
Submitted:
21 November 2024
Posted:
25 November 2024
You are already at the latest version
Abstract
Release of herbivore-induced plant volatiles (HIPVs) has been recognized to be an important strategy for plant adaptation to herbivore attack. However, whether these induced volatiles are beneficial to insect herbivores particularly insect larvae is largely unknown. We used two important highly polyphagous lepidopteran pests Spodoptera frugiperda and S. litura to evaluate their benefit for xenobiotic detoxification from larval exposure to HIPVs released by the host plant maize (Zea mays). Larval exposure of the invasive alien species S. frugiperda to maize HIPVs significantly enhanced their tolerance to all three well-known defensive compounds 2,4-dihydroxy-7-methoxy-1,4-benzoxazin-3-one (DIMBOA), chlorogenic and tannic acids in maize, and two commonly used insecticides methomyl and chlorpyrifos. The HIPVs exposure also improved larval tolerance of S. litura third instars to chlorogenic and tannic acids. Furthermore, larval exposure to either maize HIPVs or DIMBOA induced activities of cytochrome P450 enzymes (P450s), glutathione-s-transferase (GST) and carboxylesterase (CarE) in the midguts and fat bodies of the two insects, while the induction was significantly higher by two components together. In addition, expression of four genes encoding uridine diphosphate (UDP)-glycosyltransferases (UGT33F28, UGT40L8) and P450s (CYP4d8, CYP4V2) showed similar induction pattern in S. frugiperda. Cis-3-hexen-1-ol, an important component in maize HIPVs also showed the same functions as maize HIPVs, its exposure increased larval xenobiotic tolerance and induced detoxification enzymes and genes. Our findings demonstrate that HIPVs released by the pest-infested host plants are conductive to xenobiotic tolerance of lepidopteran insect larvae. Hijacking the host plant HIPVs is an important strategy of the invasive alien polyphagous lepidopteran pest to counter-defense host plant chemical defense.
Keywords:
Herbivore-induced plant volatiles
; Spodoptera frugiperda
; xenobiotic
; cytochrome P450
; DIMBOA
1. Introduction
Plants and insects contribute the majority of biodiversity on Earth. During long history of coevolution of the two major groups of organisms, plants take advantage of easy synthesis of organic compounds to produce numerous toxic secondary metabolites to defend against insect herbivores. Upon herbivore attack plants perceive damage-associated and herbivore-associated molecular patterns and immediately activate early signaling components such as Ca2+, reactive oxygen species, and MAP kinases. Subsequently plants initiate their signaling networks including activation of phytohormones and transcription factors, leading to transcriptional reprogramming and a series of metabolic, physiological, and biochemical changes including the production of secondary metabolites [1].
During co-evolution with insect herbivores, plants have developed both constitutive and inducible plant defenses at multiple morphological, molecular, and biochemical layers [2,3,4]. Production of defensive secondary metabolites such as DIMBOA, diterpenoid glycosides, and pyrethrins are a key strategy for plant defense against insect herbivores [5,6,7]. In response, herbivorous insects have evolved intricate strategies to evade toxicity of defensive compounds produced by host plants, including chelation, excretion, metabolic degradation, and target resistance mutations [8]. The main detoxification enzymes involved in insect metabolic resistance are cytochrome P450 monooxygenase (P450), UDP-glucuronide transferase (UGT), glutathione-S-transferase (GST), and carboxylesterase (CarE), which play vital roles in the development of insect metabolic resistance to xenobiotics including various phytochemicals and synthetic insecticides [8,9].
Furthermore, upon insect herbivory plants rapidly synthesize and release a complex blend of volatile chemicals named herbivore-induced plant volatiles (HIPVs) to either directly repel and intoxicate the enemies or indirectly attract the natural enemies of insect herbivores [10,11,12]. HIPVs are important transmitter of plant communication with other organisms in the environment, mainly consisting of green-leaf volatiles, terpenes and aromatic compounds. Timely emission of HIPVs acts as a key strategy of plant adaptation to insect herbivory. Importantly, the HIPVs can serve as important agents for induction and priming of plant defense against insect pests, showing a potential in management of agricultural pests [13,14,15]. However, little is known about counter defense of insect herbivores in response to plant HIPVs.
The fall armyworm Spodoptera frugiperda is a new alien invasive insect pest in Asia from Americas [16]. The insect is a highly polyphagous lepidopteran pest with more than 300 host plants. It has rapidly spread in new regions and become one of the most destructive pests due to its broad host range, high reproductive potential, and swift migration [17]. The recently published genomic data of S. frugiperda showed that the P450 gene family is notably expanded, with 425 members, of which 283 are unique in S. frugiperda when compared to its related species, Spodoptera litura, a native polyphagous lepidopteran pest in Asia [16]. This expansion may confer S. frugiperda capacity to exploit plant volatiles to augment its detoxification.
Notably, UGT33F28 and UGT40L8 have been identified as pivotal genes encoding glycosyltransferases involved in the detoxification of 2,4-dihydroxy-7-methoxy-1,4-benzoxazin-3-one (DIMBOA), the major defensive chemical in maize and other cereals [18]. The upregulation of UGT33F28 and UGT40L8 has been demonstrated to bolster the detoxification capabilities of S. frugiperda.
Herbivorous insects that feed on plants have the ability to stimulate the production of volatile compounds in plants, known as herbivore-induced plant volatiles (HIPVs) [19,20]. HIPVs are typically complex mixtures of compounds derived from various biosynthetic pathways, primarily including terpenes, green leaf volatiles, and aromatic compounds [21]. There is evidence suggesting that HIPVs from different biosynthetic sources can elicit resistance and immunity to parasites and pathogens in lepidoptera [20]. Notable active volatiles identified include cis-3-Hexen-1-ol, indole, β-basil, and β-farnesene [22,23,24]. Furthermore, recent research has demonstrated that HIPVs can induce adaptation to tomato chemical defenses in S. litura [25]. Moreover, tolerance to chemical insecticides can also be induced in Helicoverpa armigera [26]. However, there is limited research on the mechanisms of S. frugiperda's adaptation to resistance in maize hosts. Additionally, there have been no reports on the novel ecological adaptation strategies of S. frugiperda mediated by HIPVs.
In this study, we conducted a systematic evaluation of the impact of maize HIPVs and their key component cis-3-hexen-1-ol on larval detoxification of main defensive compounds in maize plants and insecticides methomyl and chlorpyrifos in S. frugiperda and S. litura. We observed changes in tolerance to methomyl and chlorpyrifos in S. frugiperda and S. litura larvae following exposure to maize HIPVs. Furthermore, exposure to HIPVs prompted S. frugiperda to overcome host resistance mediated by DIMBOA in maize. This response is likely attributed to increased activity of detoxification enzymes and upregulation in the expression of UGT and P450 family genes in the midgut and fat body of S. frugiperda. Additionally, we conducted an exposure experiment using the volatile compound cis-3-Hexen-1-ol, a prominent component of maize HIPVs. Our findings revealed that cis-3-Hexen-1-ol could induce resistance in S. frugiperda to DIMBOA. Notably, when exposed to cis-3-Hexen-1-ol and fed a diet containing DIMBOA 24 h, the weight gain (%) and activity of detoxification enzymes in S. frugiperda were significantly higher compared to those in S. litura, moreover, the gene expression of the UGT family has been upregulated
In conclusion, our findings propose a novel mechanism for the invasion of S. frugiperda. This mechanism involves the olfactory detection of HIPVs, which induces resistance in S. frugiperda to Bxs-mediated chemical defenses in maize, thereby facilitating its invasion. Understanding the ecological adaptation and resistance of S. frugiperda, facilitated by HIPVs, holds considerable importance for the efficient control of invasive pests and the advancement of environmentally sustainable methods for managing invasive insects.
2. Materials and Methods
2.1. Plants and Insects
Seeds of maize cv. Zhengdan 958 obtained from Shandong Luyan Agricultural Seed Co., Ltd. (China) were disinfected using a 2% NaClO solution for 10 minutes and then placed in peat soil (German Dahan type: 413, particle size: 0-6 mm) for germination. After 7 days, uniform and healthy seedlings displaying consistent growth were chosen for transplantation for subsequent experiments. The maize plants were cultivated in a greenhouse (14 hours light/10 hours dark) with 70 % relative humidity and a temperature regimen of 30 ℃ during the day and 25 ℃ at night. The maize plants were adequately irrigated with a low-phosphorus Hoagland nutrient solution. The initial population of S. frugiperda was provided by Professor Lin Jintian at Zhongkai University of Agriculture and Engineering. The insects were reared and propagated for more than 10 generations on artificial diets. The moths were nurtured in a 10% honey water solution, and the rearing chamber was upheld at constant environmental conditions (25 ± 2 ℃, 60% relative humidity, light: dark = 16:8 hours).
2.2. Larval Exposure to Maize HIPVs and Its Effects on Xenobiotic Tolerance
A directed airflow apparatus showed in Figure S1 was used to examine effects of exposure of maize HIPVs on performance of 3rd and 4th instar larvae of S. litura and S. frugiperda on toxin-contained diets. Ten maize seedlings at the five-leaf stage (15 days old) were transplanted in a sealed box (60 cm in length, 48 cm in width, and 55 cm in height). Fourth instar larvae of S. frugiperda were inoculated on the maize seedlings and allowed feeding on plants for 24 hours. Subsequently, four treatment groups were established, including S. litura - HIPVs, S. litura + HIPVs, S. frugiperda - HIPVs, and S. frugiperda +HIPVs. The 3rd and 4th instar larvae of the two species were exposed to maize HIPVs, respectively, and reared on diets either without toxins or with toxic defensive compounds. Weight gain of 3rd and 4th instar larvae of S. litura and S. frugiperda following exposure to maize HIPVs and feeding on an artificial diet for 24 hours was measured. Susceptibility of 3rd and 4th instar larvae to phytotoxins including chlorogenic acid, tannic acid, and DIMBOA, as well as two insecticides chlorpyrifos and methomyl was assessed. Finally, the tolerance of 4th instar S. frugiperda larvae to two insecticides chlorpyrifos and methomyl was evaluated after exposure to maize HIPVs.
Furthermore, effects of HIPVs exposure on egg hatching rate, pupation rate, and emergence rate of S. litura and S. frugiperda were examined. Additionally, the activities of P450, GST, and CarE enzymes, as well as the expression levels of UGT33F28, UGT40L8, CYP4d8, CYP6B6, and CYP4V2 in the midgut and fat body of 4th instar larvae of S. litura and S. frugiperda were quantified in the presence and absence of maize HIPVs, as well as with or without 1 μg·g-1 DIMBOA in the diets.
2.3. Volatile Compounds Toxicity and Exposure to cis-3-hexene-1-ol
To determine the specific volatile compounds responsible for changing larval tolerance, cis-3-hexen-1-ol (C6H12O, CAS:928-96-1) that is present in maize HIPVs was selected [22,23,24]. The compounds were purchased from Shanghai Macklin Biochemical Co., Ltd. The concentrations of cis-3-hexene-1-ol was used based on the report by Abhinav et al. [27]. The dietary supplementation concentrations of DIMBAO and chlorogenic acid (CA) were 1.0 μg·g-1 and 3.0 μg·g-1, respectively. Larvae of the fourth instar of S. litura and S. frugiperda were exposed to cis-3-hexene-1-ol. The mortality of the larvae was assessed after 24 hours of feeding, with 10 larvae per group and 5 groups for each concentration of different substances (n=50).
A head-space volatile release apparatus showed in Figure S2 was developed to evaluate effects of volatile compounds on insect detoxification of xenobiotics. The volatile compound cis-3-hexen-1-ol was added in glass wool within a 2 mL sample vial. Subsequently, an 18# needle was utilized to puncture the rubber spacer on the blue cap of the sample bottle and the lid of the 3.5 L transparent bowl box to apply the volatile compounds to S. litura and S. frugiperda larvae at a specific release rate through a needle connection. Artificial diets containing 1 μg·g-1 DIMBOA and 3 μg·g-1 CA were placed in the bowl, and the insect larvae were reared on these diets for 24 hours. Thereafter, the midgut and fat body were dissected, and the activities of P450, GST, and CarE enzymes, and expression levels of UGT33F28 and UGT40L8 in the midguts and fat bodies were evaluated.
2.4. Determination of Pupation Rate, Emergence Rate and Egg Hatchability
The same number of larvae at pre-pupal stage of S. litura and S. frugiperda were placed in containers constructed from polypropylene (PP) material. The base of the container was covered with fine sand containing 10% water. The larvae were subjected to maize herbivore-induced plant volatiles (HIPVs) (+HIPVs) and control conditions (-HIPVs). Upon completion of pupation or failure to pupate, as well as emergence or mortality, the pupation rate and emergence rate were determined (n=100).
A delicate brush was utilized to evenly disperse the eggs deposited by S. litura and S. frugiperda. Subsequently, the dispersed eggs were placed on a sponge, which was then positioned on a square dish. The eggs were exposed to maize HIPVs (+HIPVs) and control conditions (-HIPVs). Following 96 hours of exposure, the number of hatched insects was tallied (n=100).
2.5. Enzyme Activity of P450, GST and CarE
The enzyme activities of P450, GST and CarE in insect midguts and fat bodies were determined according to the methods of Sun et al. and Tang et al. [25,28]. The tissues of midguts and fat bodies from S. litura and S. frugiperda were used for assaying activities of the detoxification enzymes. The tissues were dissected in PBS, then ground by homogenizing and were centrifuged with 4 ℃ and 10000 g for 20 min. The supernatant was immediately transferred for assaying activities of detoxification enzymes. The activities of P450, GST and CarE were measured by using a microplate analyzer with enzyme activity assay kits (Jiangsu Yutong Biological Technology Co., Ltd, Nanjing, China) according to the manufacturer instructions.
2.6. Gene Expression Analysis
Procedures used for RNA extraction and reverse transcription of plant samples were carried out as previously described [29], with slight modifications. Total RNA was extracted from ~0.1 g flash-frozen, powdered root samples using the Eastep® Super Total RNA Extraction kit (Promega Biotech Co., Ltd., China) according to the manufacturer’s instructions. Total RNA was treated with RNase-Free DNaseI (TIANGEN Biotech Co., Ltd., China), and 1 mg of total RNA was pipetted for cDNA synthesis using the GoScript Reverse Transcription System (Promega Biotech Co., Ltd., China). Real-time PCR was performed using the MonAmp ChemoHS qPCR Mix (High Rox) Kit (Monad Biotech Co., Ltd., China). Reaction conditions for thermal cycling were 95°C for 5 min, followed by 40 cycles of 95°C for 10 s, 55–65°C for 10 s, and 72°C for 30 s. Fluorescence data were collected during the cycle at 72°C. The gene expression level was normalized using the S. frugiperda housekeeping gene GAPDH and the 2-ΔΔCT method. The gene-specific primers used in this research are listed in Table S1. Biological triplicates with technical duplicates were performed.
2.7. Statistical Analysis
Data were processed and plotted using Microsoft Excel 2013 and GraphPad Prism 9 software, and significance was tested using SPSS 19. All experiments were conducted using a completely randomized experimental design. Data were checked for normality (p > 0.05) using the Shapiro-Wilk normality test and Levene’s test for homogeneity of variance (p > 0.05) prior to all statistical analyses. On the premise of satisfying the assumption of normality and homogeneity of variance, One-way ANOVA or Two-way ANOVA (Tukey’s post hoc test, p < 0.05) were used to compare significant differences between two or more treatments.
3. Results
3.1. Maize HIPVs Promote Larval Tolerance to Plant Defensive Chemicals
DIMBOA, CA and TA
To examine the potential impact of maize HIPVs on larval tolerance to plant defensive chemicals, 3rd and 4th instar larvae of S. litura and S. frugiperda were exposed to on toxin-contained diets maize HIPVs and reared on artificial diet either without toxins or with DIMBOA, chlorogenic acid (CA) and tannic acid (TA). Without toxins on diets maize HIPVs exposure did not affect larval growth of S. frugiperda, but significantly reduced larval growth of S. litura (Figure 1A,B), suggesting that S. frugiperda larvae are more adaptive to maize HIPVs. When the larvae were exposed to toxin contained diets, maize HIPVs exposure significantly improved larval growth and toxin tolerance of both 3rd and 4th instar larvae of S. frugiperda to all three tested plant defensive chemicals. Specifically, HIPVs exposure increased weight gain of 3rd and 4th instars of S. frugiperda in presence of 1 μg·g-1 DIMBOA by 62.8% and 60.6%, respectively (Figure 1C,F). The HIPVs exposure increased weight gain by 23.8% and 209.5%, respectively in presence of 3 μg·g-1 CA (Figure 3D,G), and 64.77% and 67.75% respectively in presence of 2 μg·g-1 TA (Figure 3E,H).
For S. litura larvae, maize HIPVs exposure did not improve larval growth of both 3rd and 4th instars in presence of 1 μg·g-1 DIMBOA (Figure 1C,F). It did not improve larval growth of 4th instar either in presence of CA and TA (Figure 1G,H). However, HIPVs exposure did improve larval growth of 3rd instar in presence of CA and TA (Figure 3D,E,G,H). The results also suggest that S. frugiperda is more adaptive to maize HIPVs for tolerance to plant defensive chemicals.
3.2. Maize HIPVs Promote Larval Tolerance to Insecticides Methomyl and Chlorpyrifos
Methomyl and chlorpyrifos are commonly utilized globally as broad-spectrum insecticides [30,31]. This study aimed to examine the potential of exposure to maize HIPVs to confer larval tolerance to methomyl and chlorpyrifos in S. frugiperda. Susceptibility of 4th instars of S. litura and S. frugiperda fed on diets contained 500 μg·mL-1 of methomyl and 8000 μg·mL-1 of chlorpyrifos was examined after larval exposure to maize HIPVs for 24 hours (Figure 2A,B). The mortality of 4th instar S. frugiperda exposed to maize HIPVs was significantly lower than that of unexposed individuals. However, no significant difference was observed between HIPVs-exposed and un-exposed 4th instars in S. litura.
To determine the role of P450 in HIPVs-enhanced larval tolerance to insecticides the 4th instar larvae of S. frugiperda were exposed to HIPVs for 24 hours and topically treated with piperonyl butoxide (PBO, a general inhibitor of P450 enzymes) on the thorax and abdomen 1 hour prior to transfer to a diet containing 500 μg·mL-1 methomyl and 8000 μg·mL-1 chlorpyrifos, respectively. The mortality after 24 hours are counted (Figure 2C,D). In absence of the inhibitor PBO maize HIPVs exposure reduced larval mortality of methomyl-treated S. frugiperda by 43.3%, and chlorpyrifos-treated S. frugiperda by 30.5%. In presence of PBO larval mortality of insecticide-treated S. frugiperda significantly increased. However, in presence of PBO maize HIPVs exposure did not change larval mortality for both methomyl-treated and chlorpyrifos-treated S. frugiperda. The results indicate that P450s play a key role in HIPVs-enhanced larval tolerance to insecticides.
3.3. Maize HIPVs Exposure Does Not Affect Insect Development
3.4. Maize HIPVs and DIMBOA Show Synergistic Effect on Induction of Detoxification Enzymes
The activity of detoxification enzymes Cytochrome P450 monooxygenases (P450), glutathione-S-transferases (GST), and carboxylesterases (CarE) are three main detoxification enzymes implicated in metabolism of xenobiotics[8]. We examined the activities of the three detoxification enzymes in the midguts and fat bodies of 4th instar larvae of S. litura and S. frugiperda following exposure to maize HIPVs and plant defensive chemical DIMBOA (Figure 4). Either HIPVs exposure or treatment with DIMBOA significantly enhanced activities of P450 and CarE in both midguts and fat bodies (Figure 4A–D,I–L). More importantly, simultaneous treatments with HIPVs and DIMBOA showed the strongest induction of all three tested detoxification enzymes (Figure 4A–L). Although HIPVs exposure and treatment with DIMBOA showed lower or no obvious induction of GST, simultaneous treatments with HIPVs and DIMBOA also induced activity of GST (Figure 4E–H).
3.5. Maize HIPVs and DIMBOA Show Synergistic Effect on Induction of Detoxification Associated Genes
Uridine diphosphate (UDP)-glycosyltransferases (UGTs) are important phase II detoxification enzymes in insects, which play a key role in the metabolism of xenobiotics [32]. In S. frugiperda, SfUGT33F28 and SfUGT40L8 have been demonstrated to re-glycosylate toxic benzoxazinoids into non-toxic stable glucosides to detoxify benzoxazinoids [33], the most important defensive compounds in maize and other important cereal crops. To investigate the potential impact of HIPVs exposure on expression of genes associated with detoxification, RT-pPCR was used to quantify the expression of detoxification associated genes in the midguts and fat bodies of 4th instar larvae of S. frugiperda larvae following exposure to maize HIPVs and plant defensive chemical DIMBOA for 24 hours (Figure 5).
Maize HIPVs exposure led to 5.0-, 23.8-, 19.2-, and 2.2-fold upregulation in the gene expressions of UGT33F28, UGT40L8, CYP4d8 and CYP4V2 in the midguts, respectively (Figure 5A-E). Transcript levels of UGT33F28, UGT40L8 and CYP4d8 in the midguts increased by 2.5-, 32.2-, and 4.0-fold, respectively, 24 h after feeding on DIMBOA-contained diet. More strikingly, simultaneous treatments with HIPVs and DIMBOA showed the strongest induction of all five tested detoxification genes in the midguts (Figure 5A-E), as well as fours genes in the fat bodies (Figure 5G-J). The induction of UGT33F28, UGT40L8, CYP4d8, CYP6B6 and CYP4V2 by the two components was 13.1-, 43.2-, 32.8-, 2.1- and 3.1-fold in the midguts relative to untreated control, respectively (Figure 5A-E). In the fat bodies CYP4d8 and CYP6B6 expression levels were induced 22.1- and 13.9-fold by simultaneous treatments with HIPVs and DIMBOA, respectively relative to untreated control (Figure 5H-I).
3.6. Larval Exposure to cis-3-hexen-1-ol Enhances Tolerance to Plant Defensive Chemicals
The amount of cis-3-hexen-1-ol (cis-HXO) is increased in S. frugiperda -damaged maize plants [34]. To identify the specific volatile compounds emitted from herbivore-infested maize plants that triggered xenobiotic resistance in S. frugiperda, the 4th instar larvae of S. litura and S. frugiperda were exposed to volatile cis-HXO and reared on diets contained DIMBOA and CA in the device shown in Figure S2. We found that exposure of S. frugiperda larvae to cis-3-hexen-1-ol significantly enhanced larval tolerance to both DIMBOA and CA (Figure 6A-B).
3.7. Cis-3-Hexen-1-ol and DIMBOA Show Synergistic Effect on Induction of Detoxification Enzymes
The activities of the three detoxification enzymes P450, GST and CarE in the midguts and fat bodies of 4th instar larvae of S. litura and S. frugiperda were examined following exposure to cis-3-hexen-1-ol and plant defensive chemical DIMBOA (Figure 7). In S. frugiperda either HIPVs exposure or treatment with DIMBOA significantly enhanced activities of P450 and CarE in both midguts and fat bodies (Figure 7B,D,J,L). HIPVs exposure and treatment with DIMBOA only did not showed induction of GST in the midguts of S. frugiperda (Figure 7F), but induced GST in the fat bodies (Figure 7H).
In S. litura exposure to cis-3-hexen-1-ol induced the activities of the three detoxification enzymes P450, GST and CarE in the fat bodies (Figure 7C,G,K), only induced CarE in the midguts (Figure 7I), but not P450 and GST in the midguts (Figure 7A,E). Diet supplement with DIMBOA significantly enhanced activities of P450, GST and CarE in both midguts and fat bodies (Figure 7A,C,E,G,I,K).
Similar to the results from exposure to maize HIPVs and supplement with DIMBOA, simultaneous treatments with cis-3-hexen-1-ol and DIMBOA showed strongest induction of P450, GST and CarE in both midguts fat bodies in the two insect species.
3.8. Cis-3-Hexen-1-ol and DIMBOA Upregulate UGT33F28 and UGT40L8
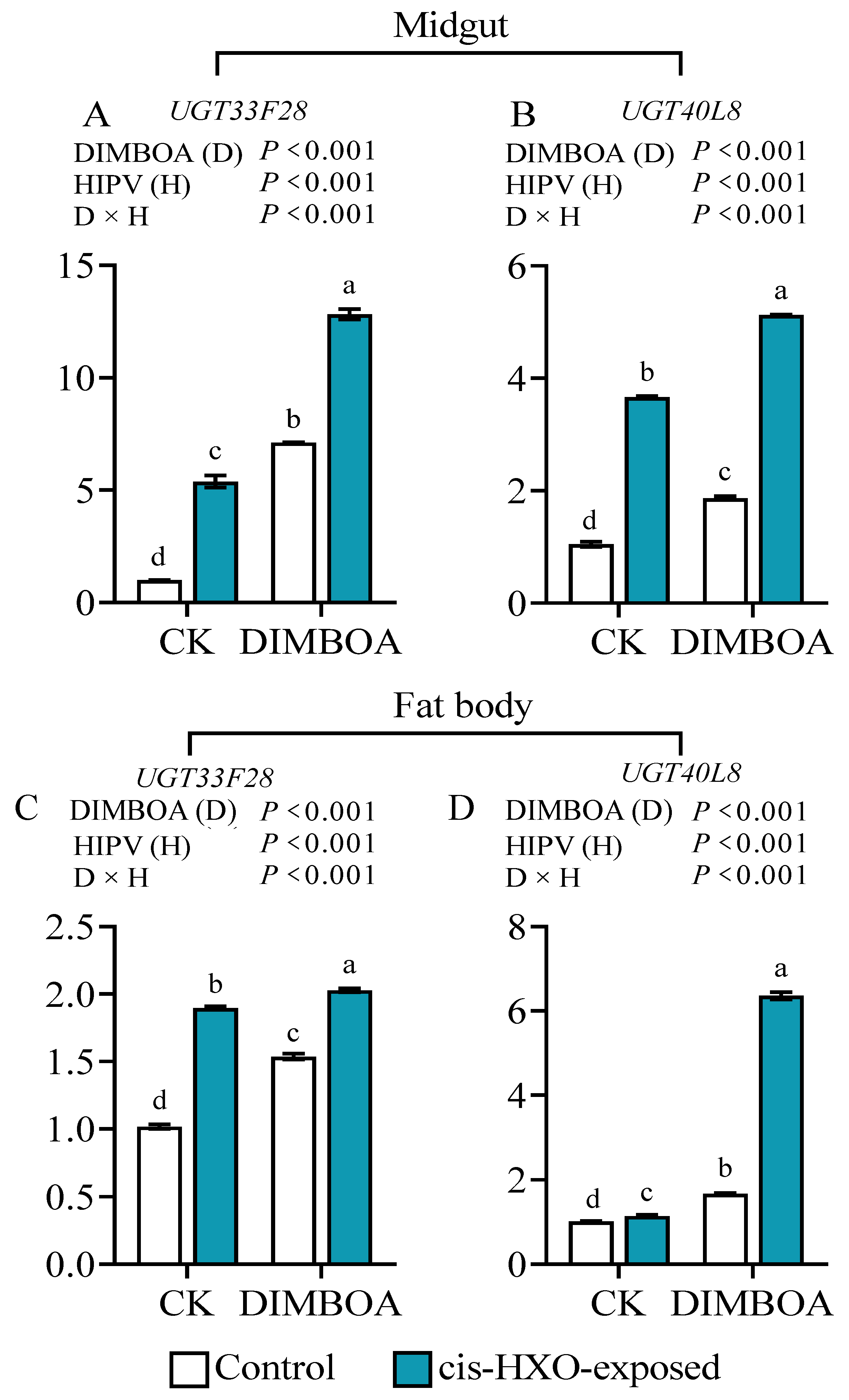
We further investigated the impact of exposure to cis-3-hexen-1-ol and DIMBOA treatments on expression of two genes encoding phase II detoxification enzymes UGT33F28 and UGT40L8 in in the midguts and fat bodies of 4th instar S. frugiperda larvae. We found that after exposure to cis-3-hexen-1-ol the gene expression levels of UGT3F28 and UGT40L8 were increased by 5.4- and 3.5-fold in the midgut (Figure 8A, 8B), and 1.9- and 1.1-fold in the fat bodies (Figure 8C, 8D), respectively, in comparison to unexposed control. Meanwhile, diet supplement with DIMBOA increased gene expressions of UGT3F28 and UGT40L8 by 6.7- and 1.8-fold in the midguts (Figure 8A, 8B), and 1.5- and 1.7- fold in the fat bodies (Figure 8C, 8D), respectively relative to untreated control. Simultaneous treatments with cis-3-hexen-1-ol and DIMBOA showed strongest induction of the two detoxification genes (Figure 8A-D).
4. Discussion
Currently, over 1,700 volatile compounds have been identified from approximate 90 plant families globally. The majority of these compounds fall into categories such as hydrocarbons, alcohols, aldehydes, ketones, esters and terpenes, with a molecular weight ranging from 100 to 200 [35]. It has been demonstrated that HIPVs display diverse ecological functions [36]. The HIPVs-mediated interactions among plants, phytophagous insects, and natural enemies has garned significant attention in recent years [37,38].
HIPVs have been found to have various effects on phytophagous insects, including growth inhibition, feeding deterrent and repelling[12,39]. Additionally, HIPVs can serve as an attractor for predatory or parasitic natural enemies, thereby indirectly protecting host plants [40,41]. Despite an array of studies on benefits of HIPVs to the plants, very limited studies have torched potential benefits of HIPVs on receiver insect herbivores [29]. One obvious benefit is that insects can use HIPVs to orientate host plants. For instance, females of Tuta absoluta utilize HIPVs emitted by tobacco plants to identify the host for oviposition [42]. A recent study revealed that symbiotic bacteria present in Acyrthosiphon pisum can play a role in suppressing the emission of HIPVs in host plants to reduces the risk of Acyrthosiphonpisum being parasitized by predators, then enhancing the adaptability of herbivorous insects [43].
This study reveals that larval exposure to maize HIPVs increased the weight gain of both 3rd and 4th instars of S. frugiperda in presence of 1 μg·g-1 DIMOBA, 1μg·g-1 CA, and 1 μg·g-1 TA compared to those without exposure to HIPVs (Figure 1C-H). The exposure only increased the weight gain of 3rd instars of S. litura in presence of CA and TA, it did not show obvious effects on fourth instars of S. litura and presence of DIMOBA (Figure 1). Furthermore, Maize HIPVs also increased larval tolerance of S. frugiperda to the two insecticides, but did not show effect on tolerance of S. litura (Figure 2). These results indicate that maize HIPVs exposure showed significantly more benefits to S. frugiperda than to S. litura. Hijacking the host plant HIPVs seems an important strategy of the invasive alien species fall armyworm to counter-defense host plant chemical defense. More importantly, maize HIPVs and main defensive chemical DIMBOA showed synergistic effect on induction of detoxification enzyme systems and detoxification genes (Figure 4, Figure 5), suggesting that the two highly polyphagous lepidopteran pests utilize both volatile HIPVs and non-volatile defensive chemicals from host plants to develop counter-defense against host plant chemical defense.
Phytophagous insects have developed various mechanisms to counteract plant defenses, such as detoxification enzyme systems, physiological tolerance, and behavioral escape [44,45]. The detoxification enzymes in insects, including cytochrome oxidase (P450), carboxylesterase (CarE) and glutathione S-transferase (GSTs), play a crucial role in metabolizing plant defense substances and are essential for insect adaptation to host plant defense [9,46,47,48]. This study revealed that exposure to maize HIPVs can lead to a significant increase in the activity of detoxification enzymes in the midgut and fat body of both S. frugiperda and S. litura (Figure 4A-L). However, the induction of detoxification enzymes was significantly higher in S. frugiperda than that in S. litura (Figure 4A-L). Furthermore, exposure to HIPVs was found to induce larval tolerance to methomyl and chlorpyrifos in S. frugiperda larvae (Figure 2A, 2B), but not in S. litura. Consequently, it is plausible to infer that S. frugiperda possesses the capability to promptly detect changes in maize HIPVs compared to S. litura, and subsequently respond physiologically to HIPVs by activating its own detoxification system to overcome the host resistance mediated by DIMBOA in maize. Our results indicate that
Green leaf volatiles (GLV) are a group of small gaseous molecules emitted by plants in response to various stressors such as mechanical damage, pathogen infection, and insect infestation. Cis-3-hexen-1-ol, identified as a main GLV in maize, plays a pivotal role in enhancing plant stress resistance [49]. This study found that exposure to cis-3-hexene-1-ol led to increased detoxification enzyme activities and UGT3F28 and UGT40L8 expression in the midgut and fat body of S. frugiperda. Our findings suggest that cis-3-hexen-1-ol plays a significant role in triggering xenobiotic tolerance of S. frugiperda to DIMBOA-mediated chemical defense in maize. This suggests that the green leaf volatile is beneficial to both plants and insect herbivores. Application of cis-3-hexen-1-ol in enhancing plant stress resistance may confer herbivore xenobiotic tolerance.
5. Conclusions
Upon larval exposure to maize HIPVs two highly polyphagous lepidopteran pests S. frugiperda and S. litura all significantly enhance their tolerance to plant defensive chemicals including two general defensive chemicals CA and TA and one specific defensive chemical DIMBOA. S. frugiperda shows more adaptive to maize HIPVs and gains more benefits. Larval HIPVs exposure also enhances their tolerance to two insecticides in S. frugiperda, but not in S. litura. Larval exposure to maize HIPVs also enhance their activities of P450s, GST and CarE in the midgut and fat body of the two insects, and the induction is significantly higher S. frugiperda than in S. litura, which may contribute to more tolerance to xenobiotics in S. frugiperda. The green leaf volatile cis-3-hexen-1-ol acts as an active component in maize HIPVs to enhance larval tolerance to xenobiotics and to induce insect detoxification enzymes.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
P.W., Q.Z., Y.Z., X.S. and Y.H. performed the experiments and analyzed data. P.W. and Y.L. drafted the manuscript. R.Z., Y.S. and D.C. conceived the study, obtained funding, and revised the final version of the manuscript. All authors read and approved the final article. All authors have read and agreed to the published version of the manuscript.
Funding
This work is the National Natural Science Foundation of China (U2005208, 32271617, 32371588, 32471656, 32260675), the Natural Science Foundation of Fujian Province, China (2021J02024).
Data Availability Statement
All the data analyzed during this study have been included in this article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Erb, M.; Reymond, P. Molecular Interactions Between Plants and Insect Herbivores. Annu. Rev. Plant Biol. 2019, 70, 527–557. [Google Scholar] [CrossRef] [PubMed]
- Ohgushi, T. Eco-evolutionary dynamics of plant-herbivore communities: Incorporating plant phenotypic plasticity. Curr. Opin. Insect Sci. 2016, 14, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Schuman, M.C.; Baldwin, I.T. The layers of plant responses to insect herbivores. Annu. Rev. Entomol. 2016, 61, 373–394. [Google Scholar] [CrossRef] [PubMed]
- Kessler, A. The information landscape of plant constitutive and induced secondary metabolite production. Curr. Opin. Insect Sci. 2015, 8, 47–53. [Google Scholar] [CrossRef]
- Zhou, S.Q.; Annett, R.; Georg, J. Beyond defense: Multiple functions of benzoxazinoids in maize metabolism. Plant Cell Physiol. 2018, 59, 1528–1537. [Google Scholar] [CrossRef]
- Li, J.C.; Halitschke, R.; Li, D.P.; Paetz, C.; Su, H.C.; Heiling, S.; Xu, S.Q.; Baldwin, I.T. Controlled hydroxylations of diterpenoids allow for plant chemical defense without autotoxicity. Science. 2021, 371, 255–260. [Google Scholar] [CrossRef]
- Lybrand, D.B.; Xu, H.Y.; Last, R.L.; Pichersky, E. How plants synthesize pyrethrins: Safe and biodegradable insecticides. Trends in Plant Sci. 2020, 25, 1245–1251. [Google Scholar] [CrossRef]
- Hanna, M.H.F.; Heiko, V. Molecular mechanisms of insect adaptation to plant secondary compounds. Curr. Opin. Insect Sci. 2015, 8, 8–14. [Google Scholar]
- Lu, K.; Cheng, Y.B.; Li, Y.M.; Li, W.R.; Zeng, R.S.; Song, Y.Y. Phytochemical flavone confers broad-spectrum tolerance to insecticides in Spodoptera litura by activating ROS/CncC-mediated xenobiotic detoxification pathways. J. Agr. Food Chem. 2021, 69, 7429–7445. [Google Scholar] [CrossRef]
- Axel, M.; Wilhelm, B. Plant defense against herbivores: chemical aspects. Annu. Rev. Plant Biol. 2012, 63, 431–450. [Google Scholar]
- Turlings, T.C.J.; Erb, M. Tritrophic interactions mediated by herbivore-induced plant volatiles: mechanisms, ecological relevance, and application potential. Annu. Rev. Entomol. 2018, 63, 433–452. [Google Scholar] [CrossRef] [PubMed]
- Veyrat, N.; Robert, C.A.M.; Turlings, T.C.J.; Erb, M. Herbivore intoxication as a potential primary function of an inducible volatile plant signal. J. Ecol. 2016, 104, 591–600. [Google Scholar] [CrossRef]
- Pérez-Hedo, M.; Alonso-Valiente, M.; Vacas, S.; Gallego, C.; Pons, C.; Arbona, V.; Rambla, J.L.; Navarro-Llopis, V.; Granell, A.; Urbaneja, A. Plant exposure to herbivore-induced plant volatiles: a sustainable approach through eliciting plant defenses. J. Pest Sci. 2021, 94, 1221–1235. [Google Scholar] [CrossRef]
- Xiao, Y.Y.; Qian, J.J.; Hou, X.L.; Zeng, L.T.; Liu, X.; Mei, G.G.; Liao,Y. Y. Diurnal emission of herbivore-induced (Z)-3-hexenyl acetate and allo-ocimene activates sweet potato defense responses to sweet potato weevils. J. Integr. Agr. 2023, 22, 1782–1796. [Google Scholar] [CrossRef]
- Guan, Y.Q.; Yang, X.M.; Chen, S.M.; Chen, F.D.; Chen, X.L.; Chen, F.; Jiang, Y.F. Infestation with chewing (Spodoptera frugiperda) and piercing-sucking (Tetranychus urticae) arthropod lead to differential emission and biosynthesis of HIPVs underscoring MeSA and terpenoids in chrysanthemum foliage. Sci. Hortic-Amsterdam. 2024, 326, 112767. [Google Scholar] [CrossRef]
- Gui, F.R.; Lan, T.M.; Zhao, Y.; Guo, W.; Dong, Y.; Fang, D.M.; Liu, H.; Li, H.M.; Wang, H.L.; Hao, R.S. Genomic and transcriptomic analysis unveils population evolution and development of pesticide resistance in fall armyworm Spodoptera frugiperda. Protein Cell. 2022, 13, 513–531. [Google Scholar] [CrossRef]
- Xiao, H.M.; Ye, X.; Xu, H.; Mei, Y.; Li, F. The genetic adaptations of fall armyworm Spodoptera frugiperda facilitated its rapid global dispersal and invasion. Mol. Ecol. Resour. 2020, 20, 1050–1068. [Google Scholar] [CrossRef]
- Israni, B.; Luck, K.; Römhild, S.C.W.; Raguschke, B.; Wielsch, N.; Hupfer, Y.; Reichelt, M.; Svatos, A.; Gershenzon, J.; Vassao, D.G. Alternative transcript splicing regulates UDP-glucosyltransferase-catalyzed detoxification of DIMBOA in the fall armyworm (Spodoptera frugiperda). Sci. Rep. 2022, 12, 10343. [Google Scholar]
- Erb, M. Volatiles as inducers and suppressors of plant defense and immunity — origins, specificity, perception and signaling. Curr. Opin. Plant Biol. 2018, 44, 117–121. [Google Scholar] [CrossRef]
- Hu, L.; Zhang, K.; Wu, Z.; Xu, J.M.; Erb, M. Plant volatiles as regulators of plant defense and herbivore immunity: molecular mechanisms and unanswered questions. Curr. Opin. Insect Sci. 2021, 44, 82–88. [Google Scholar] [CrossRef]
- Bouwmeester, H.; Schuurink, R.C.; Bleeker, P.M.; Schiestl, F. The role of volatiles in plant communication. Plant, J. 2019, 100, 892–907. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Veyrat, N.; Xu, H.; Hu, L.F.; Turlings, T.C.J.; Erb, M. An herbivore-induced plant volatile reduces parasitoid attraction by changing the smell of caterpillars. Sci. Adv. 2018, 4, eaar4767. [Google Scholar] [CrossRef] [PubMed]
- Gasmi, L.; Martinez-Solis, M.; Frattini, A.; Ye, M.; Carmen Collado, M.; Turlings, T.C.J.; Erb, M.; Herrero, S. Can herbivore-induced volatiles protect plants by increasing the herbivores' susceptibility to natural pathogens? Appl. Environ. Microb. 2019, 85, e01468–01418. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, E.; Venkatesan, R. Plant volatiles modulate immune responses of Spodoptera litura. J. Chem. Ecol. 2019, 45, 715–724. [Google Scholar] [CrossRef]
- Sun, Z.X.; Wang, R.M.; Du, Y.F.; Gao, B.Y.; Gui, F.R.; Lu, K. Olfactory perception of herbicide butachlor by GOBP2 elicits ecdysone biosynthesis and detoxification enzyme responsible for chlorpyrifos tolerance in Spodoptera litura. Env. Pollution. 2021, 285, 117409.1–117409.10. [Google Scholar] [CrossRef]
- Wu, C.F.; Ding, C.H.; Chen, S.; Wu, X.Y.; Zhang, L.Q.; Song, Y.Y.; Li, W.; Zeng, R.S. Exposure of Helicoverpa armigera larvae to plant volatile organic compounds induces cytochrome P450 monooxygenases and enhances larval tolerance to the insecticide methomyl. Insects. 2021, 12, 238–251. [Google Scholar] [CrossRef]
- Abhinav, K.M.; Rakhi, C.P.; Christopher, J. Frost Acute toxicity of the plant volatile indole depends on herbivore specialization. J. Pest Sci. 2020, 93, 1107–1117. [Google Scholar]
- Tang, B.; Cheng, Y.; Li, Y.; Li, W.; Zhou, Q.; Lu, K. Adipokinetic hormone enhances CarE-mediated chlorpyrifos resistance in the brown planthopper, Nilaparvata lugens. Insect Mol. Biol. 2020, 29, 511–522. [Google Scholar] [CrossRef]
- Sun, Z.X.; Lin, Y.B.; Wang, R.M.; Li, Q.L.; Baerson, S.R.; Chen, L.; Zeng, R.S.; Song, Y.Y. Olfactory perception of herbivore-induced plant volatiles elicits counter-defenses in larvae of the tobacco cutworm. Funct. Ecol. 2021, 35, 384–397. [Google Scholar] [CrossRef]
- Van Scoy, A.R.; Yue, M.; Deng, X.; Tjeerdema, R. Environmental fate and toxicology of methomyl. Rev. Environ. Contam. T. 2013, 222, 93–109. [Google Scholar]
- Solomon, K.R.; Williams, W.M.; Mackay, D.; Purdy, J.; Giddings, J.M.; Giesy, J.P. Properties and uses of chlorpyrifos in the United States. Rev. Environ. Contam. T. 2014, 231, 13–34. [Google Scholar]
- Li, X.; Shi, H.; Gao, X.; Liang, P. Characterization of UDP-glucuronosyltransferase genes and their possible roles in multi-insecticide resistance in Plutella xylostella (L.). Pest Manag. Sci. 2018, 74, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Israni, B.; Wouters, F.C.; Luck, K.; Seibel, E.; Ahn, S.J.; Paetz, C.; Reinert, M.; Vogel, H.; Erb, M.; Heckel, D.G.; et al. The Fall Armyworm Spodoptera frugiperda Utilizes Specific UDP-Glycosyltransferases to Inactivate Maize Defensive Benzoxazinoids. Front. Physiol. 2020, 11, 604754. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Fan, H.; Hu, R.; Huang, Y.; Sheng, C.W.; Cao, H.Q.; Wang, G.R.; Jiang, X.C. Characterization of core maize volatiles induced by Spodoptera frugiperda that alter the mating-mediated approach-avoidance behaviors of Mythimna separate. J. Integr. Agr. 2024. [Google Scholar] [CrossRef]
- Laothawornkitkul, J.; Taylor, J.E.; Paul, N.D.; Nicholashewitt, C. Biogenic volatile organic compounds in the Earth system. New Phytol. 2009, 183, 27–51. [Google Scholar] [CrossRef]
- Takabayashi, J.; Shiojiri, K. Multifunctionality of herbivoryinduced plant volatiles in chemical communication in tritrophic interactions. Curr. Opin. Insect Sci. 2019, 32, 110–117. [Google Scholar] [CrossRef]
- Mauch-Mani, B.; Baccelli, I.; Luna, E.; Flors, V. Defense Priming: An adaptive part of induced resistance. Annu. Rev. Plant Biol. 2017, 68, 485–512. [Google Scholar] [CrossRef]
- Aljbory, Z.; Chen, M.S. Indirect plant defense against insect herbivores: a review. Insect Sci. 2018, 25, 2–23. [Google Scholar] [CrossRef]
- Irmisch, S.; McCormick, A.; Günther, J.; Schmidt, A.; Boeckler, G.A.; Gershenzon, J.; Unsicker, S.B.; Koellner, T.G. Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalist caterpillar. Plant J. 2014, 80, 1095–1107. [Google Scholar] [CrossRef]
- Danner, H.; Desurmont, G.A.; Cristescu, S.M.; van Dam, N.M. Herbivore-induced plant volatiles accurately predict history of coexistence, diet breadth, and feeding mode of herbivores. New Phytol. 2018, 220, 726–738. [Google Scholar] [CrossRef]
- Xu, H.; Desurmont, G.; Degen, T.; Zhou, G.; Laplanche, D.; Henryk, L. Combined use of herbivore-induced plant volatiles and sex pheromones for mate location in braconid parasitoids. Plant Cell Environ. 2017, 40, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Anastasaki, E.; Drizou, F.; Milonas, P.G. Electrophysiological and oviposition responses of Ttuta absoluta females to herbivore-induced volatiles in tomato plants. J. Chem. Ecol. 2018, 44, 288–2981. [Google Scholar] [CrossRef] [PubMed]
- Frago, E.; Mala, M.; Weldegergis, B.T.; Yang, C.; Mclean, A.; Godfray, H.; Charles, J.; Gols, R.; Dicke, M. Symbionts protect aphids from parasitic wasps by attenuating herbivore-induced plant volatiles. Nat. Commun. 2017, 8, 1860. [Google Scholar] [CrossRef]
- Zhu, S.K.; Zeng, R. Insect response to plant defensive protease inhibitors. Annu. Rev. Entomol. 2015, 60, 233–252. [Google Scholar]
- Heidel-Fischer, H.M.; Vogel, H. Molecular mechanisms of insect adaptation to plant secondary compounds. Curr. Opin. Insect Sci. 2015, 8, 8–14. [Google Scholar] [CrossRef]
- Bass, C.; Zimmer, C.T.; Riveron, J.M.; Wilding, C.S.; Wondji, C.S.; Kaussmann, M.; Field, L.M.; Williamson, M.S.; Nauen, R. Gene amplification and microsatellite polymorphism underlie a recent insect host shift. PNAS. 2013, 110, 19460–19465. [Google Scholar] [CrossRef]
- Giraudo, M.; Hilliou, F.; Fricaux, T.; Audant, P.; Feyereisen, R.; Le, G.G. Cytochrome P450s from the fall armyworm (Spodoptera frugiperda): responses to plant allelochemicals and pesticides. Insect Mol. Biol. 2015, 24, 115–128. [Google Scholar] [CrossRef]
- Feyereisen, R. Insect CYP genes and P450 enzymes. Insect Biochem. Molec. 2012, 236–316. [Google Scholar]
- Jurgen, E.; Fabiola, C.C.; Chinmay, D.; T. ; Li, T.; Marie, E.; Martin, H. Early transcriptome analyses of Z-3-hexenol-treated Zea mays revealed distinct transcriptional networks and anti-herbivore defense potential of green leaf volatiles. PLoS One. 2013, 8, e77465. [Google Scholar]
Figure 1.
Weight gain of 3rd and 4th larvae of S. frugiperda and S. litura fed on artificial diets containing DIMBOA, chlorogenic acid and tannic acid with or without exposure to maize HIPVs. The larvae were exposed to HIPVs released from living maize plants as showed in Figure S1. Weight gain of 3rd and 4th larvae fed on artificial diets without toxin addition (A & B), containing 1 μg·g-1 DIMBOA diet (C & F), 3 μg·g-1 chlorogenic acid (D & G) and 2 μg·g-1 tannic acid (E & H) for 48 h. Data are mean ± S.D. (n=25). Asterisks indicate significant differences in comparison with control (Student’s t-test, * P<0.05, ** P<0.01).
Figure 1.
Weight gain of 3rd and 4th larvae of S. frugiperda and S. litura fed on artificial diets containing DIMBOA, chlorogenic acid and tannic acid with or without exposure to maize HIPVs. The larvae were exposed to HIPVs released from living maize plants as showed in Figure S1. Weight gain of 3rd and 4th larvae fed on artificial diets without toxin addition (A & B), containing 1 μg·g-1 DIMBOA diet (C & F), 3 μg·g-1 chlorogenic acid (D & G) and 2 μg·g-1 tannic acid (E & H) for 48 h. Data are mean ± S.D. (n=25). Asterisks indicate significant differences in comparison with control (Student’s t-test, * P<0.05, ** P<0.01).
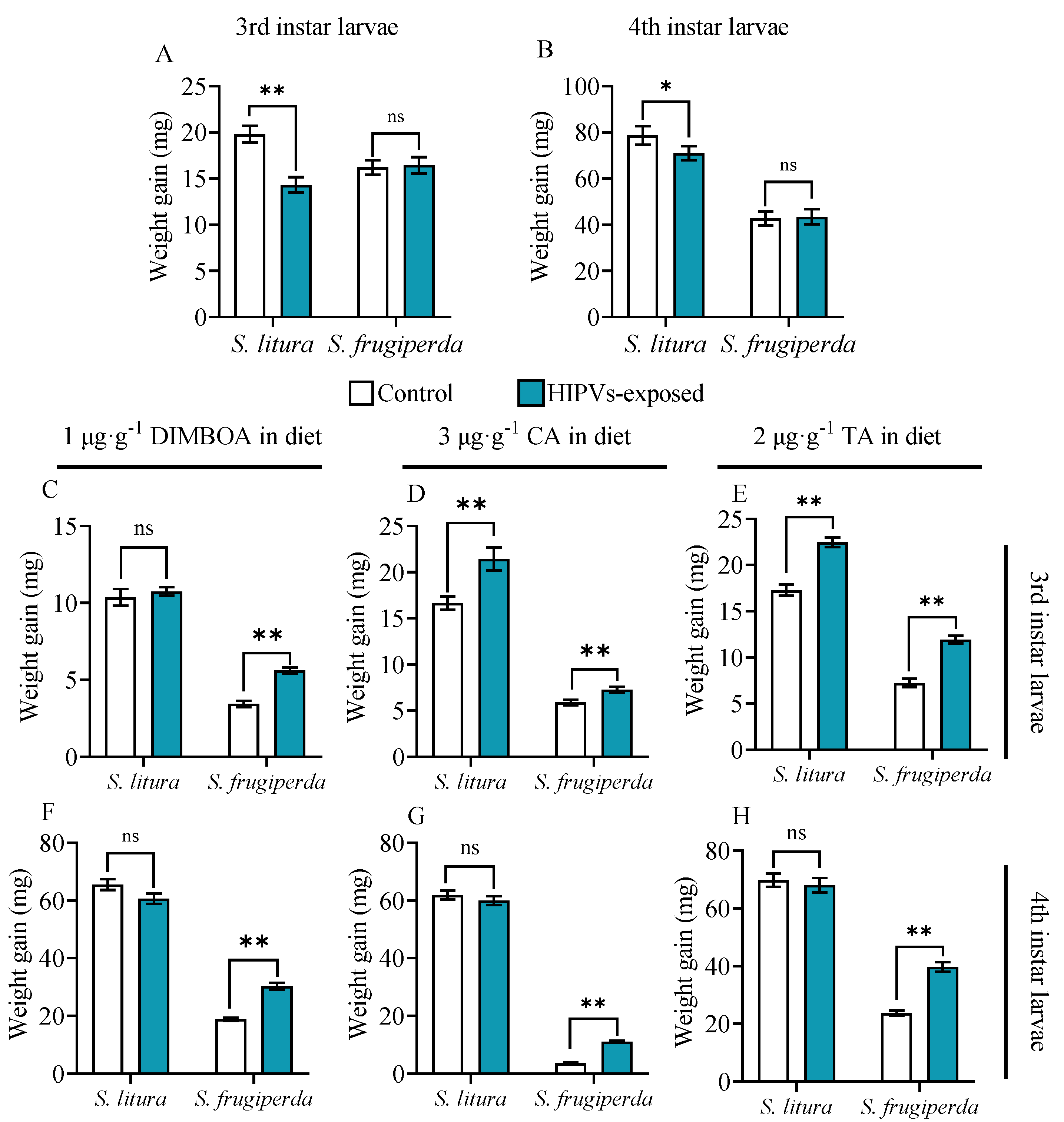
Figure 2.
Insecticide tolerance of S. frugiperda and S. litura larvae to methomyl and chlorpyrifos after exposure to maize HIPVs. The larvae were exposed to HIPVs released from living maize plants as showed in Figure S1. Larval mortality was measured 24 hours after exposure to maize HIPVs, and methomyl (A) and chlorpyrifos (B). Asterisks indicate significant differences in comparison with unexposed control (Student’s t-test, ** P<0.01); Tolerance to methomyl (C) and chlorpyrifos (D) after exposure of S. frugiperda larvae to maize HIPVs and the insecticide synergist piperonyl butoxide (PBO). Data are mean ± S.D. (n=20).
Figure 2.
Insecticide tolerance of S. frugiperda and S. litura larvae to methomyl and chlorpyrifos after exposure to maize HIPVs. The larvae were exposed to HIPVs released from living maize plants as showed in Figure S1. Larval mortality was measured 24 hours after exposure to maize HIPVs, and methomyl (A) and chlorpyrifos (B). Asterisks indicate significant differences in comparison with unexposed control (Student’s t-test, ** P<0.01); Tolerance to methomyl (C) and chlorpyrifos (D) after exposure of S. frugiperda larvae to maize HIPVs and the insecticide synergist piperonyl butoxide (PBO). Data are mean ± S.D. (n=20).
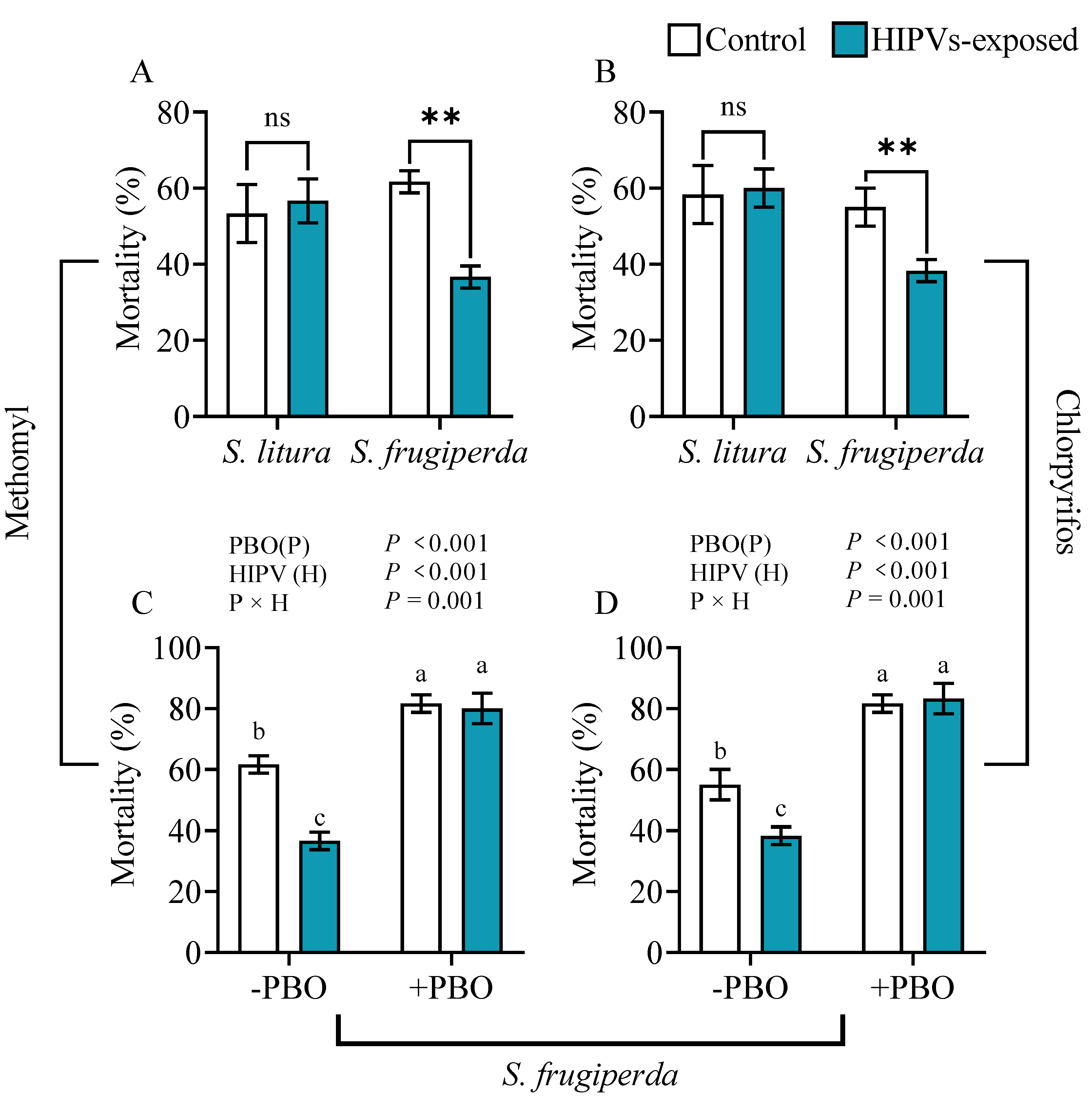
Figure 3.
Effects of larval exposure to maize HIPVs on the developmental process of S. litura and S. frugiperda. (A) egg hatching rate, (B) pupation rate, and (C) emergence rate. Data mean ± SE of (n=100). Asterisks indicate significant differences in comparison with unexposed control (ns, P>0.05).
Figure 3.
Effects of larval exposure to maize HIPVs on the developmental process of S. litura and S. frugiperda. (A) egg hatching rate, (B) pupation rate, and (C) emergence rate. Data mean ± SE of (n=100). Asterisks indicate significant differences in comparison with unexposed control (ns, P>0.05).
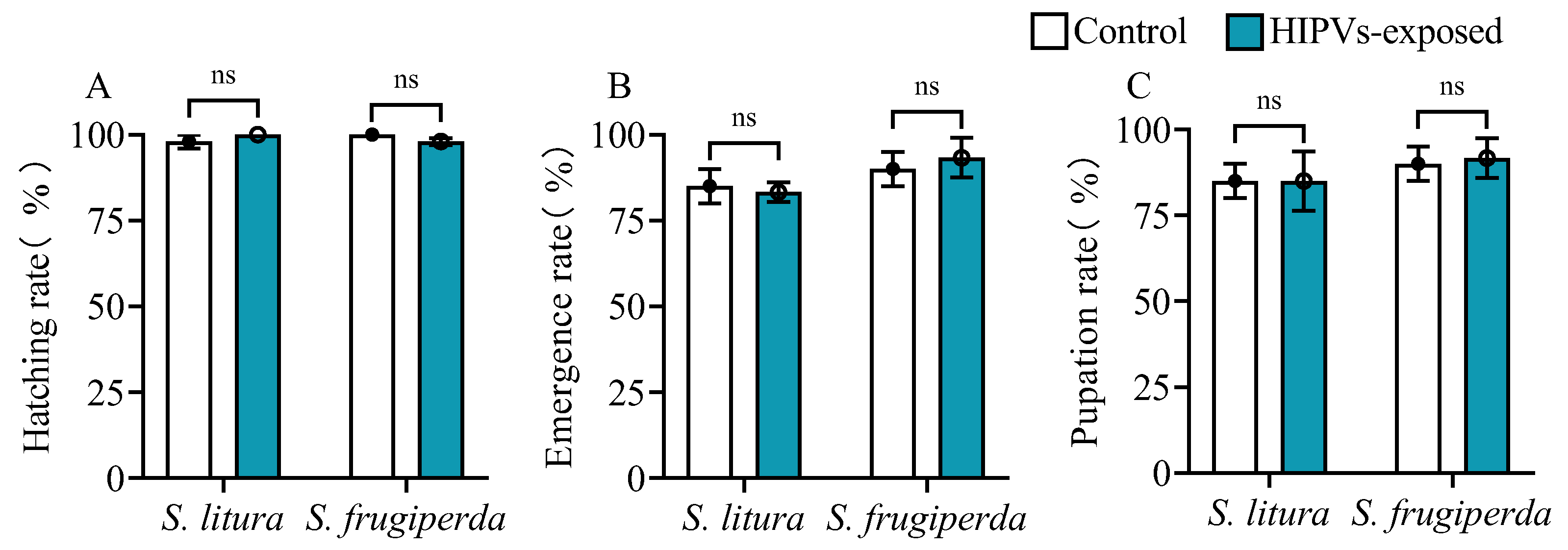
Figure 4.
Activities of cytochrome P450 enzymes (P450s), glutathione-s-transferase (GST) and carboxylesterase (CarE) in the midguts and fat bodies of 4th instars of S. litura and S. frugiperda after exposure to maize HIPVs and feeding on DIMBOA-contained diets. The larvae were exposed to maize HIPVs and 1 μg·g-1 DIMBOA diet for 24 h. Tissues dissected from five larvae were pooled and four biological replicates were run for each treatment. Data mean ± SE (n=4). Asterisks indicate significant differences in comparison with control (Student’s t-test if the interaction between HIPVs and DIMBOA was not significant, ** P<0.01). Different letters above bars indicate significant differences among treatments (p< 0.05) according to two-way ANOVA with Tukey's multiple comparison test (if the interaction between HIPVs and DIMBOA was significant).
Figure 4.
Activities of cytochrome P450 enzymes (P450s), glutathione-s-transferase (GST) and carboxylesterase (CarE) in the midguts and fat bodies of 4th instars of S. litura and S. frugiperda after exposure to maize HIPVs and feeding on DIMBOA-contained diets. The larvae were exposed to maize HIPVs and 1 μg·g-1 DIMBOA diet for 24 h. Tissues dissected from five larvae were pooled and four biological replicates were run for each treatment. Data mean ± SE (n=4). Asterisks indicate significant differences in comparison with control (Student’s t-test if the interaction between HIPVs and DIMBOA was not significant, ** P<0.01). Different letters above bars indicate significant differences among treatments (p< 0.05) according to two-way ANOVA with Tukey's multiple comparison test (if the interaction between HIPVs and DIMBOA was significant).
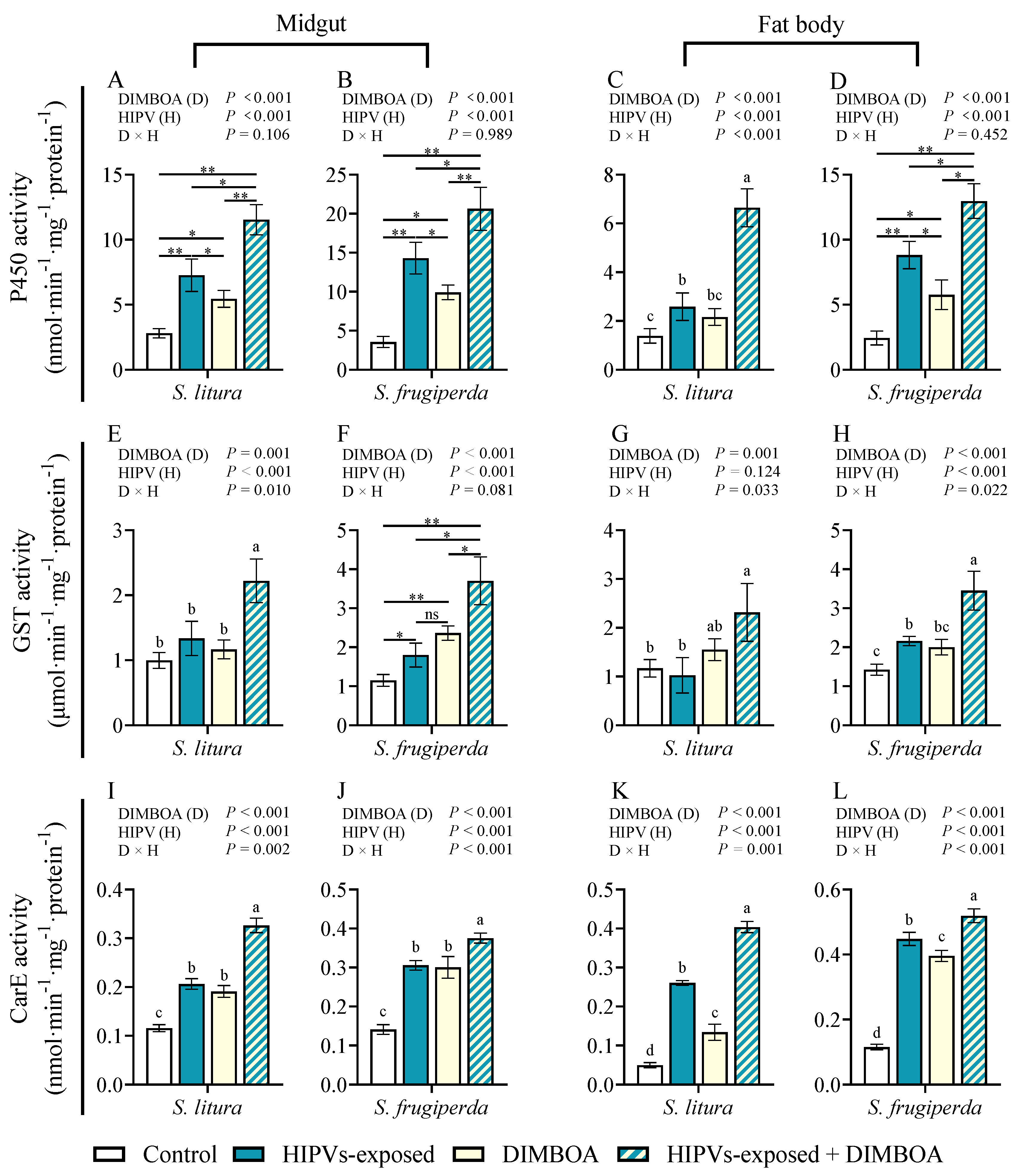
Figure 5.
Transcript levels of UGT33F28, UGT40L8, LOC118270728, CYP4d8 and CYP4V2 in the midguts and fat bodies of 4th instars of S. litura and S. frugiperda after exposure to maize HIPVs and feeding on DIMBOA-contained diets. The larvae were exposed to maize HIPVs and 1 μg·g-1 DIMBOA diet for 24 h. Tissues dissected from five larvae were pooled and three biological replicates were run for each treatment. Data mean ± SE (n=20). Asterisks indicate significant differences in comparison with control (Student’s t-test if the interaction between HIPVs and DIMBOA was not significant, ** P<0.01). Different letters above bars indicate significant differences among treatments (p < 0.05) according to two-way ANOVA with Tukey's multiple comparison test (if the interaction between HIPVs and DIMBOA was significant).
Figure 5.
Transcript levels of UGT33F28, UGT40L8, LOC118270728, CYP4d8 and CYP4V2 in the midguts and fat bodies of 4th instars of S. litura and S. frugiperda after exposure to maize HIPVs and feeding on DIMBOA-contained diets. The larvae were exposed to maize HIPVs and 1 μg·g-1 DIMBOA diet for 24 h. Tissues dissected from five larvae were pooled and three biological replicates were run for each treatment. Data mean ± SE (n=20). Asterisks indicate significant differences in comparison with control (Student’s t-test if the interaction between HIPVs and DIMBOA was not significant, ** P<0.01). Different letters above bars indicate significant differences among treatments (p < 0.05) according to two-way ANOVA with Tukey's multiple comparison test (if the interaction between HIPVs and DIMBOA was significant).

Figure 6.
Weight gain of 4th instars of S. litura and S. frugiperda fed on diets containing 1 μg·g-1 DIMBOA and 3 μg·g-1 chlorogenic acid (CA) after exposure to volatile compound cis-3-hexen-1-ol (cis-3-HXO). The larvae were exposed to volatile cis-3-HXO as showed in Figure S2. Data mean ± SE (n=50). Asterisks indicate significant differences in comparison with unexposed control (Student’s t-test, ** P<0.01).
Figure 6.
Weight gain of 4th instars of S. litura and S. frugiperda fed on diets containing 1 μg·g-1 DIMBOA and 3 μg·g-1 chlorogenic acid (CA) after exposure to volatile compound cis-3-hexen-1-ol (cis-3-HXO). The larvae were exposed to volatile cis-3-HXO as showed in Figure S2. Data mean ± SE (n=50). Asterisks indicate significant differences in comparison with unexposed control (Student’s t-test, ** P<0.01).
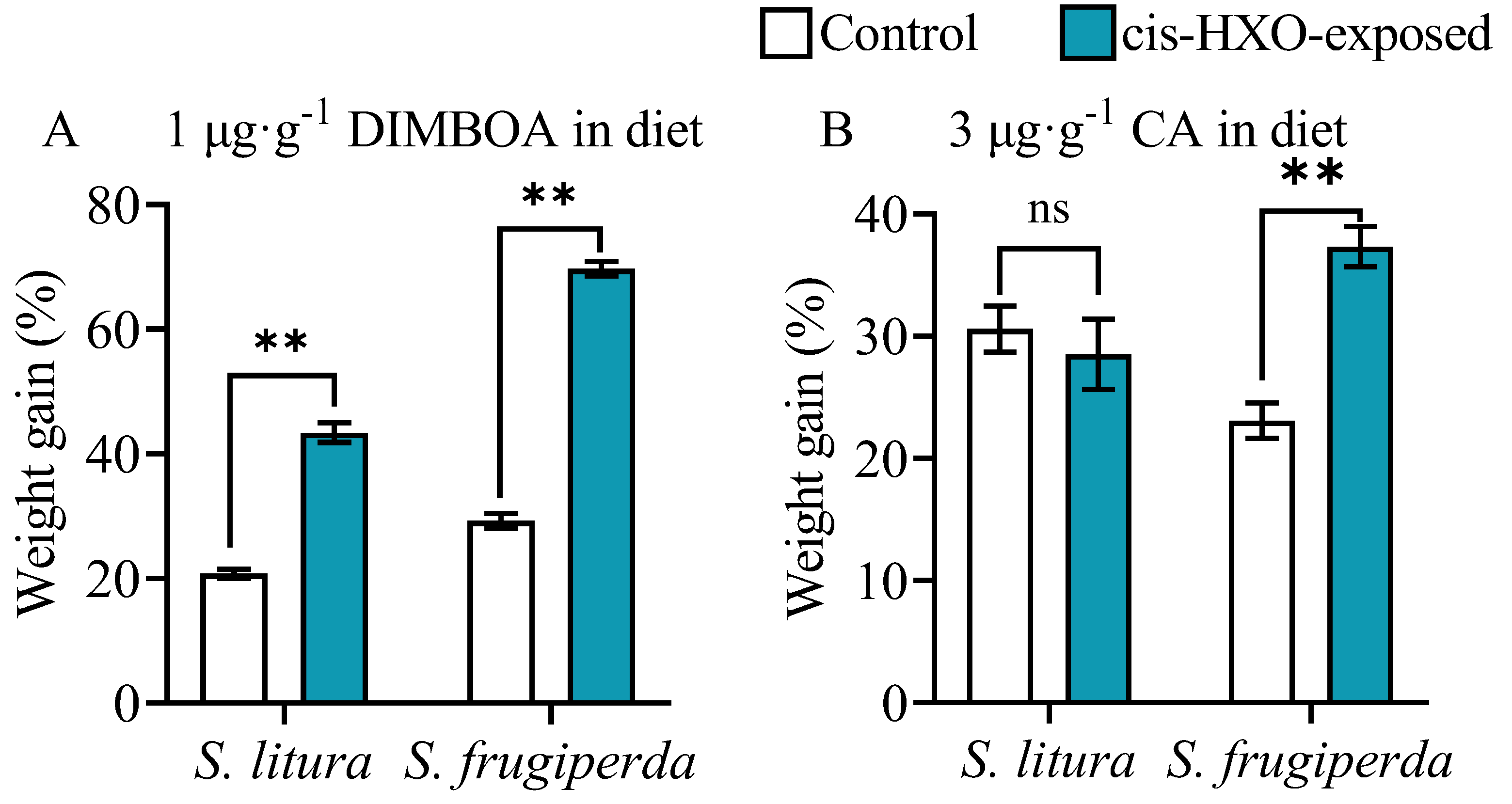
Figure 7.
Activities of cytochrome P450 enzymes (P450s), glutathione-s-transferase (GST) and carboxylesterase (CarE) in the midguts and fat bodies of 4th instars of S. litura and S. frugiperda after exposure to cis-3-hexen-1-ol (cis-3-HXO) and feeding on DIMBOA-contained diets. The larvae were exposed to cis-3-hexen-1-ol (cis-3-HXO) and 1 μg·g-1 DIMBOA diet for 24 h. The other were described in Figure 6. .
Figure 7.
Activities of cytochrome P450 enzymes (P450s), glutathione-s-transferase (GST) and carboxylesterase (CarE) in the midguts and fat bodies of 4th instars of S. litura and S. frugiperda after exposure to cis-3-hexen-1-ol (cis-3-HXO) and feeding on DIMBOA-contained diets. The larvae were exposed to cis-3-hexen-1-ol (cis-3-HXO) and 1 μg·g-1 DIMBOA diet for 24 h. The other were described in Figure 6. .
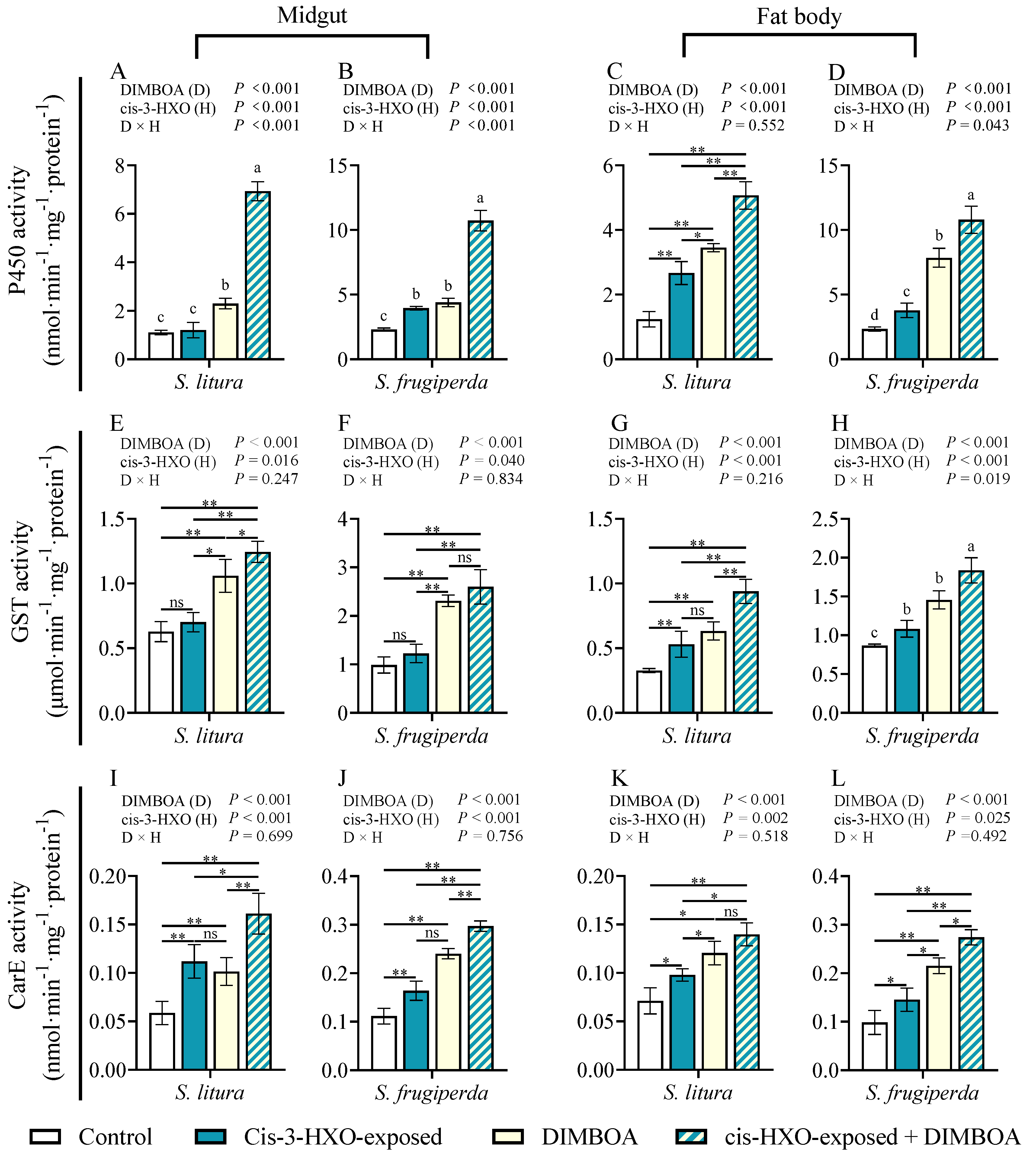
Figure 8.
Transcript levels of UGT33F28 and UGT40L8 in the midguts and fat bodies of 4th instars of S. litura and S. frugiperda after exposure to volatile cis-3-hexen-1-ol (cis-3-HXO) and feeding on DIMBOA-contained diets. The larvae were exposed to cis-3-HXO and 1 μg·g-1 DIMBOA diet for 24 h. The other were described in Figure 7.
Figure 8.
Transcript levels of UGT33F28 and UGT40L8 in the midguts and fat bodies of 4th instars of S. litura and S. frugiperda after exposure to volatile cis-3-hexen-1-ol (cis-3-HXO) and feeding on DIMBOA-contained diets. The larvae were exposed to cis-3-HXO and 1 μg·g-1 DIMBOA diet for 24 h. The other were described in Figure 7.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



