
Submitted:
29 November 2024
Posted:
02 December 2024
You are already at the latest version
Abstract
The possibility of CRISPR/Cas gene editing system regulation is an important point for creation of effective molecular biological instrument. Allosteric regulation at the level of guide RNAs is one of possible approach to regulate gene editing system. In this work, we designed chimeric guide RNAs (sgRNA, crRNA, and tracrRNA) containing an aptamer to theophylline in their structure in order to create gene editing systems whose activity can be regulated (increased or decreased) allosterically. Two approaches were used to design the guide RNAs. In the first variant, aptamer sequence was a part of the main elements of guide RNAs. In the second variant, the aptamer sequence was added to the 3'- or 5'-end of the guide RNAs. Changes in Cas9 nuclease activity in the presence of constructed chimeric guide RNAs and the effect of theophylline on the efficiency of model DNA cleavage were studied. Several promising candidates for the role of components of the theophylline-activated and deactivated CRISPR/Cas9 system have been discovered.
Keywords:
theophylline aptamer
; chemical RNA synthesis
; guide RNA
; CRISPR/Cas9 system
; allosteric regulation
1. Introduction
CRISPR/Cas systems are now widely used as universal tools for targeting DNA [1,2,3]. The CRISPR/Cas9 system consists of a Cas9 nuclease and either one relatively long guide RNA (sgRNA) or a pair of guide RNAs (crRNA and tracrRNA). The components of this system can be encoded and delivered into the cell in the form of plasmid constructs or can be synthesized chemically or biochemically and delivered using modern physicochemical methods. The CRISPR/Cas9 system is currently the most popular gene editing system among researchers in the field of molecular biology and genetic engineering. One of the problems of CRISPR systems usage is the cleavage of non-target DNA, the so-called off-target effects, which are detected during the application of this system. Regulatable CRISPR/Cas9 systems, whose activity can be controlled using various stimuli, can be considered as a promising approach to reducing off-target DNA cleavages by Cas9 nuclease.
There are different approaches for the regulation of the CRISPR/Cas9 system at the guide RNA level. Among them are the introduction of G-quadruplexes into the guide RNAs, which change conformation in the presence of certain ligands [4,5], the addition to the structure of motifs capable of forming pH-sensitive triple helices (triplexes) [6,7], introduction of light-removed photoblocking groups or chemical blocking groups removed by exposure to certain chemicals [8,9]. Moreover, aptamers or aptazymes can be introduced to the structure of guide RNAs, providing a possibility to change its spatial structure in the presence of specific ligand [10,11] or self-cleaving from the RNA sequence [12].
Aptamer-driven allosteric regulation is an interesting approach for controlling of the activity of oligonucleotide constructs (ribozymes, DNAzymes, aptazymes) [12,13,14,15]. This concept can be further widened to regulate the activity of RNA-protein ensembles. In particular, the use of aptamer sequence as a part of guide RNA opens up the possibility of allosteric regulation of CRISPR/Cas9 at the RNA level [13,14]. Such systems can be used both to regulate the system when the concentration of certain biomolecules in the system changes, and to detect these biomolecules. For allosteric regulation of the gene editing system at the RNA level, several variants of aptamers’ embedding into sgRNA have been proposed [10,11,16].
The goal of this work was to design aptamer-containing guide RNAs (sgRNA, crRNA and tracrRNA) that change their spatial structure and activity of the CRISPR/Cas system upon binding a specific ligand (theophylline). We have chosen a well-studied aptamer to theophylline for our study because of its high affinity and selectivity for the ligand.
2. Results and Discussion
2.1. Aptamer Selection for Designing Chimeric Guide RNAs
The theophylline-specific aptamer is one of the most studied [17,18,19,20]. Aptamers to theophylline are widely used for engineering of various RNA constructs, the activity of which must be changed in the presence of small molecules [17]. The aptamer contains a region that is directly involved in theophylline binding, surrounded by variable nucleotides (Figure 1). Several variants of RNA aptamers to theophylline have already been published; the most commonly used is the 33-mer aptamer shown in Figure 1B.
For this work, we chose a truncated 25-mer aptamer to theophylline, presented in Figure 1C. Shorter aptamer fits better for embedding into guide RNA sequences, making easier the chemical synthesis of the whole RNA construct.
The binding constant of truncated RNA aptamer to theophylline has not been previously published in the literature. We quantitatively characterized the binding of theophylline to the aptamer by a microscopic thermophoresis (MST) method, which requires the presence of a fluorescent label in one of the components of the complex. The advantages of the (MST) are the ability to register the complex formation directly in solution, the possibility to carry out measurements at a given temperature in the reaction buffer, and small quantity of the aptamer required for measurement.
According to the obtained results, the dissociation constant of the 25-mer RNA aptamer bearing Cy5 label on the 5'-end to theophylline was 15 ± 2 μM (Figure S1, S2). This value lays in a typical range of the dissociation constants for complexes of aptamers with small molecules.
We also wondered if the additional stem in the 33-mer RNA aptamer structure affects its affinity to theophylline (Figure 1B). In this case, we unexpectedly failed to register aptamer-theophylline binding by MST, probably due to higher molecular weight of the aptamer. Therefore, we applied another in-solution binding method, an isothermal titration calorimetry (ITC) technique (Figure S3). The dissociation constant of the 33-mer RNA aptamer to theophylline was determined as 2,6 ± 1,1 μM . This value nicely coincides with KD obtained previously by X. Cui et al. using the biolayer interferometry (1.3 μM) [21].
2.2. Design of Guide RNAs Containing the Theophylline Aptamer
Currently, there are few publications devoted to the engineering of sgRNA molecules supplied by theophylline-specific to create regulatable CRISPR/Cas9 systems [10,11,16]. The authors of these works proposed two alternative approaches to the design of such RNAs. First of them involves insertion of the aptamer sequence into functional motifs of sgRNA (hairpins, loops) [11]. The second approach exploits adding an aptamer and trigger sequences to the 3'-end fragment of the guide sgRNA [10].
We aimed to create two variants of CRISPR/Cas9 systems with theophylline-triggered activation or deactivation. In contrast to previously published studies, we proposed to use crRNA/tracrRNA pair in our design. We also developed aptamer-supplied sgRNAs similar to those described early, but differing by spacer sequence and, in some cases, by aptamer sequence. By analogy with the approaches described in the literature, we designed two types of guide RNAs, containing the aptamer sequence both in the structural elements of the guide RNA or in the 3'- or 5'-terminal fragments.
As a first approach to structure design, we inserted an aptamer RNA into functional motifs of the guide RNA (Figure 2). When designing the structures, we accounted the results of Kundert K. et al. [11]. We selected several variants of aptamer-containing sgRNAs studied in this work that changed the activity of the CRISPR/Cas9 system in the presence of theophylline. Based on them, we designed seven sgRNA variants with an aptamer in different fragments of the guide RNA (Figure 2). Five of them were intended to be activated in the presence of theophylline, and two sgRNAs were constructed to be deactivated. When designing guide RNAs, we used a spacer corresponding to the model DNA-target. Started from these sgRNAs, we designed corresponding pairs of guide crRNAs and tracrRNAs (Table 1, Figure 2). Binding of a specific ligand to the designed chimeric guide RNAs was supposed to cause a structural change that activate or inhibit the entire CRISPR/Cas9 system.
We constructed sgRNA (sgRNA_1, sgRNA_2 and sgRNA_3) and crRNA (crRNA_1 and crRNA_2) containing the aptamer as a part of the upper stem, and sgRNA_4 and tracrRNA_4 with the aptamer inside one of the 3'-terminal hairpins. We expected these constructs to increase the activity of the system in the presence of theophylline. The designed sgRNA_5 and tracrRNA_5 contained an aptamer motif within the nexus; theoretically, the addition of theophylline should reduce the activity of these constructs. Guide RNAs sgRNA_6 and sgRNA_7 contained the aptamer in the 3'-terminal hairpin structure. In this case, theophylline binding with the aptamer was expected to provoke the strand displacement and activation or inactivation of the system.
Table 1 shows the sequences of all designed guide RNAs containing an aptamer inserted into structural fragments, and control parent guide RNAs (sgRNA_c, crRNA_c, tracrRNA_c).
At the second stage, we designed guide RNAs containing an aptamer to theophylline at the 3'-terminus. The design was carried out based on data from the research of Lin B. et al. [10]. The authors proposed a design of a guide RNA consisting of a blocking motif, an aptamer, and a trigger motif. The blocking motif is complementary to the upper stem, and the trigger motif is complementary to part of the blocking motif (Figure 3). In the absence of theophylline, the blocking motif forms a duplex with the upper stem sequence, thus reducing the system activity. Theophylline addition induces the change of the aptamer tertiary structure, causing the blocking motif to leave the upper stem and form a hairpin that links the ends of the aptamer, further stabilized by the trigger motif.
Based on these data, we constructed the sgRNA_8, which differs from the sequence proposed by the authors in [10] in the spacer and aptamer sequence (Table 2, Figure 3). This sgRNA should be inactive without theophylline and change the secondary structure upon ligand addition with the activation of the CRISPR/Cas system.
In addition to sgRNA, we also engineered crRNA and tracrRNA molecules containing an aptamer to theophylline at the 3'-end. The design based on the abovementioned principles of sgRNA design. In the structure of crRNA with a similar sequence at the 3'-end, the blocking motif is complementary to the spacer sequence (crRNA_8). Since tracrRNA lacks an upper stem, we made the blocking motif complementary to the nexus (tracrRNA_8).
In all cases, we used the blocking motif of 18 nucleotides, since the authors of [10], using the guide sgRNA with a blocking motif complementary to the upper stem, showed that the 18-nt length is optimal.
We also designed guide RNAs (crRNA_9 and crRNA_10) containing 25-mer or 31-mer aptamer to theophylline at the 5'-terminus of the RNA, by analogy with [12], where the authors made allosterically regulated guide RNAs with an aptazyme at the 5'-terminus (Table 2, Figure 3).
We combined the approaches from two articles cited above and designed the crRNA containing a blocking motif, an aptamer and a trigger motif (crRNA_9) at the 5'-end. This guide RNA contained a blocking motif complementary to the spacer. In the absence of theophylline, the blocking motif and spacer form a hairpin. We also added a sequence of four uridines between spacer and blocking motif, which forms a hairpin loop, allowing the RNA to make a turn. Another variant, crRNA_10, contained a blocking motif complementary to the upper stem.
Therefore, we have constructed a series of guide RNAs (crRNA, sgRNA, tracrRNA) containing theophylline-binding aptamer motif in various positions. The next stage was the chemical synthesis of guide RNAs.
2.3. Chemical Synthesis of Guide RNAs Containing the Theophylline Aptamer
The length of designed RNAs was in the range from 42 to 145 nucleotides. The RNA chemical synthesis was carried out using 2′-O-TBDMS-protected phosphoramidite of ribonucleotides according to a standard automated solid-phase phosphoramidite method (Figure S5), including deblocking, condensation step, oxidation and capping. As a condensing reagent, we used the Activator 42, which provides high yields and minimal amounts of by-products as compared to other tetrazole derivatives [22,23]. Synthetic protocol was optimized for the synthesis of long RNAs especially for the Biosset RNA/DNA synthesizer. Reaction mixtures were analyzed by denaturing gel electrophoresis in PAGE (Figure S6). All RNAs were purified by a preparative gel electrophoresis in denaturing PAGE.
Next, we estimated the influence of additional motifs in the guide RNAs on the efficiency of DNA cleavage by Cas9 nuclease.
2.4. Cleavage of Model DNA by the Cas9 Nuclease in the Presence of Engineered Guide RNAs
We compared all synthesized guide RNAs in a model DNA cleavage by the Cas9 nuclease. As a DNA target, we used the pBlueScript II SK(−) plasmid containing a well-characterized Psp2 protospacer [1] and a TGG protospacer adjacent motif (PAM). Earlier, we optimized the conditions for cleavage of this plasmid [24,25]. We used a 20-fold excess of the effector complex (guide RNA + Cas9 nuclease) relative to the plasmid, a standard buffer and 1 hour incubation at 37 °C.
The introduction of an aptamer into guide RNAs sequences affected differently on the DNA cleavage efficiency by the Cas9 nuclease (Figure 4). In some cases, the activity of such systems decreased significantly, but for other variants of guide RNA the cleavage extent changed only slightly as compared to the positive control (parent aptamer-free sgRNA_c or a pair of guide RNA crRNA_c/tracrRNA_c).
The extent of plasmid cleavage by the Cas9 nuclease guided by sgRNA decreased in all cases. In particular, sgRNA_4 and sgRNA_5 showed lower cleavage extents than the control (Figure 4A), as well as tracrRNA_4 and tracrRNA_5 (Figure 4B). DNA cleavage in the presence of crRNA_1 and crRNA_2 was significantly lower than in the presence of corresponding sgRNA_1 and sgRNA_2. We hypothesized that this effect could be explained by the fact that in these systems, the aptamer was introduced into the upper stem of the sgRNA, which corresponds to the site of interaction between crRNA and tracrRNA. The systems with two guide RNAs do not contain this particular hairpin, so the aptamer was introduced into the crRNA, in the same site and a standard fragment was added to the 3'-end for interaction of crRNA with tracrRNA. The disruption of crRNA/tracrRNA complex has lead to sufficient decrease of DNA cleavage.
On the contrary, sgRNA_8 and tracrRNA_8, similar in structure (Figure 3), carrying an aptamer sequence at the 3'-end, had the same activity that corresponded approximately to the half of the original one. For all crRNA variants (crRNA_8, crRNA_9, crRNA_10) containing an aptamer at the 5'- or 3'-end, we did not observe a significant decrease in activity.
Therefore, in the majority cases, we registered a decrease in the extent of target DNA cleavage by Cas9 nuclease in the presence of guide RNAs containing a theophylline aptamer. Next, it was necessary to study the effect of theophylline on the cleavage of target DNA by Cas9 nuclease in the presence of guide RNAs containing an aptamer to theophylline.
2.5. Effect of Theophylline on DNA Cleavage by Cas9 Nuclease in the Presence of Engineered Guide RNAs
A comparative study of the effects of theophylline on DNA cleavage by Cas9 nuclease in the presence of theophylline-driven guide RNAs was also carried out using the same plasmid. The cleavage was carried out under conditions described above. Theophylline was added at the stage of assembly of the effector complex, prior to the addition of the Cas9. To form the theophylline/aptamer complex, guide RNAs were annealed in the presence of theophylline.
For the two types of designed guide RNAs, in most cases we expected an increase of extent of target DNA cleavage by Cas9 nuclease upon theophylline addition.
Figure 5A shows the extent of DNA cleavage by the systems with sgRNAs containing theophylline aptamer. According to the results, only sgRNA_5, sgRNA_6, and sgRNA_8 provided a significant change of activity upon theophylline addition. The sgRNA_5 and sgRNA_7 guided systems in the presence of theophylline showed a loss of activity by 21-27%, which coincides with the literature data. As we expected, the system with sgRNA_6 increased the activity for 30% upon the addition of theophylline.
Figure 5B shows the efficiencies of DNA cleavage by the systems with guide crRNAs and tracrRNA. In the presence of theophylline, crRNA_2, crRNA_8, crRNA_9, crRNA_10 and tracrRNA_4 did not show a significant change in activity. The crRNA_1 and tracrRNA_8 system showed a slight increase in activity of 7% and 9%, correspondingly, with the addition of theophylline, while the tracrRNA_5 system demonstrated a 20% loss of activity.
3. Materials and Methods
3.1. Chemicals, Enzymes and Plasmids
The 5'-O-(4,4'-dimethoxytrytyl), 2'-O-tert-Butyldimethylsilyl, N-acetyl protected ribonucleoside-3'-phosphoramidite (ChemGenes, USA) and polymers with the first immobilized nucleoside (GlenResearch, USA) were used for oligoribonucleotide synthesis on an ASM-800 DNA/RNA synthesizer (Bioset, Novosibirsk, Russia). Theophylline, triethylamine, triethylamine trihydrofluoride, ethoxytrimethylsilane, Activator 42 were purchased from Sigma-Aldrich (Sigma-Aldrich, USA). Recombinant Cas9 endonuclease and pBS2SKM Psp2 plasmid were obtained according to the published protocols [26]. The Cas9/gRNA (sgRNA or crRNA/tracrRNA pair) complex was assembled by adding Cas9 nuclease (0.27 pmole) to the preliminary annealed sgRNA or crRNA/tracrRNA duplex (0.27 pmole) in the cleavage buffer containing 20 mM HEPES–KOH (pH 7.5), 100 mM KCl, 2 mM MgCl2, 1 mM DTT, 0.5 mM Na2EDTA, and 5% glycerol. The mixture was vortexed, kept for 15 min at 37°C, and used immediately.
3.2. Guide RNA Synthesis
Guide RNA synthesis was carried out on an ASM-800 DNA/RNA synthesizer (Biosset, Novosibirsk, Russia) using the protocols optimized for this instrument. The 0.1 M solutions of ribonucleotide phosphoramidites in an absolute acetonitrile were used for the synthesis. The condensation was carried out using 0.25 M solution of the Activator 42 (Sigma, USA) in absolute acetonitrile during 7 min. The last 5′-O-dimetoxytrityl group was removed and oligonucleotides were deprotected by the 40% aqueous methylamine for 15 min at 65° and evaporation to solid residue, followed by the treatment with a NMP:TEA:TEA·3HF mixture for 1.5 h at 65°C, addition of ethoxytrimethylsilane and precipitation with ethyl ether. All guide RNA were purified by denaturating PAGE.
3.3. Plasmid Digestion by CRISPR/Cas9
Plasmid cleavage reactions were carried out by analogy with [24]. Briefly, a 20-fold molar excess of the effector complex was added to 100 ng of the plasmid and incubated for 1 h at 37°C in 10 μl of the buffer containing 20 mM HEPES–KOH (pH 7.5), 100 mM KCl, 2 mM MgCl2, 1 mM DTT, 0.5 mM Na2EDTA, and 5% glycerol. The reaction was stopped by addition of 2.5 µL of quenching solution (250 mM Na2EDTA, 1.2% SDS, 0.01% bromophenol blue in 30% glycerol). Cleavage products were analyzed by 1% agarose gel electrophoresis with subsequent ethidium bromide staining and visualizing using the E-Box-CX5 gel documentation system (Vilber Lourmat, France) and quantification with Quantity One v4.6.8 software (Bio-Rad Laboratories, Hercules, CA, USA). The fraction of cleaved plasmid (N) was calculated using the equation
3.4. Investigation of Theophylline Influence to DNA Digestion
To estimate the influence of theophylline to the extent of DNA digestion, plasmid cleavage reactions were carried out using gRNA or crRNA/tracrRNA annealed in the presence of 1 mM theophylline. The next steps of effector complex assemblage and plasmid digestion were caried out as described early.
3.5. Cy5-Labelled Aptamer Preparation
3'-Amino modified aptamer to theophylline 5'-AUACCAGCCGAAAGGCCCUUGGCAG-(CH2)6NH2 was synthesized as described above (Section 3.2) using amino-modified CPG support (3'-PT-Amino-Modifier C6 CPG, Glen Research, USA). NHS ester of Cy5 fluorophore (Lumiprobe, Moscow, Russia) was used for the 3'-modification by the manufacturer’s protocols for this reagent.
3.6. Determination of Dissociation Constant of Aptamer to Theophylline by Microscale Thermophoresis (MST) Assay
MST experiments were conducted using a Monolith NT.115 instrument (NanoTemper Technologies, Germany). The Cy5-labelled aptamer (10 nM, 100 μL) was mixed with a series of concentrations of theophylline in a buffer (20 mM HEPES-KOH (pH 7,5), 100 mM KCl, 1 mM DTT, 0.5 mM Na2EDTA, 2 mM MgCl2, 5% glicerol) annealed (2 min, 90° C) and place in MST-grade capillaries. Dissociation constant (Kd values) was calculated based on changes in the thermophoretic movement of the fluorescent aptamer in response to a gradient of infrared laser-induced temperature change. Each capillary was measured in triplicate to ensure reproducibility and accuracy.
3.7. Determination of Dissociation Constant of Aptamer to Theophylline by Isothermal Titration Calorimetry (ITC) Technique
The interaction of theophylline with the aptamer (5'-GGCGAUACCAGCCGAAAGGCCCUUGGCAGCGUC-3') was analyzed using the Nano ITC isothermal titration calorimeter (TA Instruments, New Castle, DE, USA). All ITC measurements were carried out at 25 °C with 300 rpm stirring and 300 s delay between successive injections at 2 μL using a syringe 50 μL in volume to the sample cell (190 μL). Measurements were run in the overfilled mode to prevent air bubbles, liquid evaporation or the presence of the vapor phase. All experiments were performed in the buffer containing 20 mM HEPES–KOH (pH 7.5), 100 mM KCl, 2 mM MgCl2, 1 mM DTT, 0.5 mM Na2EDTA, and 5% glycerol.
During the key experiments, the cell contained the aptamer at concentration 5 or 10 μM. The theophylline at a concentration of 100 μM was gradually added to the cell. In the qualitive experiment, the cell contained aptamer at concentration 10 μM to which theophylline in solution 1 mM was added.
We obtained the parameters of binding theophylline with aptamer (enthalpy of binding, stoichiometry, constant of association).
The data was processed with the NanoAnalyze software (TA Instruments, USA). Models of independent binding were applied to determine thermodynamic parameters and stoichiometry of binding.
3.8. Statistical Treatment
Each experiment was repeated at least three times. Statistical analysis was carried out using GraphPad Prism 7.00 software (GraphPad). One-way ANOVA was used to estimate the significance of the differences as indicated in figure legends. The differences were considered significant at p < 0.05.
4. Conclusions
We developed new guide RNAs for CRIPSR/Cas9 system (sgRNA, crRNA and tracrRNA) containing the theophylline-binding aptamer, having in mind the possibility to make the system theophylline-regulatable. For the first time, we designed and synthesized chimeric guide crRNAs and tracrRNAs with theophylline aptamer motifs. We exploited two approaches to the design of chimeric guide RNA: (1) an introduction of the aptamer into the structural elements of guide RNA and (2) addition of aptamer sequence to the 3'- or 5'-terminal fragments to restrict the possibility of effector complex formation before theophylline addition. Short 25-mer aptamer to theophylline was introduced into the chimeric guide RNA. An ability of this aptamer to bind theophylline was proved by microscopic thermophoresis (MST) assay. The addition of stem to the aptamer structure increases the affinity of aptamer to theophylline. The differences in molecular mass and structure of these two aptamers does not permit to use the same method to analyze affinity of second aptamer to theophylline. In this case we used isothermal titration calorimetry (ITC) method.
Six variants of obtained constructions change the activity of CRISPR/Cas9 system in the presence of theophylline. Three of them decrease the extent of model plasmid cleavage and other three variants, inversely, increase the extent of cleavage.
To summarize, several promising candidates for the role of components of the theophylline-activated and deactivated CRISPR/Cas9 system have been discovered. In further studies they will be used to create effective CRISPR/Cas9 systems that respond to the presence of theophylline.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org., Figure S1: Scheme of the synthesis of 3'-Cy5-labelled theophylline aptamer; Figure S2: MST data and theoretical curve of theophylline binding to the aptamer; Figure S3: ITC data and theoretical curve of theophylline binding to the 33-mer aptamer; Figure S4: Possible duplex homodimer formation by 25-mer aptamer to theophylline; Figure S5: Scheme of solid phase phosphoramidite RNA synthesis; Figure S6: Electrophoregram of reaction mixtures and isolated guide RNA; Figure S7: The example of electrophoregram of plasmid cleavage products; Figure S8: Supposed mechanism of theophylline interaction with chimeric guide RNA with theophylline aptamer; Figure S9: Supposed mechanism of theophylline interaction with CRISPR/Cas9 system containing chimeric guide RNA with theophylline aptamer.
Author Contributions
Conceptualization, D.N..; methodology, D.N. and M.M.; validation, O.D., A.E. and V.G.; investigation, O.D.; writing—original draft preparation, O.D.; writing—review and editing, D.N. and M.V.; supervision, M.V. All authors have read and agreed to the published version of the manuscript.
Funding
The reported study was funded by RSF, project number 22-14-00294. The works for support of DNA/RNA synthesizer in active status were funded by the Russian state-funded project for ICBFM SB RAS (grant number 121031300042-1).
Data Availability Statement
There are no available data.
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable Dual-RNA–Guided DNA Endonuclease in Adaptive Bacterial Immunity. Science (1979) 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Y.; Pausch, P.; Doudna, J.A. Structural Biology of CRISPR–Cas Immunity and Genome Editing Enzymes. Nat Rev Microbiol 2022, 20, 641–656. [Google Scholar] [CrossRef]
- Makarova, K.S.; Wolf, Y.I.; Iranzo, J.; Shmakov, S.A.; Alkhnbashi, O.S.; Brouns, S.J.J.; Charpentier, E.; Cheng, D.; Haft, D.H.; Horvath, P.; et al. Evolutionary Classification of CRISPR–Cas Systems: A Burst of Class 2 and Derived Variants. Nat Rev Microbiol 2020, 18, 67–83. [Google Scholar] [CrossRef]
- Deng, H.; Xu, H.; Wang, Y.; Jia, R.; Ma, X.; Feng, Y.; Chen, H. G-Quadruplex-Based CRISPR Photoswitch for Spatiotemporal Control of Genomic Modulation. Nucleic Acids Res 2023, 51, 4064–4077. [Google Scholar] [CrossRef]
- Liu, X.; Cui, S.; Qi, Q.; Lei, H.; Zhang, Y.; Shen, W.; Fu, F.; Tian, T.; Zhou, X. G-Quadruplex-Guided RNA Engineering to Modulate CRISPR-Based Genomic Regulation. Nucleic Acids Res 2022, 50, 11387–11400. [Google Scholar] [CrossRef]
- Thompson, I.A.P.; Zheng, L.; Eisenstein, M.; Soh, H.T. Rational Design of Aptamer Switches with Programmable PH Response. Nat Commun 2020, 11, 2946. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, Y.; Zhu, J.; Su, X.; Lin, X.; Xu, L.; Xing, X. Employing PH-Responsive RNA Triplex to Control CRISPR/Cas9-Mediated Gene Manipulation in Mammalian Cells. Chinese Chemical Letters 2024, 35, 109427. [Google Scholar] [CrossRef]
- Wang, S.-R.; Wu, L.-Y.; Huang, H.-Y.; Xiong, W.; Liu, J.; Wei, L.; Yin, P.; Tian, T.; Zhou, X. Conditional Control of RNA-Guided Nucleic Acid Cleavage and Gene Editing. Nat Commun 2020, 11, 91. [Google Scholar] [CrossRef]
- Habibian, M.; McKinlay, C.; Blake, T.R.; Kietrys, A.M.; Waymouth, R.M.; Wender, P.A.; Kool, E.T. Reversible RNA Acylation for Control of CRISPR–Cas9 Gene Editing. Chem Sci 2020, 11, 1011–1016. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.; An, Y.; Meng, L.; Zhang, H.; Song, J.; Zhu, Z.; Liu, W.; Song, Y.; Yang, C. Control of CRISPR-Cas9 with Small Molecule-Activated Allosteric Aptamer Regulating SgRNAs. Chemical Communications 2019, 55, 12223–12226. [Google Scholar] [CrossRef] [PubMed]
- Kundert, K.; Lucas, J.E.; Watters, K.E.; Fellmann, C.; Ng, A.H.; Heineike, B.M.; Fitzsimmons, C.M.; Oakes, B.L.; Qu, J.; Prasad, N.; et al. Controlling CRISPR-Cas9 with Ligand-Activated and Ligand-Deactivated SgRNAs. Nat Commun 2019, 10, 2127. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Hu, J.H.; Liu, D.R. Aptazyme-Embedded Guide RNAs Enable Ligand-Responsive Genome Editing and Transcriptional Activation. Nat Commun 2017, 8, 15939. [Google Scholar] [CrossRef]
- Shanidze, N.; Lenkeit, F.; Hartig, J.S.; Funck, D. A Theophylline-Responsive Riboswitch Regulates Expression of Nuclear-Encoded Genes. Plant Physiol 2020, 182, 123–135. [Google Scholar] [CrossRef]
- Harding, B.I.; Pollak, N.M.; Stefanovic, D.; Macdonald, J. Complexing Deoxyribozymes with RNA Aptamers for Detection of the Small Molecule Theophylline. Biosens Bioelectron 2022, 198, 113774. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Jensen, M.C.; Smolke, C.D. Genetic Control of Mammalian T-Cell Proliferation with Synthetic RNA Regulatory Systems. Proceedings of the National Academy of Sciences 2010, 107, 8531–8536. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Y.; Lin, J.; Xu, L. Theophylline-Induced Synergic Activation of Guide RNA to Control CRISPR/Cas9 Function. Chemical Communications 2021, 57, 5418–5421. [Google Scholar] [CrossRef]
- Wrist, A.; Sun, W.; Summers, R.M. The Theophylline Aptamer: 25 Years as an Important Tool in Cellular Engineering Research. ACS Synth Biol 2020, 9, 682–697. [Google Scholar] [CrossRef] [PubMed]
- ZIMMERMANN, G.R.; WICK, C.L.; SHIELDS, T.P.; JENISON, R.D.; PARDI, A. Molecular Interactions and Metal Binding in the Theophylline-Binding Core of an RNA Aptamer. RNA 2000, 6, S1355838200000169. [Google Scholar] [CrossRef]
- Cui, X.; Song, M.; Liu, Y.; Yuan, Y.; Huang, Q.; Cao, Y.; Lu, F. Identifying Conformational Changes of Aptamer Binding to Theophylline: A Combined Biolayer Interferometry, Surface-Enhanced Raman Spectroscopy, and Molecular Dynamics Study. Talanta 2020, 217, 121073. [Google Scholar] [CrossRef] [PubMed]
- Jenison, R.D.; Gill, S.C.; Pardi, A.; Polisky, B. High-Resolution Molecular Discrimination by RNA. Science (1979) 1994, 263, 1425–1429. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Song, M.; Liu, Y.; Yuan, Y.; Huang, Q.; Cao, Y.; Lu, F. Identifying Conformational Changes of Aptamer Binding to Theophylline: A Combined Biolayer Interferometry, Surface-Enhanced Raman Spectroscopy, and Molecular Dynamics Study. Talanta 2020, 217, 121073. [Google Scholar] [CrossRef] [PubMed]
- Threlfall, R.N.; Torres, A.G.; Krivenko, A.; Gait, M.J.; Caruthers, M.H. Synthesis and Biological Activity of Phosphonoacetate- and Thiophosphonoacetate-Modified 2′-O-Methyl Oligoribonucleotides. Org. Biomol. Chem. 2012, 10, 746–754. [Google Scholar] [CrossRef]
- Wei, X. Coupling Activators for the Oligonucleotide Synthesis via Phosphoramidite Approach. Tetrahedron 2013, 69, 3615–3637. [Google Scholar] [CrossRef]
- Sakovina, L.; Vokhtantsev, I.; Vorobyeva, M.; Vorobyev, P.; Novopashina, D. Improving Stability and Specificity of CRISPR/Cas9 System by Selective Modification of Guide RNAs with 2′-Fluoro and Locked Nucleic Acid Nucleotides. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Semikolenova, O.; Sakovina, L.; Akhmetova, E.; Kim, D.; Vokhtantsev, I.; Golyshev, V.; Vorobyeva, M.; Novopashin, S.; Novopashina, D. Photoactivatable Nanocrispr/Cas9 System Based on Crrna Reversibly Immobilized on Carbon Nanoparticles. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Anders, C.; Jinek, M. In Vitro Enzymology of Cas9. In Methods Enzymol.; Doudna, J.A., Sontheimer, E.J., Eds.; 2014; Vol. 546, pp. 1–20.
- Shubsda, M.F.; Goodisman, J.; Dabrowiak, J.C. Quantitation of Ethidium-Stained Closed Circular DNA in Agarose Gels. J Biochem Biophys Methods 1997, 34, 73–79. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Structure of theophylline-specific RNA aptamers. (A) RNA aptamer with variable regions [20]. (B) The most studied RNA aptamer to theophylline [18]. (С) Shortened 25-mer RNA aptamer to theophylline. The blue box indicates the region involved in the ligand binding.
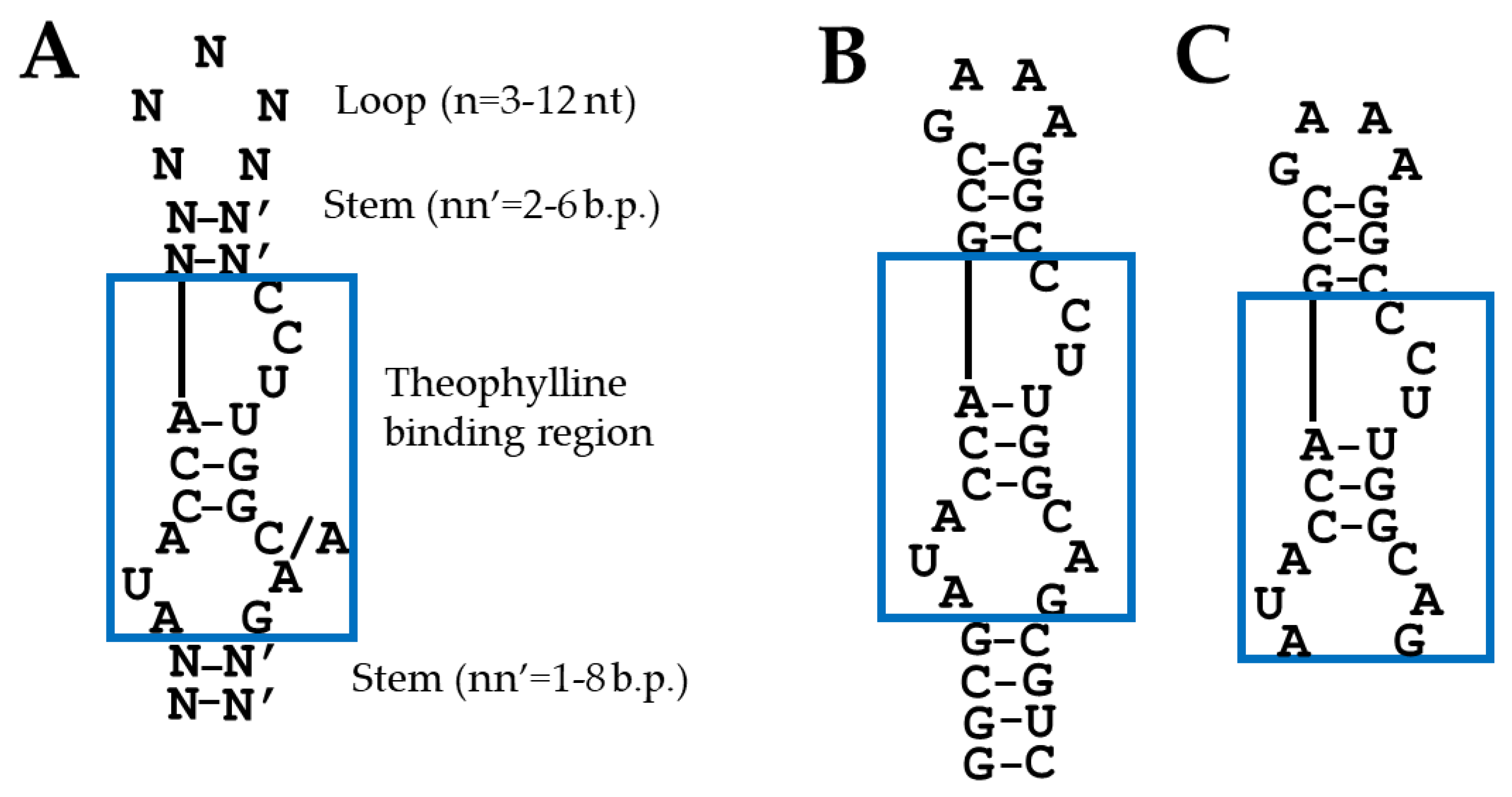
Figure 2.
Structure of designed guide RNAs contained theophylline aptamer into functional motifs. The purple color indicates spacer sequence, grey rose indicates upper stem or contact between crRNA and tracrRNA, green color indicated nexus and hairpins, orange color indicates theophylline aptamer.
Figure 2.
Structure of designed guide RNAs contained theophylline aptamer into functional motifs. The purple color indicates spacer sequence, grey rose indicates upper stem or contact between crRNA and tracrRNA, green color indicated nexus and hairpins, orange color indicates theophylline aptamer.
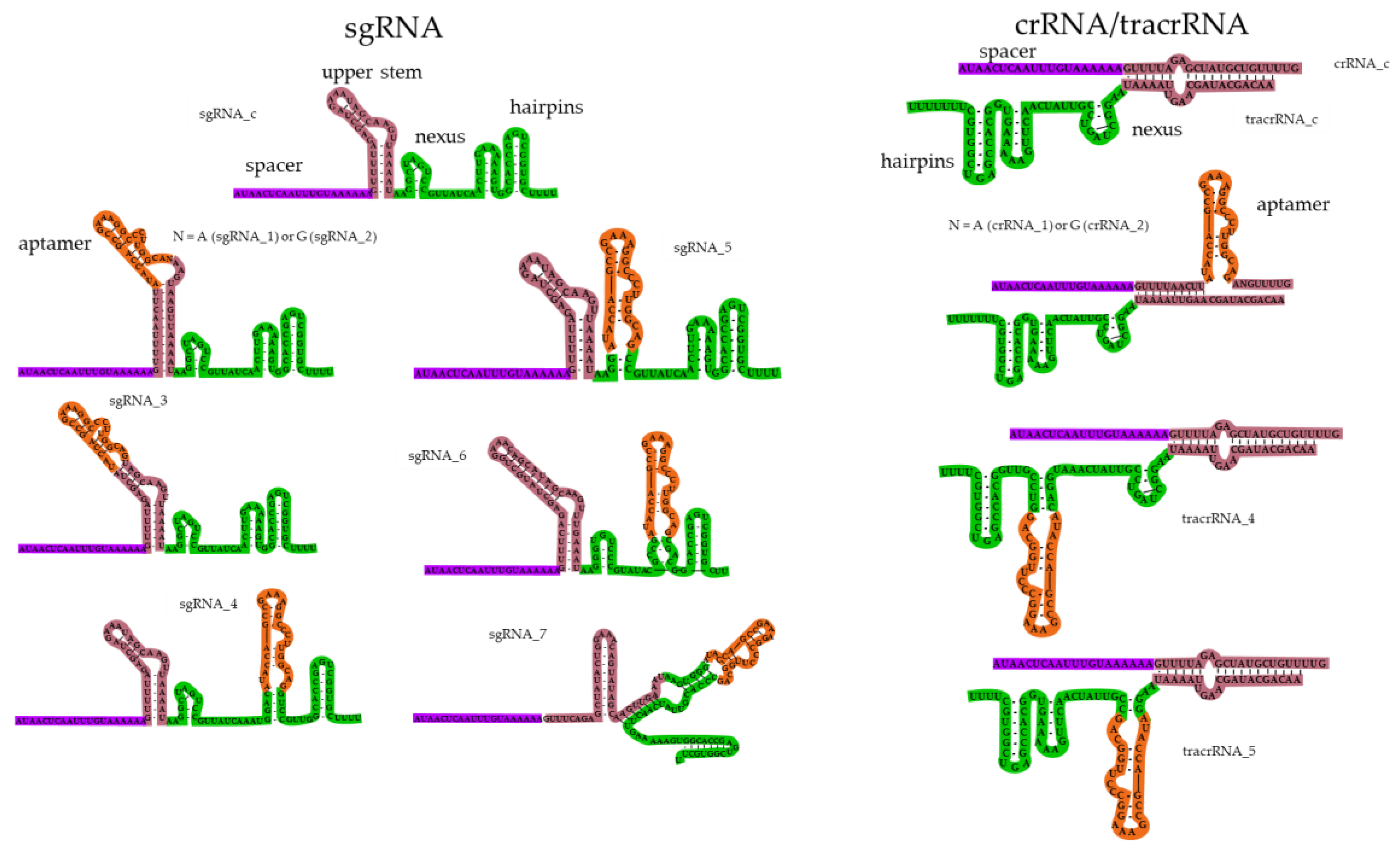
Figure 3.
Structure of designed guide RNAs contained theophylline aptamer within 3'- or 5'-terminal fragments. The purple color indicates spacer sequence, grey rose indicates upper stem or contact between crRNA and tracrRNA, green color indicated nexus and hairpins, orange color indicates theophylline aptamer, blue color indicates blocking motif.
Figure 3.
Structure of designed guide RNAs contained theophylline aptamer within 3'- or 5'-terminal fragments. The purple color indicates spacer sequence, grey rose indicates upper stem or contact between crRNA and tracrRNA, green color indicated nexus and hairpins, orange color indicates theophylline aptamer, blue color indicates blocking motif.
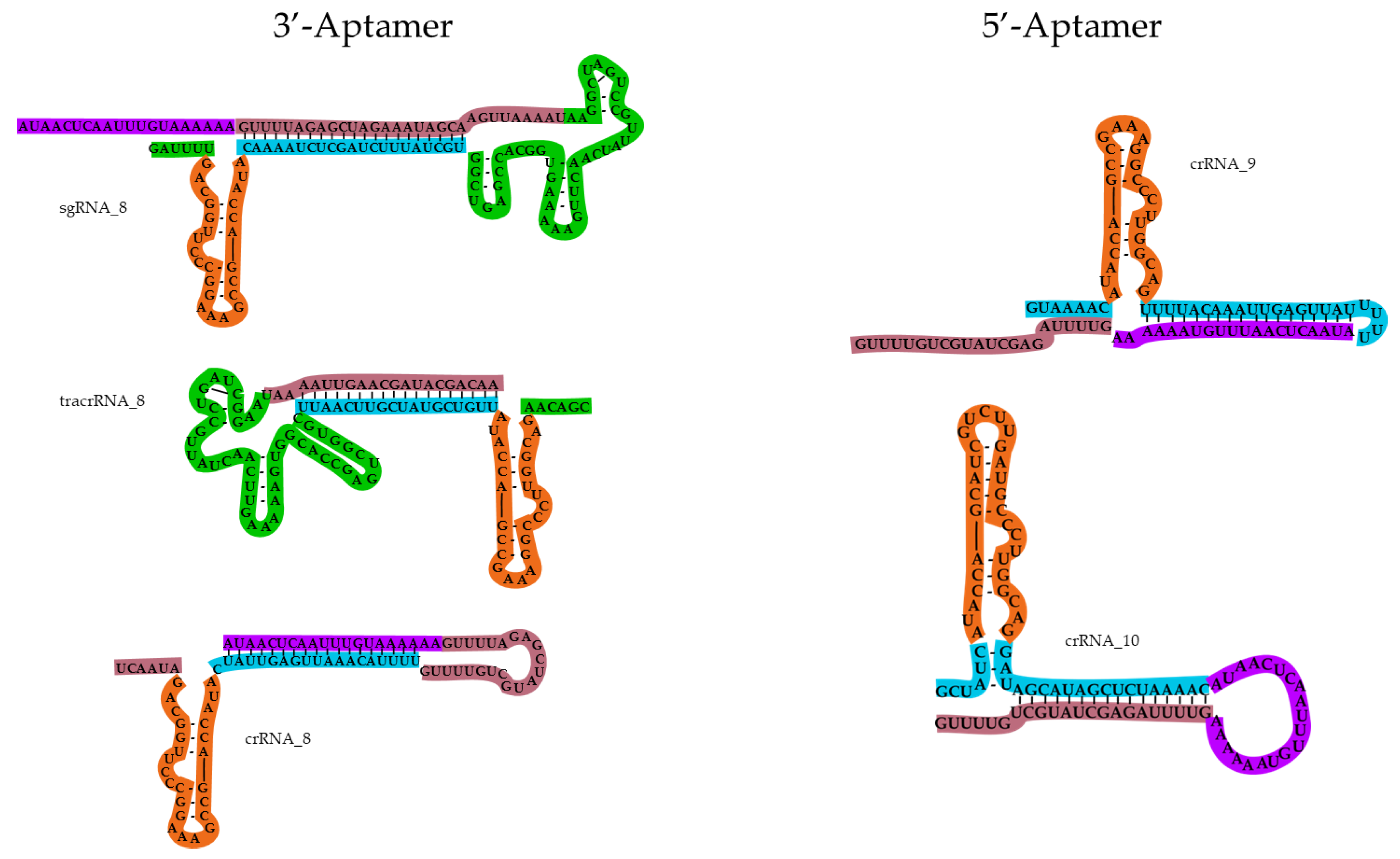
Figure 4.
The extent of plasmid DNA cleavage by the Cas9 nuclease in the presence of guide RNAs containing the theophylline aptamer in the absence of theophylline. (A) Guide RNA - sgRNA containing the aptamer. (B) Guide RNA – complex of crRNA+tracrRNA, were crRNA or tracrRNA containing the aptamer. The sequences of guide RNA are presented on Figure 2 and Figure 3 and in the Table 1 and Table 2. Reaction conditions: sgRNA(crRNA/tracrRNA):Cas9:plasmid = 20:20:20:1 in a buffer containing 20 mM HEPES-KOH (pH 7.5), 100 mM KCl, 0.5 mM Na2EDTA, 2 mM MgCl2, 1 mM DTT for 1 hour at 37 °С.
Figure 4.
The extent of plasmid DNA cleavage by the Cas9 nuclease in the presence of guide RNAs containing the theophylline aptamer in the absence of theophylline. (A) Guide RNA - sgRNA containing the aptamer. (B) Guide RNA – complex of crRNA+tracrRNA, were crRNA or tracrRNA containing the aptamer. The sequences of guide RNA are presented on Figure 2 and Figure 3 and in the Table 1 and Table 2. Reaction conditions: sgRNA(crRNA/tracrRNA):Cas9:plasmid = 20:20:20:1 in a buffer containing 20 mM HEPES-KOH (pH 7.5), 100 mM KCl, 0.5 mM Na2EDTA, 2 mM MgCl2, 1 mM DTT for 1 hour at 37 °С.
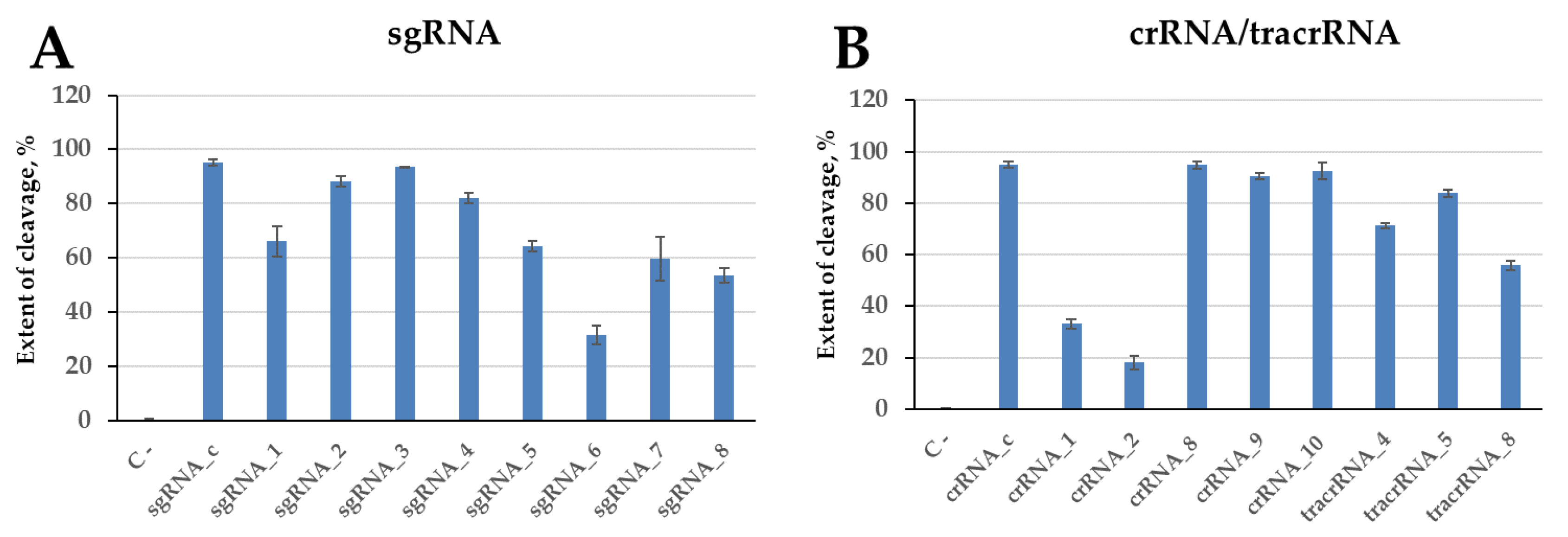
Figure 5.
Influence of theophylline to the DNA cleavage by the Cas9 nuclease guided by RNAs containing the theophylline aptamer. (A) Guide RNA - sgRNA containing the aptamer. (B) Guide RNA – complex of crRNA+tracrRNA, were crRNA or tracrRNA containing the aptamer. The sequences of guide RNA are presented on Figure 2 and Figure 3 and in the Table 1 and Table 2. Reaction conditions: sgRNA(crRNA/tracrRNA):Cas9:plasmid = 20:20:20:1 in a buffer containing 20 mM HEPES-KOH (pH 7.5), 100 mM KCl, 0.5 mM Na2EDTA, 2 mM MgCl2, 1 mM DTT for 1 hour at 37 °С. Theophylline concentration was 1 mM. Mean ± SD is shown (n = 3); *, p < 0.05; **, p < 0.01; ***, p < 0.001 (one-way ANOVA) in comparison with DNA cleavage in the absence of theophylline.
Figure 5.
Influence of theophylline to the DNA cleavage by the Cas9 nuclease guided by RNAs containing the theophylline aptamer. (A) Guide RNA - sgRNA containing the aptamer. (B) Guide RNA – complex of crRNA+tracrRNA, were crRNA or tracrRNA containing the aptamer. The sequences of guide RNA are presented on Figure 2 and Figure 3 and in the Table 1 and Table 2. Reaction conditions: sgRNA(crRNA/tracrRNA):Cas9:plasmid = 20:20:20:1 in a buffer containing 20 mM HEPES-KOH (pH 7.5), 100 mM KCl, 0.5 mM Na2EDTA, 2 mM MgCl2, 1 mM DTT for 1 hour at 37 °С. Theophylline concentration was 1 mM. Mean ± SD is shown (n = 3); *, p < 0.05; **, p < 0.01; ***, p < 0.001 (one-way ANOVA) in comparison with DNA cleavage in the absence of theophylline.
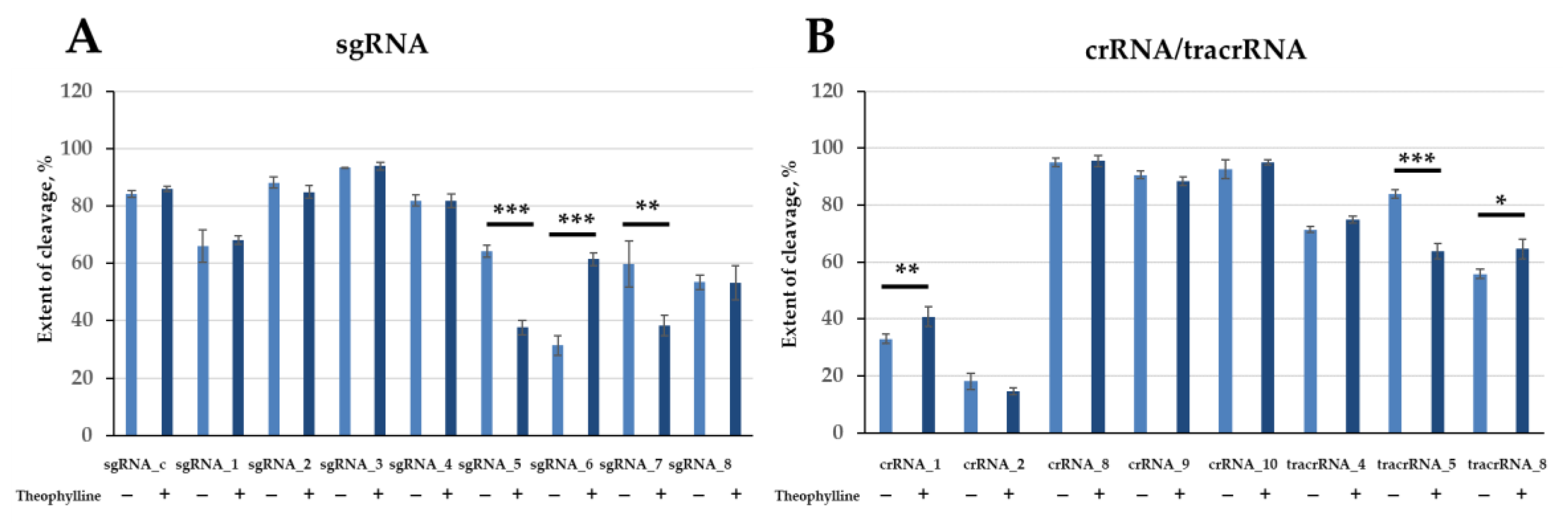

Table 1.
Sequences of guide RNA containing the theophylline aptamer in functional motifs, and planned mode of regulation.
Table 1.
Sequences of guide RNA containing the theophylline aptamer in functional motifs, and planned mode of regulation.
| Name | Sequence, 5'-3' | Length, nt | Activation/ Deactivation |
|---|---|---|---|
| sgRNA | |||
| sgRNA_c | AUAACUCAAUUUGUAAAAAAGUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCA-CCGAGUCGGUGCUUUU | 100 | none |
| sgRNA_1 | AUAACUCAAUUUGUAAAAAAGUUUUAACUUAUACCAGCC-GAAAGGCCCUUGGCAGAAGUAAGUUAAAAUAAGGCUAGU-CCGUUAUCAACUUGAAAAAGUGGCACCGAGUC-GGUGCUUUU | 119 | activation |
| sgRNA_2 | AUAACUCAAUUUGUAAAAAAGUUUUAACUUAUACCAGCCG-AAAGGCCCUUGGCAGAGGUAAGUUAAAAUAAGGCUAGUC-CGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGCUUUU | 119 | activation |
| sgРНК_3 | AUAACUCAAUUUGUAAAAAAGUUUUAGAGCUAAUACCAGCC-GAAAGGCCCUUGGCAGUAGCAAGUUAAAAUAAGGCUAGUCC-GUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGCUUUU | 121 | activation |
| sgRNA_4 | AUAACUCAAUUUGUAAAAAAGUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAAAUGGACAUACCAGCC-GAAAGGCCCUUGGCAGGUCCGUUGGCACCGAGUCGGUGCUUUU | 127 | activation |
| sgRNA_5 | AUAACUCAAUUUGUAAAAAAGUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGAUACCAGCCGAAAGGCCCUUGGCAGCCGUU-AUCAACUUGAAAAAGUGGCACCGAGUCGGUGCUUUU | 120 | deactivation |
| sgRNA_6 | AUAACUCAAUUUGUAAAAAAGUUUCAGAGCUAUGCUGGAAACA-GCAUAGCAAGUUGAAATAAGGGUGUCCCGUAUACGCCGAUACC-AGCCGAAAGGCCCUUGGCAGCGACGGC-ACCGAGUCGGUGCUU | 128 | activation |
| sgRNA_7 | AUAACUCAAUUUGUAAAAAAGUUUCAGAGCUAUGCUGGAAAC-AGCAUAGCAAGUUGAAATAAGUGGGAUACCAGCCGAAAGGCC-CUUGGCAGCCUACGUUAUCAACUUGAAAAAGUGGCACCGAGU-CGGUGCUU | 134 | deactivation |
| crRNA | |||
| crRNA_c | AUAACUCAAUUUGUAAAAAAGUUUUAGAGCUAUGCUGUUUUG | 42 | none |
| crRNA_1 | AUAACUCAAUUUGUAAAAAAGUUUUAACUUAUACCAGCCGA-AAGGCCCUUGGCAGAAGUUUUG | 63 | activation |
| crRNA_2 | AUAACUCAAUUUGUAAAAAAGUUUUAACUUAUACCAGCCGA-AAGGCCCUUGGCAGAGGUUUUG | 63 | activation |
| tracrRNA | |||
| tracrRNA_c | AACAGCAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACC-GAGUCGGUGCUUUUUUU | 74 | none |
| tracrRNA_4 | AACAGCAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAAAUGGACAUACCAGCCGAAAGGCCCUUGGCAGGUCCGUUGGCACCGA-GUCGGUGCUUUU | 98 | activation |
| tracrRNA_5 | AACAGCAUAGCAAGUUAAAAUAAGGAUACCAGCCGAAAGGCC-CUUGGCAGCCGUUAUCAACUUGAAAAAGUGGCACCGAGUC-GGUGCUUUU | 91 | deactivation |
* The sequence targeted to the DNA is underlined; the aptamer sequence is bold italic style.
Table 2.
Sequences of guide RNA containing the theophylline aptamer 3'- or 5'-terminus and planned mode of regulation.
Table 2.
Sequences of guide RNA containing the theophylline aptamer 3'- or 5'-terminus and planned mode of regulation.
| Name | Sequence, 5'-3' | Length, nt | Activation/ Deactivation |
|---|---|---|---|
| sgRNA | |||
| sgRNA_8 | AUAACUCAAUUUGUAAAAAAGUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAA GGCUAGUCCGUUAUCAACUUGAAAAAGUGGCA-CCGAGUCGGUGCUAUUUCUAGCUCUAAAACAUACCAGCCGAAA-GGCCCUUGGCAGUUUUAG | 145 | activation |
| crRNA | |||
| crRNA_8 | AUAACUCAAUUUGUAAAAAAGUUUUAGAGCUAUGCUGUUUUG-UUUUACAAAUUGAGUUAUCAUACCAGCCGAAAGGCCCUUG-GCAGAUAACU | 92 | activation |
| crRNA_9 | GUAAAACAUACAGCCGAAAGGCCCUUGGCAGUUUUACAAAUUGAGUUAUUUUUAUAACUCAAUUUGUAAAAAAGUUUUAGAGCUAUGCUGUUUUG | 96 | activation |
| crRNA_10 | GCUAUCAUACCAGCAUCGUCUUGAUGCCCUUGGCAGGAUAGCAUAGCUCUAAAACAUAACUCAAUUUGUAAAAAAGUUUUAGA-GCUAUGCUGUUUUG | 97 | activation |
| tracrRNA | |||
| tracrRNA_8 | AACAGCAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGG CACCGAGUCGGUGCUUAACUUGCUAUGCU-GUUAUACCAGCCGAAAGGCCCUUGGCAGAACAGC | 116 | activation |
* The sequence targeted to the DNA is underlined; the aptamer sequence is bold italic style.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



