
Submitted:
28 November 2024
Posted:
29 November 2024
You are already at the latest version
Abstract
In natural environment plants are simultaneously exposed to multivariable abiotic and biotic stresses. Typical abiotic stresses are changes in temperature, light intensity and quality, water stress (drought, flood), microelements availability, salinity, air pollutants and others. Biotic stresses are caused by other organisms such as pathogenic bacteria and viruses, or by parasitic insects and plants. This review presents the state of the art of the programmed cell death in the cross-tolerance phenomena and its conditional molecular and physiological regulators, which are involved in simultaneous regulation of plant acclimation, defense and developmental responses. It highlights the role of the absorbed energy in excess and its dissipation as heat in induction of the chloroplast-retrograde electrical, phytohormonal, reactive oxygen species signaling and heat shock related pathways in connection to activation of the molecular regulators of plant growth, yield and cell wall development. It also discusses how systemic and network-acquired acclimation and acquired systemic resistance are mutually regulated and demonstrates the role of non-photochemical quenching and the dissipation of absorbed energy in excess as heat in the cross-tolerance phenomenon. Finally new evidences that in plants evolved one molecular system to regulate cell death, acclimatization and cross-tolerance to abiotic and biotic stress are presented and discussed.
Keywords:
absorbed energy in excess and cellular light memory
; electrical
; reactive oxygen species (ROS) and phytohormones signaling
; hypersensitive disease defense response and systemic acquired resistance
; network and systemic acquired acclimation
; non-photochemical quenching and cross-tolerance
; transcription factors
1. Introduction
One of the greatest challenges of the twenty-first century is feeding a growing world population while dealing with an overheated planet and progressive desertification of fertile land. Climate extremes are becoming more common, disrupting agricultural production and posing continuous dangers of starvation [1,2]. It was estimated that more than 800 million people worldwide did not have access to enough food in 2021 [1,3]. It is anticipated that global warming would increase the frequency of extreme weather, endangering agricultural crop production [4]. According to calculations that desert area will increase from 3 to 10% of the total land area [5]. Crucial agricultural crops were anticipated to experience an overall yield loss of about 70% due to adverse conditions in the fields and from environment, which means only 30% of the yield was produced when compared to its yield from genetic potentials [6,7,8,9,10]. Growing demand for crop production and yield maintaining in adverse global warming conditions necessitates vastly expanded new plant stress resistance research initiatives.
The frequency and severity of several environmental stresses, such absorbed energy in excess (AEE), ultraviolet (UV) radiation, severe droughts, heat waves, and restrictions on water accessibility for plants, crops, and global ecosystems, are increasing due to global warming. When taken as a whole, these stressors cause significant losses in agricultural productivity. Stress response phytohormones and signaling molecules, including ethylene (ET), salicylic acid (SA), abscisic acid (ABA), jasmonic acid (JA), reactive oxygen species (ROS) have been widely described in terms of regulation of plant stress responses and productivity. During heat and osmotic stress response these factors, together with electrical signaling regulate a mechanism known as programmed cell death (PCD), an eventual completion of the cell cycle. Induced PCD in some cells is required for effective stress response, leading to acclimatization or induction of disease resistance, thus optimizing plant survival and production under stress condition [11,12,13,14,15,16,17,18,19,20]. Systemic and network acquired acclimation (SAA and NAA) chloroplast retrograde signaling within and between plants is important for induction of PCD [11,12,21,22,23]. It is important to know that stress response to episode of AEE stress applied to low light acclimated plants is able to induce disease resistance to the virulent bacterial pathogen and better tolerance to UV-C irradiation episode due to induction of the cellular light memory [22,24]. This memory involves molecular, physiological, biochemical and biophisical changes that are lasting for several days after AEE stress and is specific for quantity and quality of AEE. Therefore, the current knowledge and understandings of the cross-tolerance phenomena, plant stress responses (SAA, SAR, NAA) and PCD signaling mechanisms is much better than it was a decade before. The roles of conditional molecular and transcriptional regulators of PCD and stress responses such as: LESION SIMULATING DISEASE 1 (LSD1), ENHANCED DISEASE SUSCEPTIBILITY 1 (EDS1), and CYSTEINE-RICH RLK (RECEPTOR-LIKE PROTEIN KINASE) 5 (CRK5), METACASPASES (MC4 and MC8), SIGNAL RECOGINTION PARTICLES (cpSRP43 and 54), 22 kDa photosystem II protein (PsbS), Ca2+ and other ion channels, transcription factors (WRKY, DREB - CBF2 subfamily A-1 of nvironmentRF/AP2) ROS scavenging/generating enzymes and others involved in regulation of NAAs, SAAs, SAR, cellular light memory, cross-tolerance and PCD induction and/or inhibition are better, although not fully understood [11,12,13,14,15,16,20,21,22,23,24]. This review concentrate on the current understanding of plant stress responses, cross-tolerance to both biotic and abiotic, and on molecular and physiological mechanisms that allow plants to conditionally optimize their growth, development, and seed yield in multi-stress environment.
2. Abiotic and Biotic Stresses
In the classical view environmental stress factors can be classified into two major categories: abiotic and biotic stresses. Plants are constantly revealed to a mixture of both biotic and abiotic stresses, which are significantly impacting their growth, development, and overall fitness [6,12,25,26,27,28]. Abiotic stresses include factors such as: changes in temperature extremes (low or high), light intensity and quality (low light, high light, red/blue/UV ratio), water and osmotic stress, heavy metal toxicity (i.e. Hg, Cd, Pb), and nutrient deficiency (organic/inorganic). In the natural environment abiotic stresses are usually clustered, for example, shift from low to high light intensity is associated with sudden increase of UV radiation and immediate foliar temperature increase due to absorbed energy in excess and its immediate dissipation as heat via non-photochemical quenching (NPQ) mechanism [29]. This usually can change plant physiology and biochemistry (metabolism), leading to altered retrograde signaling (ROS, phytohormonal and electrical) [21,22,24] and changes in gene expression. As a result of NPQ induction plants increase foliar heat production from photosystems and inhibit stomatal conductance, leading to impaired foliar cooling (transpiration), decreased CO2 assimilation, water and nutrient uptake and induced photorespiration [17,19,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47]. Mittler and colleagues (2022) discussed the influence of various abiotic stress factors on plant signaling networks, highlighting the role of ROS and stress-responsive genes in stress perception and adaptation [20].
Abiotic stresses, such as high light, heat and drought, often occur in combination and are difficult to separate from one another. In fact AEE alone is able to induce heat shock response due to higher dissipation of AEE as heat [29] and Arabidopsis thaliana subjected to a combination of high light and heat stress exhibit a unique metabolic response, including increased accumulation of sugars and amino acids, as well as decreased levels of metabolites participating in the tricarboxylic acid (TCA) cycle [48]. The combined stress in Arabidopsis also leads to the accumulation of non-proteinogenic amino acid γ-aminobutyric acid (GABA), which contributes significantly to plants' adaptation to intense light and heat stress, possibly via encouraging autophagy [49]. In Arabidopsis, the combination of heat and drought stress led to changes in plant metabolomics and root bacterial microbiota, indicating an orchestrated modulation of the whole holobiont [50]. Wheat crops also experience combined heat and drought stress, which has more detrimental effects on growth and yield compared to individual stresses. Wheat has developed advanced responses at various levels to tolerate these combined stressors [51]. In Arabidopsis roots, due to drought and heat stress cell wall suberization rise and this led to alterations to the biosynthesis and assembly of major cell wall components [52].
Biotic stresses, arising from interactions with living organisms, pose significant threats to plant health and agricultural productivity [53,54]. These stressors include pathogens, pests, herbivores and others, which can cause diseases, nutrient deficiency, physical damage, and induce defense responses in plants [55,56,57,58,59,60,61,62]. Plants have evolved intricate mechanisms to detect and respond to pathogens - a two-branched innate immune system. It means firstly recognition and responsiveness to the molecules of microbs, including to non-pathogenic ones. Secondly to respond to the pathogen virulence factors, by directly or through their effects on host targets [63,64,65,66,67]. Plants do not have mobile “killer” cells (eg. T4 cells) and a somatic adaptive immune system like in animals. Instead, they rely on the innate immunity of each cell, mediated by gene pairs (R genes in plants and avr genes in pathogens) that induce programmed cell death in infected and noninfected cells, known as the hypersensitive disease defense response (HR) and systemic acquired resistance [63,64,65,66,67].
An oxidative burst, manifested by a rapid ROS generation, is typically present in conjunction with R gene-mediated resistance and is necessary for HR, a sort of PCD that is believed to restrict pathogen's access to nutrients and water. The stimulation of SA-dependent signaling pathway resulting in the development of particular pathogenesis-related (PR) proteins is also linked to R gene-mediated resistance. Some defensive mechanisms in plants are regulated by systems that rely on ET and/or JA. Different classes of R proteins, such as those with nucleotide-binding (NB) domains and leucine-rich repeats (LRRs), play crucial roles in basal defense regulation and resistance to various pathogens, showcasing the diversity of plant immune responses [64,68,69,70].
Plant pathogens are frequently classified as either necrotrophs or biotrophs based on their life-cycle and how they propagate. While necrotrophs lives on death host tissues, biotrophs needs alive host tissue to propagate [71]. It’s interesting to see how SA signaling and R gene-mediated resistance may lead to resistance to avirulent biotrophs [72]. Such pathogens would have no feeding supply as a result of the HR response due to PCD induction. But in the case of necrotrophs, it appears that the pathogen would simply have an easier time existing in the host due to cell death [68]. Studies screening Arabidopsis mutants for deficiencies in resistance to different diseases using defective signaling pathways important in in defense gave some evidence to it, like, resistance to the necrotrophic fungus Alternaria brassicicola is unaffected by the mutation npr1 and the transgene NahG, which disrupt SA signaling, however, they cause resistance to the biotrophic oomycete Peronospora parasitica to be abolished. On the other hand, resistance to the necrotrophic fungus A. brassicicola is significantly compromised by the coi1 mutation, which disrupts JA signaling, whereas resistance to P. parasitica is unaffected. These findings raised the possibility that plant defensive mechanisms may be modified depending on the pathogen invading, with SA-dependent defenses targeting biotrophs and JA- and ET-dependent responses targeting necrotrophs [68,73,74,75]
3. Cross-Tolerance
When plants are exposed to one kind of stress, it can trigger shared signals and pathways, which increases their ability to withstand other kinds of stress. The process called as cross-tolerance and is associated with increased adaptive fitness [17,21,26,76,77,78]. It can be accomplished through a co-activation of the plant's innate immune system, which involves a network of non-specific stress-responsive pathways that bridge biotic-abiotic stress borders. Heat, chilling, drought and salt stress are frequent abiotic stressors with cross tolerance effects [79,80]. Additionally, different stress signaling networks interact with one another, which might cause plants to develop cross-tolerance. For instance, Epichloë endophyte-infected grasses exhibit greater resistance to abiotic stressors like to drought through increased root biomass, high stomatal conductance with rich accumulation of solutes for osmotic regulation, to cold by Upregulation of unsaturated fatty acids, to salinity by keeping anatomical changes (like increase xylem, phloem and vascular bundles size) and to pathogen infections by increasing antioxidants and phenolic compounds that are known to be important for plant defenses [81,82]. According to Capiati and colleagues (2006) showed that wounding boosts tomato plants' resistance to salt, through a mechanism that involves the systemins and JA. They showed that calmodulin-like activities are necessary for the downstream signaling events that lead to cross-tolerance between wounding and salt stress [83]. Lima and colleagues (2018) evaluated cross-tolerance induced by heat stress and water stress in common beans and observed improved germination under osmotic stress following heat stress [84]. These findings suggest that agricultural plants may be engineered or bred to be resistant to many stresses either abiotic or biotic [15,26,42].
Cross tolerance involves chloroplast retrograde and nuclear anterograde signaling, which are involved in intra- and inter-cellular communication between chloroplasts and the nucleus in response to stress. This signaling pathways are mediated by redox signal, dependent on ROS, electrical and phytohormonal signaling which regulates in concerted maner expression of the chloroplast- and nuclear-encoded genes required for appropriate stress responses [11,19,20,21,22,23,24,26,42,64,77,78,85,86]. The WHIRLY family of proteins and the REDOX-RESPONSIVE TRANSCRIPTION FACTOR 1 (RRTF1) have been identified as potential mediators of chloroplast-to-nucleus retrograde signaling, leading to cross tolerance [11,19,20,21,22,23,24,26,42,64,77,78,85,86]. Hydrogen peroxide (H2O2), a byproduct of various aerobic pathways, has also been implicated in retrograde signaling and the induction of cross tolerance [86]. The chloroplast acts as an environmental sensor and communicates with other cell compartments through retrograde signaling to regulate nuclear gene expression in response to developmental cues and stresses by Ca2+ and ROS [11,19,20,21,22,23,24,26,85,88].
The mechanism by which plant chloroplasts, and more especially the LHC and oxygen-evolving reaction centers of PSII in chloroplast membranes, sense stress stimuli is the quickest process that happens faster than pico- or nano-seconds (e.g., AEE dissipation as heat and fluorescence, singlet oxygen production, absorbed energy transfer, electron transfer in photosystem II) (Figure 1). Redox reactions in photosystems, such as electrical charge separation, pH gradients imbalances across thylakoid membranes, stimulation of the xanthophyll cycle through NPQ, redox responses involving the photosynthetic electron carriers eg. Plastoquinone pool (PQ) (linear electron transport), and formation of ROS which can be in pico-, nano- to milli-seconds, can be triggered by AEE that rely on the state and size of the antenna in PSII [77,88]. On an interval of microseconds to seconds, ROS deposits can impacts hormones, sugars, ion signaling pathways, and the redox state of the PQ and glutathione/ascorbate pools (Figure 1). Certain signals, such as CRK5 and
MITOGEN-ACTIVATED PROTEIN KINASE 4 (MPK4), regulate stomatal closing within seconds, leading to enhanced photorespiration, triggering ROS generation, and ET synthesis [41,89] (Figure 1). In the following minutes to hours or days, LSD1 regulates genetically depending stress memory, acclimation and defense reactions and PCD by limiting ROS and ET production in conjunction with ROS scavenging enzymes (SODs, CA’s PXs) through controlled EDS1/PAD4/ETHYLENE INSENSITIVE2 (EIN2) - dependent signal transduction which can happen in seconds to minutes [11,17,23,89]. Consequently, these triggers physiologically controlled redox reactions in stromal metabolism and stress memory to occur in time-related (minutes, hours, days) manner in the environment of various cells, tissues, organs, and even various plants thriving in the vicinity (through SAA, SAR, NAA). According to recent studies, plants subjected to AEE may experience shifts in their local and systemic net photosynthesis quantity, as well as photo-electrochemical-redox retrograde signaling for acclimation priming and AEE light stress memory. These responses are essential for conditionally optimal plants' photosynthesis, transpiration, light acclimation, defense actions, and ultimately plant health (Figure 1 and Figure 2) [15,16,17,22,23,24,89,90]. SAA, NAA, and stress memory serve as a training process of "naive" not acclimated plant cells, chloroplasts, and PSII. Peak and colleagues (2004) [91] suggested that foliar stomata conductance and chlorophyll fluorescence are regulated by the cellular automaton algorithm that relies on collective dynamics and emergent, distributed biological computation in the leaves of plants that on the contrary regulates chlorophyll a fluorescence and stomata conductance. Later on we suggested that ES-, ROS- and NPQ-wave-like-changes propagated in SAA and NAA could be a process regulated in the identical manner like Peak and colleagues (2004) suggested [22,23,24,89] (Figure 1 and Figure 2). Plant cells have the capacity to selectively memorize AEE episodes and the spectral quality of light (either physiologically or molecularly) [22,23,24]. Overall, the interplay between chloroplast retrograde NPQ- and PQ-dependent signaling and other cellular signaling pathways is crucial for the plant's ability to acclimate and defense thus for cross-tolerance.
Peak and colleagues (2004) [91] presented confirmation for complex, joint dynamics and emergent, distributed computation in plans regulating foliar fluorescence and stomatal conductance. In short, stomatal conductance, under some condition, become synchronized and foliar regions with lower chlorophyll a fluorescence have higher while regions with higher fluorescence have lower conductance. This regulation can be describe by mathematical cellular automata algorithm [91]. Consequently, conditional optimization of transpiration and CO2 assimilation is possible. Transmission of electrical, Ca2+, phytohormonal, and ROS signals between local and systemic tissues of the same plant is necessary for wound signaling and induction of SAA [85]. Recently our group discovered that the PsbS protein (22 kDa protein of photosystem II) and NPQ are the hardware of the cellular automata algorithms known as cellular light (quanta) memory. Light harvesting antennas (LHC) versus photosystem reaction centers (P680 or P700) undergo supramolecular modifications that affect the fate of AEE between fluorescence, photochemistry (electrical charge separation), and heat channels. This memory is dependent on the communication network of all active photosystems II within a plant and between plants [23,24,88,92]. This communication network is controlled and regulated by wavy-like, discrete and spatially in time electrical- and ROS-signaling and NPQ changes, that depends on PsbS protein level, [22,23,24,88,92] proton motive force and chlorophyll a fluorescence decay time changes [88]. This hardware controls the fate of absorbed energy in photosystems thus control chloroplast retrograde signaling across the whole plant and between plants. As a result, it discreetly and spatially controls stomatal conductance values over time, causing further NPQ wave-like adjustments thus regulate AEE dissipation as heat [24,88,92,93].
In Górecka and colleagues (2020) we showed that prior induction of the cellular light memory in response to AEE episode developed cross-tolerance to subsequent UV-C episode in PsbS-dependent manner [24]. PsbS is an essential protein in plants that regulate NPQ thus regulate balance between absorbed energy dissipated as heat and that actually required for photochemistry [24,93,94,95]. In this study npq4-1 mutant lacking functional PsbS protein and overexpressing PsbS gene in transgenic Arabidopsis line (oePsbS), responded differently to episode of AEE and in induction of the cellular light memory and subsequent induction of cross-tolerance to UV-C. Untreated oePsbS plants were more resilient than wild type and npq4-1 plants. Resilience was measured by cellular ion leakage due to foliar PCD induction after UV-C episode. After AEE episode wild type and oePsbS but not npq4-1 mutant plants were able to induce light memory and subsequent cross-tolerance to UV-C. The study's findings revealed a new, significant function for PsbS and NPQ as a regulators of chloroplast retrograde signaling for PCD, light acclimation, light (quanta) memory, disease defense and UV cross-tolerance [24].
SAA is an essential light acclimatory molecular and physiological process that is depending on chloroplast retrograde signaling and on whole plant regulation of NPQ [22,23,24,88,92,96]. Ciszak and colleagues (2015) [88] studied the time-resolved fluorescence of chlorophyll a in Arabidopsis thaliana npq4-1 mutant (without functional PsbS protein thus deregulated in NPQ) in leaves exposed to excess light and in leaves undergoing SAA in ambient light condition. This experiment was done since the chlorophyll a fluorescence decay (FD) time regulations in leaves directly exposed to excess light episode and in leaves undergoing SAA were unknown. These experiments demonstrated that leaves undergoing SAA in ambient low light and leaves directly exposed to AEE showed regulation of the FD. Wild type Arabidopsis leaves exposed to AEE had significantly shorter FD time than control leaves and leaves undergoing SAA in ambient low light. However, SAA leaves show smaller but significant decrease in FD time in comparison to control low light leaves. On the other hand, leaves treated with electron transport inhibitor 3-(3,4-dichlorophenyl)-1,1-dimethylurea (DCMU) that significantly reduced NPQ and cause photoinhibition of electrical charge separation, and leaves of the npq4–1 mutant from ambient low light had circa 2-times longer FD time. Leaves of npq4-1 mutant plants exposed to AEE and undergoing SAA in low light had significantly longer FD times. Subsequently npq4-1 leaves showed deregulated (abnormal) FD times up to 2 h after AEE episode, These findings suggest that SAA signaling regulates the quantum-molecular properties of supramolecular complex of photosystem II, and that PsbS-dependent light memory processing is necessary for the regulation of SAA [88]. It is important to point out that synthetic (transgenic) improvement of NPQ and its relaxation in tobacco and soybean lead to improvement of biomass and seed yield productivity in field conditions [94,95].
Electrical signaling shows a crucial part in development of cross-tolerance. It was reported that when a single dandelion leaf is subjected to punctual wounding or punctual high light stress, the plant generates surface foliar electrical signal that then spread to the nearby in-contact plants in the vicinity [23]. This can cause systemic NPQ and physiological alterations in both plants [23,92]. This electrical signaling can be transmitted from plant to plant in a network of plants connected successively by physical touch. Also it is happening regardless of nearby plant belongs to a different species, these signals can still cause reactions in the recipient plant. This new physiological phenomenon was named as network-acquired acclimation (NAA) [23]. These findings strongly imply that the transmission of direct surface electrical signals between the receiver plant (leaves) and the stressed host plant (or one leaf) results in discrete and spatial changes in the absorbed energy fate (NPQ) as well as the induction of common retrograde signaling for defense responses and acclimatization in the host plant and the entire plant community (Figure 2) [23].
4. Signaling Networks During Biotic and Abiotic Stresses
Abiotic factors have been proven to be more detrimental to crop yield when they occur simultaneously [26,97,98]. It is well established that factors, such as drought, heat waves and higher salinity have an impact on the development and spread of diseases and insects [97,99,100,101,102,103,104]. Through modifications to plant physiology and defense mechanisms, these stress conditions also have a direct impact on plant-pest interactions [100]. Many weeds use water more effectively than crops and in that case abiotic stress like drought, can favor these interactions [97,102,105]. The cumulative impact of stressors on crops is not necessarily positive since the final result is often determined by the way these stress variables interplay [106,107,108,109,110].
Cross-talk between abiotic and biotic stresses is occurring on physiological and molecular levels. Reduced photosynthesis, decreased water use efficiency (WUE), and changes in stomatal opening are affected by biotic stresses, but are essential for a plant's ability to withstand an abiotic stress [111,112,113]. Therefore It was revealed that Na+ and Cl- levels in Phaseolus vulgaris shoots increased in salinity after being exposed to a root pathogen Macrophomina phaseolina [111,114]. ABA signaling, which coordinates plant adaptation to abiotic stress, can be diminished by SA signaling, which is generated following invasion with Pseudomonas syringae pv. tomato [115]. ABA synthesis and signaling is depending on zeaxantin a direct precursor of ABA synthesis triggered by NPQ changes in the chloroplast. HopAM1, a type III effector of Pseudomonas syringae that targets HSP70 is involved in stress-induced closure of stomata in ABA-dependent manner [116,117]. In Arabidopsis the HopAM1 overexpressed lines enhance the sensitivity towards ABA for stomatal closure and germination arrest [118]. Tolerance to abiotic stress such as drought and freezing can potentially result from plant-microbe interactions, like exposing plants to a viral strain which includes Tobacco rattle virusI (TRV), Tobacco mosaic virus (TMV), Cucumber mosaic virus (CMV), Brome mosaic virus (BMV). Infection of these strains is udeful against drought stress. On the other hand, CMV strain works in freezing stress in Beet due elevated levels of antioxidant and osmoprotectant [111,119,120]. By interacting with hormonal processes, maintaining water and source-sink relationships, and increasing plant vigor under stress, fungi such as fungal endophytes and non-pathogenic rhizobacteria and mycorrhizal fungi can benefit plants [111,121,122,123].
Research of the impacts of external use of compounds that activate plant defense mechanism known as priming, provide more evidence for the overlap between abiotic and biotic stress signaling pathways [124]. In Arabidopsis increased resilience to heat, drought, and salt stresses, as well as increased resistance to both biotrophic and necrotrophic fungi were found when β-aminobutyric acid (β-ABA), SA, and jasmonates were applied [111,125]. H2O2 and NO priming for salt tolerance moderately increased the abundance of oxidized and S-nitrosylated proteins, which then remained relatively similar after the application of stress. Non-treated plants were more sensitive and exhibited increased protein carbonylation and oxidation, and higher antioxidant enzymes activities [126,127]. In cauliflower seedlings the enrichment of H2O2 and superoxide anion is due to pre-treatment of H2O2, this in turns enhance the MDA content. This pre-treatment of H2O2 produce changes in antioxidant systems, which includes enzymatic (SOD, CAT, GPX and APX) and non-enzymatic systems (AsA, GSH and proline) [128]. In durum wheat seeds primed with H2O2, induced the resistance against salinity stress by increasing enzymatic and non-enzymatic antioxidant defense systems [129]. Recently, epigenetic modifications and specifically chromatin-regulated gene activation have been proposed to govern priming responses [130].
4.1. Transcription Factors Role During Abiotic and Biotic Stress
Because of their functions as key regulators of various stress-related genes, transcription factors (TFs), such as WRKY, NAC, ERF, LSD1, DREB, CBF2, EDS1 and PAD4 are intriguing prospects for genetic modification [11,130]. The molecular function of LESION SIMULATING DISEASE 1 (LSD1) is currently unknown, but the mutant lsd1 phenotype is described as runaway cell death (RCD). RCD in LSD1 mutants is usually elicited due to AEE, root hypoxia, disturbed stomatal conductance [11,17,132], cold [133], drought [11,17], UV radiation [39,134] and pathogen infections [64]. Due to its involvement in different signaling pathways and in regulation of plant CD due to different kind of stresses (both abiotic and biotic), LSD1 is an important TF in cross-tolerance [11]. Importantly, the RCD of lsd1 is closely involved ENHANCED DISEASE SUSCEPTIBILITY 1 (EDS1) and PHYTOALEXIN DEFICIENT 4 (PAD4). These two TFs have a specific domain called triacylglycerol lipase and this domains are important mediation of different genes and disease resistance [135,136]. Both are important in RCD, as in eds1/lsd1 and pad4/lsd1 mutants RCD was not found even by applying different stresses to the plants [21,39,64,77,132]. Ablation of LSD1 and EDS1/PAD4 behave differently toward different stresses and in result the enrichments on ROS and SA is also different [11,17,39,133]. Therefore, LSD1 is the gative regulator of EDS1- and PAD4-dependent cellular pathways that lead to CD. Alongside EDS1 and PAD4, LSD1 plays a significant role in both biotic and abiotic stress responses and contributes to the conditional control of different processes that will have an affect plant, this includes: photosynthesis, maximal photochemical efficiency of PSII, cellular ROS/hormonal homeostasis, seed yield, water use efficiency, cell wall lignin content and cellulose fibers polymerization degree (in hybrid Aspen transgenic trees with silenced LSD1 and EDS1 cultivated 4 years in the field conditions) [11,15,17,19,39,137,138,139]. It is proved that LSD1 is a redox-sensing and transcription regulator which is involve in diverse cellular signaling pathways like xylem tissue differentiation (wood formation) [138]. LSD1 also have a big impact on cell proliferation and modification processes, especially in non-oxidative stress conditions which at the helps the plants in growth and development, but if the conditions are oxidative then LSD1responce to it by regulating different stress mechanisms like, proteolysis, methylation and energy processes, which will ultimately reduce the efficiency of the pathways important for plants development which can at the end leads to CD which means LSD1 is important to keep balancing the cell division and CD depends on conditions and redox status [11,15,17,64,137]. Silenced PAD4 or EDS1 genes in transgenic hybrid Aspen trees displayed increased growth (wood production) 6-8% and altered xylem cells development in comparison to control trees in the field conditions [140,141]. In addition, silenced EDS1 trees displayed higher photosynthesis and transpiration, and higher branching in comparison to control plants [141].
WRKY, NAC, ERF, have been related to improved tolerance in agricultural crops and model plants, since they initiate defense-responsive gene expression [142]. Around 1922 TFs are been shown to be involved in various activities which belongs to 30 TF families in Arabidopsis thaliana [143]. Sorghum has reported 2448 TFs in total, together with 1611 in rice and 3337 in maize [144]. In fact most of these the genes are consider to be stress-responsive and control a large sum of downstream genes, that’s why enhancing the plant towards stress tolerance by altering the TF genes expression is a popular approach in current biotechnology [145]. Here we mentioned some previous studies related to TFs, previous results have shown that MPK3/6 phosphorylated the transcription factors WRKY33 and ERF6 to promote the synthesis of camalexin and activate genes linked to defense to mediate resistance to Botrytis cinerea [146,147]. Another example was reported in Lin and colleagues (2022) study which showed that a negative regulator of lignin biosynthesis called MYB4 was found to be regulated by MAPK-mediated phosphorylation in its modulation of vascular-lignification-caused immunity [148]. GhMAP3K15-GhMKK4-GhMPK6 a drought stress-activated MAPK cascade phosphorylated in cotton, can then activate GhWRKY59 to control the plant's response to the drought stress [149]. In Zhao et al., study it’s found that the ZmWRKY104 phosphorylation by ZmMPK6is important for its role in ABA-induced antioxidant defense and drought tolerance in maize. When H2O2 levels rise, the TF ANAC017, which is located in the ER, releases its N-terminus into the nucleus, which in turn controls primary responses [56]. It has recently been shown that ANAC017 also contributes to aluminum tolerance by controlling genes that changes the cell wall synthesis [150]. Along with EDS1 and WRKY33, which react to H2O2 quickly and transiently, a TF belonging to the ERF family, RRTF1 functions as a key redox signaling network [151,152]. RRTF1 also studied in its role in JA and auxin biosynthesis crosstalk, PCD, salt-tolerance and pathogen resistance [153,154,155,156].
Generally speaking, hypoxia lowers stomatal conductance and CO2 assimilation, which causes photorespiration, which is a major factor in the induction of CD. Recent studies revealed that PsbS and plant β carbonic anhydrases (βCAs), except their obvious functions, are also involved in regulation of plant stress responses [157]. One of our recent studies by Białas and colleagues (2024) suggested that simultaneous overexpression of βCA1 and/or βCA2 with PsbS genes leads to improved photoprotection, acclimation to variable light conditions, and WUE. Due to their extremely high amounts of βCA1 and/or βCA2 transcripts, double and triple overexpression lines react better to hypoxic stress than the other genotypes. Similarly, elevated mRNA levels of βCA1 and/or βCA2, in conjunction with bicarbonate treatment, activated the DREB-CBF2 subfamily A-1 of ERF/AP2, which was suppressed in oePsbS plants, suggesting that the cellular concentrations of βCAs and oePsbS are essential for the precise modulation of stress responses. This suggests that many plant species rely on cellular CBF subfamily genes to respond to abiotic stressors. The analysis of Arabidopsis mutants showed that in response to low-temperature stress, CBF genes control 134 genes [158]. Six genes implicated in freezing tolerance were identified by transcriptome analysis as being controlled by DREB and ERF/AP2 this includes (CBF1, ERF105, ZAT6, ZAT12, WRKY33 and WRKY40). These findings point to the presence of a novel signaling route and chloroplast retrograde regulatory hotspot that are reliant on bicarbonate absorption, βCAs, and PsbS protein relative levels [157]. Taking together these diverse studies show the importance of TFs when it comes to the cross-tolerance and shows a high potential for the future research in the field.
4.2. Reactive Oxygen Species Role During Abiotic and Biotic Stress Condition
Higher plants have developed specialized mechanisms to defend themselves from ROS toxicity and to use ROS as signaling molecules. The chloroplast, mitochondria, peroxisomes, and apoplast are the principal ROS-initiating sites under abiotic stress [19,92,159,160,161,162,163]. If left uncontrolled, ROS contents would rise in cells and result in oxidative damage [20]. Antioxidant enzymes, such as superoxide dismutase (SOD), ascorbate peroxidase (APX), catalase (CAT), glutathione peroxidase (GPX), and peroxiredoxin (PRX) represent a major ROS-scavenging force and are crucial for stress tolerance in plants [164].
Environmental factors, such as high light, UV irradiation, drought, cold, high temperatures, root hypoxia can cause a disruption in cellular redox homeostasis, which can result in cell death and limit the amount of produced biomass [15,19]. It is also widely known that ROS play a role in the plant's response to a pathogen invasion. HR, which is triggered by ROS burst generated by NADPH oxidases or apoplastic enzymes, can cause PCD in different cells at the infection sites and because of their biotrophic nature of infections (feed on live cells), this HR mechanism can limit their invasion, while the surrounding cells develop the capacity to avoid cell death via the propagation of ROS [66]. Callose deposition and cell wall cross-linking, mediated by apoplastic ROS, strengthen the cell wall and prevent pathogens from penetrating it. [165]. Plant NADPH oxidases produce ROS in the apoplast, but their accumulation has also been shown in mitochondria, chloroplasts, and even nuclei [166]. Additionally, many abiotic stressors can directly or indirectly activate additional signaling pathways to cause ROS generation in the plasma membrane by Rboh-signaling pathway, in chloroplasts by Calvin-Benson pathway, in mitochondria through ubiquinone pathway and in peroxisomes by β-oxidation pathway [167]. The cross-tolerance is achieved through the cross-talk between ROS signaling mechanisms and other pathways in activating defense responses [168]. Plants respond to abiotic stress by increasing the accumulation of antioxidant defense systems, including secondary metabolites, such as flavonoids, which scavenge excess ROS and help balance cellular redox state [169]. The interplay between ROS, redox signals, and antioxidative pathways is important for plant acclimation to stress and acquisition of cross-tolerance [170]. Additionally, ROS are involved in organelle-to-organelle and cell-to-cell signaling [171]. Crosstalk between ROS and other signaling molecules, such as reactive nitrogen species (RNS), protein kinases, phytohormones, and secondary messengers like calcium, is important for plant responses to environmental stresses [172] (Figure 2).
4.3. Hormonal Response During Abiotic and Biotic Stress
In the control of plant immunological responses, ET, JA, and SA are thought to be the key molecules. According to several studies [173,174,175,176], the JA signaling pathway always functions as a significant stress hormone route that frequently communicates with various plant hormones to create an extensive signaling network. Recently, a growing number of common elements among JA and several other plant hormone signaling pathways been discovered. JA and SA act antagonistically towards each other but they often combine to make plants more resilient to stressors [177]. Recently, processes underlying JA-SA crosstalk have been widely studied and demonstrated that a variety of genes function in SA-JA antagonism, including MYC2, PDF 1.2 (plant defensin 1.2), TGAs (TF family) [178,179], MAPK (mitogen-activated protein kinase), NPR1, ERF1, WRKY62, WRKY70, GRX480 (glutaredoxin 480), ORA59 (octadecanoid-responsive Arabidopsis AP2/ERF 59), and JAZs [180]. The discovery of the ortholog NPR1 in the progenitor of all terrestrial plants raises the possibility that JA-SA crosstalk occurs in nearly every plant [181].
According to studies by Khan et al., and Peng et al., SA acts to reduce the amount of ET produced as a result of various abiotic factors. For instance, exogenous SA reduces heat sensitivity in heat-stressed plants by decreasing the ACS activity in wheat and promoting proline metabolism while limiting ET production [1,182]. On the contrary, during ozone (O3) stress, ET and SA work together to control cell death in Arabidopsis and tobacco leaves [183,184,185]. By increasing the transcription of the genes encoding CHORISMATE MUTASE and PAL, O3-induced ethylene enhances SA biosynthesis [185]. While ET oversupply worsens cell death, inhibition of either SA or ethylene biosynthesis in Arabidopsis reverses the O3-induced hypersensitive response phenotype [184]. It is widely known that endogenous SA levels and plant defense mechanisms against biotrophic and hemibiotrophic diseases are positively correlated in plants, as discussed by Glazebrook and colleagues (2005). Additionally, the external SA application causes multiple plant species, such as Fusarium oxysporum, Alternaria alternata, Magnaporthe grisea, Colletotrichum gloeosporides, Xanthomonas spp, to develop local and systemic acquired resistance against a variety of pathogens [68,186,187,188,189,190,191,192,193,194,195,196].
JA and ET play an important, synergistic role in plant resistance towards necrotrophic diseases [197,198]. The synergies among ET-stabilized EIN3/EIL1 and JA-activated MYC2 control plant growth and insect resistance. Because MYC2 inhibits EIN3/EIL1, the transcription of ERF1 (downstream gene of EIN3/EIL1) is suppressed, reducing plant tolerance to necrotrophic fungi [199]. VSP2 and CYP79B3 (a wound and herbivore responsive genes respectively) is suppressed via JA signaling pathway when EIN3/EIL1 and MYC2 interact, however, on the other side, weakening MYC2's repression of MYC2 protects towards broad range of herbivores [200,201]. ET and JA can operate oppositely to control a number of abiotic stress genes, besides their combined effects on the functions of EIN3/EIL1 which also includes their targets in rice and Arabidopsis [202]. The transcription level of EIN3/EIL1-inducible target genes is decreased in etiolated Arabidopsis which is because of the already JA-activated MYC TFs includes (MYC2, MYC3, and MYC4) may have been actively interacted with EIN3/EIL1 and thus have blocked their ability to bind a DNA target [183,199,202]. The transcription factor gene ERF1 is a well-known target of MYC2 and also EIN3 which modulates responses to various abiotic stressors [203,204,205]. According to Cheng et al., and Lorenzo et al., Arabidopsis ERF1 regulates the synthesis of many sets of genes that are triggered by JA, drought, salinity, and high temperature. Actually, it has been demonstrated that plants overexpressing ERF1 are more resilient to salt, heat, and drought stress [203,204]. Also Chen and colleagues in (2021) discussed that JA-triggered reduction in EIN3 activity has shown to occur at the level of post-transcriptional stage at the creation of the Arabidopsis apical hook in addition to this phase of transcriptional regulation [183]. These findings imply that JA and ET can control plant immunity in the opposite manner and their role in defense responses (Figure 2).
5. Conclusions
In general, biotic and abiotic environmental stresses induce foliar stomata closure and inhibition of photosynthesis thus induce AEE and its dissipation as heat in NPQ mechanism. AEE alone is able to simultaneously induce five different types of abiotic stresses: heat shock stress, photooxidative stress, photoinhibitory stress, photorespiratory stress and osmotic stress. This in turn activate chloroplast retrograde signaling pathways that are depending on NPQ-, ROS-, heat-, osmotic-, electrical potential- and redox-changes of the photosynthetic electron carriers, like PQ-pool, and on ROS production and scavenging activity. Chloroplast and cellular redox potential is regulated by glutathione and ascorbate cycles and by other chloroplast redox-potential regulators, for example, ferredoxin-NADP+ reductase, thioredoxin and glutaredoxin. These redox changes induce synthesis of direct and indirect precursors of phytohormones like: ABA, SA, ET, JA, IAA and other signaling molecules in the chloroplasts of local and directly stressed cells, and in not directly stressed cells and chloroplasts of systemic cells, tissues and organs by self-propagating waves of electrical- and ROS- heat- and phytohormonal-signaling of SAA and SAR. These signaling waves activate or deactivate various phosphatases, kinases cascades and transcription factors like, for example, LSD1, EDS1, PAD4, WYRKY, NAC, ERF and others. As a result of these some foliar cells induce PCD due to stomata closure and induction of photorespiratory burst of ROS and in other cells induce acclimation and defense responses. Algorithmic changes in ES-, NPQ- ROS- heat- and phytohormones-waves amplitude and frequency regulate stomatal conductance in the cellular automaton manner. All these changes within one plant can be communicated to the other neighboring plants by NAA mechanisms that propagate SAA and presumably SAR within plant community. Therefore, cross-tolerance between abiotic and biotic stress responses is possible not only within one stressed plant but also within plant community, and it is a scientific fact.
Given a few leaves per plant and hundreds of plants in contact in a meadow, thousands of cells per leaf, several dozen chloroplasts per cell, and thousands of PSII per chloroplasts, which are potentially involved in the network of connections, this reveals the complexity of the SAA and NAA signaling and communication network mechanisms that involves trillions of possible communications routes in several m2 of a meadow. Plants are environmentally smart and intelligent, they communicate in sophisticated ways, globally (within community of plants) regulate absorbed energy fate, regulate foliar temperature, are able to count absorbed photons, have quantum-molecular (NPQ-value) memory of AEE episodes and physiological (redox) cellular memory, they process this memory differentially at the same time in various foliar cells and tissues in order to acclimatize and immunize. In other words, it can be said that plants have developed the characteristics of smart, intelligent and communicating organisms. Is this a result of blind and random evolution or intelligent design?
Author Contributions
Conceptualization, S.K.; resources, M.K.; writing—original draft preparation, M.K.; writing—review and editing, P.B. and S.K.; visualization, P.B.; supervision, S.K.; project administration, S.K.; funding acquisition, S.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Polish National Science Centre (Opus 20 UMO-2020/39/B/NZ3/02103) project to SK.
Institutional Review Board Statement
Not applicable
Informed Consent Statement
Not applicable
Data Availability Statement
Not applicable
Conflicts of Interest
The authors declare no conflict of interest.
References
- Peng, P.; Li, R.; Chen, Z.-H.; Wang, Y. Stomata at the Crossroad of Molecular Interaction between Biotic and Abiotic Stress Responses in Plants. Front. Plant Sci. 2022, 13, 1031891. [Google Scholar] [CrossRef] [PubMed]
- Hatfield, J.L.; Prueger, J.H. Temperature Extremes: Effect on Plant Growth and Development. Weather Clim. Extrem. 2015, 10, 4–10. [Google Scholar] [CrossRef]
- World Health Organization The State of Food Security and Nutrition in the World 2022: Repurposing Food and Agricultural Policies to Make Healthy Diets More Affordable; Food & Agriculture Org., 2022; Vol. 2022; ISBN 92-5-136499-0. [CrossRef]
- Fedoroff, N.V.; Battisti, D.S.; Beachy, R.N.; Cooper, P.J.; Fischhoff, D.A.; Hodges, C.; Knauf, V.C.; Lobell, D.; Mazur, B.J.; Molden, D. Radically Rethinking Agriculture for the 21st Century. science 2010, 327, 833–834. [Google Scholar] [CrossRef]
- Wilby, R.; Whitehead, P.; Wade, A.; Butterfield, D.; Davis, R.; Watts, G. Integrated Modelling of Climate Change Impacts on Water Resources and Quality in a Lowland Catchment: River Kennet, UK. J. Hydrol. 2006, 330, 204–220. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, Y.; Zhu, J.-K. Thriving under Stress: How Plants Balance Growth and the Stress Response. Dev. Cell 2020, 55, 529–543. [Google Scholar] [CrossRef] [PubMed]
- Boyer, J.S. Plant Productivity and Environment. Science 1982, 218, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Shewry, P.R. Biochemistry & Molecular Biology of Plants. BB Buchanan, W. Gruissem and RL Jones (Eds), 2000. 2001. [CrossRef]
- Vij, S.; Tyagi, A.K. Emerging Trends in the Functional Genomics of the Abiotic Stress Response in Crop Plants. Plant Biotechnol. J. 2007, 5, 361–380. [Google Scholar] [CrossRef] [PubMed]
- Zurbriggen, M.D.; Hajirezaei, M.-R.; Carrillo, N. Engineering the Future. Development of Transgenic Plants with Enhanced Tolerance to Adverse Environments. Biotechnol. Genet. Eng. Rev. 2010, 27, 33–56. [Google Scholar] [CrossRef]
- Karpiński, S.; SZECHYŃSKA-HEBDA, M.; Wituszyńska, W.; Burdiak, P. Light Acclimation, Retrograde Signalling, Cell Death and Immune Defences in Plants. Plant Cell Environ. 2013, 36, 736–744. [Google Scholar] [CrossRef]
- Zhang, H.; Zhu, J.; Gong, Z.; Zhu, J.-K. Abiotic Stress Responses in Plants. Nat. Rev. Genet. 2022, 23, 104–119. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S. Crop Production under Drought and Heat Stress: Plant Responses and Management Options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [PubMed]
- Minhas, P.S.; Rane, J.; Pasala, R.K. Abiotic Stresses in Agriculture: An Overview. Abiotic Stress Manag. Resilient Agric. 2017, 3–8. [Google Scholar] [CrossRef]
- Bernacki, M.J.; Czarnocka, W.; Szechyńska-Hebda, M.; Mittler, R.; Karpiński, S. Biotechnological Potential of LSD1, EDS1, and PAD4 in the Improvement of Crops and Industrial Plants. Plants 2019, 8, 290. [Google Scholar] [CrossRef] [PubMed]
- Gawroński, P.; Burdiak, P.; Scharff, L.B.; Mielecki, J.; Górecka, M.; Zaborowska, M.; Leister, D.; Waszczak, C.; Karpiński, S. CIA2 and CIA2-LIKE Are Required for Optimal Photosynthesis and Stress Responses in Arabidopsis thaliana. Plant J. 2021, 105, 619–638. [Google Scholar] [CrossRef]
- Wituszyńska, W.; Ślesak, I.; Vanderauwera, S.; Szechyńska-Hebda, M.; Kornaś, A.; Van Der Kelen, K.; Mühlenbock, P.; Karpińska, B.; Maćkowski, S.; Van Breusegem, F.; Karpiński, S. Lesion Simulating Disease1, Enhanced Disease Susceptibility1, and Phytoalexin Deficient4 Conditionally Regulate Cellular Signaling Homeostasis, Photosynthesis, Water Use Efficiency, and Seed Yield in Arabidopsis. Plant Physiol. 2013, 161, 1795–1805. [Google Scholar] [CrossRef] [PubMed]
- Petrov, V.; Hille, J.; Mueller-Roeber, B.; Gechev, T.S. ROS-Mediated Abiotic Stress-Induced Programmed Cell Death in Plants. Front. Plant Sci. 2015, 6, 69. [Google Scholar] [CrossRef] [PubMed]
- Czarnocka, W.; Karpiński, S. Friend or Foe? Reactive Oxygen Species Production, Scavenging and Signaling in Plant Response to Environmental Stresses. Free Radic. Biol. Med. 2018, 122, 4–20. [Google Scholar] [CrossRef]
- Mittler, R.; Zandalinas, S.I.; Fichman, Y.; Van Breusegem, F. Reactive Oxygen Species Signalling in Plant Stress Responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Muhlenbock, P.; Szechynska-Hebda, M.; Płaszczyca, M.; Baudo, M.; Mateo, A.; Mullineaux, P.M.; Parker, J.E.; Karpinska, B.; Karpiński, S. Chloroplast Signaling and LESION SIMULATING DISEASE1 Regulate Crosstalk between Light Acclimation and Immunity in Arabidopsis. Plant Cell 2008, 20, 2339–2356. [Google Scholar] [CrossRef] [PubMed]
- Szechyńska-Hebda, M.; Kruk, J.; Górecka, M.; Karpińska, B.; Karpiński, S. Evidence for Light Wavelength-Specific Photoelectrophysiological Signaling and Memory of Excess Light Episodes in Arabidopsis. Plant Cell 2010, 22, 2201–2218. [Google Scholar] [CrossRef]
- Szechyńska-Hebda, M.; Lewandowska, M.; Witoń, D.; Fichman, Y.; Mittler, R.; Karpiński, S.M. Aboveground Plant-to-Plant Electrical Signaling Mediates Network Acquired Acclimation. Plant Cell 2022, 34, 3047–3065. [Google Scholar] [CrossRef]
- Górecka, M.; Lewandowska, M.; Dąbrowska-Bronk, J.; Białasek, M.; Barczak-Brzyżek, A.; Kulasek, M.; Mielecki, J.; Kozłowska-Makulska, A.; Gawroński, P.; Karpiński, S. Photosystem II 22kDa Protein Level-a Prerequisite for Excess Light-inducible Memory, Cross-tolerance to UV-C and Regulation of Electrical Signalling. Plant Cell Environ. 2020, 43, 649–661. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Fujita, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. ABA-Mediated Transcriptional Regulation in Response to Osmotic Stress in Plants. J. Plant Res. 2011, 124, 509–525. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Abiotic Stress, the Field Environment and Stress Combination. Trends Plant Sci. 2006, 11, 15–19. [Google Scholar] [CrossRef]
- Mohanta, T.K.; Bashir, T.; Hashem, A.; Abd_Allah, E.F. Systems Biology Approach in Plant Abiotic Stresses. Plant Physiol. Biochem. 2017, 121, 58–73. [Google Scholar] [CrossRef] [PubMed]
- Zagorchev, L.; Seal, C.E.; Kranner, I.; Odjakova, M. A Central Role for Thiols in Plant Tolerance to Abiotic Stress. Int. J. Mol. Sci. 2013, 14, 7405–7432. [Google Scholar] [CrossRef] [PubMed]
- Kulasek, M.; Bernacki, M.J.; Ciszak, K.; Witoń, D.; Karpiński, S. Contribution of PsbS Function and Stomatal Conductance to Foliar Temperature in Higher Plants. Plant Cell Physiol. 2016, 57, 1495–1509. [Google Scholar] [CrossRef] [PubMed]
- Kul, R.; Ekinci, M.; Turan, M.; Ors, S.; Yildirim, E. How Abiotic Stress Conditions Affects Plant Roots. In Plant roots; IntechOpen, 2020 ISBN 1-83968-276-0. [CrossRef]
- Sanchez, A.; Shin, J.; Davis, S.J. Abiotic Stress and the Plant Circadian Clock. Plant Signal. Behav. 2011, 6, 223–231. [Google Scholar] [CrossRef]
- ElSayed, A.; Rafudeen, M.; Golldack, D. Physiological Aspects of Raffinose Family Oligosaccharides in Plants: Protection against Abiotic Stress. Plant Biol. 2014, 16, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ziogas, V.; Tanou, G.; Filippou, P.; Diamantidis, G.; Vasilakakis, M.; Fotopoulos, V.; Molassiotis, A. Nitrosative Responses in Citrus Plants Exposed to Six Abiotic Stress Conditions. Plant Physiol. Biochem. 2013, 68, 118–126. [Google Scholar] [CrossRef]
- Mayrose, M.; Kane, N.C.; Mayrose, I.; Dlugosch, K.M.; Rieseberg, L.H. Increased Growth in Sunflower Correlates with Reduced Defences and Altered Gene Expression in Response to Biotic and Abiotic Stress. Mol. Ecol. 2011, 20, 4683–4694. [Google Scholar] [CrossRef]
- Waters, A.J.; Makarevitch, I.; Noshay, J.; Burghardt, L.T.; Hirsch, C.N.; Hirsch, C.D.; Springer, N.M. Natural Variation for Gene Expression Responses to Abiotic Stress in Maize. Plant J. 2017, 89, 706–717. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, P. Plant Abiotic Stress Responses and microRNAs. Adv. Agric. 2020, 119–118. [Google Scholar] [CrossRef]
- Dong, S.; Adams, K.L. Differential Contributions to the Transcriptome of Duplicated Genes in Response to Abiotic Stresses in Natural and Synthetic Polyploids. New Phytol. 2011, 190, 1045–1057. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, V.; Shahzad, B.; Ramakrishnan, M.; Singh Sidhu, G.P.; Bali, A.S.; Handa, N.; Kapoor, D.; Yadav, P.; Khanna, K. Photosynthetic Response of Plants under Different Abiotic Stresses: A Review. J. Plant Growth Regul. 2020, 39, 509–531. [Google Scholar] [CrossRef]
- Rusaczonek, A.; Czarnocka, W.; Kacprzak, S.; Witoń, D.; Ślesak, I.; Szechyńska-Hebda, M.; Gawroński, P.; Karpiński, S. Role of Phytochromes A and B in the Regulation of Cell Death and Acclimatory Responses to UV Stress in Arabidopsis thaliana. J. Exp. Bot. 2015, 66, 6679–6695. [Google Scholar] [CrossRef] [PubMed]
- Bourdais, G.; Burdiak, P.; Gauthier, A.; Nitsch, L.; Salojärvi, J.; Rayapuram, C.; Idänheimo, N.; Hunter, K.; Kimura, S.; Merilo, E. Large-Scale Phenomics Identifies Primary and Fine-Tuning Roles for CRKs in Responses Related to Oxidative Stress. PLoS Genet. 2015, 11, e1005373. [Google Scholar] [CrossRef] [PubMed]
- Burdiak, P.; Mielecki, J.; Gawroński, P.; Karpiński, S. The CRK5 and WRKY53 Are Conditional Regulators of Senescence and Stomatal Conductance in Arabidopsis. Cells 2022, 11, 3558. [Google Scholar] [CrossRef] [PubMed]
- Wituszyńska, W.; SZECHYŃSKA-HEBDA, M.; Sobczak, M.; Rusaczonek, A.; KOZŁOWSKA-MAKULSKA, A.; Witoń, D.; Karpiński, S. LESION SIMULATING DISEASE 1 and ENHANCED DISEASE SUSCEPTIBILITY 1 Differentially Regulate UV-C-induced Photooxidative Stress Signalling and Programmed Cell Death in Arabidopsis thaliana. Plant Cell Environ. 2015, 38, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Gechev, T.; Petrov, V. Reactive Oxygen Species and Abiotic Stress in Plants. Int. J. Mol. Sci. 2020, 21, 7433. [Google Scholar] [CrossRef]
- Bali, A.S.; Sidhu, G.P.S. Abiotic Stress-Induced Oxidative Stress in Wheat. Wheat Prod. Chang. Environ. Responses Adapt. Toler. 2019, 225–239. [Google Scholar] [CrossRef]
- Yadav, S.; Modi, P.; Dave, A.; Vijapura, A.; Patel, D.; Patel, M. Effect of Abiotic Stress on Crops. Sustain. Crop Prod. 2020, 3, 5–16. [Google Scholar] [CrossRef]
- Liang, B.-W.; Li, C.; Bai, T.-H.; Wang, P. Nutrient Use Efficiency of Plants under Abiotic Stress. Front. Plant Sci. 2023, 14, 1179842. [Google Scholar] [CrossRef] [PubMed]
- Mitra, G. Molecular Approaches to Nutrient Uptake and Cellular Homeostasis in Plants under Abiotic Stress. Plant Nutr. Abiotic Stress Toler. 2018, 525–590. [Google Scholar] [CrossRef]
- Balfagón, D.; Gómez-Cadenas, A.; Rambla, J.L.; Granell, A.; de Ollas, C.; Bassham, D.C.; Mittler, R.; Zandalinas, S.I. γ-Aminobutyric Acid Plays a Key Role in Plant Acclimation to a Combination of High Light and Heat Stress. Plant Physiol. 2022, 188, 2026–2038. [Google Scholar] [CrossRef]
- Pereira, A. Plant Abiotic Stress Challenges from the Changing Environment. Front. Plant Sci. 2016, 7, 1123. [Google Scholar] [CrossRef]
- Kolb, S.; Senizza, B.; Araniti, F.; Lewin, S.; Wende, S.; Lucini, L. A Multi-Omics Approach to Unravel the Interaction between Heat and Drought Stress in the Arabidopsis thaliana Holobiont. Authorea Prepr. 2022. [Google Scholar] [CrossRef]
- Jha, A.; Khobra, R.; Masthigowda, M.H.; Dohrey, P.; Wadhwa, Z.; Deswal, K.; Singh, G.; Singh, G.P. Understanding Heat and Drought Stress Adaptation Mechanisms in Wheat: A Combined Approach. J. Cereal Res. 2022, 14. [Google Scholar] [CrossRef]
- Leal, A.R.; Belo, J.; Beeckman, T.; Barros, P.M.; Oliveira, M.M. The Combined Effect of Heat and Osmotic Stress on Suberization of Arabidopsis Roots. Cells 2022, 11, 2341. [Google Scholar] [CrossRef]
- Gonzalez Guzman, M.; Cellini, F.; Fotopoulos, V.; Balestrini, R.; Arbona, V. New Approaches to Improve Crop Tolerance to Biotic and Abiotic Stresses. Physiol. Plant. 2022, 174, e13547. [Google Scholar] [CrossRef] [PubMed]
- Dresselhaus, T.; Hückelhoven, R. Biotic and Abiotic Stress Responses in Crop Plants. Agronomy 2018, 8, 267. [Google Scholar] [CrossRef]
- Enebe, M.C.; Babalola, O.O. The Impact of Microbes in the Orchestration of Plants’ Resistance to Biotic Stress: A Disease Management Approach. Appl. Microbiol. Biotechnol. 2019, 103, 9–25. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Wang, H.; Chen, S.; Yu, D.; Reiter, R.J. Phytomelatonin: An Emerging Regulator of Plant Biotic Stress Resistance. Trends Plant Sci. 2021, 26, 70–82. [Google Scholar] [CrossRef]
- Yan, X.; Talbot, N.J. Investigating the Cell Biology of Plant Infection by the Rice Blast Fungus Magnaporthe oryzae. Curr. Opin. Microbiol. 2016, 34, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Andreo-Jimenez, B.; Ruyter-Spira, C.; Bouwmeester, H.J.; Lopez-Raez, J.A. Ecological Relevance of Strigolactones in Nutrient Uptake and Other Abiotic Stresses, and in Plant-Microbe Interactions below-Ground. Plant Soil 2015, 394, 1–19. [Google Scholar] [CrossRef]
- Sundin, G.W.; Castiblanco, L.F.; Yuan, X.; Zeng, Q.; Yang, C. Bacterial Disease Management: Challenges, Experience, Innovation and Future Prospects: Challenges in Bacterial Molecular Plant Pathology. Mol. Plant Pathol. 2016, 17, 1506–1518. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Lou, Y.-R.; Tzin, V.; Jander, G. Alteration of Plant Primary Metabolism in Response to Insect Herbivory. Plant Physiol. 2015, 169, 1488–1498. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, F.E.; Peiffer, M.; Tan, C.-W.; Stanley, B.A.; Stanley, A.; Wang, J.; Jones, A.G.; Hoover, K.; Rosa, C.; Luthe, D. Fall Armyworm-Associated Gut Bacteria Modulate Plant Defense Responses. Mol. Plant. Microbe Interact. 2017, 30, 127–137. [Google Scholar] [CrossRef]
- Wonglom, P.; Ito, S.; Sunpapao, A. Volatile Organic Compounds Emitted from Endophytic Fungus Trichoderma asperellum T1 Mediate Antifungal Activity, Defense Response and Promote Plant Growth in Lettuce (Lactuca sativa). Fungal Ecol. 2020, 43, 100867. [Google Scholar] [CrossRef]
- Jones, J.D.; Dangl, J.L. The Plant Immune System. nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Rustérucci, C.; Aviv, D.H.; Holt III, B.F.; Dangl, J.L.; Parker, J.E. The Disease Resistance Signaling Components EDS1 and PAD4 Are Essential Regulators of the Cell Death Pathway Controlled by LSD1 in Arabidopsis. Plant Cell 2001, 13, 2211–2224. [Google Scholar] [CrossRef] [PubMed]
- Kaur, B.; Bhatia, D.; Mavi, G. Eighty Years of Gene-for-Gene Relationship and Its Applications in Identification and Utilization of R Genes. J. Genet. 2021, 100, 50. [Google Scholar] [CrossRef]
- Wu, L.; Chen, H.; Curtis, C.; Fu, Z.Q. Go in for the Kill: How Plants Deploy Effector-Triggered Immunity to Combat Pathogens. Virulence 2014, 5, 710–721. [Google Scholar] [CrossRef]
- Zhang, S.; Li, C.; Si, J.; Han, Z.; Chen, D. Action Mechanisms of Effectors in Plant-Pathogen Interaction. Int. J. Mol. Sci. 2022, 23, 6758. [Google Scholar] [CrossRef] [PubMed]
- Glazebrook, J. Contrasting Mechanisms of Defense against Biotrophic and Necrotrophic Pathogens. Annu Rev Phytopathol 2005, 43, 205–227. [Google Scholar] [CrossRef]
- Nadarajah, K.K. Defensive Strategies of ROS in Plant–Pathogen Interactions. In Plant Pathogen Interaction; Springer, 2024; pp. 163–183. [CrossRef]
- Ding, L.-N.; Li, Y.-T.; Wu, Y.-Z.; Li, T.; Geng, R.; Cao, J.; Zhang, W.; Tan, X.-L. Plant Disease Resistance-Related Signaling Pathways: Recent Progress and Future Prospects. Int. J. Mol. Sci. 2022, 23, 16200. [Google Scholar] [CrossRef] [PubMed]
- Fei, W.; Liu, Y. Biotrophic Fungal Pathogens: A Critical Overview. Appl. Biochem. Biotechnol. 2023, 195, 1–16. [Google Scholar] [CrossRef]
- Tsuda, K.; Glazebrook, J.; Katagiri, F. The Interplay between MAMP and SA Signaling. Plant Signal. Behav. 2008, 3, 359–361. [Google Scholar] [CrossRef]
- Thomma, B.P.; Eggermont, K.; Penninckx, I.A.; Mauch-Mani, B.; Vogelsang, R.; Cammue, B.P.; Broekaert, W.F. Separate Jasmonate-Dependent and Salicylate-Dependent Defense-Response Pathways in Arabidopsis Are Essential for Resistance to Distinct Microbial Pathogens. Proc. Natl. Acad. Sci. 1998, 95, 15107–15111. [Google Scholar] [CrossRef]
- McDowell, J.M.; Dangl, J.L. Signal Transduction in the Plant Immune Response. Trends Biochem. Sci. 2000, 25, 79–82. [Google Scholar] [CrossRef] [PubMed]
- Ghozlan, M.H.; Eman, E.-A.; Tokgöz, S.; Lakshman, D.K.; Mitra, A. Plant Defense against Necrotrophic Pathogens. Am. J. Plant Sci. 2020, 11, 2122–2138. [Google Scholar] [CrossRef]
- Noctor, G.; Foyer, C.H. Intracellular Redox Compartmentation and ROS-Related Communication in Regulation and Signaling. Plant Physiol. 2016, 171, 1581–1592. [Google Scholar] [CrossRef]
- Mateo, A.; Mühlenbock, P.; Rustérucci, C.; Chang, C.C.-C.; Miszalski, Z.; Karpinska, B.; Parker, J.E.; Mullineaux, P.M.; Karpinski, S. LESION SIMULATING DISEASE 1 Is Required for Acclimation to Conditions That Promote Excess Excitation Energy. Plant Physiol. 2004, 136, 2818–2830. [Google Scholar] [CrossRef]
- Gawroński, P.; Witoń, D.; Vashutina, K.; Bederska, M.; Betliński, B.; Rusaczonek, A.; Karpiński, S. Mitogen-Activated Protein Kinase 4 Is a Salicylic Acid-Independent Regulator of Growth but Not of Photosynthesis in Arabidopsis. Mol. Plant 2014, 7, 1151–1166. [Google Scholar] [CrossRef]
- Johnson, R.; Puthur, J.T. Seed Priming as a Cost Effective Technique for Developing Plants with Cross Tolerance to Salinity Stress. Plant Physiol. Biochem. 2021, 162, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Arora, R. Dynamics of the Antioxidant System during Seed Osmopriming, Post-Priming Germination, and Seedling Establishment in Spinach (Spinacia oleracea). Plant Sci. 2011, 180, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Ramegowda, V.; Da Costa, M.V.J.; Harihar, S.; Karaba, N.N.; Sreeman, S.M. Abiotic and Biotic Stress Interactions in Plants: A Cross-Tolerance Perspective. In Priming-mediated stress and cross-stress tolerance in crop plants; Elsevier, 2020; pp. 267–302. [CrossRef]
- Lee, K.; Missaoui, A.; Mahmud, K.; Presley, H.; Lonnee, M. Interaction between Grasses and Epichloë Endophytes and Its Significance to Biotic and Abiotic Stress Tolerance and the Rhizosphere. Microorganisms 2021, 9, 2186. [Google Scholar] [CrossRef] [PubMed]
- Capiati, D.A.; País, S.M.; Téllez-Iñón, M.T. Wounding Increases Salt Tolerance in Tomato Plants: Evidence on the Participation of Calmodulin-like Activities in Cross-Tolerance Signalling. J. Exp. Bot. 2006, 57, 2391–2400. [Google Scholar] [CrossRef] [PubMed]
- de Lima, J.D.; Dedido, D.B.; Volpato, L.A.; da Silva, A.P.; Lima, G.O.; Pinheiro, C.R.; da Silva, G.J. Cross Tolerance by Heat and Water Stress at Germination of Common Beans. Sci. Agrar. Parana. 2018, 515–520. [Google Scholar] [CrossRef]
- Carvalho, F.E.; Silveira, J.A. H2O2-Retrograde Signaling as a Pivotal Mechanism to Understand Priming and Cross Stress Tolerance in Plants. In Priming-mediated stress and cross-stress tolerance in crop plants; Elsevier, 2020; pp. 57–78. [CrossRef]
- Jan, M.; Liu, Z.; Rochaix, J.-D.; Sun, X. Retrograde and Anterograde Signaling in the Crosstalk between Chloroplast and Nucleus. Front. Plant Sci. 2022, 13, 980237. [Google Scholar] [CrossRef] [PubMed]
- Corti, F.; Festa, M.; Szabo, I. Mitochondria–Chloroplast Cross Talk: A Possible Role for Calcium and Reactive Oxygen Species? Bioelectricity 2023, 5, 39–46. [Google Scholar] [CrossRef]
- Ciszak, K.; Kulasek, M.; Barczak, A.; Grzelak, J.; Maćkowski, S.; Karpiński, S. PsbS Is Required for Systemic Acquired Acclimation and Post-Excess-Light-Stress Optimization of Chlorophyll Fluorescence Decay Times in Arabidopsis. Plant Signal. Behav. 2015, 10, e982018. [Google Scholar] [CrossRef] [PubMed]
- Witoń, D.; Sujkowska-Rybkowska, M.; Dąbrowska-Bronk, J.; Czarnocka, W.; Bernacki, M.; Szechyńska-Hebda, M.; Karpiński, S. MITOGEN-ACTIVATED PROTEIN KINASE 4 Impacts Leaf Development, Temperature, and Stomatal Movement in Hybrid Aspen. Plant Physiol. 2021, 186, 2190–2204. [Google Scholar] [CrossRef] [PubMed]
- Ball, L.; Accotto, G.-P.; Bechtold, U.; Creissen, G.; Funck, D.; Jimenez, A.; Kular, B.; Leyland, N.; Mejia-Carranza, J.; Reynolds, H. Evidence for a Direct Link between Glutathione Biosynthesis and Stress Defense Gene Expression in Arabidopsis. Plant Cell 2004, 16, 2448–2462. [Google Scholar] [CrossRef] [PubMed]
- Peak, D.; West, J.D.; Messinger, S.M.; Mott, K.A. Evidence for Complex, Collective Dynamics and Emergent, Distributed Computation in Plants. Proc. Natl. Acad. Sci. 2004, 101, 918–922. [Google Scholar] [CrossRef] [PubMed]
- Gilroy, S.; Białasek, M.; Suzuki, N.; Górecka, M.; Devireddy, A.R.; Karpiński, S.; Mittler, R. ROS, Calcium, and Electric Signals: Key Mediators of Rapid Systemic Signaling in Plants. Plant Physiol. 2016, 171, 1606–1615. [Google Scholar] [CrossRef]
- Kulheim, C.; Ågren, J.; Jansson, S. Rapid Regulation of Light Harvesting and Plant Fitness in the Field. Science 2002, 297, 91–93. [Google Scholar] [CrossRef]
- Kromdijk, J.; Głowacka, K.; Leonelli, L.; Gabilly, S.T.; Iwai, M.; Niyogi, K.K.; Long, S.P. Improving Photosynthesis and Crop Productivity by Accelerating Recovery from Photoprotection. Science 2016, 354, 857–861. [Google Scholar] [CrossRef]
- De Souza, A.P.; Burgess, S.J.; Doran, L.; Hansen, J.; Manukyan, L.; Maryn, N.; Gotarkar, D.; Leonelli, L.; Niyogi, K.K.; Long, S.P. Soybean Photosynthesis and Crop Yield Are Improved by Accelerating Recovery from Photoprotection. Science 2022, 377, 851–854. [Google Scholar] [CrossRef] [PubMed]
- Karpinski, S.; Reynolds, H.; Karpinska, B.; Wingsle, G.; Creissen, G.; Mullineaux, P. Systemic Signaling and Acclimation in Response to Excess Excitation Energy in Arabidopsis. science 1999, 284, 654–657. [Google Scholar] [CrossRef]
- Pandey, P.; Irulappan, V.; Bagavathiannan, M.V.; Senthil-Kumar, M. Impact of Combined Abiotic and Biotic Stresses on Plant Growth and Avenues for Crop Improvement by Exploiting Physio-Morphological Traits. Front. Plant Sci. 2017, 8, 537. [Google Scholar] [CrossRef] [PubMed]
- Prasad, P.; Pisipati, S.; Momčilović, I.; Ristic, Z. Independent and Combined Effects of High Temperature and Drought Stress during Grain Filling on Plant Yield and Chloroplast EF-Tu Expression in Spring Wheat. J. Agron. Crop Sci. 2011, 197, 430–441. [Google Scholar] [CrossRef]
- Coakley, S.M.; Scherm, H.; Chakraborty, S. Climate Change and Plant Disease Management. Annu. Rev. Phytopathol. 1999, 37, 399–426. [Google Scholar] [CrossRef] [PubMed]
- Seherm, H.; Coakley, S.M. Plant Pathogens in a Changing World. Australas. Plant Pathol. 2003, 32, 157–165. [Google Scholar] [CrossRef]
- McDonald, A.; Riha, S.; DiTommaso, A.; DeGaetano, A. Climate Change and the Geography of Weed Damage: Analysis of US Maize Systems Suggests the Potential for Significant Range Transformations. Agric. Ecosyst. Environ. 2009, 130, 131–140. [Google Scholar] [CrossRef]
- Ziska, L.H.; Tomecek, M.B.; Gealy, D.R. Competitive Interactions between Cultivated and Red Rice as a Function of Recent and Projected Increases in Atmospheric Carbon Dioxide. Agron. J. 2010, 102, 118–123. [Google Scholar] [CrossRef]
- Peters, K.; Breitsameter, L.; Gerowitt, B. Impact of Climate Change on Weeds in Agriculture: A Review. Agron. Sustain. Dev. 2014, 34, 707–721. [Google Scholar] [CrossRef]
- Duveiller, E.; Singh, R.P.; Nicol, J.M. The Challenges of Maintaining Wheat Productivity: Pests, Diseases, and Potential Epidemics. Euphytica 2007, 157, 417–430. [Google Scholar] [CrossRef]
- Valerio, M.; Lovelli, S.; Perniola, M.; Di Tommaso, T.; Ziska, L. The Role of Water Availability on Weed–Crop Interactions in Processing Tomato for Southern Italy. Acta Agric. Scand. Sect. B–Soil Plant Sci. 2013, 63, 62–68. [Google Scholar] [CrossRef]
- Atkinson, N.J.; Urwin, P.E. The Interaction of Plant Biotic and Abiotic Stresses: From Genes to the Field. J. Exp. Bot. 2012, 63, 3523–3543. [Google Scholar] [CrossRef]
- Prasch, C.M.; Sonnewald, U. Simultaneous Application of Heat, Drought, and Virus to Arabidopsis Plants Reveals Significant Shifts in Signaling Networks. Plant Physiol. 2013, 162, 1849–1866. [Google Scholar] [CrossRef]
- Pandey, P.; Ramegowda, V.; Senthil-Kumar, M. Shared and Unique Responses of Plants to Multiple Individual Stresses and Stress Combinations: Physiological and Molecular Mechanisms. Front. Plant Sci. 2015, 6, 723. [Google Scholar] [CrossRef]
- Choudhary, A.; Pandey, P.; Senthil-Kumar, M. Tailored Responses to Simultaneous Drought Stress and Pathogen Infection in Plants. Drought Stress Toler. Plants Vol 1 Physiol. Biochem. 2016, 427–438. [Google Scholar] [CrossRef]
- Ramu, V.S.; Paramanantham, A.; Ramegowda, V.; Mohan-Raju, B.; Udayakumar, M.; Senthil-Kumar, M. Transcriptome Analysis of Sunflower Genotypes with Contrasting Oxidative Stress Tolerance Reveals Individual-and Combined-Biotic and Abiotic Stress Tolerance Mechanisms. PloS One 2016, 11, e0157522. [Google Scholar] [CrossRef] [PubMed]
- Kissoudis, C.; van de Wiel, C.; Visser, R.G.; van der Linden, G. Enhancing Crop Resilience to Combined Abiotic and Biotic Stress through the Dissection of Physiological and Molecular Crosstalk. Front. Plant Sci. 2014, 5, 207. [Google Scholar] [CrossRef]
- Bilgin, D.D.; Zavala, J.A.; Zhu, J.; Clough, S.J.; Ort, D.R.; DeLUCIA, E.H. Biotic Stress Globally Downregulates Photosynthesis Genes. Plant Cell Environ. 2010, 33, 1597–1613. [Google Scholar] [CrossRef]
- Grimmer, M.K.; John Foulkes, M.; Paveley, N.D. Foliar Pathogenesis and Plant Water Relations: A Review. J. Exp. Bot. 2012, 63, 4321–4331. [Google Scholar] [CrossRef]
- You, M.P.; Colmer, T.D.; Barbetti, M.J. Salinity Drives Host Reaction in Phaseolus vulgaris (Common Bean) to Macrophomina phaseolina. Funct. Plant Biol. 2011, 38, 984–992. [Google Scholar] [CrossRef]
- Kim, T.-H.; Hauser, F.; Ha, T.; Xue, S.; Böhmer, M.; Nishimura, N.; Munemasa, S.; Hubbard, K.; Peine, N.; Lee, B. Chemical Genetics Reveals Negative Regulation of Abscisic Acid Signaling by a Plant Immune Response Pathway. Curr. Biol. 2011, 21, 990–997. [Google Scholar] [CrossRef] [PubMed]
- Jelenska, J.; Van Hal, J.A.; Greenberg, J.T. Pseudomonas Syringae Hijacks Plant Stress Chaperone Machinery for Virulence. Proc. Natl. Acad. Sci. 2010, 107, 13177–13182. [Google Scholar] [CrossRef] [PubMed]
- Clément, M.; Leonhardt, N.; Droillard, M.-J.; Reiter, I.; Montillet, J.-L.; Genty, B.; Lauriere, C.; Nussaume, L.; Noël, L.D. The Cytosolic/Nuclear HSC70 and HSP90 Molecular Chaperones Are Important for Stomatal Closure and Modulate Abscisic Acid-Dependent Physiological Responses in Arabidopsis. Plant Physiol. 2011, 156, 1481–1492. [Google Scholar] [CrossRef]
- Goel, A.K.; Lundberg, D.; Torres, M.A.; Matthews, R.; Akimoto-Tomiyama, C.; Farmer, L.; Dangl, J.L.; Grant, S.R. The Pseudomonas Syringae Type III Effector HopAM1 Enhances Virulence on Water-Stressed Plants. Mol. Plant. Microbe Interact. 2008, 21, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Chen, F.; Mannas, J.P.; Feldman, T.; Sumner, L.W.; Roossinck, M.J. Virus Infection Improves Drought Tolerance. New Phytol. 2008, 180, 911–921. [Google Scholar] [CrossRef]
- Reusche, M.; Thole, K.; Janz, D.; Truskina, J.; Rindfleisch, S.; Drübert, C.; Polle, A.; Lipka, V.; Teichmann, T. Verticillium Infection Triggers VASCULAR-RELATED NAC DOMAIN7–Dependent de Novo Xylem Formation and Enhances Drought Tolerance in Arabidopsis. Plant Cell 2012, 24, 3823–3837. [Google Scholar] [CrossRef]
- Dodd, I.C.; Pérez-Alfocea, F. Microbial Amelioration of Crop Salinity Stress. J. Exp. Bot. 2012, 63, 3415–3428. [Google Scholar] [CrossRef]
- Márquez, L.M.; Redman, R.S.; Rodriguez, R.J.; Roossinck, M.J. A Virus in a Fungus in a Plant: Three-Way Symbiosis Required for Thermal Tolerance. science 2007, 315, 513–515. [Google Scholar] [CrossRef] [PubMed]
- Berendsen, R.L.; Pieterse, C.M.; Bakker, P.A. The Rhizosphere Microbiome and Plant Health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Goellner, K.; Conrath, U. Priming: It’s All the World to Induced Disease Resistance. Sustain. Dis. Manag. Eur. Context 2008, 233–242. [Google Scholar] [CrossRef]
- Kissoudis, C. Genetics and Regulation of Combined Abiotic and Biotic Stress Tolerance in Tomato. 2016. [CrossRef]
- Karimi, Z.; Khara, J.; Habibi, G. Combined Hydrogen Peroxide and Nitric Oxide Priming Modulate Salt Stress Tolerance in Acclimated and Non-Acclimated Oilseed Rape (Brassica napus L.) Plants. J. Plant Physiol. Breed. 2020, 10, 27–43. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Nafees, M.; Chen, J.; Darras, A.; Ferrante, A.; Hancock, J.T.; Ashraf, M.; Zaid, A.; Latif, N.; Corpas, F.J. Chemical Priming Enhances Plant Tolerance to Salt Stress. Front. Plant Sci. 2022, 13, 946922. [Google Scholar] [CrossRef] [PubMed]
- Ellouzi, H.; Oueslati, S.; Hessini, K.; Rabhi, M.; Abdelly, C. Seed-Priming with H2O2 Alleviates Subsequent Salt Stress by Preventing ROS Production and Amplifying Antioxidant Defense in Cauliflower Seeds and Seedlings. Sci. Hortic. 2021, 288, 110360. [Google Scholar] [CrossRef]
- Momeni, M.M.; Kalantar, M.; Dehghani-Zahedani, M. H2O2 Seed Priming Improves Tolerance to Salinity Stress in Durum Wheat. Cereal Res. Commun. 2023, 51, 391–401. [Google Scholar] [CrossRef]
- Pillay, L.C.; Nekati, L.; Makhwitine, P.J.; Ndlovu, S.I. Epigenetic Activation of Silent Biosynthetic Gene Clusters in Endophytic Fungi Using Small Molecular Modifiers. Front. Microbiol. 2022, 13, 815008. [Google Scholar] [CrossRef] [PubMed]
- Baillo, E.H.; Kimotho, R.N.; Zhang, Z.; Xu, P. Transcription Factors Associated with Abiotic and Biotic Stress Tolerance and Their Potential for Crops Improvement. Genes 2019, 10, 771. [Google Scholar] [CrossRef]
- Muhlenbock, P.; Plaszczyca, M.; Plaszczyca, M.; Mellerowicz, E.; Karpinski, S. Lysigenous Aerenchyma Formation in Arabidopsis Is Controlled by LESION SIMULATING DISEASE1. Plant Cell 2007, 19, 3819–3830. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Li, Y.; Zhang, X.; Zuo, J.; Yang, S. The Arabidopsis LSD1 Gene Plays an Important Role in the Regulation of Low Temperature-dependent Cell Death. New Phytol. 2010, 187, 301–312. [Google Scholar] [CrossRef]
- Czarnocka, W.; Van Der Kelen, K.; Willems, P.; Szechyńska-Hebda, M.; Shahnejat-Bushehri, S.; Balazadeh, S.; Rusaczonek, A.; Mueller-Roeber, B.; Van Breusegem, F.; Karpiński, S. The Dual Role of LESION SIMULATING DISEASE 1 as a Condition-dependent Scaffold Protein and Transcription Regulator. Plant Cell Environ. 2017, 40, 2644–2662. [Google Scholar] [CrossRef]
- Parker, J.E.; Holub, E.B.; Frost, L.N.; Falk, A.; Gunn, N.D.; Daniels, M.J. Characterization of Eds1, a Mutation in Arabidopsis Suppressing Resistance to Peronospora parasitica Specified by Several Different RPP Genes. Plant Cell 1996, 8, 2033–2046. [Google Scholar] [CrossRef] [PubMed]
- Glazebrook, J.; Zook, M.; Mert, F.; Kagan, I.; Rogers, E.E.; Crute, I.R.; Holub, E.B.; Hammerschmidt, R.; Ausubel, F.M. Phytoalexin-Deficient Mutants of Arabidopsis Reveal That PAD4 Encodes a Regulatory Factor and That Four PAD Genes Contribute to Downy Mildew Resistance. Genetics 1997, 146, 381–392. [Google Scholar] [CrossRef]
- Bernacki, M.J.; Rusaczonek, A.; Czarnocka, W.; Karpiński, S. Salicylic Acid Accumulation Controlled by LSD1 Is Essential in Triggering Cell Death in Response to Abiotic Stress. Cells 2021, 10, 962. [Google Scholar] [CrossRef]
- Bernacki, M.J.; Mielecki, J.; Antczak, A.; Drożdżek, M.; Witoń, D.; Dąbrowska-Bronk, J.; Gawroński, P.; Burdiak, P.; Marchwicka, M.; Rusaczonek, A. Biotechnological Potential of the Stress Response and Plant Cell Death Regulators Proteins in the Biofuel Industry. Cells 2023, 12, 2018. [Google Scholar] [CrossRef]
- Bernacki, M.J.; Rusaczonek, A.; Gołębiewska, K.; Majewska-Fala, A.B.; Czarnocka, W.; Karpiński, S.M. METACASPASE8 (MC8) Is a Crucial Protein in the LSD1-Dependent Cell Death Pathway in Response to Ultraviolet Stress. Int. J. Mol. Sci. 2024, 25, 3195. [Google Scholar] [CrossRef] [PubMed]
- Ślesak, I.; SZECHYŃSKA-HEBDA, M.; Fedak, H.; Sidoruk, N.; DĄBROWSKA-BRONK, J.; Witoń, D.; Rusaczonek, A.; Antczak, A.; Drożdżek, M.; Karpińska, B. PHYTOALEXIN DEFICIENT 4 Affects Reactive Oxygen Species Metabolism, Cell Wall and Wood Properties in Hybrid Aspen (Populus tremula L.× tremuloides). Plant Cell Environ. 2015, 38, 1275–1284. [Google Scholar] [CrossRef]
- Bernacki, M.J.; Czarnocka, W.; Witoń, D.; Rusaczonek, A.; Szechyńska-Hebda, M.; Ślesak, I.; Dąbrowska-Bronk, J.; Karpiński, S. ENHANCED DISEASE SUSCEPTIBILITY 1 (EDS1) Affects Development, Photosynthesis, and Hormonal Homeostasis in Hybrid Aspen (Populus tremula L.× P. tremuloides). J. Plant Physiol. 2018, 226, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, H.; Shao, H.; Tang, X. Recent Advances in Utilizing Transcription Factors to Improve Plant Abiotic Stress Tolerance by Transgenic Technology. Front. Plant Sci. 2016, 7, 67. [Google Scholar] [CrossRef]
- Aglawe, S.; Fakrudin, B.; Patole, C.; Bhairappanavar, S.; Koti, R.; Krishnaraj, P. Quantitative RT-PCR Analysis of 20 Transcription Factor Genes of MADS, ARF, HAP2, MBF and HB Families in Moisture Stressed Shoot and Root Tissues of Sorghum. Physiol. Mol. Biol. Plants 2012, 18, 287–300. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, J.; Zhang, B.; Vanitha, J.; Ramachandran, S.; Jiang, S. Genome-wide Expansion and Expression Divergence of the Basic Leucine Zipper Transcription Factors in Higher Plants with an Emphasis on Sorghum F. J. Integr. Plant Biol. 2011, 53, 212–231. [Google Scholar] [CrossRef] [PubMed]
- Lan Thi Hoang, X.; Ngoc Hai Nhi, D.; Binh Anh Thu, N.; Phuong Thao, N.; Phan Tran, L.-S. Transcription Factors and Their Roles in Signal Transduction in Plants under Abiotic Stresses. Curr. Genomics 2017, 18, 483–497. [Google Scholar] [CrossRef]
- Mao, G.; Meng, X.; Liu, Y.; Zheng, Z.; Chen, Z.; Zhang, S. Phosphorylation of a WRKY Transcription Factor by Two Pathogen-Responsive MAPKs Drives Phytoalexin Biosynthesis in Arabidopsis. Plant Cell 2011, 23, 1639–1653. [Google Scholar] [CrossRef]
- Meng, X.; Xu, J.; He, Y.; Yang, K.-Y.; Mordorski, B.; Liu, Y.; Zhang, S. Phosphorylation of an ERF Transcription Factor by Arabidopsis MPK3/MPK6 Regulates Plant Defense Gene Induction and Fungal Resistance. Plant Cell 2013, 25, 1126–1142. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Wang, M.; Chen, Y.; Nomura, K.; Hui, S.; Gui, J.; Zhang, X.; Wu, Y.; Liu, J.; Li, Q. An MKP-MAPK Protein Phosphorylation Cascade Controls Vascular Immunity in Plants. Sci. Adv. 2022, 8, eabg8723. [Google Scholar] [CrossRef]
- Li, F.; Li, M.; Wang, P.; Cox Jr, K.L.; Duan, L.; Dever, J.K.; Shan, L.; Li, Z.; He, P. Regulation of Cotton (Gossypium hirsutum) Drought Responses by Mitogen-activated Protein (MAP) Kinase Cascade-mediated Phosphorylation of Gh WRKY 59. New Phytol. 2017, 215, 1462–1475. [Google Scholar] [CrossRef]
- Tao, Y.; Wan, J.X.; Liu, Y.S.; Yang, X.Z.; Shen, R.F.; Zhu, X.F. The NAC Transcription Factor ANAC017 Regulates Aluminum Tolerance by Regulating the Cell Wall-Modifying Genes. Plant Physiol. 2022, 189, 2517–2534. [Google Scholar] [CrossRef]
- Khandelwal, A.; Elvitigala, T.; Ghosh, B.; Quatrano, R.S. Arabidopsis Transcriptome Reveals Control Circuits Regulating Redox Homeostasis and the Role of an AP2 Transcription Factor. Plant Physiol. 2008, 148, 2050–2058. [Google Scholar] [CrossRef]
- Matsuo, M.; Johnson, J.M.; Hieno, A.; Tokizawa, M.; Nomoto, M.; Tada, Y.; Godfrey, R.; Obokata, J.; Sherameti, I.; Yamamoto, Y.Y. High REDOX RESPONSIVE TRANSCRIPTION FACTOR1 Levels Result in Accumulation of Reactive Oxygen Species in Arabidopsis thaliana Shoots and Roots. Mol. Plant 2015, 8, 1253–1273. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.-T.; Xu, P.; Zhao, P.-X.; Liu, R.; Yu, L.-H.; Xiang, C.-B. Arabidopsis ERF109 Mediates Cross-Talk between Jasmonic Acid and Auxin Biosynthesis during Lateral Root Formation. Nat. Commun. 2014, 5, 5833. [Google Scholar] [CrossRef]
- Bahieldin, A.; Atef, A.; Edris, S.; Gadalla, N.O.; Ali, H.M.; Hassan, S.M.; Al-Kordy, M.A.; Ramadan, A.M.; Makki, R.M.; Al-Hajar, A.S. Ethylene Responsive Transcription Factor ERF109 Retards PCD and Improves Salt Tolerance in Plant. BMC Plant Biol. 2016, 16, 1–9. [Google Scholar] [CrossRef]
- Cantabella, D.; Karpinska, B.; Teixidó, N.; Dolcet-Sanjuan, R.; Foyer, C.H. Non-Volatile Signals and Redox Mechanisms Are Required for the Responses of Arabidopsis Roots to Pseudomonas oryzihabitans. J. Exp. Bot. 2022, 73, 6971–6982. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Patil, V.; Kumar, R.; Gautam, J.K.; Singh, V.; Nandi, A.K. RSI1/FLD and Its Epigenetic Target RRTF1 Are Essential for the Retention of Infection Memory in Arabidopsis thaliana. Plant J. 2023, 115, 662–677. [Google Scholar] [CrossRef] [PubMed]
- Białas, A.; Dąbrowska-Bronk, J.; Gawroński, P.; Karpiński, S. The New Role of Carbonic Anhydrases and Bicarbonate in Acclimatization and Hypoxia Stress Responses in Arabidopsis thaliana. bioRxiv 2024, 2024–08. [Google Scholar] [CrossRef]
- Jia, Y.; Ding, Y.; Shi, Y.; Zhang, X.; Gong, Z.; Yang, S. The Cbfs Triple Mutants Reveal the Essential Functions of CBF s in Cold Acclimation and Allow the Definition of CBF Regulons in Arabidopsis. New Phytol. 2016, 212, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Dietz, K.-J.; Turkan, I.; Krieger-Liszkay, A. Redox-and Reactive Oxygen Species-Dependent Signaling into and out of the Photosynthesizing Chloroplast. Plant Physiol. 2016, 171, 1541–1550. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Van Aken, O.; Schwarzländer, M.; Belt, K.; Millar, A.H. The Roles of Mitochondrial Reactive Oxygen Species in Cellular Signaling and Stress Response in Plants. Plant Physiol. 2016, 171, 1551–1559. [Google Scholar] [CrossRef]
- Kerchev, P.; Waszczak, C.; Lewandowska, A.; Willems, P.; Shapiguzov, A.; Li, Z.; Alseekh, S.; Mühlenbock, P.; Hoeberichts, F.A.; Huang, J. Lack of GLYCOLATE OXIDASE1, but Not GLYCOLATE OXIDASE2, Attenuates the Photorespiratory Phenotype of CATALASE2-Deficient Arabidopsis. Plant Physiol. 2016, 171, 1704–1719. [Google Scholar] [CrossRef]
- Rodríguez-Serrano, M.; Romero-Puertas, M.C.; Sanz-Fernández, M.; Hu, J.; Sandalio, L.M. Peroxisomes Extend Peroxules in a Fast Response to Stress via a Reactive Oxygen Species-Mediated Induction of the Peroxin PEX11a. Plant Physiol. 2016, 171, 1665–1674. [Google Scholar] [CrossRef]
- Takagi, D.; Takumi, S.; Hashiguchi, M.; Sejima, T.; Miyake, C. Superoxide and Singlet Oxygen Produced within the Thylakoid Membranes Both Cause Photosystem I Photoinhibition. Plant Physiol. 2016, 171, 1626–1634. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive Oxygen Gene Network of Plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef]
- O’Brien, J.A.; Daudi, A.; Butt, V.S.; Paul Bolwell, G. Reactive Oxygen Species and Their Role in Plant Defence and Cell Wall Metabolism. Planta 2012, 236, 765–779. [Google Scholar] [CrossRef]
- Van Breusegem, F.; Bailey-Serres, J.; Mittler, R. Unraveling the Tapestry of Networks Involving Reactive Oxygen Species in Plants. Plant Physiol. 2008, 147, 978–984. [Google Scholar] [CrossRef] [PubMed]
- Corpas, F.J.; Gupta, D.K.; Palma, J.M. Production Sites of Reactive Oxygen Species (ROS) in Organelles from Plant Cells. React. Oxyg. Species Oxidative Damage Plants Stress 2015, 1–22. [Google Scholar] [CrossRef]
- Imran, Q.M.; Shahid, M.; Hussain, A.; Yun, B.-W. NO and ROS Crosstalk and Acquisition of Abiotic Stress Tolerance. In Nitric Oxide in Plant Biology; Elsevier, 2022; pp. 477–491. [CrossRef]
- Chen, Z.; Raji, M. Role of Reactive Oxygen Species in Modulating Cross Tolerance in Plants via Flavonoids. In Priming-mediated stress and cross-stress tolerance in crop plants; Elsevier, 2020; pp. 203–214. [CrossRef]
- Locato, V.; Cimini, S.; De Gara, L. ROS and Redox Balance as Multifaceted Players of Cross-Tolerance: Epigenetic and Retrograde Control of Gene Expression. J. Exp. Bot. 2018, 69, 3373–3391. [Google Scholar] [CrossRef]
- Mhamdi, A.; Van Breusegem, F. Reactive Oxygen Species in Plant Development. Development 2018, 145, dev164376. [Google Scholar] [CrossRef]
- Nafees, M.; Fahad, S.; Shah, A.N.; Bukhari, M.A.; Maryam; Ahmed, I.; Ahmad, S.; Hussain, S. Reactive Oxygen Species Signaling in Plants. Plant Abiotic Stress Toler. Agron. Mol. Biotechnol. Approaches 2019, 259–272. [CrossRef]
- Kazan, K.; Manners, J.M. Jasmonate Signaling: Toward an Integrated View. Plant Physiol. 2008, 146, 1459–1468. [Google Scholar] [CrossRef]
- Raza, A.; Charagh, S.; Zahid, Z.; Mubarik, M.S.; Javed, R.; Siddiqui, M.H.; Hasanuzzaman, M. Jasmonic Acid: A Key Frontier in Conferring Abiotic Stress Tolerance in Plants. Plant Cell Rep. 2021, 40, 1513–1541. [Google Scholar] [CrossRef] [PubMed]
- Wasternack, C.; Strnad, M. Jasmonate Signaling in Plant Stress Responses and Development–Active and Inactive Compounds. New Biotechnol. 2016, 33, 604–613. [Google Scholar] [CrossRef] [PubMed]
- Santner, A.; Estelle, M. Recent Advances and Emerging Trends in Plant Hormone Signalling. Nature 2009, 459, 1071–1078. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-X.; J Ahammed, G.; Wu, C.; Fan, S.; Zhou, Y.-H. Crosstalk among Jasmonate, Salicylate and Ethylene Signaling Pathways in Plant Disease and Immune Responses. Curr. Protein Pept. Sci. 2015, 16, 450–461. [Google Scholar] [CrossRef] [PubMed]
- Gatz, C. From Pioneers to Team Players: TGA Transcription Factors Provide a Molecular Link between Different Stress Pathways. Mol. Plant. Microbe Interact. 2013, 26, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, X.; Lv, Z.; Khanzada, A.; Huang, M.; Cai, J.; Zhou, Q.; Huo, Z.; Jiang, D. Effects of Cold and Salicylic Acid Priming on Free Proline and Sucrose Accumulation in Winter Wheat under Freezing Stress. J. Plant Growth Regul. 2022, 41, 2171–2184. [Google Scholar] [CrossRef]
- Cristina, M.S.; Petersen, M.; Mundy, J. Mitogen-Activated Protein Kinase Signaling in Plants. Annu. Rev. Plant Biol. 2010, 61, 621–649. [Google Scholar] [CrossRef] [PubMed]
- de Vries, S.; de Vries, J.; von Dahlen, J.K.; Gould, S.B.; Archibald, J.M.; Rose, L.E.; Slamovits, C.H. On Plant Defense Signaling Networks and Early Land Plant Evolution. Commun. Integr. Biol. 2018, 11, 1–14. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Iqbal, N.; Masood, A.; Per, T.S.; Khan, N.A. Salicylic Acid Alleviates Adverse Effects of Heat Stress on Photosynthesis through Changes in Proline Production and Ethylene Formation. Plant Signal. Behav. 2013, 8, e26374. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Bullock Jr, D.A.; Alonso, J.M.; Stepanova, A.N. To Fight or to Grow: The Balancing Role of Ethylene in Plant Abiotic Stress Responses. Plants 2021, 11, 33. [Google Scholar] [CrossRef]
- Rao, M.V.; Lee, H.; Davis, K.R. Ozone-induced Ethylene Production Is Dependent on Salicylic Acid, and Both Salicylic Acid and Ethylene Act in Concert to Regulate Ozone-induced Cell Death. Plant J. 2002, 32, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, D.; Nakajima, N.; Sano, T.; Tamaoki, M.; Aono, M.; Kubo, A.; Kanna, M.; Ioki, M.; Kamada, H.; Saji, H. Salicylic Acid Accumulation under O3 Exposure Is Regulated by Ethylene in Tobacco Plants. Plant Cell Physiol. 2005, 46, 1062–1072. [Google Scholar] [CrossRef]
- Koo, Y.M.; Heo, A.Y.; Choi, H.W. Salicylic Acid as a Safe Plant Protector and Growth Regulator. Plant Pathol. J. 2020, 36, 1. [Google Scholar] [CrossRef]
- Daw, B.; Zhang, L.; Wang, Z. Salicylic Acid Enhances Antifungal Resistance to Magnaporthe grisea in Rice Plants. Australas. Plant Pathol. 2008, 37, 637–644. [Google Scholar] [CrossRef]
- Esmailzadeh, M.; Soleimani, M.; Rouhani, H. Exogenous Applications of Salicylic Acid for Inducing Systemic Acquired Resistance against Tomato Stem Canker Disease. 2008. [CrossRef]
- Jendoubi, W.; Harbaoui, K.; Hamada, W. Salicylic Acid-Induced Resistance against Fusarium oxysporumf. s. Pradicis lycopercisi in Hydroponic Grown Tomato Plants. J. New Sci. 2015, 21. https://www.jnsciences.org/agri-biotech/29-volume-21.html.
- Kundu, S.; Chakraborty, D.; Pal, A. Proteomic Analysis of Salicylic Acid Induced Resistance to Mungbean Yellow Mosaic India Virus in Vigna Mungo. J. Proteomics 2011, 74, 337–349. [Google Scholar] [CrossRef]
- Le Thanh, T.; Thumanu, K.; Wongkaew, S.; Boonkerd, N.; Teaumroong, N.; Phansak, P.; Buensanteai, N. Salicylic Acid-Induced Accumulation of Biochemical Components Associated with Resistance against Xanthomonas oryzae Pv. oryzae in Rice. J. Plant Interact. 2017, 12, 108–120. [Google Scholar] [CrossRef]
- Mohan Babu, R.; Sajeena, A.; Vijaya Samundeeswari, A.; Sreedhar, A.; Vidhyasekaran, P.; Seetharaman, K.; Reddy, M. Induktion Einer Systemischen Resistenz Gegen Xanthomonas oryzae Pv. oryzae Durch Salicylsäure in Oryza sativa (L.). J. Plant Dis. Prot. 2003, 110, 419–431. [Google Scholar] [CrossRef]
- Radwan, D.E.M.; Fayez, K.A.; Mahmoud, S.Y.; Hamad, A.; Lu, G. Physiological and Metabolic Changes of Cucurbita Pepo Leaves in Response to Zucchini Yellow Mosaic Virus (ZYMV) Infection and Salicylic Acid Treatments. Plant Physiol. Biochem. 2007, 45, 480–489. [Google Scholar] [CrossRef]
- Saikia, R.; Singh, T.; Kumar, R.; Srivastava, J.; Srivastava, A.K.; Singh, K.; Arora, D.K. Role of Salicylic Acid in Systemic Resistance Induced by Pseudomonas Fluorescens against Fusarium oxysporum f. Sp. ciceri in Chickpea. Microbiol. Res. 2003, 158, 203–213. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, J.-H. Exogenous Treatment with Salicylic Acid Attenuates Occurrence of Citrus Canker in Susceptible Navel Orange (Citrus Sinensis Osbeck). J. Plant Physiol. 2012, 169, 1143–1149. [Google Scholar] [CrossRef]
- Wang Jun, W.J.; Chen ShaoHong, C.S.; Huang YongFang, H.Y.; Sun Si, S.S. Induced Resistance to Anthracnose of Camellia oleifera by Salicylic Acid. 2006. https://www.cabidigitallibrary.org/doi/full/10.5555/20073031288.
- Erb, M.; Meldau, S.; Howe, G.A. Role of Phytohormones in Insect-Specific Plant Reactions. Trends Plant Sci. 2012, 17, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Liu, Y.; Li, S.-S.; Han, G.-Z. Insights into the Origin and Evolution of the Plant Hormone Signaling Machinery. Plant Physiol. 2015, 167, 872–886. [Google Scholar] [CrossRef]
- Song, S.; Huang, H.; Gao, H.; Wang, J.; Wu, D.; Liu, X.; Yang, S.; Zhai, Q.; Li, C.; Qi, T. Interaction between MYC2 and ETHYLENE INSENSITIVE3 Modulates Antagonism between Jasmonate and Ethylene Signaling in Arabidopsis. Plant Cell 2014, 26, 263–279. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Lee, B. Friends or Foes: New Insights in Jasmonate and Ethylene Co-Actions. Plant Cell Physiol. 2015, 56, 414–420. [Google Scholar] [CrossRef]
- Wasternack, C.; Song, S. Jasmonates: Biosynthesis, Metabolism, and Signaling by Proteins Activating and Repressing Transcription. J. Exp. Bot. 2017, 68, 1303–1321. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhu, Z.; An, F.; Hao, D.; Li, P.; Song, J.; Yi, C.; Guo, H. Jasmonate-Activated MYC2 Represses ETHYLENE INSENSITIVE3 Activity to Antagonize Ethylene-Promoted Apical Hook Formation in Arabidopsis. Plant Cell 2014, 26, 1105–1117. [Google Scholar] [CrossRef]
- Cheng, M.-C.; Liao, P.-M.; Kuo, W.-W.; Lin, T.-P. The Arabidopsis ETHYLENE RESPONSE FACTOR1 Regulates Abiotic Stress-Responsive Gene Expression by Binding to Different Cis-Acting Elements in Response to Different Stress Signals. Plant Physiol. 2013, 162, 1566–1582. [Google Scholar] [CrossRef]
- Lorenzo, O.; Piqueras, R.; Sánchez-Serrano, J.J.; Solano, R. ETHYLENE RESPONSE FACTOR1 Integrates Signals from Ethylene and Jasmonate Pathways in Plant Defense. Plant Cell 2003, 15, 165–178. [Google Scholar] [CrossRef]
- Zander, M.; Lewsey, M.G.; Clark, N.M.; Yin, L.; Bartlett, A.; Saldierna Guzmán, J.P.; Hann, E.; Langford, A.E.; Jow, B.; Wise, A. Integrated Multi-Omics Framework of the Plant Response to Jasmonic Acid. Nat. Plants 2020, 6, 290–302. [Google Scholar] [CrossRef]
Figure 1.
Model of the chloroplast retrograde signaling pathways that regulate cell death (CD) and integrate immune defenses and acclimation responses in plants – cross-tolerance to biotic and abiotic stresses. Red lighting-like symbol on the top demonstrate absorption of energy in excess (AEE) of that actually required for electrical charge separation in P680. AEE induce overproduction of electrons and protons thus induce non-photochemical quenching (NPQ), heat and electrical signaling waves. Excess of electrons lead to increase reactive oxygen species (ROS) production and to higher reduction of the plastoquinone (PQ) pool. This trigger redox changes in stroma in ascorbate, glutathione and others redox pairs components pools which regulate carbonic anhydrases (CA) activities thus CO2/O2 ratio nearby RuBisCO and sugar production. This in turn trigger phytohormones precursor synthesis and photorespiration as a photosynthetic electron sink which further increase ROS production in peroxisomes and mitochondria and regulate stomatal conductance and programmed cell death (PCD). Chloroplast stroma can be directly connected via stormless with nucleus and ES-, ROS-, and heat- and other-signaling molecules or dual transcription factors can be directly transduced to the nucleus and specifically induce/inhibit nuclear-encoded gene expression and cross-tolerance to abiotic and biotic stresses. This happening in the majority of foliar cells but some cells induce PCD when chloroplast outer membrane is disintegrated due to AEE, heat, SA and ET signaling. Red wavy-like and black lightning-like arrows shows heat and electrical signaling waves respectively. Solid lines shows the confirmed regulatory signaling pathways while dashed lines shows hypothetical signaling pathways. Modified version from Karpiński and colleagues (2014) [11] on the basis of new data [15,16,17,18,19,20,21,22,23,24,41,42,78,89,90,91,92,93,94,95,96,138].
Figure 1.
Model of the chloroplast retrograde signaling pathways that regulate cell death (CD) and integrate immune defenses and acclimation responses in plants – cross-tolerance to biotic and abiotic stresses. Red lighting-like symbol on the top demonstrate absorption of energy in excess (AEE) of that actually required for electrical charge separation in P680. AEE induce overproduction of electrons and protons thus induce non-photochemical quenching (NPQ), heat and electrical signaling waves. Excess of electrons lead to increase reactive oxygen species (ROS) production and to higher reduction of the plastoquinone (PQ) pool. This trigger redox changes in stroma in ascorbate, glutathione and others redox pairs components pools which regulate carbonic anhydrases (CA) activities thus CO2/O2 ratio nearby RuBisCO and sugar production. This in turn trigger phytohormones precursor synthesis and photorespiration as a photosynthetic electron sink which further increase ROS production in peroxisomes and mitochondria and regulate stomatal conductance and programmed cell death (PCD). Chloroplast stroma can be directly connected via stormless with nucleus and ES-, ROS-, and heat- and other-signaling molecules or dual transcription factors can be directly transduced to the nucleus and specifically induce/inhibit nuclear-encoded gene expression and cross-tolerance to abiotic and biotic stresses. This happening in the majority of foliar cells but some cells induce PCD when chloroplast outer membrane is disintegrated due to AEE, heat, SA and ET signaling. Red wavy-like and black lightning-like arrows shows heat and electrical signaling waves respectively. Solid lines shows the confirmed regulatory signaling pathways while dashed lines shows hypothetical signaling pathways. Modified version from Karpiński and colleagues (2014) [11] on the basis of new data [15,16,17,18,19,20,21,22,23,24,41,42,78,89,90,91,92,93,94,95,96,138].
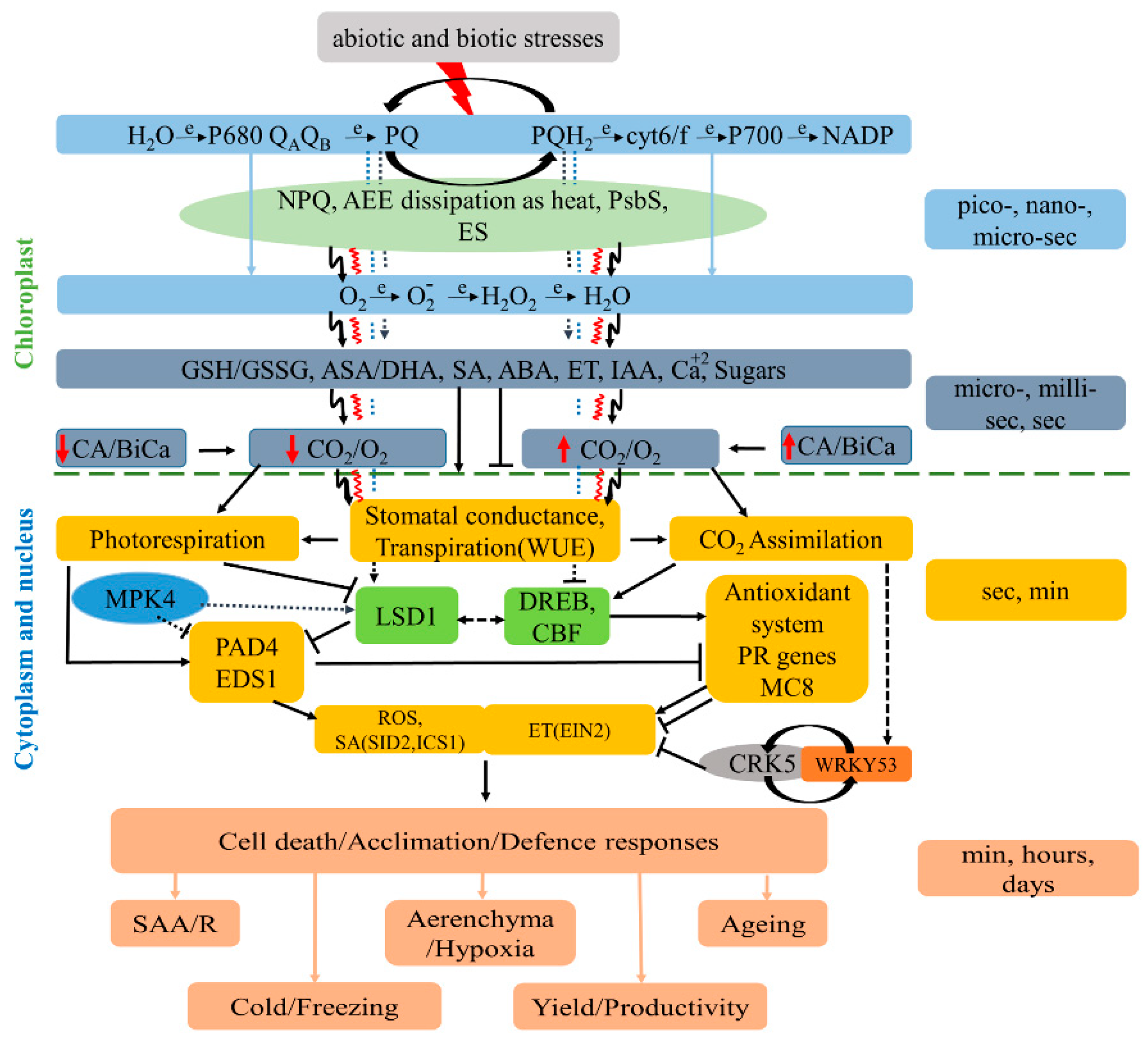
Figure 2.
Systemic and network communication of non-photochemical quenching (NPQ) changes between each photosystem II and mesophyll cells within and between plants, respectively. Electrical signaling (ES) and reactive oxygen species (ROS) waves can induce wavey-like changes in NPQ value within a chloroplast, between all chloroplasts in the cell, in a whole leaf, in all leaves of a plant and within the community-dwelling plants, such as dandelion or Arabidopsis, when leaves of neighboring plants are in physical contact [18,19,20,21,22,23,24]. ES and ROS waves transmitted from stressed cells across the whole plant and between plants regulate NPQ- and plastoqionone pool-dependent chloroplast retrograde signaling to induce systemic acquired acclimation (SAA) in a plant and NAA between plants. Physical contact and electrical conductivity (e.g., a drop of water or high relative humidity) are necessary for NAA. In both transmitter and receiver plants, ES causes spatiotemporal variations in energy quenching (NPQ) (Solid red line), the following induction of the ROS wave (solid green line), and retrograde signaling (RS) (solid blue line). Its variable amplitude is caused by interdependent ion fluxes in the plants. (Model modified from Szechyńska-Hebda et al., 2022 [23]).
Figure 2.
Systemic and network communication of non-photochemical quenching (NPQ) changes between each photosystem II and mesophyll cells within and between plants, respectively. Electrical signaling (ES) and reactive oxygen species (ROS) waves can induce wavey-like changes in NPQ value within a chloroplast, between all chloroplasts in the cell, in a whole leaf, in all leaves of a plant and within the community-dwelling plants, such as dandelion or Arabidopsis, when leaves of neighboring plants are in physical contact [18,19,20,21,22,23,24]. ES and ROS waves transmitted from stressed cells across the whole plant and between plants regulate NPQ- and plastoqionone pool-dependent chloroplast retrograde signaling to induce systemic acquired acclimation (SAA) in a plant and NAA between plants. Physical contact and electrical conductivity (e.g., a drop of water or high relative humidity) are necessary for NAA. In both transmitter and receiver plants, ES causes spatiotemporal variations in energy quenching (NPQ) (Solid red line), the following induction of the ROS wave (solid green line), and retrograde signaling (RS) (solid blue line). Its variable amplitude is caused by interdependent ion fluxes in the plants. (Model modified from Szechyńska-Hebda et al., 2022 [23]).
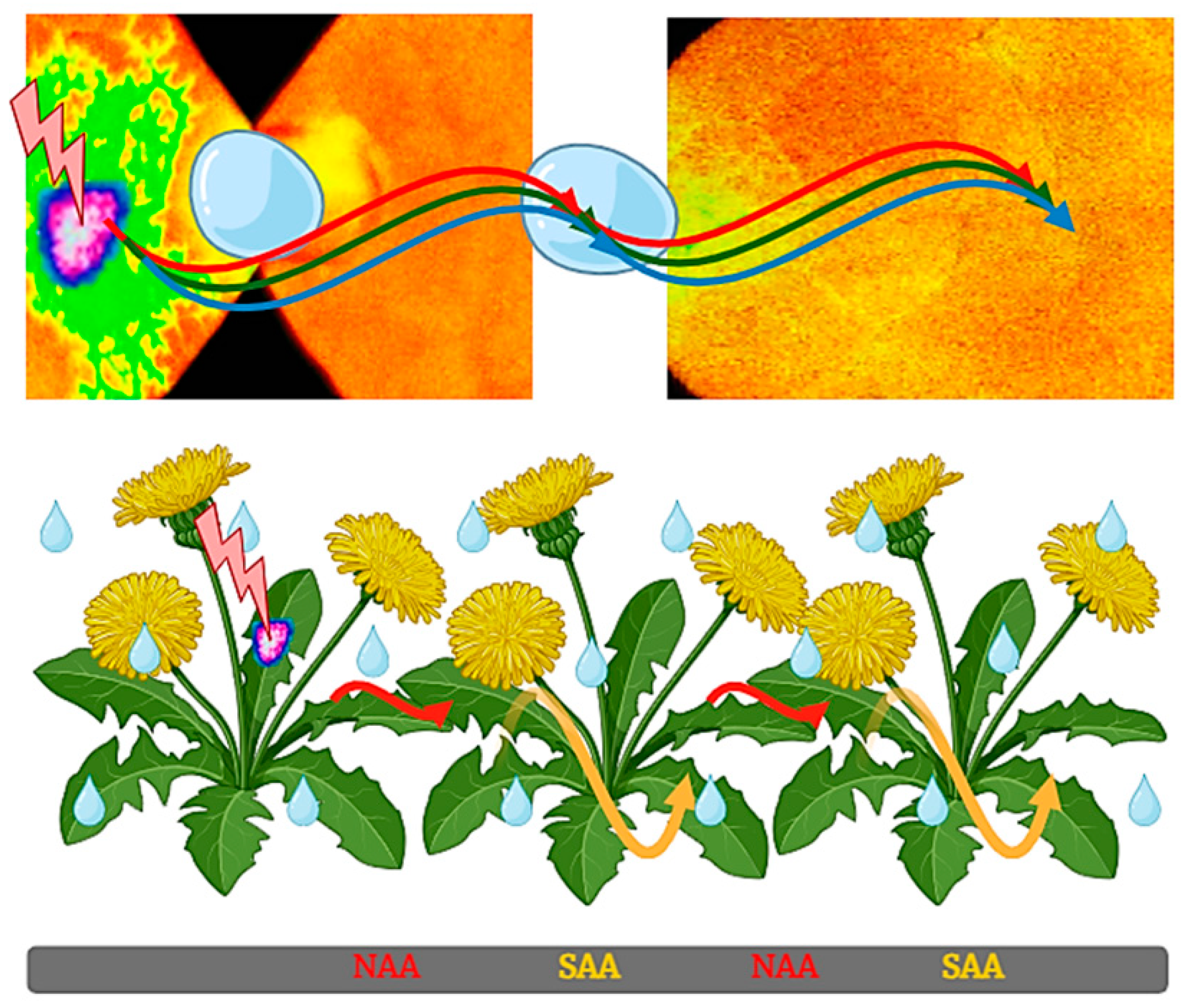
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



