
Submitted:
28 November 2024
Posted:
29 November 2024
You are already at the latest version
Abstract
Enterococci, commonly found in the normal intestinal flora of humans and animals, have emerged as an important human pathogen. A total of 184 isolates (88 isolates in 2015 and 96 isolates in 2016) were collected from 46 flocks. Two predominant enterococcus species were identified: Enterococcus faecalis (59%) and Enterococcus faecium (~39%). Resistance to penicillin was significantly decreased in the overall enterococci community, while it remained unchanged in the multi-class resistant (MCR) community. We identified emeA and efrAB genes, which encode efflux pump systems, in 93% (26/28) of the multi-class resistant (MCR) isolates with (intermediate) resistance to levofloxacin. The ermB gene was present in all MCR strains with resistance to erythromycin. The lsa gene was detected in 87% (84/97) of the MCR isolates with resistance to quinupristin/dalfopristin. About 82.2% of MCR strains in 2015 and 100% of MCR strains in 2016 carried insertion sequence IS256, which is known to be associated with AMR genes conferring resistance to erythromycin, gentamicin and vancomycin in enteroccocci. These results support the need for monitoring AMR in Gram positive bacteria in poultry production, specifically in broiler chicken farms, to complement current AMR data, and developing a timely intervention framework.
Keywords:
Enterococci
; Antimicrobial resistance
; Poultry production
; Resistance genes
; Multidrug resistance
1. Introduction
The broiler chicken sector has continued to grow in Canada since 2012 and contributed $6.8 billion to Canada’s Gross Domestic Product (2016) [1]. Over the past 30 years, per capita consumption of broiler chicken in Canada has been continuously increasing from 19.9 kg to 32.8 kg. In Alberta, the poultry industry provided 8,031 total jobs, contributed $623 million to the GDP and paid $203 million in taxes in 2016 [2].
Enterococci, previously classified as group D in the genus Streptococcus, have now been recognized as a distinct and unique genus [3]. They are gram positive bacteria that mainly inhabit the gastrointestinal tract of animal and humans [4]. However, they can be found anywhere in the environment (soil, surface water, sewage, plants, dairy products and raw retail meats, etc.) due to their great adaptability and persistent nature [5,6]. There was evidence of a transmission from pigs to human during an outbreak of E. faecium related-sepsis in China [7]. Thousands of domestic pigs died of hemorrhagic shock, while 40 local farmers who were in contact with them were hospitalized with severe illness. A more recent study showed the relatedness of vancomycin-resistant E. faecium strains originating from humans and animals [8].
Some species and strains can cause life-threatening infections in people due to virulent traits such as aggregation, adhesins, cytolysin, gelatinase, extracellular surface proteins and pheromones [9]. Two enterococci species that are of great human clinical concern are Enterococcus faecalis and Enterococcus faecium [10]. The estimated number of hospitalization related to enterococcal infection in the US was 130,922 in 2003 and 125,134 in 2004 [11]. Nosocomial enterococci are associated with various infections, such as urinary tract infections, intra-abdominal and pelvic abscesses (post-surgery wound infections), bacteremia, central nervous system and neonatal infections and, in rare cases, respiratory tract infections, osteomyelitis, or cellulitis [12]. Not only do enterococci cause nosocomial infection in humans, but they also cause systemic infection in a variety of birds [13,14]. E. faecium led to septicemia in ducklings, goslings, chickens, etc., while E. faecalis caused amyloid arthropathy in chickens. E. hirae was associated with encephalomalacia with vascular thrombosis and meningitis in broiler chicks. More recently, Enterococcus cecorum, primarily responsible for vertebral canal osteomyelitis, has emerged in Canadian flocks [15].
Intrinsic resistance in enterococci include the resistance to β-lactams and cephalosporins, aminoglycosides, trimethoprim-sulfamethoxazole, lincosamides and streptogramins [10,16]. This could occur due to chromosomal resistance genes, such as the lsa gene in E. faecalis. E. faecalis and E. faecium have different mechanisms of intrinsic resistance; hence, their resistance phenotypes differ [16]. For instance, E. faecalis is intrinsically resistant to clindamycin, streptogrammin A and B because it carries the lsa gene encoding ATP-binding cassette efflux pump on its chromosome [16]. E. faecium harbors another putative efflux pump encoded by the intrinsic msrC gene which confers low-level resistance (MIC 1–2 mg/ml) to streptogramin B compounds only [16]. However, not all resistance mechanisms in enterococci are well-understood. In addition to their intrinsic resistance to some antimicrobial agents, enterococci have shown the ability to acquire new AMR genes via plasmids and other mobile genetic elements or by chromosomal mutations [17]. Resistance against β-lactams, aminoglycosides (high level resistance), glycopeptides and streptogramins has been reported to be acquired via mobile genetic elements [10].
Mobile genetic elements are nucleic acid elements such as plasmids, and transposons which play a role in horizontal dissemination of AMR genes among bacterial community members [17]. Pheromone-responsive plasmids, which are self-transferable (conjugative) narrow-host-range plasmids such as pADI, pAM373 and pCF10, have been thoroughly studied in enterococci [10,18]. Enterococcal pheromones are small peptide fragments which are essential for cell-cell communication and high-frequency plasmid transfer in enterococci [19]. Their host range appears to be restricted to E. faecalis. Among them, only pCF10 has been shown to carry the tet(M)-containing conjugative transposon Tn925. Well-known broad-host-range Inc18 plasmids including pAMβ1 and pIP501 also contribute to disseminating AMR genes. Plasmid pAMβ1 harbors macrolide resistance genes, and pIP501 encodes genes for macrolides and chloramphenicol resistance. These plasmids have been shown to be transferable among enterococci species and between enterococci with other species (staphylococci and streptococci).
Enterococci transposons are fairly versatile and plastic elements which play a crucial role in the emergence and dissemination of AMR genes among enterococci [18]. Based on their composition and structure, they can be categorized into three groups: composite transposons, Tn3 family transposons and conjugative transposons [10,18]. Composite transposons contain flanking insertion sequence (IS) elements which confer mobility for this transposon group [20]. The typical structure of composite transposons has two IS elements in a direct or inverted orientation with resistance genes in between. The Tn3 family transposons carry two enzymes: a transposase (TnpA) and a resolvase (TnpR) which promotes the intracellular movement of transposons within or between different replicons on a single genome of a strain [21]. IS256 is located as part of multiple composite transposons such as Tn4001 (Staphylococcus aureus), Tn4031 (Staphylococcus epidermidis), Tn5281 (E. faecalis), Tn5384 (E. faecalis). Tn5281 confers resistance to gentamicin, while Tn5384 confers resistance to erythromycin and high levels of gentamicin [22,23,24]. The last group, conjugative transposons, are the most important group of transposons in AMR dissemination since they are able to move between various species, particularly between enterococci and streptococci. These transposons harbor all essential genes including recombinase/integrase gene (int) and excisionase gene (xis) which facilitate their intercellular transfer.
The Canadian Integrated Program for Antimicrobial Resistance Surveillance (CIPARS) monitors AMR in food-animals including poultry (turkeys and broiler chickens) at various points along the poultry production continuum. The vast majority of the antimicrobials used in Canadian poultry target Gram positive bacteria, though CIPARS does not focus on AMR research for Gram positive bacteria in poultry. To complement CIPARS research, this study characterized AMR patterns in fecal enterococci isolates from Alberta broiler chicken farms in 2015-2016. It also identified phenotype-associated resistance genes and associated mobile genetic elements.
2. Materials & Methods
2.1. Description of Sampling Method, Bacterial Isolation and Growth Condition
Pooled fecal samples were collected from a total of 46 flocks according to routine CIPARS methodology [25]. Briefly, the barn was arbitrarily divided into quadrants and 1 pooled fecal sample that consisted of 10 individual droppings was collected from each quadrant. A total of 4 pooled fecal samples were collected per farm. Enterococci isolation from broiler chicken feces was performed by Alberta Agriculture and Forestry in 2015 and 2016. The fecal sample was diluted (1 in 10) into Buffer Peptone Water (BPW) and incubated at 35°C for 24h. The primary enrichment was then plated onto selective agar, ENT (Enterococcosel Agar) and SBA (Slanetz and Bartley Agar) and incubated at 35°C and 42°C, respectively, for 24h. Typical colonies were streaked onto BAP (Blood Agar) for purity and incubated at 35°C for 24h. Isolates resembling enterococci were re-streaked to ENT and SBA and incubated at 35°C and 42°C, respectively, for 24-48hr. Typical colonies were confirmed biochemically with sugars (L-arabinose, D-mannitol, and α-methyl-D-glucoside), 6.5% NaCl and catalase testing. Four isolates per flock were then shipped to the University of Calgary for further characterization.
Tryptic soy agar (TSA) and tryptic soy broth (TSB) were purchased for enterococci cultivation from Criterion (1430 West McCoy Lane, Santa Maria, CA 93455, USA) and Becton Dickinson (2100 Derry Road West Suite 100, Mississauga, ON L5N 0B3). Enterococci were cultured at 37°C under aerobic conditions. Mueller-Hinton (MH) agar was purchased from Becton Dickinson company to be used for antimicrobial susceptibility tests.
2.2. Antimicrobial Susceptibility Assays
Disc diffusion was performed to characterize the AMR profile of isolates according to the Clinical and Laboratory Standards Institute (CLSI) guidelines [26]. Ready-to-use antibiotic discs were purchased from BD and Oxoid companies (1926 Merivale Rd, Nepean, ON K2G 1E8). Zone diameters were recorded and used to interpret resistance phenotypes based on zone diameter breakpoints of each drug in CLSI guidelines [26]. Screening tests for high-level aminoglycoside resistance (gentamicin and streptomycin) were performed using 120-µg gentamicin and 300-µg streptomycin discs which were made by adding antibiotic solution to blank discs at appropriate concentrations [27].
Isolates considered MCR (resistance to ≥ 3 drug classes) were subjected to susceptibility testing for daptomycin using daptomycin 256 - 0.015µg/ml M.I.C. Evaluator Strips (MA0125) purchased from the Oxoid company.
2.3. Enterococci Speciation
Enterococci were identified using a PCR assay as described previously [28]. In this method, species-specific genes encoding D-alanyl-D-alanine ligase (ddlE. faecalisor ddlE. faecium) or alternatively the ligase responsible for intrinsic vancomycin resistance (vanC-1 in E. gallinarum and vanC-2/3 in E. casseliflavus/E. flavescens) [28]. Isolates that were not able to be identified using above method were subsequently identified using the method described by Zaheer et al. [29]. This method targets groES-EL spacer region which is hypervariable among Enterococcus species.
2.4. Detection and Identification of Resistance Genes
A PCR assay was used to identify various AMR genes in MCR isolates based on their phenotypes as described previously [30,31,32,33,34,35,36,37,38]. G-Biosciences Taq DNA Polymerase, purchased through the VWR biobar (HSC building, University of Calgary), was used in all PCR reactions. For each type of AMR gene, we randomly picked one or two PCR products at desired sizes, purified them, and sent them for sequencing (http://www.ucalgary.ca/dnalab/sequencing) to confirm their sequences. Primers are listed in Table 1.
2.5. Insertion Sequence Detection
PCR was used to determine the presence of insertion sequence IS256 as previously described [39]. Additionally, a new primer set (IS256t_F & IS256t_R) was designed to target the internal region of IS256. PCR assay was carried out with the following thermal cycling profile: initial denaturation at 95°C for 5 min, 30 cycles of amplification (denaturation at 94°C for 1 min, annealing at 55°C for 1 min, and extension at 72°C for 1.30 min), and a final extension at 72°C for 5 min in a Bio-Rad T100TM Thermal Cycler (Mississauga, Ontario L5T 1C9).
2.6. Statistical Analysis of Data
Associations between antibiotics, antibiotic classes, year and species of Enterococcus were examined using two sample tests of proportions, taking into account clustering of isolates within flocks when appropriate. Rho was calculated based on the Intraclass Correlation Coefficient (ICC) computed following a mixed effects unconditional logistic regression of the drug of interest taking clustering within flock into account. Logistic regression using robust variance estimation allowing for clustering of isolates within flocks was used to examine the association of variables such as species and year with the outcomes of interest. Robust variance estimation relaxes the standard errors allowing for independence within flocks. A P value of 0.05 was considered statistically significant in all statistical analyses. All analyses were completed in Stata 15 (StataCorp. 2017. Stata Statistical Software: Release 15. College Station, TX: StataCorp LLC).
3. Results
3.1. Surveillance of AMR in Enterococci Isolates from Broiler Chicken Farms in 2015-2016 (Fig. 1)
Enterococci were present in all fecal samples, with a recovery rate of 100%. There were 34/88 (39) and 50/96 (52%) isolates susceptible to all drugs tested or resistant to up to two drug classes in 2015 and 2016 respectively. In contrast, in 2015 and 2016 there were 54/88 (61%) and 46/96 (48%) isolates MCR (resistant ≥ three drug classes) respectively.
Figure 1.
Comparison of AMR patterns between enterococci in year 2015 and 2016.
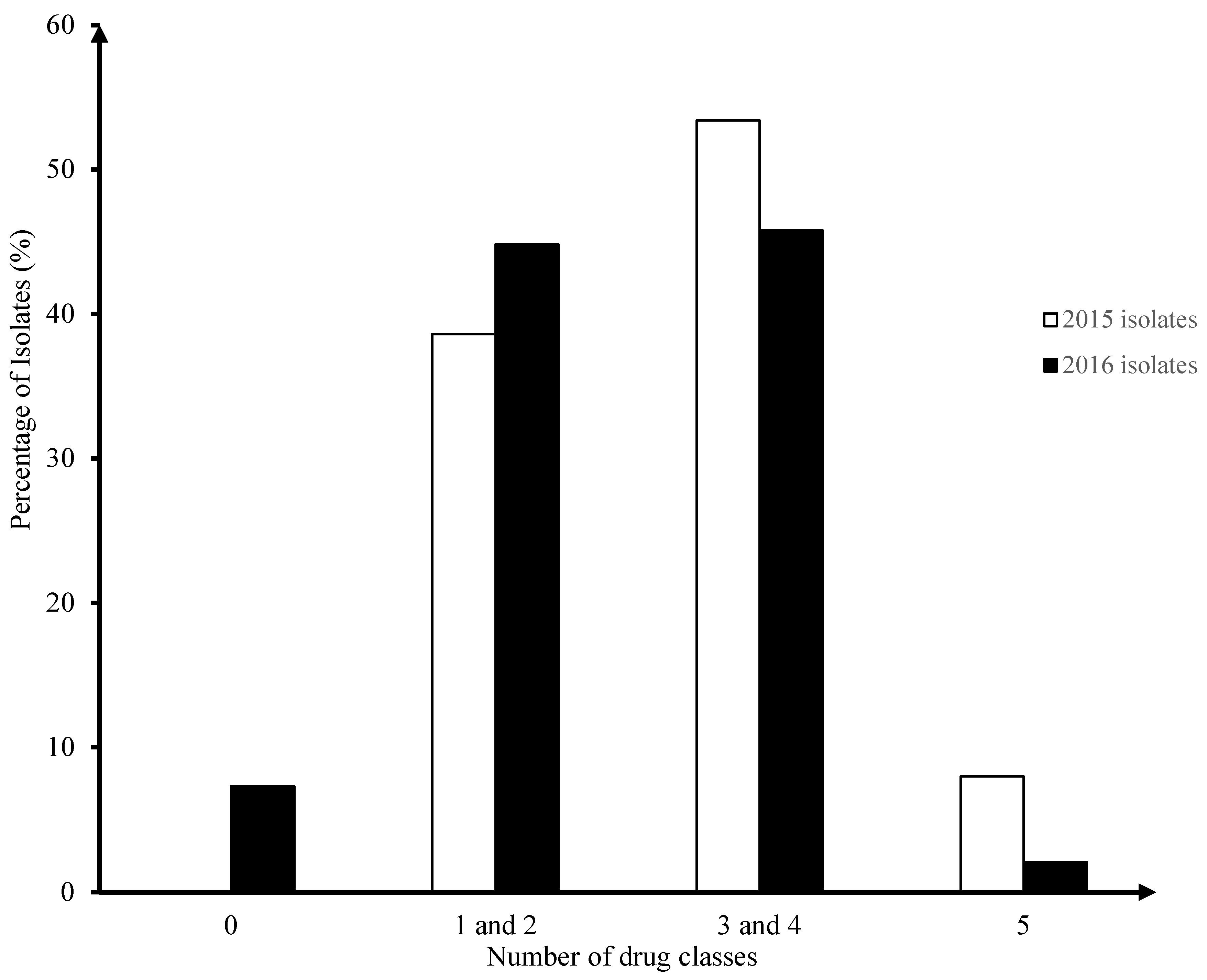
Resistance to 0-2 classes of antibiotics was compared with resistance to three or more classes of antibiotics using logistic regression while controlling for year. There was no significant difference in the number of isolates resistant to three or more classes from 2015 to 2016 (OR = 0.58, P = 0.160, 95% CI 0.27-1.24). All 2015 isolates were resistant to at least one drug class. The proportion of isolates resistant to five drug classes in 2015 (7/88, 8%) compared with 2016 (2/96, 2.1%) was not statistically significantly different (OR = 0.25, P = 0.08, 95%, CI = 0.05 – 1.20).
3.2. Speciation of Enterococcus Isolates from Bbroiler Chicken Farms (Table 2)
The two predominant enterococci species that inhabit the chicken GI tract are Enterococcus faecalis (n = 109) and Enterococcus faecium (n = 72). The percentage of these enterococci species was similar in 2015 (E. faecalis = 52 (48%), E. faecium = 35 (49%)) and 2016 (E. faecalis = 57 (52%), E. faecium = 37 (51%) among our isolates. The percentage of E. faecalis isolates was found to be 20% higher than that of E. faecium isolates in both years’ isolate collections. Other enterococci species identified included Enterococcus durans (one isolate in 2015) and Enterococcus hirae (two isolates in 2016).

Table 2.
Species of enterococci isolated from poultry farms in 2015-2016.
| Enterococcus species | 2015 | 2016 | ||||||
|---|---|---|---|---|---|---|---|---|
| Number of isolates | Percentage | Number of MCR isolates | Percentage | Number of isolates | Percentage | Number of MCR isolates | Percentage | |
| Enterococcus faecalis | 52 | 59% | 37 | 43% | 57 | 59% | 29 | 30% |
| Enterococcus faecium | 35 | 40% | 17 | 20% | 37 | 39% | 16 | 17% |
| Enterococcus durans | 1 | 1% | - | - | - | - | - | - |
| Enterococcus hirae | - | - | - | - | 2 | 2% | 1 | 1% |
| Total | 88 | 100% | 54 | 63% | 96 | 100% | 46 | 48% |
In both years, the percentage of MCR E. faecalis isolates was about as twice as that of MCR E. faecium isolates. E. faecalis decreased from 37/52 (42.5%) in 2015 to 29/57 (31%) in 2016. Meanwhile, a slight decrease in MCR isolates of E. faecium species (3%) was observed. There was no significant change in the odds of an isolate being MCR between E. faecium and E. faecalis when also controlling for year (OR = 0.54, P = 0.112, 95% CI = 0.25-1.15). The only E. durans isolate was identified in 2015 and was not MCR. Among two E. hirae isolated in 2016, one showed resistance to 3 drug classes (aminoglycoside, macrolide and nitrofurantoin).
3.3. Characterization of Overall AMR Profile – Frequency Distribution per Antimicrobial Agents in Year 2015-2016 Categorized by Drug Classes (Fig. 2)
Among tested drug classes, 157/184 (85%), 128/184 (70 %) and 90/184 (49%) of isolates in both years were resistant to streptogramin, macrolide and tetracycline respectively. Resistance to streptogramins was evenly distributed across both years (~85%). Resistance to macrolides declined from 65/88 (74 %) in 2015 to 63/96 (66 %) in 2016, and resistance to tetracyclines declined from 49/88 (56%) in 2015 to 41/96 (43%) in 2016. The reduction in resistance to each of these drugs over the two year period was not significant.
Figure 2.
The distribution of enterococcus isolates’ AMR phenotype by drug classes in year 2015-2016.
Figure 2.
The distribution of enterococcus isolates’ AMR phenotype by drug classes in year 2015-2016.
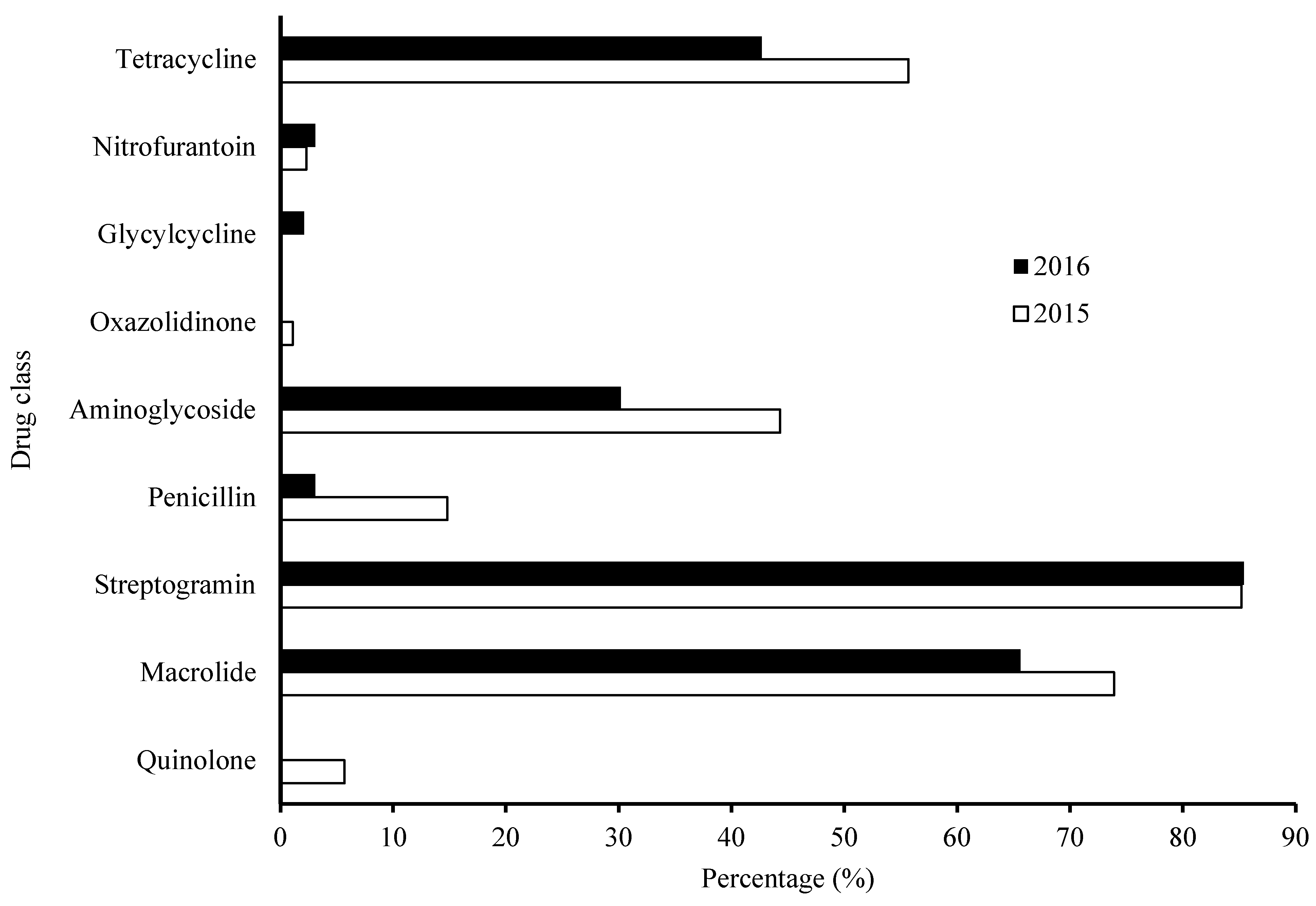
Thirty-seven percent of isolates (n = 68) were resistant to aminoglycosides. There was a declining trend of resistance to aminoglycosides (approximately 14% decrease) between 2015 (n = 39/88, 44%) and 2016 (n = 29/96, 30%). Aminoglycosides were the only drug class that had two antibiotics used in the susceptibility test: gentamicin and streptomycin. A significantly larger proportion of samples were resistant to streptomycin than gentamicin in 2015 and 2016 (Two sample test of proportions, Z = 4.3728, P value = <0.0001 in both years). Most isolates were resistant to either streptomycin or gentamicin; only two isolates in 2015 and one in 2016 showed resistance to both of them.
Only 13/88 (15%) and 3/96 (3%) of isolates were resistant to penicillin (drug: ampicillin) in 2015 and 2016 respectively. The decrease in resistance between the two years was statistically significant (Fisher’s exact P value = 0.007).
Very few enterococci (<10%) showed resistance to nitrofurantoin, glycylcycline, oxazolidinone and quinolone. One isolate in 2015 was resistant to oxazolidinone and five to quinolones, but none were found in 2016. Conversely, only 2 isolates in 2016 were resistant to glycylcycline, while there was none in 2015. In both years, no isolates were resistant to glycopeptide and cyclic lipopeptide. Due to high costs, only MCR isolates were tested for resistance to cyclic lipopeptides.
3.4. Characterization of AMR Profile – Frequency Distribution per Antimicrobial Agents in MCR Community in Year 2015-2016 Categorized by Drug Classes (Fig. 3)
Only MCR isolates (resistant to ≥ 3 drug classes) were selected for further investigation of their AMR profile. Around 93% of MCR isolates were resistant to either streptogramins or macrolides. The percentage of MCR isolates exhibiting resistance to aminoglycosides increased slightly from 57% in 2015 to 59% in 2016. Very few MCR isolates were resistant to nitrofurantoin (n = 4), glycylcycline (n = 1), oxazolidinone (n = 1) and quinolone (n =5).
Figure 3.
The distribution of enterococcus MCR isolates’ AMR phenotype by drug classes in year 2015-2016.
Figure 3.
The distribution of enterococcus MCR isolates’ AMR phenotype by drug classes in year 2015-2016.
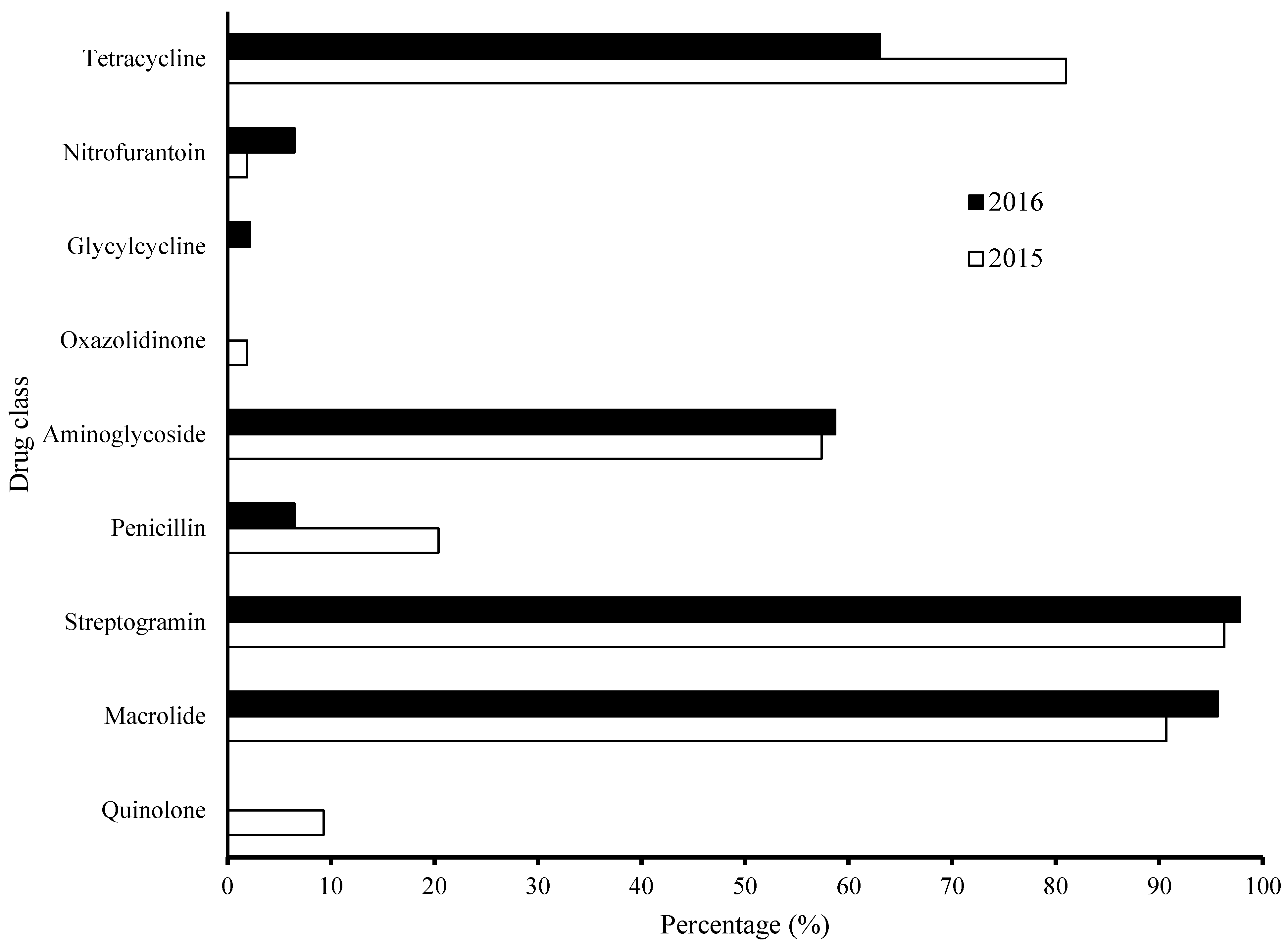
Only 14 isolates were MCR when considering penicillin drug class (11 in 2015, 3 in 2016). The decrease from 2015 to 2016 was not significant.
The percentage of MCR isolates exhibiting resistance to tetracycline decreased from 81.5% in 2015 to 63% in 2016. The odds of an isolate being resistant to tetracycline was 10.3 times higher when the isolate was MCR, while also controlling for year (OR = 10.3, P = <0.0001, 95% CI = 4.4-24.1).
3.5. Characterization of AMR Patterns of Isolates Resistant to a Combination of Three or More Antimicrobials
Resistance to a combination of doxycycline, erythromycin and quinupristin was the most common AMR pattern in both years (n = 14 in 2015, n = 18 in 2016) (Table 3). E. faecalis isolates were predominant in this pattern; there was no E. faecium in 2016 displaying this pattern. The second most common resistance pattern was the above pattern with the addition of streptomycin (Table 3). Twelve isolates exhibited this resistance pattern in 2015 and six in 2016. Similarly, E. faecalis isolates were also predominant in this pattern, and none of E. faecium in 2016 displayed this pattern. Resistance to a combination of streptomycin, erythromycin and quinupristin was also a common resistance pattern with four isolates resistant in 2015 and 11 in 2016. Only E. faecium isolates were found to have this resistance pattern. There were no significant changes in resistance between 2015 and 2016 in any of these resistance patterns.
Interestingly, the other enterococci species in our study displayed different AMR patterns. The E. durans isolate found in 2015 was resistant to both streptomycin and nitrofuratoins, and intermediate to both doxycycline and quinupristin. One out of two E. hirae in 2016 displayed a combined resistance to streptomycin, erythromycin and nitrofurantoins. The other E. hirae isolate was susceptible to all tested antimicrobials.
3.6. Characterization of AMR Patterns of Isolates with Intermediate Resistance in Combinations of Three or More Antimicrobials (Table 4)
When considering intermediate resistance phenotypes, resistance to levofloxacin and vancomycin appeared quite often in a variety of patterns which included intermediate resistance. Levofloxacin (drug class: quinolones) resistance appeared in eight out of 11 different patterns; vancomycin (drug class: glycopeptides) resistance appeared in five out of 11 different patterns. But these patterns occurred only among 2015 isolates.
Table 4.
The number of isolates (intermediate) resistant to a combination of 3 or more antimicrobials.
Table 4.
The number of isolates (intermediate) resistant to a combination of 3 or more antimicrobials.
| a. All AMR patterns of intermediate phenotypes only | |||||
| Pattern | 2015 | 2016 | Total | ||
| E. faecalis | E. faecium | E. faecalis | E. faecium | ||
| dox-ery-lzd | 1 | 0 | 0 | 0 | 1 |
| dox-ery-van | 1 | 0 | 0 | 0 | 1 |
| dox-lvx_qd | 0 | 1 | 0 | 0 | 1 |
| dox-lvx-qd-str | 0 | 1 | 0 | 0 | 1 |
| dox-lvx-str | 0 | 2 | 0 | 0 | 2 |
| ery-lvx-lzd-tgc | 1 | 0 | 0 | 0 | 1 |
| ery-str-van | 2 | 0 | 0 | 0 | 2 |
| lvx-lzd-str-tgc-van | 1 | 0 | 0 | 0 | 1 |
| lvx-lzd-van | 1 | 0 | 0 | 0 | 1 |
| lvx-nit-van | 1 | 0 | 0 | 0 | 1 |
| lvx-str-van | 1 | 0 | 0 | 0 | 1 |
| Total | 9 | 4 | 0 | 0 | 13 |
| b. Common AMR patterns of both intermediate and resistant phenotypes | |||||
| Pattern | 2015 | 2016 | Total | ||
| DOX-ERY-lvx-QD | 3 | 0 | 3 | ||
| DOX-ERY-lvx-QD-STR | 3 | 0 | 3 | ||
| DOX-ery-QD | 0 | 6 | 0 | ||
| ERY-QD-str | 0 | 5 | 5 | ||
| dox-ERY-QD | 0 | 6 | 6 | ||
| dox-ERY-lvx-QD-str | 3 | 0 | 3 | ||
Antimicrobials in uppercase represent resistant phenotypes. Antimicrobials in lowercase represent intermediate phenotypes. Antimicrobial abbreviation: GEN = gentamicin, STR = Streptomycin, AMP = Ampicillin, ERY = erythromycin, LZD = Linezolid, VAN = Vancomycin, QD = Quinupristin, TGC = Tigecycline, DAP = Daptomycin, LVX = Levofloxacin, NIT = Nitrofurantoins, DOX = Doxycycline.
3.7. The Association of Phenotype and Genotype in MCR Isolates 2015-2016
There were five MCR strains in 2015 that showed resistance to levofloxacin (class: quinolone). The emeA gene was detected in all five of these strains, while efrA and efrB genes were present in four of five. We also tested 20 out of 21 MCR strains in 2015 that showed intermediate resistance to levofloxacin. They all carried efflux pump genes; emeA, efrA and efrB. Two strains in 2016 displayed intermediate resistance to levofloxacin, and they both harbored emeA, efrA and efrB genes.
Two genes, ermB and mefA, were screened in MCR strains that showed resistance to erythromycin (class: macrolide). The ermB gene were present in all 49 MCR strains from 2015 and 44 MCR strains from 2016 batch (100%). The mefA gene was not detected in these isolates.
Two genes lsa and mefA were screened in MCR strains that showed resistance to quinupristin (QD). The results showed that 96% (50/52) of the MCR strains in 2015 and 84% (38/45) MCR strains in the 2016 batch harbored lsa gene. Among 50 lsa-detected strains in 2015, 37 and 13 strains were E. faecalis and E. faecium, respectively. The other two strains that did not carry lsa gene were speciated as E. faecalis and E. faecium. Twenty-nine E. faecalis and 16 E. faecium in 2016 carried the lsa gene. The remaining 7 strains from 2016 that did not carry lsa gene were all E. faecium. Four PCR products of lsa gene amplification from both E. faecalis and E. faecium were purified and sent for sequencing. Multiple sequence alignment revealed that they were all distinct in sequence and the single nucleotide polymorphisms (SNPs) varied between isolates (Figure 4). Although the SNPs were all different, the sites of the SNPs were all the same. Also, the lsa-like gene amplified from E. faecium (53.4) shared 99% nucleotide sequence identity with the E. faecalis lsa gene from Genbank database (Access No. AY587982.1). None of these strains had the mefA gene detected. None of MCR E. faecium isolates that were resistant to QD had any vat/vga genes detected.
3.8. Insertion Sequence
IS256 were PCR-amplified in MCR strains from 2015 and 2016. No PCR products were obtained using flanking primers that were designed and used in a previous study [39]. However, we were able to amplify IS256 products when using new primer set which targeted the internal region of IS256. The sequence of PCR product was confirmed by Sanger sequencing. Our sequencing results matched with IS256 sequence (EF_0125) on Tn4001 in E. faecalis (strain ATCC 700802 / V583). IS256 was present in 82.2% of MCR strains in 2015 and 100% of MCR strains in 2016.
4. Discussion
Resistance to individual antimicrobial agents identified in this study were lower than that reported in The Netherlands in 2002 for gentamicin (7% versus 44%), quinupristin (92% versus 88%), vancomycin (0% versus 80%) and erythromycin (70% versus 80%) [40]. Enterococci isolated from poultry litter samples collected from poultry houses in the eastern United States in 2004 reported resistance to erythromycin of 69% in E. faecalis, comparable to 62% in E. faecalis in the current study. In this study however, the prevalence of erythromycin resistance in E. faecium was higher at 82% compared with 34% in the US study. Resistance to ampicillin in both E. faecalis and E. faecium was low in both studies. High levels of resistance to quinupristin were recorded in both studies [41]. However, there are variations in methodology for isolation and antibiotic susceptibility testing of enterococci to consider.
There were no statistically significant differences in the number of MCR strains and general overall AMR profiles between 2015 and 2016. A slight decrease in monensin use as feed additives was also observed over two years (data not shown). With such a small sample size, it is not feasible to draw any conclusions from statistical analysis of these isolates. The association between ionophores used in agricultural and antibiotic resistance in human health is not yet clear [42,43]. Therefore, precautions in using these compounds as feed additives still need to be taken seriously. Future ongoing surveillance will help unravel their effects on antibiotic resistance issues and ultimately on animal/human health.
AMR profiles showed that the top three prevalent distribution frequencies were individually considering resistances towards streptogramin, macrolide and tetracycline. Our results showed some similarities with a previous study where they looked at enterococci from broiler chickens in the Fraser Valley of British Columbia, Canada [44]. However, upon a closer comparison, there were some discrepancies between their study and ours. The percentage of resistant isolates towards streptogramin, macrolide and tetracycline were 85%, ~70%, and ~50%, respectively. Meanwhile, in their study, they were ~75%, ~65%, and 90%, respectively. The percentage of poultry isolates resistant to tetracycline was higher among enterococcus collected in the study conducted in BC (90%) versus those collected in our study conducted in AB (50%).
Enterococcus species in poultry were mainly comprised of two species E. faecalis and E. faecium [45]. These two species are the most prevalent enterococci species in human infection. They are pathogenic due to their AMR, extracellular proteins (toxins), extra chromosome, mobile genetic elements, cell wall components, biofilm formation, adherence factors, and colonization factor [9]. Among the isolates collected in both years of our study the proportion of these two species (3/2) remained unchanged, with a higher proportion of E. faecalis than E. faecium. These results are in agreement with a study of enterococci species on a poultry farm in North Georgia [45]. Additionally, they found that antimicrobial usage on this farm had no effect on enterococci species. In contrast, E. faecium isolates were more common than E. faecalis isolates in other studies [40,44].
Two efflux pumps, EmeA, a NorA homolog of Staphylococcus aureus, and EfrAB, a heterodimeric ABC transporter, were present in most MCR isolates that showed (intermediate) resistance to quinolones (26/28). Quinolone resistance in enterococci can be due to either mutation in genes encoding gyrase and topoisomerase IV, or efflux pump systems that actively pump antimicrobials out of cell [10]. Efflux pump systems are the main mechanism of resistance to different types of antimicrobial agents in many bacteria. The gene emeA has been shown to be responsible for a decreasing susceptibility to several structurally unrelated compounds, including quinolones, in an emeA mutant E. faecalis strain [30,31]. In another study, the emeA gene was detected in 26.3% of E. faecalis and 72.6% of E. faecium isolates [35]. The second efflux pump associated with quinolone resistance in enterococci is EfrAB, an ABC-type multidrug efflux pump. This efflux pump was characterized in drug-hypersensitive E. coli, and showed the ability to excrete many compounds, such as norfloxacin, ciprofloxacin, doxycycline, acriflavine, ,4’,6-diamidino-2-phenylindole, tetraphenylphosphonium chloride, daunorubicin, and doxorubicin [32]. In this study, all but one MCR quinolone-resistant strains carried genes for both efflux pumps. More interestingly, MCR strains that displayed intermediate resistance to quinolone also carried these efflux pump genes. This suggests that these efflux pumps may not be the main determinant of quinolone resistance in our isolates, assuming that these pumps were functional.
Macrolide resistance mechanisms in enterococci include both enzyme activity (ermB gene) and efflux pumps (mefA gene). The erm gene encodes for a 23S rRNA methylase which modifies ribosome structure, rendering the strain resistant to macrolides [46]. The mefA gene product, an efflux pump, is known to pump macrolide out of the cell, but this mechanism confers resistance less efficiently than the mechanism of target modification (ermB gene) [10]. None of the tested strains in our study had the mefA gene detected, while all of them harbored ermB gene. Hence the main macrolide resistance mechanism in this study was very likely due to the ribosome target modification activity of the ermB gene product. The methylation of a specific adenine in the 23S rRNA of the 50S ribosomal subunit results in decreasing binding affinity of the macrolide for the ribosome [34]. A correlation between the presence of ermB gene and macrolide resistance strains was also reported in previous studies [34,46]. The ermB gene was present in 39 of 40 highly erythromycin-resistant enterococcus isolates (MIC >128 g/ml) of different species including E. faecium, E. faecalis, E. durans, E. avium, E. hirae and E. gallinarum [34].
Only E. faecalis exhibits intrinsic resistance to QD while E. faecium is normally QD-susceptible [36]. The lsa gene encoding a putative ABC transporter is located only in the E. faecalis genome, not in the genomes of other enterococci [36]. Further investigations continued to support its role in QD-resistance in enterococci [36]. However, full knowledge of its action mechanism remains undiscovered. In our study, a high portion of MCR strains with resistance to QD carried the lsa gene (96% in 2015 and 84% in 2016). The number of MCR E. faecalis with resistance to QD always exceeded that of MCR E. faecium. Surprisingly, we identified the lsa gene in a majority of the MCR E. faecium resistant to QD among our isolates. This is interesting because, to our knowledge, there has not been any report of an E. faecalis lsa homolog found in E. faecium. Recently the lsa(E) gene located on a novel plasmid-borne multidrug resistance gene cluster was reported in E. faecium [47]. However, this gene does not have much sequence identity in common with the lsa (also known as lsa(A)) gene intrinsically present in E. faecalis [48]..
Acquired resistance mechanisms in QD-resistant E. faecium were known to be involved with vat/vga genes. The vat genes encoding streptogramin acetyltransferase enzymes, such as vatD (satA) and vatE (satG), were proven to be necessary for expression of QD resistance in E. faecium [49]. Other newly identified genes, vgaD and vatG, also conferred resistance to QD in E. faecium [37]. The vga genes, including vgaA, vgaB, and vgaC, encoding ATP-binding ABC transporter, were well-characterized in QD-resistant staphylococci [50,51,52,53]. The vgaD gene, however, was first identified as an ABC transporter in QD-resistant E. faecium [37]. However, none of those genes were detected in our MCR E. faecium with resistance to QD.
Dissemination of AMR genes in enterococci occurs due to the involvement of mobile genetic elements such as transposons, plasmids, and IS elements. IS256 is one of the three most common IS elements in enterococci [54]. The presence of this IS element was shown to be associated with various AMR genes conferring resistance to erythromycin, gentamicin and vancomycin [55,56]. In a previous study, they designed primers complementary to unique DNA sequences flanking IS256 copies (six on the chromosome and four on plasmids) [39]. None of PCR products for IS256 were detected using primers published in this previous study. Nevertheless, IS256 was detected in both E. faecalis and E. faecium isolates using our newly designed primers complementary to a highly conserved internal region of IS256. This suggests that the locations of IS256 in our isolate genomes were different from those in the fully sequenced genome of E. faecalis V583. Future whole genome sequencing work will help us know how many copies there are in our isolate’s genomes and pinpoint their locations. The prevalence of the IS256 in our MCR isolates (82% of MCR isolates in 2015 and 100% of MCR isolates in 2016) might explain the high distribution frequencies of macrolide and aminoglycoside resistance among our MCR isolates.
5. Conclusions
Characterization of various AMR profiles and patterns in enterococci from broiler chickens in Alberta in 2015-2016 provided insights into the diversity and prevalence of AMR across different enterococci species in broiler chickens in Alberta. The study further emphasized the role of broiler chickens as reservoir of resistant Gram positive bacteria and their genetic elements. The lsa gene, which is normally located only in the E. faecalis genome, was also found in QD-resistant E. faecium isolates. IS256 was detected in both E. faecalis and E. faecium isolates, but its location got shifted compared to those reported in a previous study. Future studies are required to elucidate molecular mechanisms of quinolone resistance in our MCR isolates, the location of IS256 as well as the function of the lsa gene in QD-resistant E. faecium isolates of this study. This study could serve as a baseline for future surveillance of AMR in enterococci and to complement current AMR surveillance.
Authors’ contributions
SC, KL, SG, CM conceptualized the research idea and obtained research funding. AAF (PI :SC). SG and AA developed the CIPARS AMU-AMR farm surveillance framework, farm surveillance tools (questionnaire) and protocols, and validated the recovery and AMR datasets. Bacterial isolation and initial antimicrobial susceptibility testing were performed by RC. TT and KL were responsible for experimental design. TT, HG and JW conducted research and laboratory analysis. NC conducted statistical analysis. TT and NC designed and drafted the manuscript. All authors contributed to manuscript development. The authors read and approved the final manuscript.
Funding
This study was funded by Alberta Agriculture and Forestry (AAF) [grant number 2015R025R] with significant in a kind support from PHAC and the AAF, Agri-Food Laboratories.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Availability of data and materials
All data generated or analyzed during this study are included in this published article.
Acknowledgments
We would like to thank the poultry veterinarians and producers who voluntarily participated in the CIPARS farm surveillance program and enabled data and sample collection. We are grateful to the Chicken Farmers of Canada and the Alberta Chicken Producers for their valuable input to the framework development and technical discussions. We sincerely thank Dr. Rahat Zaheer, Agriculture and Agri-Food Canada in Lethbridge, for her technical support. TT also wishes to thank Odd-Gunnar Wikmark and the Norwegian Research Council (MARMIB - project number 315812) for their support in finishing this work.
Competing interests
The authors declare that they have no competing interests.
References
- Chicken Farmers of Canada. 2016.
- Grier, K. The 2015 Economic Impact of the Poultry and Egg Industries in Canada. 2016.
- Hardie, J.M.; Whiley, R.A. Classification and overview of the genera Streptococcus and Enterococcus. Society for Applied Bacteriology symposium series 1997, 26, 1S–11S. [Google Scholar] [CrossRef]
- Ruzauskas, M.; Siugzdiniene, R.; Spakauskas, V.; Povilonis, J.; Seputiene, V.; Suziedeliene, E.; Daugelavicius, R.; Pavilonis, A. Susceptibility of bacteria of the Enterococcus genus isolated on Lithuanian poultry farms. Veterinarni Medicina 2009, 54, 577–582. [Google Scholar] [CrossRef]
- Iversen, A.; Kühn, I.; Franklin, A.; Möllby, R. High Prevalence of Vancomycin-Resistant Enterococci in Swedish Sewage High Prevalence of Vancomycin-Resistant Enterococci in Swedish Sewage 2002, 68, 2838–2842. [CrossRef]
- Byappanahalli, M.N.; Nevers, M.B.; Korajkic, A.; Staley, Z.R.; Harwood, V.J. Enterococci in the Environment. Microbiology and Molecular Biology Reviews 2012, 76, 685–706. [Google Scholar] [CrossRef]
- Lu, H.; Weng, X.; Li, H.; Yin, Y.; Pang, M.; Tang, Y. Enterococcus faecium -Related Outbreak with Molecular Evidence of Transmission from Pigs to Humans 2002, 40, 913–917.
- Freitas, A.R.; Coque, T.M.; Novais, C.; Hammerum, A.M.; Lester, C.H.; Zervos, M.J.; Donabedian, S.; Jensen, L.B.; Francia, M.V.; Baquero, F.; Peixe, L. Human and swine hosts share vancomycin-resistant Enterococcus faecium CC17 and CC5 and Enterococcus faecalis CC2 clonal clusters harboring Tn1546 on indistinguishable plasmids. Journal of Clinical Microbiology 2011, 49, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.P. The pathogenicity of enterococci. Journal of Antimicrobial Chemotherapy 1994, 33, 1083–1089. [Google Scholar] [CrossRef] [PubMed]
- Kristich, C.J.; Rice, L.B. Enterococcal Infection — Treatment and Antibiotic Resistance. 2009:1–62.
- Reik, R.; Tenover, F.C.; Klein, E.; McDonald, L.C. The burden of vancomycin-resistant enterococcal infections in US hospitals, 2003 to 2004. Diagnostic Microbiology and Infectious Disease 2008, 62, 81–85. [Google Scholar] [CrossRef]
- Low, D.E.; Keller, N.; Barth, A.; Jones, R.N. Clinical Prevalence, Antimicrobial Susceptibility, and Geographic Resistance Patterns of Enterococci: Results from the SENTRY Antimicrobial Surveillance Program, 1997-1999. Clinical Infectious Diseases 2001, 32 (Suppl. 2), 2):S133–S145. [Google Scholar] [CrossRef] [PubMed]
- Shivaprasad, H.L. An overview of anatomy, physiology and pathology of urinary system in birds. AAV Proceedings 1998:201–205.
- Jung, A.; Rautenschlein, S. Comprehensive report of an Enterococcus cecorum infection in a broiler flock in Northern Germany. BMC Veterinary Research 2014, 10, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Boerlin, P.; Nicholson, V.; Brash, M.; Slavic, D.; Boyen, F.; Sanei, B.; Butaye, P. Diversity of Enterococcus cecorum from chickens. Veterinary Microbiology 2012, 157, 405–411. [Google Scholar] [CrossRef]
- Hollenbeck, B.L.; Rice, L.B. Intrinsic and acquired resistance mechanisms in enterococcus. Virulence 2012, 3, 421–569. [Google Scholar] [CrossRef]
- Aslam, M.; Diarra, M.S.; Checkley, S.; Bohaychuk, V.; Masson, L. Characterization of antimicrobial resistance and virulence genes in Enterococcus spp. isolated from retail meats in Alberta, Canada. International Journal of Food Microbiology 2012, 156, 222–230. [Google Scholar] [CrossRef]
- Hegstad, K.; Mikalsen, T.; Coque, T.M.; Werner, G.; Sundsfjord, A. Mobile genetic elements and their contribution to the emergence of antimicrobial resistant Enterococcus faecalis and Enterococcus faecium. Clinical Microbiology and Infection 2010, 16, 541–554. [Google Scholar] [CrossRef] [PubMed]
- Dunny, G.M.; Berntsson, R.P.A. Enterococcal sex pheromones: Evolutionary Pathways to Complex, Two-Signal Systems. Journal of bacteriology 2016, 198, 1556–1562. [Google Scholar] [CrossRef] [PubMed]
- Eva, B. On Mobile Genetic Elements in Enterococci ; Adding More Facets to the Complexity. October 2010(July).
- Lett, M. Tn3-1ike elements : molecular structure, evolution. Biochimie 1988, 70, 167–176. [Google Scholar] [CrossRef]
- Rice, L.B.; Carias, L.L.; Marshall, S.H. Tn5384, a composite enterococcal mobile element conferring resistance to erythromycin and gentamicin whose ends are directly repeated copies of IS256. Antimicrobial Agents and Chemotherapy 1995, 39, 1147–1153. [Google Scholar] [CrossRef]
- Hodel-Christian, S.L.; Murray, B.E. Characterization of the gentamicin resistance transposon Tn5281 from Enterococcus faecalis and comparison to staphylococcal transposons Tn4001 and Tn4031. Antimicrobial Agents and Chemotherapy 1991, 35, 1147–1152. [Google Scholar] [CrossRef] [PubMed]
- Bonafede, M.E.; Carias, L.L.; Rice, L.B. Enterococcal transposon Tn5384: Evolution of a composite transposon through cointegration of enterococcal and staphylococcal plasmids. Antimicrobial Agents and Chemotherapy 1997, 41, 1854–1858. [Google Scholar] [CrossRef]
- Public Health Agency of Canada (PHAC): Canadian Antimicrobial Resistance Surveillance System – Report 2016. 2016.
- Clinical and Laboratory Standards Institute: Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Third Informational Supplement. M100-S23 edition. Wayne, PA; 2013.
- Vineetha, N.; Vignesh, R.; Sridhar, D. Preparation, Standardization of Antibiotic Discs and Study of Resistance Pattern for First-Line Antibiotics in Isolates from Clinical Samples. International Journal of Applied Research 2015, 1, 624–631. [Google Scholar]
- Woodford, N.; Egelton, C.M.M.D. Comparison of PCR with Phenotypic Methods for the Speciation of Enterococci. In Streptococci and the Host Advances in Experimental Medicine and Biology. Edited by Horaud T., Bouvet A., Leclercq R., de Montclos H. SM. Boston, MA; Springer; 1997:405–406.
- Zaheer, R.; Yanke, L.J.; Church, D.; Topp, E.; Read, R.R.; McAllister, T.A. High-throughput species identification of enterococci using pyrosequencing. Journal of Microbiological Methods 2012, 89, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Suchiya, T.T. Functional Cloning and Expression of emeA, and Characterization of EmeA, a Multidrug Efflux Pump from Enterococcus faecalis. Biol Pharm Bull 2003, 26, 266–270. [Google Scholar]
- Jonas, B.M.; Murray, B.E.; Weinstock, G.M. Characterization of emeA,a norA Homolog and Multidrug Resistance Efflux Pump, in Enterococcus faecalis. Antimicrobial Agents and Chemotherapy 2001, 45, 3574–3579. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.; Huda, M.N.; Kuroda, T.; Mizushima, T.; Tsuchiya, T. EfrAB, an ABC Multidrug Efflux Pump in Enterococcus faecalis. Antimicrobial Agents and Chemotherapy 2003, 47, 3733–3738. [Google Scholar] [CrossRef] [PubMed]
- Lavilla Lerma, L.; Benomar, N.; Sánchez Valenzuela, A.; Casado Muñoz M del, C.; Gálvez, A.; Abriouel, H. Role of EfrAB efflux pump in biocide tolerance and antibiotic resistance of Enterococcus faecalis and Enterococcus faecium isolated from traditional fermented foods and the effect of EDTA as EfrAB inhibitor. Food Microbiology 2014, 44, 249–257. [Google Scholar] [CrossRef]
- Portillo, A.; Ruiz-Larrea, F.; Zarazaga, M.; Alonso, A.; Martinez, J.L.; Torres, C. Macrolide Resistance Genes in Enterococcus spp. Antimicrobial Agents and Chemotherapy 2000, 44, 967–971. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Li, G.; Wang, W. Prevalence and antimicrobial resistance of Enterococcus species: A hospital-based study in China. International Journal of Environmental Research and Public Health 2014, 11, 3424–3442. [Google Scholar] [CrossRef]
- Singh, K.V.; Weinstock, G.M.; Murray, B.E. An Enterococcus faecalis ABC homologue (Lsa) is required for the resistance of this species to clindamycin and quinupristin-dalfopristin. Antimicrobial Agents and Chemotherapy 2002, 46, 1845–1850. [Google Scholar] [CrossRef]
- Jung, Y.H.; Shin, E.S.; Kim, O.; Yoo, J.S.; Lee, K.M.; Yoo, J.; Chung, G.T.; Lee, Y.S. Characterization of two newly identified genes, vgaD and vatG, conferring resistance to streptogramin A in Enterococcus faecium. Antimicrobial Agents and Chemotherapy 2010, 54, 4744–4749. [Google Scholar] [CrossRef]
- Soltani, M.; Beighton, D.; Philpott-howard, J.; Soltani, M.; Beighton, D.; Philpott-howard, J. Mechanisms of Resistance to Quinupristin-Dalfopristin among Isolates of Enterococcus faecium from Animals, Raw Meat, and Hospital Patients in Western Europe Mechanisms of Resistance to Quinupristin-Dalfopristin among Isolates of Enterococcus faecium fro. Antimicrobial Agents and Chemotherapy 2000, 44, 433–436. [Google Scholar] [CrossRef]
- Manson, J.M.; Hancock, L.E.; Gilmore, M.S. Mechanism of chromosomal transfer of Enterococcus faecalis pathogenicity island, capsule, antimicrobial resistance, and other traits. Proceedings of the National Academy of Sciences 2010, 107, 12269–12274. [Google Scholar] [CrossRef]
- Bogaard, A.E.; Willems, R.; London, N.; Top, J.; Stobberingh, E.E. Antibiotic resistance of faecal enterococci in poultry, poultry farmers and poultry slaughterers. Journal of Antimicrobial Chemotherapy 2002, 49, 497–505. [Google Scholar] [CrossRef]
- Hayes, J.R.; English, L.L.; Carr, L.E.; Wagner, D.D.; Joseph, S.W. Multiple-Antibiotic Resistance of Enterococcus spp. Isolated from Comemercial Poultry Production Environments. Apllied and Environmental Microbiology 2004, 70, 6005–6011. [Google Scholar] [CrossRef]
- Science, E.; Wong, A. Unknown Risk on the Farm : Does Agricultural Use of Ionophores Contribute to the Burden of Antimicrobial. mSphere 2019(September):1–6.
- Russell, J.B.; Houlihan, A.J. Ionophore resistance of ruminal bacteria and its potential impact on. FEMS microbiology reviews 2003, 27, 65–74. [Google Scholar] [CrossRef]
- Diarra, M.S.; Rempel, H.; Champagne, J.; Masson, L.; Pritchard, J.; Topp, E. Distribution of antimicrobial resistance and virulence genes in Enterococcus spp. and characterization of isolates from broiler chickens. Applied and Environmental Microbiology 2010, 76, 8033–8043. [Google Scholar] [CrossRef] [PubMed]
- Debnam, A.L.; Jackson, C.R.; Avellaneda, G.E.; Barrett, J.B.; Hofacre, C.L. Effect of Growth Promotant Usage on Enterococci Species on a Poultry Farm. Avian Diseases 2005, 49, 361–365. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, F.J.; Sadurski, R.; Kray, A.; Boos, M.; Geisel, R.; Koöhrer, K.; Verhoef, J.; Fluit, A.C. Prevalence of macrolide-resistance genes in Staphylococcus aureus and Enterococcus faecium isolates from 24 European university hospitals. Journal of Antimicrobial Chemotherapy 2000, 45, 891–894. [Google Scholar] [CrossRef]
- Si, H.; Zhang, W.J.; Chu, S.; Wang, X.M.; Dai, L.; Hua, X.; Dong, Z.; Schwarz, S.; Liu, S. Novel plasmid-borne multidrug resistance gene cluster including lsa(E) from a Linezolid-Resistant Enterococcus faecium Isolate of Swine Origin. Antimicrobial Agents and Chemotherapy 2015, 59, 7113–7116. [Google Scholar] [CrossRef] [PubMed]
- Wendlandt, S.; Lozano, C.; Kadlec, K.; Gómez-Sanz, E.; Zarazaga, M.; Torres, C.; Schwarz, S. The enterococcal ABC transporter gene lsa(E) confers combined resistance to lincosamides, pleuromutilins and streptogramin A antibiotics in methicillin-susceptible and methicillinresistant Staphylococcus aureus. Journal of Antimicrobial Chemotherapy 2013, 68, 473–475. [Google Scholar] [CrossRef]
- Werner, G.; Klare, I.; Heier, H.; Hinz, K.H.; Böhme, G.; Wendt, M.; Witte, W. Quinupristin/dalfopristin-resistant enterococci of the satA (vatD) and satG (vatE) genotypes from different ecological origins in Germany. Microbial drug resistance 2000, 6, 37–47. [Google Scholar] [CrossRef]
- Strain, S.T.; Kadlec, K.; Schwarz, S. Novel ABC Transporter Gene, vga(C), Located on a Multiresistance Plasmid from a Porcine Methicillin-Resistant Staphylococcus aureus. AntimicrobAgents Chemother 2009, 53, 3589–3591. [Google Scholar]
- Allignet, J.; Solh, E. Characterization of a new staphylococcal gene, vgaB, encoding a putative ABC transporter conferring resistance to streptogramin A and related compounds 1997, 202, 133–138. [CrossRef]
- Science, E.; All, P.B.V.; Cedex, P. Sequence of a staphylococcal plasmid gene, vga, encoding a putative ATP-binding protein involved in resistance to virginiamyein A-like antibiotics. Gene 1992, 117, 45–51. [Google Scholar]
- Haroche, J.; Allignet, J.; Buchrieser, C.; Pathoge, M. Characterization of a Variant of vga(A) Conferring Resistance to Streptogramin A and Related Compounds. AntimicrobAgents Chemother 2000, 44, 2271–2275. [Google Scholar] [CrossRef] [PubMed]
- Schramm, V.L.; Shi, W.; Opin, C.; Biol, S.; Allard, J.; Grochulski, P.; Sygusch, J.; Acad, P.N.; Lett, O. Role of Mobile DNA in the Evolution of Vancomycin-Resistant Enterococcus faecalis. Science 2003, 299, 2071–2075. [Google Scholar]
- Quintiliani, R.; Courvalin, P. Characterization of Tn1547, a composite transposon flanked by the IS16 and IS256-like elements, that confers vancomycin resistance in Enterococcus faecalis BM4281. Gene 1996, 172, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Rice, L.B.; Thorisdottir, A.S. The prevalence of sequences homologous to IS256 in clinical enterococcal isolates. Plasmid 1994, 32, 344–349. [Google Scholar] [CrossRef]
Figure 4.
Multiple sequence alignment of lsa gene amplification from four isolates against lsa gene from Genbank database (Access No. AY587982.1) using online tool (http://multalin.toulouse.inra.fr/multalin/). Two of them (92.4 and 100.4) were E. faecalis isolates from 2016 batch, and the other two (37.4 and 53.4) were E. faealis and E. faecium, respectively, from 2015 batch.
Figure 4.
Multiple sequence alignment of lsa gene amplification from four isolates against lsa gene from Genbank database (Access No. AY587982.1) using online tool (http://multalin.toulouse.inra.fr/multalin/). Two of them (92.4 and 100.4) were E. faecalis isolates from 2016 batch, and the other two (37.4 and 53.4) were E. faealis and E. faecium, respectively, from 2015 batch.
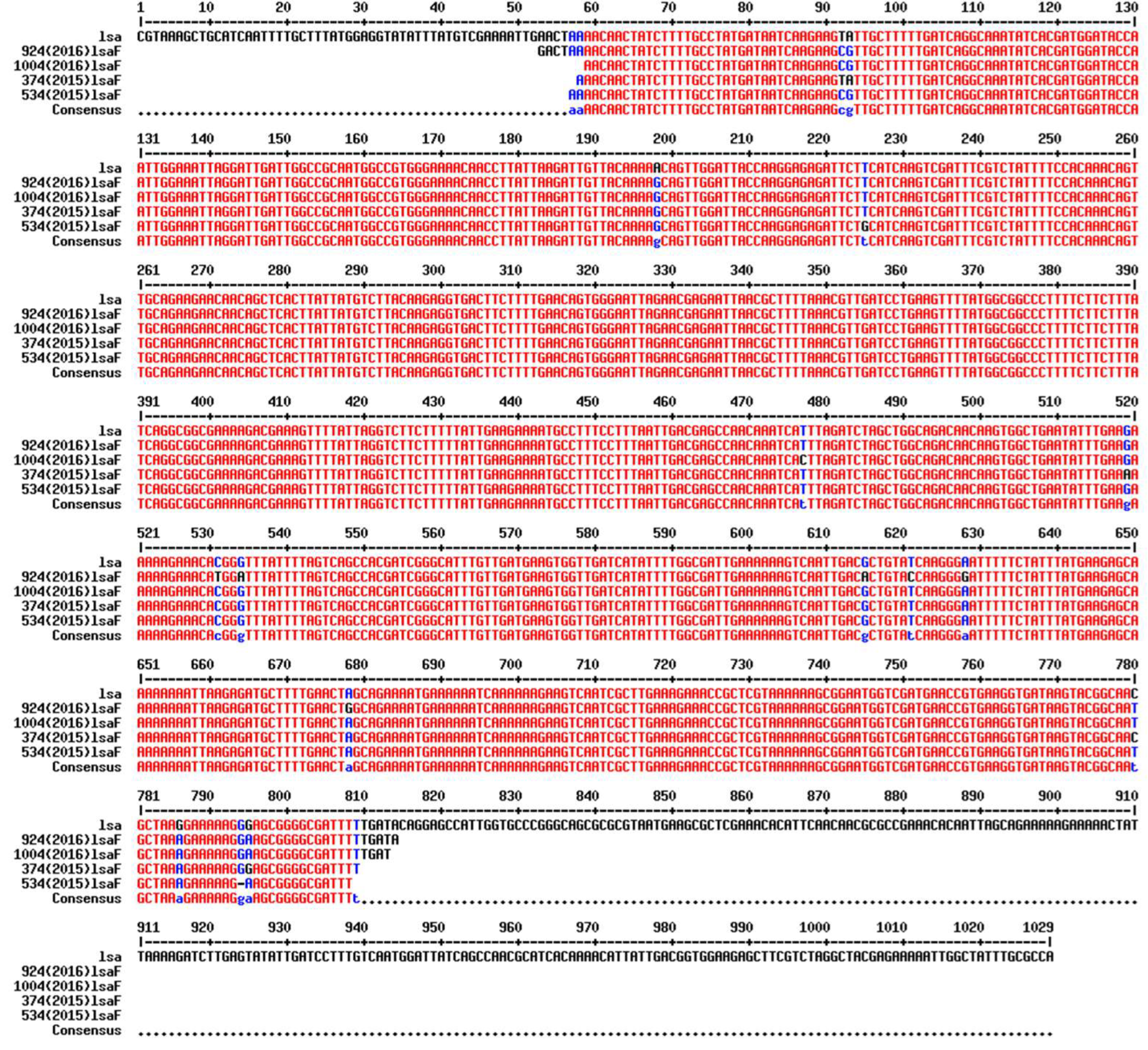
Table 1.
List of primers used in this study.
| Primers | PCR Product | Size (bp) | Reference |
|---|---|---|---|
| ddl1_F 5’-ATCAAGTACAGTTAGTCTT-3’ ddl1_R 5’-ACGATTCAAAGCTAACTG-3’ |
ddlE.faecalis FOd |
941 |
[28] |
| ddl2_F 5’-GCAAGGCTTCTTAGAGA-3’ ddl2_R 5’-CATCGTGTAAGCTAACTTC-3’ vanC1_F 5’- GGTATCAAGGAAACCTC-3’ vanC1_R 5’- CTTCCGCCATCATAGCT-3’ vanC2-F 5’-CTCCTACGATTCTCTTG-3’ vanC2-R 5’-CGAGCAAGACCTTTAAG-3’ |
ddlE.faecium vanC1E.gallinarum vanC2 E.casseliflavus & E.flavescens |
550 822 439 |
[28] [28] [28] |
| Ent-ES-211-233-F 5’- GHACAGAAGTRAAATAYGAAGG-3’ Ent-EL-74-95-R 5’- GGNCCTAABGTHACTTTNACTG-3’ emeA_F 5’-AGTATGATGTACTTAGCAATTTC-3’ emeA_R 5’-CATCTTATTTCGATTTAAAAATAAC-3’ efrA_F 5’-TTGGCTTTATGACGCCAGTG-3’ efrA_R 5’-CGTGCGATAGCTAAACGTTG-3’ efrB_F 5’-CCTTATTTAACTGGATTACCAAC-3’ efrB_R 5’-GAATAGTTGATAGGCGGTGG-3’ ermB_F 5’-ATTCTCAAAACTTTTTAACGAGTG -3’ ermB_R 5’- CCTCCCGTTAAATAATAGATAAC-3’ lsa_F 5’-CGTAAAGCTGCATCAATTTTGC -3’ lsa_R 5’- AATGGCTCCTGTATCAAAAATC -3’ mefA_F 5’- GGCAAGCAGTATCATTAATCAC -3’ mefA_R 5’- CATTATTGCACAGCAAACTACG -3’ vatG_F 5’-GTGGGAAAAGCATACACCT-3’ vatG_R 5’-TTGCAGGATTACCACCAAC-3’ vgaD_F 5’-CAACTGGAGCGAGCTGTTA-3’ vgaD_R 5’-GACAGCCGGATAATCTTTTG-3’ vatD_F 5’-GCTCAATAGGACCAGGTGTA-3’ vatD_R 5’-TCCAGCTAACATGTATGGCG-3’ vatE_F 5’-ACTATACCTGACGCAAATGC-3’ vatE_R 5’-GGTTCAAATCTTGGTCCG-3’ IS256c-F 5’-CATTGGTAAATTGGAATGGAAATC-3’ IS256c-R 5’-ATTCAAACATTTTTTCCTCTCC-3’ IS256d_F 5’-GATCAACTGGAGAATTAGTGTT-3’ IS256d-R 5’-CTCTAATATCCCCTAATGAAAATAATG-3’ |
groES-EL spacer region emeA efrA efrB ermB lsa mefA vatG vgaD vatD/ satA vatE/ satG Primers flanking ef0125 Primers flanking ef0529 |
variable (~200bp) 1137 1048 1513 713 825 911 200 201 272 512 ~1173 ~1173 |
[29] [30,31] [32,33] [32,33] [34,35] [36] [34,35] [37] [37] [38] [38] [39] [39] |
| IS256e_F 5’-GGCTATTTTTTAGCAAACTATGTAT-3’ IS256e_R 5’-CACAGCAACTATTGGTAACG-3’ |
Primers flanking ef2187 | ~1173 | [39] |
| IS256f_F 5’-TGTCTAGCTAAAACGAAGCC-3’ IS256f-R 5’-GACCCAACAAAAGTAACTCG-3’ |
Primers flanking ef2632 | ~1173 | [39] |
| IS256g-F 5’-CTGTTTTGTCTCGTCATTATATGA-3’ |
Primers flanking ef3100 | ~1173 | [39] |
| IS256g-R 5’-GGTTATAGTAGGAATAATTTTGCC-3’ IS256h-F 5’-CTGAACTGACACAATTCATTAAAT-3’ |
Primers flanking ef3215 |
~1173 |
[39] |
| IS256h-R 5’-AATTTAGCAACATCTTTCATTGG-3’ |
|||
| IS256t_F 5’-CTGAAAAGCGAAGAGATTCAAAGC-3’ |
IS256 transposase | 748 | This study |
| IS256t_R 5’-GAACTTGGCATCTTTGCCAACTTAC-3’ |
Table 3.
AMR patterns of isolates resistant to a combination of three or more antimicrobials.
| Pattern | 2015 | 2016 | Total | ||
|---|---|---|---|---|---|
| E. faecalis | E. faecium | E. faecalis | E. faecium | ||
| AMP-DOX-ERY | 0 | 1 | 0 | 0 | 1 |
| AMP-DOX-ERY-GEN-QD | 1 | 0 | 0 | 0 | 1 |
| AMP-DOX-ERY-STR-QD | 1 | 1 | 0 | 1 | 3 |
| AMP-DOX-LVX-NIT-QD | 1 | 0 | 0 | 0 | 1 |
| AMP-DOX- LVX-QD | 1 | 0 | 0 | 0 | 1 |
| AMP-ERY-QD | 3 | 2 | 0 | 0 | 5 |
| AMP-ERY- STR-QD | 0 | 0 | 0 | 2 | 2 |
| DOX-ERY-GEN-QD | 3 | 0 | 1 | 0 | 4 |
| DOX-ERY-GEN-STR-QD | 2 | 0 | 1 | 0 | 3 |
| DOX-ERY-LVX-STR-QD | 2 | 0 | 0 | 0 | 2 |
| DOX-ERY-LZD-STR-QD | 1 | 0 | 0 | 0 | 1 |
| DOX-ERY-QD | 9 | 5 | 18 | 0 | 32 |
| DOX-ERY-STR | 0 | 1 | 0 | 0 | 1 |
| DOX-ERY-STR-QD | 11 | 1 | 6 | 0 | 18 |
| DOX-GEN-QD | 2 | 0 | 1 | 0 | 3 |
| DOX-STR-QD | 0 | 1 | 1 | 0 | 2 |
| ERY-LVX-QD | 0 | 1 | 0 | 0 | 1 |
| ERY-NIT-QD | 0 | 0 | 0 | 1 | 1 |
| ERY-STR-QD | 0 | 4 | 0 | 11 | 15 |
| ERY-NIT-QD-STR-TGC | 0 | 0 | 0 | 1 | 1 |
| GEN-ERY-QD | 0 | 0 | 1 | 0 | 1 |
| Total | 37 | 17 | 29 | 16 | 99 |
Antimicrobial abbreviation: GEN = gentamicin, STR = Streptomycin, AMP = Ampicillin, ERY = erythromycin, LZD = Linezolid, VAN = Vancomycin, QD = Quinupristin, TGC = Tigecycline, DAP = Daptomycin, LVX = Levofloxacin, NIT = Nitrofurantoins, DOX = Doxycycline.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



