
Submitted:
25 November 2024
Posted:
25 November 2024
You are already at the latest version
Abstract
The emergence of cognition requires a framework that bridges evolutionary principles with neuro-computational mechanisms. This paper introduces the novel “thoughtseed” framework, proposing that cognition arises from dynamic interactions of self-organizing units of embodied knowledge called “thoughtseeds” within the Global Workspace of consciousness. By leveraging evolutionary theory, neuronal packets, and the Free Energy Principle, we propose a hierarchical model of cognitive states comprising Neuronal Packet Domains (NPDs), Knowledge Domains (KDs), thoughtseed networks, and meta-cognition. This hierarchical interplay, mediated by nested Markov blankets and reciprocal message passing, facilitates the emergence of thoughtseeds as coherent neural activity patterns that guide perception, action, and learning. Thoughtseeds, posited as fundamental units of thought, compete for dominance within the Global Workspace, with the dominant thoughtseed shaping conscious experiences and guiding behavior. We present an illustrative mathematical framework grounded in active inference and dynamical systems theory to model thoughtseed dynamics and their contribution to the unitary nature of consciousness. This biologically grounded model offers a novel approach to understanding the organizing principles and emergence of embodied cognition, with potential applications in consciousness, attention, and decision making.
Keywords:
global workspace
; evolution
; embodied cognition
; Markov blanket
; thoughtseed
; free energy principle
; neuronal packet
; metacognition
1. Introduction
1.1. Embodied Cognition: An Evolutionary and Free Energy Perspective
Survival depends not only on adaptation but also on the ability of living systems to actively shape their environments [1]. Evolutionary mechanisms such as the Baldwin effect and natural selection explain how learning and environmental pressures influence evolutionary processes [2,3]. Beyond genetic evolution, modern perspectives have recognized the role of epigenetic, behavioral, and cultural inheritance in shaping living systems and their environments [4,5]. These interactions underscore how living systems dynamically regulate their relationship with the environment, not just responding to it but influencing it through actions and perceptions [6,7]. This dynamic, adaptive relationship is central to the concept of embodied cognition, in which understanding emerges through interactions between the brain, body, and the environment [8,9].
In line with this view, autopoiesis describes the self-organizing nature of living systems [10], allowing them to maintain a non-equilibrium steady state (NESS) by continuously exchanging energy and matter with their surroundings [11]. These systems resist the natural tendency towards disorder (entropy), exhibit emergent properties [12,13], and actively regulate internal states to ensure persistence. Building on the idea that effective regulation requires an internal model [14], the Free Energy Principle (FEP) provides a unifying framework to explain how living systems adapt by minimizing surprise or variational free energy [15]. Through a process called active inference, living systems continuously refine their internal models and take actions that align their predictions with incoming sensory data [16,17]. The Markov blanket concept, central to FEP, separates internal states from the external world, enabling computational autonomy and conditional independence while mediating interactions with the environment [15,17].
FEP’s scale-free modeling approach integrates both evolutionary and cognitive dynamics, offering insights into the emergence and evolution of adaptive systems [18,19]. Furthermore, the Hierarchically Mechanistic Mind (HMM) hypothesis views the brain as an adaptive control system that minimizes free energy through recursive interactions between neurocognitive processes, which emerge as a result of evolutionary pressure and self-organization [20].
1.2. Neuronal Representations
Efforts to understand the neural basis of cognition have explored representational units beyond individual neurons, such as cognits [21], engrams [22], and cell assemblies [23]. Furthermore, recent research suggests that cognitive processes may operate in a discrete manner, with distinct packets of information or “cognitive atoms” forming the building blocks of thought and consciousness [24]. These discrete units of cognitive processing compete for access to a global workspace, where they can be broadcast across the brain to generate a unified and coherent conscious experience [25,26].
The Neuronal Packet Hypothesis (NPH) [27,28,29] posits that neuronal packets (NPs), which are self-organizing ensembles of neurons, serve as fundamental units of neuronal representation in the brain [30]. Superordinate ensembles (SEs) emerge from the coordinated activities of multiple NPs, enabling the representation of complex and abstract concepts. Through their interactions and emitted signals, NPs allow the system to infer probabilistic beliefs about external states and continuously update these beliefs to minimize free energy [29].
Each NP and SE possesses its own Markov blanket that defines its boundaries and interactions. The Markov blankets of lower-level NPs are nested within those of higher-level SEs, integrating information through a shared generative model [28]. This hierarchical structure facilitates the brain’s ability to represent knowledge across multiple scales from sensory details to abstract categories. Nested SEs interact via reciprocal message passing, where top-down predictions from higher-level SEs modulate lower-level SEs and NPs, whereas bottom-up sensory evidence and prediction errors update the higher-level representations. This continuous exchange of information supports adaptive behavior in response to a changing environment [18,28,31]. The FEP also has implications for the nature of the representation itself, suggesting that the content of neural representations lies in their role in guiding actions and minimizing surprise [32,33].
NPH lays a robust foundation for exploring several key questions: How do evolutionary pressures shape the emergence and organization of neuronal packets and their hierarchical ensembles? How do individual experiences interact with inherited predispositions to refine and adapt to these representations? How do these representational units collaborate within a network to generate complex cognition and adaptive behavior?
2. Architectural Foundations of Embodied Cognition
2.1. The Thoughtseed Hypothesis
A fundamental challenge in elucidating the neural basis of cognition lies in the inherent opacity of the Markov blanket, which can be referred to as the “black box” problem. While this statistical construct offers a robust framework for modeling the exchange of information between an agent and its environment, it inherently obscures the internal generative model responsible for behavior [15,34]. See Figure 1.

Table 1.
Key Notations of Thoughtseed Framework.
| Concept | Symbol | Explanation |
| Neuronal Packet (NP) | 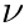 |
The fundamental unit of neuronal representation, a self-organizing ensemble of neurons that encodes a specific feature or aspect of the world. |
| Core Attractor | 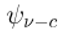 |
The most probable and stable pattern of neural activity within a manifested NP, embodying its core functionality. |
| Subordinate Attractor | 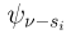 |
Less dominant patterns of neural activity within an NP that may become active under specific conditions or in response to novel stimuli, offering flexibility and adaptability. |
| Encapsulated Knowledge Structure | 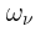 |
The structured knowledge content within a NP’s Markov Blanket, shaping its activity and contribution to KDs. |
| Superordinate Ensemble (SE) | 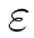 |
A higher-order organization emerging from the coordinated activity of multiple NPs, or even KDs, enabling the representation of more complex and abstract concepts. |
| Neuronal Packet Domain (NPD) | 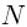 |
A functional unit within the brain, comprised of interconnected SEs, specialized for specific cognitive processes or tasks. |
| Knowledge Domain (KD) | 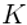 |
A large-scale, organized structure within the brain’s internal model, representing interconnected networks of concepts, categories, and relationships that constitute a specific area of knowledge or expertise. |
| Thoughtseed | 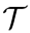 |
A higher-order construct with agency, emerging from the coordinated activity of SEs across different KDs. It represents a unified and meaningful representation of a concept, idea, or percept and guides perception, action, and decision-making. |
| Thoughtseed Activation Level | 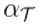 |
A measure of a thoughtseed’s prominence in the current cognitive landscape, calculated by weighing the probabilities that the brain’s state aligns with the thoughtseed’s dominant or subordinate attractor states. |
| Global Activation Threshold | 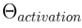 |
A global parameter, influenced by the consciousness state and arousal levels that determines the minimum activation level a thoughtseed must cross to enter the active thoughtseed pool. |
| Active Thoughtseed Pool | 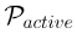 |
The set of active thoughtseeds whose activation levels surpass the activation threshold at a given time. |
| Dominant Thoughtseed | 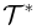 |
The thoughtseed within the active thoughtseed pool that has the highest activation level and is primarily shaping the content of consciousness at a given moment. |
| Thoughtseed Network | 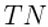 |
The collection of interconnected thoughtseeds within the brain’s internal states, hypothesized to encode a generative model of the environment. |
To address these challenges, we propose the “thoughtseed” framework, which builds upon the concept of neuronal packets (NPs) and neuronal packet hypothesis (NPH). This framework integrates insights from the free-energy principle (FEP) [15,18], evolutionary theory, and Global Workspace Theory (GWT) [35,36].
The thoughtseed hypothesis posits that cognition arises from the dynamic interaction of self-organizing units of embodied knowledge termed “thoughtseeds.” These thoughtseeds, conceptualized as emergent agents with their own Markov blankets, are shaped by a hierarchy of evolutionary priors that encompasses both inherited predispositions and learned experiences. This framework proposes a four-level hierarchical/heterarchical model of the cognitive agent’s internal states: Neuronal Packet Domains (NPDs), Knowledge Domains (KDs), Thoughtseed Network (TN), and meta-cognition.
Each level in this model consists of nested Markov blankets encoding information about the world at varying levels of abstraction, with deeper levels representing more domain-general and abstract knowledge [33,37]. The dynamic interplay within this hierarchy facilitates the emergence of thoughtseeds as coherent patterns of activities that guide perception, action, and learning. The framework emphasizes the embodied nature of cognition, where the living system’s interactions with its Umwelt [38] and the knowledge encoded in the KDs shape the Thoughtseed Network.
In the following sections, we elaborate on the architectural foundations of the thoughtseed framework, exploring the details of NPDs, KDs, and their dynamic interplay in the emergence of thoughtseeds and shaping of conscious experience through nested Markov blankets.
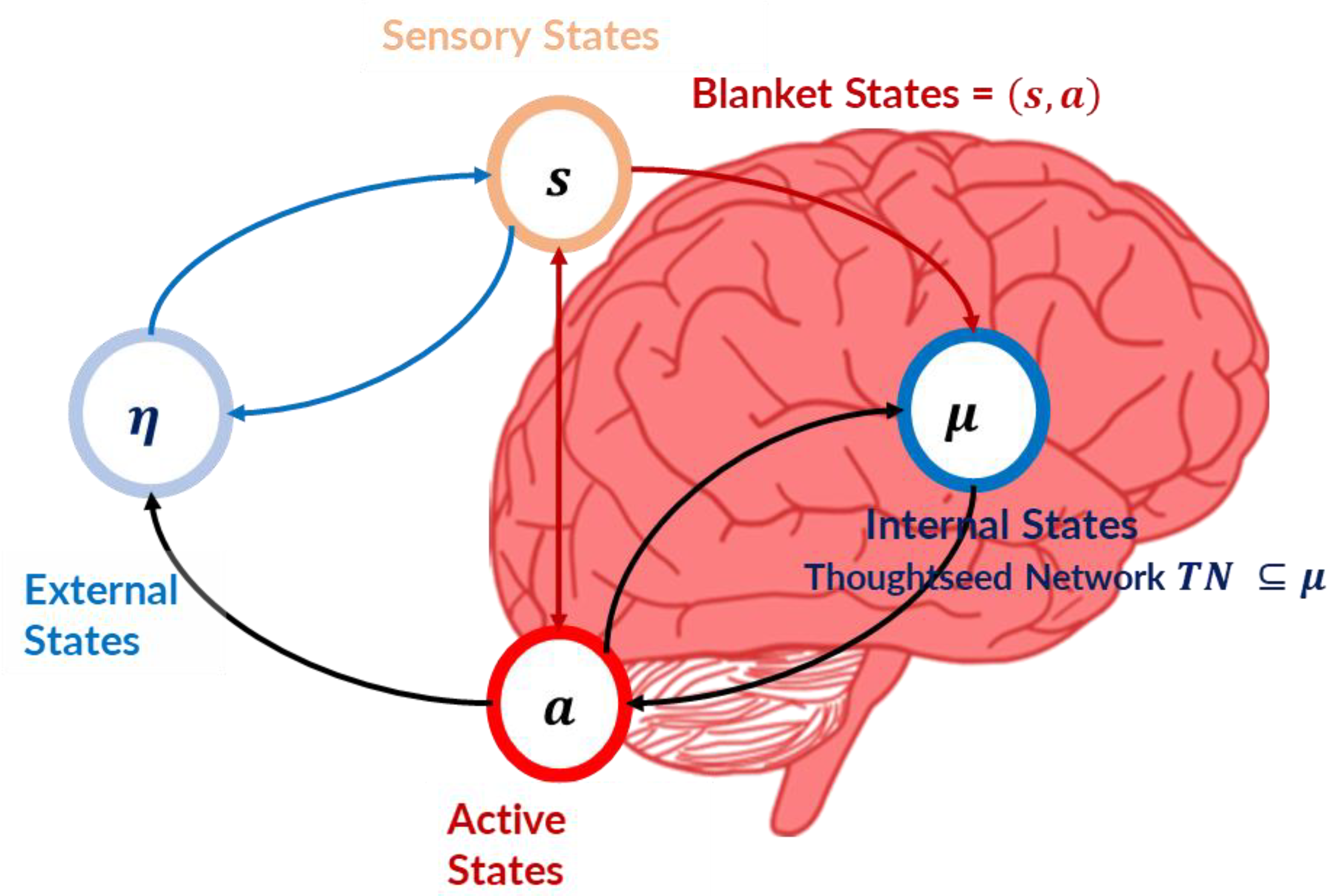
Figure 1.
Markov Blanket of an Agent. Adapted from Friston et al. [39], this diagram illustrates the partitioning of an agent’s states into internal states (μ), encompassing the Thoughtseed Network (TN), and external states (η). The Markov blanket, comprising sensory (s) and active states (a), mediates the interaction between the internal and external states. The internal states, housing the TN, can only influence active states, while external states can only influence sensory states. This separation allows the TN to operate with a degree of autonomy, generating predictions and selecting actions based on its internal model of the world.
Figure 1.
Markov Blanket of an Agent. Adapted from Friston et al. [39], this diagram illustrates the partitioning of an agent’s states into internal states (μ), encompassing the Thoughtseed Network (TN), and external states (η). The Markov blanket, comprising sensory (s) and active states (a), mediates the interaction between the internal and external states. The internal states, housing the TN, can only influence active states, while external states can only influence sensory states. This separation allows the TN to operate with a degree of autonomy, generating predictions and selecting actions based on its internal model of the world.
2.2. Neuronal Packet Domains
The sparse architecture of the brain, characterized by a limited number of active neurons and low connection density, supports localized functional units for specific cognitive tasks [40,41]. NPDs are hypothesized to emerge through self-organizing interconnected NPs. Their hierarchical organization, via nested Markov blankets, reflects “ascending scales of canonical microcircuits” [42,43,44], enabling complex computations from simpler units. This structure facilitates efficient information processing and adaptive behavior [18,28,31].
NPD formation is guided by evolutionary priors that favor adaptive neural architectures suited for specific functions [45]. For instance, the visual cortex has distinct NPDs for color, motion, and form [46,47], and these specializations are evolutionarily conserved across mammalian species [48]. Hebbian learning further shapes their specific organization.
NPs compete for resources by minimizing variational free energy (VFE), resulting in efficient internal models. This competition, described as “neural Darwinism,” produces specialized NPs, each representing a different aspect of the world [49]. The Markov blanket structure supports this decentralized modular organization, enhancing both robustness and adaptability.
NPs dynamically interact to form superordinate ensembles (SEs) within NPDs, representing higher-order concepts and allowing for greater complexity in cognitive representations. The hierarchical structure, where the Markov blankets of lower-level NPs are nested within those of higher-level SEs, enables the brain to encode knowledge across different scales from sensory details to abstract categories [29,50]. SEs may emerge as stable entities when they accurately predict and explain sensory input [18]. Nested SEs interact through reciprocal message passing, facilitating flexible and adaptive behavior in response to changing environments.
Furthermore, the formation of SEs within NPDs is influenced by the alignment of knowledge representations through vector rotations. When multiple NPs contribute to a shared generative model, their encapsulated knowledge representations undergo alignment via vector rotation [27]. This rotation facilitates the integration of knowledge from different NPs by bringing their vectors closer within the knowledge space. The rotation angle is determined based on the similarity between the generative models of the participating NPs. This alignment process is an integral part of the formation of SEs, ensuring that the constituent NPs have a coherent and integrated representation of knowledge relevant to the shared generative model [29].
2.3. Neuronal Packets (NPs): The Fundamental Units of Neuronal Representation
Within each NPD, NPs serve as fundamental units of neuronal representation [27,29,30]. An NP can exist in three states. See Figure 2.
- Unmanifested State: Represents a potential configuration of neural activity with high prior probability under specific conditions, shaped by evolutionary priors. It can be viewed as a sparsely connected neural ensemble with low precision, corresponding to a shallow local minimum in the free energy landscape, indicating low stability and high potential for change.
- Manifested State: Emerges from the unmanifested state upon repeated exposure to relevant stimuli, leading to a phase transition [30] and the formation of a Markov Blanket stabilized by an energy barrier [30]. This Markov blanket structure may enable local computation and autonomy within the NP, while maintaining informational boundaries. It is characterized by increased coherence of neural activity, resulting in a stable state with a core attractor representing the most probable and stable pattern of neural activity [11,17]. This state embodies the NP’s core functionality with high certainty and can be interpreted as the mode of posterior distribution over the NP’s internal states given its Markov blanket [18]. This corresponds to a deeper local minimum in the free energy landscape, reflecting high stability and low surprise [51]. The depth of this global minimum can indicate the NP binding energy, reflecting the overall stability and resistance to changes in the core representation [30]. Subordinate attractors represent less dominant patterns of neural activity, which may become active under specific conditions or in response to novel stimuli [52]. They can be viewed as shallower local minima in the free energy landscape, separated from the core attractor by energy barriers. The existence of alternative attractors, along with the energy barriers that separate them, allows flexibility and adaptability in the NP response to changing inputs.
- Activated (or Spiking) State: A transient state characterized by heightened neural activity within the manifested NP ensemble triggered by specific inputs that resonate with the NP’s internal model. NP generates a response that may influence the behavior or cognition of a living system. This response can be interpreted as a consequence of the NP’s internal dynamics and its attempt to minimize free energy rather than deliberate or planned action. The active state can be seen as a temporary shift in the free energy landscape, where the core attractor becomes even more pronounced.
2.4. Knowledge Domains (KDs)
Knowledge Domains (KDs) are conceptualized as large-scale, organized structures within the brain’s internal model that function as knowledge hypergraphs [53]. These domains serve as nested levels of “knowledge repositories” that provide conceptual scaffolding for interpreting sensory information and generating the content of consciousness within the Global Workspace. KDs encompass not only sensory representations but also retrieved memories, beliefs, experiences, policies, emotions, and learning patterns.
KDs are hypothesized to exhibit both hierarchical and heterarchical structures, enabling flexible and context-dependent knowledge retrieval for adaptive behavior [54]. The hierarchical nature represents layers of knowledge from concrete to abstract [42,55,56], whereas the heterarchical structure highlights cross-domain interactions and the integration of diverse knowledge sources [30].
In contrast to Neuronal Packet Domains (NPDs) that process raw sensory data, KDs contextualize this information within the brain’s broader neural architecture. Their formation is influenced by evolutionary priors, which provide a foundational structure [57], whereas continuous learning refines KDs throughout life. See Figure 3. Moreover, KDs possess an intrinsic affective dimension that reflects the emotional valence and arousal associated with their content [58], shaping subjective experiences, and influencing behavior and decision-making processes [59,60]. This emotional component is vital in guiding behavior, although certain KDs related to abstract knowledge such as mathematical rules may lack affective content.
Dynamic, context-dependent binding within KDs integrates information across NPDs, helping form coherent percepts in the Global Workspace. This integration is supported by neural synchrony [61], superordinate ensembles spanning multiple NPs, and attentional mechanisms that prioritize relevant information [62]. Enhanced neuronal firing rates may facilitate binding across NPDs [63]. This process is reinforced by reciprocal message passing and attentional selection, which reflect circular causality within the FEP, where hierarchical levels influence each other through prediction and error correction [64].
Furthermore, KDs can be conceptualized as abstract spaces in which thoughtseeds interact to form complex representations and narratives. This aligns with the perspective that intelligence stems from navigating physical space and subsequently generalizes to “navigating abstract spaces” for higher-level cognition [65]. In the thoughtseed framework, KDs provide a scaffold for organizing knowledge, facilitating the integration of diverse modalities into coherent cognitive frameworks.
A contrived illustration of potential brain areas that have a greater influence on KDs might include the following:
The thoughtseed framework offers a biologically grounded model of cognition within the broader context of mortal computation [73], elucidating how cognitive processes emerge from the dynamic interplay between neuronal packets, knowledge domains, and higher-order structures. An illustrative example of Visual Object Recognition KD is provided in Supplementary Section.
3. Thoughtseeds Network and Meta-Cognition
3.1. Formal Definition of a Thoughtseed
The thoughtseed framework posits that thoughtseeds are self-organizing higher-order cognitive constructs that emerge from the coordinated activity of distributed neural networks, aligning with research on large-scale brain networks and the Global Workspace Theory of consciousness (GWT) [74].
Thoughtseeds can be understood as transient units within this global workspace, representing temporary coalitions of neural activity that form, compete, and dissolve as an individual interacts with the world. These dynamic patterns of neuronal activity can be interpreted as metastable states within the brain’s pullback attractor landscape [11,15,75].
The content and structure of thoughtseeds emerge from knowledge and beliefs embedded within Knowledge Domains (KDs), reflecting the evolutionary history and individual experiences of the living system. The dynamic interplay between thoughtseeds and KDs shapes cognition, behavior, and conscious experiences.
3.2. Key Characteristics of Thoughtseeds
- Sub-Agents: Thoughtseeds act as autonomous sub-agents within the cognitive system, engaging in active inference to generate predictions, influence actions, and update internal models based on sensory feedback [76]. This agency allows them to explore the environment and develop affordances.
- Pullback Attractor Dynamics: When active, thoughtseeds function as pullback attractors [11,77] by integrating information from multiple KDs to form coherent representations. This establishes a transient Markov blanket that maintains autonomy and computational independence. Each thoughtseed is associated with a core attractor, representing its most probable and stable pattern of neural activity, and may also have subordinate attractors, offering flexibility and adaptability.
- Goal-Directed Behavior: Thoughtseeds are inherently goal-directed and driven by the imperative to minimize Expected Free Energy (EFE ) [78]. They anticipate action consequences and select those that minimize EFE. The interaction between thoughtseeds and KDs results in affordances, or potential actions within the environment [79]. These affordances can be categorized as epistemic (opportunities for learning or exploration) or pragmatic (opportunities for goal fulfillment or exploitation) [80,81].
- Learning and adaptation: Thoughtseeds continuously engage in learning and adaptation through a process of Bayesian belief updating. Each thoughtseed possesses a generative model that predicts its future state and the content of consciousness it generates. As the thoughtseed interacts with the environment and receives sensory feedback, it updates its internal model and beliefs, thereby refining its predictions and contributing to more adaptive behavior [103]. This dynamic updating process enables thoughtseeds to maintain responsiveness to environmental changes and to continuously enhance their representation of the world.
3.2.1. Thoughtseed States
Thoughtseeds exist in the following states: unmanifested, manifested (inactive, active, dominant, dissipated).
- Unmanifested: The thoughtseed exists as a potential configuration within the neural network, latent within interconnected knowledge domains, primed by prior experiences and learning. It is yet to actively influence cognition or behavior.
-
Manifested: The thoughtseed has emerged and is now part of the cognitive landscape, with the potential to influence thought and action. This state is further divided into the following substates:
- o
- Inactive: The thoughtseed is present but does not currently contribute to conscious experience. It resides in the background, with its core attractor and associated dynamics intact and potentially primed for activation.
- o
- Active: The thoughtseed is in the “active thoughtseed pool” and contributes to the content of consciousness, competing with other active thoughtseeds for dominance.
- o
- Dominant: The dominant thoughtseed is selected through an ongoing process of cumulative EFE minimization, in which thoughtseeds within the active thoughtseed pool compete in a winner-take-all dynamic. The thought seed that minimizes cumulative EFE over time gains dominance, enters the Global Workspace, and shapes the unitary conscious experience. This process ensures that the most relevant, predictive, and adaptive thoughtseed guides cognition and actions.
- o
- Dissipated: The thoughtseed has lost its influence and its core attractor has become significantly weakened or dissolved. It no longer contributes to the active thoughtseed pool and its constituent elements may be reabsorbed into the broader neural network or contribute to the formation of new thoughtseeds.
Formal Definition: A “thoughtseed” is a higher-order, transient, Markov-blanketed unit with agency that emerges from the coordinated activity of neuronal ensembles across different knowledge domains (KDs). It is characterized by a core attractor and subordinate attractors that shape its content and dynamics. Thoughtseeds exhibit repetitive behavior, revisiting the brain’s cognitive landscape even when not explicitly required, contributing to phenomena such as mind-wandering. Through continuous competition with other thoughtseeds via a winner-take-all dynamic, a dominant thoughtseed emerges, enters the Global Workspace, and contributes to the unitary conscious experience, enabling active exploration of the environment and development of affordances.
An illustrative example of a “dog thoughtseed” is shown in Supplementary Section.
3.3. Thoughtseeds and the Global Workspace
Global Workspace Theory (GWT) proposes that consciousness arises from a “global workspace, “ a central information exchange hub within the brain [25,35]. This workspace facilitates communication and integration among the specialized modular processing units. Within the thoughtseed framework, Knowledge Domains (KDs) can be conceptualized as specialized modules, each encapsulating a specific domain of knowledge or expertise. These KDs operate in parallel, processing their respective inputs until the selection process determines which information will enter the global workspace for conscious access.
3.3.1. Thoughtseeds as Information Processors
Thoughtseeds, in turn, act as information processors in the GWT framework. They dynamically integrate information from multiple KDs, forming transient coalitions of neural activity that compete for access to a Global Workspace. Each thoughtseed, with its associated core and subordinate attractors, represents a candidate for conscious access and vying to shape the content of awareness. These attractors can be seen as metastable states, aligning with the brain’s pullback attractor dynamics [11].
3.3.2. Competition for Dominance
Thoughtseeds compete in a winner-take-all dynamic, where the one that minimizes the cumulative expected free energy (EFE), balancing both epistemic (uncertainty reduction) and pragmatic (goal achievement) considerations [78], gains access to the Global Workspace. This process is akin to the “ignition” of a neural assembly [25], in which a critical threshold of neural activity is reached, leading to a self-sustaining pattern of activation.
Once a thoughtseed becomes dominant, it triggers a shift in global brain activity, reflecting a change in the content of consciousness. This mechanism aligns with the theory of discrete conscious states, in which consciousness is seen as a sequence of transient attractor states that emerge through coordinated neural activity [26,82]. The winning thoughtseed, after gaining access to the Global Workspace, enables the “broadcast” of its content across the cognitive system, influencing processes such as attention, decision-making, and motor planning, facilitating adaptive behavior [24,83].
In this way, the global workspace functions as a “broadcasting platform,” while KDs act as the “sources” of specialized knowledge, and thoughtseeds serve as the “processors” that dynamically integrate this knowledge to form the content of consciousness [74]. Through this mechanism, thoughtseeds shape both perception and action, allowing the cognitive system to respond flexibly to changing environmental demands.
3.3.3. The Unitary Nature of Conscious Experience
The unitary nature of conscious experience, the subjective feeling of a single, coherent stream of consciousness, emerges from the dominance of a single thoughtseed at any given moment. This dominance reflects the winner-take-all dynamic inherent in the Global Workspace Theory (GWT), ensuring that only one thoughtseed broadcasts its content globally, while other competing thoughtseeds remain inactive or latent. Continuous competition maintains the coherence and integration of conscious experience, preventing fragmented or conflicting streams of awareness from simultaneously entering the workspace [36].
Thus, thoughtseeds do not merely compete for access to the global workspace, but actively contribute to the unitary and seamless nature of conscious experience. Once a thoughtseed gains dominance, it effectively “controls”‘ the global workspace, shaping the current content of consciousness, and guiding future actions and decisions. This guidance is achieved through policy selection that balances epistemic (learning) and pragmatic (goal-driven) affordances [80].
This cyclical process of competition, selection, and dominance underpins the seamless flow of conscious experience, aligning with the brain’s overarching goal of minimizing surprise and maximizing adaptive efficiency through active inference.
3.4. Meta-Cognition and Agency in Thoughtseed Dynamics
Meta-cognition refers to the ability to monitor, evaluate, and regulate one’s cognitive processes. In the context of thoughtseeds and the GWT, meta-cognition plays a crucial role in the orchestration of thoughtseed dynamics, influencing which thoughtseed is selected as dominant and how it interacts with broader agent-level goals and policies [84]. This suggests that conscious access to information is a selective process mediated by prefrontal cortical activity [36].
3.4.1. Meta-Cognitive Regulation of Thoughtseeds
Meta-cognitive processes, potentially implemented by higher-order thoughtseeds, help modulate activation and competition among lower-level thoughtseeds by dynamically adjusting attention, prediction, and error signals. These higher-order thoughtseeds can be seen as “observers” or “regulators” within the cognitive system, evaluating the relevance and coherence of lower-level thoughtseeds and influencing their access to the Global Workspace.
Thoughtseeds compete for dominance based on their ability to minimize free energy, but metacognition introduces a higher level of control by shaping attention and prior expectations. For instance, attentional control can amplify the precision of certain sensory signals, enhancing the chance of relevant thoughtseeds being selected for conscious access [85]. The attentional precision parameter plays a crucial role in this modulation, affecting the sensitivity of lower-level thoughts to sensory evidence during the free energy minimization process. By adjusting this parameter, higher-order thoughtseeds can influence which lower-order thoughtseeds gain prominence in competition for conscious access. This selection process, contributes to the discrete yet unitary nature of conscious experience, where transitions between dominant thoughts correspond to distinct shifts in conscious content.
Moreover, meta-cognition adjusts the activation threshold that determines which thoughtseeds enter the active thoughtseed pool, thereby influencing the pool’s composition. This threshold can vary based on current cognitive or emotional states. For example, in states of high arousal or focused attention, the threshold may be lowered, thereby allowing a broader range of thoughtseeds to manifest. Conversely, in states of reduced attentional control, only highly salient thoughtseeds surpass the threshold [86]. The meta-awareness parameter enriches this process by representing the higher-order thoughtseed’s awareness of its influence on lower-level thoughtseeds and attentional focus. This meta-awareness can be modeled as a probabilistic belief about the degree of opacity in accessing and understanding lower-level thoughtseeds and their associated Knowledge Domains (KDs). It serves as a metacognitive monitoring and control mechanism [87,88].
3.4.2. Agency and Goal-Directed Thoughtseed Dynamics
Agency in this context refers to the ability of the cognitive system to act intentionally and adaptively by selecting the most appropriate thoughtseeds based on its goals. Thoughtseeds, acting as sub-agents within the larger cognitive architecture, contribute to the agent’s overarching goals by driving behavior that aligns with the minimization of long-term surprise or free energy [76].
Higher-level thoughtseeds, which represent more abstract or long-term goals, act as agents of control that guide the system’s overall direction. These thoughtseeds exert a top-down influence on the selection and competition process, ensuring that the selected thoughtseed aligns with the global goals and broader objectives of the agent. For example, a higher-order goal (e.g., finding food) might bias the selection of thoughtseeds that prioritize related affordances within the environment, such as detecting food sources or planning routes to them [89].
3.4.3. Policies, Goals, and Affordances at the Agent Level
At the agent level, thoughtseeds help shape the perceptions of policies (strategies), goals, and affordances (opportunities for action) within the environment. These elements guide the adaptive behavior of the living system by integrating sensory data with internal models of the world. The active inference framework posits that the agent constantly selects policies that minimize EFE, balancing epistemic affordances (opportunities to reduce uncertainty or gain knowledge) with pragmatic affordances (opportunities to fulfill goals or needs) [90].
From this perspective, meta-cognition plays a key role in determining which goals are prioritized and how policies are enacted. As the agent navigates its environment, thoughtseeds help to represent and update predictions regarding the most relevant actions and affordances. This aligns with the notion of a hierarchical policy structure, in which lower-level thoughtseeds handle immediate, task-specific actions and higher-level thoughtseeds manage broader, long-term goals and strategies [91]. The brain’s ergodic principles ensure that these higher-level goals are regularly revisited, influencing future behavior and keeping the system aligned with long-term adaptive objectives [92].
An illustrative mathematical formalism is discussed in Supplementary Section.
4. Discussion and Future Directions
4.1. Towards a General Theory of Embodied Cognition
The Thoughtseed Framework, based on the concept of neuronal packets [27,29,30,94] and the Free Energy Principle (FEP) [15], offers a promising foundation for a general theory of embodied cognition [95,96,97]. Within this framework, thoughtseeds—emergent higher-order constructs—arise from coordinated activity across knowledge domains (KDs). These thoughtseeds represent coherent patterns of neural activity that serve as agentic constructs, driving actions, perception, and cognitive function. This aligns closely with the Global Workspace Theory (GWT) [25,35] and perspectives that emphasize the emergence of intelligence through the navigation of both physical and abstract spaces [65]. Thoughtseeds can be seen as high-salience representations that gain access to the global workspace, thereby influencing conscious experiences. Furthermore, the thoughtseed framework aligns with the notion of discrete thought processes, in which each dominant thoughtseed represents a distinct cognitive moment or “atom” of conscious experience [24].
The thoughtseed framework introduces the novel concept of thoughtseeds as self-organizing units with their own internal states and dynamics, which sets it apart from other cognitive theories. These self-organizing dynamics, driven by the principles of active inference, allow thoughtseeds to actively engage with the environment, predict future states, and shape the ongoing flow of conscious experience. The active and embodied nature of thoughtseeds provides a unique perspective on how cognition emerges from the interaction between the living system and its environment.
As seen in this framework, cognition is not merely an internal process. It is inherently embodied and situated, and deeply rooted in the interactions between the internal states of the living system, its body, and the environment [97]. Thoughtseeds emerge from this reciprocal relationship and shape the living system’s perception and decision-making processes by forming predictions, selecting actions, and integrating sensory feedback.
4.2. The Hierarchical Structure of the Thoughtseed Framework
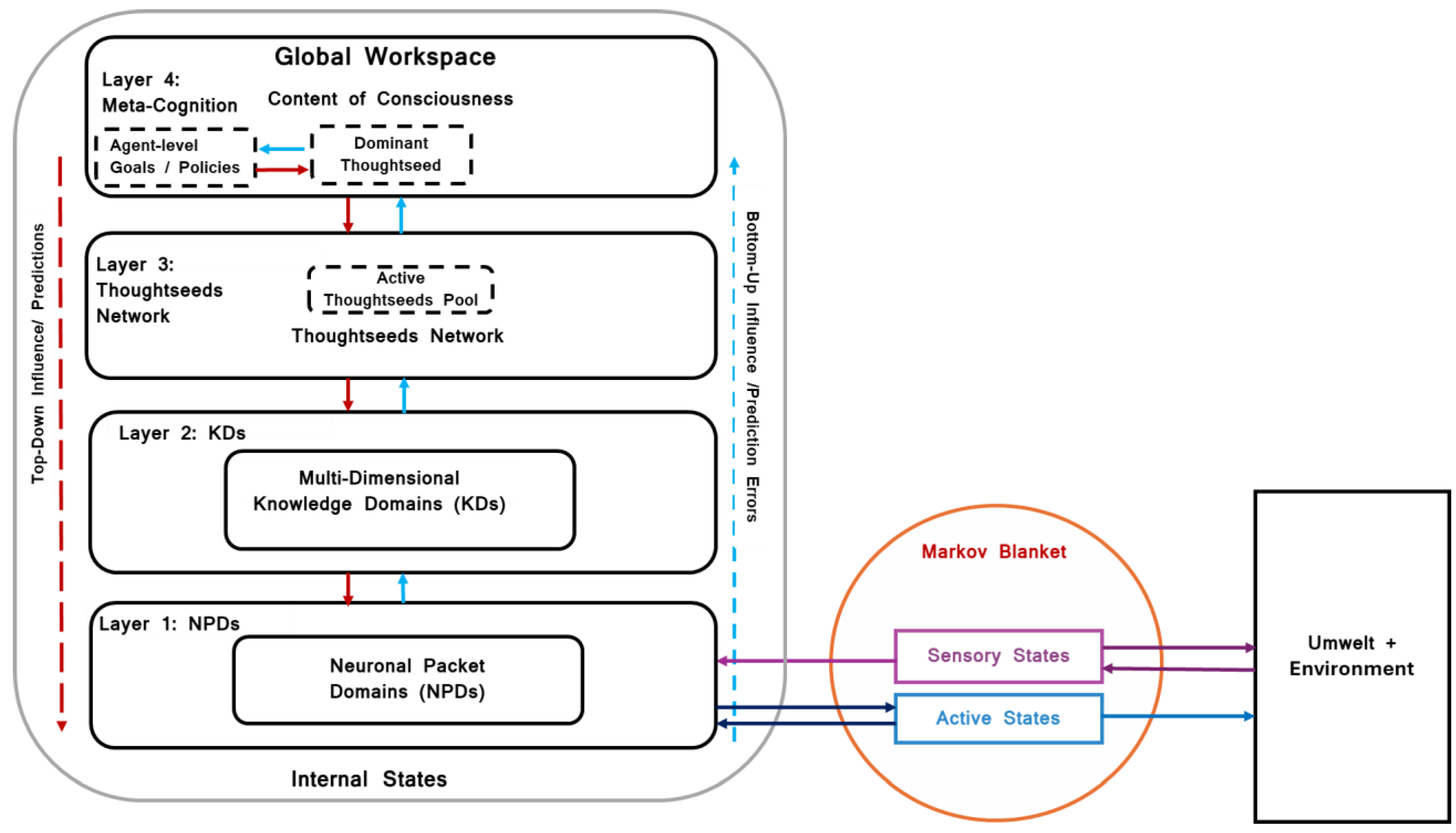
The Thoughtseed Framework proposes a hierarchical organization of cognitive processes (see Figure 4), offering a mechanistic explanation of how the brain organizes and manages complex functions such as decision-making, problem-solving, and planning. These processes can be understood within the Global Workspace Theory (GWT) paradigm, where information from various subsystems becomes globally accessible to the living system when it reaches a level of conscious awareness. Below is an outline of this hierarchy:
- Neuronal Packet Domains (NPDs): NPDs form the foundation of the framework and consist of groups of neurons that process sensory input or generate potential actions. Within the GWT, these subsystems provide foundational and localized information processing. NPDs can be divided into domains responsible for sensory processing, motor planning, and internal state regulation, which contribute raw data to higher-order representations.
- Knowledge Domains (KDs): KDs represent large-scale networks that integrate knowledge, memories, and conceptual categories. They serve as structured repositories from which thoughtseeds are drawn. Within the GWT, KDs act as functional modules that hold specialized knowledge and are accessible to the global workspace when relevant. For example, recognizing a dog involves KDs related to visual processing, auditory associations (barking), and emotional contexts (companionship).
- Thoughtseeds Network: Thoughtseeds represent dynamic, self-organizing entities that emerge from the coordinated activity within and across KDs. In the GWT context, thoughtseeds represent potential candidates for entry into the global workspace, where they can shape conscious experiences and direct behavior. Each thoughtseed is a coherent pattern of neural activity associated with specific concepts, percepts, or possible actions. Thoughtseeds act as cognitive sub-agents, actively generating predictions and selecting actions to minimize free energy, and constantly updating based on sensory feedback.
- Meta-Cognition, Dominant Thoughtseed and Higher-Order Thoughtseeds: This level involves higher-order thoughtseeds that modulate attention and guide goal-directed behavior. Higher-order thoughtseeds reflect the agent’s global goals, influencing which lower-level thoughtseeds gain prominence in the global workspace. In terms of GWT, these higher-order constructs govern the selection of conscious content by modulating attentional precision and prioritizing relevant information. This helps shape global policies that determine the actions of the living system in a goal-oriented manner. The selection of a dominant thoughtseed can be seen as a discrete event, leading to a distinct shift in conscious content.
The culmination of these hierarchical processes is the projection of content onto the Global Workspace–the metaphorical stage of conscious experience. The dominant thoughtseed at any moment, selected through competitive processes based on free energy minimization and influenced by metacognitive processes, shapes the conscious content. This content includes sensory input, memories, concepts, and potential actions, reflecting the current understanding of the living system and its interactions with the environment. The dynamic interplay between thoughtseeds and knowledge domains creates the fluid “stream of thoughts” that manifests as the content of consciousness, guiding adaptive behavior and decision-making.
4.3. Limitations and Future Research Directions
Investigating thoughtseed formation requires a multi-scale approach, encompassing neuronal co-activation and the development of stable Markov blankets representing distinct attractor states. The assumption of supervenience—that consciousness is solely reliant on physical states—presents limitations by challenging the empirical distinguishability required for theories of consciousness to advance scientifically [99]. This suggests that research on thoughtseed formation could benefit from integrating non-reductive frameworks, where consciousness emerges from, yet is not entirely reducible to, physical systems.
Moreover, creating biologically plausible computational models capable of simulating spiking neural networks with synaptic plasticity and homeostatic mechanisms remains a complex challenge. Such models could allow researchers to track the evolution of attractor state stability and precision, particularly as these states are embedded within nested Markov blankets that support thoughtseed resilience [30,42,100]. Extending these models to capture ontological coherence and empirical discernibility could help identify conditions under which thoughtseeds transition between states, drawing on dynamics observed in memory models [102,103]. Future work should aim to incorporate emergent, hierarchical structures into computational frameworks to enhance the measurability and discernibility of conscious experience [99,101]. Identifying neural signatures such as spatiotemporal activity patterns, connectivity motifs, or oscillatory dynamics may provide critical insights into thoughtseed formation and resilience [104,105,106].
4.4. Key Limitations
- Metastability of Thoughtseeds: Thoughtseed dynamics are inherently metastable with rapid transitions between states. This makes it difficult to isolate core attractors representing stable knowledge or behaviors, posing challenges in empirically capturing and measuring the dynamic properties of thoughtseeds [52,107,108,109].
- Mapping Distributed Neural Activity: Given the complexity of the brain’s distributed networks, linking specific patterns of neural activity to cognitive processes (e.g., thoughtseeds) is fraught with difficulty. Misinterpretation can arise without a clear framework that maps distributed signals to well-defined cognitive domains [110,111].
- Hierarchical Complexity: The nested Markov blanket structure of thoughtseeds introduces additional complexity in understanding how higher- and lower-order processes interact. This complicates the experimental design and analysis, especially when attempting to link abstract thought processes to neurobiological signatures [31,34].
- Vast Repertoire of Brain States: The extensive diversity of potential brain states and thoughtseed configurations presents a significant challenge for both empirical investigation and computational modeling. The isolation and characterization of specific thoughtseed dynamics within this vast repertoire necessitates meticulous experimental design and sophisticated analytical techniques [107,108,109,110,111]. This limitation underscores the need for focused research paradigms, such as the study of meditation or deep focus states, wherein the repertoire of brain states may be more constrained and specific thoughtseed trajectories can be more readily identified.
4.5. Future Directions
Future research should focus on several key areas for further development and validation of the thoughtseed framework:
- Computational Modeling: Given the vast repertoire of potential brain states, developing a comprehensive computational model that captures the full spectrum of thoughtseed dynamics presents a significant challenge. Consequently, the analysis of thoughtseed dynamics is particularly suited to focused attention contexts, such as meditation or deeply focussed actions. These conditions, characterized by reduced mind-wandering and enhanced attentional precision, facilitate more precise observation of thoughtseed trajectories and their interactions. Simulating thoughtseed transitions as multi-dimensional trajectories could enable the generation of testable hypotheses regarding their neural correlates, facilitating comparisons with fMRI data, particularly in relation to neural assembly ignition [24]. By adopting the Leading Eigenvector Dynamics Analysis (LEiDA) and Probabilistic Metastable Substate (PMS) framework [112], thoughtseeds can be represented as dynamic trajectories in a multi-dimensional state space derived from neuroimaging data. This biologically grounded and empirically testable approach allows researchers to align hypothesized thoughtseeds—such as mind-wandering, breath focus, and meta-awareness—with specific neural patterns. Such alignment could provide critical insights into how meditation practices influence cognitive dynamics and conscious experience.
An alternative approach could involve focusing on specific aspects of thoughtseed dynamics, integrating insights from computational phenomenology [88] and higher-order Partially Observable Markov Decision Processes (POMDPs). This approach would provide a robust framework for modeling the probabilistic and decision-based nature of thoughtseed transitions, complementing renormalization group (RG) approaches that remain valuable for understanding multi-scale dynamics.
- Cognitive Development: Investigate how the thoughtseed framework can enhance our understanding of cognitive development across the lifespan, particularly in learning, memory, and complex skill acquisition. Early research could target specific domains, such as object permanence [113] or numerical cognition [114], to observe how thoughtseeds emerge and evolve in these processes. Additionally, the development of thoughtseeds may also be investigated in the context of the emergence of discrete symbols and language [115,116].
- Clinical Applications: Explore the thoughtseed framework’s potential in understanding and treating attention disorders by examining how disruptions in thoughtseed dynamics may contribute to specific clinical conditions. Additionally, investigate whether interventions targeting thoughtseed regulation can enhance cognitive function in these contexts.
By pursuing these research directions, the thoughtseed framework can be further refined and validated, leading to a deeper understanding of cognition, consciousness, and their underlying neural mechanisms.
5. Conclusion
The thoughtseed framework offers a novel and biologically plausible model for understanding the emergence and the organizing principles of embodied cognition. By integrating evolutionary principles, active inference, and Global Workspace Theory, the framework provides a comprehensive account of how cognitive processes arise from the dynamic interplay of neuronal packets, knowledge domains, and higher-order thoughtseeds. The framework’s emphasis on self-organization, competition, and free energy minimization sheds light on the mechanisms underlying thought formation, conscious experience, and adaptive behavior.
The hierarchical structure of the framework, with its nested Markov blankets and reciprocal message passing, provides a compelling explanation of the brain’s ability to integrate information across different levels of representation and generate coherent cognitive states. The concept of thoughtseeds as self-organizing units of embodied knowledge offers a promising new perspective on the nature of thought and its role in shaping conscious experiences.
While still in its theoretical stages, the thoughtseed framework provides fertile ground for future research, with potential applications in understanding cognitive development, psychopathology, and artificial intelligence. By continuing to explore the dynamics of thoughtseeds and their interactions within the Global Workspace, we can gain a deeper understanding of the organizing principles governing cognition and consciousness.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Institutional Review Board Statement
NA
Author Contributions
Conceptualization, Prakash Kavi (Thoughtseeds) and Daniel Friedman (Active Inference Formlization); Methodology, Prakash Kavi; Writing – original draft, Prakash Kavi; Writing – review & editing, Prakash Kavi, Gorka Zamora-López and Daniel Friedman.
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author(s).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Futuyma, D.J. Evolutionary Biology. Trends Cogn. Sci. 2017, 21, 500–501. [Google Scholar]
- Baldwin, J.M. A New Factor in Evolution. Am. Nat. 1896, 30, 441–451. [Google Scholar] [CrossRef]
- Darwin, C. On the Origin of Species; John Murray, 1859. [Google Scholar]
- Brooke, J.L.; Larsen, C.S. The Nurture of Nature: Genetics, Epigenetics, and Environment in Human Biohistory. The American Historical Review 2014, 119, 1500–1513. [Google Scholar] [CrossRef]
- Boyd, R.; Richerson, P.J. Culture and the Evolutionary Process; University of Chicago Press, 1988. [Google Scholar]
- Laland, K.N.; Odling-Smee, J.; Feldman, M.W. Niche Construction, Biological Evolution, and Cultural Change. Behavioral and Brain Sciences 2000, 23, 131–146. [Google Scholar] [CrossRef]
- Odling-Smee, F.J.; Laland, K.N.; Feldman, M.W. Niche Construction: The Neglected Process in Evolution; Princeton University Press, 2003. [Google Scholar]
- Varela, F.J.; Thompson, E.; Rosch, E. The Embodied Mind: Cognitive Science and Human Experience; MIT Press, 1991. [Google Scholar]
- Noë, A. Action in Perception; MIT Press, 2004. [Google Scholar]
- Maturana, H.R.; Varela, F.J. Autopoiesis and Cognition: The Realization of the Living; D. Reidel Publishing Company, 1980. [Google Scholar]
- Friston, K.; Ao, P. Free Energy, Value, and Attractors. Comput. Math. Methods Med. 2012, 2012, 937860. [Google Scholar] [CrossRef] [PubMed]
- Schrödinger, E. What Is Life? The Physical Aspect of the Living Cell; Cambridge University Press, 1944. [Google Scholar]
- Nicolis, G.; Prigogine, I. Self-Organization in Nonequilibrium Systems; Wiley, 1977. [Google Scholar]
- Conant, R.C.; Ross Ashby, W.R. Every Good Regulator of a System Must Be a Model of That System. Int. J. Syst. Sci. 1970, 1, 89–97. [Google Scholar] [CrossRef]
- Friston, K.J. The Free Energy Principle: A Unified Approach to Biological Systems. Philos. Trans. R. Soc. Lond. B 2010, 366, 2257–2268. [Google Scholar]
- Friston, K.; Kilner, J.; Frith, C. A Free Energy Formulation of Active Inference. Front. Comp. Neurosci. 2010, 4, 168. [Google Scholar]
- Parr, T.; Friston, K.J. Working Memory as Active Inference. Biol. Cybern. 2018, 112, 417–443. [Google Scholar]
- Friston, K.; Parr, T.; De Vries, P. The Graphical Brain: A Framework for Understanding Brain Function. Neurosci. Biobehav. Rev. 2021, 127, 16–30. [Google Scholar]
- Friston, K.; Heins, C.; Verbelen, T.; Da Costa, L.; Salvatori, T.; Markovic, D.; Tschantz, A.; Koudahl, M.; Buckley, C.; Parr, T. From Pixels to Planning: Scale-Free Active Inference. arXiv 2024. [Google Scholar] [CrossRef]
- Badcock, P.B.; Friston, K.J.; Ramstead, M.J.D.; Ploeger, A.; Hohwy, J. The Hierarchically Mechanistic Mind: An Evolutionary Systems Theory of the Human Brain, Cognition, and Behavior. Cogn. Affect. Behav. Neurosci. 2019, 19, 1319–1351. [Google Scholar] [CrossRef] [PubMed]
- Fodor, J.A. The Modularity of Mind: An Essay on Faculty Psychology; MIT Press, 1983. [Google Scholar]
- Josselyn, S.A.; Tonegawa, S. Memory Engrams: Recalling the past and Imagining the Future. Science 2020, 367, eaaw4325. [Google Scholar] [CrossRef]
- Hebb, D.O. The Organization of Behavior; Wiley & Sons, 1949. [Google Scholar]
- Dehaene, S. Consciousness and the Brain: Deciphering How the Brain Codes Our Thoughts; Viking Press, 2014. [Google Scholar]
- Dehaene, S.; Changeux, J.-P. Experimental and Theoretical Approaches to Conscious Processing. Neuron 2011, 70, 200–227. [Google Scholar] [CrossRef]
- Dehaene, S.; Sergent, C.; Changeux, J.-P. A Neuronal Network Model Linking Subjective Reports and Objective Physiological Data During Conscious Perception. Proc. Natl. Acad. Sci. U. S. A. 2003, 100, 8520–8525. [Google Scholar] [CrossRef] [PubMed]
- Yufik, Y.M.; Friston, K. Life and Understanding: The Origins of ‘Understanding‘ in Self-Organizing Nervous Systems. Front. Syst. Neurosci. 2016, 10, 98. [Google Scholar] [CrossRef]
- Palacios, E.R.; Razi, A.; Parr, T.; Kirchhoff, M.; Friston, K. On Markov Blankets and Hierarchical Self-Organisation. J. Theor. Biol. 2020, 486, 110089. [Google Scholar] [CrossRef]
- Ramstead, M.J.D.; Hesp, C.; Tschantz, A.; Smith, R.; Constant, A.; Friston, K. Neural and Phenotypic Representation Under the Free-Energy Principle. Neurosci. Biobehav. Rev. 2021, 120, 109–122. [Google Scholar] [CrossRef]
- Yufik, Y.M. The Understanding Capacity and Information Dynamics in the Human Brain. Entropy (Basel, Switzerland) 2019, 21, 308. [Google Scholar] [CrossRef]
- Kirchhoff, M.; Parr, T.; Palacios, E.; Friston, K.; Kiverstein, J. The Markov Blankets of Life: Autonomy, Active Inference and the Free Energy Principle. J. R. Soc. Interface 2018, 15, 20170792. [Google Scholar] [CrossRef]
- Ramstead, M.J.D.; Kirchhoff, M.D.; Friston, K.J. A Tale of Two Densities: Active Inference Is Enactive Inference. Adapt. Behav. 2020, 28, 225–239. [Google Scholar] [CrossRef] [PubMed]
- Ramstead, M.J.; Albarracin, A.M.; Kiefer, A.; Klein, B.; Fields, C.; Friston, K.; Safron, A. The Inner Screen Model of Consciousness: Applying the Free Energy Principle Directly to the Study of Conscious Experience. Neurosci. Conscious. 2023, 2023, niac013. [Google Scholar]
- Friston, K.J.; Wiese, W.; Hobson, J.A. Sentience and the Origins of Consciousness: From Cartesian Duality to Markovian Monism. Entropy 2020, 22, 516. [Google Scholar] [CrossRef] [PubMed]
- Baars, B.J. the Theater of Consciousness: The Workspace of the Mind; Oxford University Press, 1997. [Google Scholar] [CrossRef]
- Dehaene, S.; Naccache, L. Towards a Cognitive Neuroscience of Consciousness: Basic Evidence and a Workspace Framework. Cognition 2001, 79, 1–37. [Google Scholar] [CrossRef]
- Fields, C.; Glazebrook, J.F.; Levin, M. Minimal physicalism as a scale-free substrate for cognition and consciousness. Neuroscience of Consciousness 2021, 2021, niab013. [Google Scholar] [CrossRef]
- von Uexküll, J. A Stroll Through the Worlds of Animals and Men: A Picture Book of Invisible Worlds. In. Semiotica; Springer, 1934; 89 (4), (5–80). [Google Scholar] [CrossRef]
- Friston, K.; Da Costa, L.; Sakthivadivel, D.A.R.; Heins, C.; Pavliotis, G.A.; Ramstead, M.; Parr, T. Path Integrals, Particular Kinds, and Strange Things. Phys. Life Rev. 2023, 47, 35–62. [Google Scholar] [CrossRef]
- Bullmore, E.; Sporns, O. Complex Brain Networks: Graph Theoretical Analysis of Structural and Functional Systems. Nat. Rev. Neurosci. 2009, 10, 186–198. [Google Scholar] [CrossRef]
- Sporns, O.; Betzel, R.F. Modular Brain Networks. Annu. Rev. Psychol. 2016, 67, 613–640. [Google Scholar] [CrossRef]
- Hipólito, I.; Ramstead, M.J.D.; Convertino, L.; Bhat, A.; Friston, K.; Parr, T. Markov Blankets in the Brain. Neurosci. Biobehav. Rev. 2021, 125, 88–97. [Google Scholar] [CrossRef]
- Douglas, R.J.; Martin, K.A. A Functional Microcircuit for Cat Visual Cortex. J. Physiol. 1991, 440, 735–769. [Google Scholar] [CrossRef]
- Mountcastle, V.B. The Columnar Organization of the Neocortex. Brain 1997, 120, 701–722. [Google Scholar] [CrossRef] [PubMed]
- Quartz, S.R.; Sejnowski, T.J. The Neural Basis of Cognitive Development: A Constructivist Manifesto. Behav. Brain Sci. 1997, 20, 537–56. [Google Scholar] [CrossRef]
- Zeki, S. A Vision of the Brain; Blackwell Scientific Publications, 1993. [Google Scholar]
- DiCarlo, J.J.; Zoccolan, D.; Rust, N.C. How Does the Brain Solve Visual Object Recognition? Neuron 2012, 73, 415–434. [Google Scholar] [CrossRef] [PubMed]
- Markov, N.T.; Ercsey-Ravasz, M.M.; Ribeiro Gomes, A.R.; Lamy, C.; Magrou, L.; Vezoli, J.; Misery, P.; Falchier, A.; Quilodran, R.; Gariel, M.A.; Sallet, J.; Gamanut, R.; Huissoud, C.; Clavagnier, S.; Giroud, P.; Sappey-Marinier, D.; Barone, P.; Dehay, C.; Toroczkai, Z.; Knoblauch, K.; Van Essen, D.C.; Kennedy, H. A Weighted and Directed Interareal Connectivity Matrix for Macaque Cerebral Cortex. Cereb. Cortex 2014, 24, 17–36. [Google Scholar] [CrossRef]
- Edelman, G.M. Neural Darwinism: The Theory of Neuronal Group Selection; Basic Books, 1987. [Google Scholar]
- Riesenhuber, M.; Poggio, T. Hierarchical Models of Object Recognition in Cortex. Nat. Neurosci. 1999, 2, 1019–1025. [Google Scholar] [CrossRef] [PubMed]
- Kiebel, S.J.; Daunizeau, J.; Friston, K.J. A Hierarchy of Time-Scales and the Brain. PLoS Comput. Biol. 2008, 4, e1000209. [Google Scholar] [CrossRef]
- Rabinovich, M.I.; Huerta, R.; Laurent, G. Neuroscience. Transient Dynamics for Neural Processing. Science 2008, 321, 48–50. [Google Scholar] [CrossRef]
- Pisarchik, A.N.; Serrano, N.P.; Jaimes-Reátegui, R. Brain Connectivity Hypergraphs. 2024 8th Scientific School Dynamics of Complex Networks and their Applications (DCNA), Kaliningrad, Russian Federation, 2024, pp. 190-193. [CrossRef]
- Pessoa, L. Understanding Brain Networks and Brain Organization. Phys. Life Rev. 2014, 11, 400–435. [Google Scholar] [CrossRef]
- Friston, K. Hierarchical Models in the Brain. PLOS Comp. Biol. 2008, 4, e1000211. [Google Scholar] [CrossRef]
- Zamora-López, G.; Zhou, C.S.; Kurths, J. Exploring Brain Function from Anatomical Connectivity. Front. Neurosci. 2011, 5, 83. [Google Scholar] [CrossRef]
- Spelke, E.S.; Kinzler, K.D. Core Knowledge. Dev. Sci. 2007, 10, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Pattisapu, C.; Verbelen, T.; Pitliya, R.J.; Kiefer, A.B.; Albarracin, M. Free Energy in a Circumplex Model of Emotion. arXiv Preprint 2024, arXiv:2407.02474. [Google Scholar]
- Sladky, R.; Hahn, A.; Karl, I.L.; Geissberger, N.; Kranz, G.S.; Tik, M.; Kraus, C.; Pfabigan, D.M.; Gartus, A.; Lanzenberger, R.; Lamm, C. Dynamic Causal Modeling of the Prefrontal/Amygdala Network During Processing of Emotional Faces. Brain Connect. 2022, 12, 670–682. [Google Scholar] [CrossRef]
- Solms, M. The Hidden Spring: A Journey to the Source of Consciousness; Profile Books, 2021. [Google Scholar]
- Singer, W. Neuronal Synchrony: A Versatile Code for the Definition of Relations? Neuron 1999, 24, 49–65. [Google Scholar] [CrossRef]
- Palacios, E.R.; Isomura, T.; Parr, T.; Friston, K. The Emergence of Synchrony in Networks of Mutually Inferring Neurons. Sci. Rep. 2019, 9, 6412. [Google Scholar] [CrossRef] [PubMed]
- Roelfsema, P.R. Solving the Binding Problem: Assemblies Form When Neurons Enhance Their Firing Rate-They Don’t Need to Oscillate or Synchronize. Neuron 2023, 111, 1003–1019. [Google Scholar] [CrossRef] [PubMed]
- Friston, K.J.; Kiebel, S.J. Predictive Coding Under the Free-Energy Principle. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2009, 364, 1211–1221. [Google Scholar] [CrossRef] [PubMed]
- Safron, A.; Çatal, O.; Verbelen, T. Generalized Simultaneous Localization and Mapping (G-SLAM) as Unification Framework for Natural and Artificial Intelligences: Towards Reverse Engineering the Hippocampal/Entorhinal System and Principles of High-Level Cognition. Front. Syst. Neurosci. 2022, 16, 787659. [Google Scholar] [CrossRef]
- Squire, L.R.; Wixted, J.T.; Clark, R.E. Recognition Memory and the Medial Temporal Lobe: A New Perspective. Nat. Rev. Neurosci. 2007, 8, 872–883. [Google Scholar] [CrossRef]
- Eichenbaum, H. The Hippocampus and Its Role in Navigation and Memory: A Twenty-Five-Year Perspective. Neuron 2017, 95, 644–661. [Google Scholar]
- Stachenfeld, K.L.; Botvinick, M.M.; Gershman, S.J. The Hippocampus as a Predictive Map. Nat. Neurosci. 2017, 20, 1643–1653. [Google Scholar] [CrossRef] [PubMed]
- LeDoux, J.E.; Brown, R. A Higher-Order Theory of Emotional Consciousness. Proc. Natl. Acad. Sci. U. S. A. 2017, 114, E2016–E2025. [Google Scholar] [CrossRef] [PubMed]
- Phelps, E.A.; LeDoux, J.E. Contributions of the Amygdala to Emotion Processing: From Animal Models to Human Behavior. Neuron 2005, 48, 175–187. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.K.; Cohen, J.D. An Integrative Theory of Prefrontal Cortex Function. Annu. Rev. Neurosci. 2001, 24, 167–202. [Google Scholar] [CrossRef] [PubMed]
- Fuster, J.M. The Prefrontal Cortex, 5th ed.; Academic Press, 2015. [Google Scholar]
- Ororbia, A.; Friston, K. Mortal Computation: A Foundation for Biomimetic Intelligence. arXiv Preprint 2023, arXiv:2311.09589. [Google Scholar]
- Mashour, G.A.; Roelfsema, P.; Changeux, J.-P.; Dehaene, S. Conscious Processing and the Global Neuronal Workspace Hypothesis. Neuron 2020, 105, 776–798. [Google Scholar] [CrossRef]
- Friston, K.; SenGupta, B.; Auletta, G. Cognitive Dynamics: From Attractors to Active Inference. Proc. IEEE 2014, 102, 427–445. [Google Scholar] [CrossRef]
- Seth, A.K. The Cybernetic Bayesian Brain: From Interoceptive Inference to Sensorimotor Contingencies. In Open MIND; Metzinger, T.K., Windt, J.M., Eds.; MIND Group, 2015; pp. 1–24. [Google Scholar]
- Haken, H. Synergetics: An Introduction. Non-equilibrium Phase Transitions and Self-Organization in Physics, Chemistry, and Biology (3rd Rev. enl. Ed.); Springer-Verlag, 1983. [Google Scholar]
- Parr, T.; Friston, K.J. Generalised Free Energy and Active Inference. Biol. Cybern. 2019, 113, (5–6). [Google Scholar] [CrossRef]
- Gibson, J.J. The Theory of Affordances. Hilldale, USA 1977, 1, 67–82. [Google Scholar]
- Friston, K.; Frith, C. A Duet for One. Conscious. Cogn. 2015, 36, 390–405. [Google Scholar] [CrossRef]
- Friston, K. Affordance and Active Inference. In Affordances in Everyday Life: A Multidisciplinary Collection of Essays; Springer International Publishing, 2022. [Google Scholar]
- Deco, G.; Ponce-Alvarez, A.; Mantini, D.; Romani, G.L.; Hagmann, P.; Corbetta, M. Resting-State Functional Connectivity Emerges from Structurally and Dynamically Shaped Slow Linear Fluctuations. J. Neurosci. 2013, 33, 11239–11252. [Google Scholar] [CrossRef] [PubMed]
- Deco, G.; Tononi, G.; Boly, M.; Kringelbach, M.L. Rethinking Segregation and Integration: Contributions of Whole-Brain Modelling. Nat. Rev. Neurosci. 2015, 16, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Desimone, R.; Duncan, J. Neural Mechanisms of Selective Visual Attention. Annu. Rev. Neurosci. 1995, 18, 193–222. [Google Scholar] [CrossRef] [PubMed]
- Feldman, H.; Friston, K.J. Attention, Uncertainty, and Free-Energy. Front. Hum. Neurosci. 2010, 4, 215. [Google Scholar] [CrossRef]
- Corbetta, M.; Shulman, G.L. Control of Goal-Directed and Stimulus-Driven Attention in the Brain. Nat. Rev. Neurosci. 2002, 3, 201–215. [Google Scholar] [CrossRef]
- Metzinger, T. Phenomenal Transparency and Cognitive Self-Reference. Phenomenol. Cogn. Sci. 2003, 2, 353–393. [Google Scholar] [CrossRef]
- Sandved-Smith, L.; Hesp, C.; Mattout, J.; Friston, K.; Lutz, A.; Ramstead, M.J.D. Towards a Computational Phenomenology of Mental Action: Modelling Meta-awareness and Attentional Control with Deep Parametric Active Inference. Neurosci. Conscious. 2021, 2021, niab018. [Google Scholar] [CrossRef]
- Friston, K. Life as We Know It. J. R. Soc. Interface 2013, 10, 20130475. [Google Scholar] [CrossRef]
- Friston, K.; Rigoli, F.; Ognibene, D.; Mathys, C.; Fitzgerald, T.; Pezzulo, G. Active Inference and Epistemic Value. Cogn. Neurosci. 2015, 6, 187–214. [Google Scholar] [CrossRef]
- Pezzulo, G.; Rigoli, F.; Friston, K. Active Inference, Homeostatic Regulation and the Origins of Consciousness. J. Conscious. Stud. 2013, 20, 42–64. [Google Scholar]
- Friston, K.J.; Fagerholm, E.D.; Zarghami, T.S.; Parr, T.; Hipólito, I.; Magrou, L.; Razi, A. Parcels and Particles: Markov Blankets in the Brain. Netw. Neurosci. 2021, 5, 211–251. [Google Scholar] [CrossRef] [PubMed]
- Chalmers, D.J. Facing up to the Problem of Consciousness. J. Conscious. Stud. 1995, 2, 200–219. [Google Scholar]
- Yufik, Y.M. Understanding, Consciousness and Thermodynamics of Cognition. Chaos Solitons Fract. 2013, 55, 44–59. [Google Scholar] [CrossRef]
- Allen, M.; Friston, K.J. From Cognitivism to Autopoiesis: Towards a Computational Framework for the Embodied Mind. Synthese 2018, 195, 2459–2482. [Google Scholar] [CrossRef]
- Foglia, L.; Wilson, R.A. Embodied Cognition. Wiley Interdiscip. Rev. Cogn. Sci. 2013, 4, 319–325. [Google Scholar] [CrossRef]
- Pezzulo, G.; D‘Amato, L.; Mannella, F.; Priorelli, M.; Van de Maele, T.; Stoianov, I.P.; Friston, K. Neural Representation in Active Inference: Using Generative Models to Interact With—And Understand—The Lived World. Ann. N. Y. Acad. Sci. 2024, 1534, 45–68. [Google Scholar] [CrossRef] [PubMed]
- Thompson, E. Mind in Life: Biology, Phenomenology, and the Sciences of Mind; Harvard University Press, 2007. [Google Scholar]
- Kleiner, J. Supervenience, Consciousness and Scientific Practice. Presented at the International Association for the Scientific Study of Consciousness (IASSC) Conference, 2022. 2022. [Google Scholar]
- Litwin-Kumar, A.; Doiron, B. Slow Dynamics and High Variability in Balanced Cortical Networks with Clustered Connections. Nat. Neurosci. 2012, 15, 1498–1505. [Google Scholar] [CrossRef]
- Kleiner, J.; Ludwig, T. What Is a Mathematical Structure of Conscious Experience? Synthese 2024, 203, 89. [Google Scholar] [CrossRef]
- Boscaglia, M.; Gastaldi, C.; Gerstner, W.; Quian Quiroga, R. A Dynamic Attractor Network Model of Memory Formation, Reinforcement and Forgetting. PLOS Comp. Biol. 2023, 19, e1011727. [Google Scholar] [CrossRef]
- Parr, T.; Da Costa, L.; Heins, C.; Ramstead, M.J.D.; Friston, K.J. Memory and Markov Blankets. Entropy (Basel) 2021, 23, 1105. [Google Scholar] [CrossRef]
- Buzsáki, G. Rhythms of the Brain; Oxford University Press, 2006. [Google Scholar]
- Fries, P. Rhythms for Cognition: Communication Through Coherence. Neuron 2015, 88, 220–235. [Google Scholar] [CrossRef] [PubMed]
- Sporns, O. Networks of the Brain; MIT Press, 2011. [Google Scholar]
- Tognoli, E.; Kelso, J.A.S. The Metastable Brain. Neuron 2014, 81, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Deco, G.; Kringelbach, M.L. Metastability and Coherence in Brain Dynamics. Trends Cogn. Sci. 2016, 20, 251–257. [Google Scholar]
- Deco, G.; Kringelbach, M.L. Hierarchy of Information Processing in the Brain: A Novel ‘Intrinsic Ignition’ Framework. Neuron 2017, 94, 961–968. [Google Scholar] [CrossRef]
- deCharms, R.C.; Zador, A. Neural Representation and the Cortical Code. Annu. Rev. Neurosci. 2000, 23, 613–647. [Google Scholar] [CrossRef]
- Rué-Queralt, J.; Stevner, A.; Tagliazucchi, E.; Laufs, H.; Kringelbach, M.L.; Deco, G.; Atasoy, S. Decoding Brain States on the Intrinsic Manifold of Human Brain Dynamics Across Wakefulness and Sleep. Commun. Biol. 2021, 4, 854. [Google Scholar] [CrossRef]
- Escrichs, A.; Sanz Perl, Y.; Martínez-Molina, N.; Biarnes, C.; Garre-Olmo, J.; Fernández-Real, J.M.; Ramos, R.; Martí, R.; Pamplona, R.; Brugada, R.; Serena, J.; Ramió-Torrentà, L.; Coll-De-Tuero, G.; Gallart, L.; Barretina, J.; Vilanova, J.C.; Mayneris-Perxachs, J.; Saba, L.; Pedraza, S.; Kringelbach, M.L.; Puig, J.; Deco, G. The Effect of External Stimulation on Functional Networks in the Aging Healthy Human Brain. Cereb. Cortex 2022, 33, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Piaget, J.; Cook, M.T. The Development of Object Concept; 1954. [Google Scholar]
- Nieder, A. The Neuronal Code for Number. Nat. Rev. Neurosci. 2016, 17, 366–382. [Google Scholar] [CrossRef]
- Dehaene, S.; Al Roumi, F.; Lakretz, Y.; Planton, S.; Sablé-Meyer, M. Symbols and mental programs: A hypothesis about human singularity. Trends in Cognitive Sciences 2022, 26, 751–766. [Google Scholar] [CrossRef]
- Dehaene, S. How We Learn: Why Brains Learn Better than Any Machine... for Now; Viking Press, 2020. [Google Scholar]
Figure 2.
Illustrative Free Energy Landscapes of a Neuronal Packet The figure depicts the illustrative free energy landscapes associated with the three potential states of a neuronal packet (NP). The x-axis represents the hypothetical internal states of the NP, while the y-axis represents the free energy associated with each state. The blue curve depicts the unmanifested state, characterized by a relatively flat landscape with shallow local minima, indicating low stability and high susceptibility to change. Upon undergoing a phase transition from the unmanifested state to the manifested state, the NP can be represented by a red curve, featuring a deep local minimum (the core attractor) and a shallower local minimum (a subordinate attractor). The energy barrier associated with the manifested state is depicted by the vertical dashed line, highlighting the energy required for the transition between the core attractor and subordinate attractor. The binding energy, representing the overall stability of the manifested NP, is visualized as the vertical distance between the core attractor and the zero free energy level (horizontal dashed line). The green curve illustrates the activated state, where the core attractor is further deepened, reflecting heightened neural activity and a stronger focus on the NP’s core representation.
Figure 2.
Illustrative Free Energy Landscapes of a Neuronal Packet The figure depicts the illustrative free energy landscapes associated with the three potential states of a neuronal packet (NP). The x-axis represents the hypothetical internal states of the NP, while the y-axis represents the free energy associated with each state. The blue curve depicts the unmanifested state, characterized by a relatively flat landscape with shallow local minima, indicating low stability and high susceptibility to change. Upon undergoing a phase transition from the unmanifested state to the manifested state, the NP can be represented by a red curve, featuring a deep local minimum (the core attractor) and a shallower local minimum (a subordinate attractor). The energy barrier associated with the manifested state is depicted by the vertical dashed line, highlighting the energy required for the transition between the core attractor and subordinate attractor. The binding energy, representing the overall stability of the manifested NP, is visualized as the vertical distance between the core attractor and the zero free energy level (horizontal dashed line). The green curve illustrates the activated state, where the core attractor is further deepened, reflecting heightened neural activity and a stronger focus on the NP’s core representation.
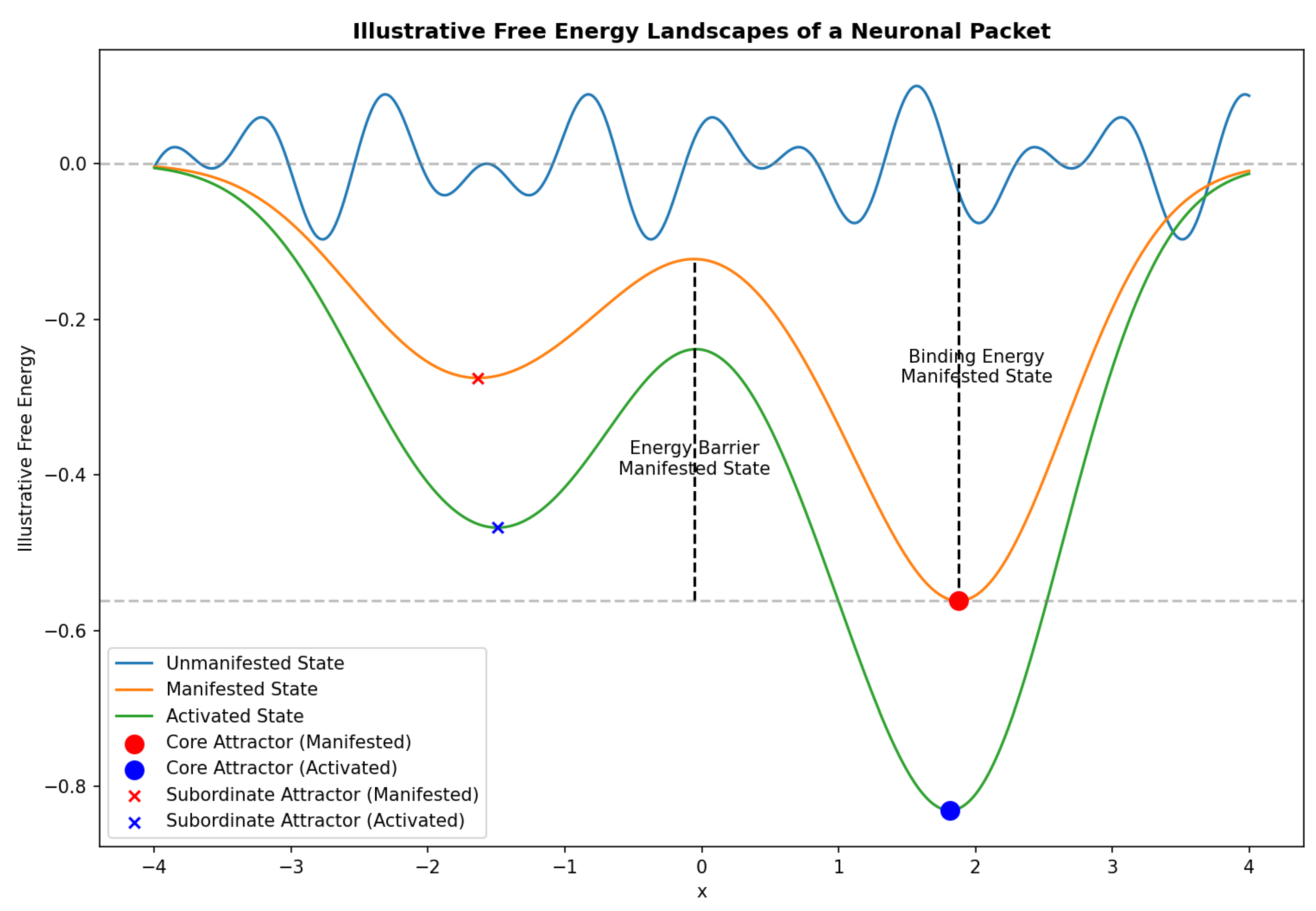
Figure 3.
Neuronal Packet Domains (NPDs) and Knowledge Domains (KDs). This figure provides a static illustration of the relationship between NPDs and KDs at a given time snapshot, highlighting their roles in the thoughtseed framework. KDs: The four distinct colors represent different KDs, each specializing in a particular domain of knowledge or expertise. The unfilled squares symbolize localized KDs, formed as superordinate ensembles of NPDs, integrating information from specific functional units within the brain. The filled squares represent a superordinate ensemble of non-local, heterarchical KDs, suggesting the integration of knowledge across multiple domains. NPDs: The circles depict NPDs, the functional units of the brain responsible for processing specific types of information. They act as “projectors,” providing raw sensory data or generating potential actions. Hierarchical/Heterarchical Organization: The arrangement of NPDs within or connected to KDs visually represents the hierarchical nature of knowledge representation, where KDs integrate and interpret information from multiple NPDs to form meaningful and complex representations. The presence of a higher-order KD further suggests the potential for hierarchical/heterarchical organization of knowledge domains themselves. Dynamic Interplay: While the figure provides a static snapshot, it implies the dynamic interplay between NPDs and KDs. The connections between them suggest the flow of information and influence, where NPDs provide raw data, and KDs interpret and contextualize this information, contributing to the formation of thoughtseeds. Thoughtseed Formation and the Global Workspace: Thoughtseeds emerge from the dynamic interplay between NPDs and KDs, drawing upon the knowledge and information represented within these domains. These thoughtseeds then compete for dominance within the Global Workspace, with the winning thoughtseed shaping the “content of conscious experience.”.
Figure 3.
Neuronal Packet Domains (NPDs) and Knowledge Domains (KDs). This figure provides a static illustration of the relationship between NPDs and KDs at a given time snapshot, highlighting their roles in the thoughtseed framework. KDs: The four distinct colors represent different KDs, each specializing in a particular domain of knowledge or expertise. The unfilled squares symbolize localized KDs, formed as superordinate ensembles of NPDs, integrating information from specific functional units within the brain. The filled squares represent a superordinate ensemble of non-local, heterarchical KDs, suggesting the integration of knowledge across multiple domains. NPDs: The circles depict NPDs, the functional units of the brain responsible for processing specific types of information. They act as “projectors,” providing raw sensory data or generating potential actions. Hierarchical/Heterarchical Organization: The arrangement of NPDs within or connected to KDs visually represents the hierarchical nature of knowledge representation, where KDs integrate and interpret information from multiple NPDs to form meaningful and complex representations. The presence of a higher-order KD further suggests the potential for hierarchical/heterarchical organization of knowledge domains themselves. Dynamic Interplay: While the figure provides a static snapshot, it implies the dynamic interplay between NPDs and KDs. The connections between them suggest the flow of information and influence, where NPDs provide raw data, and KDs interpret and contextualize this information, contributing to the formation of thoughtseeds. Thoughtseed Formation and the Global Workspace: Thoughtseeds emerge from the dynamic interplay between NPDs and KDs, drawing upon the knowledge and information represented within these domains. These thoughtseeds then compete for dominance within the Global Workspace, with the winning thoughtseed shaping the “content of conscious experience.”.
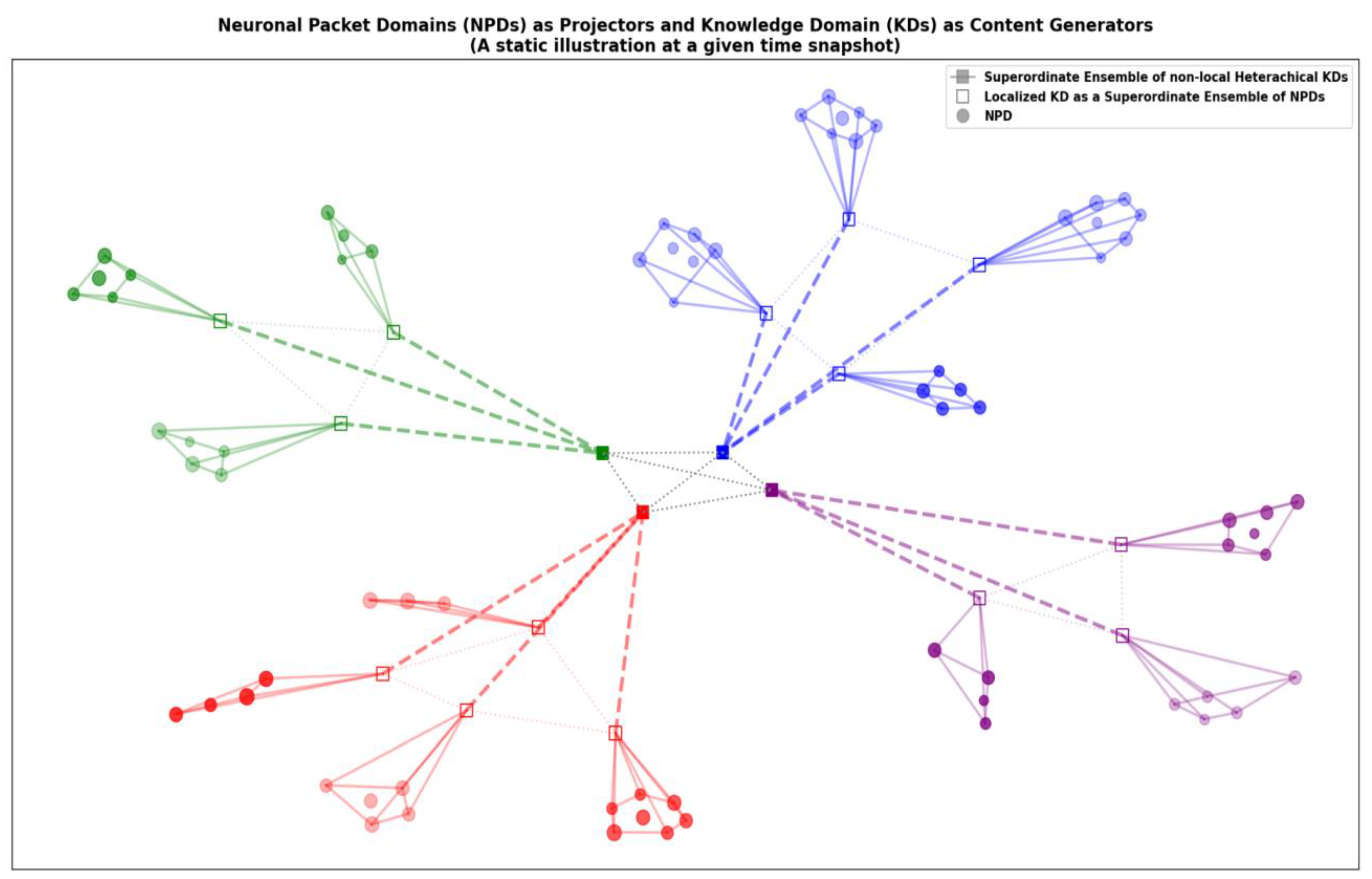
Figure 4.
Illustrative Thoughtseed Framework. This diagram provides a high-level conceptual overview of the internal states within the Thoughtseed Framework, structured across four nested levels. It employs an active inference formalism framed within the principles of Global Workspace Theory.
Figure 4.
Illustrative Thoughtseed Framework. This diagram provides a high-level conceptual overview of the internal states within the Thoughtseed Framework, structured across four nested levels. It employs an active inference formalism framed within the principles of Global Workspace Theory.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



