
Submitted:
18 November 2024
Posted:
20 November 2024
You are already at the latest version
Abstract
Viruses in the Circoviridae family can infect both mammals and birds. Porcine circovirus type 2 (PCV2) significantly affects the livestock industry by causing porcine circovirus-associated diseases, such as postweaning multisystem wasting syndrome, respiratory disease complex, and dermatitis nephropathy syndrome. Additionally, beak and feather disease virus in parrots, canine circovirus in dogs, and columbid circovirus (pigeon circovirus) in racing pigeons induce immunosuppression, followed by secondary infections in these hosts. Although the PCV2 capsid protein has been demonstrated to inhibit type I interferon (IFN) signaling, the molecular mechanisms of Circoviridae-induced immunosuppression are largely unknown. In this study, we examined whether these functions are conserved across Circoviridae capsid proteins. Our results illustrated that although the nuclear localization of capsid proteins is conserved, their effects on IFN-β signaling vary by species, revealing the diverse roles of Circoviridae capsid proteins in modulating immune responses.
Keywords:
Circoviridae
; Capsid proteins
; Interferon β signaling
1. Introduction
The Circoviridae family consists of small icosahedral viruses without an envelope protein, and they feature a circular single-stranded DNA genome of 1600–2200 nucleotides in length. The Circoviridae family consists of two genera, namely Circovirus and Cyclovirus (Virus Taxonomy: 2023 Release; https://ictv.global/taxonomy, accessed on 13 November 2024). Several viruses in the Circoviridae family cause disease in mammals and birds, and in particular, porcine circovirus type 2 (PCV2) has significantly impacted the livestock industry. PCV2 causes several conditions collectively termed porcine circovirus-associated diseases (PCVADs), including postweaning multisystemic wasting syndrome, porcine respiratory disease complex, and porcine dermatitis nephropathy syndrome [1,2,3,4]. By contrast, porcine circovirus type 1 (PCV1), discovered in the porcine kidney-derived PK-15 cell line approximately 50 years ago, is considered nonpathogenic [1]. PCV3 was also identified in the US in 2015 in a case of porcine dermatitis and nephropathy syndrome (PDNS) [5,6], followed by the detection of PCV4 in pigs with signs of PDNS in China in 2019 [7]. Thus, porcine circoviruses are of high economic importance to the livestock industry.
Beak and feather disease virus (BFDV) is an important pathogen in parrots with psittacine beak and feather disease, and it has been present in Australia for at least 10 million years [8]. This virus significantly hampers conservation efforts for endangered parrots, causing feather damage, abnormal beak formation, and immunodeficiency [9]. Canine circovirus (CanineCV) causes severe symptoms in the presence of other pathogens, such as co-infection of CanineCV and canine parvovirus type 2 in puppies [10]. In addition, columbid circovirus (pigeon circovirus: PiCV) poses the greatest health and economic damage in young racing pigeons, inducing young pigeon disease syndrome with a secondary infection attributable to immunosuppression [11]. Therefore, immunosuppression is a common pathogenic feature of circovirus infections.
However, the molecular mechanism of immunosuppression following circovirus infection is largely unknown. Previous studies indicated that the PCV2 capsid protein inhibits type I interferon (IFN) signaling [12]. Specifically, it suppresses IFN-β promoter activity driven by stimulator of interferon genes, TANK-binding kinase 1, and interferon regulatory factor 3 (IRF3) [13], and it reduces phosphorylated IRF3 protein levels in the nucleus [14,15,16]. Supporting the mechanism of action, PCV2 capsid protein localizes in the nucleus [17,18]. However, whether these phenotypes are PCV2-specific remains unclear.
In this study, we investigated whether these functions are conserved across Circoviridae family capsid proteins. Our results demonstrated that although nuclear localization of the capsid protein is conserved among the Circoviridae family, the effect on IFN-β signaling differs among the virus species. These findings highlight the complex and divergent effects of Circoviridae family capsid proteins on IFN-β signaling.
2. Materials and Methods
2.1. Plasmids
The IFN-Beta_pGL3 plasmid was a gift from Nicolas Manel (plasmid #102597; http://n2t.net/addgene:102597; accessed on 13 November 2024; RRID: Addgene_102597, Addgene, Watertown, MA, USA). pCAGGS–humanTRIF–Myc [19] and pCAGGS–pigTRIF–Myc [20], in which the pCAGGS vector (Addgene) encoded Myc–tagged human TRIF or pig TRIF, respectively, and the pRL–TK vector (Promega, Madison, WI, USA, Cat# E2241) were used to evaluate IFN-β signaling. The coding sequence of the Circoviridae capsid plasmids was synthesized according to the amino acid sequences deposited in GenBank with codon optimization to human cells (Integrated DNA Technologies, Inc., Coralville, IA, USA). The synthesized DNA sequence is summarized in Table S1. Synthesized DNA was cloned into the pCAGGS–DsRed–monomer plasmid [21], which was prelinearized with AgeI–HF (New England Biolabs (NEB), Ipswich, MA, USA, Cat# R3552L) and NheI–HF (NEB, Cat# R3131L) using an In-Fusion HD Cloning Kit (TaKaRa, Kusatsu, Japan, Cat# Z9633N). The plasmids were amplified using NEB 5–alpha F’ Iq competent Escherichia coli (NEB, Cat# C2992H) and extracted using the PureYield Plasmid Miniprep System (Promega, Cat# A1222). The sequences of all plasmids were verified using the SupreDye v3.1 Cycle Sequencing Kit (M&S TechnoSystems, Osaka, Japan, Cat# 063001) with the Spectrum Compact CE System (Promega).
To generate pCAGGS–DsRed–monomer plasmids expressing capsid proteins with deletions or chimeric capsid proteins between PCV2 and BFDV capsid proteins, each fragment was generated with PrimeSTAR® GXL DNA Polymerase (TaKaRa, Cat# R050A). The primers used for PCR amplification and the amino acid sequences are listed in Table S2 and Table S3, respectively. The amplified fragments were cloned into the pCAGGS–DsRed–monomer plasmid and the sequences of all plasmids were verified as previously described.
2.2. Cell Culture
Lenti-X 293T cells (TaKaRa, Cat# Z2180N) were cultured in Dulbecco’s modified Eagle’s medium (Nacalai Tesque, Kyoto, Japan, Cat# 08458-16) supplemented with 10% fetal bovine serum (Cytiva, Marlborough, MA, USA, Cat# SH30396) and 1× penicillin–streptomycin (Nacalai Tesque, Cat# 09367-34) at 37°C in a humidified incubator with 5% CO2.
2.3. Nuclear Localization of Circoviridae Capsid Proteins
Lenti-X 293T cells were seeded into a 24-well plate (FUJIFILM Wako Pure Chemical, Osaka, Japan, Cat# 630-28441) at 1.25 × 105 cells/well, cultured overnight, and transfected with 500 ng of pCAGGS–DsRed–monomer Circoviridae capsid plasmids using TransIT-293 Transfection Reagent (TaKaRa, Cat# V2700) in Opti-MEM (Thermo Fisher Scientific, Waltham, MA, USA, Cat# 31985062). NucBlue Live ReadyProbes Reagent (Hoechst 33342, Thermo Fisher Scientific, Cat# R37605) was used to probe the nucleus. The localization of Circoviridae capsid proteins was evaluated 48 h after transfection using the EVOS M7000 Imaging System (Thermo Fisher Scientific).
2.4. IFN-β Luciferase Reporter Assay
Lenti-X 293T cells were seeded in a 96-well plate (FUJIFILM Wako Pure Chemical, Cat# 63528511) at 3 × 104 cells/well, cultured overnight, and co-transfected with 50 ng of pCAGGS–DsRed–monomer Circoviridae capsid plasmids, 2.5 ng of IFN-Beta_pGL3 plasmid, 45 ng of pRL–TK plasmid, and 2.5 ng of pCAGGS–humanTRIF–Myc or pCAGGS–pigTRIF–Myc plasmid using TransIT-293 Transfection Reagent. Two days after transfection, the luciferase activity was measured using a Dual-Glo Luciferase assay (Promega, Cat# E2920) and a GloMax Explorer Multimode Microplate Reader to investigate the effect of Circoviridae capsid proteins on IFN-β signaling. Firefly luciferase activities were normalized according to Renilla luciferase activities. The percentage relative activities were calculated by comparing the normalized luciferase data of Circoviridae capsid protein plasmid-transfected cells and empty plasmid-transfected cells. The assays were repeated at least three times. The data are presented as the mean ± SD of one representative experiment.
2.5. Western Blotting
The cellular pellets of Lenti-X 293T cells were lysed with 2× Bolt LDS sample buffer (Thermo Fisher Scientific, Cat# B0008) containing 2% β-mercaptoethanol (Bio-Rad, Hercules, CA, USA, Cat# 1610710) and incubated at 70°C for 10 min. The expression of Circoviridae capsid proteins was assessed using SimpleWestern Abby (ProteinSimple, San Jose, CA, USA) with mouse monoclonal anti–DsRed–monomer antibody (clone OTI4c8, Thermo Fisher Scientific, Cat# TA180084) and Anti–Mouse Detection Module (ProteinSimple, Cat# DM-002). The amount of input protein was measured using the Total Protein Detection Module (ProteinSimple, Cat# DM-TP01). The predicted sizes of the DsRed–monomer Circoviridae capsid proteins were calculated according to the Protein Molecular Weight website (https://www.bioinformatics.org/sms/prot_mw.html, accessed on 13 November 2024).
2.6. Phylogenic Analysis of Circoviridae Capsid Proteins
A phylogenetic tree was constructed using 68 Circoviridae capsid protein sequences obtained from GenBank. The tree was created using the MUSCLE algorithm within MEGA X software [22]. We subsequently constructed a phylogenetic tree from the aligned amino acid sequences obtained from public databases. Evolutionary analysis was performed using the maximum likelihood and neighbor-joining methods and employing the Jones–Taylor–Thornton matrix-based model.
2.7. Calculation of Capsid Identity in the Circoviridae Family among Animal Species
The level of conservation among the capsid proteins within the Circoviridae family was examined using The Sequence Manipulation Suite (http://imed.med.ucm.es/Tools/SMS/ident_sim.html, accessed on 13 November 2024), as described in Table S4.
2.8. Statistical Analysis
Unless stated otherwise, the data are presented as the mean ± SD of six measurements from a single assay, reflecting the results from at least three independent experiments. Variations in the relative values between the circovirus capsid proteins and the plasmid encoding only DsRed–monomer were analyzed using one-way ANOVA, followed by Dunnett’s post hoc test for multiple comparisons. A statistically significant difference was indicated by p ≤0.05.
3. Results
3.1. The Divergent Effects of Five Circoviridae Capsid Proteins on IFN-β Signaling
Immunosuppression is a common pathology observed in Circoviridae infections, including PCVAD and PBFD [12,23]. However, the molecular basis of immunosuppression remains unknown. Previous reports indicated that the PCV2 capsid protein localizes to the nucleus and inhibits IFN signaling [12]. To determine whether this feature is conserved throughout the Circoviridae family, we first performed experiments using capsid proteins derived from five Circoviridae viruses, specifically PCV1, PCV2, PiCV, CanineCV, and BFDV. First, we tested the expression of DsRed–monomer–tagged Circoviridae capsid proteins in Lenti-X 293T cells by western blotting. The expected molecular weights of DsRed–monomer–tagged Circoviridae capsid proteins ranged 53.1–57.5 kDa according to the Protein Molecular Weight website, and the results of western blotting were consistent with the expected sizes (Figure 1a). Next, we tested the cellular localization of these capsid proteins. The PCV2 capsid protein localized to the nucleus, consistent with previous reports (Figure 1b) [17]. The other four capsid proteins also displayed nuclear localization (Figure 1b).
Next, we observed the effects of the five circoviruses on IFN-β signaling. Consistent with previous reports [15], PCV2 suppressed IFN-β signaling by up to 32.0% versus the control (DsRed–monomer, Figure 1c). Similarly, CanineCV, PiCV, and PCV1 inhibited IFN-β signaling by as much as 75.8%, 60.7%, and 23.5%, respectively. In sharp contrast, BFDV enhanced IFN-β signaling by up to 147% (Figure 1c).
3.2. Regions/motifs Determining the Phenotypes of the BFDV and PCV2 Capsid Proteins
To determine the domains/motifs responsible for the opposing effects of BFDV and PCV2 on IFN-β signaling, we first created an alignment between BFDV and PCV2 (Figure 2a). Based on the alignment, we made mutant capsid proteins of BFDV and PCV2 (Figure 2b,c). First, we generated mutant capsid proteins with deletion of the N-terminal or C-terminal region (Figure 2b). Specifically, we generated plasmids expressing BFDV(del–50) and PCV2(del–50) proteins with deletion of 50 N-terminal amino acids because the N-terminus features a 41-amino acid nuclear localization signal (NLS) responsible for nuclear localization [12].
Because the location of the NLS is unknown in the BFDV capsid protein, we used the cNLS Mapper website (https://nls-mapper.iab.keio.ac.jp/cgi-bin/NLS_Mapper_form.cgi, accessed on 13 November 2024) to identify the possible NLS. We found that 27 amino acid sequences starting from amino acid 208 in the C-terminus could be a possible NLS (Figure S1). Based on this simulation, we generated plasmids expressing BFDV(del191–) and PCV2(del191–) proteins featuring deletion of the C-terminus after amino acid 190. the nuclear localization of the PCV2(del–50) protein was slightly weakened compared with that of the wild-type PCV2 (PCV2(WT)) protein (Figure 2d). Although no effect on nuclear localization was observed with BFDV(del–50) protein, we observed slightly weaker nuclear localization for BFDV(del191–) protein compared with that of the wild-type BFDV (BFDV(WT)) protein.
Next, we examined the effect of the five mutant capsid proteins on IFN-β signaling. We observed that both BFDV(del–50) and BFDV(del191–) proteins lost ability to enhance IFN-β signaling (Figure 2e). In the case of PCV2, although PCV2(del–50) lost its suppressing effect, PCV2(del191–) retained its suppressing effect (Figure 2f).
We further generated chimeric capsid proteins between BFDV and PCV2 (Figure 2c). Although both the PCV2(–100)/BFDV(101–) and BFDV(–100)/PCV2(101–) proteins localized to the nucleus (Figure 2d), the BFDV(–100)/PCV2(101–) protein exhibited an enhancing effect similar to that of the BFDV(WT) protein (Figure 2g).
Taken together, although both BFDV and PCV2 capsid proteins localized to the nucleus, their localization was dependent on different regions/motifs. Furthermore, these viruses utilized different regions/motifs to modulate IFN-β signaling.
3.3. Conserved Nuclear Localization and Divergent Effects on IFN-β Signaling of Circoviridae Capsid Proteins
We demonstrated that the BFDV capsid protein had a distinct effect on IFN-β signaling (Figure 1c). To elucidate whether this phenotype is specific for the BFDV capsid protein, we expanded the analysis to include additional Circoviridae capsid proteins. We constructed a phylogenetic tree of 68 Circoviridae capsid proteins and generated plasmids encoding 25 Circoviridae capsid proteins representing each cluster (Figure 3). Although PCV1 and PCV2 capsid proteins were grouped into the same cluster, PCV3 and PCV4 were grouped into different clusters. Interestingly, all bat-associated circoviruses (BatACV) were categorized into different clusters (Figure 3).
The nuclear localization of capsid proteins was conserved among the Circoviridae family (Figure 4a). In sharp contrast, we found a divergent effect on IFN-β signaling. Although capsid proteins from chimpanzee stool avian-like circovirus (ChimpACV), BatACV3, canary circovirus (CaCV), dipodfec virus UA04Rod_4537 (DipV_4537), mulard duck circovirus (MDuCV), and finch circovirus (FiCV) enhanced IFN-β signaling, IFN-β signaling as supposed by BatACV1, BatACV2, equine circovirus 1 (EquCV), and Culex circovirus-like virus (MosACV1) capsid proteins (Figure 4b). These observations suggested that the enhancing effect on IFN-β signaling was not BFDV capsid protein-specific.
3.4. Effect of Circoviridae Capsid Proteins on IFN-β Signaling Induced by the Pig-derived TRIF Protein
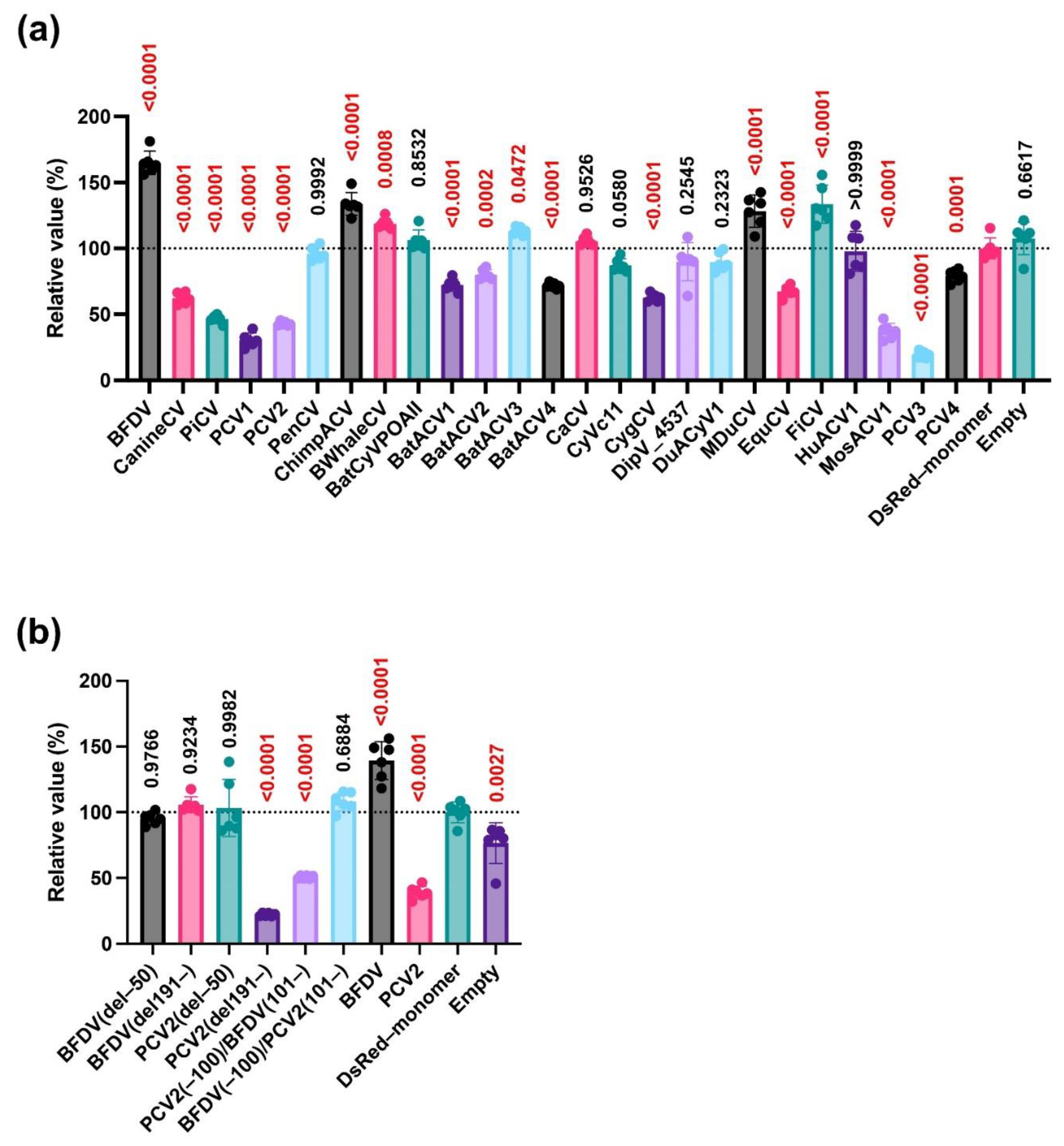
Because the aforementioned experiments were performed using human-derived TRIF protein (Figure 1c, Figure 2e–g, and Figure 4b), we repeated the experiments using pig-derived TRIF protein. We evaluated the effects of 25 Circoviridae capsid proteins, including mutant capsid proteins, in Lenti-X 293-T cells. The results were similar to those obtained with human-derived TRIF protein (Figure 5a). However, although the PCV2(–100)/BFDV(101–) protein did not affect IFN-β signaling induced by human-derived TRIF protein (Figure 2g), the PCV2(–100)/BFDV(101–) protein suppressed IFN-β signaling induced by pig-derived TRIF protein (Figure 5b). This result suggests that the effects of Circoviridae capsid proteins differ depending on the source of TRIF.
4. Discussion
Immunosuppression, a prevalent pathology in Circoviridae infections, was the focus of our study. We investigated the conservation of several functions of Circoviridae capsid proteins. Our findings revealed significant divergence in their effects of different virus species on IFN signaling. This discovery sheds new light on the complex interactions within the Circoviridae family.
Our research began with a comprehensive examination of the intracellular localization of five Circoviridae capsid proteins derived from PCV1, PCV2, CanineCV, PiCV, and BFDV. We found that all capsid proteins consistently localized to the nucleus (Figure 1b). This thorough analysis was expanded using capsid proteins from genetically distant viruses within the Circoviridae family (Figure 3). The results illustrated that all 31 Circoviridae capsid proteins localized to the nucleus (Figure 4a), further confirming the conserved nuclear localization of the capsid proteins.
The results using deletion mutants of the PCV2 capsid protein illustrated that nuclear localization was slightly weakened when the N-terminal 50 amino acids were deleted (Figure 2d). This result is consistent with a previous study revealing that the N-terminal 41 amino acids of the PCV2 capsid protein are associated with nuclear localization [17]. Contrarily, the BFDV(del191–) protein exhibited weaker nuclear localization (Figure 2d), suggesting that the C-terminal region is responsible for nuclear localization of the BFDV capsid protein. However, because the impact of the deletion in these regions was partial (Figure 2d), it is possible that other domains and/or motifs of the capsid proteins were also involved in their nuclear localization. Thus, it will be intriguing to test the impact of these regions on the localization of other Circoviridae capsid proteins. Further studies can address these points.
Next, we examined the effects of the Circoviridae capsid proteins on IFN-β signaling, specifically examining five Circoviridae capsid proteins derived from PCV1, PCV2, CanineCV, PiCV, and BFDV (Figure 1c). PCV1 and PCV2 capsid proteins suppressed IFN-β signaling. Although the phenotype of the PCV2 capsid protein was consistent with previous findings [16], the BFDV capsid protein enhanced IFN-β signaling.
Our analysis using deletion mutants of capsid proteins suggested that both the N- and C-termini of the BFDV capsid protein are required for its enhancing effect (Figure 2e). In the case of the PCV2 capsid protein, the C-terminus appeared dispensable for its inhibitory effects (Figure 2f). Further analysis using a chimeric protein between PCV2 and BFDV capsid proteins demonstrated that the BFDV(–100)/PCV2(101–) protein had an enhancing effect, albeit weaker than that of the BFDV(WT) protein (Figure 2g). This suggests that the effect of the BFDV capsid protein was dominant over that of the PCV2 capsid protein. Furthermore, these results suggest that whereas the PCV2 capsid protein does not require the C-terminal region to suppress IFN-β signaling, both the N- and C-termini are involved in the IFN-β signaling-stimulating effect of the BFDV capsid protein, and the C-terminal region of the PCV2 capsid protein can complement the activity of its C-terminal region.
Our experiment using 25 Circoviridae capsid proteins revealed that the effects of these proteins on IFN-β signaling varied greatly among the virus species (Figure 4b). Similarly as the PCV2 capsid protein (Figure 1c), several Circoviridae capsid proteins such as BatACV2, equine circovirus, Culex circovirus, and PCV3 capsid proteins suppressed IFN-β signaling (Figure 1c and Figure 4b). Contrarily, capsid proteins from chimpanzee circovirus, BatACV3, canary circovirus, dipodfec virus, and FiCV, especially the BFDV capsid protein, markedly enhanced IFN-β signaling (Figure 1c and Figure 4b).
Among PCV1, PCV2, PCV3, and PCV4, only the PCV4 capsid protein failed to suppress IFN-β signaling (Figure 4b). Given that the PCV3 capsid protein was more distinct from the PCV2 capsid protein than the PCV4 capsid protein in the phylogenetic tree (Figure 3), this result suggests that the function of IFN-β signaling is independent of the genetic similarity of Circoviridae capsid proteins. Similarly, in the case of the bat-derived circovirus capsid protein, whereas BatACV1 and BatACV2 capsid proteins suppressed IFN-β signaling (Figure 4b), an enhancing effect was observed for the BatACV3 capsid protein, and no effect was observed for the BatACV4 capsid protein. This finding supports our hypothesis that the effects of Circoviridae capsid proteins on IFN-β signaling are independent of genetic similarity.
When we focused on the Circoviridae capsid proteins that enhanced IFN-β signaling (Figure 4b), the chimpanzee circovirus, canary circovirus, BFDV, and FiCV capsid proteins were found to be genetically similar (Figure 3). However, PiCV, which is genetically similar to BFDV, oppositely suppressed IFN-β signaling (Figure 4b). These findings suggest that the effect of capsid proteins on IFN-β signaling is not associated with the genetic distance.
This study had several limitations. First, we were unable to identify the region(s) responsible for the divergent effects of Circoviridae capsid proteins on IFN-β signaling. Second, we did not elucidate the correlation between the suppressing effect of capsid proteins on IFN-β signaling and clinical severity. Because both PCV2 and BFDV infections induce immunosuppression, other viral proteins of BFDV might be responsible for its immunosuppressive effect. These points should be investigated in future studies. Third, we used a luciferase-based reporter system in human-derived Lenti-X 293-T cells to test the effect on IFN-β signaling. Although similar results were obtained using both human- and pig-derived TRIF proteins (Figure 4b and Figure 5a), we must use cell lines derived from other animals, including pigs and parrots, to clarify the significance of PCV2 and BFDV capsid proteins on IFN-β signaling.
In summary, our study revealed the diverse effects of Circoviridae capsid proteins on IFN-β signaling, finding that these effects were virus species-specific. This highlights the intricate pathophysiology induced by viruses in the Circoviridae family. These findings both deepen our understanding of these viruses and provide a foundation for future research to develop effective treatments for animals infected by Circoviridae viruses.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Possible NLS sequences in PCV2 and PBDFV capsid proteins; Table S1: Synthesized DNAs for generating plasmids encoding capsid protein; Table S2: Primers used for generating plasmids encoding mutant Circoviridae capsid proteins; Table S3: Amino acid sequence of mutant Circoviridae capsid proteins; Table S4: Genetic distance of Circoviridae capsid based on the amino acid sequence.
Author Contributions
Conceptualization, A.H.K., and A.S.; methodology, A.H.K., A.C., R.S.R., and A.S.; formal analysis, A.H.K., C.-Y.H., Z.-Y.L., H.-Z.H., A.C., R.S.R., K.-P.C., and A.S.; investigation, A.H.K., C.-Y.H., Z.-Y.L., H.-Z.H., A.C., and R.S.R.; writing – original draft, A.H.K.; writing – review & editing C.-Y.H., Z.-Y.L., H.-Z.H., A.C., R.S.R., K.-P.C., and A.S.; supervision, A.S.; funding acquisition, A.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the Japan Agency for Medical Research and Development (AMED) Research Program on HIV/AIDS JP24fk0410047, JP24fk0410056, and JP24fk0410058 (to A.S.); the AMED Research Program on Emerging and Re-emerging Infectious Diseases JP22fk0108511, JP22fk0108506, and JP23fk0108583 (to A.S.); the AMED the Research Project for Practical Applications of Regenerative Medicine JP24bk0104177 (to A.S.); the JSPS KAKENHI Grant-in-Aid for Scientific Research (C) JP24K09227 (to A.S.); the JSPS KAKENHI Grant-in-Aid for Scientific Research (B) JP22H02500 (to A.S.) and JP21H02361 (to A.S.); the JSPS Bilateral Program JPJSBP120245706 (to A.S.); the JSPS Fund for the Promotion of Joint International Research (International Leading Research) JP23K20041 (to A.S.); the G-7 Grant (to A.S.); and the Ito Foundation Research Grant R6 KEN119 (to A.S.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Acknowledgments
The authors thank Miki Kawano, Natsumi Matsubara, Tomoko Nishiuchi, and the staff of CADIC, University of Miyazaki, for their assistance.
Conflicts of Interest
The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Iizuka, N.; Kohara, M.; Hagino-Yamagishi, K.; Abe, S.; Komatsu, T.; Tago, K.; Arita, M.; Nomoto, A. Construction of Less Neurovirulent Polioviruses by Introducing Deletions into the 5’ Noncoding Sequence of the Genome. Journal of Virology 1989, 63, 5354–5363. [Google Scholar] [CrossRef] [PubMed]
- Nayar, G.P.; Hamel, A.; Lin, L. Detection and Characterization of Porcine Circovirus Associated with Postweaning Multisystemic Wasting Syndrome in Pigs. Can Vet J 1997, 38, 385–386. [Google Scholar] [PubMed]
- Allan, G.M.; McNeilly, F.; Kennedy, S.; Daft, B.; Clarke, E.G.; Ellis, J.A.; Haines, D.M.; Meehan, B.M.; Adair, B.M. Isolation of Porcine Circovirus-like Viruses from Pigs with a Wasting Disease in the USA and Europe. J Vet Diagn Invest 1998, 10, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.; Hassard, L.; Clark, E.; Harding, J.; Allan, G.; Willson, P.; Strokappe, J.; Martin, K.; McNeilly, F.; Meehan, B.; et al. Isolation of Circovirus from Lesions of Pigs with Postweaning Multisystemic Wasting Syndrome. Can Vet J 1998, 39, 44–51. [Google Scholar] [PubMed]
- Palinski, R.; Piñeyro, P.; Shang, P.; Yuan, F.; Guo, R.; Fang, Y.; Byers, E.; Hause, B.M. A Novel Porcine Circovirus Distantly Related to Known Circoviruses Is Associated with Porcine Dermatitis and Nephropathy Syndrome and Reproductive Failure. J Virol 2017, 91, e01879-16. [Google Scholar] [CrossRef]
- Klaumann, F.; Correa-Fiz, F.; Franzo, G.; Sibila, M.; Núñez, J.I.; Segalés, J. Current Knowledge on Porcine Circovirus 3 (PCV-3): A Novel Virus With a Yet Unknown Impact on the Swine Industry. Frontiers in Veterinary Science 2018, 5, 315. [Google Scholar] [CrossRef]
- Zhang, P.; Shen, H.; Liu, X.; Wang, S.; Liu, Y.; Xu, Z.; Song, C. Porcine Circovirus Type 3 Cap Inhibits Type I Interferon Induction Through Interaction With G3BP1. Front. Vet. Sci. 2020, 7, 594438. [Google Scholar] [CrossRef]
- Martens, J.M.; Stokes, H.S.; Berg, M.L.; Walder, K.; Bennett, A.T.D. Seasonal Fluctuation of Beak and Feather Disease Virus (BFDV) Infection in Wild Crimson Rosellas (Platycercus Elegans). Sci Rep 2020, 10, 7894. [Google Scholar] [CrossRef]
- Raidal, S.R.; Peters, A. Psittacine Beak and Feather Disease: Ecology and Implications for Conservation. Emu - Austral Ornithology 2018, 118, 80–93. [Google Scholar] [CrossRef]
- Hao, X.; Li, Y.; Chen, H.; Chen, B.; Liu, R.; Wu, Y.; Xiao, X.; Zhou, P.; Li, S. Canine Circovirus Suppresses the Type I Interferon Response and Protein Expression but Promotes CPV-2 Replication. IJMS 2022, 23, 6382. [Google Scholar] [CrossRef]
- Raue, R.; Schmidt, V.; Freick, M.; Reinhardt, B.; Johne, R.; Kamphausen, L.; Kaleta, E.F.; Müller, H.; Krautwald-Junghanns, M.-E. A Disease Complex Associated with Pigeon Circovirus Infection, Young Pigeon Disease Syndrome. Avian Pathology 2005, 34, 418–425. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Chen, J.; Zhang, Q.; Huang, K.; Ma, D.; Du, Q.; Tong, D.; Huang, Y. Porcine Circovirus Type 2 Infection Inhibits the Activation of Type I Interferon Signaling via Capsid Protein and Host gC1qR. Veterinary Microbiology 2022, 266, 109354. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Wang, Z.; Qiao, D.; Yuan, Y.; Han, C.; Yang, N.; Li, R.; Du, Q.; Tong, D.; Huang, Y. Porcine Circovirus Type 2 Infection Attenuates the K63-Linked Ubiquitination of STING to Inhibit IFN-β Induction via P38-MAPK Pathway. Veterinary Microbiology 2021, 258, 109098. [Google Scholar] [CrossRef]
- Liu, S.; Cai, X.; Wu, J.; Cong, Q.; Chen, X.; Li, T.; Du, F.; Ren, J.; Wu, Y.-T.; Grishin, N.V.; et al. Phosphorylation of Innate Immune Adaptor Proteins MAVS, STING, and TRIF Induces IRF3 Activation. Science 2015, 347, aaa2630. [Google Scholar] [CrossRef]
- Li, J.; Lu, M.; Huang, B.; Lv, Y. Porcine Circovirus Type 2 Inhibits Interferon-β Expression by Targeting Karyopherin Alpha-3 in PK-15 Cells. Virology 2018, 520, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Chen, J.; Wu, X.; Ma, D.; Zhang, X.; Li, R.; Han, C.; Liu, H.; Yin, X.; Du, Q.; et al. PCV2 Targets cGAS to Inhibit Type I Interferon Induction to Promote Other DNA Virus Infection. PLoS Pathog 2021, 17, e1009940. [Google Scholar] [CrossRef]
- Liu, Q.; Tikoo, S.K.; Babiuk, L.A. Nuclear Localization of the ORF2 Protein Encoded by Porcine Circovirus Type 2. Virology 2001, 285, 91–99. [Google Scholar] [CrossRef]
- Hou, Q.; Hou, S.; Chen, Q.; Jia, H.; Xin, T.; Jiang, Y.; Guo, X.; Zhu, H. Nuclear Localization Signal Regulates Porcine Circovirus Type 2 Capsid Protein Nuclear Export through Phosphorylation. Virus Research 2018, 246, 12–22. [Google Scholar] [CrossRef]
- Choonnasard, A.; Shofa, M.; Okabayashi, T.; Saito, A. Conserved Functions of Orthohepadnavirus X Proteins to Inhibit Type-I Interferon Signaling. Int J Mol Sci 2024, 25, 3753. [Google Scholar] [CrossRef]
- Ringo, R.S.; Choonnasard, A.; Okabayashi, T.; Saito, A. Conserved Antagonization of Type I Interferon Signaling by Arterivirus GP5 Proteins. Viruses 2024, 16, 1240. [Google Scholar] [CrossRef]
- Niwa, H.; Yamamura, K.; Miyazaki, J. Efficient Selection for High-Expression Transfectants with a Novel Eukaryotic Vector. Gene 1991, 108, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol Biol Evol 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Katoh, H.; Ogawa, H.; Ohya, K.; Fukushi, H. A Review of DNA Viral Infections in Psittacine Birds. J Vet Med Sci 2010, 72, 1099–1106. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Divergent effects of five Circoviridae capsid proteins on human TRIF-mediated IFN-β signaling. (a) Expression of five Circoviridae capsid proteins in Lenti-X 293-T cells. The expected DsRed–monomer–tagged Circoviridae capsid proteins had expected sizes of 53.07–57.54 kDa according to the Protein Molecular Weight website (https://www.bioinformatics.org/sms/prot_mw.html, accessed on 13 November 2024). (b) The cellular localization of the five DsRed–monomer–tagged Circoviridae capsid proteins. (c) Effects of the five Circoviridae capsid proteins on human TRIF-mediated IFN-β signaling as determined using the IFN-β luciferase reporter assay. Differences between cells transfected with plasmids expressing Circoviridae capsid proteins and DsRed–monomer plasmids were examined by one-way ANOVA followed by Dunnett’s multiple comparison test.
Figure 1.
Divergent effects of five Circoviridae capsid proteins on human TRIF-mediated IFN-β signaling. (a) Expression of five Circoviridae capsid proteins in Lenti-X 293-T cells. The expected DsRed–monomer–tagged Circoviridae capsid proteins had expected sizes of 53.07–57.54 kDa according to the Protein Molecular Weight website (https://www.bioinformatics.org/sms/prot_mw.html, accessed on 13 November 2024). (b) The cellular localization of the five DsRed–monomer–tagged Circoviridae capsid proteins. (c) Effects of the five Circoviridae capsid proteins on human TRIF-mediated IFN-β signaling as determined using the IFN-β luciferase reporter assay. Differences between cells transfected with plasmids expressing Circoviridae capsid proteins and DsRed–monomer plasmids were examined by one-way ANOVA followed by Dunnett’s multiple comparison test.
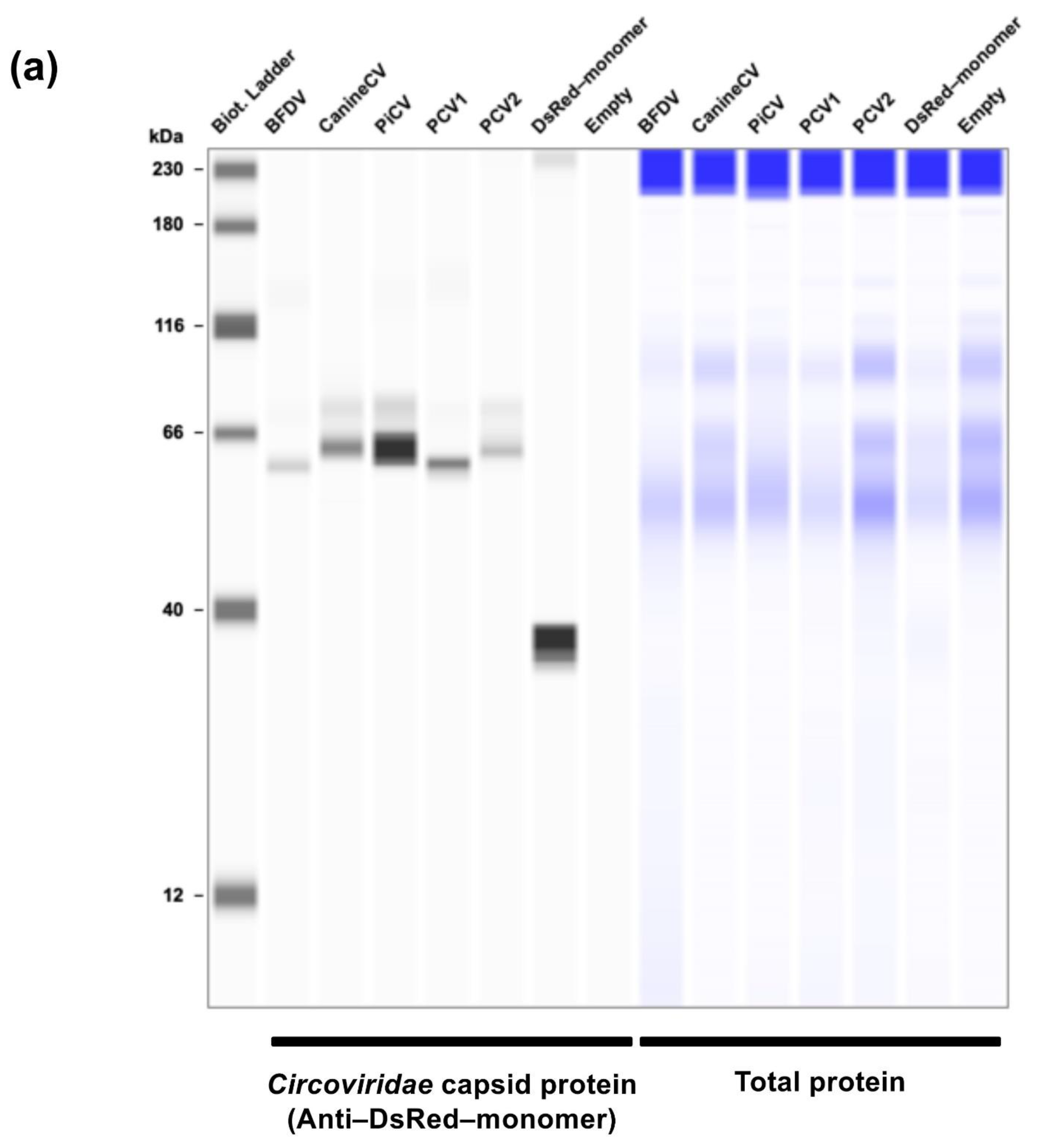
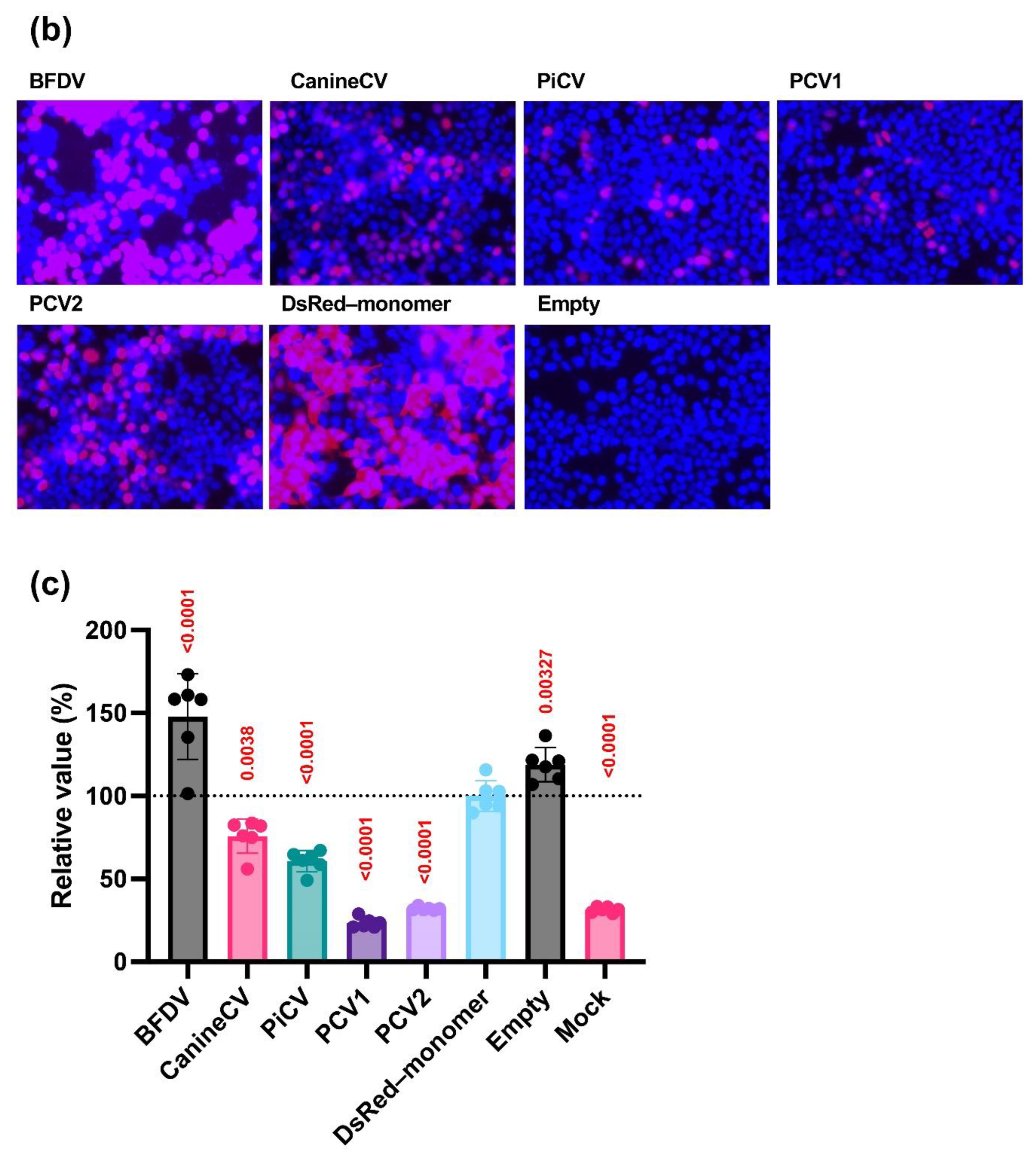
Figure 2.
The phenotypes of deletion mutants of PCV2 and BFDV capsid proteins and chimeric proteins between PCV2 and BFDV capsid proteins. (a) Amino acid alignment of PCV2 and BFDV capsid proteins. (b) Schematic representation of the deletion mutants of PCV2 and BFDV capsid proteins. (c) Schematic representation of chimeric proteins between PCV2 and BFDV capsid proteins. (d) The cellular localization of the DsRed–monomer–tagged capsid proteins. (e–g) Effects of capsid proteins on human TRIF-mediated IFN-β signaling. Relative value of the IFN-β luciferase reporter assay. Differences between cells transfected with plasmids expressing capsid proteins or DsRed–monomer plasmids were examined by one-way ANOVA followed by Dunnett’s multiple comparison test.
Figure 2.
The phenotypes of deletion mutants of PCV2 and BFDV capsid proteins and chimeric proteins between PCV2 and BFDV capsid proteins. (a) Amino acid alignment of PCV2 and BFDV capsid proteins. (b) Schematic representation of the deletion mutants of PCV2 and BFDV capsid proteins. (c) Schematic representation of chimeric proteins between PCV2 and BFDV capsid proteins. (d) The cellular localization of the DsRed–monomer–tagged capsid proteins. (e–g) Effects of capsid proteins on human TRIF-mediated IFN-β signaling. Relative value of the IFN-β luciferase reporter assay. Differences between cells transfected with plasmids expressing capsid proteins or DsRed–monomer plasmids were examined by one-way ANOVA followed by Dunnett’s multiple comparison test.
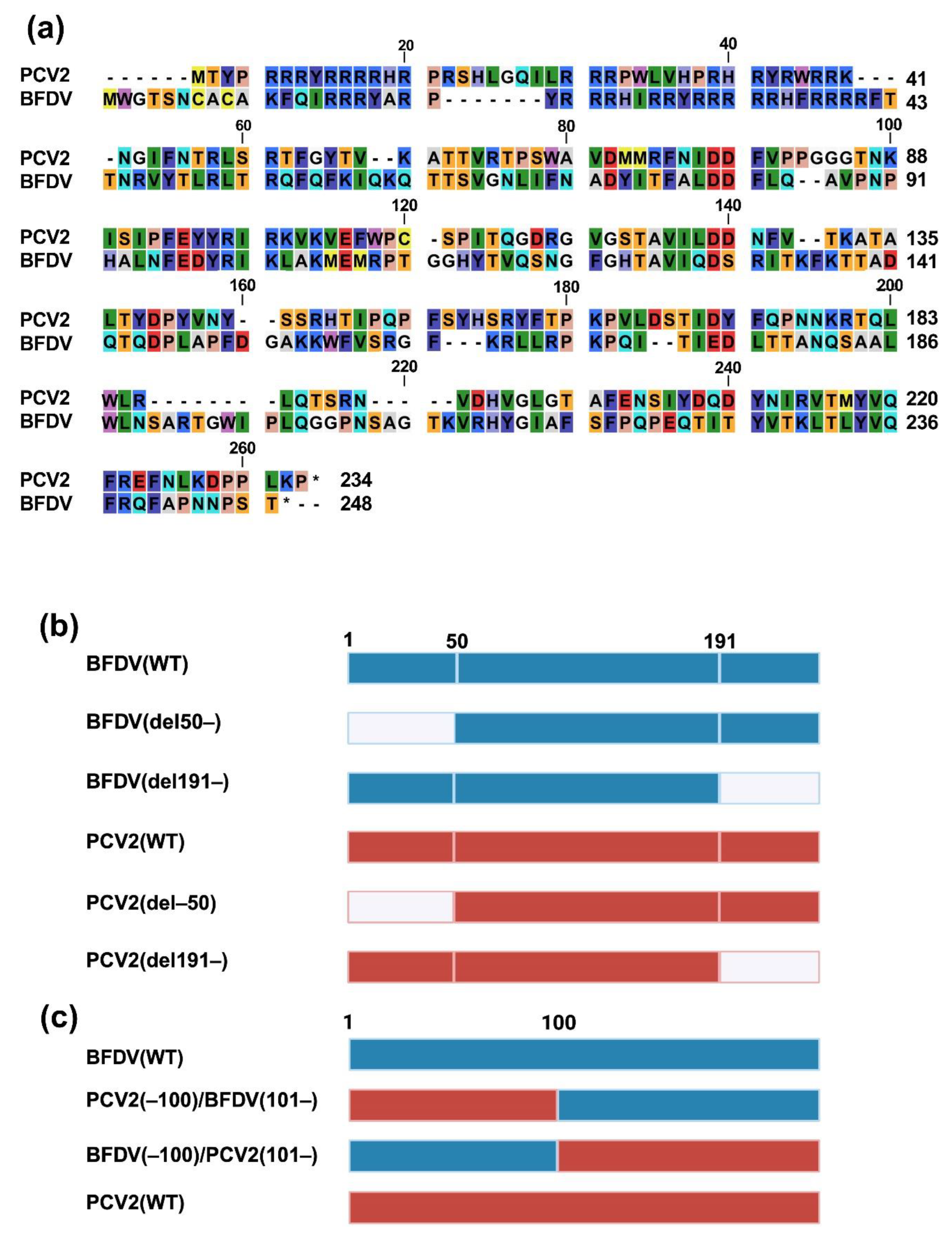
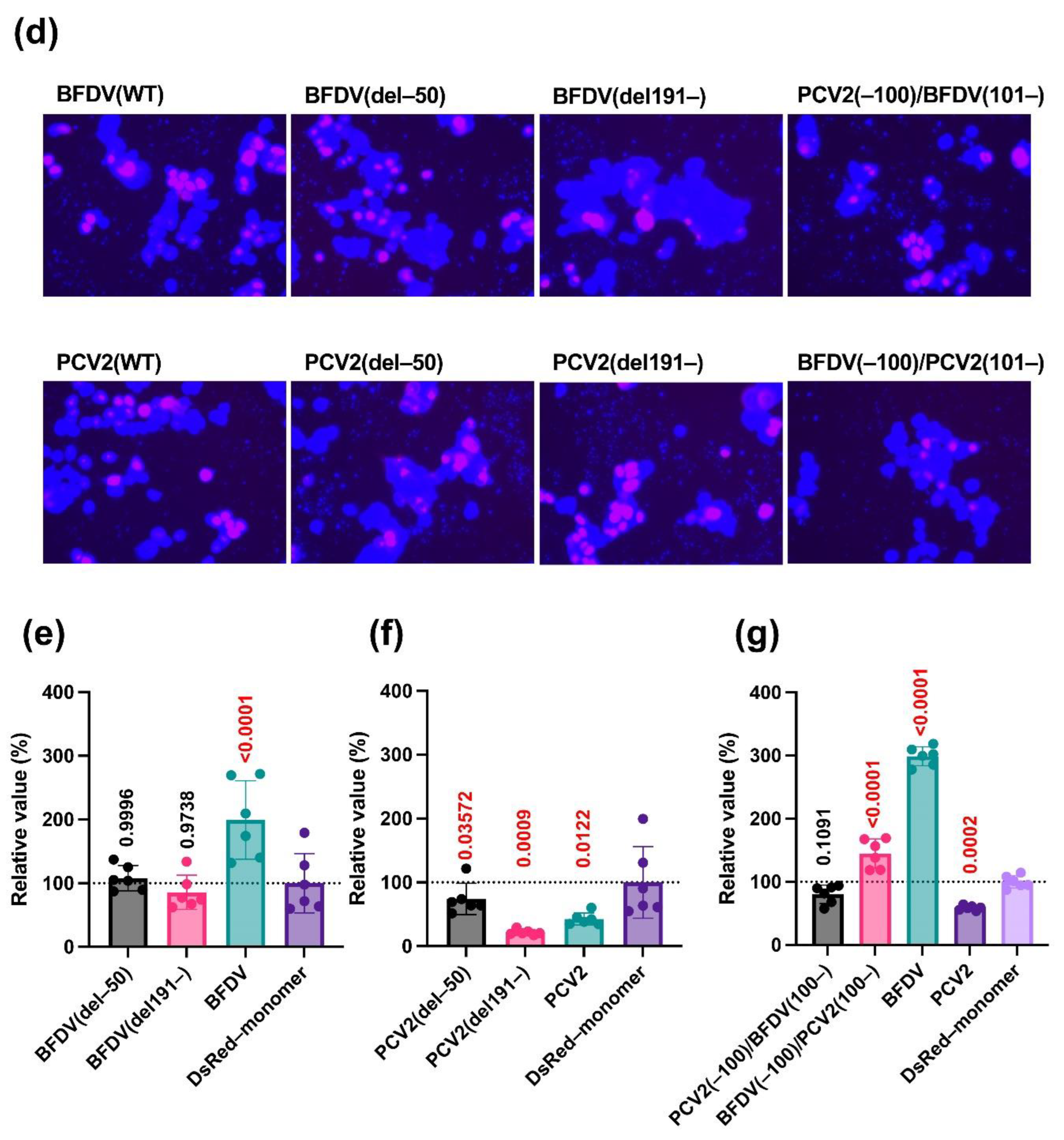
Figure 3.
Phylogenetic tree of Circoviridae capsid proteins. The phylogenetic tree was constructed using MEGA software [22], and evolutionary analysis was conducted using the maximum likelihood and neighbor-joining methods based on the Jones–Taylor–Thornton matrix-based model with 1000 bootstrap replicates. In the phylogenetic tree, the capsid proteins characterized in this experiment are marked with red circles.
Figure 3.
Phylogenetic tree of Circoviridae capsid proteins. The phylogenetic tree was constructed using MEGA software [22], and evolutionary analysis was conducted using the maximum likelihood and neighbor-joining methods based on the Jones–Taylor–Thornton matrix-based model with 1000 bootstrap replicates. In the phylogenetic tree, the capsid proteins characterized in this experiment are marked with red circles.
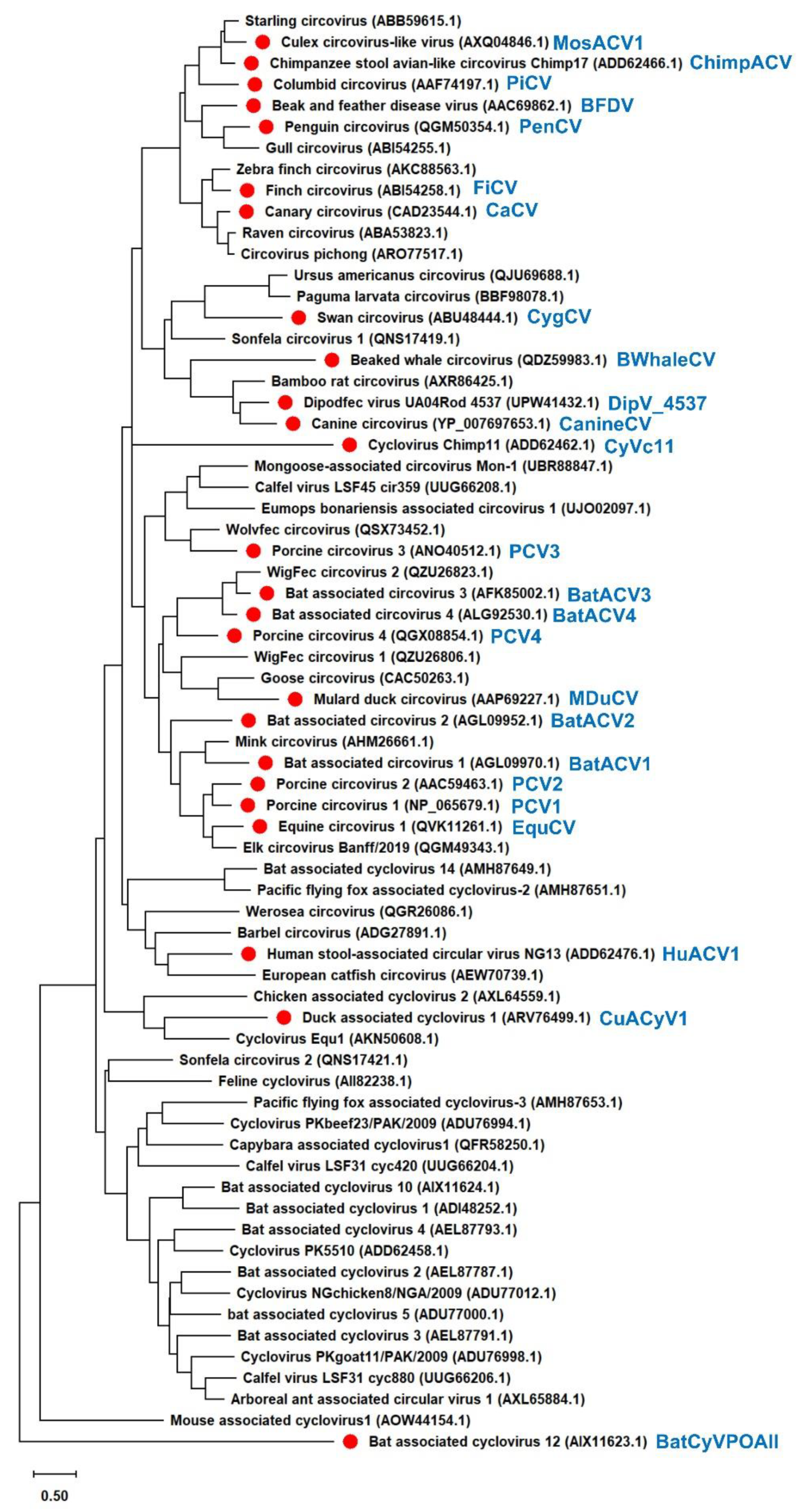
Figure 4.
Effects of Circoviridae capsid proteins on human TRIF-mediated IFN-β signaling. (a) The cellular localization of DsRed–monomer–tagged Circoviridae capsid proteins. (b) Effects of Circoviridae capsid proteins on human TRIF-mediated IFN-β signaling. The relative values determined by the IFN-β luciferase reporter assay are presented. Differences between cells transfected with plasmids expressing Circoviridae capsid proteins or DsRed–monomer plasmids were examined by one-way ANOVA followed by Dunnett’s multiple comparison test.
Figure 4.
Effects of Circoviridae capsid proteins on human TRIF-mediated IFN-β signaling. (a) The cellular localization of DsRed–monomer–tagged Circoviridae capsid proteins. (b) Effects of Circoviridae capsid proteins on human TRIF-mediated IFN-β signaling. The relative values determined by the IFN-β luciferase reporter assay are presented. Differences between cells transfected with plasmids expressing Circoviridae capsid proteins or DsRed–monomer plasmids were examined by one-way ANOVA followed by Dunnett’s multiple comparison test.
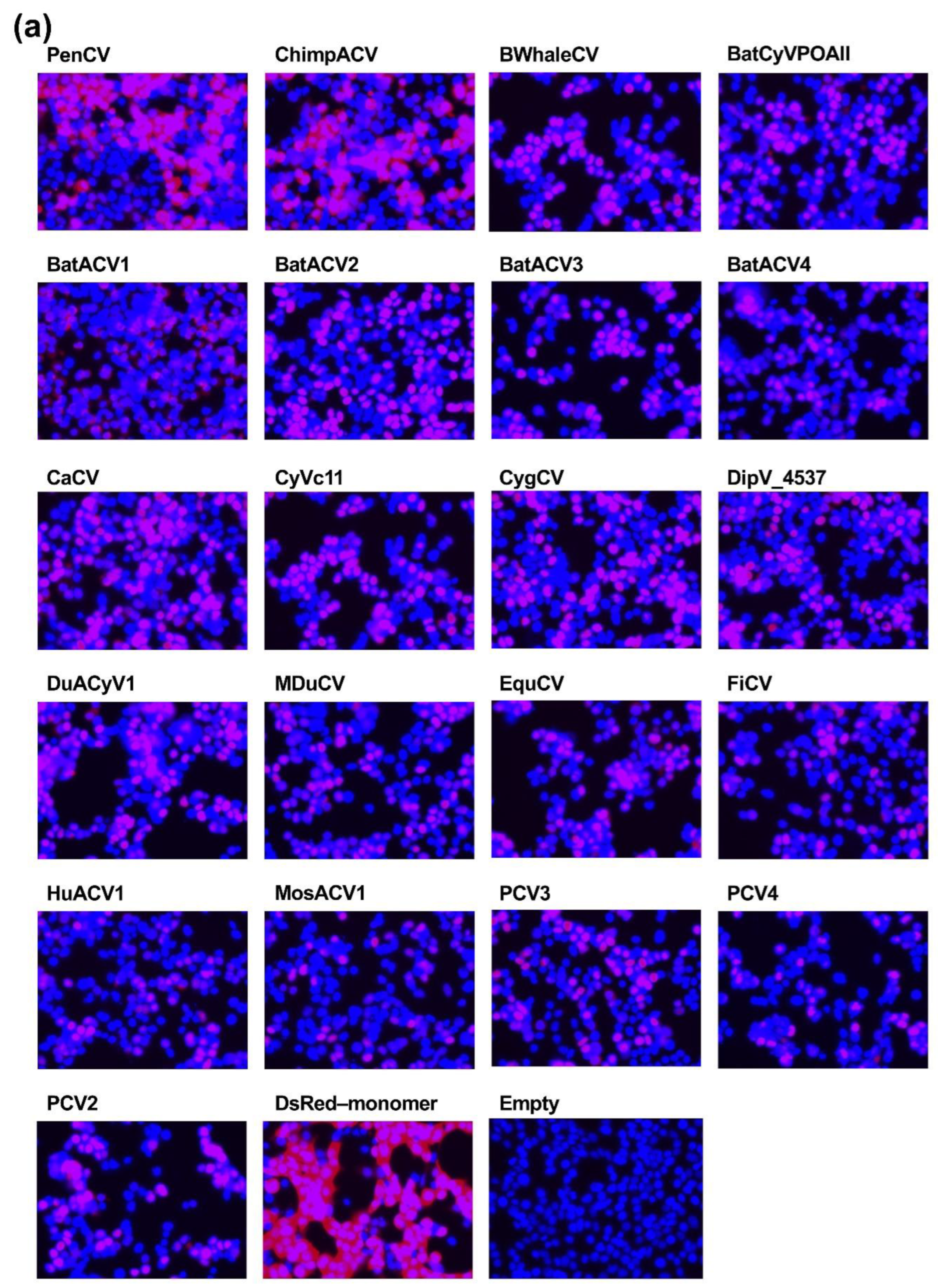
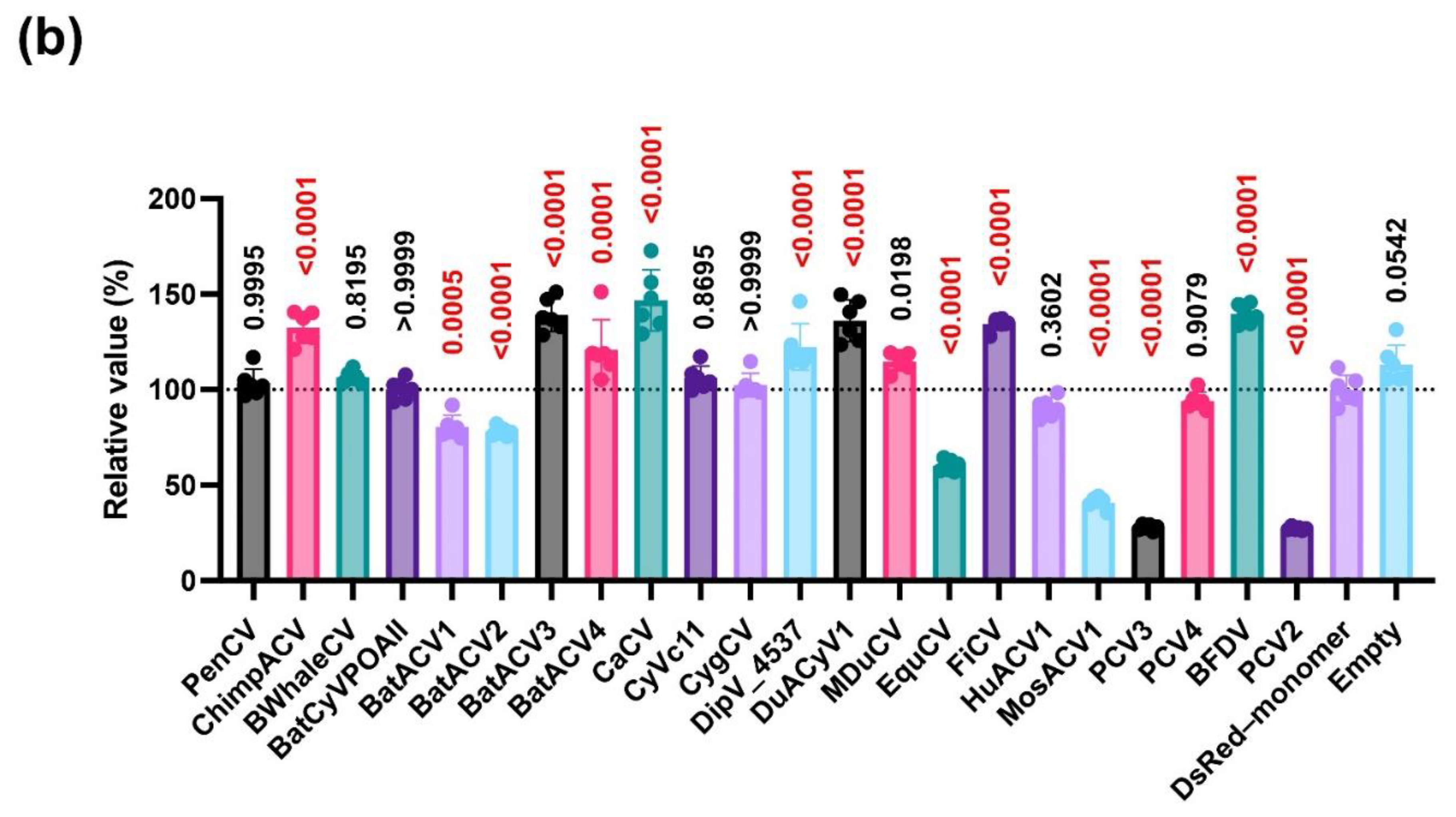
Figure 5.
Effects of Circoviridae capsid proteins on pig TRIF-mediated IFN-β signaling. (a) Comparisons of the effects of capsid proteins from different Circoviridae species on pig TRIF-mediated IFN-β signaling. (b) Comparisons of the effects of deletion/chimeric constructs of Circoviridae capsid proteins on pig TRIF-mediated IFN-β signaling. The relative values determined by the IFN-β luciferase reporter assay are presented. Differences between cells transfected with plasmids expressing Circoviridae capsid proteins or DsRed–monomer plasmids were examined by one-way ANOVA followed by Dunnett’s multiple comparison test.
Figure 5.
Effects of Circoviridae capsid proteins on pig TRIF-mediated IFN-β signaling. (a) Comparisons of the effects of capsid proteins from different Circoviridae species on pig TRIF-mediated IFN-β signaling. (b) Comparisons of the effects of deletion/chimeric constructs of Circoviridae capsid proteins on pig TRIF-mediated IFN-β signaling. The relative values determined by the IFN-β luciferase reporter assay are presented. Differences between cells transfected with plasmids expressing Circoviridae capsid proteins or DsRed–monomer plasmids were examined by one-way ANOVA followed by Dunnett’s multiple comparison test.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



