
Submitted:
26 November 2024
Posted:
27 November 2024
You are already at the latest version
Abstract
First believed to be a simple intermediary between the information encoded in deoxyribonucleic acid and that functionally displayed in proteins, ribonucleic acid (RNA) is now known to have many functions through its abundance and intricate, ubiquitous, diverse, and dynamic structure. About 70-90% of the human genome is transcribed into protein-coding and noncoding RNAs as main determinants along with regulatory sequences of cellular to populational biological diversity. From the nucleotide sequence or primary structure, through Watson-Crick pairing self-folding or secondary structure, to compaction via longer distance Watson-Crick and non-Watson-Crick interactions or tertiary structure, and interactions with other biopolymers or quaternary structure, or with metabolites or quinary structure, RNA structure plays a critical role in RNA’s lifecycle from transcription to decay. In contrast to the success with 3-dimensional protein structure prediction using AlphaFold, determining and predicting RNA tertiary and beyond structures remains challenging. However, many approaches have been introduced or are being worked on relying on the use of machine learning and artificial intelligence, sequencing of RNA and its modifications, and structural analyses at the single cell and intact tissue levels, among others. These approaches provide an optimistic outlook for the continued development and refinement of RNA-based applications in medicine, agriculture, and industry.
Keywords:
RNA structure
; Tertiary structure
; Helix structure
1. RNA as an Organic Code
The seven-note musical alphabet played with variations in number, tempo, intensity, rhythm, pitch, or instrument has produced myriads of musical melodies over millennia. Similarly, the organic codes of nucleic acid, protein, polysaccharide, and lipid biomolecules [1-3] use relatively few symbols to generate vast information-storing combinations with context-dependent meanings, underlying the diversity and complexity of Earthly life [3-5]. Underscoring the analogy between the human-made music code and organic codes, sonification tools provide auditory displays of individual and collective biomolecule sequence information as an adjunct to visual and analytical bioinformatics tools [6-8].
The hypothesis of the emergence of overlapping organic codes heralding the living organisms (biotic) era is in line with the intricate interdependence between deoxyribonucleic acid (DNA), ribonucleic acid (RNA), proteins, carbohydrates, lipids, and metabolites in cell- or capsid (viruses)-based organisms [9-12].
The genetic code is the core of life [13], and DNA is its blueprint [14]. The 1800s saw crucial developments in this field. Justus Liebig reported an acidic material in a beef muscle filtrate. Friedrich Miescher discovered ‘nuclein’ in leukocyte nuclei as a protein-degradation-resistant, phosphorus-rich, natural living system’s chemical and chromosome structural component [15]. Richard Altmann coined the term nucleic acid [16].
By 1910, two kinds of nucleic acids were distinguished based on sources and isolation methods: the thymonucleic or zoonucleic acid, now termed DNA, from thymus or animals, and the phytonucleic acid, now termed RNA, from yeast and plants [17,18]. Later, both were found ubiquitously in living organisms. Levene and collaborators [19-21] identified the planar aromatic ring structures of DNA’s constituent purine (adenine [A] and guanine [G]) and pyrimidine (thymine [T] and cytosine [C]) nitrogenous bases, and that RNA has the pyrimidine uracil [U], instead of thymine, and pentose instead of hexose as carbohydrate.
In 1944, Oswald Avery [22] proposed that DNA is the genetic information carrier. In 1950, Erwin Chargaff [23] deciphered the consistent proportions of DNA’s constituent bases and the A-T and C-G base pairing rules. In 1953, Maurice Wilkins [24,25] and Rosalind Franklin with Raymond Gosling [26,27] conducted the X-ray diffraction and crystallography studies that led James Watson and Francis Crick to discover the double-helical structure of DNA in 1953 [28-30] as the foundation for the DNA theory of inheritance [31]. Each nucleotide interacts with water, ions, amino acids, small molecules, and every other nucleotide, stabilizing the structure [32].
The Watson-Crick right-handed helical B-DNA is the native form of DNA in cells. However, DNA’s helical structure and biological properties can vary transiently along short repetitive tracts, as in left-handed Z-DNA [33-36], or reversibly en masse, as in the transition between B- and A-DNA in microorganisms in extreme temperatures and pH [37,38]. Other higher-order variations include supercoils (double helix ends join in bacterial genomes), bubbles, hairpins and cruciforms (when palindromes are present), slipped loops, three-stranded triple helices (H-DNA), and tetrameric i-motifs (over 50,000 in the human genome) and related four-stranded G-quadruplexes [39-41]. Tertiary DNA structures vary from person to person in critical genes like the insulin gene, constituting therapeutic targets [42].
First proposed by Mitsui et al in 1970 [43] and later proven by Wang AH et al [33], Z-DNA is a left-handed helix in equilibrium with the lower energy right-handed B-DNA. Flipons, typically involving an alternating purine/pyrimidine motif, can flip between B- and Z-DNA conformations under physiological conditions aided by binding proteins, introducing diversity to transcriptomes, particularly in immunity and transcription functions [35,36,44].
Discovered by Franklin and Gosling in 1953 [45] in DNA crystals after dehydration, A-DNA, also derived from protein binding to DNA, is a right-handed double helix but with a shorter and more compact helical structure than B-DNA, resulting in slightly more base pairs per turn, a smaller twist angle, and a shorter rise per base pair. The major groove of A-DNA is deep and narrow, the minor groove is wide and shallow, and the base pairs are not perpendicular to the helix-axis as in B-DNA. A-DNA can occur in DNA-RNA hybrid double helices and double-stranded RNAs. RNA can only form an A-type double helix because of the steric restrictions imposed on ribose by the 2’ hydroxyl residue [40].
After Z-DNA was discovered and named after its sugar-phosphate backbone’s zig-zag course as an alternative to the more common Watson-Crick B-DNA, nuclear magnetic resonance, and other studies showed that the common A-RNAs, particularly those with higher Guanine/Cytosine content, could similarly undergo the right-to-left-handed conformational change to the higher energy Z-RNA [44,46-48]. Z-binding proteins specifically recognize and bind Z-DNA [49-51] and Z-RNA [52,53]. Z-DNA and Z-RNA encoded by flipons under physiological conditions are implicated in various biological processes, including transcription and immunity [44]. Z-RNA has been studied less than Z-DNA and both are challenging to detect in vivo.
Structural similarities between RNA and DNA allow the formation of RNA-DNA hybrids, such as the R loops, which also include a displaced single-stranded DNA [54]. Antisense noncoding RNAs may form R lops. R loops accumulate throughout the genome in pericentromeric DNA, telomeres, ribosomal DNA, or transcription termination regions, among others, and are involved in transcription and chromatin structure. Because they can also adversely affect genome stability and replication, several DNA and RNA metabolism factors, such as ribonucleases, RNA-DNA helicases, RNA processing factors, and topoisomerase I, degrade R-loops or prevent their formation [54].
As an example of a protein that interacts with DNA and RNA, the topoisomerase I enzyme prevents genomic instability by alleviating DNA torsional strain. Topoisomerase I introduces transient single-strand breaks that prevent the accumulation of supercoiling and torsional stress, which could otherwise lead to damage and instability of DNA, and cell death [55]. Interactions between RNA and Topoisomerase I regulate DNA during transcription by modulating Topoisomerase I-mediated relaxation. In cancer cells, for instance, DNA transcription is often elevated, necessitating increased levels of Topoisomerase I activity to relax the DNA and maintain proper gene expression. RNA opposes Topoisomerase I activity. Inhibiting RNA binding of Topoisomerase I may work similarly to antineoplastic Topoisomerase I inhibitors like camptothecin by increasing Topoisomerase I catalytic complexes on DNA [55].
Beyond cancer, dysfunction of R loop-interacting factors in several genetic diseases leads to replication stress, genome instability, chromatin alterations, or gene silencing [54]. Furthermore, many chromatin-associated complexes, including histone modifiers, transcription factors, and DNA methyltransferase, interact with RNA [56]. RNA can also promote the repair of double-strand breaks in DNA, by helping position and holding the broken DNA ends in place and guiding the cellular repair machinery, thereby contributing to genome integrity [57].
2. RNA Has Many Functions Through Its Intricate, Ubiquitous, Diverse, And Dynamic Structure
RNA has emerged as a central biomolecule in the multidirectional flow of genetic information for phenotype and biological diversity generation [12,58]. RNA is no longer considered simply an intermediary between the data stored in DNA and that functionally displayed in proteins. Although about 70–90% of the human [59] and 85–90% of the yeast genome [60] is transcribed into RNA, much remains unknown about RNA functions in cells [61].
RNA structure plays critical roles in every step of RNA’s lifecycle, including transcription, splicing, localization [62,63], translation [64,65], and RNA decay [66]. However, RNA structure differs among individual cells and provides an additional layer of information in defining cellular identities by, for instance, informing RNA-binding protein binding and gene regulation [67]. To this end, overall RNA structure profiles better discriminate cell type identity and differentiation stage than gene expression profiles alone. For instance, RNA structure is more homogeneous in human embryonic stem cells than differentiating neurons, with the greatest homogeneity found in coding regions. More extensive heterogeneity is found within 3’ untranslated regions and is determined by specific RNA-binding proteins. Moreover, the cell-type variable region of 18S ribosomal RNA is associated with cell cycle and translation control. It is therefore important to systematically characterize RNA structure-function relationships at single-cell resolution using approaches such as single-cell structure probing of RNA transcripts [67].
RNA accomplishes its many functions through various structural levels beyond its primary and secondary structural ones, which are defined by nucleotide sequence and Watson-Crick pairing-based folding, respectively [68-71]. Along with the staggering number of noncoding RNA genes, RNA structural versatility underlies biological diversity from the organismal to population levels.
RNA’s bases closely stack on each other like ‘coins in a roll’ via noncovalent interactions, exposing their charged exocyclic groups to water molecules and ions, underlying RNA’s solvability and helical conformations unrelated to Watson-Crick pairing [72]. RNA’s conformation also varies with environmental changes, liquid-liquid phase separation, or interactions with other biomolecules[73-75].
Underlying its compactness, RNA intrinsically tends to form A-U, G-C, and G-U Watson-Crick base pairs in short and long-range structures, higher-order architectures, and RNA-RNA interactions in picoseconds to seconds [76-79], which are fundamental to its diverse functions [80]. As many as 40% of the nucleotides of an RNA molecule can be part of hairpins and multi-helix junction loops [81], and 30% to 40% of RNA duplexes in living cells involve sequences over 200 nucleotides apart [80].
3. RNA's Structure Is Defined at Primary, Secondary, Tertiary, Quaternary, And Quinary Levels
3.1. Primary, Secondary, and Tertiary RNA STRUCTURES
The primary structure is the RNA’s linear nucleotide sequence (Figure 1). The secondary structure describes the paired and unpaired elements of stems, loops, and bulges that form as the single-stranded RNA molecule folds back on itself via Watson-Crick pairs and interacts via hydrogen bonding and stacking as soon as it is synthesized [74,78] (Figure 1).
The tertiary or 3D structure, which typically compacts the RNA, is achieved by longer-distance Watson-Crick and non-Watson-Crick interactions of elements within the preformed secondary structures [70] (Figure 1). These interactions give rise to structural elements, including pseudoknots, which lock together two stem-loops by base pairing and sugar-phosphate interactions, often in a so-called kissing interaction.
Many RNA loops are characterized by structural modules with highly organized networks of noncanonical interactions comprising ordered non-Watson-Crick base pairs embedded between Watson-Crick base pairs [82]. Non-Watson–Crick pairs are key for folding and binding to proteins or other ligands [83-85].
RNA three-dimensional (3D) motifs occupy places in structured RNA molecules corresponding to the hairpin, internal, and multi-helix junction loops of their 2D structure representations [81]. These 3D structural RNA modules, with specific loop geometries, contribute to structural stability, have central roles as architectural organizers of catalytic activity and ligand binding sites in RNA molecules, and are recurrently observed in RNA families throughout phylogeny [81,86-91].
Among RNA 3D motifs are pseudoknots, which are minimally composed of two helical segments connected by single-stranded regions or loops (Figure 2). Pseudoknots form the catalytic core of various ribozymes, self-splicing introns, and telomerase, and alter gene expression by inducing ribosomal frameshifting in many viruses (reviewed in [92]). The best characterized is the H-type pseudoknot (Figure 2).
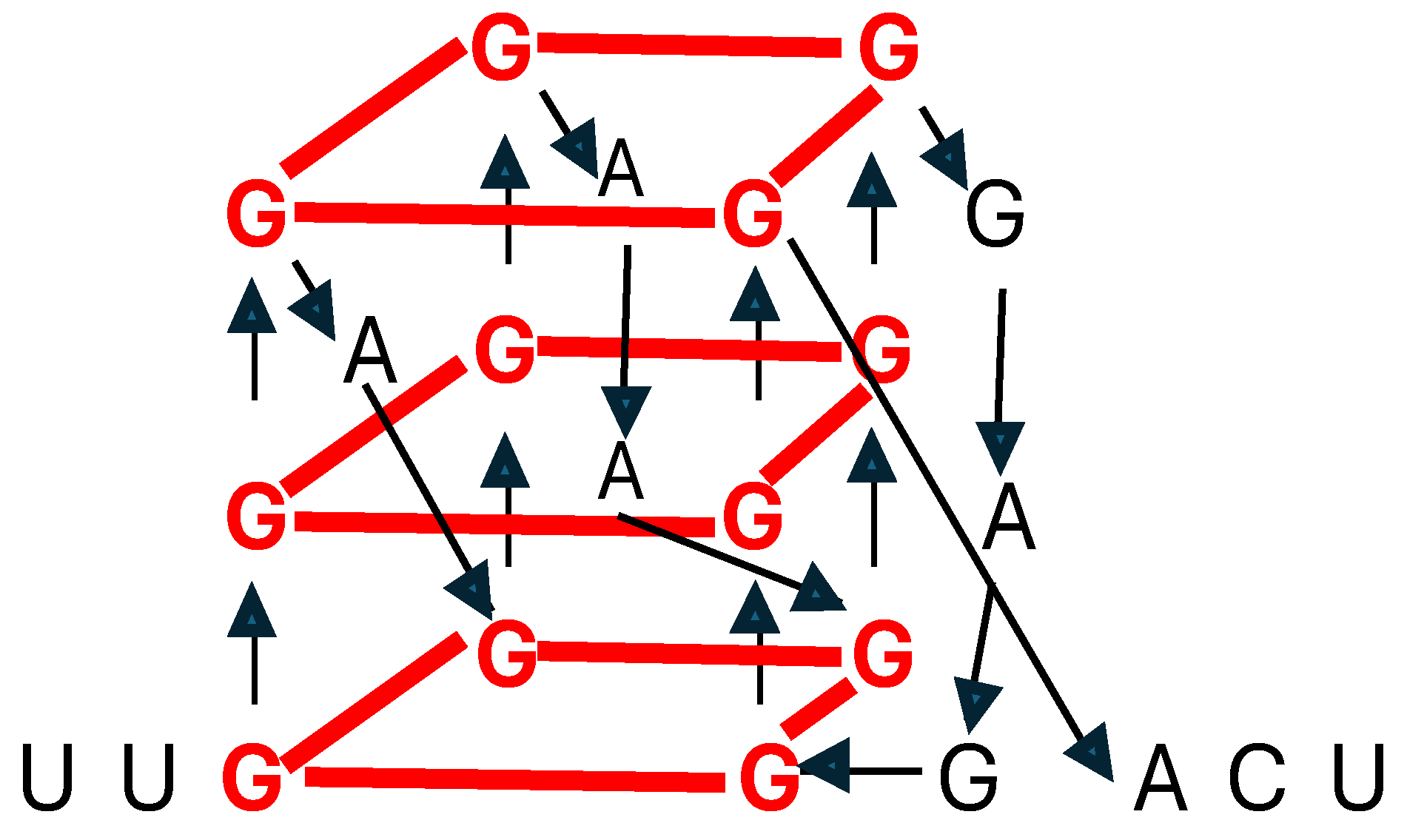
Guanine-rich regions in RNA and DNA can form noncanonical G-quadruplex structures encompassing stacked guanine tetrads, a square planar structure formed by four guanine residues [93] (Figure 3). RNA G-quadruplexes participate in translation, splicing, RNA stability, and cellular stress responses, among other functions mediated by the RNA binding proteins with which they interact [93].
3.2. Quaternary and Quinary RNA Structures
Similar to the interactions of DNA and histones, RNA’s quaternary structures result from a folded RNA's interaction with other biopolymers, such as proteins and RNAs. The quinary structure of RNA results from its weak and nonspecific interaction with cellular metabolites, such as osmolytes, accumulated by cells in response to osmotic stress [75]. Understanding the effects of osmolytes on RNA's tertiary structure, whether stabilizing or destabilizing, is crucial to comprehend the intricacies of RNA [94]. For instance, as hydrated magnesium ions neutralize a notable fraction of the negative charge of an RNA tertiary structure, the RNA becomes less responsive to stabilizing osmolytes and may even be destabilized [94].
4. Determining RNA Tertiary and Beyond Structures Remains Challenging
The complex biological functions of RNA molecules are underpinned by their specific sustained 3D structures, with or without the help of proteins or other RNAs in multimolecular complexes [95]. However, the study of RNA 3D structure is often hindered by the scarcity of atomic coordinates, a significant challenge in the field. These determinations are typically low-resolution or miss atoms due to the limitations of the low-throughput and costly structure determination methods, i.e., X-ray crystallography, nuclear magnetic resonance, and cryo-electron microscopy [96], which also creates a significant gap between the number of RNAs sequenced and the number of structures defined. Moreover, RNA’s shifting into diverse forms according to environmental conditions renders structural studies challenging. Traditional imaging methods, such as cryo-electron microscopy single-particle averaging analysis, rely on averaging data from thousands of selected molecules with common shapes, making it difficult to capture the unique shapes of individual RNA molecules.
Developed during the last two decades [97], some RNA 3D structure prediction computational tools use high-resolution homologs' more precise structural information to annotate the base-pairing interactions in low-resolution structures in coarse-grained models/simulations [98,99] or in imaging data missing atoms [100]. Moreover, a machine-learning approach identifies accurate structural models without assumptions about their defining characteristics despite being trained with the atomic coordinates of only 18 known RNA structures [101].
Structural imaging studies are complemented by gel or capillary electrophoresis based on in-line probing, i.e., structural sensitivity to spontaneous degradation, nucleases targeting either single- or double-stranded regions, or chemical probes, such as dimethyl sulfoxide (DMS). DMS, for instance, is used to probe unpaired adenines and cytidines, and 1- metho-p-toluenesulfonate (CMCT) to probe unpaired uridines in chemical inference of RNA structures sequencing (CRIS-seq) [102-104]. DMS is also used in RNA structure sequencing (Structure-seq and STRucture-seq2) [105-108], dimethyl sulfate-modified RNA sequencing (DMS-seq) [109], dimethyl sulfate mutational profiling with sequencing (DMS-MaPseq) [110], and transfer RNA structure sequencing (tRNA structure-seq) [111]. Pyrdiostatin, the chemical probe in RNA GQ sequencing (rG4-seq) [112], and selective 2’-hydroxyl acylation analyzed by primer extension (SHAPE) [113-118] have also extended chemical probing to the entire transcriptome [70,119-122].
Even if RNA structures are accurately determined, they may not represent the one(s) relevant in vivo. Many factors influence RNA structure in the living cell, including variations in organelle environments and interactions with proteins or other macromolecules, which render the elucidation of RNA structure in vivo particularly challenging. For instance, in silico modeling provides the most thermodynamically stable structure of an RNA sequence, while RNAs can become trapped in vivo in alternative structures [70,123]. Moreover, processing the low abundance, long nascent, or precursor RNAs, including splicing and polyadenylation, entails pathway networks that determine mature isoform composition and control gene expression, further adding to the complexity of studying RNA structure [124].
The success of Alphafold [125,126] in predicting protein 3D structures has not yet extended to RNA [127]. This is due to differences in building blocks (amino acids vs. nucleotides), diversity of sequence range (up to tens of thousands of nucleotides for RNA vs. a few hundred amino acids for proteins), number of available structure data (orders of magnitude greater for proteins), and folding stability (multiple conformations for RNA vs. usually one for proteins) [97].
Readily available RNA 3D structural prediction tools often rely on the primary sequence and canonical 2D structures formed by A-U, G-C, and G-U Watson-Crick pairs to detect structural RNA modules from primary sequence data and identify recurrent interaction networks [128-133]. Several databases contain RNA structural information [91,134-136].
During the last decade, computational RNA structure predictions have evolved from the earliest thermodynamic and molecular dynamic-based approaches to deep learning-based conformation approaches [137]. Earlier deep learning models for RNA structure have been competitive but not consistently better than traditional 3D structure prediction methods, including ab initio physics-based methods using various levels of granularity in nucleotide representation, template-based methods that try to map sequences to structural motifs before merging them into a whole structure, or hybrid methods, combining ab initio and template-based methods [97,138]. However, platforms such as the RNA3DB dataset [139], which arranges the RNA 3D chains into distinct non-redundant groups (Components), and Dfold, which combines an autoregressive Deep Generative Model, Monte Carlo Tree Search, and a scoring model [140] have been developed to improve RNA 3D structure prediction.
An innovative technique to study the 3D structure of individual molecules without averaging builds on advanced Individual-Particle cryo-Electron Tomography (IPET) to focus on single-molecule 3D imaging in cryopreserved samples. IPET captures a snapshot of RNA's folding landscape by capturing molecules in various stages of folding, from immature states to their optimal shape. This approach may allow engineering the folding of more effective RNA vaccines and dynamic sensors for molecular medicine [141].
5. Conclusions
Since the central molecular biology dogma was formulated almost seven decades ago, mounting evidence revealed that RNA is the main determinant of biological diversity. This is driven by RNA’s abundance, modifications, and structural versatility of its coding and noncoding versions, which occasionally overlap. RNA structure-function relationships also vary among cells in an organism, determining cellular identity. However, understanding RNA’s tertiary, quaternary, and quinary structures and their functional relationships, remains challenging. Advances in approaches including the use of machine learning and artificial intelligence [101] and data gatherings, such as the United States National Institutes of Health’s and National Academies of Sciences’ RNA sequencing initiative, provide an optimistic outlook for the continued development and refinement of disruptive RNA-based approaches for medical therapy, diagnosis, and prevention, and agriculture and industrial applications [142]. To this end, novel imaging methods enable detailed RNA analysis within single cells and intact tissues [143].
Author Contributions
Both authors (W.A.H. and R.P.) contributed to writing—original draft preparation, writing—review, and editing. Both authors have read and agreed to the published version of the manuscript.
Funding
This review received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Maraldi, N.M. In search of a primitive signaling code. Biosystems 2019, 183, 103984. [Google Scholar] [CrossRef] [PubMed]
- Melero, A. , Jiménez-Rojo, N. Cracking the membrane lipid code. Curr Opin Cell Biol 2023, 83, 102203. [Google Scholar] [CrossRef] [PubMed]
- Gabius, H.J. , Cudic, M., Diercks, T., Kaltner, H., Kopitz, J., Mayo, K.H., Murphy, P.V., Oscarson, S., Roy, R., Schedlbauer, A., Toegel, S., Romero, A. What is the Sugar Code? Chembiochem 2022, 23, e202100327. [Google Scholar] [CrossRef]
- Chatterjee, S. , Yadav, S. The Origin of Prebiotic Information System in the Peptide/RNA World: A Simulation Model of the Evolution of Translation and the Genetic Code. Life (Basel) 2019, 9, 25. [Google Scholar] [CrossRef]
- Chatterjee, S. , Yadav, S. The Coevolution of Biomolecules and Prebiotic Information Systems in the Origin of Life: A Visualization Model for Assembling the First Gene. Life (Basel) 2022, 12, 834. [Google Scholar] [CrossRef]
- Riego, E. , Silva, A., De la Fuente, J. The sound of the DNA language. Biol Res 1995, 28, 197–204. [Google Scholar]
- Sánchez Sousa, A. , Baquero, F., Nombela, C. The making of "The Genoma Music". Rev Iberoam Micol 2005, 22, 242–248. [Google Scholar] [CrossRef]
- Temple, M.D. An auditory display tool for DNA sequence analysis. BMC Bioinformatics 2017, 18, 221. [Google Scholar] [CrossRef]
- de Farias, S.T. , Prosdocimi, F., Caponi, G. Organic Codes: A Unifying Concept for Life. Acta Biotheor 2021, 69, 769–782. [Google Scholar] [CrossRef]
- Kondratyeva, L.G. , Dyachkova, M.S., Galchenko, A.V. The Origin of Genetic Code and Translation in the Framework of Current Concepts on the Origin of Life. Biochemistry (Mosc) 2022, 87, 150–169. [Google Scholar] [CrossRef]
- Pavlinova, P. , Lambert, C.N., Malaterre, C., Nghe, P. Abiogenesis through gradual evolution of autocatalysis into template-based replication. FEBS Lett 2023, 597, 344–379. [Google Scholar] [CrossRef] [PubMed]
- Haseltine, W.A. , Patarca, R. The RNA revolution in the central molecular biology dogma evolution. Preprints 2024, 2024110983. [Google Scholar] [CrossRef]
- Crick, F.H. , Barnett, L., Brenner, S., Watts-Tobin, R.J. General nature of the genetic code for proteins. Nature 1961, 192, 1227–1232. [Google Scholar] [CrossRef] [PubMed]
- Portin, P. The birth and development of the DNA theory of inheritance: sixty years since the discovery of the structure of DNA. J Genet 2014, 93, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Dahm, R. Friedrich Miescher and the discovery of DNA. Dev Biol 2005, 278, 274–288. [Google Scholar] [CrossRef]
- Altmann, R. Ueber nucleinsäuren. Arch. f. Anat. u. Physiol. Physiol. Abt. 1889, 524–536.Smýkal, P., K Varshney, R., K Singh, V., Coyne, C.J., Domoney, C., Kejnovský, E., Warkentin, T. From Mendel's discovery on pea to today's plant genetics and breeding : Commemorating the 150th anniversary of the reading of Mendel's discovery. Theor Appl Genet 2016, 129, 2267–2280. [Google Scholar] [CrossRef]
- Levene, P.A. On the biochemistry of nucleic acids. J Am Chem Soc 1910, 32, 231–240. [Google Scholar] [CrossRef]
- Frixione, E. , Ruiz-Zamarripa, L. The "scientific catastrophe" in nucleic acids research that boosted molecular biology. J Biol Chem 2019, 294, 2249–2255. [Google Scholar] [CrossRef]
- Levene, P.A., and Bass, L.W. (1931) Nucleic Acids, Chemical Catalog Company, New York: Available online at: https://babel.hathitrust.org/cgi/pt?id=uc1.b4165245;view=1up;seq=5 (Accessed August 8, 2024).
- Levene, P.A. , Tipson, R.S. The ring structure of adenosine. Science 1931, 74, 521. [Google Scholar] [CrossRef]
- Levene, P.A. , Tipson, R.S. The ring structure of thymidine. Science 1935, 81, 98. [Google Scholar] [CrossRef]
- Avery, O.T. , Macleod, C.M., McCarty, M. Studies on the chemical nature of the substance inducing transformation of Pneumococcal types: Induction of transformation by a desoxyribonucleic acid fraction isolated from Pneumococcus type III. J Exp Med 1944, 79, 137–58. [Google Scholar] [CrossRef] [PubMed]
- Chargaff, E. Chemical specificity of nucleic acids and mechanism of their enzymatic degradation. Experientia 1950, 6, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, M.H. , Stokes, A.R., Wilson, H.R. Molecular structure of deoxypentose nucleic acids. Nature 1953, 171, 738–740. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, M.H. , Stokes, A.R., Wilson, H.R. Molecular structure of nucleic acids. Molecular structure of deoxypentose nucleic acids. Ann N Y Acad Sci 1995, 758, 13–16. [Google Scholar] [CrossRef]
- Franklin, R.E. , Gosling, R.G. Molecular configuration in sodium thymonucleate. Nature 1953, 171, 740–741. [Google Scholar] [CrossRef]
- Franklin, R.E. , Gosling, R.G. Evidence for 2-chain helix in crystalline structure of sodium deoxyribonucleate. Nature 1953, 172, 156–157. [Google Scholar] [CrossRef]
- Watson, J.D. , Crick, F.H. Molecular structure of nucleic acids; a structure for deoxyribose nucleic acid. Nature 1953, 171, 737–738. [Google Scholar] [CrossRef]
- Watson, J.D. , Crick, F.H. The structure of DNA. Cold Spring Harb Symp Quant Biol 1953, 18, 123–131. [Google Scholar] [CrossRef]
- Watson, J.D. , 1928-. The Double Helix: a Personal Account of the Discovery of the Structure of DNA. London: Weidenfield and Nicolson, 1981.
- Watson, J.D. , Crick, F.H. Genetical implications of the structure of deoxyribonucleic acid. Nature 1953, 171, 964–967. [Google Scholar] [CrossRef]
- Martel, P. Base crystallization and base stacking in water. Eur J Biochem 1979, 96, 213–219. [Google Scholar] [CrossRef]
- Wang, A.H. , Quigley, G.J., Kolpak, F.J., Crawford, J.L., van Boom, J.H., van der Marel, G., Rich, A. Molecular structure of a left-handed double helical DNA fragment at atomic resolution. Nature 1979, 282, 680–686. [Google Scholar] [CrossRef] [PubMed]
- Wing, R. , Drew, H., Takano, T., Broka, C., Tanaka, S., Itakura, K., Dickerson, R.E. Crystal structure analysis of a complete turn of B-DNA. Nature 1980, 287, 755–758. [Google Scholar] [CrossRef] [PubMed]
- Herbert, A. A Genetic Instruction Code Based on DNA Conformation. Trends Genet 2019, 35, 887–890. [Google Scholar] [CrossRef] [PubMed]
- Herbert, A. The ancient Z-DNA and Z-RNA specific Zα fold has evolved modern roles in immunity and transcription through the natural selection of flipons. R Soc Open Sci 2024, 11, 240080. [Google Scholar] [CrossRef]
- Whelan, D.R. , Hiscox, T.J., Rood, J.I., Bambery, K.R., McNaughton, D., Wood, B.R. Detection of an en masse and reversible B- to A-DNA conformational transition in prokaryotes in response to desiccation. J R Soc Interface 2014, 11, 20140454. [Google Scholar] [CrossRef]
- DiMaio, F. , Yu, X., Rensen, E., Krupovic, M., Prangishvili, D., Egelman, E.H. Virology. A virus that infects a hyperthermophile encapsidates A-form DNA. Science 2015, 348, 914–917. [Google Scholar] [CrossRef]
- Choi, J. , Majima, T. Conformational changes of non-B DNA. Chem Soc Rev 2011, 40, 5893–5909. [Google Scholar] [CrossRef]
- Travers, A. , Muskhelishvili, G. DNA structure and function. FEBS J 2015, 282, 2279–2295. [Google Scholar] [CrossRef]
- Peña Martinez, C.D., Zeraati, M., Rouet, R., Mazigi, O., Henry, J.Y., Gloss, B., Kretzmann, J.A., Evans, C.W., Ruggiero, E., Zanin, I., Marušič, M., Plavec, J., Richter, S.N., Bryan, T.M., Smith, N.M., Dinger, M.E., Kummerfeld, S., Christ, D. Human genomic DNA is widely interspersed with i-motif structures. The EMBO J 2024. [CrossRef]
- Guneri, D. , Alexandrou, E., El Omari, K., Dvořáková, Z., Chikhale, R.V., Pike, D.T.S., Waudby, C.A., Morris, C.J., Haider, S., Parkinson, G.N., Waller, Z.A.E. Structural insights into i-motif DNA structures in sequences from the insulin-linked polymorphic region. Nat Commun 2024, 15, 7119. [Google Scholar] [CrossRef]
- Mitsui, Y. , Langridge, R., Shortle, B.E., Cantor, C.R., Grant, R.C., Kodama, M., Wells, R.D. "Physical and enzymatic studies on poly d(I–C)·poly d(I–C), an unusual double-helical DNA". Nature 1970, 228, 1166–1169. [Google Scholar] [CrossRef]
- Krall, J.B. , Nichols, P.J., Henen, M.A., Vicens, Q., Vögeli, B. Structure and Formation of Z-DNA and Z-RNA. Molecules 2023, 28, 843. [Google Scholar] [CrossRef] [PubMed]
- Franklin, R.E. , Gosling, R.G. The structure of sodium thermonucleate fibres. I. The influence of water content. Acta Cryst 1953, 6, 673–377. [Google Scholar] [CrossRef]
- Hall, K. , Cruz, P., Tinoco, I. Jr., Jovin, T.M., van de Sande, J.H. 'Z-RNA'--a left-handed RNA double helix. Nature 1984, 311, 584–586. [Google Scholar] [CrossRef] [PubMed]
- Davis, P.W. , Adamiak, R.W., Tinoco, I. Jr. Z-RNA: the solution NMR structure of r(CGCGCG). Biopolymers 1990, 29, 109–122. [Google Scholar] [CrossRef]
- Popenda, M. , Milecki, J., Adamiak, R.W. High salt solution structure of a left-handed RNA double helix. Nucleic Acids Res 2004, 32, 4044–4054. [Google Scholar] [CrossRef]
- Herbert, A. , Alfken, J., Kim, Y.G., Mian, I.S., Nishikura, K., Rich, A. A Z-DNA binding domain present in the human editing enzyme, double-stranded RNA adenosine deaminase. Proc Natl Acad Sci U S A 1997, 94, 8421–8426. [Google Scholar] [CrossRef]
- Kim, Y.G. , Lowenhaupt, K., Oh, D.B., Kim, K.K., Rich, A. Evidence that vaccinia virulence factor E3L binds to Z-DNA in vivo: Implications for development of a therapy for poxvirus infection. Proc Natl Acad Sci U S A 2004, 101, 1514–1518. [Google Scholar] [CrossRef]
- Schwartz, T. , Rould, M.A., Lowenhaupt, K., Herbert, A., Rich, A. Crystal structure of the Zalpha domain of the human editing enzyme ADAR1 bound to left-handed Z-DNA. Science 1999, 284, 1841–1845. [Google Scholar] [CrossRef]
- Brown, B.A. 2nd, Lowenhaupt, K., Wilbert, C.M., Hanlon, E.B., Rich, A. The zalpha domain of the editing enzyme dsRNA adenosine deaminase binds left-handed Z-RNA as well as Z-DNA. Proc Natl Acad Sci U S A 2000, 97, 13532–13536. [Google Scholar] [CrossRef]
- Placido, D. , Brown, B.A. 2nd, Lowenhaupt, K., Rich, A., Athanasiadis, A. A left-handed RNA double helix bound by the Z alpha domain of the RNA-editing enzyme ADAR1. Structure 2007, 15, 395–404. [Google Scholar] [CrossRef]
- Santos-Pereira, J.M. , Aguilera, A. R loops: new modulators of genome dynamics and function. Nat Rev Genet 2015, 16, 583–597. [Google Scholar] [CrossRef] [PubMed]
- Bhola, M. , Abe, K., Orozco, P., Rahnamoun, H., Avila-Lopez, P., Taylor, E., Muhammad, N., Liu, B., Patel, P., Marko, J.F., Starner, A.C., He, C., Van Nostrand, E.L., Mondragón, A., Lauberth, S.M. RNA interacts with topoisomerase I to adjust DNA topology. Mol Cell 2024, 84, 3192–3208.e11. [Google Scholar] [CrossRef] [PubMed]
- Song, J. , Gooding, A.R., Hemphill, W.O., Love, B.D., Robertson, A., Yao, L., Zon, L.I., North, T.E., Kasinath, V., Cech, T.R. Structural basis for inactivation of PRC2 by G-quadruplex RNA. Science 2023, 381, 1331–1337. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y. , Lu, Y., Ferrari, M.M., Channagiri, T., Xu, P., Meers, C., Zhang, Y., Balachander, S., Park, V.S., Marsili, S., Pursell, Z.F., Jonoska, N., Storici, F. RNA-mediated double-strand break repair by end-joining mechanisms. Nat Commun 2024, 15, 7935. [Google Scholar] [CrossRef]
- Jones, C.H. , Androsavich, J.R., So, N., Jenkins, M.P., MacCormack, D., Prigodich, A., Welch, V., True, J.M., Dolsten, M. Breaking the mold with RNA-a "RNAissance" of life science. NPJ Genom Med 2024, 9, 2. [Google Scholar] [CrossRef]
- Pertea, M. The human transcriptome: an unfinished story. Genes (Basel) 2012, 3, 344–360. [Google Scholar] [CrossRef]
- David, L. , Huber, W., Granovskaia, M., Toedling, J., Palm, C.J., Bofkin, L., Jones, T., Davis, R.W., Steinmetz, L.M. A high-resolution map of transcription in the yeast genome. Proc Natl Acad Sci U S A 2006, 103, 5320–5325. [Google Scholar] [CrossRef]
- Kazimierczyk, M. , Kasprowicz, M.K., Kasprzyk, M.E., Wrzesinski, J. Human Long Noncoding RNA Interactome: Detection, Characterization and Function. Int J Mol Sci 2020, 21, 1027. [Google Scholar] [CrossRef]
- Martin, K.C. , Ephrussi, A. mRNA localization: gene expression in the spatial dimension. Cell 2009, 136, 719–730. [Google Scholar] [CrossRef]
- Wan, Y. , Kertesz, M., Spitale, R.C., Segal, E., Chang, H.Y. Understanding the transcriptome through RNA structure. Nat Rev Genet 2011, 12, 641–655. [Google Scholar] [CrossRef]
- Mao, Y. , Liu, H., Liu, Y., Tao, S. Deciphering the rules by which dynamics of mRNA secondary structure affect translation efficiency in Saccharomyces cerevisiae. Nucleic Acids Res 2014, 42, 4813–4822. [Google Scholar] [CrossRef] [PubMed]
- Clancy, S., Brown, W. Translation: DNA to mRNA to protein. Nat Educ 2008, 1, 101.
- Garneau, N.L. , Wilusz, J., Wilusz, C.J. The highways and byways of mRNA decay. Nat Rev Mol Cell Biol 2007, 8, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Wang, J., Zhang, Y., Zhang, T., Tan, W.T., Lambert, F., Darmawan, J., Huber, R., Wan, Y. RNA structure profiling at single-cell resolution reveals new determinants of cell identity. Nat Methods 2024, 21, 411-422. [CrossRef]
- Tomezsko, P.J. , Corbin, V.D.A., Gupta, P., Swaminathan, H., Glasgow, M., Persad, S., Edwards, M.D., McIntosh, L., Papenfuss, A.T., Emery, A, et al.. Determination of RNA structural diversity and its role in HIV-1 RNA splicing. Nature 2020, 582, 438–442. [Google Scholar] [CrossRef] [PubMed]
- Vicens, Q. , Kieft, J.S. Thoughts on how to think (and talk) about RNA structure. Proc Natl Acad Sci U S A 2022, 119, e2112677119. [Google Scholar] [CrossRef] [PubMed]
- Assmann, S.M. , Chou, H.L., Bevilacqua, P.C. Rock, scissors, paper: How RNA structure informs function. Plant Cell 2023, 35, 1671–1707. [Google Scholar] [CrossRef]
- Shine, M. , Gordon, J., Schärfen, L., Zigackova, D., Herzel, L., Neugebauer, K.M. Co-transcriptional gene regulation in eukaryotes and prokaryotes. Nat Rev Mol Cell Biol 2024, 25, 534–554. [Google Scholar] [CrossRef]
- Westhof, E. , Fritsch, V. RNA folding: beyond Watson-Crick pairs. Structure 2000, 8, R55–65. [Google Scholar] [CrossRef]
- Cruz, J.A. , Westhof, E. The dynamic landscapes of RNA architecture. Cell 2009, 136, 604–609. [Google Scholar] [CrossRef]
- Ganser, L.R. , Kelly, M.L., Herschlag, D., Al-Hashimi, H.M. The roles of structural dynamics in the cellular functions of RNAs. Nat Rev Mol Cell Biol 2019, 20, 474–489. [Google Scholar] [CrossRef]
- Alfano, C. , Fichou, Y., Huber, K., Weiss, M., Spruijt, E., Ebbinghaus, S., De Luca, G., Morando, M.A., Vetri, V., Temussi, P.A., Pastore, A. Molecular Crowding: The History and Development of a Scientific Paradigm. Chem Rev 2024, 124, 3186–3219. [Google Scholar] [CrossRef]
- Schultes, E.A. , Spasic, A., Mohanty, U., Bartel, D.P. Compact and ordered collapse of randomly generated RNA sequences. Nat Struct Mol Biol 2005, 12, 1130–1136. [Google Scholar] [CrossRef] [PubMed]
- Dethoff, E.A. , Chugh, J., Mustoe, A.M., Al-Hashimi, H.M. Functional complexity and regulation through RNA dynamics. Nature 2012, 482, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Mustoe, A.M. , Brooks, C.L., Al-Hashimi, H.M. Hierarchy of RNA functional dynamics. Annu Rev Biochem 2014, 83, 441–466. [Google Scholar] [CrossRef] [PubMed]
- Bose, R. , Saleem, I., Mustoe, A.M. Causes, functions, and therapeutic possibilities of RNA secondary structure ensembles and alternative states. Cell Chem Biol 2024, 31, 17–35. [Google Scholar] [CrossRef]
- Lu, Z. , Zhang, Q.C., Lee, B., Flynn, R.A., Smith, M.A., Robinson, J.T., Davidovich, C., Gooding, A.R., Goodrich, K.J., Mattick, J.S., Mesirovm J.P., Cech, T.R., Chang, H.Y. RNA Duplex Map in Living Cells Reveals Higher-Order Transcriptome Structure. Cell 2016, 165, 1267–1279. [Google Scholar] [CrossRef]
- Parlea, L.G. , Sweeney, B.A., Hosseini-Asanjan, M., Zirbel, C.L., Leontis, N.B. The RNA 3D Motif Atlas: Computational methods for extraction, organization and evaluation of RNA motifs. Methods 2016, 103, 99–119. [Google Scholar] [CrossRef]
- Sarrazin-Gendron, R. , Waldispühl, J., Reinharz, V. Classification and Identification of Non-canonical Base Pairs and Structural Motifs. Methods Mol Biol 2024, 2726, 143–168. [Google Scholar] [CrossRef]
- Hermann, T. , Westhof, E. Non-Watson-Crick base pairs in RNA-protein recognition. Chem Biol 1999, 6, R335–43. [Google Scholar] [CrossRef]
- Hermann, T. , Patel, D.J. Adaptive recognition by nucleic acid aptamers. Science 2000, 287, 820–825. [Google Scholar] [CrossRef]
- Leontis, N.B. , Westhof, E. Analysis of RNA motifs. Curr Opin Struct Biol 2003, 13, 300–308. [Google Scholar] [CrossRef]
- Cruz, J.A. , Westhof, E. Sequence-based identification of 3D structural modules in RNA with RMDetect. Nat Methods 2011, 8, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Hamdani, H.Y. , Appasamy, S.D., Willett, P., Artymiuk, P.J., Firdaus-Raih, M. NASSAM: a server to search for and annotate tertiary interactions and motifs in three-dimensional structures of complex RNA molecules. Nucleic Acids Res 2012, 40, issue):W35–41. [Google Scholar] [CrossRef] [PubMed]
- Yen, C.Y. , Lin, J.C., Chen, K.T., Lu, C.L. R3D-BLAST2: an improved search tool for similar RNA 3D substructures. BMC Bioinformatics 2017, 18, 574. [Google Scholar] [CrossRef] [PubMed]
- Emrizal, R. , Hamdani, H.Y., Firdaus-Raih, M. Graph Theoretical Methods and Workflows for Searching and Annotation of RNA Tertiary Base Motifs and Substructures. Int J Mol Sci 2021, 22, 8553. [Google Scholar] [CrossRef]
- Ghani, N.S.A. , Emrizal, R., Moffit, S.M., Hamdani, H.Y., Ramlan, E.I., Firdaus-Raih, M. GrAfSS: a webserver for substructure similarity searching and comparisons in the structures of proteins and RNA. Nucleic Acids Res 2022, 50, W375–W383. [Google Scholar] [CrossRef]
- Lawson, C.L., Berman, H.M., Chen, L., Vallat, B., Zirbel, C.L. The Nucleic Acid Knowledgebase: a new portal for 3D structural information about nucleic acids. Nucleic Acids Res 2024, 52, D245-D254. [CrossRef]
- Staple, D.W. , Butcher, S.E. Pseudoknots: RNA structures with diverse functions. PLoS Biol 2005, 3, e213. [Google Scholar] [CrossRef]
- Kharel, P. , Ivanov, P. RNA G-quadruplexes and stress: emerging mechanisms and functions. Trends Cell Biol 2024, 34, 771–784. [Google Scholar] [CrossRef]
- Lambert, D. , Draper, D.E. Effects of osmolytes on RNA secondary and tertiary structure stabilities and RNA-Mg2+ interactions. J Mol Biol 2007, 370, 993–1005. [Google Scholar] [CrossRef]
- Bugnon, L.A. , Edera, A.A., Prochetto, S., Gerard, M., Raad, J., Fenoy, E., Rubiolo, M., Chorostecki, U., Gabaldón, T., Ariel, F., Di Persia, L.E., Milone, D.H., Stegmayer, G. Secondary structure prediction of long noncoding RNA: review and experimental comparison of existing approaches. Brief Bioinform 2022, 23, bbac205. [Google Scholar] [CrossRef]
- Zhang, K. , Li, S., Kappel, K., Pintilie, G., Su, Z., Mou, T.C., Schmid, M.F., Das, R., Chiu, W. Cryo-EM structure of a 40 kDa SAM-IV riboswitch RNA at 3.7 Å resolution. Nat Commun 2019, 10, 5511. [Google Scholar] [CrossRef]
- Bernard, C. , Postic, G., Ghannay, S., Tahi, F. State-of-the-RNArt: benchmarking current methods for RNA 3D structure prediction. NAR Genom Bioinform 2024, 6, lqae048. [Google Scholar] [CrossRef]
- Islam, S. , Ge, P., Zhang, S. CompAnnotate: a comparative approach to annotate base-pairing interactions in RNA 3D structures. Nucleic Acids Res 2017, 45, e136. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.Z. , Wu, H., Li, S.S., Li, H.Z., Zhang, B.G., Tan, Y.L. ABC2A: A Straightforward and Fast Method for the Accurate Backmapping of RNA Coarse-Grained Models to All-Atom Structures. Molecules 2024, 29, 1244. [Google Scholar] [CrossRef]
- Perry, Z.R. , Pyle, A.M., Zhang, C. Arena: Rapid and Accurate Reconstruction of Full Atomic RNA Structures From Coarse-grained Models. J Mol Biol 2023, 435, 168210. [Google Scholar] [CrossRef] [PubMed]
- Townshend, R.J.L., Eismann, S., Watkins, A.M., Rangan, R., Karelina, M., Das, R., Dror, R.O. Geometric deep learning of RNA structure. Science (New York, N.Y.) 2021, 373, 1047–1051. Erratum in: Science 2023, 379, eadg6616. 2021, 373, 1047–1051. [CrossRef]
- Peattie, D.A. , Gilbert, W. Chemical probes for higher-order structure in RNA. Proc Natl Acad Sci U S A 1980, 77, 4679–4682. [Google Scholar] [CrossRef]
- Ehresmann, C. , Baudin, F., Mougel, M., Romby, P., Ebel, J.P., Ehresmann, B. Probing the structure of RNAs in solution. Nucleic Acids Res 1987, 15, 9109–9128. [Google Scholar] [CrossRef]
- Incarnato, D. , Neri, F., Anselmi, F., Oliviero, S. Genome-wide profiling of mouse RNA secondary structures reveals key features of the mammalian transcriptome. Genome Biol 2014, 15, 491. [Google Scholar] [CrossRef]
- Ding, Y. , Tang, Y., Kwok, C.K., Zhang, Y., Bevilacqua, P.C., Assmann, SM. In vivo genome-wide profiling of RNA secondary structure reveals novel regulatory features. Nature 2014, 505, 696–700. [Google Scholar] [CrossRef]
- Ritchey, L.E. , Su, Z., Tang, Y., Tack, D.C., Assmann, S.M., Bevilacqua, P.C. Structure-seq2: sensitive and accurate genome-wide profiling of RNA structure in vivo. Nucleic Acids Res 2017, 45, e135. [Google Scholar] [CrossRef]
- Ritchey, L.E. , Su, Z., Assmann, S.M., Bevilacqua, P.C. In Vivo Genome-Wide RNA Structure Probing with Structure-seq. Methods Mol Biol 2019, 1933, 305–341. [Google Scholar] [CrossRef] [PubMed]
- Ritchey, L.E. , Tack, D.C., Yakhnin, H., Jolley, E.A., Assmann, S.M., Bevilacqua, P.C., Babitzke, P. Structure-seq2 probing of RNA structure upon amino acid starvation reveals both known and novel RNA switches in Bacillus subtilis. RNA 2020, 26, 1431–1447. [Google Scholar] [CrossRef] [PubMed]
- Rouskin, S. , Zubradt, M., Washietl, S., Kellis, M., Weissman, J.S. Genome-wide probing of RNA structure reveals active unfolding of mRNA structures in vivo. Nature 2014, 505, 701–705. [Google Scholar] [CrossRef] [PubMed]
- Zubradt, M. , Gupta, P., Persad, S., Lambowitz, A.M., Weissman, J.S., Rouskin, S. DMS-MaPseq for genome-wide or targeted RNA structure probing in vivo. Nat Methods 2017, 14, 75–82. [Google Scholar] [CrossRef]
- Yamagami, R. , Sieg, J.P., Assmann, S.M., Bevilacqua, P.C. Genome-wide analysis of the in vivo tRNA structurome reveals RNA structural and modification dynamics under heat stress. Proc Natl Acad Sci U S A 2022, 119, e2201237119. [Google Scholar] [CrossRef]
- Kwok, C.K. , Marsico, G., Sahakyan, A.B., Chambers, V.S., Balasubramanian, S. rG4-seq reveals widespread formation of G-quadruplex structures in the human transcriptome. Nat Methods 2016, 13, 841–844. [Google Scholar] [CrossRef]
- Siegfried, N.A. , Busan, S., Rice, G.M., Nelson, J.A., Weeks, K.M. RNA motif discovery by SHAPE and mutational profiling (SHAPE-MaP). Nat Methods 2014, 11, 959–965. [Google Scholar] [CrossRef]
- Spitale, R.C., Flynn, R.A., Zhang, Q.C., Crisalli, P., Lee, B., Jung, J.W., Kuchelmeister, H.Y., Batista, P.J., Torre, E.A., Kool, E.T., Chang, H.Y. Structural imprints in vivo decode RNA regulatory mechanisms. Nature 2015, 519, 486-490. https://doi.org/10.1038/nature14263. Erratum in: Nature 2015, 527, 264. [CrossRef]
- Spasic, A. , Assmann, S.M., Bevilacqua, P.C., Mathews, D.H. Modeling RNA secondary structure folding ensembles using SHAPE mapping data. Nucleic Acids Res 2018, 46, 314–323. [Google Scholar] [CrossRef]
- Yang, M. , Woolfenden, H.C., Zhang, Y., Fang, X., Liu, Q., Vigh, M.L., Cheema, J., Yang, X., Norris, M., Yu, S., Carbonell, A., Brodersen, P., Wang, J., Ding, Y. Intact RNA structurome reveals mRNA structure-mediated regulation of miRNA cleavage in vivo. Nucleic Acids Res 2020, 48, 8767–8781. [Google Scholar] [CrossRef]
- Yang, X. , Cheema, J., Zhang, Y., Deng, H., Duncan, S., Umar, M.I., Zhao, J., Liu, Q., Cao, X., Kwok, C.K., Ding, Y. RNA G-quadruplex structures exist and function in vivo in plants. Genome Biol 2020, 21, 226. [Google Scholar] [CrossRef]
- Luo, Q.J. , Zhang, J., Li, P., Wang, Q., Zhang, Y., Roy-Chaudhuri, B., Xu, J., Kay, M.A., Zhang, Q.C. RNA structure probing reveals the structural basis of Dicer binding and cleavage. Nat Commun 2021, 12, 3397. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D. 3rd., Ritchey, L.E., Park, H., Babitzke, P., Assmann, S.M., Bevilacqua, P.C. Glyoxals as in vivo RNA structural probes of guanine base-pairing. RNA 2018, 24, 114–124. [CrossRef]
- Mitchell, D. 3rd., Renda, A.J., Douds, C.A., Babitzke, P., Assmann, S.M., Bevilacqua, P.C. In vivo RNA structural probing of uracil and guanine base-pairing by 1-ethyl-3-(3-dimethylaminopropyl)carbodiimide (EDC). RNA 2019, 25, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.Y. , Sexton, A.N., Culligan, W.J., Simon, M.D. Carbodiimide reagents for the chemical probing of RNA structure in cells. RNA 2019, 25, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Spitale, R.C. , Crisalli, P., Flynn, R.A., Torre, E.A., Kool, E.T., Chang, H.Y. RNA SHAPE analysis in living cells. Nat Chem Biol 2013, 9, 18–20. [Google Scholar] [CrossRef]
- Bushhouse, D.Z. , Choi, E.K., Hertz, L.M., Lucks, J.B. How does RNA fold dynamically? J Mol Biol 2022, 434, 167665. [Google Scholar] [CrossRef]
- Herbert, A. , Hatfield, A., Lackey, L. How does precursor RNA structure influence RNA processing and gene expression? Biosci Rep 2023, 43, BSR20220149. [Google Scholar] [CrossRef]
- Senior, A.W. , Evans, R., Jumper, J., Kirkpatrick, J., Sifre, L., Green, T., Qin, C., Žídek, A., Nelson, A.W.R., Bridgland, A., Penedones, H., Petersen, S., Simonyan, K., Crossan, S., Kohli, P., Jones, D.T., Silver, D., Kavukcuoglu, K., Hassabis, D. Improved protein structure prediction using potentials from deep learning. Nature 2020, 577, 706–710. [Google Scholar] [CrossRef]
- Jumper, J. , Evans, R., Pritzel, A., Green, T., Figurnov, M., Ronneberger, O., Tunyasuvunakool, K., Bates, R., Žídek, A., Potapenko, A., Bridgland, A., Meyer, C., Kohl, S. A. A., Ballard, A. J., Cowie, A., Romera-Paredes, B., Nikolov, S., Jain, R., Adler, J., Back, T., … Hassabis, D. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Schneider, B. , Sweeney, B.A., Bateman, A., Cerny, J., Zok, T., Szachniuk, M. When will RNA get its AlphaFold moment? Nucleic Acids Res 2023, 51, 9522–9532. [Google Scholar] [CrossRef]
- Miao, Z. , Westhof, E. RNA Structure: Advances and Assessment of 3D Structure Prediction. Annu Rev Biophys 2017, 46, 483–503. [Google Scholar] [CrossRef]
- Piao, M. , Sun, L., Zhang, Q.C. RNA Regulations and Functions Decoded by Transcriptome-wide RNA Structure Probing. Genomics Proteomics Bioinformatics 2017, 15, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Li, J. , Zhu, W., Wang, J., Li, W., Gong, S., Zhang, J., Wang, W. RNA3DCNN: Local and global quality assessments of RNA 3D structures using 3D deep convolutional neural networks. PLoS computational biology 2018, 14, e1006514. [Google Scholar] [CrossRef] [PubMed]
- Li, J. , Zhang, S., Zhang, D., Chen, S.J. Vfold-Pipeline: a web server for RNA 3D structure prediction from sequences. Bioinformatics 2022, 38, 4042–4043. [Google Scholar] [CrossRef] [PubMed]
- Tieng, F.Y.F. , Abdullah-Zawawi, M.R., Md Shahri, N.A.A., Mohamed-Hussein, Z.A., Lee, L.H., Mutalib, N.A. A Hitchhiker's guide to RNA-RNA structure and interaction prediction tools. Brief Bioinform 2023, 25, bbad421. [Google Scholar] [CrossRef]
- Wang, X. , Yu, S., Lou, E., Tan, Y.L., Tan, Z.J. RNA 3D Structure Prediction: Progress and Perspective. Molecules 2023, 28, 5532. [Google Scholar] [CrossRef]
- Murthy, V.L. , Rose, G.D. RNABase: an annotated database of RNA structures. Nucleic Acids Res 2003, 31, 502–504. [Google Scholar] [CrossRef]
- Khan, N.S. , Rahaman, M.M., Islam, S., Zhang, S. RNA-NRD: a non-redundant RNA structural dataset for benchmarking and functional analysis. NAR Genom Bioinform 2023, 5, lqad040. [Google Scholar] [CrossRef]
- Rigden, D.J. , Fernández, X.M. The 2024 Nucleic Acids Research database issue and the online molecular biology database collection. Nucleic Acids Res 2024, 52, D1–D9. [Google Scholar] [CrossRef]
- Ma, J., Tsuboi, T. Efficient Prediction Model of mRNA End-to-End Distance and Conformation: Three-Dimensional RNA Illustration Program (TRIP). Methods Mol Biol 2024, 2784, 191-200. [CrossRef]
- Wang, W. , Feng, C., Han, R., Wang, Z., Ye, L., Du, Z., Wei, H., Zhang, F., Peng, Z., Yang, J. trRosettaRNA: automated prediction of RNA 3D structure with transformer network. Nat Commun 2023, 14, 7266. [Google Scholar] [CrossRef]
- Szikszai, M. , Magnus, M., Sanghi, S., Kadyan, S., Bouatta, N., Rivas, E. RNA3DB: A structurally-dissimilar dataset split for training and benchmarking deep learning models for RNA structure prediction. J Mol Biol 2024, 168552. [Google Scholar] [CrossRef]
- Ramakers, J., Blum, C.F., König, S., Harmeling, S., Kollmann, M. De novo prediction of RNA 3D structures with deep generative models. PLoS One 2024, 19, e0297105. [CrossRef]
- Liu, J. , McRae, E.K.S., Zhang, M., Geary, C., Andersen, E.S., Ren, G. Non-averaged single-molecule tertiary structures reveal RNA self-folding through individual-particle cryo-electron tomography. Nat Commun 2024, 15, 9084. [Google Scholar] [CrossRef]
- National Academies of Sciences, Engineering, and Medicine; Health and Medicine Division; Division on Earth and Life Studies; Board on Health Sciences Policy; Board on Life Sciences; Toward Sequencing and Mapping of RNA Modifications Committee. Charting a Future for Sequencing RNA and Its Modifications: A New Era for Biology and Medicine. Washington (DC): National Academies Press (US); 2024 Jul 22.
- Kanatani, S. , Kreutzmann, J.C., Li, Y., West, Z., Larsen, L.L., Nikou, D.V., Eidhof, I., Walton, A., Zhang, S., Rodríguez-Kirby, L.R., Skytte, J.L., Salinas, C.G., Takamatsu, K., Li, X., Tanaka, D.H., Kaczynska, D., Fukumoto, K., Karamzadeh, R., Xiang, Y., Uesaka, N., Tanabe, T., Adner, M., Hartman, J., Miyakawa, A., Sundström, E., Castelo-Branco, G., Roostalu, U., Hecksher-Sørensen, J., Uhlén, P. Whole-brain spatial transcriptional analysis at cellular resolution. Science 2024, 386, 907–915. [Google Scholar] [CrossRef]
Figure 1.
Primary, secondary, and tertiary structural levels of RNA.
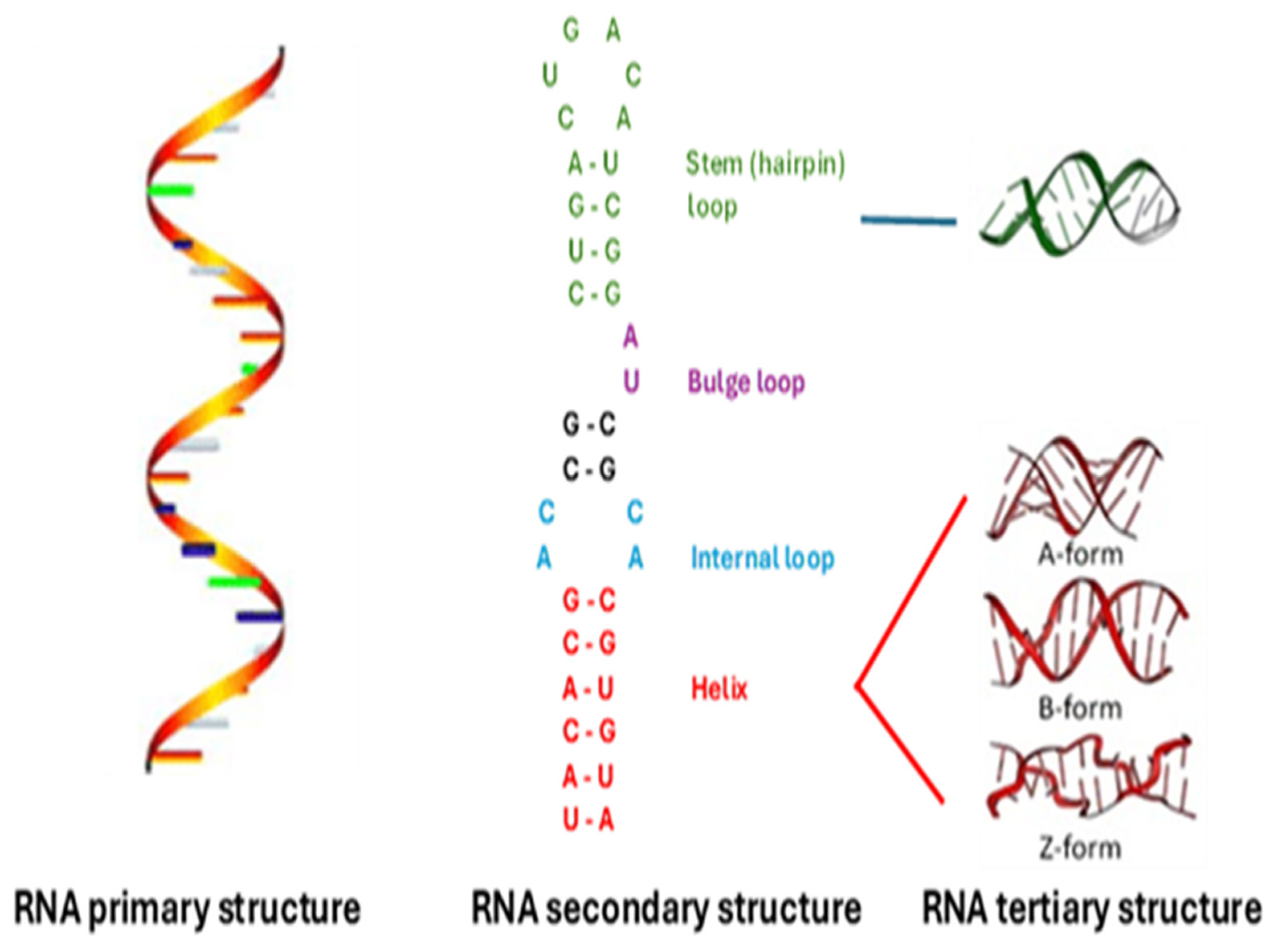
Figure 2.
H-type pseudoknot. Complementarity regions are shown as overlapping boxes.
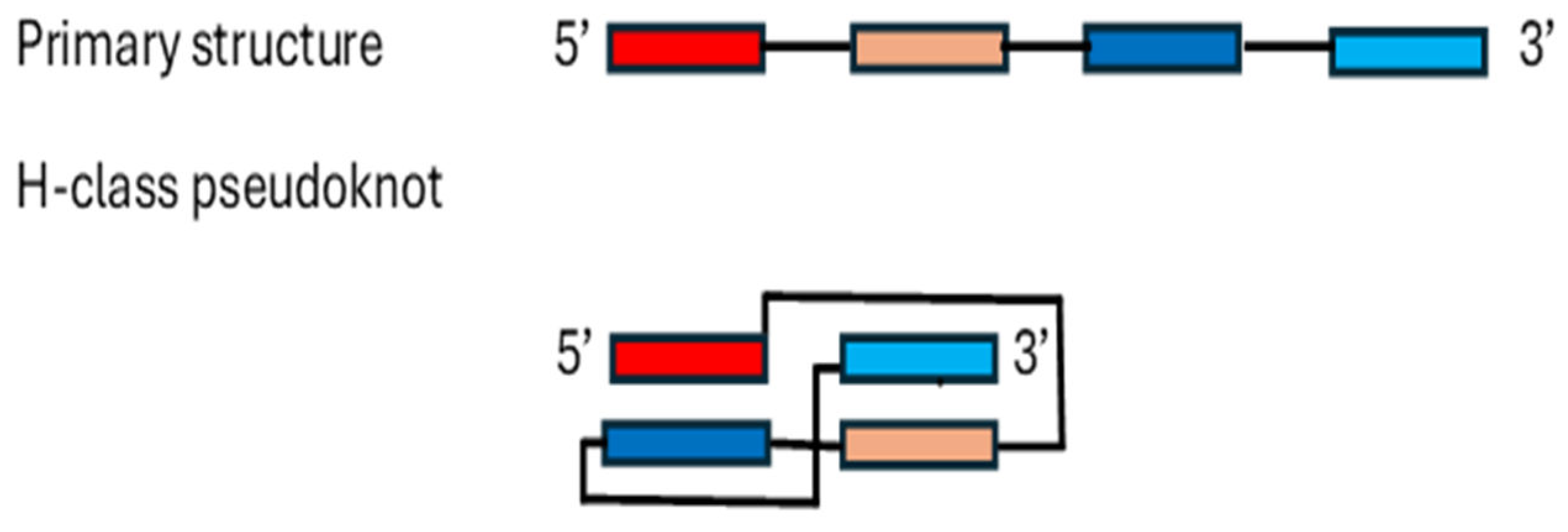
Figure 2.
RNA G-quadruplex. Guanine residues forming stacked tetrads are in red. Arrows follow the primary sequence.
Figure 2.
RNA G-quadruplex. Guanine residues forming stacked tetrads are in red. Arrows follow the primary sequence.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



