
Submitted:
20 November 2024
Posted:
21 November 2024
You are already at the latest version
Abstract
In 2022, an outbreak of H5N1 highly pathogenic avian influenza (HPAI) killed 60% of the largest breeding colony of Dalmatian pelican (DP) in the world, at Mikri Prespa Lake (Greece), prompting a multidisciplinary study on HPAI and other pathogens. This study determines the antimicrobial resistance rates of cloacal enterococci and Escherichia coli in DPs. Fifty-two blood and cloacal swab samples were collected from 31 nestlings (20 DP/11 great white pelicans) hatched after the H5N1 outbreak at the Prespa colony and 21 subadult/adult DPs captured at a spring migration stopover. Swabs were inoculated in non-selective and chromogenic selective media. Identification was performed by MALDI-TOF, and antimicrobial susceptibility was tested. The genetic content was characterized by PCR-sequencing and the clonality of extended-spectrum-betalactamase (ESBL)-producing-E. coli isolates by Multi-Locus-Sequence-Typing. Twenty-eight non-repetitive E. coli and 45 enterococci isolates were recovered in non-selective media; most of them were susceptible to all antibiotics tested (85.7% E. coli/91.1% enterococci). Three of 52 samples (6%, all adults) contained ESBL-E. coli isolates (detected in chromogenic ESBL plates), all carrying the blaCTX-M-15 gene and belonging to the lineage ST69. Despite the susceptibility of most fecal E. coli and enterococci isolates to all antibiotics tested, the identification of E. coli of lineage ST69 carrying blaCTX-M-15 is of concern. This high-risk clone needs further investigation to elucidate its primary sources and address the growing threat of antimicrobial resistance from an integrated 'One Health' perspective. Furthermore, it is imperative to further study the potential impacts of ESBL-E. coli on the endangered DP.
Keywords:
Dalmatian pelican
; antimicrobial resistance
; ESBL-E. coli
; One Health
1. Introduction
The Dalmatian pelican (Pelecanus crispus), hereafter referred to as “DP,” is an emblematic Palearctic bird species, ranging from Eastern Europe to China [1,2]. They live mainly in inland freshwater wetlands, rivers, lakes, deltas and estuaries, where they feed almost exclusively on fish [3]. The South-Eastern Europe (SEE) population of DP consists of short distance migrating populations from Greece, Albania, Montenegro, Bulgaria, Romania, Ukraine and Turkey [2,4]. In Greece, the birds belonging to the eastern group of populations (from four breeding colonies: Prespa, Chimaditida, Kerkini and Karla lakes) winter close to their breeding grounds, and usually migrate to wetlands of north/northeastern Greece and western Turkey [4,5]. The largest breeding colony of DPs in the world is located at Mikri Prespa Lake, Greece [6,7], where they breed side by side with the great white pelican Pelecanus onocrotalus (GWP) [8,9], which shows a similar feeding behavior and habitat preferences [3]. However, GWPs are long-distance migrants [10,11] that fly through many important stop-over sites in their southward migration to African wetlands along the Rift Valley for wintering in the eastern Africa region [10,11,12].
Prior to 2022, due to enormous conservation efforts, the SEE DP population was growing, with an increasing population estimated at 2,831-3,094 breeding pairs, [11]. From February to April 2022, a HPAI outbreak (H5N1, clade 2.3.4.4b) resulted in the death of 1,734 adult DPs only at Prespa Lake (60% of the colony), and a total of 2,286 DPs deceased in Greece, turning this tragic event into the worst ecological disaster for Greek wildlife. Over 40% of the SEE breeding population was lost, corresponding to approximately 10% of the species’ global population. GWPs also died in small numbers, leading to the conclusion that DPs are more susceptible to H5N1 than their congener species [13]. Multidisciplinary research into HPAI and other pathogen carriage in DPs, including bacterial diversity and antimicrobial resistance (AMR) rates, was implemented to investigate the underlying reasons.
AMR has become one of the significant challenges to human and animal health globally [14], and multidrug-resistant (MDR) bacteria, particularly Escherichia coli, have been detected in the gastrointestinal tract of wild birds living in a wide range of environments, from very anthropized locations like landfills to remote and less human-influenced ecosystems like the Arctic [15,16,17]. Extended-spectrum beta-lactamase (ESBL)-producing E. coli has been designated as critical priority in the World Health Organization's list of AMR "priority pathogens". This is attributed to its resistance to a wide range of antibiotics and its significant clinical relevance, as it is known to cause severe, life-threatening infections. [14]. Wild birds harboring ESBL-producing bacteria have been described by several authors in recent years, including other pelican species [17,18,19,20], which puts the spotlight on migratory birds and the role they could play as dispersers due to their ability to connect different ecosystems and remote locations [21,22,23,24]. It has been reported that DPs visit many different wetlands along their annual cycle [5,25], so even if their potential role as dispersers remains unclear, these birds could be quite useful as sentinels of AMR in the aquatic environment. From a conservation point of view, AMR in bacteria can be linked to strains with pathogenic potential that could negatively impact pelican health.
It will take decades for the DP to recover from the devastating effects of the recent avian influenza outbreak [13], so determining antibiotic-resistant bacteria that pelicans can carry and excrete is essential to have a better understanding of their epidemiology, besides highlighting other possible threats for these animals. The present study aims to determine the susceptibility of enteric E. coli and Enterococcus and the prevalence of ESBL and/or carbapenemase-producing (CP) E. coli in pelicans, as well as to understand the role of this species as sentinels of AMR.
2. Results
2.1. Recovery of E. coli and enterococci from cloacal samples of pelicans in non-supplemented media
After overnight culture on MacConckey agar (MCA) plates, MALDI-TOF identification, and antibiotic susceptibility testing, a collection of 28 non-repetitive E. coli isolates were obtained from 6/21 adults (28.6%) and 20/31 (64.5%) nestlings (10 DP/10 GWP). Moreover, after inoculating the cloacal samples in Slanetz-Bartley (S-B) plates, a total of 45 non-repetitive enterococci isolates were obtained from 11/21 adults (52.4%) and 30/31 (96.8%) nestlings (19 DP/11 GWP) (Table 1).
E. coli isolates were frequently retrieved from adult DP (n = 8, 28.6% of samples) and out of all the enterococci isolates (n = 11, 52.4%), E. faecalis, E. faecium (n = 4, 19% each), and E. hirae (n = 3, 14.3%) were the detected species. In the case of nestlings, four enterococcal species were identified with E. faecalis being the most frequent one (n = 19, 61.3%), followed by E. faecium (n = 10, 32.2%), E. hirae (n = 4, 12.9%) and E. mundtii (n = 1, 3.2%). E. coli was also frequently recovered (n = 20, 64.5%). Detailed information about prevalences and differences found in nestlings of the two pelican species, as well as total prevalences for both age groups of DP, are summarized in Table 1.
2.2. Phenotypes and genotypes Of Non-Repetitive E. coli and Enterococci Isolates Recovered In Non-Supplemented Media
Most of the E. coli isolates recovered in MCA media (n=24, 85.7%) were susceptible to all antibiotics tested (Figure 1), with only one isolate from a DP nestling showing resistance to tetracycline (carrying the tet(A) gene) and two isolates from adult DP with resistance to gentamicin. In addition, one isolate from an adult DP showed a multidrug-resistant pattern (MDR, resistant to at least 3 antimicrobial classes) with phenotypic ESBL production; this isolate carried the blaCTX-M15 gene, encoding the ESBL CTX-M-15 (Table 2).
Similarly, most enterococci recovered in S-B plates were fully susceptible to all antibiotics tested (n=41, 91.1%) (Figure 2), with four isolates belonging to three adults and from one DP nestling showing a resistance pattern to tetracycline (tet(M), n=4; tet(L), n=2) and to streptomycin (n=1) (Table 2). AMR rates for each antibiotic tested among the E. coli and enterococci isolates are detailed in Figure 3.
2.3. Prevalence of ESBL-producing E. coli (ESBL-Ec)
Subsequent seeding of the samples in ESBL- and carbapenem-resistance chromogenic media, resulted in the detection of ESBL-Ec isolates in three of the 21 adult pelicans (14.3%); as described above in one of these pelicans, ESBL-Ec had also been detected in the non-supplemented MCA media. After Multilocus-Sequence-Typing (MLST) of ESBL-Ec isolates all of them were assigned to the lineage ST69 (Table 3). None of the nestling samples were positive for ESBL-Ec. None of the pelicans tested (adults or nestlings) gave a growth in the carbapenem-chromogenic media, indicating the absence of Enterobacterales with this specific resistant phenotype.
2.4. Sexing
From a total of 21 adult pelicans, molecular sexing determined 12 males and 9 females. DP nestlings were identified as 9 males and 11 females while GWP were 5 males and 6 females.
2.5. Differences Between Sex, Age Groups and Species
A significantly higher prevalence of E. faecalis (χ2 = 6.304, d.f. = 1, p = 0.012) and E. coli (χ2 = 5.188, d.f. = 1, p = 0.023) was found in GWP nestlings, while the occurrence of E. faecium was significantly higher in DP nestlings (χ2 = 4.188, d.f. = 1, p = 0.041) (Table 1). The occurrence of AMR bacteria in nestlings did not differ between the two pelican species. To compare age groups, both nestling species were considered as a whole group without distinction. The prevalence of E. faecalis was significantly higher in nestlings (χ2 = 9.057, d.f. = 1, p = 0.03) and the prevalence of E. coli showed also a trend to be higher in this age group (χ2 = 3.516, d.f. = 1, p = 0.061). ESBL-Ec isolates were found only in adults (two males and one female), these being also the only isolates with a MDR phenotype, a finding that was statistically significant (χ2 = 4.7, d.f. = 1, p = 0.03) and the prevalence of resistant E. coli showed a trend to be higher in adults than in nestlings (χ2 = 3.606, d.f. = 1, p = 0.058) (Table 1). No differences were found between sexes in adults or nestlings from both species for prevalence of AMR/MDR bacteria, nor for the occurrence of any of the bacterial species isolated.
3. Discussion
This is the first general microbiological screening performed for AMR E. coli and enterococci in cloacal samples of free-living Dalmatian pelicans; so to our knowledge, all our findings can be potentially considered novel for this species. Except for the MDR isolates found in adults (carrying ESBL-Ec), most of the E. coli and enterococci recovered were pan-susceptible, showing low rates of resistance for the other antibiotics screened. Differences between the two species of nestlings were scarce, with similar occurrence of AMR and a higher prevalence of E. coli in GWP. The reason or clinical significance of the difference in the predominant species of enterococci has not been elucidated, with E. faecalis more present in GWP while E. faecium was more common in DP. Few significant differences were observed between adults and nestlings, highlighting the ESBL strains mentioned above and the tendency (p < 0.1) for E. coli with any resistant phenotype, to be more prevalent in adults.
As previously mentioned, one important result obtained in this study is the high proportion of pan-susceptible E. coli and enterococci isolates (susceptibility to all antibiotics tested) obtained when non-supplemented media were used. This is relevant data that could reflect a relatively limited environmental contamination with MDR/AMR bacteria where the animals live. Nevertheless, despite this fact, another remarkable result obtained in this study was the isolation of ESBL-Ec with the blaCTX-M-15 gene and belonging to the lineage ST69 in three adult birds.
Third-generation cephalosporin-resistant Enterobacteriaceae are included in the critical group of the WHO Bacterial Priority Pathogen List [14] and, therefore, are considered a highly important finding due to their clinical impact on public health. Among the CTX-M family, CTX-M15 ranks first in prevalence in most parts of the world [26], except Greece, where SHV-type was most frequent [27]. Regarding ST69, it is considered a pandemic lineage like ST131, ST95 or ST73, and it has been frequently reported from both community and nosocomial infections [28,29]. After ST131, this lineage is the most frequent clone, present in both humans and animals [29,20], and its effective transmission was highlighted in a household transfer of CTX-M group 9 E. coli ST69 strain between pet dogs and humans [31]. The dissemination of this high-risk clone can contribute to an accelerated spread of CTX-M15 beta-lactamase globally and could additionally be spread through avian migration, as was suggested for wintering rooks (Corvus frugilegus) [32].
In 2022, ECDC reported that in Greece, antimicrobial consumption was nearly twofold the average total consumption of European Union (EU) countries [33]. Having one of the highest rates of antibiotic consumption explains rates of antibiotic resistance consistently higher than EU average [34]. ESBL in human isolates have been frequently described in Greece [27]; regarding food and food-producing animals, EFSA/ECDC described an ESBL prevalence of 33.8% in fattening pigs in Greece in 2021/2022, and lower prevalences in pig and cattle meat (5.3% and 4%, respectively) [35].
Considering the aforementioned AMR prevalences, it could be reasonable to think that these bacteria and genes could transfer to Greek aquatic ecosystems, where pelicans and other wildlife are exposed to them. Antibiotic-resistant bacteria (ARB) reach rivers and lakes through many sources, such as hospital effluents and wastewater treatment plant discharges and leaching from proximate farms [36,37,38]. Once in the aquatic ecosystem, ARB can persist and spread despite the natural processes of dilution and degradation [37]. Moreover, antibiotic-resistant genes (ARGs) remain and reach other bacteria of public health concern through horizontal gene transfer [39]. In both freshwater and seawater from Greece, several ARBs and ARGs have been described, even in uninhabited coastal sites with presumptively less influx of wastewater [40,41,42]. It is worth mentioning that Greece is one of the major European marine aquaculture producers [43], and this important sector is generally related to a considerable use of antibiotics [44,45]. However, pelicans primarily use freshwater wetlands, and these are not related to any aquaculture activity.
One important pathway for acquiring ARGs is through livestock, which is considered a significant reservoir of ESBL and has a high prevalence of CTX-M clusters [46,47]. Leachates from farms located near wetlands or lakes can end up in water; even the cattle sometimes come to the lakes to graze and drink and could contribute to AMR/ARGs contamination of the aquatic environment [37,38], where it can eventually get into contact with wildlife. Many ESBL-producing Enterobacteriaceae have also been reported in a wide diversity of wildlife species [48], although most detections belong to wild birds with family CTX-M as the most prevalent. In Greece, ESBL genes in fecal samples from wild birds were first detected in Eurasian magpies (Pica pica) [47]. The same research also analyzed samples from livestock, and results from cattle, pigs, and magpies corroborated the global tendency with all ESBL-Ec harboring blaCTX-M-1/15. In a more recent survey including 47 different species of wild birds in the same country [20], the blaCTX-M-1 group (along with blaTEM) was again isolated in 3.3% of samples. To our knowledge, this is the first report of CTX-M15 E. coli in DPs; the presence of these strains of clinical interest in their gut suggests considering their usefulness as sentinels of AMR contamination of aquatic environments to which pelicans are closely related.
Finding an epidemic clone like ST69 in these animals is also important due to their spatial ecology. DPs use numerous wetlands around the year [5,25,49], connecting even five different countries according to previous GPS tracking of the Greek breeding population (Figure 4). In addition, GWP is a long-distance migratory bird that interconnects the European, Asian and African continents [12]. The migratory movements of both species of pelicans, making use of diverse wetlands across several countries and continents, and getting in contact with resident pelicans and other birds highlight the potential capacity of these animals to carry and disperse AMR and ARGs, as described for other wild birds [16, 18, 50].
Finally, ESBL-Ec prevalence found in this study (14.3% of adults sampled) is in accordance with previous reports for other adult wild birds [51,52,53], although they differ with other studies reporting prevalences higher than 40% [15,54,55]. However, a much lower prevalence was observed in wild birds in Greece [20], and in other European surveys [56,57]. Regional differences in some cases could explain this discrepancy in the results, but another possible explanation suggested by other authors [20] could be the species studied, as some researchers reporting high prevalences have focused on species strongly associated with anthropogenic diet and environments, like seagulls and storks that forage in human wastes or landfills [15,57,58]. A lower prevalence was expected in nestlings from our study, and no ESBL-Ec was found, although some authors have also described it in wild nestlings [17]. Further investigations are needed to look for temporal patterns of ESBL-Ec prevalence and thus persistence of carriage in DP and GWP. Expanding sampling to other breeding colonies (including those where DPs do not come in contact with GWPs) will allow to obtain more data and elucidate different factors involved in the acquisition of these critical bacteria in pelicans.
4. Material and Methods
4.1. Study Sites
Samples of pelicans from two different locations in Greece [Mikri Prespa Lake (N40°45′, E21°06′) and Kerkini Lake (N41°12′, E23°09′)] were collected (Figure 4), and both locations are quite connected through frequent movements of adult pelicans [49]. Mikri Prespa Lake is the largest breeding colony of DP in the world with 1,226-1,585 pairs until 2022 [59] and is also the nesting site for the majority of GWPs in Greece, with 500-780 pairs [60]. Both species nest side by side, forming large colonies that allow them to strengthen their sense of safety and consequently increase their breeding success [8]. In 2022, after the massive mortality caused by the HPAI outbreak, around 100 pairs of DPs were able to complete the reproductive process and raise 90 chicks [13], while GWPs were not affected. A total of 31 cloacal samples (20 DP/11 GWP) were obtained in July 2022 from nestlings belonging to these remaining breeding pairs.
The second location was Kerkini Lake, that supports the greatest diversity of waterbird species in freshwater in Greece, besides being an important wintering and stop-over site for many species, including many of the DPs that breed at Prespa [4,5,49]. The HPAI outbreak also affected the DP population in Kerkini, but to a lesser extent than in Mikri Prespa Lake [13]. In January 2023, a total of 21 DPs were sampled at Kerkini Lake, mostly adults but also some second and third-year immatures.
4.2. Sampling
Adults were captured using leg-hold traps set on the shore, or by hand from a boat, after being attracted very close to the boat with fish as bait, while nestlings were approached in their nests and gently restrained by manual handling or trapped from a boat using a fishing net pole. Cloacal samples were taken and were stored in Amies transport medium (Aptaca, Caneli, Italy), kept refrigerated in the field and frozen at -20oC for sending to the laboratory, where they were frozen at -80oC until further processing. Blood samples were taken from the medial metatarsal vein and kept in sterile tubes with heparin lithium as anticoagulant. Tubes were spun at 10000 rpm for two minutes to separate plasma from bottom pellet and both were kept separated and frozen at -20oC for further analysis.
4.3. Bacterial Isolation And Identification
Cloacal swabs were inoculated in 3ml of brain heart infusion (BHI) broth for enrichment and the resulting suspension was incubated at 37oC for 24 hours. For enterobacteria and enterococci isolation, MCA and S-B agar plates were used, adding 50 µL of enriched suspension in each one for streaking over the plates; from these plates E. coli and enterococci were recovered in media not supplemented with antibiotics to obtain a collection of non-selected isolates. After incubation at 37oC for 24 hours, one or two isolated colonies morphologically compatible with E. coli (MCA) or enterococci (S-B) were selected and identified by MALDI-TOF mass spectrometry (MALDI Biotyper®, Bruker).
Moreover, 50-100 µL of suspension was also spread over chromogenic selective media for detection of ESBL producing bacteria (Brilliance™ ESBL Agar; Oxoid) and carbapenem resistant (CP) bacteria (Brilliance™ CRE Medium; Oxoid), to analyze the prevalence of samples with E. coli carrying these specific resistance mechanisms. Plates were incubated at 37oC for 24h and colonies with the compatible morphology were identified by MALDI-TOF, as previously indicated.
4.4. Antimicrobial Susceptibility Testing
All identified isolates as E. coli or enterococci were tested for antimicrobial susceptibility by the disc-diffusion method, following methodology and breakpoints of the Clinical Laboratory Standards Institute (CLSI) [61]. Twelve antibiotics were tested for E. coli: ampicillin, amoxicillin/clavulanate, cefoxitin, cefotaxime, ceftazidime, imipenem, gentamicin, tobramycin trimethoprim/sulfamethoxazole, chloramphenicol, ciprofloxacin, and tetracycline. The double disc synergy test, using cefotaxime, ceftazidime, and amoxicillin/clavulanate discs, was used for the detection of phenotypic ESBL production [61]. In the case of enterococci, eleven different antibiotics were tested: vancomycin, teicoplanin, penicillin, ampicillin, tetracycline, erythromycin, gentamicin, streptomycin, linezolid, chloramphenicol and ciprofloxacin. In the collection of isolates recovered in MCA and S-B plates, non-repetitive isolates were considered for further analysis, and they represented one isolate of each species per sample, or more than one if they presented different antibiotic resistance phenotypes. For ESBL-Ec isolates recovered in chromogenic Brilliance™ ESBL Agar media, one isolate per sample was considered and further characterized.
4.5. E. coli and Enterococci DNA Extraction
The DNA extraction of E. coli isolates was performed suspending fresh isolated colonies in 1 ml of sterile Milli-Q water and incubating in boiled water at 100oC for 8 minutes. After that, suspension was thoroughly mixed by vortex and centrifuged at 12000 revolutions per minute (rpm) for 2 min. The supernatant with DNA was stored at -20 oC. For DNA extraction of enterococci isolates, fresh colonies were suspended in 1 ml of sterile Milli-Q water, mixed by vortex and centrifuged at 12000 rpm for 3 min. After removing the supernatant, 20 µL of Insta-Gene matrix (Bio-Rad Laboratories, Hercules, CA, USA) was added to the sediment, mixed by vortex and incubated in bath at 56oC for 20 min. Another incubation was repeated at 100o C for 8 min and centrifuged at 12000 rpm for 3 min. The resultant supernatant with DNA was stored at -20oC.
4.6. Molecular Characterization Of Antibiotic-Resistant Genes
Based on the resistant phenotype, genes conferring resistance to tetracycline (tet(A)/tet(B) for E. coli and tet(K)/tet(M)/tet(L) for enterococci), aminoglycosides (aac(3’)-II), vancomycin (vanA/vanB) and streptomycin (str) were studied by PCR. In addition, all phenotypic ESBL-Ec isolates were selected for blaSHV, blaCTX-M, blaTEM and blaOXA screening, and the resultant PCR products were further analyzed using sanger sequencing and confirmed by mapping against the NCBI database. The clonality of ESBL-Ec isolates were characterized by MultiLocus Sequence Typing (MLST) based on the Achtman scheme, amplifying and sequencing the seven housekeeping genes (adk, fumC, icd, gyrB, mdh, purA and recA) and comparing them with the MLST database to assign sequenced types (ST) (https://pubmlst.org/organisms/escherichia-spp). Conditions of PCRs and primers for genes tested, as well as the molecular typing, are described in Supplementary Tables S1 and S2.
4.7. Sex Determination
DNA was extracted from centrifuged blood, taking 5 µL of bottom pellet diluted in 1 ml of Milli-Q water, and boiled at 100oC for 10 minutes. The resultant DNA was used in a modified version of the PCR described by Fridolfsson and Ellegren [62], using the commercial kit Phusion blood direct PCR (ThermoFisher, Madrid, Spain). Horizontal electrophoresis in a 1.5% agarose gel was carried out to allow for separation and posterior visualization of DNA bands.
4.8. Statistical Analysis
Data analysis was run in the Statistical Package for Social Sciences (SPSS) Version 28 (IBM, California, USA). The association between the carriage rate of different bacteria and the bird species/age/sex groups was compared using the chi-square test at a 95% confidence interval. All analyses were considered statistically significant with probability values less than 0.05.
5. Conclusions
The present study describes the prevalence of enterococcal cloacal microbiota and E. coli in pelican nestlings from two different species and DP adults. Specifically, this study focused on phenotypic and genotypic characteristics of isolates with clinical interest, with clonality characterization of ESBL-producing E. coli due to its importance for Public Health.
We can outline that the high rate of susceptibility of most of the E. coli and enterococci recovered from the tested pelicans is in line with the habitat they live in. Nevertheless, at the same time, the detection of ESBL-producing E. coli isolates of the epidemic clone ST69 carrying the gene encoding CTX-M-15 in species like DPs, feeding quite restricted diets and under a moderate influence of anthropogenic activities is of relevance. This fact makes further investigation mandatory to clarify the main sources and face the AMR threat from a 'One Health' approach.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Primers and protocols employed in the PCR analyses to detect genes encoding for resistance to different antibiotics selected based on resistant phenotype.; Table S2: Primers and protocols employed in the PCR analyses to detect MLST genes. References [63-71] are cited in the Supplementary Materials.
Author Contributions
Conceptualization, T.C., U.H. and C.T.; methodology, T.C., S.M., C.G. and C.J.G.; formal analysis, T.C., S.M, C.G. and C.T.; investigation, O.A, G.C. and P.A.; data curation, T.C., U.H. and C.T.; writing—original draft preparation, T.C., U.H. and C.T.; writing—review and editing, S.M., C.G., O.A., G.C. and P.A.; supervision, U.H., C.T. and G.C.; funding acquisition, U.H., O.A. and G.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the project Ecoepidemiology of avian influenza and other diseases in Dalmatian pelicans between the Society for the Protection of Prespa and CSIC (Contract No. 20230896), and by the project PID2022-139591OB-I00 financed by MICIU/AEI/10.13039/501100011033 and FEDER, EU.
Institutional Review Board Statement
Capturing, handling and sampling wild birds complied with European and national legislation. Official permissions for capturing and sampling pelicans were provided by the Hellenic Ministry of Environment and Energy (74506/2354/20-7-2022 and 123240/4001/6-12-2022).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are reported in the manuscript.
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Crivelli, A.J.; Vizi, O. The Dalmatian pelican Pelecanus crispus Bruch 1832, a recently world-endangered bird species. Biol. Conserv. 1981, 20, 297–310. [CrossRef]
- Catsadorakis, G.; Portolou, D. International single species action plan for the conservation of the Dalmatian Pelican (Pelecanus crispus). CMS Technical Series No. 39, AEWA Technical Series No. 69. EAAFP Technical Report No. 1. Bonn, Germany and Incheon, South Korea. 2018.
- Del Hoyo, J.; Elliott, A.; Sargatal, J. Handbook of the Birds of the World; Lynx Editions: Barcelona, Spain, 1992; Volume 1, pp. 290-311.
- Catsadorakis, G.; Onmuş, O.; Bugariu, S.; Gül, O.; Hatzilacou, D.; Hatzofe, O.; Malakou, M.; Michev, T.; Naziridis, T.; Nikolaou, H.; Rudenko, A.; Saveljic, D.; Shumka, S.; Siki, M.; Crivelli, A.J. Current status of the Dalmatian pelican and the great white pelican populations of the Black Sea-Mediterranean flyway. Endanger Species Res. 2015, 27, 119–130. [CrossRef]
- Georgopoulou, E.; Alexandrou, O.; Manolopoulos, A.; Xirouchakis, S.; Catsadorakis, G. Home range of the Dalmatian pelican in south-east Europe. Eur J Wildl Res 2023, 69. [CrossRef]
- Crivelli, AJ.; Catsadorakis, G.; Hatzilacou, D.; Hulea, D.; Malakou, M.; Marinov, M.; Michev, T.; Nazirides, T.; Peja, N.; Sarigul, G.; Sıkı, M. Status and population development of Great white and Dalmatian pelicans, Pelecanus onocrotalus and P. crispus breeding in the Palearctic. 5th Medmaravis Pan-Mediterranean Seabird Symposium ‘Monitoring and conservation of birds, mammals and sea turtles of the Mediterranean and Black Seas’, Gozo, Malta, 2000; pp 38–45.
- Catsadorakis, G.; Crivelli, A.J. Nesting Habitat Characteristics and Breeding Performance of Dalmatian Pelicans in Lake Mikri Prespa, NW Greece. Waterbirds: The International Journal of Waterbird Biology 2001, 24, 386-393. [CrossRef]
- Crivelli, A.J.; Hatzilacou, D.; Catsadorakis, G. The breeding biology of Dalmatian Pelican Pelecanus crispus. Ibis 1998, 140, 472-481. [CrossRef]
- BirdLife International. Pelecanus onocrotalus. The IUCN Red List of Threatened Species 2021: e.T22697590A177120498. Available online: https://dx.doi.org/10.2305/IUCN.UK.2021-3.RLTS.T22697590A177120498.en (Accessed on 12 September 2024).
- Crivelli, A.J.; Leshem, Y.; Mitchev, T.; Jerrentrup, H. Where do Palaearctic great white pelicans (Pelecanus onocrotalus) presently overwinter?. Revue d’Ecologie,Terre et Vie 1991, 46, 145-171. [CrossRef]
- Catsadorakis, G. An update of the two Pelecanus species in the Mediterranean–Black Sea region. In: Conservation of marine and coastal birds in the Mediterranean. Proc. UNEP-MAPRAC/SPA Symposium, Hamammet, Tunisia, 20 to 22 February 2015; Yésou, P.; Sultana, J.; Walmsley J.; Azafzaf, H. (Eds), 2016, pp. 47–52.
- Izhaki, I.; Shmueli, M.; Arad, Z.; Steinberg, Y.; Crivelli, A.J. Satellite tracking of migratory and ranging behaviour of inmature Great white pelicans. Waterbirds 2002, 25, 295-304. [CrossRef]
- Alexandrou, O.; Malakou, M.; Catsadorakis, G. The impact of avian influenza 2022 on Dalmatian pelicans was the worst ever wildlife disaster in Greece. Oryx 2022, 56, 813-813. [CrossRef]
- WHO Bacterial Priority Pathogens List, 2024: bacterial pathogens of public health importance to guide research, development and strategies to prevent and control antimicrobial resistance. Geneva: World Health Organization; 2024. License: CC BY-NC-SA 3.0 IGO. Available online: https://iris.who.int/bitstream/handle/10665/376776/9789240093461-eng.pdf (Accessed on 13 September 2024).
- Höfle, U.; Gonzalez-Lopez, J.; Camacho, M.C.; Solà-Ginés, M.; Moreno-Mingorance, A.; Manuel Hernández, J.; De La Puente, J.; Pineda-Pampliega, J.; Aguirre J.I.; Torres-Medina, F.; Ramis, A.; Majó, N.; Blas, J.; Migura-García, L. Foraging at Solid Urban Waste Disposal Sites as Risk Factor for Cephalosporin and Colistin Resistant Escherichia coli Carriage in White Storks (Ciconia ciconia). Front Microbiol 2020, 11, 1397. [CrossRef]
- Akhil Prakash, E.; Hromádková, T.; Jabir, T.; Vipindas, P.V.; Krishnan, K.P.; Mohamed Hatha, A.A.; Briedis, M. Dissemination of multidrug resistant bacteria to the polar environment - Role of the longest migratory bird Arctic tern (Sterna paradisaea). Sci Total Environ 2022, 815, 152727. [CrossRef]
- Martínez-Álvarez, S.; Châtre, P.; Cardona-Cabrera, T.; François, P.; Sánchez-Cano, A.; Höfle, U.; Zarazaga, M.; Madec, J-Y.; Haenni, M.; Torres, C. Detection and genetic characterization of blaESBL-carrying plasmids of cloacal Escherichia coli isolates from white stork nestlings (Ciconia ciconia) in Spain. J Glob Antimicrob Resist 2023, 34, 186-194. [CrossRef]
- Suarez-Yana, T.; Salgado-Caxito, M.; Hayer, J.; Rojas-Sereno, Z.E.; Pino-Hurtado, M.S.; Campaña-Burguet, A.; Caparrós, C.; Torres, C.; Benavides, J.A. ESBL-producing Escherichia coli prevalence and sharing across seabirds of central Chile. Sci Total Environ 2024, 951, 175475. [CrossRef]
- Fuentes-Castillo, D.; Farfán-López, M.; Esposito, F.; Moura, Q; Fernandes, M.R.; Lopes, R.; Cardoso, B.; Muñoz, M.E.; Cerdeira, L.; Najle, I.; Muñoz, P.M.; Catão-Dias, J.L.; González-Acuña, D.; Lincopan, N. Wild owls colonized by international clones of extended-spectrum β-lactamase (CTX-M)-producing Escherichia coli and Salmonella Infantis in the Southern Cone of America. Sci Total Environ 2019, 674, 554-562. [CrossRef]
- Athanasakopoulou, Z.; Diezel, C.; Braun, S.D.; Sofia, M.; Giannakopoulos, A.; Monecke, S.; Gary, D.; Krähmer, D.; Chatzopoulos, D.C.; Touloudi, A.; et al. Occurrence and Characteristics of ESBL- and Carbapenemase- Producing Escherichia coli from Wild and Feral Birds in Greece. Microorganisms 2022, 10, 1217. [CrossRef]
- Ahlstrom, C.A.; van Toor, M.L.; Woksepp, H.; Chandler, J.C.; Reed, J.A.; Reeves, A.B.; Waldenström, J.; Franklin, A.B.; Douglas, D.C.; Bonnedahl, J.; Ramey, A.M. Evidence for continental-scale dispersal of antimicrobial resistant bacteria by landfill foraging gulls. Sci Total Environ 2021, 764, 144551. [CrossRef]
- Ahlstrom, C.A.; Woksepp, H.; Sandegren, L.; Mohsin, M.; Hasan, B.; Muzyka, D.; Hernandez, J.; Aguirre, F.; Tok, A.; Söderman, J.; Olsen, B.; Ramey, A.M.; Bonnedahl, J. Genomically diverse carbapenem resistant Enterobacteriaceae from wild birds provide insight into global patterns of spatiotemporal dissemination. Sci Total Environ 2022, 824, 153632. [CrossRef]
- Jarma, D.; Sánchez, M.I.; Green, A.J.; Peralta-Sánchez, J.M.; Hortas, F.; Sánchez-Melsió, A.; Borrego, C.M. Faecal microbiota and antibiotic resistance genes in migratory waterbirds with contrasting habitat use. Sci Total Environ 2021, 783, 146872. [CrossRef]
- Martínez-Álvarez, S.; Höfle, U.; Châtre, P.; Alonso, C. A.; Asencio-Egea, M. A.; François, P.; Cardona-Cabrera, T.; Zarazaga, M.; Madec, J.; Haenni, M.; Torres, C. One Health bottom-up analysis of the dissemination pathways concerning critical priority carbapenemase- and ESBL-producing Enterobacterales from storks and beyond. J Antimicrob Chemother 2024, in press. [CrossRef]
- Efrat, R.; Harel, R.; Alexandrou, O.; Catsadorakis, G.; Nathan, R. Seasonal differences in energy expenditure, flight characteristics and spatial utilization of Dalmatian pelicans Pelecanus crispus in Greece. Ibis 2018, 161, 415–427. [CrossRef]
- Peirano, G.; Pitout, J.D.D. Extended-Spectrum β-Lactamase-Producing Enterobacteriaceae: Update on Molecular Epidemiology and Treatment Options. Drugs 2019, 79, 1529–1541. [CrossRef]
- Kazmierczak, K.M.; de Jonge, B.L.M.; Stone, G.G.; Sahm, D.F. Longitudinal analysis of ESBL and carbapenemase carriage among Enterobacterales and Pseudomonas aeruginosa isolates collected in Europe as part of the International Network for Optimal Resistance Monitoring (INFORM) global surveillance programme, 2013-17. J Antimicrob Chemother 2020, 75, 1165–1173.
- Riley L.W. (2014). Pandemic lineages of extraintestinal pathogenic Escherichia coli. Clin Microbiol Infect 2014, 20, 380–390. [CrossRef]
- Kocsis, B.; Gulyás, D.; Szabó, D. Emergence and Dissemination of Extraintestinal Pathogenic High-Risk International Clones of Escherichia coli. Life 2022, 12, 2077. [CrossRef]
- Day, M.J.; Rodríguez, I.; van Essen-Zandbergen, A.; Dierikx, C.; Kadlec, K.; Schink, A.K.; Wu, G.; Chattaway, M.A.; DoNascimento, V.; Wain, J.; Helmuth, R.; Guerra, B.; Schwarz, S.; Threlfall, J.; Woodward, M.J.; Coldham, N.; Mevius, D.; Woodford, N. Diversity of STs, plasmids and ESBL genes among Escherichia coli from humans, animals and food in Germany, the Netherlands and the UK. J Antimicrob Chemother 2016, 71, 1178–1182. [CrossRef]
- Grönthal, T.; Österblad, M.; Eklund, M.; Jalava, J.; Nykäsenoja, S.; Pekkanen, K.; Rantala, M. Sharing more than friendship - transmission of NDM-5 ST167 and CTX-M-9 ST69 Escherichia coli between dogs and humans in a family, Finland, 2015. Euro Surveill 2018, 23, 1700497. [CrossRef]
- Jamborova, I.; Dolejska, M.; Vojtech, J.; Guenther, S.; Uricariu, R.; Drozdowska, J.; Papousek, I.; Pasekova, K.; Meissner, W.; Hordowski, J.; Cizek, A.; Literak, I. Plasmid-mediated resistance to cephalosporins and fluoroquinolones in various Escherichia coli sequence types isolated from rooks wintering in Europe. Appl Environ Microb 2015, 81, 648–657. [CrossRef]
- European Centre for Disease Prevention and Control (ECDC). Antimicrobial consumption in the EU/EEA (ESAC-Net) - Annual Epidemiological Report 2022. Stockholm: ECDC; 2023. Available online: https://www.ecdc.europa.eu/sites/default/files/documents/AER-antimicrobial-consumption.pdf (Accessed on 16 September 2024).
- Kopsidas, I.; Theodosiadis, D.; Triantafyllou, C.; Koupidis, S.; Fanou, A.; Hatzianastasiou S. EVIPNet evidence brief for policy: preventing antimicrobial resistance and promoting appropriate antimicrobial use in inpatient health care in Greece. Copenhagen: WHO Regional Office for Europe; 2022. Available online: https://iris.who.int/bitstream/handle/10665/361842/WHO-EURO-2022-5837-45602-65411-eng.pdf (Accessed on 8 July 2024).
- EFSA (European Food Safety Authority) and ECDC (European Centre for Disease Prevention and Control), 2024. The European Union Summary Report on Antimicrobial Resistance in zoonotic and indicator bacteria from humans, animals and food in 2021/2022. EFSA J 2024, 22, e8583. [CrossRef]
- Marti, E.; Variatza, E.; Balcazar, J.L. The role of aquatic ecosystems as reservoirs of antibiotic resistance. Trends Microbiol 2014, 22, 36–41. [CrossRef]
- Nnadozie, C.F.; Odume, O.N. Freshwater environments as reservoirs of antibiotic resistant bacteria and their role in the dissemination of antibiotic resistance genes. Environ Pollut 2019, 254(Pt B), 113067. [CrossRef]
- Samreen; Ahmad, I.; Malak, H.A.; Abulreesh, H.H. Environmental antimicrobial resistance and its drivers: a potential threat to public health. J Glob Antimicrob Resist 2021, 27, 101–111. [CrossRef]
- Pepi, M.; Focardi, S. Antibiotic-Resistant Bacteria in Aquaculture and Climate Change: A Challenge for Health in the Mediterranean Area. Int J Environ Res Public Health 2021, 18, 5723. [CrossRef]
- Nikolakopoulou, T.L.; Giannoutsou, E.P.; Karabatsou, A.A.; Karagouni, A.D. Prevalence of tetracycline resistance genes in Greek seawater habitats. J Microbiol 2008, 46, 633–640. [CrossRef]
- Kalantzi, I.; Rico, A.; Mylona, K.; Pergantis, S.A.; Tsapakis, M. Fish farming, metals and antibiotics in the eastern Mediterranean Sea: Is there a threat to sediment wildlife?. Sci total environ 2021, 764, 142843. [CrossRef]
- Anagnostopoulos, D. A.; Parlapani, F.F.; Natoudi, S.; Syropoulou, F.; Kyritsi, M.; Vergos, I.; Hadjichristodoulou, C.; Kagalou, I.; Boziaris, I.S. Bacterial Communities and Antibiotic Resistance of Potential Pathogens Involved in Food Safety and Public Health in Fish and Water of Lake Karla, Thessaly, Greece. Pathogens 2022, 11, 1473. [CrossRef]
- FAO. 2024. The State of World Fisheries and Aquaculture 2024 – Blue Transformation in action. Rome. Available online: https://openknowledge.fao.org/handle/20.500.14283/cd0683en (Accessed on 10 July 2024). [CrossRef]
- Lulijwa, R.; Rupia, Emmanuel.; Alfaro, Andrea. Antibiotic use in aquaculture, policies and regulation, health and environmental risks: a review of the top 15 major producers. Rev Aquac 2019, 12, 10.1111/raq.12344. [CrossRef]
- Preena, P.G.; Swaminathan, T.R.; Kumar, V.J.R.; Singh, I.S.B. (2020) Antimicrobial resistance in aquaculture: a crisis for concern. Biologia 2020,75, 1497–1517. [CrossRef]
- Ejaz, H.; Younas, S.; Abosalif, K.O.A.; Junaid, K.; Alzahrani, B.; Alsrhani, A.; Abdalla, A.E.; Ullah, M.I.; Qamar, M.U.; Hamam, S.S.M. Molecular analysis of blaSHV, blaTEM, and blaCTX-M in extended-spectrum β-lactamase producing Enterobacteriaceae recovered from fecal specimens of animals. PloS One 2021, 16, e0245126. [CrossRef]
- Athanasakopoulou, Z.; Reinicke, M.; Diezel, C.; Sofia, M.; Chatzopoulos, D.C.; Braun, S.D.; Reissig, A.; Spyrou, V.; Monecke, S.; Ehricht, R.; Tsilipounidaki, K.; Giannakopoulos, A.; Petinaki, E.; Billinis, C. Antimicrobial Resistance Genes in ESBL-Producing Escherichia coli Isolates from Animals in Greece. Antibiotics 2021, 10, 389. [CrossRef]
- Wang, J.; Ma, Z. B.; Zeng, Z.L.; Yang, X.W.; Huang, Y.; Liu, J.H. The role of wildlife (wild birds) in the global transmission of antimicrobial resistance genes. Zool Res 2017, 38, 55–80. [CrossRef]
- Bounas, A.; Catsadorakis, G.; Naziridis, T.; Bino, T.; Hatzilacou, D.; Malakou, M.; Onmus, O.; Siki, M.; Simeonov, P.; Crivelli, A. J. Site fidelity and determinants of wintering decisions in the Dalmatian pelican (Pelecanus crispus). Ethol Ecol Evol 2022, 35, 434–448. [CrossRef]
- Sandegren, L.; Stedt, J.; Lustig, U.; Bonnedahl, J.; Andersson, D.I.; Järhult, J.D. Long-term carriage and rapid transmission of extended spectrum beta-lactamase-producing E. coli within a flock of Mallards in the absence of antibiotic selection. Environ Microbiol Rep 2018, 10, 576-582. [CrossRef]
- Veldman, K.; van Tulden, P.; Kant, A.; Testerink, J.; Mevius, D. Characteristics of Cefotaxime-Resistant Escherichia coli from Wild Birds in The Netherlands. Appl Environ Microbiol 2013, 79, 7556–7561. [CrossRef]
- Alcalá, L.; Alonso, C.A.; Simón, C.; González-Esteban, C.; Orós, J.; Rezusta, A.; Ortega, C.; Torres, C. Wild Birds, Frequent Carriers of Extended-Spectrum β-Lactamase (ESBL) Producing Escherichia coli of CTX-M and SHV-12 Types. Microb Ecol 2016, 72, 861-869. [CrossRef]
- Prandi, I.; Bellato, A.; Nebbia, P.; Stella, M.C.; Ala, U.; von Degerfeld, M.M.; Quaranta, G.; Robino, P. Antibiotic resistant Escherichia coli in wild birds hospitalised in a wildlife rescue centre. Comp Immunol Microbiol Infect Dis 2023, 93, 101945. [CrossRef]
- Aires-de-Sousa, M.; Fournier, C.; Lopes, E.; de Lencastre, H.; Nordmann, P.; Poirel, L. High Colonization Rate and Heterogeneity of ESBL- and Carbapenemase-Producing Enterobacteriaceae Isolated from Gull Feces in Lisbon, Portugal. Microorganisms 2020, 8, 1487. [CrossRef]
- Vergara, A.; Pitart, C.; Montalvo, T.; Roca, I.; Sabaté, S.; Hurtado, J.C.; Planell, R.; Marco, F.; Ramírez, B.; Peracho, V.; de Simón, M.; Vila, J. Prevalence of Extended-Spectrum-β-Lactamase- and/or Carbapenemase-Producing Escherichia coli Isolated from Yellow-Legged Gulls from Barcelona, Spain. Antimicrob Agents Chemother 2017, 61, 10.1128/aac.02071-16. [CrossRef]
- Dotto, G.; Menandro, M.L.; Mondin, A.; Martini, M.; Tonellato, F.R.; Pasotto, D. Wild Birds as Carriers of Antimicrobial-Resistant and ESBL-Producing Enterobacteriaceae. Int J Infect Dis 2016, 53, 59. [CrossRef]
- Oteo, J.; Mencía, A.; Bautista, V.; Pastor, N.; Lara, N.; González-González, F.; García-Peña, F.J.; Campos, J. Colonization with Enterobacteriaceae-Producing ESBLs, AmpCs, and OXA-48 in Wild Avian Species, Spain 2015-2016. Microb Drug Resist 2018, 24, 932–938. [CrossRef]
- Jarma, D.; Sánchez, M.I.; Green, A.J.; Peralta-Sánchez, J.M.; Hortas, F.; Sánchez-Melsió, A.; Borrego, C.M. Faecal microbiota and antibiotic resistance genes in migratory waterbirds with contrasting habitat use. Sci Total Environ 2021, 783, 146872. [CrossRef]
- Catsadorakis, G.; Alexandrou, O.; Hatzilacou, D.; Kasvikis, I.; Katsikatsou, M.; Konstas, S.; Malakou, M.; Michalakis, D.; Naziridis, Th.; Nikolaou, H.; Noulas, N.; Portolou, D.; Roussopoulos, Y.; Vergos, I.; Crivelli, A.J. Breeding colonies, population growth and breeding success of the Dalmatian pelican (Pelecanus crispus) in Greece: a country-wide perspective, 1967-2021. Eur J Ecol 2024, 10, 63-86. [CrossRef]
- Handrinos, G.; Catsadorakis, G. The historical and current distribution of Dalmatian Pelican Pelecanus crispus and Great White Pelican Pelecanus onocrotalus in Greece and adjacent areas: 1830-2019. Avocetta 2020, 44, 11-20. [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing. 32nd ed. CLSI Supplement M100-Ed32. National Committee for Clinical Laboratory Standards, USA, 2022.
- Fridolfsson, A.K.; Ellegren, H. A simple and universal method for molecular sexing of non-ratite birds. J Avian Biol 1999, 30, 116–121. [CrossRef]
- Pagani, L.; Dell’Amico, E.; Migliavacca, R.; D’Andrea, M.M.; Giacobone, E.; Amicosante, G.; Romero, E.; Rossolini, G.M. Multiple CTX-M-type extended-spectrum β-lactamases innosocomial isolates of Enterobacteriaceae from a hospital in Northern Italy. J Clin Microbiol 2003, 41, 4264–4269. [CrossRef]
- Coque, T.M.; Oliver, A.; Pérez-Díaz, J.C.; Baquero, F.; Cantón, R. Genes encoding TEM-4, SHV-2, and CTX-M-10 extended-spectrum β-lactamases are carried by multiple Klebsiella pneumoniae clones in a single hospital (Madrid, 1989 to 2000). Antimicrob Agents Chemother 2002, 46, 500–510. [CrossRef]
- Pitout, J.D.D.; Thomson, K.S.; Hanson, N.D.; Ehrhardt, A.F.; Moland, E.S.; Sanders, C.C. β-lactamases responsible for resistance to expanded-spectrum cephalosporinsin Klebsiella pneumoniae, Escherichia coli, and Proteus mirabilis isolates recovered in South Africa. Antimicrob Agents Chemother 1998, 42, 1350–1354. [CrossRef]
- Belaaouaj, A.; Lapoumeroulie, C.; Caniça, M.M.; Vedel, G.; Névot, P.; Krishnamoorthy, R.; Paul, G. Nucleotide sequences of the genes coding for the TEM-like β-lactamases IRT-1 and IRT-2 (formerly called TRI-1 and TRI-2). FEMS Microbiol Lett 1994, 120, 75-80. [CrossRef]
- Steward, C.D.; Rasheed, J.K.; Hubert, S.K.; Biddle, J.W.; Raney, P.M.; Anderson, G.J.; Williams, P.P.; Brittain, K.L.; Oliver, A.; McGowan, J.E.Jr.; Tenover, F.C. Characterization of clinical isolates of Klebsiella pneumoniae from 19 laboratories using the National Committee for Clinical Laboratory Standards extended-spectrum beta-lactamase detection methods. J Clin Microbiol 2001, 39, 2864–2872. [CrossRef]
- Guardabassi, L.; Dijkshoorn, L.; Collard, J.M.; Olsen, J.E.; Dalsgaard, A. Distribution and in-vitro transfer of tetracycline resistance determinants in clinical and aquatic Acinetobacter strains. J Med Microbiol 2000, 49, 929–936. [CrossRef]
- Aarestrup, F.M.; AgersŁ, Y.; Ahrens, P.; JŁrgensen, J.C.; Madsen, M.; Jensen, L.B. Antimicrobial susceptibility and presence of resistance genes in staphylococci from poultry. Vet Microbiol 2000, 74, 353–364. [CrossRef]
- Schnellmann, C.; Gerber, V.; Rossano, A.; Jaquier, V.; Panchaud, Y.; Doherr, M.G.; Thomann, A.; Straub, R.; Perreten, V. Presence of new mecA and mph(C) variants conferring antibiotic resistance in Staphylococcus spp. isolated from the skin of horses before and after clinic admission. J Clin Microbiol 2006,44, 4444–4454. [CrossRef]
- Tartof, S.Y.; Solberg, O.D.; Manges, A.R.; Riley, L.W. Analysis of a uropathogenic Escherichia coli clonal group by multilocus sequence typing. J Clin Microbiol 2005, 43, 5860–5864. [CrossRef]
Figure 1.
Heat map showing antibiotic susceptibility/resistance of E. coli isolated in MacConkey media from cloacal swabs from great white and Dalmatian pelicans. Legend: DP, Dalmatian pelican; GWP, great white pelican; AMP, ampicillin; AMC, amoxicillin/clavulanate; FOX, cefoxitin; CTX, cefotaxime; CAZ, ceftazidime; IMP, imipenem; GEN, gentamicin; TOB, tobramycin; TET, tetracycline; CHL, chloramphenicol; SXT, trimethoprim/sulfamethoxazole and CIP, ciprofloxacin. Black squares, antibiotic resistant phenotype; grey squares, intermediate resistant phenotype; white squares, wild type/susceptible phenotype.
Figure 1.
Heat map showing antibiotic susceptibility/resistance of E. coli isolated in MacConkey media from cloacal swabs from great white and Dalmatian pelicans. Legend: DP, Dalmatian pelican; GWP, great white pelican; AMP, ampicillin; AMC, amoxicillin/clavulanate; FOX, cefoxitin; CTX, cefotaxime; CAZ, ceftazidime; IMP, imipenem; GEN, gentamicin; TOB, tobramycin; TET, tetracycline; CHL, chloramphenicol; SXT, trimethoprim/sulfamethoxazole and CIP, ciprofloxacin. Black squares, antibiotic resistant phenotype; grey squares, intermediate resistant phenotype; white squares, wild type/susceptible phenotype.

Figure 2.
Heat map showing antibiotic susceptibility of all Enterococcus strains from Great White and Dalmatian pelican cloacal swabs isolated in Slanetz-Bartley media. Legend: DP, Dalmatian pelican; GWP, great white pelican; VAN, vancomycin; TEC, teicoplanin; PEN, penicillin; AMP, ampicillin; TET, tetracycline; ERY, erythromycin; GEN, gentamicin; STR, streptomycin; LNZ, linezolid; CHL, chloramphenicol; and CIP, ciprofloxacin. Black squares, antibiotic resistant phenotype; grey squares, intermediate resistant phenotype; white squares, susceptible phenotype.
Figure 2.
Heat map showing antibiotic susceptibility of all Enterococcus strains from Great White and Dalmatian pelican cloacal swabs isolated in Slanetz-Bartley media. Legend: DP, Dalmatian pelican; GWP, great white pelican; VAN, vancomycin; TEC, teicoplanin; PEN, penicillin; AMP, ampicillin; TET, tetracycline; ERY, erythromycin; GEN, gentamicin; STR, streptomycin; LNZ, linezolid; CHL, chloramphenicol; and CIP, ciprofloxacin. Black squares, antibiotic resistant phenotype; grey squares, intermediate resistant phenotype; white squares, susceptible phenotype.
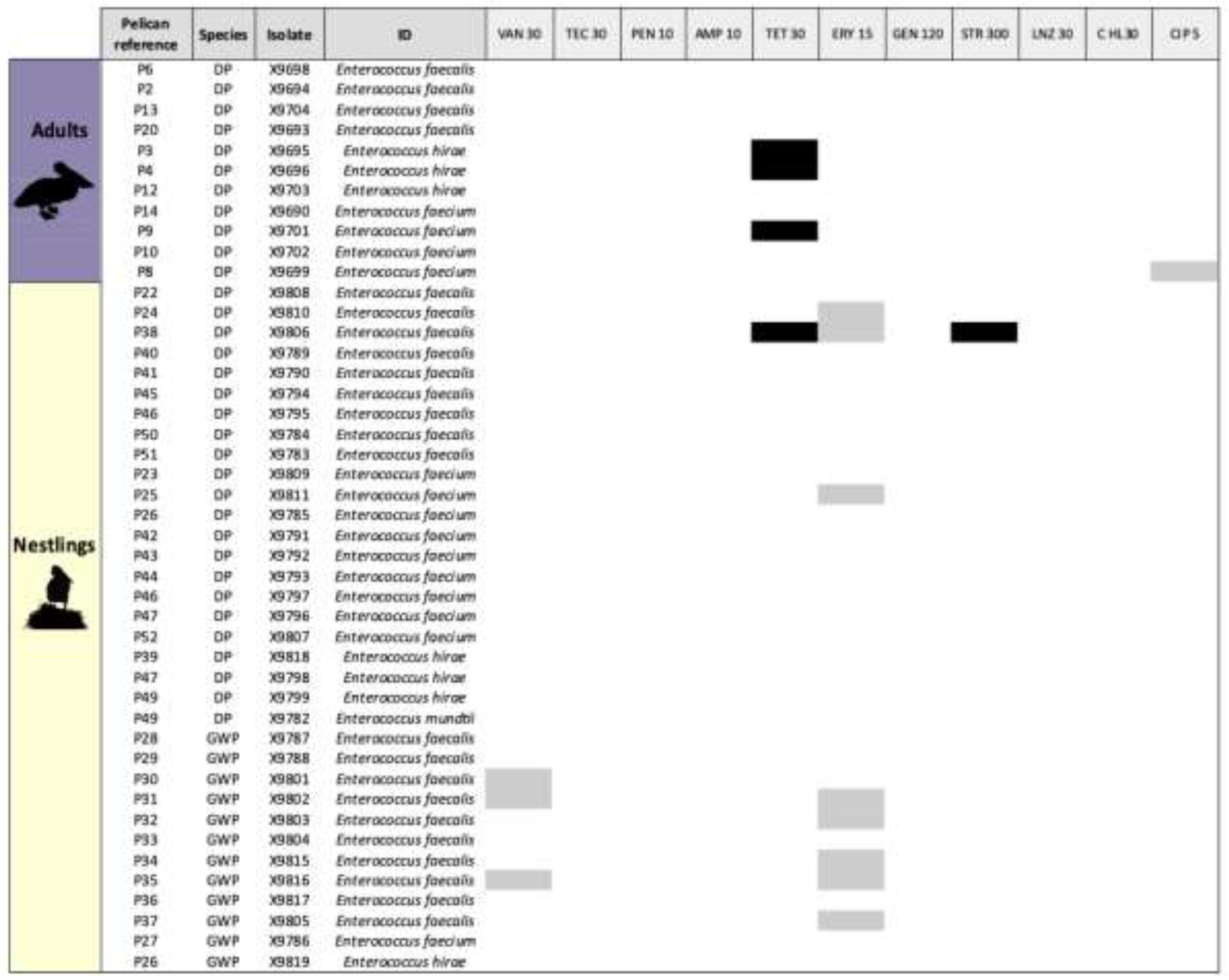
Figure 3.
Frequency of antimicrobial resistance exhibited by E. coli isolated in MAC (left) and enterococci isolated in S-B (right). Antibiotics with no resistance detected among the isolates are excluded.
Figure 3.
Frequency of antimicrobial resistance exhibited by E. coli isolated in MAC (left) and enterococci isolated in S-B (right). Antibiotics with no resistance detected among the isolates are excluded.
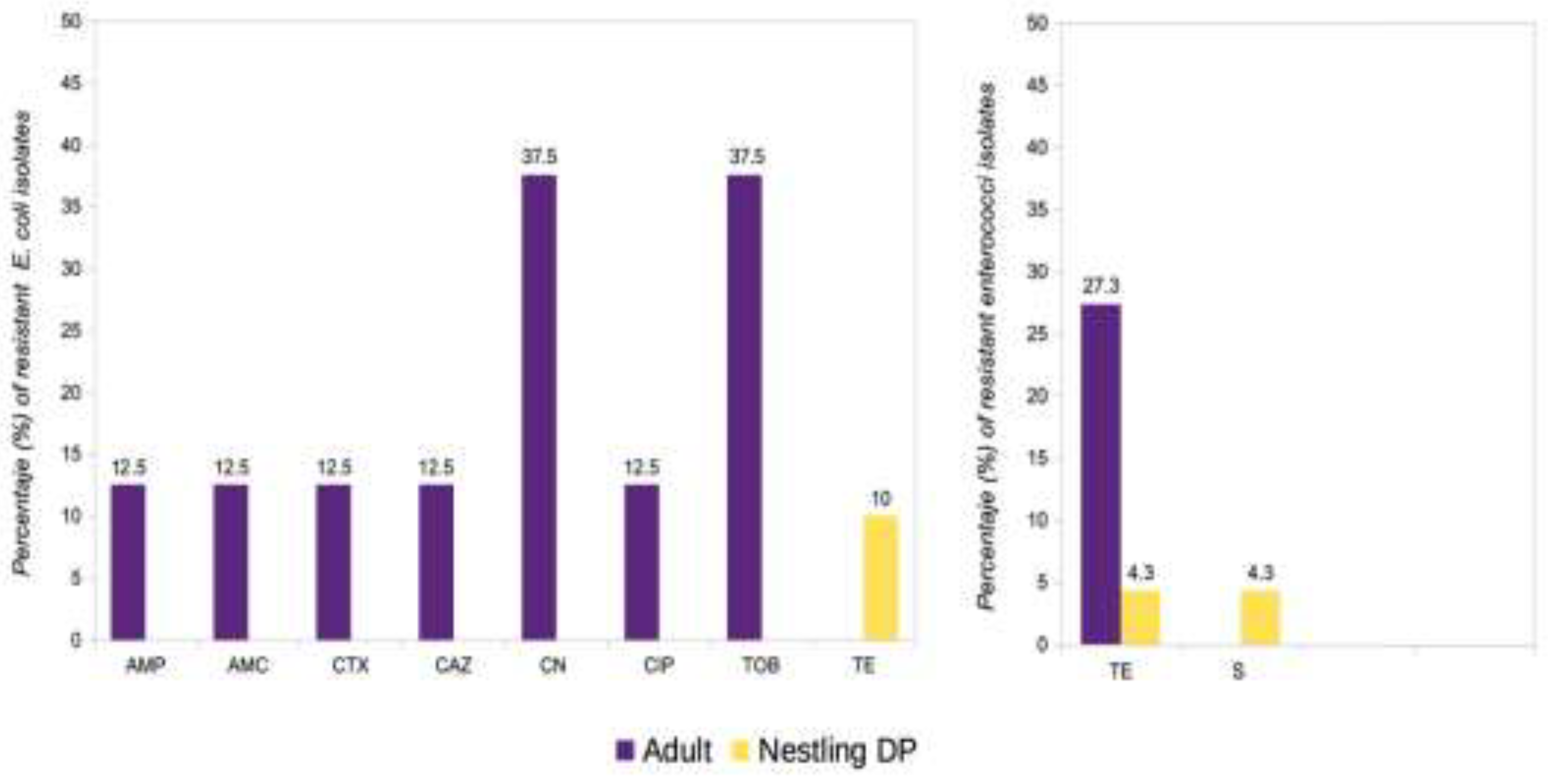
Figure 4.
Map showing sampling sites, and movements of GPS tagged DP from Greek eastern DP population (n=53; 2012-2024). Tracking data provided by Society for the Protection of Prespa. Available online: https://www.movebank.org/cms/movebank-main
Figure 4.
Map showing sampling sites, and movements of GPS tagged DP from Greek eastern DP population (n=53; 2012-2024). Tracking data provided by Society for the Protection of Prespa. Available online: https://www.movebank.org/cms/movebank-main
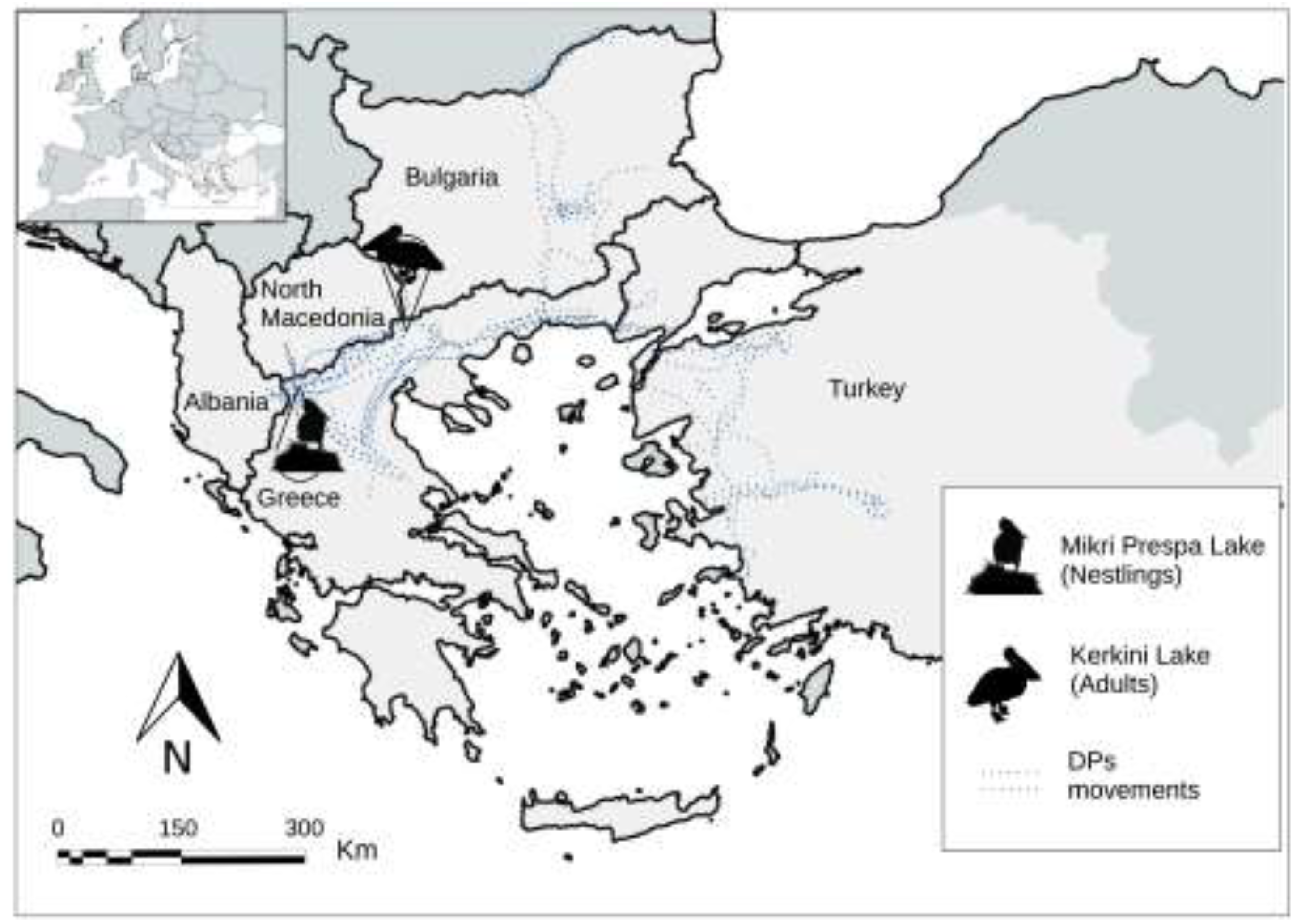

Table 1.
Occurrence of E. coli and Enterococcus isolated in non-selective media (MAC and S-B, respectively) in cloacal samples from adults and nestlings.
Table 1.
Occurrence of E. coli and Enterococcus isolated in non-selective media (MAC and S-B, respectively) in cloacal samples from adults and nestlings.
| Isolate | Number/Percentage of positive samples (n.º of non-repetitive isolates1) | |||
| Adults n=21 |
Nestlings n=31 |
Nestling DP n=20 |
Nestling GWP n=11 |
|
| Escherichia coli | 6/28.6 (8) | 20/64.5 (20) | 10/50 (10) | 10/90.9 (10)* |
| Enterococcus faecalis | 4/19 (4) | 19/61.3 (19)* | 9/45 (9) | 10/90.9 (10)* |
| Enterococcus faecium | 4/19 (4) | 10/32.2 (10) | 9/45 (9)* | 1/9.1 (1) |
| Enterococcus hirae | 3/14.3 (3) | 4/12.9 (4) | 4/20 (4) | - |
| Enterococcus mundtii | - | 1/ 3.2 (1) | 1/5 (1) | - |
| Total Enterococcus | 11/52.4 (11) | 30/96.8 (34) | 19/95 (23) | 11/100 (11) |
| Total non-repetitive isolates | 19 | 54 | 33 | 21 |
1One isolate of each species and sample or more than one if they presented different resistance phenotype. Significant differences between age groups are marked in red. Significant differences between nestling species are marked in bold. * p < 0.05
Table 2.
Isolates with resistance phenotypes obtained from non-selective media, with detection of resistance genes.
Table 2.
Isolates with resistance phenotypes obtained from non-selective media, with detection of resistance genes.
| Isolate | ID | Pelican reference | Age group |
Species | Resistance phenotype | Resistance genes |
|---|---|---|---|---|---|---|
| X9689 | E. coli | P12 | A | DP | CTX-AMC-CAZ-GEN-AMP-CIP-TOB | blaCTX-M15 |
| X9688 | E. coli | P12 | A | DP | GEN-TOB | |
| X9686 | E. coli | P17 | A | DP | GEN-TOB | |
| X9774 | E. coli | P41 | N | DP | TET | tet(A) |
| X9695 | E. hirae | P3 | A | DP | TET | tet(M) |
| X9696 | E. hirae | P4 | A | DP | TET | tet(M), tet(L) |
| X9701 | E. faecium | P9 | A | DP | TET | tet(M), tet(L) |
| X9806 | E. faecalis | P38 | N | DP | STR-TET | tet(M) |
Legend: A, adult; N, nestling; DP, Dalmatian pelican; GEN, gentamicin; TOB, tobramycin; TET, tetracycline; STR, streptomycin.
Table 3.
Characteristics of ESBL producing E. coli isolates recovered in cloacal swabs of adult DP.
| E. coli isolate | Pelican reference | Resistance phenotype | ESBL resistance genes | ST |
|---|---|---|---|---|
| X9682 | P11 | CTX-AMC-CAZ-GEN-AMP-CIP-TOB | CTX-M15 | 69 |
| X9684* | P12 | CTX-AMC-CAZ-GEN-AMP-CIP-TOB | CTX-M15 | 69 |
| X9706 | P15 | CTX-AMC-CAZ-GEN-AMP-SXT-CIP-TOB | CTX-M15 | 69 |
*Isolated in both MAC and chromogenic media. Legend: CTX, cefotaxime; AMC, amoxicillin/clavulanate; CAZ, ceftazidime; GEN, gentamicin; AMP, ampicillin; SXT, trimethoprim/sulfamethoxazole; CIP, ciprofloxacin and TOB, tobramycin.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



