetrically arranged pores or similar apertures (arrowheads). (h) Detail of another globose structure showing 2 symmetrically arranged, short-necked apertures (arrowheads). (i) Detail of a globose structure with a slightly textured surface and a somewhat pyramidal apophysis (arrowhead). Note also its relatively thicker wall formed by at least two layers. (j) Close up of another globose structure with a rough wall and a thread of minute hyphae or rhizoids (arrowheads) through which is attached to the host petiole cell. (k) Detail of another globose to pyriform structure showing a granulated cytoplasm consisting of small circular to quadrangular subunits. (l) Close up on another globose to amoeboid, collapsed structure, partly disrupted and containing a few hyaline smaller globose structures. Bar = (a-b) 250 µm, (c-l) 15 µm.
Figure 6.
(a) Silicified adventitious diarch root of Osmundaceae showing the complete decay of its central part, where numerous globose structures are present. (b) Decayed petiole of Osmundaceae with the area of the vascular trace hollowed out and filled by additional globose structures. (c) Close up on the decayed petiole area showing numerous, simple, spherical to pyriform globose structures. (d) Additional globose structures of variable size, texture, and general appearance, included in the decayed areas of a petiole of Osmundaceae. (e) Close up on additional globose structures present among the decayed fern petiole cortex, sometimes singly placed within individual cells. (f) Close up on other fern petiole cortex cells, showing some fully filled by small globose structures. (g) Detail of a bigger globose, to somewhat amoeboid structure displaying 3 symmetrically arranged pores or similar apertures (arrowheads). (h) Detail of another globose structure showing 2 symmetrically arranged, short-necked apertures (arrowheads). (i) Detail of a globose structure with a slightly textured surface and a somewhat pyramidal apophysis (arrowhead). Note also its relatively thicker wall formed by at least two layers. (j) Close up of another globose structure with a rough wall and a thread of minute hyphae or rhizoids (arrowheads) through which is attached to the host petiole cell. (k) Detail of another globose to pyriform structure showing a granulated cytoplasm consisting of small circular to quadrangular subunits. (l) Close up on another globose to amoeboid, collapsed structure, partly disrupted and containing a few hyaline smaller globose structures. Bar = (a-b) 250 µm, (c-l) 15 µm.
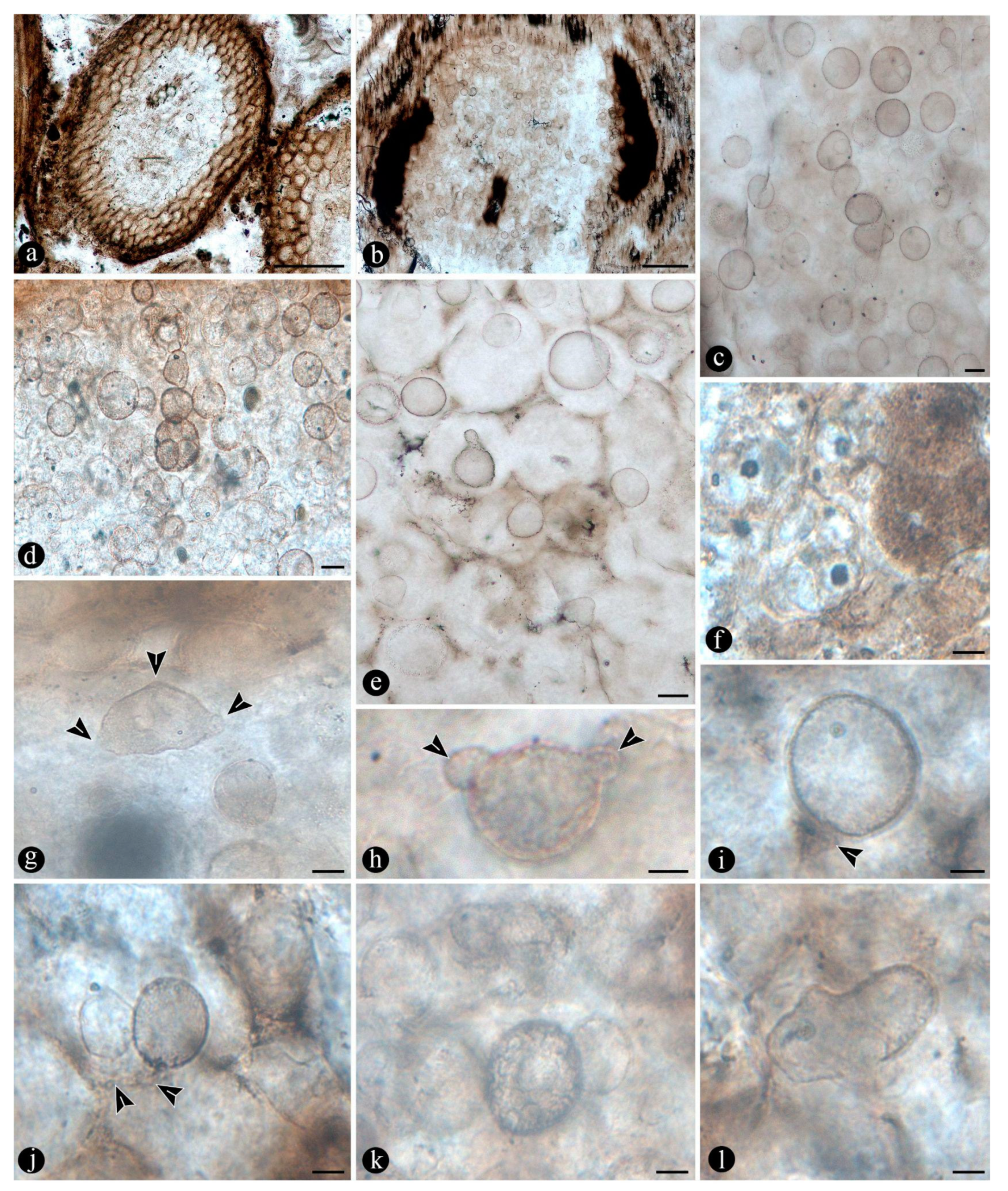
Figure 7.
(a) Close up on a group of globose structures with parts of their bodies missing, bowl-like (left arrow) and tote bag-like (right arrowheads) shaped, respectively. (b) Group of globose structures, including some with shallower parts of their bodies missing (black arrowhead) and with further, smaller globose structures, attached to their outer surfaces (white arrowheads). (c) Detail of additional globose structures with a single or a couple of smaller globose structures within their lumina (arrowheads). (d) Detail of globose structure with several smaller ones (arrowheads) attached to its outer surface. (e) Group of globose structures holding a single smaller unit inside, which partly to almost completely fill in the host’s lumina, sometimes extending a short neck to the outer border connecting to the exterior (arrowheads). Bar = (a-e) 15 µm.
Figure 7.
(a) Close up on a group of globose structures with parts of their bodies missing, bowl-like (left arrow) and tote bag-like (right arrowheads) shaped, respectively. (b) Group of globose structures, including some with shallower parts of their bodies missing (black arrowhead) and with further, smaller globose structures, attached to their outer surfaces (white arrowheads). (c) Detail of additional globose structures with a single or a couple of smaller globose structures within their lumina (arrowheads). (d) Detail of globose structure with several smaller ones (arrowheads) attached to its outer surface. (e) Group of globose structures holding a single smaller unit inside, which partly to almost completely fill in the host’s lumina, sometimes extending a short neck to the outer border connecting to the exterior (arrowheads). Bar = (a-e) 15 µm.
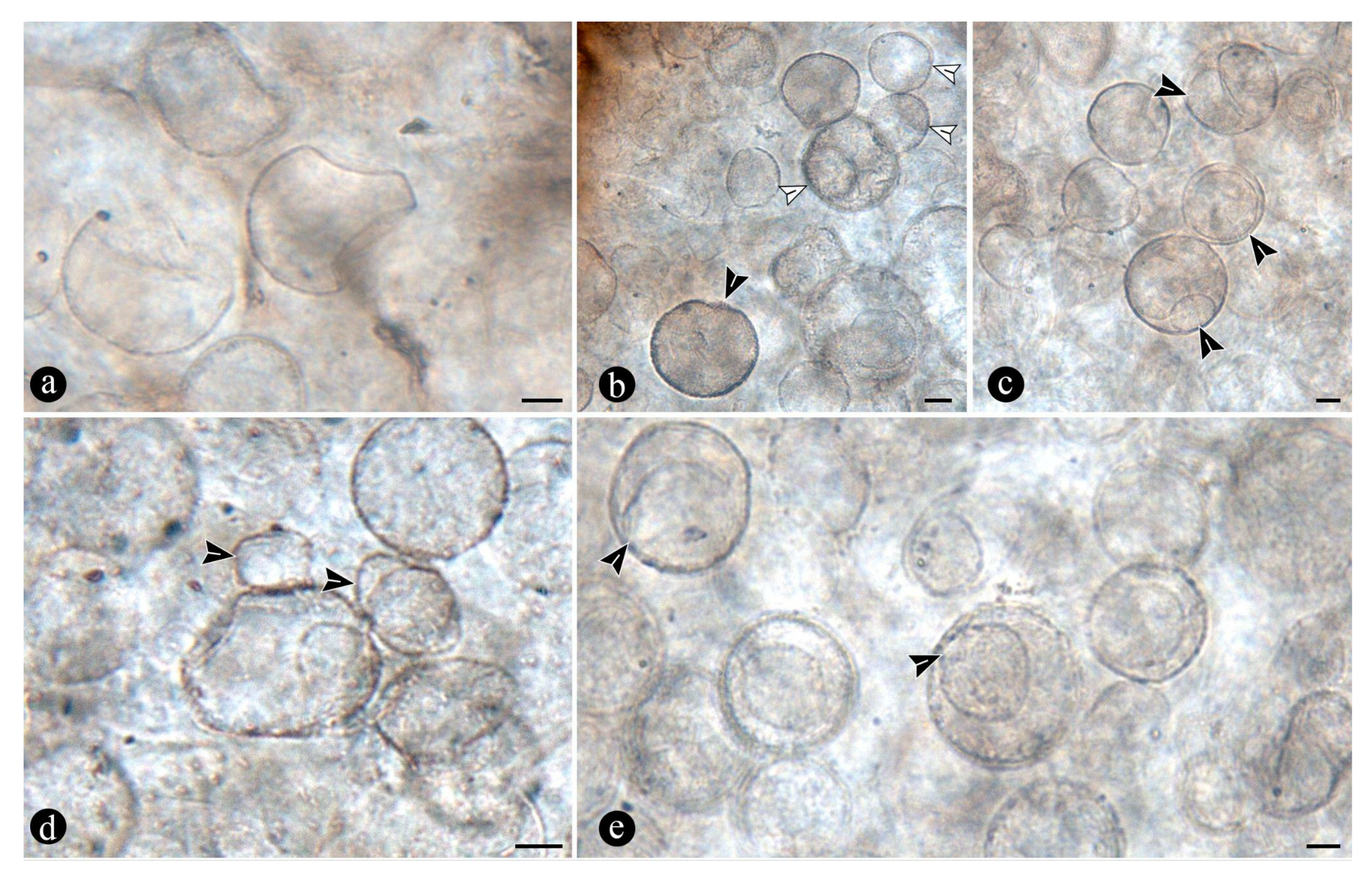
Figure 8.
(a). Filaments in degraded Osmundaceae fern tissues, including petioles, adventitious roots, and various unidentifiable remains. (b) A decayed fern sporangium with thinner, septate filaments on it. (c) Cortical cells of a fern petiole with inter and intracellular, broad, mostly coenocytic filaments. (d) Cortical cells of a fern petiole with filaments of variable width, which are set in the cellular interspaces and intracellularly. (e) Close up on a cortical cell, which is entirely filled by highly contorted, broad filaments that intersect each other, resulting in a reticulate pattern. (f) Cortical cell of fern petiole with closely septate, broad, intracellular filaments consisting of a series of consecutive cylindrical cells. (g) Close up on degraded tissues with branched, septate filaments. (h) Close up on a branched filament in degraded tissue. Note (arrow) septation of the hypha, which gives rise to consecutive cylindrical cells at the distal end of the lateral branch (arrow). (i) Another example of branched filament in degraded fern tissues, divided into plate-like cells (arrow), at the distal end of the lateral branch. (j) Close up on a filament consisting of principal filament with two symmetrical phialide-like, upright branches (arrows). (k) Close up on another filament with a single, phialide-like upright branch (arrow). (l) Close up on a phialide-bearing filament showing a broad hyphopodium (arrow) firmly attached to the host substrate. (m) Smaller, opaque, branched filaments within the lumina of a host petiole cortical cell. Note lateral branches further subdivided into small subunits in a rosary-like pattern. Bar = (a-b) 30 µm, (c-f) 15 µm, (g-m) 10 µm.
Figure 8.
(a). Filaments in degraded Osmundaceae fern tissues, including petioles, adventitious roots, and various unidentifiable remains. (b) A decayed fern sporangium with thinner, septate filaments on it. (c) Cortical cells of a fern petiole with inter and intracellular, broad, mostly coenocytic filaments. (d) Cortical cells of a fern petiole with filaments of variable width, which are set in the cellular interspaces and intracellularly. (e) Close up on a cortical cell, which is entirely filled by highly contorted, broad filaments that intersect each other, resulting in a reticulate pattern. (f) Cortical cell of fern petiole with closely septate, broad, intracellular filaments consisting of a series of consecutive cylindrical cells. (g) Close up on degraded tissues with branched, septate filaments. (h) Close up on a branched filament in degraded tissue. Note (arrow) septation of the hypha, which gives rise to consecutive cylindrical cells at the distal end of the lateral branch (arrow). (i) Another example of branched filament in degraded fern tissues, divided into plate-like cells (arrow), at the distal end of the lateral branch. (j) Close up on a filament consisting of principal filament with two symmetrical phialide-like, upright branches (arrows). (k) Close up on another filament with a single, phialide-like upright branch (arrow). (l) Close up on a phialide-bearing filament showing a broad hyphopodium (arrow) firmly attached to the host substrate. (m) Smaller, opaque, branched filaments within the lumina of a host petiole cortical cell. Note lateral branches further subdivided into small subunits in a rosary-like pattern. Bar = (a-b) 30 µm, (c-f) 15 µm, (g-m) 10 µm.
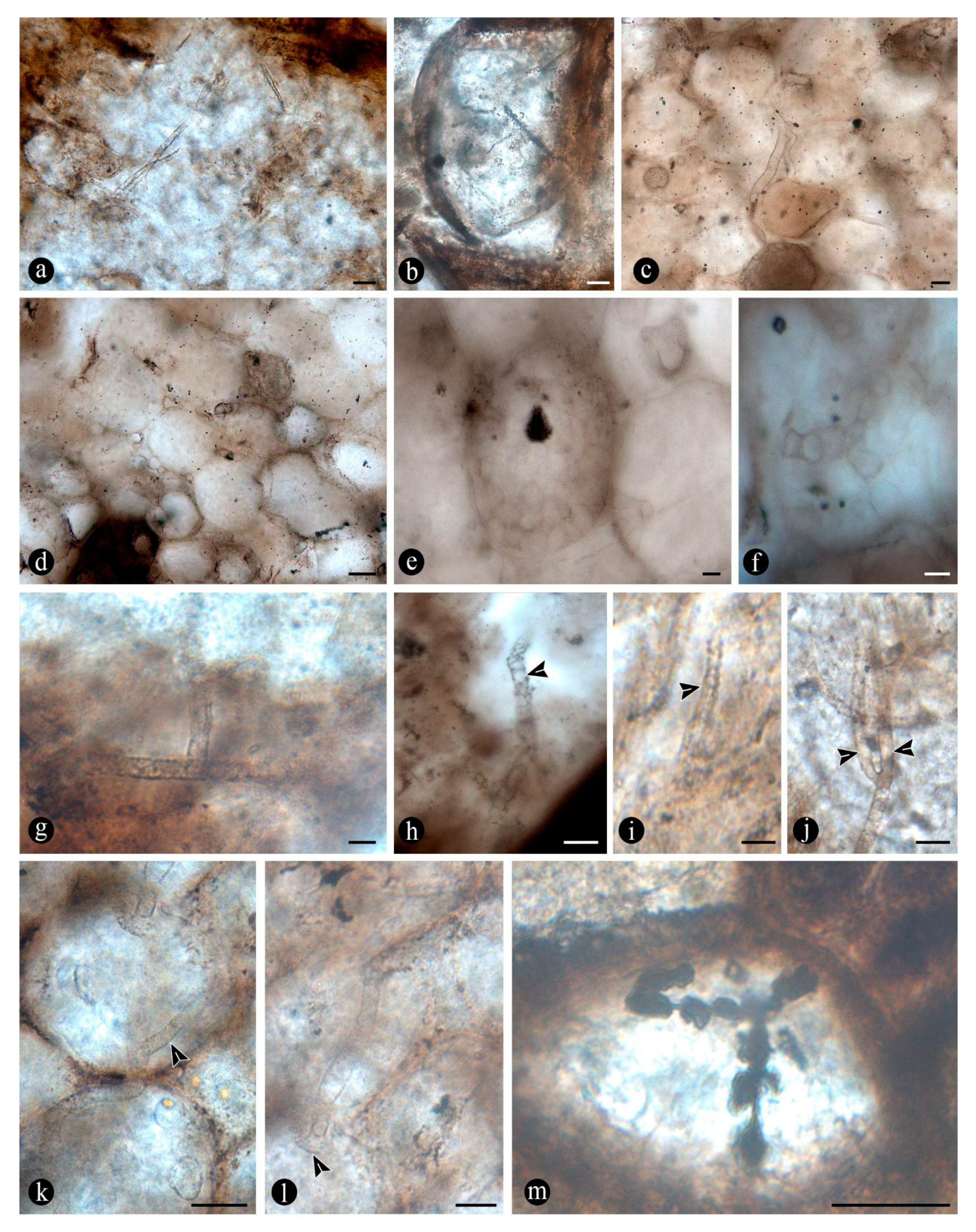
Figure 9.
(a) General view of degraded fern petioles having deformed and variously disrupted cells, sometimes filled with an opaque substance. (b) Close up on some degraded cortex cells including some with their innermost cell wall component (i.e., S3) disrupted and projected inwards into the lumen. (c) Close up on another group of cortex cells showing differential decay of the lignin-rich cell wall components (i.e., middle lamella) and variously deformed and distorted cells. (d) Close up on another group of degraded petiole cells showing decay of all cell wall components, which remain connected through their lignin-rich junctions at the cell corners. (e) Close up on additional degraded petiole showing synchronous decay of all cell wall components, including filaments present directly in contact with the decayed cells. (f) Fern petiole showing decayed area with small globose structures attached to the disrupted tissues. (g) Example of irregularly shaped, decayed areas, some with a characteristic furry texture, on the cortex cells of a decayed fern petiole. (h) Another fern petiole showing a cluster of smaller, regular, approximately circular decay areas, with a characteristic furry, radiating texture. (i) Close up on some of the furry decay areas showing variously eroded cells having a soft surface formed by longitudinally oriented features. (j) Close up on other furry decay areas showing further decayed cells having a soft texture and radiating appearance. (k) Close up on another furry decay area with even hardly recognizable cells, resulting in a soft texture in a radiating pattern. (l) Close up on another furry decay area where individual cells are not recognizable, having a pattern consisting of radially oriented, fusiform features. Bar = (a, g-h) 250 µm, (b-e) 25 µm, (f, i-l) 50 µm.
Figure 9.
(a) General view of degraded fern petioles having deformed and variously disrupted cells, sometimes filled with an opaque substance. (b) Close up on some degraded cortex cells including some with their innermost cell wall component (i.e., S3) disrupted and projected inwards into the lumen. (c) Close up on another group of cortex cells showing differential decay of the lignin-rich cell wall components (i.e., middle lamella) and variously deformed and distorted cells. (d) Close up on another group of degraded petiole cells showing decay of all cell wall components, which remain connected through their lignin-rich junctions at the cell corners. (e) Close up on additional degraded petiole showing synchronous decay of all cell wall components, including filaments present directly in contact with the decayed cells. (f) Fern petiole showing decayed area with small globose structures attached to the disrupted tissues. (g) Example of irregularly shaped, decayed areas, some with a characteristic furry texture, on the cortex cells of a decayed fern petiole. (h) Another fern petiole showing a cluster of smaller, regular, approximately circular decay areas, with a characteristic furry, radiating texture. (i) Close up on some of the furry decay areas showing variously eroded cells having a soft surface formed by longitudinally oriented features. (j) Close up on other furry decay areas showing further decayed cells having a soft texture and radiating appearance. (k) Close up on another furry decay area with even hardly recognizable cells, resulting in a soft texture in a radiating pattern. (l) Close up on another furry decay area where individual cells are not recognizable, having a pattern consisting of radially oriented, fusiform features. Bar = (a, g-h) 250 µm, (b-e) 25 µm, (f, i-l) 50 µm.
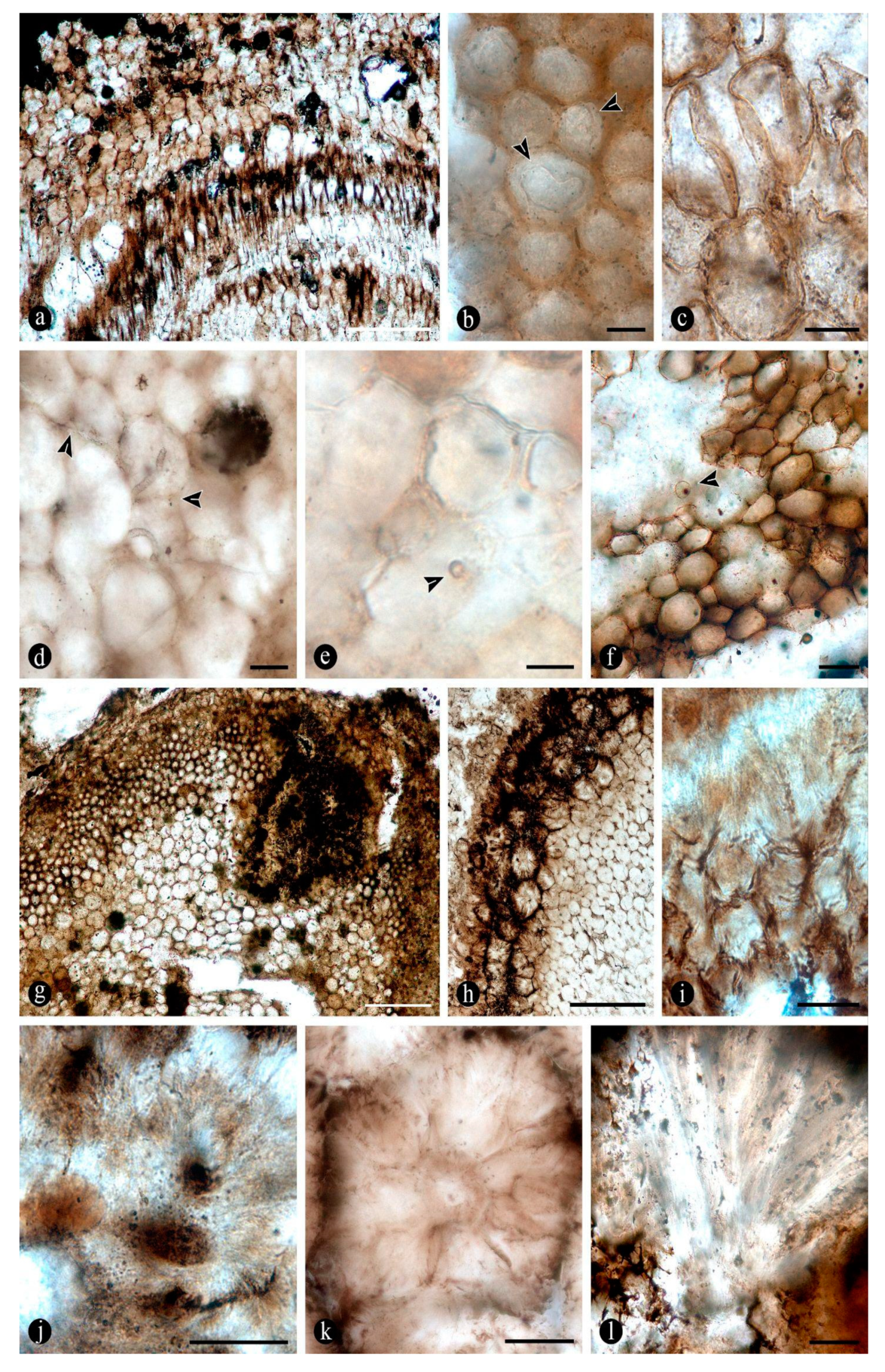
Figure 10.
(a) General view of degraded fern petiole centered in the vascular trace area, which is partly decayed and filled with clusters of small coprolites. (b) General view of a degraded fern petiole having some big coprolites set within a cell-free area in the cortex. (c) View of a degraded adventitious root completely hollowed out and filled with big cylindrical coprolites. (d) Another view of a degraded fern petiole showing an extensive irregular decay area with all tissues consumed away. (e) General view of another decayed fern petiole having an opaque, amorphous, organic residue and some small coprolites in the degraded area. (f) Close up on a group of small coprolites included in a degraded fern petiole showing its soft texture of highly comminuted particles and filaments directly associated with some of them (arrow). (g) Close up on a big coprolite associated with the degraded fern petioles showing globose structures attached to its surface. (h) Close up on a cluster of small coprolites with a characteristic elliptical shape and circular cross section. (i) Close up on a bigger coprolite showing included organic elements of various shapes. Bar = (a) 50 µm, (b-c) 200 µm, (d) 250 µm, (e-i) 100 µm.
Figure 10.
(a) General view of degraded fern petiole centered in the vascular trace area, which is partly decayed and filled with clusters of small coprolites. (b) General view of a degraded fern petiole having some big coprolites set within a cell-free area in the cortex. (c) View of a degraded adventitious root completely hollowed out and filled with big cylindrical coprolites. (d) Another view of a degraded fern petiole showing an extensive irregular decay area with all tissues consumed away. (e) General view of another decayed fern petiole having an opaque, amorphous, organic residue and some small coprolites in the degraded area. (f) Close up on a group of small coprolites included in a degraded fern petiole showing its soft texture of highly comminuted particles and filaments directly associated with some of them (arrow). (g) Close up on a big coprolite associated with the degraded fern petioles showing globose structures attached to its surface. (h) Close up on a cluster of small coprolites with a characteristic elliptical shape and circular cross section. (i) Close up on a bigger coprolite showing included organic elements of various shapes. Bar = (a) 50 µm, (b-c) 200 µm, (d) 250 µm, (e-i) 100 µm.
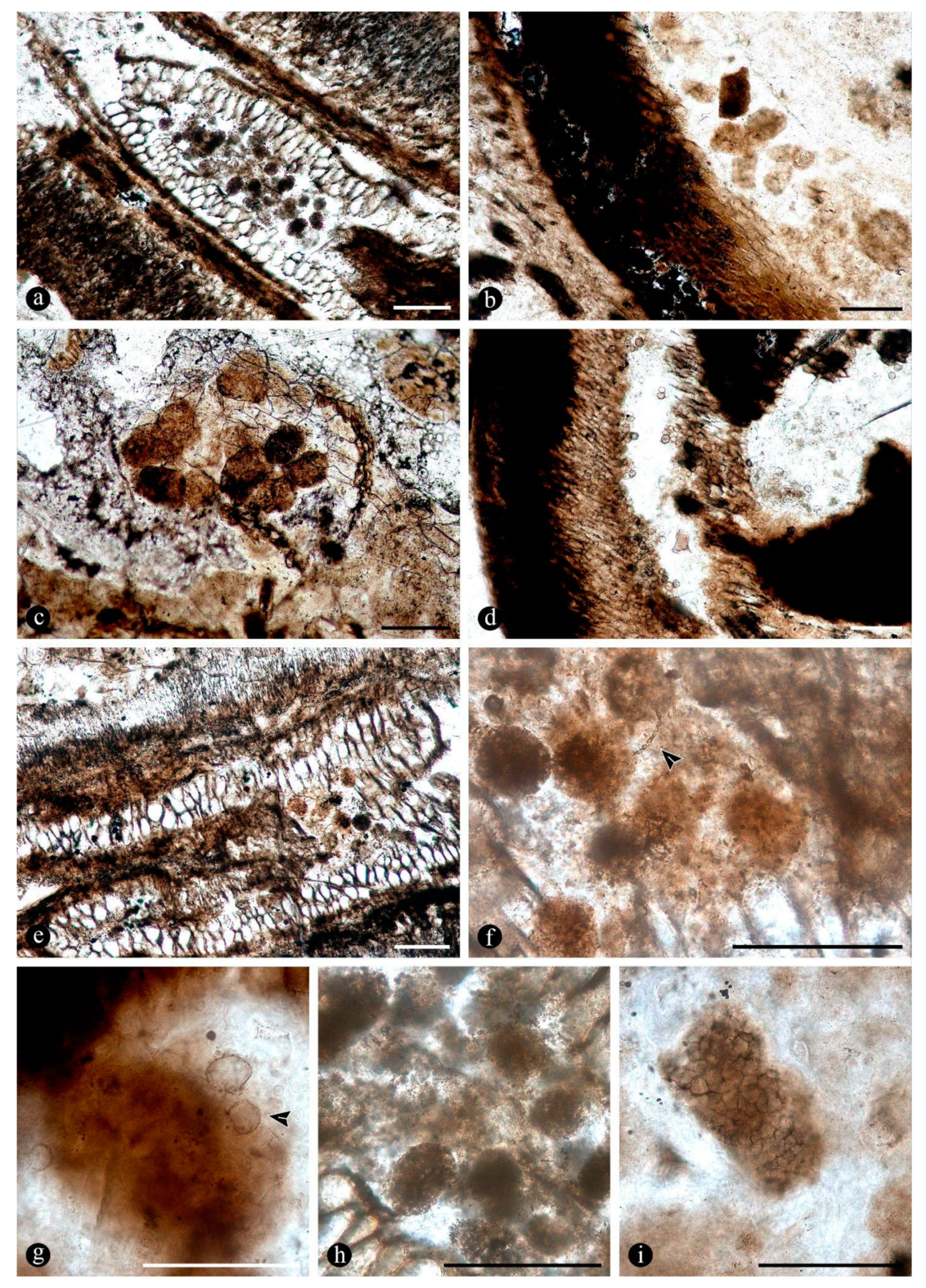
Figure 11.
(a) Globose to elliptical structures of different sizes, with characteristic thick-walled and variously layered walls, encysted in degraded fern tissues. (b) Close up on a pyriform structure encysted in decayed tissues with a multilayered wall and possible opposite broad apertures at each pole. (c) Endobiotic elliptical structure within a fern petiole cortex cell. Note its thick wall and granulated contents. (d) Structure of circular profile in the matrix surrounding the rhizomatous stems. Note characteristic thick wall and approximately trapezoidal external ornaments (arrowheads). (e) Broadly pyriform structure within decayed fern tissues, with a thick and multilayered wall. Note the presence of a centrally placed, hyaline spore (arrow) and broad subtending hypha (arrow). (f) Globose, thick-walled structure ornamented with short spines, embedded within degraded organic debris. Note central structure (arrow). (g) Globose structure with spiny ornamentation, encysted in decayed organic debris. Note the opaque, central spore (arrow). (h) Helicoidal, multicelled fungal spore associated with decayed fern tissues. (i) Multicelled dictyospore preserved along decayed fern tissues. Bar = (a) 100 µm, (b-g) 50 µm, (h) 10 µm, (i) 25 µm.
Figure 11.
(a) Globose to elliptical structures of different sizes, with characteristic thick-walled and variously layered walls, encysted in degraded fern tissues. (b) Close up on a pyriform structure encysted in decayed tissues with a multilayered wall and possible opposite broad apertures at each pole. (c) Endobiotic elliptical structure within a fern petiole cortex cell. Note its thick wall and granulated contents. (d) Structure of circular profile in the matrix surrounding the rhizomatous stems. Note characteristic thick wall and approximately trapezoidal external ornaments (arrowheads). (e) Broadly pyriform structure within decayed fern tissues, with a thick and multilayered wall. Note the presence of a centrally placed, hyaline spore (arrow) and broad subtending hypha (arrow). (f) Globose, thick-walled structure ornamented with short spines, embedded within degraded organic debris. Note central structure (arrow). (g) Globose structure with spiny ornamentation, encysted in decayed organic debris. Note the opaque, central spore (arrow). (h) Helicoidal, multicelled fungal spore associated with decayed fern tissues. (i) Multicelled dictyospore preserved along decayed fern tissues. Bar = (a) 100 µm, (b-g) 50 µm, (h) 10 µm, (i) 25 µm.
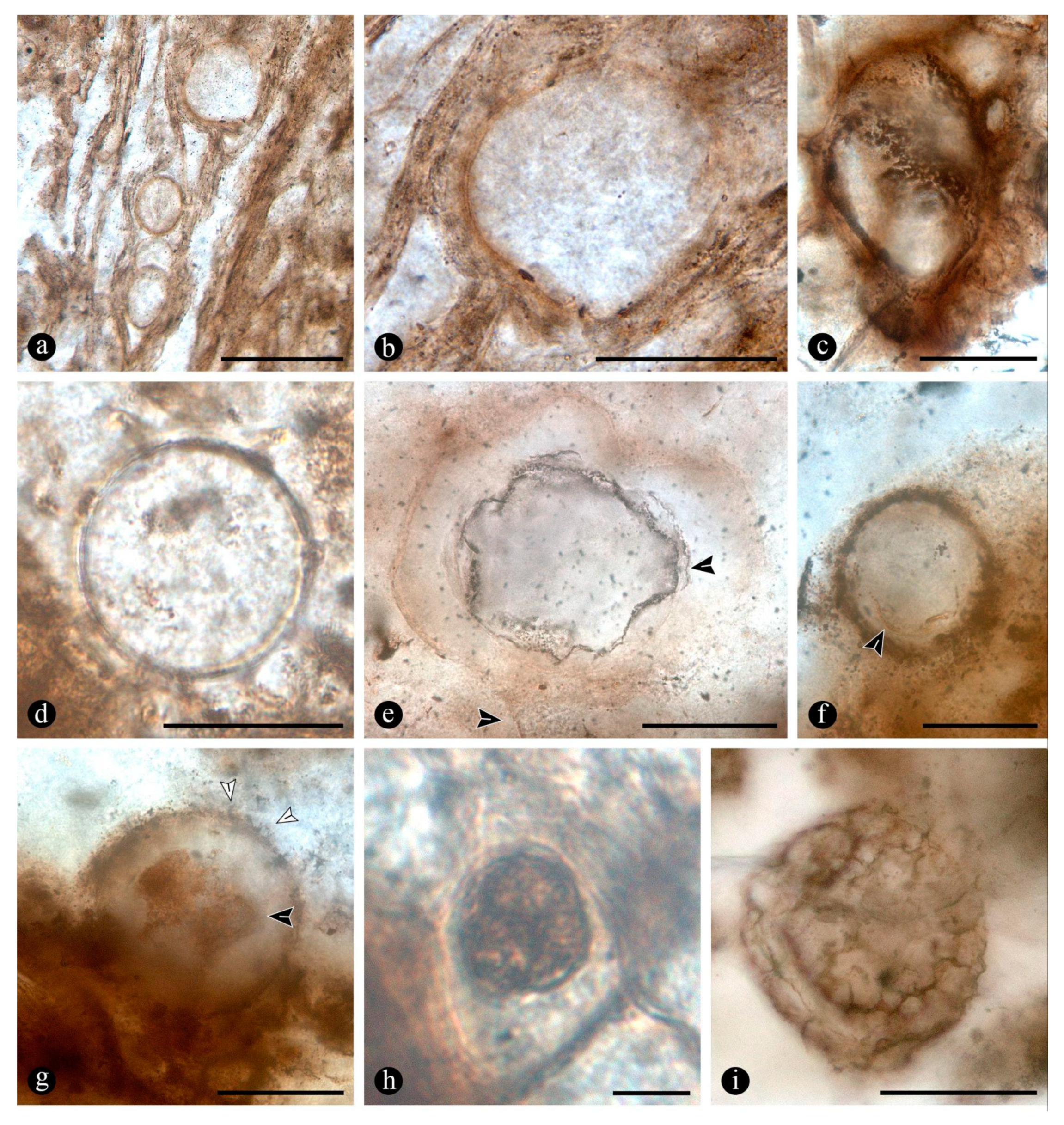
Figure 12.
(a) General view of a section of decayed conifer wood with a typical mottle-pattern with large, irregular decay areas. (b) General view of a section of decayed conifer wood with a mottle-rot pattern, characterized by numerous approximately circular decay areas distributed over sound tissues. (c) Close up on decay areas characterizing the mottles of the mottle-rot patterns showing deformed hyaline xylem cells. (d) Close up on the margins of other mottles showing progressive decay of the xylem cell inwards, towards the center of the decay area. (e) General view of conifer wood with large, irregular decay areas, where xylem tissues are replaced by a homogeneous lightly opaque, mass of disrupted cells along with cellular contents and other organic debris. (f) General view of a decay area with variously distorted cells and massive decay spots filled with a gooey, opaque substance. (g) General view of wood with radially arranged decay features, consisting of regular deposits of an opaque substance along ray cells and tracheids. Note that they are set parallel and in close association with each other. (h) Close up on wood showing xylem cells with their lumina filled by an opaque substance. (i) General view of decayed conifer wood with cells with deposits of an opaque substance aligned with the growth rings. Note the decay areas around the growth rings. Bar = (a) 750 µm, (b, g-i) 500 µm, (c) 250 µm, (d-f) 100 µm.
Figure 12.
(a) General view of a section of decayed conifer wood with a typical mottle-pattern with large, irregular decay areas. (b) General view of a section of decayed conifer wood with a mottle-rot pattern, characterized by numerous approximately circular decay areas distributed over sound tissues. (c) Close up on decay areas characterizing the mottles of the mottle-rot patterns showing deformed hyaline xylem cells. (d) Close up on the margins of other mottles showing progressive decay of the xylem cell inwards, towards the center of the decay area. (e) General view of conifer wood with large, irregular decay areas, where xylem tissues are replaced by a homogeneous lightly opaque, mass of disrupted cells along with cellular contents and other organic debris. (f) General view of a decay area with variously distorted cells and massive decay spots filled with a gooey, opaque substance. (g) General view of wood with radially arranged decay features, consisting of regular deposits of an opaque substance along ray cells and tracheids. Note that they are set parallel and in close association with each other. (h) Close up on wood showing xylem cells with their lumina filled by an opaque substance. (i) General view of decayed conifer wood with cells with deposits of an opaque substance aligned with the growth rings. Note the decay areas around the growth rings. Bar = (a) 750 µm, (b, g-i) 500 µm, (c) 250 µm, (d-f) 100 µm.

Figure 13.
(a) General view of a section of another decayed conifer wood showing traumatic resin ducts of different sizes aligned parallel to the growth rings and discrete spread of an opaque substance that connects them to each other. (b) Close up on a diamond-shaped traumatic resin duct with a characteristic rim of hyaline subsidiary cells and a lumen partly filled by an opaque substance. (c) Close up on a smaller, elliptical traumatic resin duct with an abundant opaque substance distributed around it and completely filling its lumen. (d) Bigger, irregular to diamond shaped traumatic resin duct with several concentrical deposits of an opaque substance around it and with its lumen empty. (e) Medium size empty traumatic resin duct surrounded by an amorphous opaque substance that is continuous all the way to growth rings and local xylem cells among sound tissues. (f) Smaller, elliptical, partly filled traumatic resin duct placed within decayed wood and connected to surrounding xylem cells and growth rings by an opaque substance. (g) General view of traumatic resin ducts and parallel growth rings, connected by an opaque substance, forming a tangential barrier zone. (h) Close up on opaque substances at the barrier zone formed showing variously preserved cells around it. (i) Close on a group of xylem cells showing synchronous decay of all wall components and resulting, cell-free spot (arrowhead). (j) Close up on another group of xylem cells showing differential decay of intermediate cell wall components (arrows) resulting in the progressive detachment of the innermost component (i.e., S3). (k) Close up on a partly disrupted traumatic resin duct showing a series of globose structures trapped among an opaque substance within its lumen (arrowhead). (l) View of a group of variously preserved xylem cells showing the presence of several, few-celled traumatic resin ducts, clustered or closely placed to each other (white arrowheads). (m) Close up on some of the few-celled traumatic resin ducts showing its typical radial arrangement and filling with an opaque substance. Bar = (a) 750 µm, (b-g) 500 µm, (h-i, l-m) 100 µm, (j-k) 50 µm.
Figure 13.
(a) General view of a section of another decayed conifer wood showing traumatic resin ducts of different sizes aligned parallel to the growth rings and discrete spread of an opaque substance that connects them to each other. (b) Close up on a diamond-shaped traumatic resin duct with a characteristic rim of hyaline subsidiary cells and a lumen partly filled by an opaque substance. (c) Close up on a smaller, elliptical traumatic resin duct with an abundant opaque substance distributed around it and completely filling its lumen. (d) Bigger, irregular to diamond shaped traumatic resin duct with several concentrical deposits of an opaque substance around it and with its lumen empty. (e) Medium size empty traumatic resin duct surrounded by an amorphous opaque substance that is continuous all the way to growth rings and local xylem cells among sound tissues. (f) Smaller, elliptical, partly filled traumatic resin duct placed within decayed wood and connected to surrounding xylem cells and growth rings by an opaque substance. (g) General view of traumatic resin ducts and parallel growth rings, connected by an opaque substance, forming a tangential barrier zone. (h) Close up on opaque substances at the barrier zone formed showing variously preserved cells around it. (i) Close on a group of xylem cells showing synchronous decay of all wall components and resulting, cell-free spot (arrowhead). (j) Close up on another group of xylem cells showing differential decay of intermediate cell wall components (arrows) resulting in the progressive detachment of the innermost component (i.e., S3). (k) Close up on a partly disrupted traumatic resin duct showing a series of globose structures trapped among an opaque substance within its lumen (arrowhead). (l) View of a group of variously preserved xylem cells showing the presence of several, few-celled traumatic resin ducts, clustered or closely placed to each other (white arrowheads). (m) Close up on some of the few-celled traumatic resin ducts showing its typical radial arrangement and filling with an opaque substance. Bar = (a) 750 µm, (b-g) 500 µm, (h-i, l-m) 100 µm, (j-k) 50 µm.


Table 1.
Comparative measurements of the rhizome between fossil samples studied in this article and current specimens of Osmunda spectabilis from Misiones (Argentina).
Table 1.
Comparative measurements of the rhizome between fossil samples studied in this article and current specimens of Osmunda spectabilis from Misiones (Argentina).
| Samples* | Distance between individuals from center to center (cm) | Diameter (cm) | Number of petiole cycles | Ramifications measurements (smaller individuals) | Petioles diameter (cm) | Roots diameter (cm) | ||
|---|---|---|---|---|---|---|---|---|
| Ramification | Diameter | Number of petiole cycles | ||||||
| MPM-Pb 16096 | 15 | 7 | 7 | - | - | - | - | - |
| 12 | 9 | - | - | - | - | - | ||
| MPM-Pb 16097 | 20 | 13 | 15 | 1 | 3 | 5 | - | - |
| - | - | - | 2 | 3 | 5 | - | - | |
| - | - | - | 3 | 4 | 6 | - | - | |
| - | 7 | 10 | 1 | - | - | - | - | |
| - | 6 | 8 | 1 | - | - | - | - | |
| MPM-Pb 16084-16085 | - | 14 | 10 | - | - | - | - | - |
| MPM-Pb 16086 | - | 5 | 5 | - | - | - | 1.5-10 | 0.6-1.8 |
| MPM-Pb 16087 | - | 13 | 10 | 1 | - | - | - | - |
| MPM-Pb 16088 | 10 | 10 | - | - | - | 1.8-73 | - | |
| MPM-Pb 16089 | - | 2.5 | 5 | - | - | - | - | - |
| - | 1.5 | 3 | - | - | - | - | - | |
| MPM-Pb 16090 | - | 8 | 9 | - | - | - | - | - |
| MPM-Pb 16091 | - | 14 | 13 | - | - | - | - | - |
| MPM-Pb 16092 | - | - | - | 1 | 1.5 | 3 | - | - |
| MPM-Pb 16093 | - | 16 | 12 | - | - | - | - | - |
| MPM-Pb 16094 | - | 11 | 10 | - | - | - | - | - |
| MPM-Pb 16095 | - | 12 | 9 | - | - | - | - | - |
| Yañez et al. 596 | 6.2 | 4.6-7 | 5-7 | 1 | 1.2 | 2 | 4-10.2 | 0.35-3 |
* Lack of measurement of certain features in some specimens is related with the preservation (e.g., fragmentation or sectioning, orientation of the specimen, incompleteness, deformation), or it is not applicable (e.g., there are no ramifications).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



