
Submitted:
28 November 2024
Posted:
29 November 2024
You are already at the latest version
Abstract
Modern agriculture must reverse its degrading influence on soils, the natural environment, and the climate. No-till soil cultivation technologies, which have been used for many years and are constantly being improved, are an example of such activities. Our research assumes that soil samples taken from a faba bean plantation in the twentieth year of the static field experiment will show differences in the composition of bacterial communities at the level of phylum, orders, classes, and species. The differences would be manifested by the environmental and agricultural functionality of the microorganisms in the soil cultivation system-dependent manner and they would indicate the bacterial species with environment-friendly properties. As a result, five variants of the soil samples were distinguished: 1 – plantless, fallow buffer zone of the field experiment – sample ‘0’, 2 – conventional tillage (ploughing), 3 – reduced tillage (stubble cultivator), 4 – strip tillage,5 – no tillage (direct drilling into stubble). Our study showed that the no-till cultivation technologies, mainly the strip till and no-till methods, applied at the horse bean plantation had a positive influence on microbial communities. The number of OTUs of species such as Gemmatimonas aurantiaca and Aeromicrobium ponti increased. The abundance of these species determines soil fertility and yield of crops. They are also environment and climate friendly, and therefore very important for sustainable or even regenerative agriculture. Further investigations of soil samples collected from another plantation of crops in different tillage systems may indicate functional species of microorganisms which are important for climate change and for the fertility of the soil.
Keywords:
biodiversity
; the abundance of microorganisms
; Gemmatimonas aurantiaca
; Aeromicrobium ponti
1. Introduction
Modern agriculture faces two major challenges. On the one hand, it must feed the growing world population. On the other hand, it must reverse its current degrading influence on soils, the natural environment, and the climate [1,2]. It is possible to achieve these goals simultaneously by following one of many parallel paths, i.e. the development of conservation and regenerative agriculture, and especially the rational implementation of the widest possible spectrum of its principles into practices of intensive commercial agriculture. No-till soil cultivation technologies, which have been used for many years and are constantly being improved, are an example of such activities. They limit soil degradation and help to preserve the production, biological, environmental, and climatic potential of soils [3,4].
No-till technologies involve the use of simplified soil cultivation methods or even complete abandonment of tillage. As a result, it is possible to reduce input costs, including fuel consumption [5]. No-till systems have a positive influence on the physical, chemical, and biological properties of soils because they limit their aeration and drying. In consequence, the rate of mineralisation of organic matter and CO2 emissions are reduced. Leaving some or all of the crop residues on the surface or in the subsurface layer of soil increases its protection from drying and erosion. Apart from that, it improves the transformation of soil organic matter and carbon sequestration [6,7].
However, these technologies sometimes also have negative effects such as excessive soil compaction, big differences in the properties of individual layers of the soil humus horizon – significantly higher C and N content and higher acidification due to nitrogen fertilisation of the topsoil. These phenomena may negatively influence the yield of some crops [5]. Moreover, the absence of tillage usually favors the expansion of some weeds, which may cause the need to use larger amounts of herbicides [8,9].
The specific physical and chemical properties of soils cultivated in different systems also result in their characteristic biological properties, i.e. mainly the size and diversity of the edaphon, especially the population of microorganisms and the enzyme activity [10]. Individual cultivation systems significantly influence soil aeration and moisture as well as the amount and distribution of the biomass of crop residue [11]. Soil cultivation simplifications, especially the complete abandonment of tillage, usually favour greater soil bioactivity and positively affect the size and diversity of the microbial population. This applies to the total count of bacteria and fungi as well as individual taxonomic and trophic groups. At the same time, the populations of some microorganisms, e.g. bacteria of the Azotobacter genus, are larger in traditionally ploughed soils [12].
The current knowledge of the influence of the cultivation system on the soil microbiome is still very general. There have been few studies covering a wide spectrum of individual taxa of soil microorganisms and their communities [13]. Detailed studies in this field became possible only with the development and greater availability of molecular and genetic research techniques.
The assumption of our study was that the soil samples collected from a horse bean plantation in the twentieth year of a static field experiment would exhibit differences in the composition of bacterial communities at the level of phylum, order, classes, and species. The differences would be manifested by the environmental and agricultural functionality of the microorganisms in the soil cultivation system-dependent manner and they would indicate the bacterial species with environment-friendly properties.
The aim of the study was to determine the structure of bacterial communities and to assess the trend of changes in the microbiome in the soil under horse beans grown in a four-field cereal rotation system (75% cereals): horse beans, winter wheat, spring barley, winter triticale, in four different cultivation technologies: traditional (ploughing), simplified (stubble cultivator), strip till, and direct drilling into stubble.
2. Materials and Methods
2.1. Field Experiment
The research was based on soil samples collected from a horse bean plantation in the twentieth year of a static field experiment, in which four levels of the experimental factor (tillage system) were used: conventional tillage (ploughing), reduced tillage (stubble cultivator), strip tillage (strip-till seed drill), no-tillage (no-till seed drill). Horse beans was grown in a four-year crop rotation cycle: horse beans, winter wheat, spring barley, and winter triticale.
In 1999, a field experiment (randomised complete block design, four replications) was started at the Brody Agricultural Experimental Station (52°26’N; 16°17’E) belonging to the Poznań University of Life Sciences, Poland. The soil was categorised as Albic Luvisols (12% clay, 19% silt, 69% sand) and, according to the Soil Taxonomy, it was categorised as Typic Hapludalfs, due to the granulation in loamy sand underlined by loam [14].
In the autumn, a mineral fertiliser was applied at a dose of 60 kg of phosphorus and 90 kg of potassium per 1 ha. In the spring, 40 kg N·ha–1 was applied. In early April, horse beans of the Albus cultivar was sown at a density of 60 seeds·m–2 (approx. 390 kg·ha–1). Weeds were controlled with Wing P 462.5 EC (after sowing) and Corum 502.4 SL (after the emergence of plants). During the pod formation period, the Bulldock 025 EC pesticide was applied when necessary. One week before harvesting horse bean seeds, the Dessicash 20 SL herbicide was applied for desiccation. The crops were harvested with a field combine harvester in early August.
2.2. Weather Conditions
The weather conditions (rainfall and air temperatures) during the growing season (Table 1) are shown as the Selyaninov Hydrothermal Coefficient (HTC) according to the following formula [15,16]:
where: – ten-day/monthly rainfall (periods with an average daily temperature above 8°C), – ten-day/monthly average daily air temperature (periods with an average daily temperature above 8°C).
Despite its limitations, this simple-to-calculate coefficient is useful for monitoring agricultural conditions during the growing season and drought. The HTC clearly indicates periods of excessive or optimal humidity and drought. It is flexible enough for use in both monthly and ten-day monitoring of weather conditions [17].
Drought prevailed during the growing season. June was the driest month, with the extreme HTC values in all ten-day periods. There was no drought, with near-optimal conditions, only in May and September. In mid-May humidity was too high, as evidenced by the HTC value.
2.3. Soil Analyses
The composition of the soil microbiome under the horse bean plantation was analysed in 2019, i.e. in the twentieth year of the field experiment. Soil samples were collected from the topsoil (0-18 cm) once, immediately after the harvesting of horse beans. An Egner probe was used for sampling. Forty detailed samples were collected from random points of all plots representing individual experimental combinations (10 samples from each plot). They were merged into collective soil samples – each weighing about 1 kg. The same procedure was applied to collect detailed samples from the plantless, fallow buffer zone surrounding the entire site of the experiment (sample ‘0’). A collective sample was created. As a result, five variants of the soil samples were distinguished (one variant for sample ‘0’ and four variants representing tillage systems). They were used in microbiological analyses. They were marked in the following way:
- plantless, fallow buffer zone of the field experiment – sample ‘0’,
- conventional tillage,
- reduced tillage,
- strip tillage,
- no tillage.
2.4. Identification of Soil Microorganisms. DNA Extraction
Total DNA was extracted from 500 mg of each sample with a Genomic Mini AX Soil kit (A&A Biotechnology) according to the manufacturer’s instructions. The extracted DNA was quantified with a Quant-iT HS dsDNA assay kit (Invitrogen) on a Qubit2 fluorometer; 2 μl of the extracts was examined on 0.8% agarose gel.
2.5. PCR Amplification
The metagenomic analysis was based on the hypervariable region V3–V4 of the 16S rRNA gene. Specific primers 341F and 785R were used for amplification of this region and to prepare libraries. A PCR was conducted with a Q5 Hot Start High-Fidelity DNA Polymerase kit (NEB Inc., Ipswich, MA, United States). The reaction conditions were maintained according to the manufacturer’s specifications. An Illumina MiSeq PE300 sequencer (Genomed S.A., Warsaw, Poland) in 2 × 250 bp paired-end (PE) technology with a v2 Illumina chemistry kit was used for sequencing. The reactions were conducted according to the Illumina V3–V4 16S RNA amplification protocol (Illumina, San Diego, CA, United States). The data were analysed automatically with the MiSeq and in the Illumina BaseSpace cloud environment according to the 16S Metagenomics protocol (ver. 1.0.1). The libraries were prepared in an analogous way to the attached Illumina protocol.
2.6. Statistical and Bioinformatics Analyses
A comparative analysis of genus composition between experimental variants was visualized using several types of plots [18]. Differences in the mean abundance of bacteria between the variants were calculated and visualized. All analyses were conducted using the statistical package Genstat version 23.1 [19].
3. Results and Discussion
The metapopulation analysis based on the analysis of 16S rRNA sequences showed that different cultivation technologies influenced the number of operational taxonomic units (OTUs) of the bacteria found in the soil under the horse beans (variants 2-5), as compared with the plantless control soil (sample ‘0’) collected from the fallow buffer zone of the experiment (variant 1). Next-generation sequencing is an increasingly popular and extremely sensitive method of determining similarities and differences within the soil microbiome. This fact is evidenced by the results of our study and data provided in reference publications [20,21]. Depending on the soil site, there were 32 phyla, 63 classes, 133 orders, and 1900 species of bacteria. Due to such a large number of operational taxonomic units (OTUs) only those whose average abundance was above 1% were presented.
The results showed that in each of the soil samples collected from individual experimental variants of cultivation (1-5) at the horse bean plantation, the abundance of the Bacteria domain was over 99.96-99.98% of the microbiome, whereas the abundance of the Archaea domain amounted to 0.02-0.03% (Table 2).
3.1. Bacterial Phyla
The 16S rRNA metagenomic analysis of the soil metabiome revealed the dominance of three phyla of bacteria: Actinobacteria, Firmicutes, and Proteobacteria, which were found in all experimental variants. There were similar findings presented in reference publications on arable soils.
The metagenomic analyses showed that among all bacterial phyla identified in sample ‘0’ and in the soil samples collected from the horse bean plantation, Actinobacteria were predominant. The share of operational taxonomic units (OTUs) was as follows: sample ‘0’ collected before the experiment – 41.19% (variant 1), traditional tillage (variant 2) – 39.57%, reduced tillage (variant 3) – 43.10%, strip tillage (variant 4) – 50.72%, direct drilling (variant 5) – 40.04% (Figure 1). Actinobacteria grew and developed particularly abundantly in the soil where the strip tillage system was applied (variant 4) (Figure 1).
Apart from the assessment and identification of the dominant phyla in the soil samples, differences in the number of operational taxonomic units (OTU) between the soil samples collected from the horse beans grown in different cultivation systems and the control soil were analysed: 2 vs. 1, 3 vs. 1, 4 vs. 1, 5 vs. 1. They are shown in Figure 2.
All four comparisons showed that in the experimental variants, the Acidobacteria and Gemmatimonadetes phyla were more abundant than in sample ‘0’ collected from the fallow buffer zone of the experiment. An inverse relationship was observed for the Verrucomicrobia phylum. The number of OTUs in sample ‘0’ was always greater than in the soils cultivated in different systems. The largest positive difference in the number of OTUs (9.54%) in relation to sample ‘0’ was observed for the Actinobacteria phylum in the strip-till variant. In the same cultivation system, the largest negative difference (-7.39%) was observed for the Proteobacteria phylum (Figure 2).
As observed by Swędrzyńska and Małecka-Jankowiak [11], Actinobacteria are a saprophytic group of microorganisms which adapt and proliferate intensively in an environment with high availability of organic matter. This effect could be seen in the strip tillage system, as well as in the soil under a horse bean plantation. Bao et al. [22] also observed the dominance of Actinobacteria, especially in less fertile soils with a supply of additional organic matter (rice straw). The researchers indicated the important ecophysiological role of these microorganisms. Actinobacteria communities broadly adapt to the decomposition of plant residues. They are of potential significance for the decomposition of these residues and carbon sequestration in the soil. This phylum mineralises organic material. Similarly to oligotrophic microorganisms, it spares carbon in the soil, which additionally proves its ecological role in the pedon [23,24]. It is important to note that Actinobacteria can quickly adapt to unfavourable environmental conditions such as drought by producing conidia. This is why they are capable of decomposing organic matter even at low soil moisture, Our study confirmed this fact, as evidenced by the weather conditions during the experiment (Table 1).
Apart from Actinobacteria, Proteobacteria was another dominant phylum in our study. Its relative abundance ranged from 24.60% in the strip-till variant to 35.67% in the reduced tillage system (Figure 1). The study by Mahnkopp-Dirks et al. [25] also showed that Proteobacteria was the dominant phylum in agricultural soils. Fierer et al. [24] proposed a concept of bacterial classification in which Proteobacteria phyla were described as fast-growing copiotrophs, i.e. microorganisms developing in environments with a high supply of carbon. Their abundance was closely correlated with the degree of carbon mineralisation in the soil.
Our study also showed that Firmicutes, Planctomycetes, Chloroflexi, Acidobacteria, Bacteroidetes, and Verucomicrobia were dominant in all soil variants (1-5). Gemmatimonadates was also a dominant phylum but only in the soil under horse beans, in various cultivation systems. Its number of OTUs was the greatest in the direct drilling system – 3.29% (Figure 1).
The Firmicutes, Planctomycetes, Chloroflexi, Acidobacteria, Bacteroidetes, and Verrucomicrobia phyla were isolated less frequently (Figure 1).
Chloroflexi is a phylum of anaerobic or microaerophilic nitrifying bacteria which can survive in extreme and intensively changing conditions. Chloroflexi occurred more frequently in variant 1 in sample ‘0’ (fallow). This can be explained by the fact that these bacterial communities develop by using cellular compounds from dead microorganisms and their metabolites, which is characteristic of uncultivated (fallow) or exhausted soils. Tang et al. [26] observed a similar dominance of this group of microorganisms. The researchers used rice straw and biochar to effectively improve the soil quality, and thus reduce the abundance of this bacterial phylum.
The Firmicutes phylum includes bacteria of the Bacillus and Clostridium genera, which produce spores in unfavorable environmental conditions. The phylum was dominant in variant 1, whereas in the variants with horse beans grown in different cultivation systems, the number of its OTUs was on average 1-2% lower (Figure 1).
Bacteria of the Verrucomicrobia phylum are cosmopolitans in the rhizosphere. Their adaptations and functions are enigmatic because so far a low percentage of members of the phylum have been cultured. However, research has shown that Verrucomicrobia plays a key role in environmental carbon cycling and (poly)saccharide degradation because they have a high capacity to encode glycoside hydrolase genes [27].
The occurrence of the Gemmatimonadates phylum in the soil samples of the four cultivation systems collected from the horse bean plantation was higher than in the fallow soil sample ‘0’ (Figure 1). According to the data in reference publications, Gemmatimonadota are the eighth most abundant phylum of soil bacteria, with a share of about 1-2% worldwide [28]. DeBruyn et al. [29] proved that although Gemmatimonadota can be found in various environments, the maximum number of their sequences comes from soils used in different ways, including grassland soils, agricultural soils, forest soils, and contaminated soils. The cosmopolitan distribution of Gemmatimonadota in different soils suggests that they are generalist species with versatile metabolism and they can adapt to a wide range of nutrients. Mujakić et al. [30] and Liu et al. [31] found that Gemmatimonadota are a bacterial phylum whose abundance in soil increases with the presence of vegetation in the field. The relative number of OTUs within the phylum increases strongly then and exceeds 2%. Moreover, the bacteria are positively correlated with plant species richness and soil nutrients, mainly carbon. Gemmatimonadota are one of the seven more dominant bacterial phyla, with abundance above 1%. The bacteria are positively correlated with total carbon, nitrogen, and phosphorus in the soil. Studies by Deng et al. [32] and Ye et al. [33] showed that the high concentration of nutrients stimulated the abundance of Gemmatimonadota and suggested that the bacteria could play a key role in soil ecosystems. In our study, there was an increased relative number of OTUs of Gemmatimonadota due to the applied cultivation systems and the crop, i.e. horse bean (Figure 2). These results were in line with the properties of Gemmatimonadota described in the reference publications. Legumes have been widely studied due to their significant ecological benefits. They enrich the soil with organic matter by leaving a large mass of crop residues (straw and roots), i.e. 3-6 t ha–1. They are capable of biological nitrogen fixation by establishing symbiosis with bacteria from the Rhizobiaceae family. This is particularly important on light soils because it increases the sorption complex [34] and creates a better environment for the biodiversity of the soil microbiome.
The analysis of differences (Figure 2) showed that the number of OTUs of Acidobacteria in the soils cultivated in different systems was greater than in sample ‘0’. According to Lee et al. [35], the occurrence of the Acidobacteria phylum is correlated with the physicochemical properties of soils. It is characteristic of soils on which rice is grown as well as orchard soils, with limited oxygen access due to soil compaction and generally deteriorating conditions for bacteria. Li et al. [36] conducted a fifteen-year experiment, in which the no-till cultivation system was mostly used. The researchers observed an increased abundance of the Acidobacteria phylum and concluded that conservation tillage increased the biodiversity of the microbiome. Our observations of changes in the abundance of bacterial communities belonging to specific phyla in the soil samples collected from different cultivation systems under the horse bean plantation were in line with the findings of the aforementioned studies.
3.2. Dominant Classes and Orders of Soil Bacteria Under Horse Beans Cultivated in Different Systems
In further analyses, the abundance of the dominant classes and orders within the microbiome was assessed. The metagenomic analysis showed that the type of soil cultivation system used at the horse bean plantation affected the content of bacterial OTUs within classes and orders. Also here, only the orders and classes that constituted more than 1% of the microbiome were shown in the graphs. Apart from the microorganisms which were not classified into classes and whose abundance of OTUs was 6.09-7.05%, depending on the cultivation system used, there were dominant shares of Actinobacteria and Alphaproteobacteria (Figure 3).
Khmelevtsova et al. [37] observed similar dependencies related to the use of different cultivation technologies. The researchers found a large number of OTUs in the classes of Actinobacteria and Alphaproteobacteria in different soil cultivation technologies. They explained that the increased growth of Actinobacteria was caused by their greater sensitivity to physical disturbances due to their hyphal structure. Deep ploughing disrupted their mycelium, which broke the hyphae of Actinobacteria. Although the diameter of S. coelicolor cells is smaller than 1 μm, it may reach 100 μm due to the elongation of hyphae at the ends by the membrane vesicle transporter [38].
Our study showed that apart from Actinobacteria, Alphaproteobacteria was another class that was frequently isolated in the soils under horse beans grown in different cultivation systems. The abundance of this class amounted to 18.34% in sample ‘0’, 18.98% in the traditional cultivation variant (ploughing), 20.41% in the reduced tillage variant (stubble cultivator), 13.96% in the strip till variant, and 20.43% in the variant with direct drilling into stubble (Figure 3). Alphaproteobacteria are classified as oligotrophs as they use organic resources very sparingly. Apart from oligotrophic bacteria, this class also includes bacteria of agricultural significance, capable of fixing nitrogen in symbiosis with plants [39,40]. Batut et al. [41] observed that many Alphaproteobacteria interacted with higher eukaryotes. These interactions range from pericellular colonisation, through facultative intracellular multiplication, to enforcing an intracellular lifestyle. This wide range of interactions has a common feature – the modulation of proliferation of host cells. This sometimes leads to the formation of structures known as nodules in which bacteria can grow.
Apart from the assessment and indication of the dominant classes in the soil samples, relative differences in the number of OTUs between a specific soil variant collected from under horse beans grown in different cultivation systems (traditional tillage, reduced tillage, strip tillage, direct drilling) and the control soil sample were also analysed (2 vs. 1, 3 vs. 1, 4 vs. 1, 5 vs. 1). The results are shown in Figure 4. In all four comparisons, the abundance of Actinobacteria and Gemmatimonadetes classes in the soils cultivated in different systems was greater than in sample ‘0’ – plantless, fallow buffer zone of the field experiment. There was an inverse relationship observed for the Acidimicrobiia, Brocadiae, and Clostridia classes. The relative number of OTUs belonging to these classes in sample ‘0’ was always greater than in the other experimental variants. The comparison with sample ‘0’ (plantless, fallow buffer zone of the field experiment) revealed the highest relative difference (10.63%) for Actinobacteria in the strip-till system (4 vs. 1). The greatest decrease in OTUs was observed for Alphaproteobacteria in the same cultivation system (Figure 4).
The share of OTUs of unclassified bacterial orders ranged from 7.18% to 9.49%, depending on the cultivation system. The dominant orders were: Actinomycetales , followed by Bacillales and Rhizobiales (Figure 5).
According to the reference publications, there are 16 orders in the Actinobacteria class. These are: Actinomycetales, Actinopolysporales, Bifidobacteriales, Catenulisporales, Corynebacteriales, Frankiales, Glycomycetales, Jiangellales, Kineosporiales, Micrococcales, Micromonosporales, Propionibacteriales, Pseudonocardiales, Streptomycetales, Streptosporangiales, and Incertae sedis [42].
In our study, Actinomycetales was the only order whose abundance exceeded 1%. The highest relative share of OTUs of this order, i.e. 44.21%, was found in variant 4 (strip-till cultivation), whereas the lowest share, i.e. 33.62% was found in sample ‘0’ (plantless, fallow buffer zone of the field experiment) (Figure 5). The results of our study are in line with the findings of the research conducted by Bao et al. [22], who observed the dominance of this group of bacteria, especially in less fertile soils with the inflow of additional organic matter. According to Mokni-Tlili et al. [43], the dominance (abundance and distribution) of Actinomycetales, which play an important role in the decomposition of organic matter, may indicate good soil fertility.
Apart from the Actinomycetales order, there was also a large number of OTUs of the Bacillales order, which belongs to the Firmicutes phylum. The dominance of this group of bacteria depends on the source of carbon rather than the pH of the soil environment due to their ability to produce spores. In our study, the largest numbers of OTUs of this order were found in variants 3, 4, and 5, in which reduced tillage technologies (reduced tillage, strip tillage, and direct drilling) were used (Figure 5). On average, the shares of this order in these variants were 1.5%-2% greater than in the conventional tillage (ploughing) variant and in soil sample ‘0’ collected from the plantless fallow buffer zone of the experiment. According to the data provided in the reference publications, the presence of these bacteria in soil ecosystems is crucial for the decomposition of organic matter, as they decompose dead plant and animal material and thus release nutrients back into the soil. The decomposition process not only enriches the soil, but also facilitates the formation of humus, a stable organic component that improves soil structure, water retention, and aeration. The activity of Bacillales bacteria in the nutrient cycle helps to maintain plant growth by creating a dynamic balance in the soil ecosystem. Many species within the Bacillales order interact with other soil organisms, which indicates the ecological importance of these bacteria. Bacillales often colonise the rhizosphere and thus establish symbiosis with plants. They facilitate the uptake of nutrients in this microenvironment by converting them into forms available to plants. In consequence, the health and immunity of plants improves. This interrelation may increase agricultural productivity, thus indicating the integral role of Bacillales in supporting sustainable agriculture [44].
Unlike Bacilalles, there was an inverse relationship observed for the Rhizobiales order. Its abundance was lower in the variants with reduced tillage (Figure 5). This order of bacteria includes both plant and animal pathogens, such as the Brucella and Bartonella genera, the Agrobacterium genus, as well as nitrogen-fixing bacteria, including the Rhizobium, Sinorhizobium, and Bradyrhizobium genera [45]. The decrease in the number of OTUs of bacteria belonging to the Rhizobiales order can be explained by the fact that the proliferation and activity of symbiotic diazotrophic bacteria are inhibited by the inflow of mineralised organic matter, especially available forms of nitrogen [46]. The results of our study were in line with the aforementioned assumptions. The abundance of the Rhizobiales order was most reduced in variant 4 (strip-till cultivation) (Figure 5).
3.3. Dominant Bacterial Species in Different Soil Cultivation Technologies Under Horse Bean Plantation
Our study also included an assessment of the dominance of specific bacterial species in the soil samples collected from individual cultivation systems (conventional tillage, reduced tillage, strip tillage, no tillage) in relation to the uncultivated soil (NC – no cultivation). The abundance of unclassified bacterial species ranged from 58.44% to 61.83% (Figure 6). The dominant species were: Arthrobacter psychrochitiniphilus, Cohnella soli, and Nocardioides islandensis. In order to interpret the results more precisely, our analyses also included finding relative differences in the abundance of the dominant species between the samples of soil collected from different cultivation systems under the horse bean plantation and the zero sample collected from the plantless, fallow buffer zone (Figure 6). The comparisons of all four variants with sample 0 revealed an increase in the OTUs of the following species: Aeromicrobium ponti, Arthrobacter psychrochitiniphilus, Cohnella soli, and Gemmatimonas aurantiaca. By contrast, the abundance of the Candidatus Scalindua brodae and Nocardioides islandensis species in all four variants was lower than in sample ‘0’. The greatest increase in the share of OTUs (3.90%) in relation to sample ‘0’ was observed for the Arthrobacter psychrochitiniphilus species in the no-till variant. The greatest decrease in the share of OTUs (-1.25%) in relation to sample ‘0’ was observed for the Nocardioides islandensis species in the reduced tillage variant (Figure 7).
Aeromicrobium ponti is a species known for its potential to produce a variety of natural products and secondary metabolites with antimicrobial, antioxidative, and plant growth-stimulating properties [47]. It is a significantly functional species of great ecological importance for growing crops, especially in sustainable agriculture.
The Arthrobacter psychrochitiniphilus species has oligocarbophilic properties, i.e. economical carbon management. The bacteria have low temperature requirements. According to the reference publications, the Arthrobacter genus belongs to the group of plant growth-promoting bacteria (PGPB) due to its metabolic activity (the production of hormones, increasing the availability of nutrients, the production of antifungal substances, colonisation of plant roots, etc.). It offers an environment-friendly approach to improved plant production and counteracting the negative effects of abiotic stress [48,49].
Gemmatimonas aurantiaca is another species with an extremely important environmental role. In our study, the most OTUs of this species were observed in variants 3 – 1.05%), 4 – 0.93%), and 5 – 2.53%, i.e. in all no-till technologies (Figure 6). This species was also found in sample ‘0’ and in the conventional cultivation variant, but the shares of OTUs were much lower, i.e. 0.12% and 0.59%, respectively. Studies have shown that Gemmatimonas aurantiaca can reduce nitrous oxide (N2O). N2O is one of the most important greenhouse gases degrading the ozone layer of the stratosphere. Since the beginning of the industrial era its concentration in the atmosphere has increased significantly (from 270 ppb to 332 ppb). One of the reasons for this increase is the widespread use of nitrogen fertilisers in agriculture [50]. Microorganisms synthesising nitrous oxide reductase (NosZ), including Gemmatimonas aurantiaca, are the only known biological factor capable of reducing the emission of soil-derived N2O to an extent that is significant for the environment [51].
4. Conclusions
It is widely known that both soil fungi and bacteria play a key role in the course of most biochemical changes occurring in the soil as well as in the maintenance of soil health. In recent years, there have been numerous studies on the influence of agrotechnical factors related to the use of new technologies, including no-till farming systems, on the composition of communities of soil bacteria. However, it is still difficult to clearly determine the influence of individual factors on the structure and diversity of the soil microbiome and to directly relate these parameters to soil fertility and crop productivity. This is due to the large diversity of soils as well as other environmental and agrotechnical factors. So far there have been controversial results of studies on the influence of a specific farming technology on communities of soil bacteria. There are a few criteria defining the optimal quantitative, trophic, and taxonomic structure of the microbiome in the context of soil health and environmental functions. Therefore, further research is necessary in order to indicate the phyla or even species of bacteria or other groups of environment-friendly microorganisms for climate change and for the fertility of the soil.
Our study showed that the no-till cultivation technologies, mainly the strip till and no-till methods, applied at the horse bean plantation had a positive influence on microbial communities. The number of OTUs of species such as Gemmatimonas aurantiaca and Aeromicrobium ponti increased. The abundance of these species determines soil fertility and yield of crops. They are also environment- and climate-friendly, and therefore very important for sustainable or even regenerative agriculture. Although at present it is impossible to make an unequivocal statement, the results of our unpublished studies in soil samples collected from the plantations of other species from the Fabaceae family (soybean, white lupine) pointed to the presence of Gemmatimonas aurantiaca only at the plantations of species representing this botanical family, regardless of the cultivation system. Further investigations of soil samples collected from the plantations of crops belonging to the Fabaceae family may indicate functional species of microorganisms which are important for climate change. A larger number of results based on further investigations would not only unambiguously confirm the fact that the cultivation of species of the Fabaceae family has a positive influence on soil fertility. They would also facilitate limiting climate change by reducing the emissions of N2O, which is one of the most important greenhouse gases and a factor degrading the stratospheric ozone layer. These emissions could be reduced by a soil biological factor, i.e. the Gemmatimonas aurantiaca species.
Author Contributions
Conceptualization, D.S., A.N., A.W-M. and J.B.; methodology, D.S., A.N., A.W-M.; software, J.B. and R.G.; validation, D.S., A.N., A.W-M. and A.P.; formal analysis, D.S., A.N., A.W-M. and A.P.; investigation, D.S., A.N., and A.W-M.; resources, R.G and J.B.; data curation, D.S., A.N., A.W-M.; writing—original draft preparation, D.S., J.B., A.W-M., A.S. and A.N; writing—review and editing, D.S., A.N., A.W-M., A.S. and J.B.; visualization, J.B, R.G. and A.P.; supervision, D.S., A.N. and J.B.; project administration, D.S., A.S. and A.N.; funding acquisition, J.B. All authors have read and agreed to the published version of the manuscript.
Funding
Project “Increasing the use of domestic feed protein for the production of high-quality animal products in sustainable development conditions” (2016–2020). Research area no. 3 “Agrotechnical methods for increasing the use of the biological potential of legumes in terms of production, environmental, and economic effects”, Ministry of Agriculture and Rural Development. Thanks to retired Professor Irena Małecka from the Department of Agronomy, Poznań University of Life Sciences.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kopittke, P.M.; Menzies, N.W.; Wang, P.; McKenna, B.A.; Lombi, E. Soil and the intensification of agriculture for global food security. Environ. Int. 2019, 132, 105078. [Google Scholar] [CrossRef] [PubMed]
- Sanderman, J.; Hengl, T.; Fiske, G.J. Soil carbon debt of 12 000 years of human land use. Proc. Natl. Acad. Sci. USA 2017, 114, 9575–9580. [Google Scholar] [CrossRef] [PubMed]
- Jha, P.; Hati, K.M.; Dalal, R.C.; Dang, Y.P.; Kopittke, P.M.; McKenna, B.A.; Menzies, N.W. Effect of 50 Years of No-Tillage, Stubble Retention, and Nitrogen Fertilization on Soil Respiration, Easily Extractable Glomalin, and Nitrogen Mineralization. Agronomy 2022, 12, 151. [Google Scholar] [CrossRef]
- Swędrzyńska, D.; Grześ, S. Microbiological parameters of soil under sugar beet as a response to the long-term application of different tillage systems. Pol. J. Environ. Stud. 2015, 24(1), 285–294. [Google Scholar] [CrossRef]
- Schlatter, D.C.; Kahl, K.; Carlson, B.; Huggins, D.R.; Paulitz, T. Soil acidification modifies soil depth-microbiome relationships in a no-till wheat cropping system. Soil Biol. Biochem. 2020, 149, 107939. [Google Scholar] [CrossRef]
- Melero, S.; Panettieri, M.; Madejón, E.; Gómez Macpherson, H.; Moreno, F.; Murillo, J.M. Implementation of chiselling and mouldboard ploughing in soil after 8 years of no-till management in SW, Spain: Effect on soil quality. Soil Tillage Res. 2011, 112, 107–113. [Google Scholar] [CrossRef]
- Zhang, Z.; Liang, S.; Wang, J.; Zhang, X.; Mahamood, M.; Yu, J.; Zhang, X.; Liang, A.; Liang, W. Tillage and crop succession effects on soil microbial metabolic activity and carbon utilization in a clay loam soil. Eur. J. Soil Biol. 2018, 88, 97–104. [Google Scholar] [CrossRef]
- Cordeau, S.; Baudron, A.; Adeux, G. Is Tillage a Suitable Option for Weed Management in Conservation Agriculture? Agronomy 2020, 10, 1746. [Google Scholar] [CrossRef]
- Kubiak, A.; Wolna-Maruwka, A.; Niewiadomska, A.; Pilarska, A.A. The Problem of Weed Infestation of Agricultural Plantations vs. the Assumptions of the European Biodiversity Strategy. Agronomy 2022, 12, 1808. [Google Scholar] [CrossRef]
- Małecka-Jankowiak, I.; Blecharczyk, A.; Swedrzynska, D.; Sawinska, Z.; Piechota, T. The effect of long-term tillage systems on some soil properties and yield of pea (Pisum sativum L.). Acta Scientiarum Polonorum. Agricultura, 2016; 15(1), 37–50. [Google Scholar]
- Swędrzyńska, D.; Małecka-Jankowiak, I. The Impact of Tillaging Spring Barley on Selected Chemical, Microbiological, and Enzymatic Soil Properties. Pol. J. Environ. Stud. 2017, 26, 303–313. [Google Scholar] [CrossRef]
- Nikitin, D.A.; Ivanova, E.A.; Zhelezova, A.D.; Semenov, M.V.; Gadzhiumarov, R.G.; Tkhakakhova, A.K.; Chernov, T.I.; Ksenofontova, N.A.; Kutovaya, O.V. Assessment of the impact of no-till and conventional tillage technologies on the microbiome of southern agrochernozems. Eurasian Soil Sci. 2020, 53, 1782–1793. [Google Scholar] [CrossRef]
- Wieczorek, R.; Zydlik, Z.; Wolna-Maruwka, A.; Niewiadomska, A.; Kayzer, D. The Effect of Biofumigation on the Microbiome Composition in Replanted Soil in a Fruit Tree Nursery. Agronomy 2023, 13(10), 2507. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World Reference Base for Soil Resources. International Soil Classification System for Naming Soil and Creating Legends for Soils and creating Legends for Soil Maps. 4th edition. International Union of Soil Sciences (IUSS) Vienna, Austria, 2022.
- Selyaninov, G.T. About climate agricultural estimation. Proceedings on Agricultural Meteorology 1928, 20, 165–177. [Google Scholar]
- Taparauskiene, L.; Miseckaite, O. Comparison of watermark soil moisture content with Selyaninov hydrothermal coefficient. Agrofor, 2017; 2(2), 106–115. [Google Scholar]
- Chmist-Sikorska, J.; Kępińska-Kasprzak, M.; Struzik, P. Agricultural drought assessment on the base of Hydro-thermal Coefficient of Selyaninov in Poland. Ital. J. Agrometeorol. 2022, 1, 3–12. [Google Scholar] [CrossRef]
- Kozak, M.; Bocianowski, J.; Sawkojć, S.; Wnuk, A. Call for more graphical elements in statistical teaching and consultancy. Biom. Lett. 2010, 47, 57–68. [Google Scholar]
- VSN International Genstat for Windows, 23rd ed.; VSN International: Hemel Hempstead, UK, 2023.
- Li, B.B.; Roley, S.S.; Duncan, D.S.; Guo, J.; Quensen, J.F.; Yu, H.-Q.; Tiedje, J.M. Long-Term Excess Nitrogen Fertilizer Increases Sensitivity of Soil Microbial Community to Seasonal Change Revealed by Ecological Network and Metagenome Analyses. Soil Biol. Biochem. 2021, 160, 108349. [Google Scholar] [CrossRef]
- Gao, W.H.; Gao, K.; Guo, Z.H.; Liu, Y.; Jiang, L.; Liu, C.; Liu, X.Y.; Wang, G.L. Different Responses of Soil Bacterial and Fungal Communities to 3 Years of Biochar Amendment in an Alkaline Soybean Soil. Front. Microbiol. 2021, 12, 630418.
- Bao, Y.Y.; Feng, Y.Z.; Stegen, J.C.; Wu, M.; Chen, R.R.; Liu, W.J.; et al. Straw chemistry links the assembly of bacterial communities to decomposition in paddy soils. Soil Biol Biochem. 2020, 148, 107866. [Google Scholar] [CrossRef]
- Liu, Y.R.; Delgado-Baquerizo, M.; Wang, J.T.; Hu, H.W.; Yang, Z.M.; He, J.Z. New insights into the role of microbial community composition in driving soil respiration rates. Soil Biol Biochem. 2018, 118, 35–41. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88(6), 1354–1364. [Google Scholar] [CrossRef]
- Mahnkopp-Dirks, F.; Radl, V.; Kublik, S.; Gschwendtner, S.; Schloter, M.; Winkelmann, T. Dynamics of bacterial root endophytes of Malus domestica plants grown in field soils affected by apple replant disease. Front. Microbiol. 2022, 13, 841558. [Google Scholar] [CrossRef]
- Tang, Z.; Zhang, L.; He, N.; Gong, D.; Gao, H.; Ma, Z.; Fu, L.; Zhao, M.; Wang, H.; Wang, C.; Zheng, W.; Zhang, W. . Soil bacterial community as impacted by addition of rice straw and 649 biochar. Sci, Rep. 2021; 11, 22185. [Google Scholar] [CrossRef]
- Bünger, W.; Jiang, X.; Müller, J.; Hurek, T.; Reinhold-Hurek, B. Novel cultivated endophytic Verrucomicrobia reveal deep-rooting traits of bacteria to associate with plants. Sci. Rep. 2020, 10, 8692. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Baquerizo, M.; Oliverio, A.M.; Brewer, T.E.; Benavent-González, A.; Eldridge, D.J.; Bardgett, R.D.; Maestre, F.T.; Singh, B.K.; Fierer, N. A global atlas of the dominant bacteria found in soil. Science 2018, 359, 320–325. [Google Scholar] [CrossRef]
- DeBruyn, J.M.; Nixon, L.T.; Fawaz, M.N.; Johnson, A.M.; Radosevich, M. Global Biogeography and Quantitative Seasonal Dynamics of Gemmatimonadetes in Soil. Appl. Environ. Microbiol. 2011, 77, 6295–6300. [Google Scholar] [CrossRef] [PubMed]
- Mujakić, I.; Piwosz, K.; Koblížek, M. Phylum Gemmatimonadota and Its Role in the Environment. Microorganisms 2022, 10(1), 151. [Google Scholar] [CrossRef]
- Liu, M.; Li, X.; Zhu, R.; Chen, N.; Ding, L.; Chen, C. Vegetation richness, species identity and soil nutrients drive the shifts in soil bacterial communities during restoration process. Environ. Microbiol. Rep. 2021, 13, 1758–2229. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Bai, X.; Zhou, Y.; Zhu, W.; Yin, Y. Variations of soil microbial communities accompanied by different vegetation restoration in an open-cut iron mining area. Sci. Total Environ. 2020, 704, 135243. [Google Scholar] [CrossRef]
- Ye, W.; Liu, X.; Lin, S.; Tan, J.; Pan, J.; Li, D.; Yang, H. The vertical distribution of bacterial and archaeal communities in the water and sediment of Lake Taihu. FEMS Microbiol. Ecol. 2009, 70, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Tang, X.; Hou, Q.; Li, T.; Xie, H.; Lu, Z.; & Wen, X. Response of soil organic carbon fractions to legume incorporation into cropping system and the factors affecting it: A global meta-analysis. Agric. Ecosyst. Environ. 2023, 342, 108231. [Google Scholar] [CrossRef]
- Lee, S.A.; Kim, J.M.; Kim, Y.; Joa, J.H.; Kang, S.S.; Ahn, J.H.; Kim, M; Song, J.; Weon, H.Y. Different types of agricultural land use drive distinct soil bacterial communities. Sci. Rep. 2020, 10(1), 17418. [Google Scholar]
- Li, M.; He, P.; Guo, X.L.; Zhang, X.; Li, L.J. Fifteen-year no tillage of a Mollisol with residue retention indirectly affects topsoil bacterial community by altering soil properties. Soil Tillage Res. 2021, 205, 104804. [Google Scholar] [CrossRef]
- Khmelevtsova, L.E.; Sazykin, I.S.; Azhogina, T.N.; Sazykina, M.A. Influence of agricultural practices on bacterial community of cultivated soils. Agriculture 2022, 12(3), 371. [Google Scholar] [CrossRef]
- Mitra, D.; Mondal, R.; Khoshru, B.; Senapati, A.; Radha, T.K.; Mahakur, B.;... & Mohapatra, P.K.D. Actinobacteria-enhanced plant growth, nutrient acquisition, and crop protection: Advances in soil, plant, and microbial multifactorial interactions. Pedosphere, 2022; 32(1), 149–170. [Google Scholar]
- Becker, A.; Overlöper, A.; Schlüter, J.P.; Reinkensmeier, J.; Robledo, M.; Giegerich, R.; Narberhaus, F.; Evguenieva-Hackenberg, E. Riboregulation in plant-associated α-proteobacteria. RNA Biology 2014, 11(5), 550–562. [Google Scholar] [CrossRef] [PubMed]
- Klūga, A.; Dubova, L.; Alsiņa, I.; Rostoks, N. Alpha-, gamma-and beta-proteobacteria detected in legume nodules in Latvia, using full-length 16S rRNA gene sequencing. Acta Agric. Scand. Sect. B 2023, 73(1), 127–141. [Google Scholar] [CrossRef]
- Batut, J.; Andersson, S.G.; O’Callaghan, D. The evolution of chronic infection strategies in the α-proteobacteria. Nat. Rev. Microbiol. 2004, 2(12), 933–945. [Google Scholar] [CrossRef]
- Anandan, R.; Dharumadurai, D.; Manogaran, G.P. An introduction to actinobacteria. In: Actinobacteria-basics and biotechnological applications. IntechOpen, 2016.
- Mokni-Tlili, S.; Mehri, I.; Ghorbel, M.; Hassen, W.; Hassen, A.; Jedidi, N.; Hamdi, H. Community-level genetic profiles of actinomycetales in long-term biowaste-amended soils. Arch. Microbiol. 2020, 202, 2607–2617. [Google Scholar] [CrossRef]
- Mandic-Mulec, I.; Stefanic, P.; van Elsas, J.D. Ecology of Bacillaceae. The bacterial spore: From molecules to systems 2016, 59–85. [CrossRef]
- Masson-Boivin, C.; Giraud, E.; Perret, X.; Batut, J. Establishing nitrogen-fixing symbiosis with legumes: how many rhizobium recipes? Trends Microbiol. 2009, 17(10), 458–466. [Google Scholar] [CrossRef]
- Li, Y.; Pan, F.; Yao, H. Response of symbiotic and asymbiotic nitrogen-fixing microorganisms to nitrogen fertilizer application. J. Soils Sediments 2019, 19, 1948–1958. [Google Scholar] [CrossRef]
- Anavadiya, B.; Chouhan, S.; Saraf, M.; Goswami, D. Exploring Endophytic Actinomycetes: A Rich Reservoir of Diverse Antimicrobial Compounds for Combatting Global Antimicrobial Resistance. The Microbe 2024, 100110. [Google Scholar] [CrossRef]
- Platamone, G.; Procacci, S.; Maccioni, O.; Borromeo, I.; Rossi, M.; Bacchetta, L.; Forni, C. Arthrobacter sp. inoculation improves cactus pear growth, quality of fruits, and nutraceutical properties of cladodes. Curr. Microbiol. 2023; 80(8), 266. [Google Scholar]
- Chhetri, G.; Kim, I.; Kang, M.; So, Y.; Kim, J.; Seo, T. An isolated Arthrobacter sp. enhances rice (Oryza sativa L.) plant growth. Microorganisms, 2022; 10(6), 1187. [Google Scholar]
- Forster, P.; Ramaswamy, V.; Artaxo, P.; Berntsen, T.; Betts, R.; Fahey, D.W.; Haywood, J.; Lean, J.; Lowe, D.C.; Myhre, G.; Nganga, J.; Prinn, R.; Raga, G.; Schulz, M.; Van Dorland, R. Changes in Atmospheric Constituents and in Radiative Forcing. In: Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change [Solomon, S.; Qin, D.; Manning, M.; Chen, Z.; Marquis, M.; Averyt, K.B.; Tignor, M.; Miller, H.L. (eds.)]. Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA, 2007.
- Park, D.; Kim, H.; Yoon, S. Nitrous oxide reduction by an obligate aerobic bacterium, Gemmatimonas aurantiaca strain T-27. Appl. Environ. Microbiol. 2017, 83(12), e00502-17. [Google Scholar] [CrossRef]
Figure 1.
The percentages of operational taxonomic units (OTUs) of dominant phyla of bacteria. The classifications with less than 1% abundance are gathered into the category “Other” (1 – plantless, fallow buffer zone of the field experiment – sample ‘0’, 2 – conventional tillage, 3 – reduced tillage, 4 – strip tillage, 5 – no tillage).
Figure 1.
The percentages of operational taxonomic units (OTUs) of dominant phyla of bacteria. The classifications with less than 1% abundance are gathered into the category “Other” (1 – plantless, fallow buffer zone of the field experiment – sample ‘0’, 2 – conventional tillage, 3 – reduced tillage, 4 – strip tillage, 5 – no tillage).
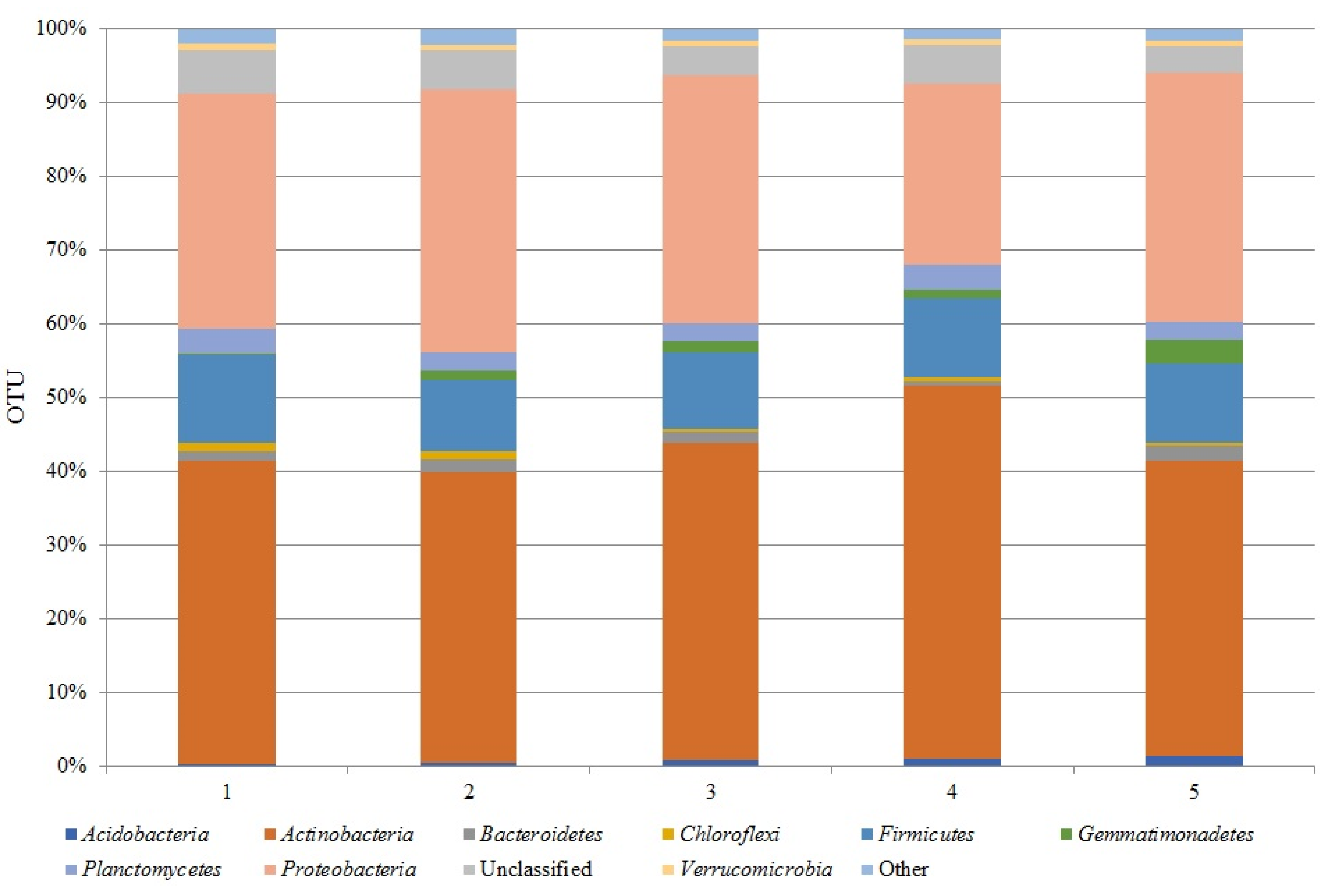
Figure 2.
Relative abundance of dominant phyla of bacteria. The classifications with less than 1% abundance are gathered into the category “other” (1 – plantless, fallow buffer zone of the field experiment – sample ‘0’, 2 – conventional tillage, 3 – reduced tillage, 4 – strip tillage, 5 – no tillage; "red" means negative difference; “green” means positive difference).
Figure 2.
Relative abundance of dominant phyla of bacteria. The classifications with less than 1% abundance are gathered into the category “other” (1 – plantless, fallow buffer zone of the field experiment – sample ‘0’, 2 – conventional tillage, 3 – reduced tillage, 4 – strip tillage, 5 – no tillage; "red" means negative difference; “green” means positive difference).
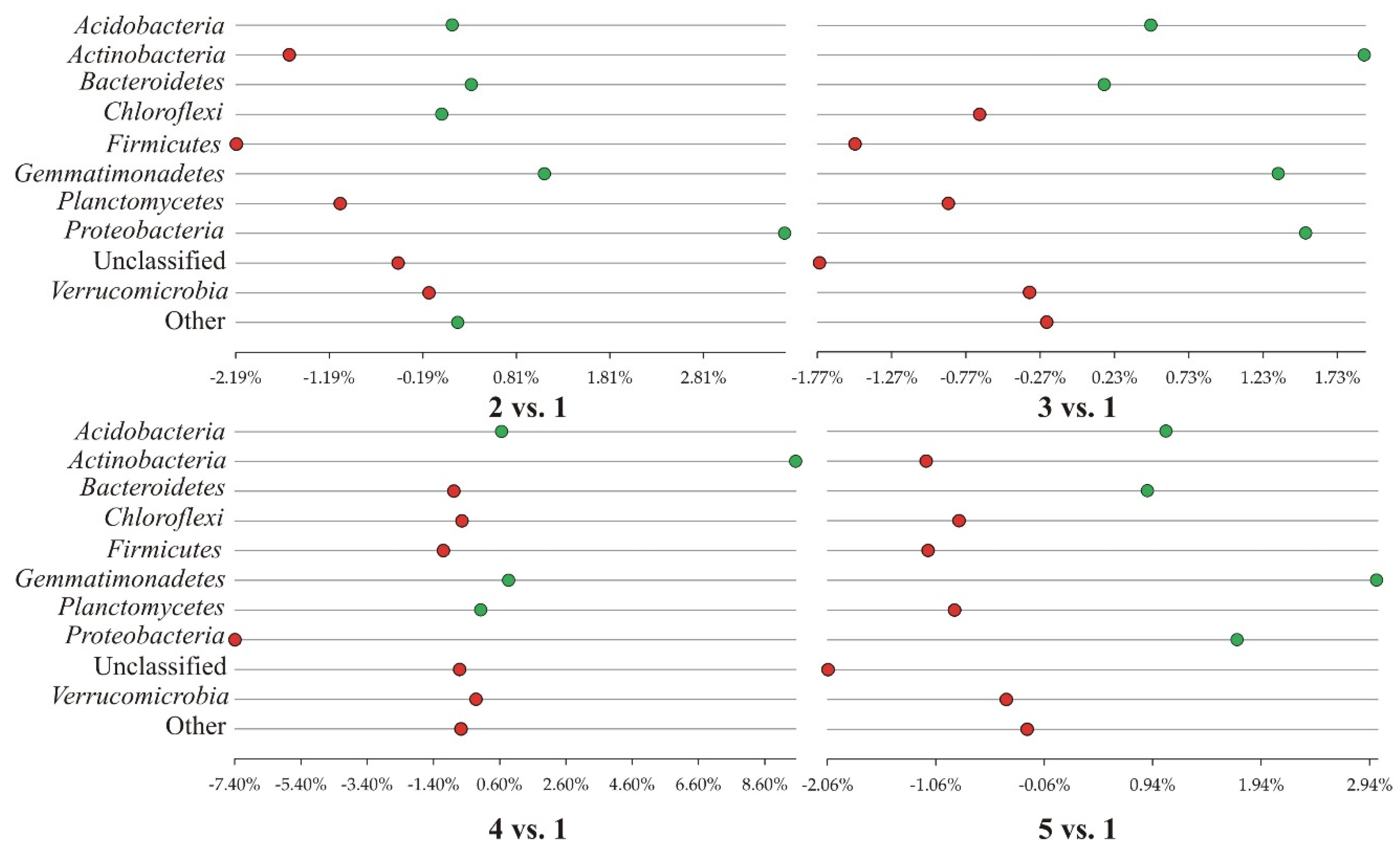
Figure 3.
The percentages of operational taxonomic units (OTUs) of the dominant classes of bacteria. The classifications with less than 1% abundance are gathered into the category “Other” (1 – plantless, fallow buffer zone of the field experiment – sample ‘0’, 2 – conventional tillage, 3 – reduced tillage, 4 – strip tillage, 5 – no tillage).
Figure 3.
The percentages of operational taxonomic units (OTUs) of the dominant classes of bacteria. The classifications with less than 1% abundance are gathered into the category “Other” (1 – plantless, fallow buffer zone of the field experiment – sample ‘0’, 2 – conventional tillage, 3 – reduced tillage, 4 – strip tillage, 5 – no tillage).
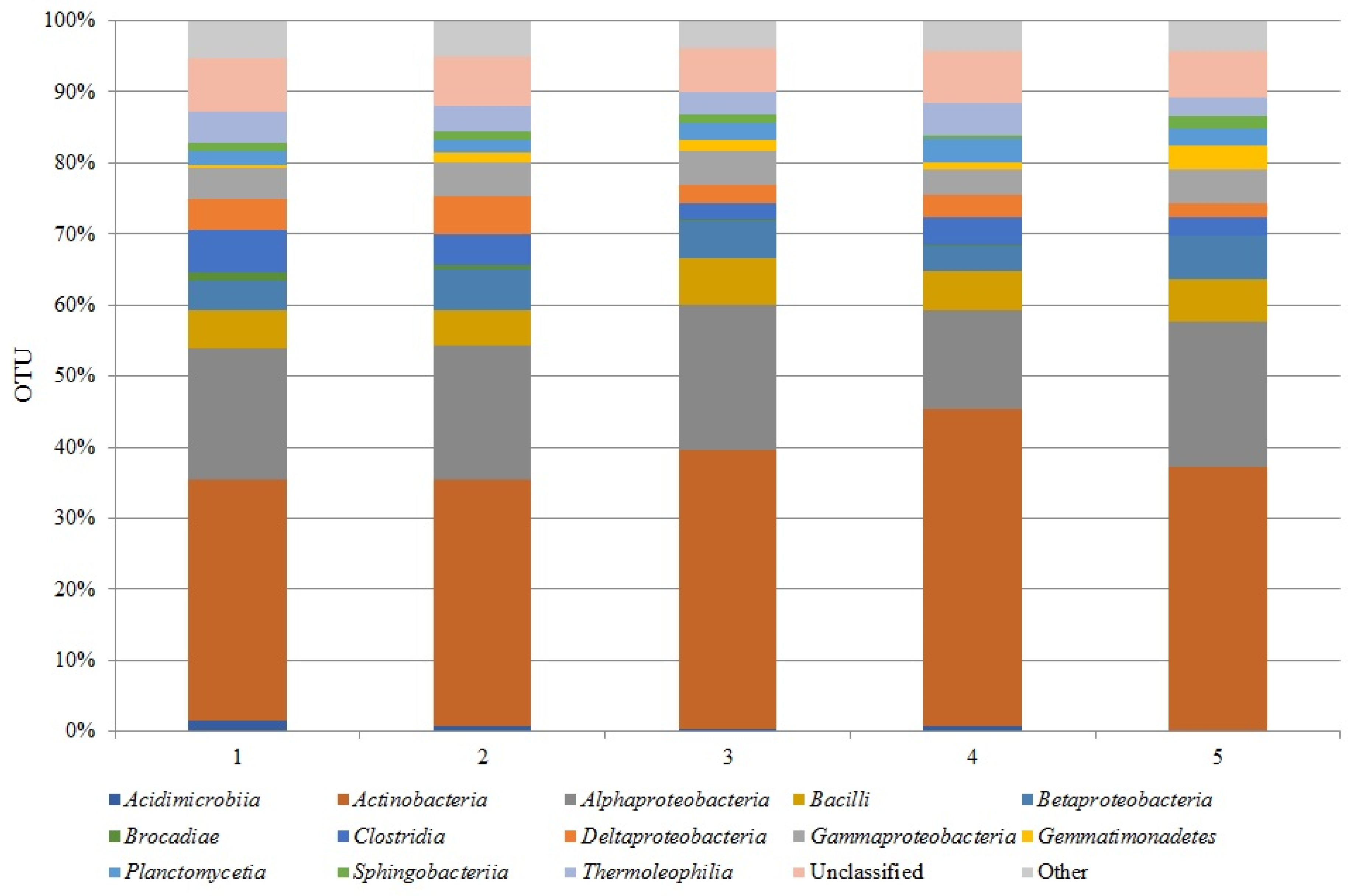
Figure 4.
Relative abundance of dominant classes of bacteria. The classifications with less than 1% abundance are gathered into the category “other” (1 – plantless, fallow buffer zone of the field experiment – sample ‘0’, 2 – conventional tillage, 3 – reduced tillage, 4 – strip tillage, 5 – no tillage; "red" means negative difference; “green” means positive difference).
Figure 4.
Relative abundance of dominant classes of bacteria. The classifications with less than 1% abundance are gathered into the category “other” (1 – plantless, fallow buffer zone of the field experiment – sample ‘0’, 2 – conventional tillage, 3 – reduced tillage, 4 – strip tillage, 5 – no tillage; "red" means negative difference; “green” means positive difference).
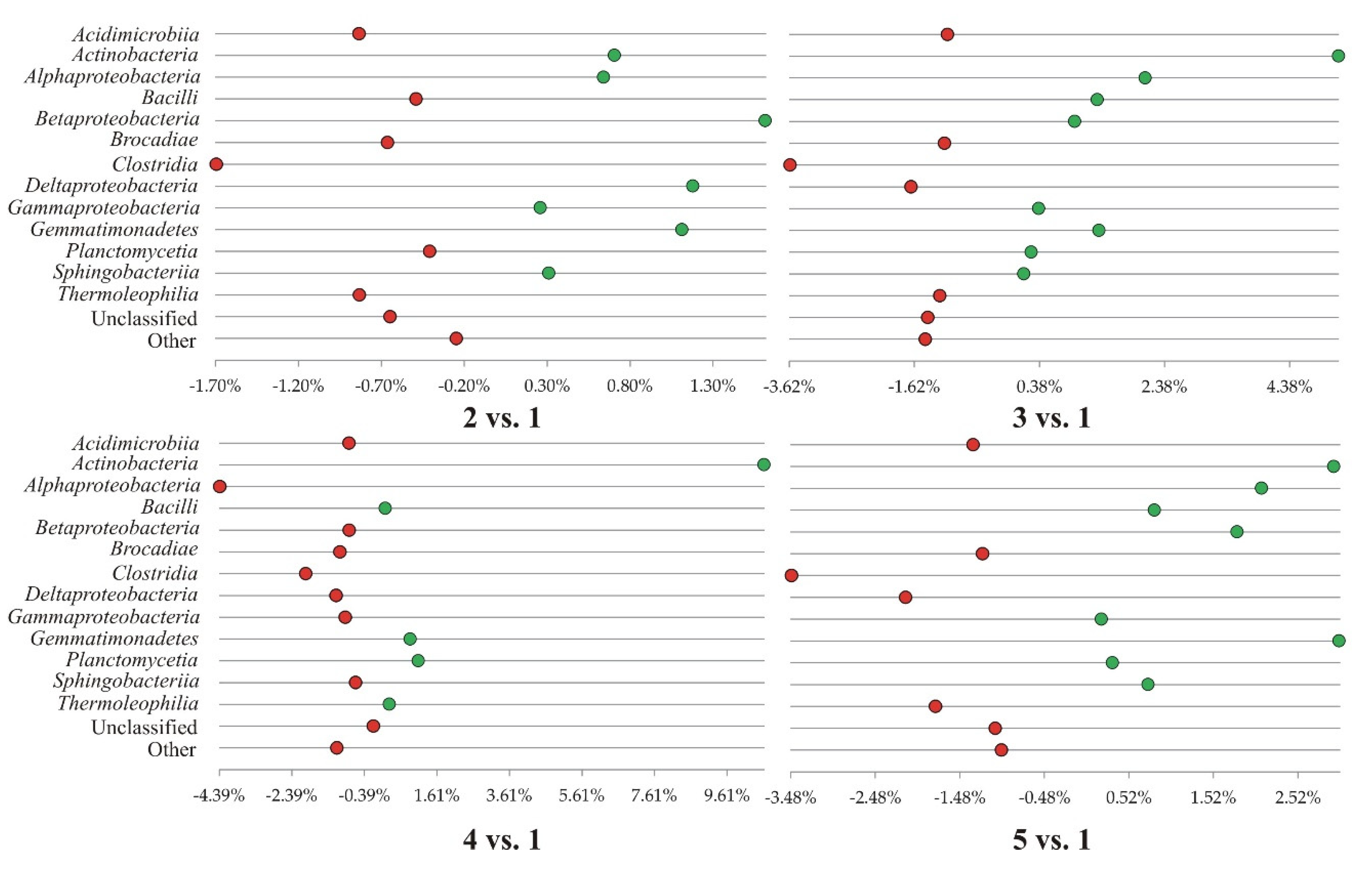
Figure 5.
The percentages of operational taxonomic units (OTUs) of the dominant orders of bacteria. The classifications with less than 1% abundance are gathered into the category “Other” (1 – plantless, fallow buffer zone of the field experiment – sample ‘0’, 2 – conventional tillage, 3 – reduced tillage, 4 – strip tillage, 5 – no tillage).
Figure 5.
The percentages of operational taxonomic units (OTUs) of the dominant orders of bacteria. The classifications with less than 1% abundance are gathered into the category “Other” (1 – plantless, fallow buffer zone of the field experiment – sample ‘0’, 2 – conventional tillage, 3 – reduced tillage, 4 – strip tillage, 5 – no tillage).
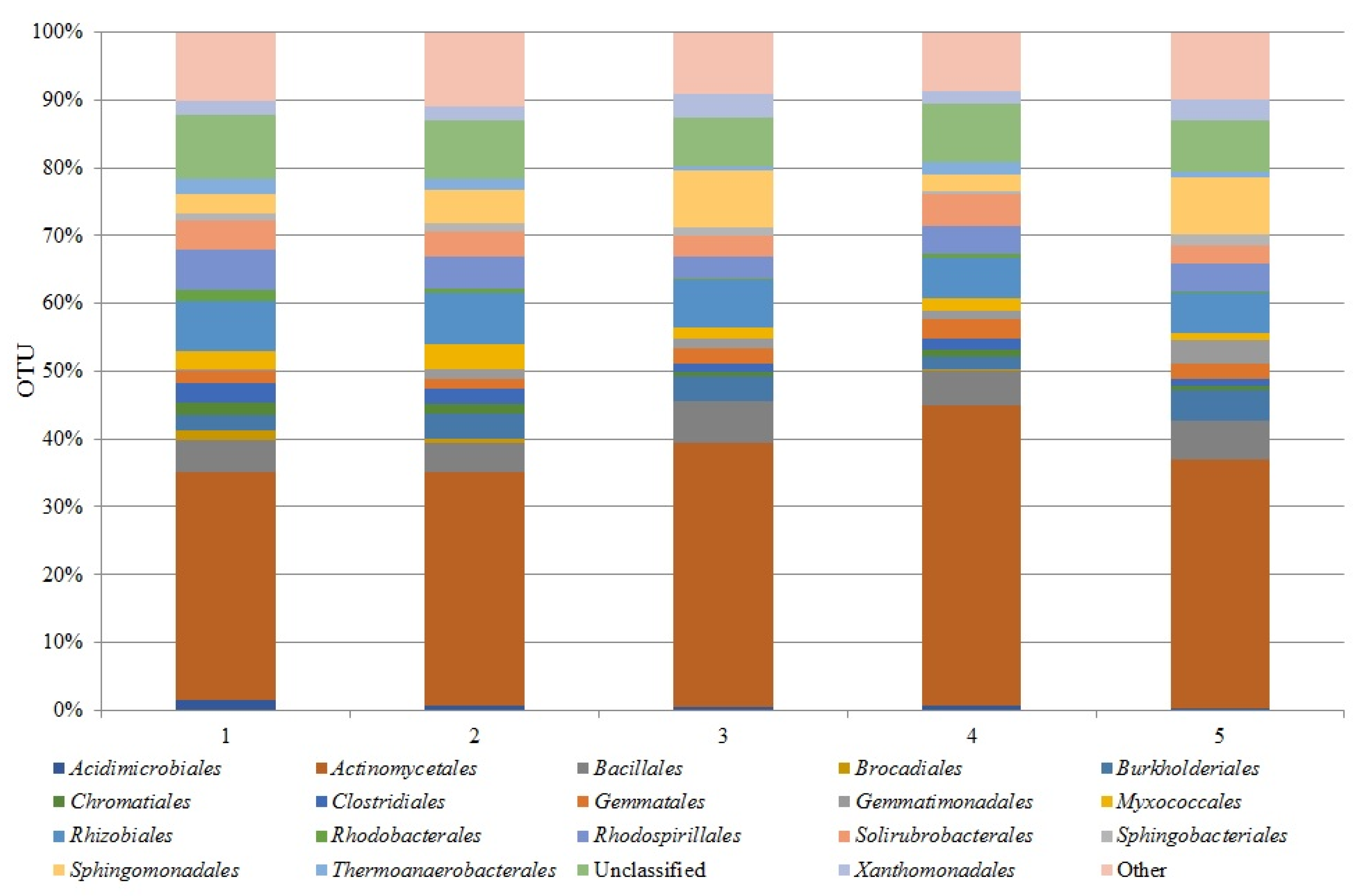
Figure 6.
The percentages of operational taxonomic units (OTUs) of the dominant species of bacteria. The classifications with less than 1% abundance are gathered into the category “Other” (1 – plantless, fallow buffer zone of the field experiment – sample ‘0’, 2 – conventional tillage, 3 – reduced tillage, 4 – strip tillage, 5 – no tillage).
Figure 6.
The percentages of operational taxonomic units (OTUs) of the dominant species of bacteria. The classifications with less than 1% abundance are gathered into the category “Other” (1 – plantless, fallow buffer zone of the field experiment – sample ‘0’, 2 – conventional tillage, 3 – reduced tillage, 4 – strip tillage, 5 – no tillage).
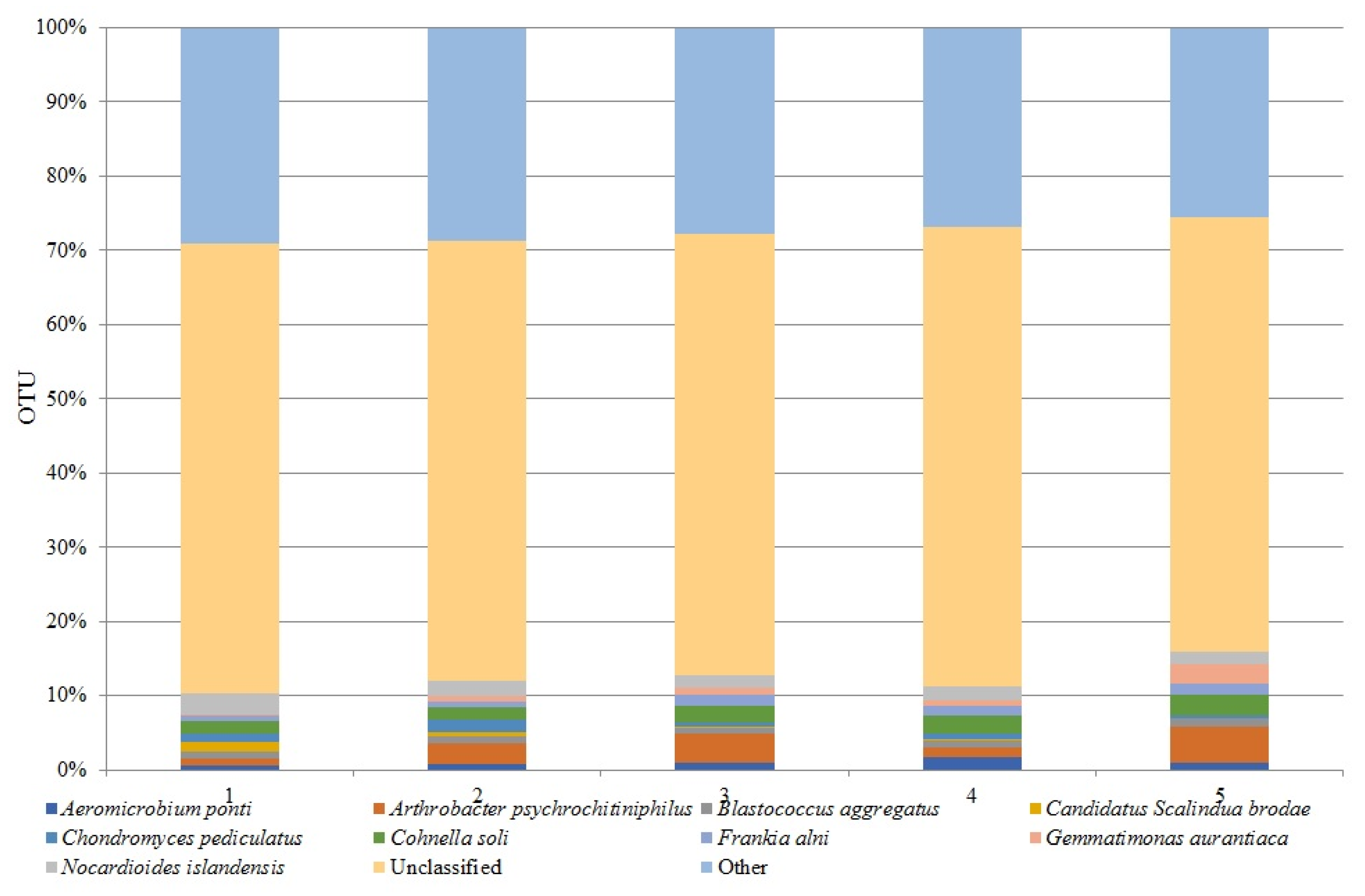
Figure 7.
Relative abundance of dominant bacteria species. The classifications with less than 1% abundance are gathered into the category “other” (1 – plantless, fallow buffer zone of the field experiment – sample ‘0’, 2 – conventional tillage, 3 – reduced tillage, 4 – strip tillage, 5 – no tillage; "red" means negative difference; “green” means positive difference).
Figure 7.
Relative abundance of dominant bacteria species. The classifications with less than 1% abundance are gathered into the category “other” (1 – plantless, fallow buffer zone of the field experiment – sample ‘0’, 2 – conventional tillage, 3 – reduced tillage, 4 – strip tillage, 5 – no tillage; "red" means negative difference; “green” means positive difference).
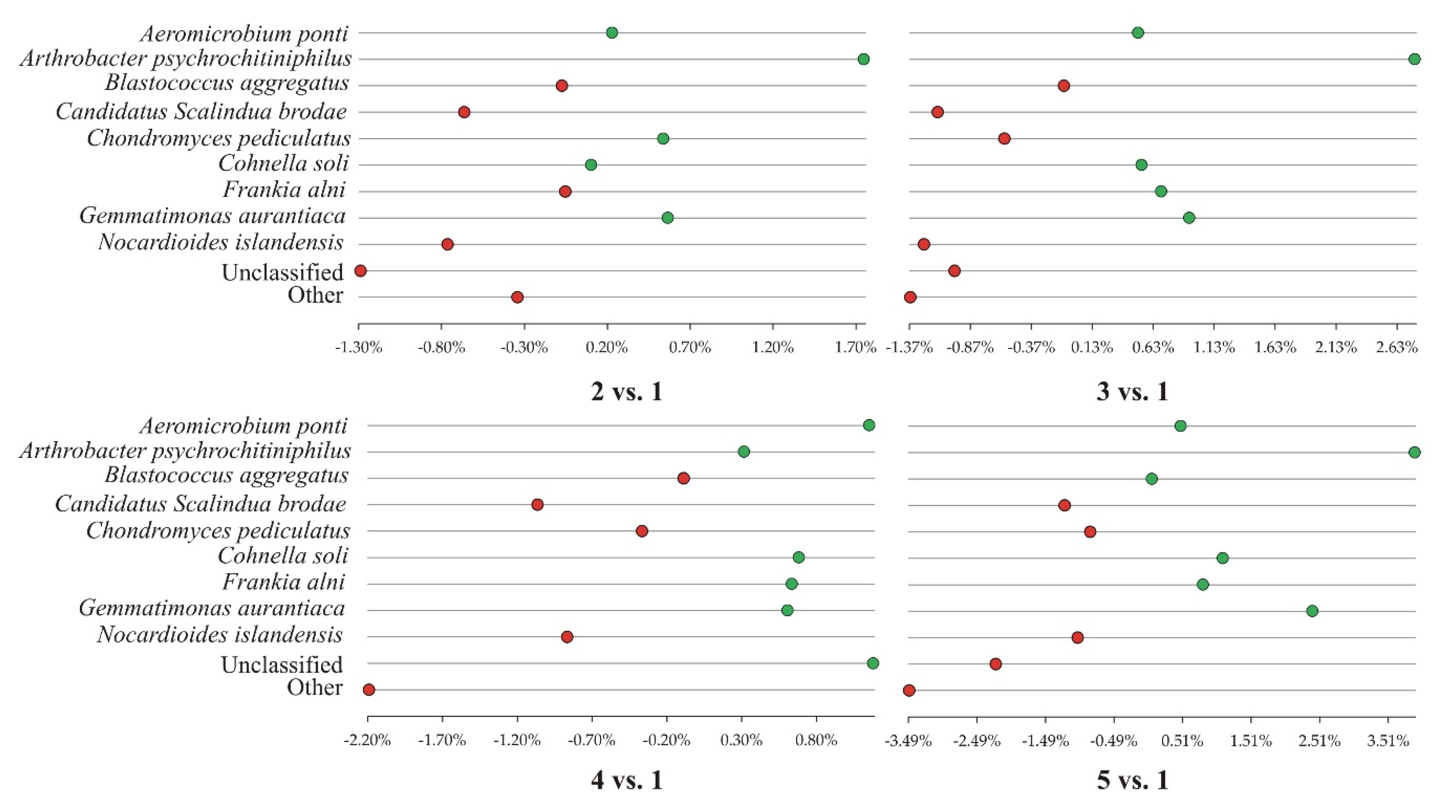

Table 1.
The monthly and ten-day values of the HTC in the growing season.
| Period | Month | |||||||||||||
| Apr | May | June | July | Aug | Sep | |||||||||
| 1st ten days | 0.00 | 1.73 | 0.03 | 0.32 | 0.65 | 1.75 | ||||||||
| 2nd ten days | 0.96 | 4.71 | 0.06 | 1.39 | 0.50 | 0.62 | ||||||||
| 3rd ten days | 0.52 | 1.67 | 0.28 | 1.32 | 0.22 | 1.94 | ||||||||
| month | 0.45 | 2.56 | 0.13 | 1.05 | 0.44 | 1.47 | ||||||||
| >3.0 | 2.6-3.0 | 2.1-2.6 | 1.7-2.1 | 1.4-1.7 | 1.1-1.4 | 0.8-1.1 | 0.4-0.8 | <0.4 | ||||||
| Extremely humid | Very humid | Humid | Quite humid | Optimum | Quite dry | Dry | Very dry | Extremely dry | ||||||
Table 2.
The Kingdom classification results.
| Classification | Experiment variants | ||||
| 1 | 2 | 3 | 4 | 5 | |
| sample ‘0’ plantless, fallow buffer zone of the field experiment | conventional tillage | reduced tillage | strip tillage, | no tillage, | |
| Total reads [%] | |||||
| Bacteria | 0.01% | 0.00% | 0.00% | 0.00% | 0.00% |
| Unclassified at Kingdom level | 99.96% | 99.97% | 99.98% | 99.96% | 99.98% |
| Archaea | 0.03% | 0.03% | 0.02% | 0.03% | 0.02% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



