
Submitted:
16 November 2024
Posted:
19 November 2024
You are already at the latest version
Abstract
The Xyloglucan endotransglucosylase/hydrolase (XTH) family, a group of cell wall-modifying enzymes, plays crucial roles in plant growth, development, and stress adaptation. The quality and yield of Chinese jujube (Ziziphus jujuba) fruit are significantly impacted by environmental stresses, including excessive salinity, drought, freezing, and disease. However, there has no report of the XTH encoding genes present in Chinese jujube genome and their response transcription level under various stresses. This study provides an in-depth analysis of ZjXTH genes in genome of Chinese jujube and elucidating their structural motifs, regulatory networks, and expression patterns under various stresses. A total of 29 ZjXTH genes were identified from Ziziphus jujuba genome. Phylogenetic analysis classifies ZjXTH genes into four distinct groups, while conserved motifs and domain analyses reveal coordinated xyloglucan modifications, highlighting key shared motifs and domains. Interaction network predictions suggest that ZjXTHs may interact with proteins such as Expansin-B1 (EXPB1) and Pectin Methylesterase 22 (PME22). Additionally, cis-regulatory element analysis enhances our understanding of Chinese jujube plant's defensive systems, where TCA- and TGACG-motifs process environmental cues and orchestrate stress responses. Expression profiling revealed differential regulation of ZjXTH genes under various stress conditions. Under salt stress, ZjXTH1 was upregulated, while ZjXTH6, and ZjXTH13 were downregulated, with ZjXTH3 and ZjXTH19 exhibiting negative regulation. In response to drought, ZjXTH5 showed upregulation, whereas ZjXTH7, -25, -13 were downregulated, and ZjXTH13 and ZjXTH19 were negatively regulated. Furthermore, under cold conditions, ZjXTH14 and ZjXTH18 were upregulated, while ZjXTH22, -23, -28, -29 were downregulated, and ZjXTH20 exhibited negative regulation. Additionally, phytoplasma infection induced the upregulation of ZjXTH1, -3, -5, with ZjXTH6, -7, -11, -16, -25 being downregulated, and ZjXTH20 showing negative regulation. Collectively, these findings deepen our understanding of the functional roles of Chinese jujube XTHs, emphasizing their regulatory function in adaptive responses in Chinese jujube plants.
Keywords:
Xyloglucan endotransglucosylase/hydrolase (XTH)
; Salt stress
; Drought stress
; Cold stress
; Chinese jujube
1. Introduction
The plant cell wall, an essential structure external to the cell membrane, is a dynamic and adaptable component that can be rapidly remodeled in response to both developmental and environmental signals. It is predominantly composed of polysaccharides, including cellulose, hemicellulose, pectin, and glycoproteins. The cell wall serves multiple critical functions, such as determining plant morphology, providing structural integrity, offering mechanical support, and defending against biotic and abiotic stresses [1,2,3]. In recent years, growing evidence has highlighted that cell wall loosening and reorganization are pivotal events in processes such as cell proliferation, volumetric expansion, morphological changes, and stress adaptation [4]. Of particular significance are cell wall-modifying proteins, which play a crucial role in structural remodeling, enabling plants to respond and adapt to fluctuating environmental conditions, including biotic and abiotic stress [4,5,6]. Xyloglucan endotransglucosylase/hydrolase (XTH) gene family plays a pivotal role in the plasticity and remodeling of the plant cell wall by mediating the cleavage and rejoining of xyloglucan polymers. Along with members of the glycoside hydrolase family 16 (GH16), XTH enzymes are thought to modulate cell wall mechanics and expansion through these catalytic activities [7,8,9]. The XTH family exhibits two primary enzymatic functions: xyloglucan endotransglucosylase (XET) activity, responsible for xyloglucan polymer rearrangement, and xyloglucan endohydrolase (XEH) activity, which hydrolyzes xyloglucan glycosidic bonds to facilitate cell wall expansion and morphogenesis [7]. Structurally, XTH proteins are categorized into four distinct groups: I/II, III A, III B, and an ancestral group. Enzymes within Group I/II primarily demonstrate glycosyltransferase activity, while those in Group III are predominantly associated with hydrolase activity [10,11]. GH16 family members characteristically contain the conserved Glyco_hydro_16 motif and an N-glycosylation site, which are essential for their enzymatic function [12]. With advancements in plant genome sequencing, the identification and functional regulation of XTH family members have been extensively studied in various plant species, including Arabidopsis thaliana [10], Oryza sativa [13], Solanum lycopersicum [14], Glycine max [15], Hordeum vulgare, and two Brassica species (Brassica rapa and Brassica oleracea) [16], as well as Camellia sinensis [17]. Additionally, various plant hormones modulate the expression of Xyloglucan Endotransglucosylase/Hydrolase (XTH) genes. Auxins, ethylene, gibberellins, abscisic acid, brassinosteroids, and cytokinins play a complex role in regulating XTH activity, thereby impacting cell wall architecture, growth processes, and defense mechanisms.
Extensive research has established the critical roles of XTHs (xyloglucan endotransglucosylase/hydrolases) in mediating plant responses to diverse environmental stresses. For example, the overexpression of CaXTH3 in both tomato and Arabidopsis has been shown to enhance salt and drought tolerance by regulating stomatal closure in transgenic plant [18,19]. the constitutive expression of PeXTH in tobacco conferred increased resistance to salt and cadmium [12]. In Arabidopsis, loss-of-function mutations in XTH15, XTH17, and XTH31 resulted in heightened aluminum tolerance compared to wild-type controls [20]. Additionally, AtXTH30 has been identified as a negative regulator of salt tolerance, influencing xyloglucan side chain modification, the concentration of xyloglucan-derived oligosaccharides, cellulose biosynthesis, and the stability of cortical microtubules [21]. Furthermore, XTH19 and XTH23 are implicated in lateral root development through the BES1-dependent pathway, facilitating the adaptation of lateral roots to salt stress [22]. Notably, the XTH19 mutant exhibited reduced freezing tolerance following cold and sub-zero acclimation, attributed to alterations in cell wall composition and structure [23]. In Brachypodium distachyon, a monocot model species, increased expression of XTH genes and those involved in Xylan biosynthesis has been closely linked to enhanced drought tolerance [5].
Chinese jujube (Ziziphus jujuba Mill.), native to China, is a vital fruit crop for commerce, the environment, and social value, especially in central and western China. With a global growing area exceeding 1.5 million hectares, China remains the sole supplier, supporting over 1,000 recognized cultivars [24]. Top cultivars are usually graft-propagated, and traits like drought and saline-alkali resistance are largely dependent on the rootstock. Cold tolerance and resistance to Jujube Witches' Broom (JWB) disease are essential for selecting robust cultivars. Under low saline-alkali stress (0–90 mM), jujube fruit shows increased sucrose, glucose, and fructose levels, along with enhanced activities of sucrose phosphate synthase. However, high saline-alkali stress (120–150 mM) inhibits sugar accumulation [25]. Drought stress, seen at soil moisture levels of 30%–50% field capacity, significantly reduces fruit redness and lowers anthocyanin and carotenoid levels [26]. Additionally, severe freezing stress enhances the galactose metabolism pathway [27]. Jujube witches' broom (JWB) disease, caused by phytoplasma infection, leads to substantial crop losses in the woody perennial fruit tree Ziziphus jujuba [18]. The presence of JWB-associated phytoplasmas decreases auxin accumulation and downregulates the expression of ZjARF4 in flower tissues, leading to phyllody symptoms. ZjTCP6 and ZjTCP16 are likely key regulatory factors involved in the jujube plant's response to JWB [28]. Additionally, PHYL1 promotes the proteasome-mediated degradation of crucial floral morphogenetic regulators in both jujube and Arabidopsis [29]. Moreover, CsXTH04 was identified as a potential candidate gene, upregulated in citrus varieties resistant to bacterial canker (Xanthomonas citri subsp.) and induced by exogenous treatments with Salicylic Acid and Methyl Jasmonate [30].
Genome-wide identification is critical to contemporary crop breeding strategies, as it helps identify candidate genes associated with traits important for agricultural productivity and nutritional enhancement. Similarly, RNA-Seq analysis is an effective approach for determining gene involvement and expression patterns across various biological processes, organs, and cell types. This method has proven valuable for studying genes involved in defense mechanisms and their specialized functions. The present research employs in-silico analysis to investigate the ZjXTH gene family in Chinese jujube and to understand how these genes contribute to the plant's defense against environmental stresses, including both biotic and abiotic factors. Examining ZjXTH expression patterns, transcriptional networks, and stress-responsive elements advances our understanding of plant adaptability and provides a foundation for genetic interventions aimed at enhancing stress tolerance in jujube cultivation.
2. Results
2.1. Identification and Sequence Analysis of XTH Genes
We successfully identified the XTH family genes in the Z. jujuba Dongzao genome available in the NCBI database under accession ASM3175591v1. Subsequent validation through BLAST-P, using 33 AtXTH proteins as references, revealed a total of 29 potential ZjXTH genes. These genes are predicted to encode proteins containing both the Gly-co_hydro_16 domain (PF00722) and the XET_C domain (PF06955). The genes were des-ignated as ZjXTH1 through ZjXTH29, based on their chromosomal locations (Table 1). The Expasy online platform was employed to analyze the properties of ZjXTH proteins, including their amino acid length (AA), aliphatic index (AI), instability index (II), molecu-lar weight (MW), and theoretical isoelectric point (pI). Signal peptides (SP) were predicted using the SignalP 6.0 Service. The results showed that the amino acid lengths ranged from 271 to 352, while the gene sequences varied from 815 to 1045 base pairs. The molecular weights of the proteins ranged from 30 to 40 kDa, corresponding to their respective lengths. The predicted isoelectric points (pI) ranged from 5.24 to 9.38, which may be at-tributed to differences in the polarity of the amino acids. Furthermore, 25 of the ZjXTH proteins were predicted to contain signal peptides, while 4 lacked any signal peptide. No-tably, an analysis of the Grand Average of Hydropathy (GRAVY) values for these proteins revealed negative scores, ranging from -0.137 to -0.713, indicating a predominantly hy-drophilic nature. Additional metrics, such as the instability index (II) and aliphatic index (AI), are also presented in Table 1.
2.2. Phylogenetic Classification of ZjXTH, Their Motif, and Gene Structure Analysis
A maximum likelihood analysis was performed to investigate the interspecific and intraspecific phylogenetic relationships among ZjXTH protein sequences. A total of 106 XTH proteins were compiled from four distinct species. To assess the evolutionary rela-tionships, a phylogenetic tree was constructed using 29 ZjXTH proteins from Ziziphus jujuba, 33 AtXTH proteins from Arabidopsis thaliana, 29 OsXTH proteins from Oryza sa-tiva, and 15 MdXTH proteins from Malus domestica. Phylogenetic analysis revealed that the 29 ZjXTH proteins were grouped into four distinct clades. Twelve ZjXTH proteins were classified under Group I/II, nine were grouped in the IIIB subgroup, six in Group IIA, and two ZjXTH proteins were placed in the ancestral group (Figure 1).
To compare the ZjXTH s at both the nucleic acid and protein levels, we conducted a com-prehensive analysis of the conserved motifs, gene structure, domains, and phylogenetic relationships among the 29 ZjXTH proteins (Figure 2). Using the MEME search tool, we iden-tified fifteen conserved motifs labeled as motifs 1–15. The results showed varying compo-sitions and distributions of these motifs among the ZjXTH proteins. Motifs 1 through 6 were identified in all ZjXTH proteins in the same arrangement. In contrast, motif 11 was found only in ZjXTH23, ZjXTH28, and ZjXTH29, while motif 14 appeared exclusively in ZjXTH1, ZjXTH6, ZjXTH7, and ZjXTH15 (Figure 2A). We also used the NCBI CD-blast tool to analyze the conserved domains of the ZjXTH proteins and found that the XTH domain was present in all 29 proteins. Interestingly, two other domains, Glyco_hydro_16 and the XET_C superfamily, were consistently present across all gene families (Figure 2B). Our in-vestigation into the genetic architecture of the XTH genes revealed differences in the num-ber of exons and introns they contain. In general, genes grouped together tend to share similar structures. For instance, all members of group 1 (ZjXTH1, ZjXTH2, ZjXTH3, ZjXTH4, ZjXTH5, ZjXTH6, ZjXTH9, ZjXTH12, ZjXTH15, ZjXTH17, ZjXTH18, ZjXTH21, ZjXTH23, ZjXTH24, ZjXTH25, ZjXTH26, ZjXTH28, and ZjXTH29) contained three exons and two introns in their coding regions. In contrast, members of group 2 (ZjXTH16, ZjXTH27, ZjXTH20, ZjXTH22, ZjXTH7, ZjXTH8, ZjXTH10, ZjXTH11, ZjXTH13, ZjXTH14, and ZjXTH19) possessed four exons and three introns. Additionally, it is noteworthy that within this group, ZjXTH16 and ZjXTH27 stand out as they possess the longest introns among the identified genes (Figure 2C).
2.3. Chromosomal Localization, Synteny Analysis, and PPI of ZjXTH Genes
To understand the chromosomal distribution of the Chinese jujube XTH genes, we physically mapped the locations of the 29 ZjXTH genes on the Chinese jujube genome. This investigation revealed a non-uniform distribution pattern across the chromosomes. Chromosome 01 harbored the highest number of ZjXTH genes [13], followed by Chromosomes 03, 06, 07, and 11, each containing one ZjXTH gene. ZjXTH genes were also located on Chromosomes 09 and 12, with three genes on each. Chromosome 10 contained two ZjXTH genes, while four ZjXTH genes were found on Chromosome 04 (Figure 3A).
Protein-protein interaction (PPI) analyses have revealed that several XTH proteins, including ZjXTH1, ZjXTH3, ZjXTH8, ZjXTH9, ZjXTH10, ZjXTH13, ZjXTH14, ZjXTH27, and ZjXTH29, are positively involved in the metabolism of polysaccharides, particularly glucuronarabinoxylan. Notably, ZjXTH1 interacts with Expansin-B1 (EXPB1), a protein crucial for cell wall loosening, which facilitates processes requiring cell wall modification. This interaction also contributes to the formation of root hairs, which are vital for efficient water and nutrient uptake. Additionally, ZjXTH3, ZjXTH9, and ZjXTH27 interact with Pectin Methylesterase 22 (PME22), which plays a pivotal role in the demethylesterification of pectin. This modification of cell wall properties is integral to the plant’s response to various environmental stresses, including salt stress, drought, and pathogen attack. These findings underscore the diverse and essential functions of XTH proteins in Ziziphus jujuba and highlight their potential roles in the physiological adaptations of Chinese jujube plants (Figure 3B)
In our current study, we utilized the MCScanX method to delve into the phenomenon of gene duplication within the ZjXTH gene family. In order to conduct a thorough investigation of the potential evolutionary pathways associated with the ZjXTH gene family, we have constructed three comparative syntenic maps. These maps include Ziziphus jujuba, a dicotyledonous plant, alongside monocotyledonous species such as Arabidopsis thaliana, Glycine max, and Oryza sativa. As seen in (Figure 3C). As a result, a comprehensive set of orthologous genes was identified, consisting of thirty-four pairs shared between Ziziphus jujuba and Glycine max, seventeen collinear gene pairs belonging to the ZjXTH gene family between Ziziphus jujuba and Arabidopsis thaliana, and four orthologous genes shared between Ziziphus jujuba and Oryza sativa. The higher frequency of orthologous events detected in GmXTHs-ZjXTHs, relative to other orthologous events, indicates a shorter evolutionary gap between Ziziphus jujuba and Glycine max. The evolutionary significance of XTH genes may have been influenced by the existence of a minimum of four pairs of orthologous genes in select OsXTH genes. The results of this investigation suggest that the ZjXTH genes identified in Ziziphus jujuba could have derived from orthologous genes present in other plant species (Figure 3C).
2.4. Cis-Regulatory Elements in the Promoter Region of ZjXTH
The upstream region, consisting of 2 kb nucleotides from the start codon, was analyzed to investigate the promoter cis-regulatory elements (CREs). The data were submitted for analysis through the PlantCARE database. The results revealed a total of 607 CREs distributed across all ZjXTH genes, with ZjXTH20 containing the highest number at 33, and ZjXTH10 possessing the fewest at 10 CREs. These elements were systematically classified into three main categories: regulatory elements involved in Growth and Development, Stress-responsive elements, and Phytohormone-responsive elements, as visually illustrated for clearer understanding (Figure 4).
(i). The regulatory elements associated with growth and development represented the largest proportion of CREs within the promoter regions of the ZjXTH genes, comprising 350 elements and accounting for 58% of the total CREs. All ZjXTH genes contained elements from this category, predominantly those linked to light responsiveness. Specifically, light-responsive CREs, such as G-box, Box II, Box 4, TCT-motif, and I-box, were highly prevalent, making up 308 elements, which constituted 88% of the total elements in this category. Additional regulatory elements included meristem expression elements (CAT-box) with 14 elements (4%), endosperm expression elements (GCN4_motif) and zein metabolism regulatory elements (O2-site), each with nine elements (2.5%). Furthermore, cis-elements associated with circadian control and seed-specific regulation were present in five elements (1.5%) each, across the promoters of all ZjXTH genes.
(ii). Second category comprises stress-responsive CREs, with a total of 132 elements (22%) identified in the promoter regions of the ZjXTH genes. At least one CRE from this category was found in every ZjXTH gene. Among these, anaerobic induction elements (ARE) were the most abundant, with 81 elements (61.4%). This was followed by drought-inducibility elements (MBS), comprising 19 elements (14.4%), while defense and stress-responsive elements (TC-rich repeats) and low-temperature responsive elements (LTR) were equally represented, each contributing 16 elements (12.1%).
(iii).Third category of CREs identified in the promoter regions of ZjXTH genes is associated with phytohormone responsiveness, comprising a total of 125 predicted CREs (20%). The most abundant of these elements is the MeJA-responsive motif (TGACG-motif), which accounts for 36 CREs (28.8%). This is followed by abscisic acid-responsive elements (ABRE) with 30 CREs (24%) and salicylic acid-responsive elements (TCA-element) with 29 CREs (23.2%). Gibberellin-responsive elements contributed 17 CREs (13.6%), while auxin-responsive elements (TGA-element) represented the smallest proportion, with 13 CREs (10.4%).
2.5. Functional GO Annotation and Ka/KS Analysis of ZjXTHs
The functional annotation of ZjXTH genes identified 14 distinct Gene Ontology (GO) terms, classified into three major categories: molecular function, biological process, and cellular component. The molecular function of ZjXTH genes was predominantly linked to xyloglucosyl transferase activity (GO:00167762), aligning with the established role of ZjXTH genes in xyloglucan modification. In terms of cellular localization, ZjXTH genes were mainly associated with the cell wall (GO:005618), external encapsulating structure (GO:0030312), and apoplast (GO:0048046), highlighting their critical role in the structural development of Chinese jujube (Ziziphus jujuba) plants. Under the biological process category, ZjXTH genes were implicated in regulating metabolic pathways, including xyloglucan (GO:0010411) and hemicellulose metabolism (GO:0010410). Additionally, several ZjXTH genes were linked to the positive regulation of cellular processes and developmental pathways. Collectively, these findings underscore the significant contributions of ZjXTH genes to the growth and development of Chinese jujube plants (Figure 5).
To gain a comprehensive insight into the evolutionary dynamics and selective pressures influencing the ZjXTH gene family, we calculated the nonsynonymous-to-synonymous substitution rate ratio (Ka/Ks) for all 29 ZjXTH genes within the Chinese jujube (Ziziphus jujuba) genome. These results are systematically detailed in Table 2. In the context of evolutionary selection, a Ka/Ks value less than 1 indicates purifying selection, while values exceeding 1 suggest positive or diversifying selection. Two distinct gene groups emerged from our analysis: the first group, including gene pairs such as ZjXTH2-ZjXTH5 and ZjXTH4-ZjXTH6, displayed Ka/Ks values ranging from 0.11 to 0.9, indicating strong purifying selection. The second group, represented by gene pairs ZjXTH15-ZjXTH26 and ZjXTH17-ZjXTH8, exhibited Ka/Ks ratios between 1.18 and 1.54, suggesting a more pronounced role of positive selection. Overall, these findings imply that purifying selection has predominantly shaped the evolutionary trajectory of ZjXTH genes, preserving their conserved structural features and highlighting their critical functional roles within the genome.
2.6. Expression Patterns of ZjXTHs on Subcellular Levels and Different Tissues
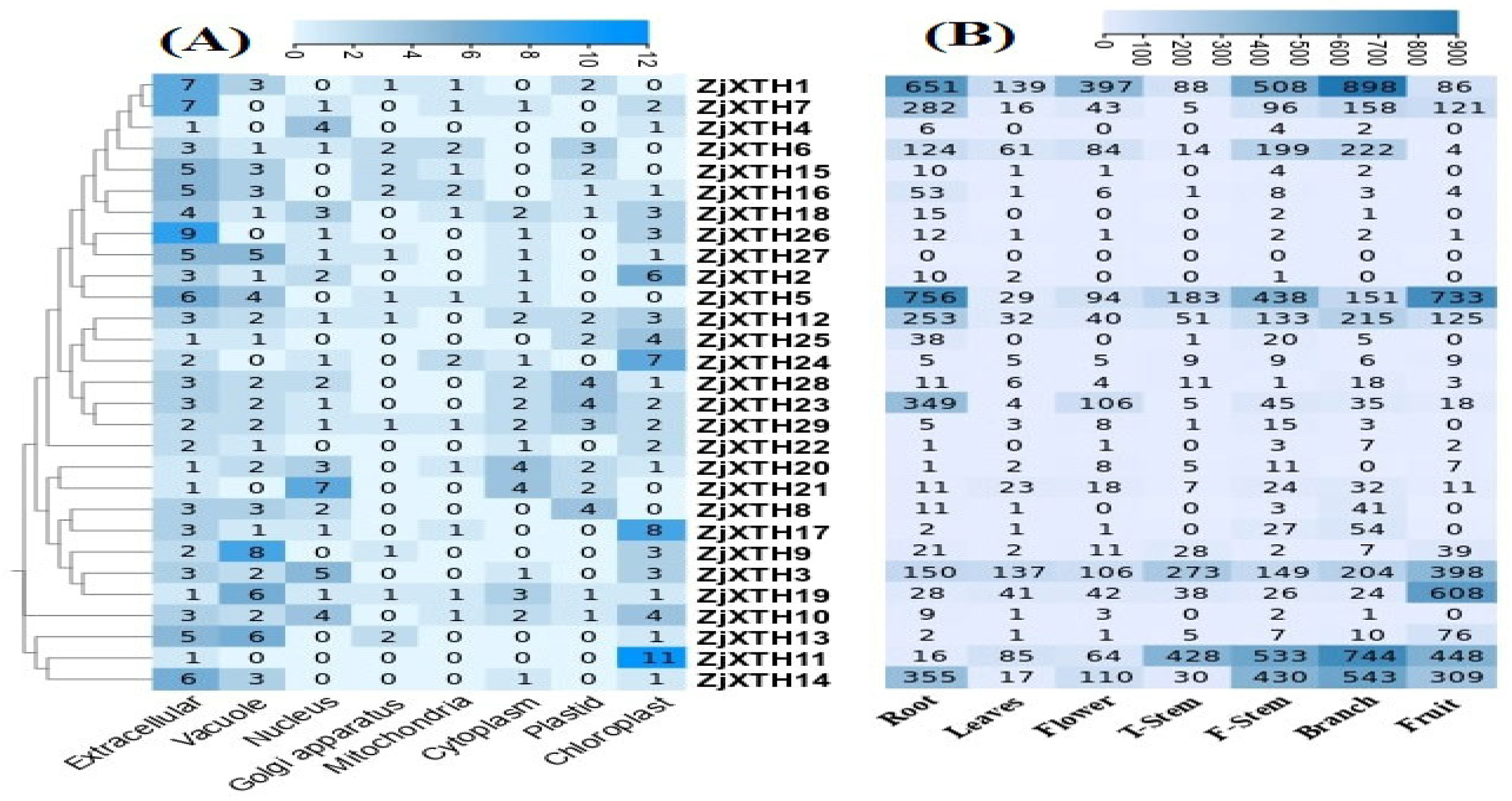
To determine the subcellular localization of ZjXTH proteins, we employed the WoLF PSORT online database. The analysis revealed that the majority of ZjXTH proteins were predominantly localized to the extracellular space (Figure 5-A), indicating their potential role as catalysts in decomposition processes. Specifically, ZjXTH1, ZjXTH5, ZjXTH7, ZjXTH14, and ZjXTH26 were predicted to be exclusively localized in the extracellular space. Additionally, certain ZjXTH proteins were predicted to localize to other subcellular compartments. For instance, ZjXTH11 and ZjXTH17 were significantly localized in the chloroplast, while ZjXTH20 and ZjXTH21 were predicted to localize in both the cytoplasm and nucleus. ZjXTH9 was primarily found in the vacuole, and ZjXTH8, ZjXTH23, and ZjXTH28 showed traces of localization within plastids.
We investigated the expression profile of ZjXTHs in various tissues using publicly available transcriptome data by RNA-Seq. The tissues analyzed included Root, Leaves, Flower, T-stem, F-stem, Branch, and Fruit as shown in (Figure 5-B), we observed significant differences in the expression patterns of ZjXTH transcripts. Most ZjXTH genes, including ZjXTH1, ZjXTH5, and ZjXTH14, exhibited high expression levels in vegetative tissues such as root, flower, F-stem, and branches. In contrast, ZjXTH5 demonstrated notably high expression in fruit, with the highest levels observed in T-stem, indicating its potential importance in tissue development. The expression of ZjXTH127 was absent across all tissues analyzed. Furthermore, ZjXTH2, ZjXTH4, ZjXTH15, ZjXTH18, ZjXTH26, and ZjXTH27 showed minimal expression, suggesting a lesser role in organ development. While most ZjXTH genes exhibited consistent tissue-specific expression patterns, ZjXTH27 was uniquely downregulated across all tissues. Notably, ZjXTH3, ZjXTH7, ZjXTH11, and ZjXTH12 were predominantly expressed in roots, leaves, flowers, T-stem, F-stem, branches, and fruit. Among these, ZjXTH19 displayed the highest expression in fruit. These results highlight a distinct, tissue-specific expression profile for ZjXTH genes, suggesting their specialized roles in various tissues and developmental processes.
Figure 6.
Heatmaps were generated to examine the expression patterns of ZjXTHs under various cellular compartments, developmental stages, and stress conditions. The heatmaps were constructed and visualized using TBTools software. (A) the sub-cellular localization of ZjXTH proteins was predicted using the WoLF PSORT web tool. (B) The tissue-specific expression profiles of ZjXTH at different developmental stages of the Chinese jujube plant were analyzed using publicly available transcriptome data and displayed in a heatmap. The normalized fragments per kilobase of transcript per million fragments (FPKM) values.
Figure 6.
Heatmaps were generated to examine the expression patterns of ZjXTHs under various cellular compartments, developmental stages, and stress conditions. The heatmaps were constructed and visualized using TBTools software. (A) the sub-cellular localization of ZjXTH proteins was predicted using the WoLF PSORT web tool. (B) The tissue-specific expression profiles of ZjXTH at different developmental stages of the Chinese jujube plant were analyzed using publicly available transcriptome data and displayed in a heatmap. The normalized fragments per kilobase of transcript per million fragments (FPKM) values.
2.7. Expression Profiles of ZjXTH Under Salt Stress
A total of 29 ZjXTH genes were identified, and their expression patterns under salt stress were thoroughly analyzed. The findings revealed significant temporal differences in the expression of these genes in response to salt stress. ZjXTH1 and ZjXTH5 were notably upregulated after one and two days of salt treatment in the salt-tolerant tetraploid sour, displaying the most rapid and robust response. These genes were the most highly expressed, suggesting their critical roles in the salt stress response. In contrast, ZjXTH6, ZjXTH7, ZjXTH12, and ZjXTH13 were downregulated under the same conditions. Some genes exhibited negatively regulated expression patterns. For instance, the expression of ZjXTH3 and ZjXTH19 remained relatively unchanged in the tolerant cultivar during both control and salt stress treatments but significantly increased in the sensitive cultivar. Additionally, ZjXTH10, ZjXTH16, ZjXTH18, ZjXTH20, and ZjXTH21 showed no detectable expression in either cultivar, indicating that they may not play a pivotal role in the salt stress response (Figure 7).
2.8. Expression Profiles of ZjXTH Under Drought Stress
The results revealed substantial temporal variations in the expression of ZjXTH genes in response to drought stress. ZjXTH1 and ZjXTH5 were highly expressed at 6, 12, and 48 hours following drought treatment in the drought-tolerant jujube cultivar, demonstrating the most rapid and pronounced response. These genes exhibited the highest expression levels, indicating their pivotal roles in the drought stress response. Conversely, ZjXTH2, ZjXTH4, ZjXTH6, ZjXTH12, and ZjXTH15 were downregulated under the same conditions. They showed relatively stable expression during drought stress treatments but were significantly upregulated in the tolerant cultivar. Furthermore, ZjXTH8 and ZjXTH19 exhibited negatively regulated expression patterns (Figure 8).
2.9. Expression Profiles of ZjXTH Under Freezing Stress
We analyzed the gene expression patterns of ZjXTHs under cold stress, as presented in Figure 9. The analysis revealed six distinct stages of differential gene expression. In the first stage, ZjXTH1, ZjXTH7, ZjXTH4, and ZjXTH6 were moderately expressed under freezing conditions, with ZjXTH1 exhibiting a notably higher expression level. In the second stage, ZjXTH6 and ZjXTH18 were moderately expressed, while ZjXTH8 showed a significant increase in expression, particularly in cold-tolerant cultivars. During the third stage, ZjXTH2, ZjXTH12, and ZjXTH25 were downregulated in cold-sensitive cultivars but upregulated in cold-tolerant cultivars. Notably, ZjXTH5 was highly expressed. In stages four and five, ZjXTH28, ZjXTH23, ZjXTH29, ZjXTH22, ZjXTH17, and ZjXTH3 exhibited downregulation in tolerant cultivars, while ZjXTH19 was negatively regulated. Additionally, in the six XTH stage, ZjXTH11 and ZjXTH14 were downregulated, except for ZjXTH14, which was highly expressed in cold-tolerant cultivars. Furthermore, ZjXTH21 showed no expression in either cultivar, suggesting that this gene may not play a critical role in the cold stress response.
2.10. Expression Profiles of ZjXTH Response to Phytoplasma Infection
To investigate the gene expression patterns of ZjXTHs under different biotic stresses, we analyzed transcriptome RNA-seq data of ZjXTHs in response to jujube witches' broom phytoplasma (JWB). Z. jujuba ‘Huping’, a sensitive cultivar (Figure 10), and Z. mauritiana ‘Cuiming’, a tolerant cultivar (Figure 10), were used for comparison, and expression heatmaps were generated. ZjXTH1, ZjXTH3, and ZjXTH5 were upregulated following JWB stress in the infected-tolerant jujube cultivar, demonstrating the most rapid and pronounced response. These genes exhibited the highest expression levels, indicating their pivotal roles in the jujube witches' broom phytoplasma stress response. Conversely, ZjXTH6, ZjXTH7, ZjXTH11, ZjXTH16, and ZjXTH25 were downregulated under the infected-tolerant conditions, showing relatively stable expression during phytoplasma stress treatments in the tolerant cultivar. Furthermore, ZjXTH20 exhibited negatively regulated expression patterns (Figure 10).
3. Discussion
Xyloglucan endotransglucosylases/hydrolases (XTH) genes have been thoroughly examined in several plants due to their crucial functions in plant growth and stress response [31]. Chinese jujube (Z. jujuba) is a plant of significant commercial value, but its growth and development are affected by several abiotic and biotic conditions, including salt [24], drought [32], cold [27], and phytoplasma infection [18]. However, jujube has not been examined in relation to this gene family.
According to our comprehensive identification methodology, 29 XTH genes were identified in the Chinese jujube genome and designated ZjXTH-1 to ZjXTH-29 [33]. The number of identified XTH genes is significantly lower than that in other species, such as Glycine max (61), Solanum lycopersicum (37), wheat (71), and tobacco (56) [15,34,35,36]. The functional properties of these genes are closely associated with their structural and physicochemical features. This analysis revealed considerable variation among the 29 ZjXTH protein members in terms of protein sequence length, molecular weight, isoelectric point (pI), and intron-exon distribution. In Z. jujuba, this variation indicates a significant amount of diversity among the XTH family members. Additionally, most ZjXTH genes were predicted to be in extracellular space, while a few were located in the golgi apparatus, vacuole, mitochondrion, and nuclear region. This contrasts with previous reports for other XTH protein members in other plant species, where the majority of XTH proteins were found in the plasma membrane rather than the extracellular space or other locations [37]. Furthermore, phylogenetic analysis indicated that ZjXTH protein families were clustered into four groups, similar to those observed for XTH proteins from cucumber [31]. Interestingly, the ZjXTH proteins belonging to the same group demonstrated similar gene structures and conserved sequence expression, which is consistent with previously documented literature, suggesting that XTH members within the same group may exhibit analogous functionalities. Moreover, most of the ZjXTH genes exhibit two main conserved domains (Glyco_hydro_16 and XET_C). This suggests a potential evolutionary divergence, highlighting the role of the XET_C domain in the evolutionary trajectory of XTH proteins in Z. jujuba.
Phylogenetic classification of XTH proteins from Z. jujuba, O. sativa, M. domestica, and A. thaliana indicates that ZjXTH genes can be classified into four distinct groups. Previous research has classified XTH gene families into specific categories across different plant species. In tobacco, eight familial groupings were identified, whereas three groups were observed in peanut [38], barley, and sweet potato. In contrast, poplar displayed four distinct categories. ZjXTH genes were observed to cluster more closely with XTH proteins from G. max than O. sativa, suggesting a closer evolutionary relationship between the XTH proteins in Z. jujuba and those of A. thaliana rather than O. sativa. A chromosomal localization study revealed that ZjXTHs were unevenly distributed across 9 of the 12 chromosomes of Z. jujuba. Previous studies have demonstrated that a group of gene functions shows significant conservation across several plant species. Therefore, it is essential to identify genuine orthologs across various plant species using synteny analysis. The results of the synteny study revealed a notable degree of synteny between the Z. jujuba genome and those of G. max and A. thaliana, showing 34 and 17 syntenic blocks of ZjXTH with G. max and A. thaliana, respectively. Conversely, only four syntenic blocks were discovered with O. sativa [39].
Protein-protein interaction analysis offers insights into potential functional associations among ZjXTH genes. ZjXTH1 plays a positive role in the metabolism of polysaccharides, particularly glucuronarabinoxylan. Notably, ZjXTH1 interacts with Expansin-B1 (EXPB1), a protein crucial for cell wall loosening, which facilitates processes that require cell wall modification. This interaction also contributes to the formation of root hairs, which are vital for efficient water and nutrient uptake. Meanwhile, the transcripts of some members exhibited strong responses to salt, low temperature, and drought stress. For example, overexpression of GsEXPB1 in soybean significantly increased the number of hairy roots, root length, root weight, and tolerance to salt stress [40]. Additionally, ZjXTH3, ZjXTH9, and ZjXTH27 interact with Pectin Methylesterase 22 (PME22), which is crucial for the demethylesterification of pectin. The alteration of cell wall characteristics is essential to the plant's response to diverse environmental conditions [41]. A further study indicates that purified recombinant CaPMEI1 protein not only inhibits PME but also exhibits antifungal action against some harmful plant fungi. Virus-induced gene silencing of CaPMEI1 in pepper increases vulnerability to Xanthomonas campestris pv., along with diminished expression of several defense-related genes [42]. Furthermore, transgenic Arabidopsis lines overexpressing CaPMEI1 show increased resistance to Pseudomonas syringae pv [43].
In several species, XTHs were predicted and identified as modulating light-associated genes and phytohormonal-regulated photomorphogenesis. Therefore, we predicted and analyzed the promoter cis-regulatory elements (CREs) in the 2k bp upstream of ZjXTHs. The results indicated that a total of 607 CREs were expected in all of the ZjXTHs. Growth and development regulatory elements, stress-responsive elements, and phytohormone-responsive elements comprised these CREs. In the promoter region of ZjXTH genes, the growth and development regulatory elements accounted for the highest proportion of CREs, with a total of 350 elements and 58% of the total CREs. CREs' mild responsiveness was exceedingly prevalent in this category, comprising 308 elements and 88% of the category's total elements. This was comparable to research conducted on Rosaceae sp. and cucumber [44]. Light significantly impacts a variety of plant processes, such as development, photosynthetic regulation, and circadian rhythm. ZjXTHs may promote chloroplast development, which may mitigate excessive light damage and enhance photosynthetic activity, as previously mentioned [45]. In turn, this leads to an increase in the conversion of light energy into chemical energy, which in turn leads to a greater accumulation of carbohydrates [46,47]. Nevertheless, this hypothesis can be further investigated by conducting additional experiments that involve the application of excessive light stress to Chinese jujube plants. Other elements in this category included the regulation of Zein metabolism, the expression of Endosperm, the regulation of circadian control, and seed-specific elements.
The second category contains stress-responsive CREs, with 132 (22%) CREs predicted in the promoter regions of ZjXTHs. This category encompasses defense and stress responsiveness, anaerobic induction, low-temperature responsiveness, MYB drought-inducibility, and defense and stress responsiveness. The most prevalent elements in this category were anaerobic induction elements, which suggests that ZjXTHs play a critical role in the resistance of Chinese jujube plants to environmental stress [48]. While the third category of CREs identified in the promoter regions of ZjXTHs is associated with phytohormone response, there are a total of 125 (20%) predicted CREs. Methyl jasmonate (MeJA)-responsive elements are the most prevalent phytohormone-responsive elements, followed by abscisic acid-responsive elements, salicylic acid-responsive elements, gibberellin-responsive elements, and auxin-responsive elements.The identification of CREs in the promoter regions of ZjXTHs offers a unique perspective on the regulation of gene expression, with a particular emphasis on phytohormone signaling, stress response, and growth and development. This information can be employed to investigate gene expression and regulation, which will facilitate the enhancement of crop yields and the ability to withstand duress. The identification of molecular targets for the enhancement and propagation of Chinese jujubes can be facilitated by an understanding of the functions of these regulatory elements in ZjXTH genes.
Throughout growth and developmental phases, plants are subjected to numerous abiotic stresses, including low temperatures, salinity, and drought. The XTH enzyme is a crucial modulator of cell wall modifying, playing a significant role in both the synthesis and degradation of cell wall components. It is essential for maintaining cell wall integrity and resilience, contributing to structural stability and stress tolerance under both optimal and adverse environmental conditions. Our research revealed that ZjXTH1 and ZjXTH5 may be essential for the regulation of salt tolerance in a jujube cultivar under lack of water and salt stress. The Arabidopsis gene XTH23, a homolog of ZjXTH1, also exhibited upregulation under salt stress, as evidenced by similar results [22]. Dhar et al. demonstrated that overexpressing AtXTH22 enhanced cell division, elongation, and primary root growth [49]. Similarly, Takeda et al. reported that the application of xyloglucans to excised pea stems resulted in increased stem rigidity [50]. Numerous studies also underscore the critical role of the XTH gene family in bolstering plant stress tolerance. For instance, overexpression of CaXTH3 in tomato and Arabidopsis thaliana enhanced resistance to drought and salinity by modulating stomatal dynamics [19,51]. Conversely, Bi et al. found that TaXTH17 negatively influenced plant resistance to salt and drought stress, suggesting a diverse functional spectrum within the XTH gene family [52]. Additionally, the heterologous expression of XTH in Populus tomentosa markedly increased total intracellular sugar content and yeast cell osmotolerance, indicating that PtoXTH27 and PtoXTH34 may be integral to osmotic stress response mechanisms [53]. Cold stress is a significant challenge for plants, with extreme instances potentially resulting in plant mortality. This research revealed that ZjXTH1, ZjXTH5, ZjXTH18, and ZjXTH14 were expressed, with ZjXTH1 demonstrating a significantly elevated expression level under freezing conditions. In cotton, Overexpression of the GhXTH22 gene resulted in higher enzyme levels and activity, contributing to increased hemicellulose and cellulose accumulation. This accumulation enhances cell wall rigidity, which, in turn, influences cell wall biosynthesis and modulates the expression of associated genes [54]. While Tan et al found that MaXTH7 was identified in banana as a contributor to tolerance to low-temperature stress [55]. Collectively, these changes contribute to improved cold tolerance in plants. It is thus hypothesized that XTH positively regulates plant cold tolerance through modulations of cell wall rigidity. Phytoplasma-associated diseases have significantly jeopardized the cultivation of several commercially vital plants. This study demonstrated that the expression levels of ZjXTH1, ZjXTH3, and ZjXTH5 increased in response to Jujube witches' broom (JWB) stress in the infected, tolerant jujube cultivar, exhibiting the most immediate and significant reaction. Li et al found that CsXTH04 is upregulated in response to citrus bacterial canker (CBC). Furthermore, overexpression of CsXTH04 increased susceptibility to CBC in transgenic citrus lines [30]. However, numerous questions remain unresolved and warrant further exploration, including the functional mechanisms and stress responses of XTH in the context of bacterial and fungal infections, as well as the pathways through which microbial presence induces XTH gene expression.
Taken together, this discovery underscores the dynamic nature of these genes and their potential involvement in the precise adjustment of Chinese jujube responses to the continuously evolving signals from their environment.
4. Materials and Methods
4.1. Genome-Wide Identification of XTH Genes
A query was performed on the NCBI database (https://www.ncbi.nlm.nih.gov/) to retrieve the protein sequences of the Chinese jujube (Ziziphus jujuba). The Hidden Markov Model (HMM) profiles for PF00722 (also known as Glyco hydro 16) and PF06955 (also known as XET C) were acquired from the Pfam database. These profiles correspond to protein domains associated with XTH. The profiles were used as queries in the HMMER3.0 program, using the default E-value, to do a search throughout the database. The researchers used the PFAM and SMART databases to identify the conserved domains that are present in the putative Xyloglucan endotransglucosylase/hydrolase (XTH) proteins of Ziziphus jujuba. In order to provide a more comprehensive examination, only proteins including both the PF00722 and PF06955 domains were retained. Protein characterization was performed using the ExPASy database (https://web.expasy.org/protparam/), using the identified ZjXTHs. The protein sequences were analyzed to determine their predicted features, such as length, molecular weight (MW), instability index (II), isoelectric point (PI), aliphatic index (AI), and grand average of hydropathicity (GRAVY). The use of WoLF PSORT online database (https://wolfpsort.hgc.jp/) was employed for the purpose of predicting cellular localization [37].
4.2. Multiple Sequence Alignment and Phylogenetic Analysis
The XTH protein sequences of A. thaliana were obtained by accessing the Arabidopsis Information Resource (TAIR), whereas OsXTHs and MdXTHs were obtained via the Phytozome database (https://phytozome-next.jgi.doe.gov/). The phylogenetic tree was constructed using the MEGA7.0 software, using the aligned sequences. The tree was constructed using the neighbor-joining (NJ) approach using the default parameters. A phylogenetic tree with 1,000 duplicates was used to evaluate the reliability of the tree via the utilization of the bootstrap method. The graphical representation of the resulting tree was shown using ITOL v3 (https://itol.embl.de/).
4.3. Motif Analysis and Exon-Intron Structure of ZjXTHs
Utilizing the default settings, the MEME 20 suite (https://meme-suite.org/meme/) was used to ascertain the preserved motifs. The TBtools software was used to view the exons and introns of each XTH that were found using the GFF file of the Z. jujuba genome, which included gene structure information.
4.4. Chromosomal Locations, Synteny and Duplications Analyses of ZjXTHs
After obtaining the genomic data of Z. jujuba from NCBI, the location information of the Z. jujuba XTH genes was used to map each gene to its matching chromosomal position. Repetitive events of ZjXTHs family genes across several species were computed using the MCscan. To compute the recurrent occurrences of ZjXTHs among Z. jujuba and the other three species (Arabidopsis, G. max, and O. sativa), the TBtools program used a Dual Synteny Plotter.
4.5. Cis-Element Putative Promoter Regions and Ka/Ks Analysis
The functions and patterns of gene expression are influenced by the cis elements present in the promoter regions. In order to ascertain the presence of cis-regulatory elements in the promoters of XTH genes in Z. jujube, NCBI-extracted 2 kb upstream sequences of the genes' coding area were used as input in the PlantCARE program (https://bioinformatics.psb.ugent.be/webtools/plantcare/html/). TBtools software was then employed to screen and display the cis-elements. In order to approximate the evolutionary trajectory of the ZjXTH genes, the TBtools program was used to compute the ratio of synonymous mutation rate to nonsynonymous mutation rate (Ka/Ks). The evolutionary calculation used ZjXTH protein findings above the 60 percent sequence similarity criterion that were obtained by blasting the proteins against the NCBI database. Following this, a tab-delimited text file containing the similarity data was generated in order to compute Ka/Ks using the TBtools program. The Ks value was used to approximate the divergence time in millions of years for each gene pair, with a substitution rate of 6.1 × 10−9 substitutions per site per year. To determine the divergence time (T), T was computed as T = Ks/(2 × 6.1 × 10−9) × 10−6 million years ago (Mya).
4.6. Protein Interaction Network and Gene Ontology
The protein interaction network was constructed using the STRING (https://cn.string-db.org/) database based on the orthologous genes between Z. jujuba and Arabidopsis. The predicted interaction network was visualized through the Cytoscape software.
ShinyGO v0.77 (http://bioinformatics.sdstate.edu/go/) was used to obtain gene ontology (GO) annotation against A. thaliana. The p-value cut-off (FDR) at 0.01 is set to calculate GO enrichment.
4.7. Gene Expression Analysis of ZjXTH Genes Under Various Stresses
The RNA-seq expression data for the Z. jujuba were obtained from the National Center for Biotechnology Information database. The accession codes for the data are GCF_000826755.1. The FPKM data of several tissues underwent hierarchical clustering analysis using Tbtools.
To measure the expression of XTH genes under stress tolerance, RNA-seq data of our research previously reported were used. Z. jujuba. var. spinosa diploid and tetraploid, representing sensitive and tolerant types respectively, were used in a salinity treatment, gradually applied at 50, 100, and finally 150 mM NaCL [24]. For drought treatment, PEG6000 was applied to diploid and tetraploid Z. jujuba var. spinosa seedlings, with concentrations of 5%, 10%, 15%, and finally 20% over 1-day intervals. For freezing treatment, branches were collected from the cold-sensitive cultivar ‘Dongzao’ and the cold-tolerant cultivar ‘Jinsixiaozao’. Some of the branches were placed at 4 °C for 10 hours and used as controls, while others were treated and maintained at freezing temperatures of -10 °C, -20 °C, -30 °C, and -40 °C, also for 10 hours. The xylem from these branches was then collected for RNA-seq library construction and sequencing [27]. For biotic treatment, ‘Cuimi’ (Z. mauritiana Lam. JWB tolerant) and ‘Huping’ (Z. jujuba JWB susceptible) scions were grafted onto the diseased ‘Jinsixiaozao’ (Z. jujuba Mill.), and the leaves were sampled for RNA-seq [18]. XTH expression data were analyzed using the FPKM values of assembled transcripts. FPKM values for different tissues were subjected to hierarchical clustering analysis using TBtools and Origin.
5. Conclusions
Our in-depth identification of the Xyloglucan Endotransglucosylase/Hydrolase (XTH) genes in Chinese jujube provides critical insights into their structure, function, and evolution. The ZjXTH genes are strategically positioned in the group I/II category, which is crucial for cell wall dynamics. Phylogenetic analysis reveals similarities among plant species, while gene architecture emphasizes critical characteristics such as the conservation of the Glyco_hydro_16 domain and Motifs 1 and 6, which regulate xyloglucan modification. For all ZjXTHs, our analysis of promoter cis-regulatory elements predicted 607 such elements. The interplay between Chinese jujube defense mechanisms and gene expression is exemplified by cis-acting elements, while environmental signal conversion is driven by TCA and TGACG motifs. ZjXTH1 functions as a key regulator in the PPI network, enabling the precise regulation of ZjXTH gene expression in conjunction with EXPB1 and PME22. These results underscore the critical role of XTH genes in the growth, response, and adaptation of Chinese jujube, providing valuable insights for innovative applications in sustainable agriculture and beyond.
Author Contributions
Writing - original draft, Methodology and formal analysis, M.R.; formal anal-ysis, M.T.; Visualization, C.Y., L.J. and X.Z.; Investigation H.Z.; Data provoding, Y.X. and S.S.; Writing - review & editing, X.P.; All authors have read and agreed to the published version of the manuscript.
Funding
This work was financially supported by the National Key Research and Development Program of China (2022YFD2200404), National XA Science and Technology innovation project (2022XACX1100), Guangdong Provincial Key Laboratory of Silviculture, Protection, and Utilization, and the open funds of the National Key Laboratory for Germplasm Innovation and Utilization of Horticultural Crops. The central government guides the local science and technology development project “Germplasm Innovation of Xinjiang Characteristic Fruit Tree”.
References
- Carpita, N. C.; Gibeaut, D. M. Structural models of primary cell walls in flowering plants: consistency of molecular structure with the physical properties of the walls during growth. The Plant Journal 1993, 3, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Somerville, C.; Bauer, S.; Brininstool, G.; Facette, M.; Hamann, T.; Milne, J.; et al. Toward a systems approach to understanding plant cell walls. Science 2004, 306, 2206–2211. [Google Scholar] [CrossRef] [PubMed]
- Novaković, L.; Guo, T.; Bacic, A.; Sampathkumar, A.; Johnson, K. L. Hitting the wall—sensing and signaling pathways involved in plant cell wall remodeling in response to abiotic stress. Plants 2018, 7, 89. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Gao, Y.; Zhang, L.; Zhou, Y. The plant cell wall: Biosynthesis, construction, and functions. Journal of Integrative Plant Biology 2021, 63, 251–272. [Google Scholar] [CrossRef]
- Lenk, I.; Fisher, L. H. C.; Vickers, M.; Akinyemi, A.; Didion, T.; Swain, M.; et al. Transcriptional and metabolomic analyses indicate that cell wall properties are associated with drought tolerance in Brachypodium distachyon. International Journal of Molecular Sciences 2019, 20, 1758. [Google Scholar] [CrossRef]
- Houston, K.; Tucker, M. R.; Chowdhury, J.; Shirley, N.; Little, A. The plant cell wall: a complex and dynamic structure as revealed by the responses of genes under stress conditions. Frontiers in plant science 2016, 7, 984. [Google Scholar] [CrossRef]
- Rose, J. K. C.; Braam, J.; Fry, S. C.; Nishitani, K. The XTH family of enzymes involved in xyloglucan endotransglucosylation and endohydrolysis: current perspectives and a new unifying nomenclature. Plant and cell physiology 2002, 43, 1421–1435. [Google Scholar] [CrossRef]
- Van Sandt, V. S. T.; Suslov, D.; Verbelen, J.-P.; Vissenberg, K. Xyloglucan endotransglucosylase activity loosens a plant cell wall. Annals of botany 2007, 100, 1467–1473. [Google Scholar] [CrossRef]
- Miedes, E.; Suslov, D.; Vandenbussche, F.; Kenobi, K.; Ivakov, A.; Van Der Straeten, D.; et al. Xyloglucan endotransglucosylase/hydrolase (XTH) overexpression affects growth and cell wall mechanics in etiolated Arabidopsis hypocotyls. Journal of experimental botany 2013, 64, 2481–2497. [Google Scholar] [CrossRef]
- Yokoyama, R.; Nishitani, K. A comprehensive expression analysis of all members of a gene family encoding cell-wall enzymes allowed us to predict cis-regulatory regions involved in cell-wall construction in specific organs of Arabidopsis. Plant and cell physiology 2001, 42, 1025–1033. [Google Scholar] [CrossRef]
- Baumann, M. J.; Eklöf, J. M.; Michel, G.; Kallas, A. M.; Teeri, T. T.; Czjzek, M.; et al. Structural Evidence for the Evolution of Xyloglucanase Activity from Xyloglucan Endo-Transglycosylases: Biological Implications for Cell Wall Metabolism. The Plant Cell 2007, 19, 1947–1963. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Wang, W.; Sun, J.; Ding, M.; Zhao, R.; Deng, S.; et al. Populus euphratica XTH overexpression enhances salinity tolerance by the development of leaf succulence in transgenic tobacco plants. Journal of experimental botany 2013, 64, 4225–4238. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, R.; Rose, J. K. C.; Nishitani, K. A surprising diversity and abundance of xyloglucan endotransglucosylase/hydrolases in rice. Classification and expression analysis. Plant Physiology 2004, 134, 1088–1099. [Google Scholar] [CrossRef] [PubMed]
- Saladié, M.; Rose, J. K. C.; Cosgrove, D. J.; Catalá, C. Characterization of a new xyloglucan endotransglucosylase/hydrolase (XTH) from ripening tomato fruit and implications for the diverse modes of enzymic action. The Plant journal : for cell and molecular biology 2006, 47, 282–295. [Google Scholar] [CrossRef]
- Song, L.; Valliyodan, B.; Prince, S.; Wan, J.; Nguyen, H. T. Characterization of the XTH gene family: new insight to the roles in soybean flooding tolerance. International Journal of Molecular Sciences 2018, 19, 2705. [Google Scholar] [CrossRef]
- Wu, D.; Liu, A.; Qu, X.; Liang, J.; Song, M. Genome-wide identification, and phylogenetic and expression profiling analyses, of XTH gene families in Brassica rapa L. and Brassica oleracea L. BMC Genomics 2020, 21, 782. [Google Scholar] [CrossRef]
- Wu, Z.; Cui, C.; Xing, A.; Xu, X.; Sun, Y.; Tian, Z.; et al. Identification and response analysis of xyloglucan endotransglycosylase/hydrolases (XTH) family to fluoride and aluminum treatment in Camellia sinensis. BMC genomics 2021, 22, 1–16. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, C.; Kong, D.; Cao, M.; Zhang, Q.; Tahir, M.; et al. Identification of High Tolerance to Jujube Witches’ Broom in Indian Jujube (Ziziphus mauritiana Lam.) and Mining Differentially Expressed Genes Related to the Tolerance through Transcriptome Analysis. Plants. [CrossRef]
- Choi, J. Y.; Seo, Y. S.; Kim, S. J.; Kim, W. T.; Shin, J. S. Constitutive expression of CaXTH3, a hot pepper xyloglucan endotransglucosylase/hydrolase, enhanced tolerance to salt and drought stresses without phenotypic defects in tomato plants (Solanum lycopersicum cv. Dotaerang). Plant cell reports 2011, 30, 867–877. [Google Scholar] [CrossRef]
- Zhu, X. F.; Shi, Y. Z.; Lei, G. J.; Fry, S. C.; Zhang, B. C.; Zhou, Y. H.; et al. XTH31, Encoding an in Vitro XEH/XET-Active Enzyme, Regulates Aluminum Sensitivity by Modulating in Vivo XET Action, Cell Wall Xyloglucan Content, and Aluminum Binding Capacity in Arabidopsis. The Plant Cell 2012, 24, 4731–4747. [Google Scholar] [CrossRef]
- Yan, J.; Huang, Y.; He, H.; Han, T.; Di, P.; Sechet, J.; et al. Xyloglucan endotransglucosylase-hydrolase30 negatively affects salt tolerance in Arabidopsis. Journal of experimental botany 2019, 70, 5495–5506. [Google Scholar] [CrossRef]
- Xu, P.; Fang, S.; Chen, H.; Cai, W. The brassinosteroid-responsive xyloglucan endotransglucosylase/hydrolase 19 (XTH19) and XTH23 genes are involved in lateral root development under salt stress in Arabidopsis. The Plant Journal 2020, 104, 59–75. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, D.; Johnson, K. L.; Hao, P.; Tuong, T.; Erban, A.; Sampathkumar, A.; et al. Cell wall modification by the xyloglucan endotransglucosylase/hydrolase XTH19 influences freezing tolerance after cold and sub-zero acclimation. Plant, Cell & Environment 2021, 44, 915–930. [Google Scholar]
- Li, M.; Guo, Y.; Liu, S.; Zhao, Y.; Pang, X.; Li, Y. Autotetraploidization in Ziziphus jujuba Mill. var. spinosa enhances salt tolerance conferred by active, diverse stress responses. Environmental and Experimental Botany 2019, 165, 92–107. [Google Scholar] [CrossRef]
- Wang, Y.; Feng, Y.; Yan, M.; Yu, J.; Zhou, X.; Bao, J.; et al. Effect of Saline–Alkali Stress on Sugar Metabolism of Jujube Fruit. Horticulturae. 2022. [CrossRef]
- Jiang, W.; Chen, L.; Han, Y.; Cao, B.; Song, L. Effects of elevated temperature and drought stress on fruit coloration in the jujube variety ‘Lingwuchangzao’ (Ziziphus jujube cv. Lingwuchangzao). Scientia Horticulturae 2020, 274, 109667. [Google Scholar] [CrossRef]
- Zhou, H.; He, Y.; Zhu, Y.; Li, M.; Song, S.; Bo, W.; et al. Comparative transcriptome profiling reveals cold stress responsiveness in two contrasting Chinese jujube cultivars. BMC Plant Biology 2020, 20. [Google Scholar] [CrossRef]
- Chen, P.; Li, J.; Ye, X.; Tan, B.; Zheng, X.; Cheng, J.; et al. Genome-wide identification of Ziziphus jujuba TCP transcription factors and their expression in response to infection with jujube witches’ broom phytoplasma. Acta Physiologiae Plantarum 2019, 41, 86. [Google Scholar] [CrossRef]
- Xue, C.; Zhang, L.; Li, H.; Liu, Z.; Zhang, Y.; Liu, M.; et al. The effector PHYL1JWB from Candidatus Phytoplasma ziziphi induces abnormal floral development by destabilising flower development proteins. Plant, Cell & Environment 2024, n/a. [Google Scholar] [CrossRef]
- Li, Q.; Hu, A.; Dou, W.; Qi, J.; Long, Q.; Zou, X.; et al. Systematic Analysis and Functional Validation of Citrus XTH Genes Reveal the Role of Csxth04 in Citrus Bacterial Canker Resistance and Tolerance. Frontiers in Plant Science 2019, 10. [Google Scholar] [CrossRef]
- Bhatti, M. H. T.; Han, S.; Sami, A.; Haider, M. Z.; Shafiq, M.; Ali, M.; et al. Genome-wide analysis of XTH gene family in cucumber (Cucumis sativus) against different insecticides to enhance defense mechanism. Plant Stress 2024, 13, 100538. [Google Scholar] [CrossRef]
- Li, M.; Zhang, C.; Hou, L.; Yang, W.; Liu, S.; Pang, X.; et al. Multiple responses contribute to the enhanced drought tolerance of the autotetraploid Ziziphus jujuba Mill. var. spinosa. Cell & Bioscience 2021, 11, 119. [Google Scholar] [CrossRef]
- Danso, B.; Ackah, M.; Jin, X.; Ayittey, D. M.; Amoako, F. K.; Zhao, W. Genome-Wide Analysis of the Xyloglucan Endotransglucosylase/Hydrolase (XTH) Gene Family: Expression Pattern during Magnesium Stress Treatment in the Mulberry Plant (Morus alba L.) Leaves. Plants. [CrossRef]
- Sarmiento-López, L. G.; López-Espinoza, M. Y.; Juárez-Verdayes, M. A.; López-Meyer, M. Genome-wide characterization of the xyloglucan endotransglucosylase/hydrolase gene family in Solanum lycopersicum L. and gene expression analysis in response to arbuscular mycorrhizal symbiosis. PeerJ 2023, 11, e15257. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Liu, Y.; Shen, Y.; Li, W. A surprising diversity of Xyloglucan Endotransglucosylase/Hydrolase in wheat: New in Sight to the roles in Drought Tolerance. International Journal of Molecular Sciences 2023, 24, 9886. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Xu, Z.; Ding, A.; Kong, Y. Genome-wide identification and expression profiling analysis of the xyloglucan endotransglucosylase/hydrolase gene family in tobacco (Nicotiana tabacum L.). Genes 2018, 9, 273. [Google Scholar] [CrossRef]
- Cheng, Z.; Zhang, X.; Yao, W.; Gao, Y.; Zhao, K.; Guo, Q.; et al. Genome-wide identification and expression analysis of the xyloglucan endotransglucosylase/hydrolase gene family in poplar. BMC Genomics 2021, 22, 804. [Google Scholar] [CrossRef]
- Zhu, J.; Tang, G.; Xu, P.; Li, G.; Ma, C.; Li, P.; et al. Genome-wide identification of xyloglucan endotransglucosylase/hydrolase gene family members in peanut and their expression profiles during seed germination. PeerJ 2022, 10, e13428. [Google Scholar] [CrossRef]
- Chen, C.; Chen, X.; Han, J.; Lu, W.; Ren, Z. Genome-wide analysis of the WRKY gene family in the cucumber genome and transcriptome-wide identification of WRKY transcription factors that respond to biotic and abiotic stresses. BMC plant biology 2020, 20, 1–19. [Google Scholar] [CrossRef]
- Feng, X.; Li, C.; He, F.; Xu, Y.; Li, L.; Wang, X.; et al. Genome-Wide Identification of Expansin Genes in Wild Soybean (Glycine soja) and Functional Characterization of Expansin B1 (GsEXPB1) in Soybean Hair Root. International Journal of Molecular Sciences. 2022. [CrossRef]
- Liu, Q.; Talbot, M.; Llewellyn, D. J. Pectin Methylesterase and Pectin Remodelling Differ in the Fibre Walls of Two Gossypium Species with Very Different Fibre Properties. PLOS ONE 2013, 8, e65131. [Google Scholar] [CrossRef]
- An, S. H.; Sohn, K. H.; Choi, H. W.; Hwang, I. S.; Lee, S. C.; Hwang, B. K. Pepper pectin methylesterase inhibitor protein CaPMEI1 is required for antifungal activity, basal disease resistance and abiotic stress tolerance. Planta 2008, 228, 61–78. [Google Scholar] [CrossRef]
- 43. Bethke, G.; Grundman, R. E.; Sreekanta, S.; Truman, W.; Katagiri, F.; Glazebrook, J. Arabidopsis PECTIN METHYLESTERASEs Contribute to Immunity against Pseudomonas syringae. Plant Physiology 2014, 164, 1093–1107. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Jia, L.; Yang, D.; Hu, Y.; Njogu, M. K.; Wang, P.; et al. Genome-Wide Identification, Phylogenetic and Expression Pattern Analysis of GATA Family Genes in Cucumber (Cucumis sativus L.). Plants, 2021. [Google Scholar] [CrossRef]
- Ma, Z.; Li, S.; Zhang, M.; Jiang, S.; Xiao, Y. Light intensity affects growth, photosynthetic capability, and total flavonoid accumulation of Anoectochilus plants. HortScience 2010, 45, 863–867. [Google Scholar] [CrossRef]
- An, Y.; Zhou, Y.; Han, X.; Shen, C.; Wang, S.; Liu, C.; et al. The GATA transcription factor GNC plays an important role in photosynthesis and growth in poplar. Journal of experimental botany 2020, 71, 1969–1984. [Google Scholar] [CrossRef] [PubMed]
- Abdulla, M. F.; Mostafa, K.; Aydin, A.; Kavas, M.; Aksoy, E. GATA transcription factor in common bean: A comprehensive genome-wide functional characterization, identification, and abiotic stress response evaluation. Plant Molecular Biology 2024, 114, 43. [Google Scholar] [CrossRef]
- Dordas, C. A.; Sioulas, C. Safflower yield, chlorophyll content, photosynthesis, and water use efficiency response to nitrogen fertilization under rainfed conditions. Industrial crops and products 2008, 27, 75–85. [Google Scholar] [CrossRef]
- Dhar, S.; Kim, J.; Yoon, E. K.; Jang, S.; Ko, K.; Lim, J. SHORT-ROOT controls cell elongation in the etiolated arabidopsis hypocotyl. Molecules and cells 2022, 45, 243–256. [Google Scholar] [CrossRef]
- Takeda, T.; Furuta, Y.; Awano, T.; Mizuno, K.; Mitsuishi, Y.; Hayashi, T. Suppression and acceleration of cell elongation by integration of xyloglucans in pea stem segments. Proceedings of the National Academy of Sciences 2002, 99, 9055–9060. [Google Scholar] [CrossRef]
- Cho, S. K.; Kim, J. E.; Park, J.-A.; Eom, T. J.; Kim, W. T. Constitutive expression of abiotic stress-inducible hot pepper CaXTH3, which encodes a xyloglucan endotransglucosylase/hydrolase homolog, improves drought and salt tolerance in transgenic Arabidopsis plants. FEBS letters 2006, 580, 3136–3144. [Google Scholar] [CrossRef]
- Bi, H.; Liu, Z.; Liu, S.; Qiao, W.; Zhang, K.; Zhao, M.; et al. Genome-wide analysis of wheat xyloglucan endotransglucosylase/hydrolase (XTH) gene family revealed TaXTH17 involved in abiotic stress responses. BMC Plant Biology 2024, 24, 640. [Google Scholar] [CrossRef]
- Jiang, Y.; Li, Y.; Lu, C.; Tang, Y.; Jiang, X.; Gai, Y. Isolation and characterization of Populus xyloglucan endotransglycosylase/hydrolase (XTH) involved in osmotic stress responses. International Journal of Biological Macromolecules 2020, 155, 1277–1287. [Google Scholar] [CrossRef]
- Wu, H.; Lian, B.; Lv, X.; Sun, M.; Wei, F.; An, L.; et al. Xyloglucan endotransglucosylase-hydrolase 22 positively regulates response to cold stress in upland cotton (Gossypium hirsutum L.). Industrial Crops and Products 2024, 220, 119273. [Google Scholar] [CrossRef]
- Tan, Y.; Zhan, H.; Chen, H.; Li, X.; Chen, C.; Liu, H.; et al. Genome-wide identification of gene family in Musa acuminata and response analyses of and xyloglucan to low temperature. Physiologia Plantarum 2024, 176, e14231. [Google Scholar] [CrossRef]
Figure 1.
Phylogenetic relationship analysis of the XTH proteins between Zizphus jujuba, Oryza sativa, Malus domestica, and Arabidopsis thaliana. Whole protein sequences of the XTH gene family were used for alignment using the MEGA X software. The phylogenetic tree was constructed using IQ-TREE 2 web tool using maximum likelihood with 1000 bootstrap replicates. Different colored branches correspond to distinct XTH subfamilies, and the XTH IDs of Arabidopsis, Apple, and Rice were assigned based on previous studies.
Figure 1.
Phylogenetic relationship analysis of the XTH proteins between Zizphus jujuba, Oryza sativa, Malus domestica, and Arabidopsis thaliana. Whole protein sequences of the XTH gene family were used for alignment using the MEGA X software. The phylogenetic tree was constructed using IQ-TREE 2 web tool using maximum likelihood with 1000 bootstrap replicates. Different colored branches correspond to distinct XTH subfamilies, and the XTH IDs of Arabidopsis, Apple, and Rice were assigned based on previous studies.
Figure 2.
Comparative analysis of the phylogenetics, exon-intron structures, and conserved motifs of the XTH family in Chinese jujube (ZjXTHs). (A) Motif composition models of 29 XTH proteins, with different motifs color-coded according to the legend. (B) Two conserved domains were identified and are represented in green and yellow. (C) The gene structures of ZjXTH were analyzed and visualized, including introns (black lines), exons (coding sequences, blue rectangles), and untranslated regions (UTRs, red rectangles).
Figure 2.
Comparative analysis of the phylogenetics, exon-intron structures, and conserved motifs of the XTH family in Chinese jujube (ZjXTHs). (A) Motif composition models of 29 XTH proteins, with different motifs color-coded according to the legend. (B) Two conserved domains were identified and are represented in green and yellow. (C) The gene structures of ZjXTH were analyzed and visualized, including introns (black lines), exons (coding sequences, blue rectangles), and untranslated regions (UTRs, red rectangles).
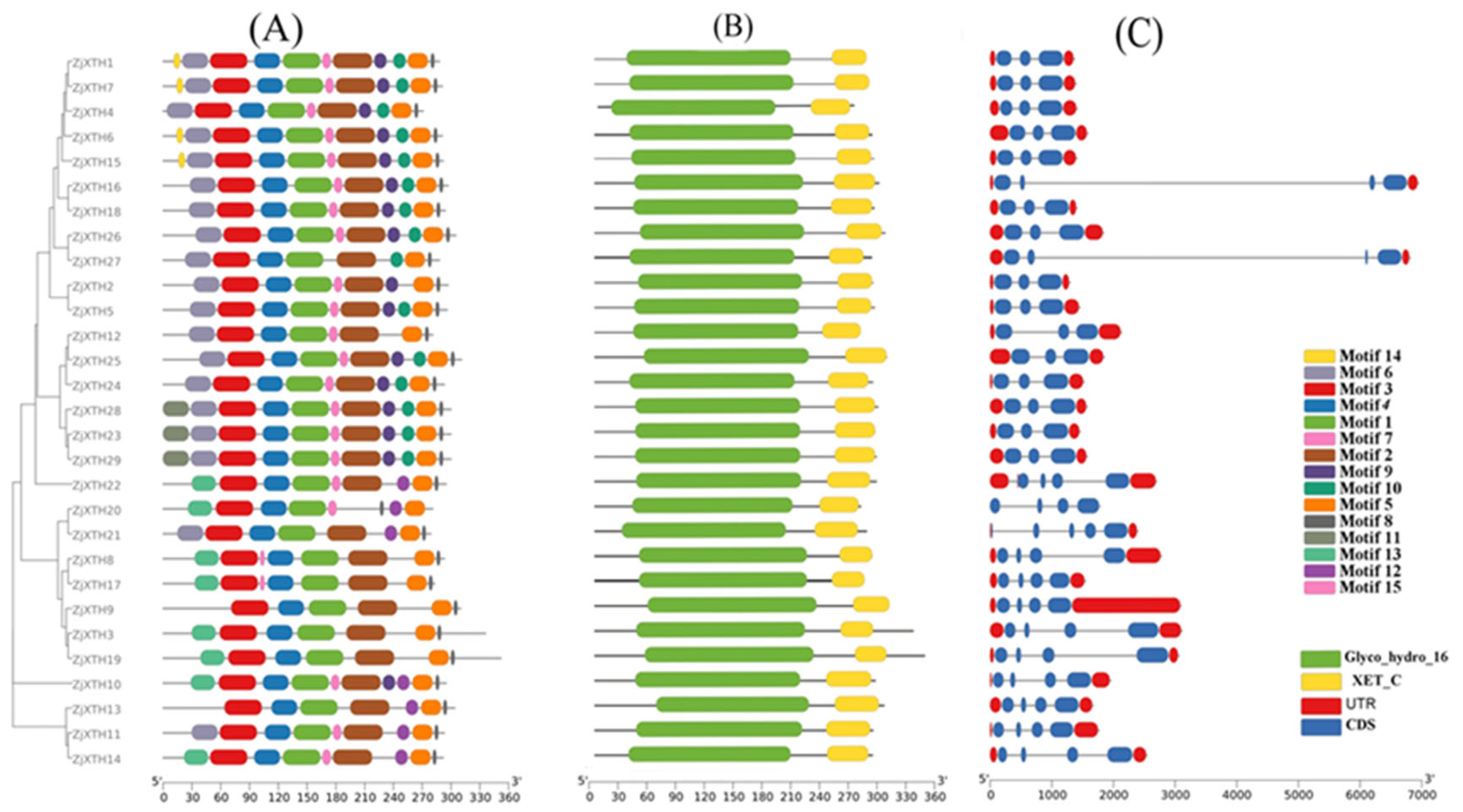
Figure 3.
Chromosomal localization and synteny analysis of ZjXTH proteins in the Chinease date genome. Genes IDs in black indicate the absence of collinearity, genes and lines colored in green indicate dispersed duplication; red indicates whole genome duplication, and blue-colored lines indicate transposed duplicated pairs (A). Protein-protein interaction analyses were performed on the String web tool and visualized using Cytoscape software. The network consists of various proteins represented as nodes, with interactions depicted by edges. Proteins labeled highlighted in yellow form key hubs with multiple interactions, suggesting their significant role in the network. Green nodes represent additional interacting proteins (B). Syntenic relationships of ZjXTH genes between Arabidopsis thaliana, Glycine max, and Oryza sativa. The browen lines in the background represent the collinear blocks within Ziziphus jujuba and other plant genomes, while the red lines highlight the syntenic ZjXTH gene pairs (C).
Figure 3.
Chromosomal localization and synteny analysis of ZjXTH proteins in the Chinease date genome. Genes IDs in black indicate the absence of collinearity, genes and lines colored in green indicate dispersed duplication; red indicates whole genome duplication, and blue-colored lines indicate transposed duplicated pairs (A). Protein-protein interaction analyses were performed on the String web tool and visualized using Cytoscape software. The network consists of various proteins represented as nodes, with interactions depicted by edges. Proteins labeled highlighted in yellow form key hubs with multiple interactions, suggesting their significant role in the network. Green nodes represent additional interacting proteins (B). Syntenic relationships of ZjXTH genes between Arabidopsis thaliana, Glycine max, and Oryza sativa. The browen lines in the background represent the collinear blocks within Ziziphus jujuba and other plant genomes, while the red lines highlight the syntenic ZjXTH gene pairs (C).
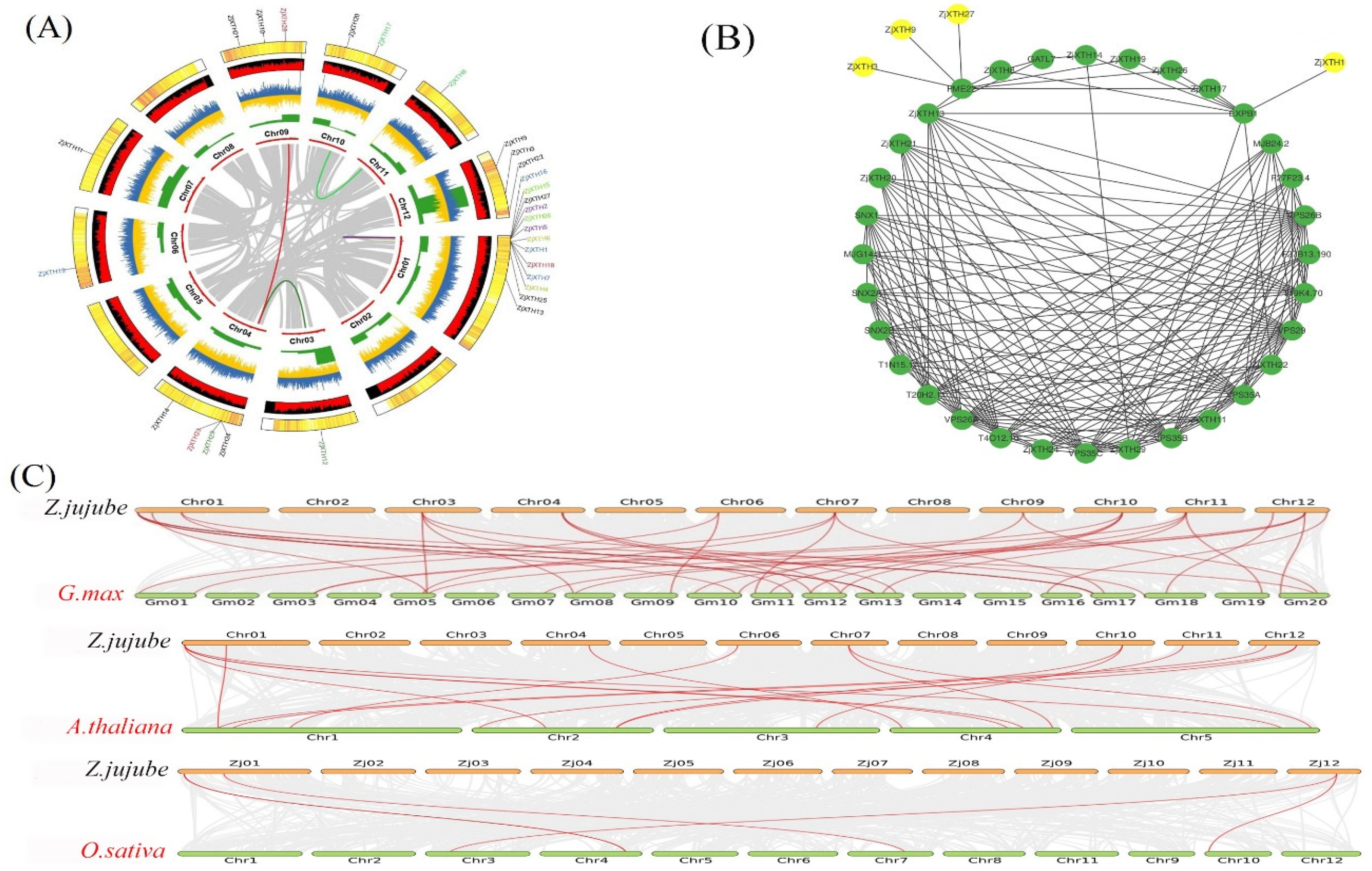
Figure 4.
Analysis of cis-regulatory elements (CREs) in the putative promoter region of ZjXTH genes using the PlantCARE database. (a) The number of predicted CREs located in the 2k bp upstream of the ZjXTH genes and the distribution of the three categories of CREs among the members of the ZjXTH gene family. (b) Venn diagram plot and pie chart showing the distribution of different functional categories of CREs identified in the ZjXTH promoter region.
Figure 4.
Analysis of cis-regulatory elements (CREs) in the putative promoter region of ZjXTH genes using the PlantCARE database. (a) The number of predicted CREs located in the 2k bp upstream of the ZjXTH genes and the distribution of the three categories of CREs among the members of the ZjXTH gene family. (b) Venn diagram plot and pie chart showing the distribution of different functional categories of CREs identified in the ZjXTH promoter region.
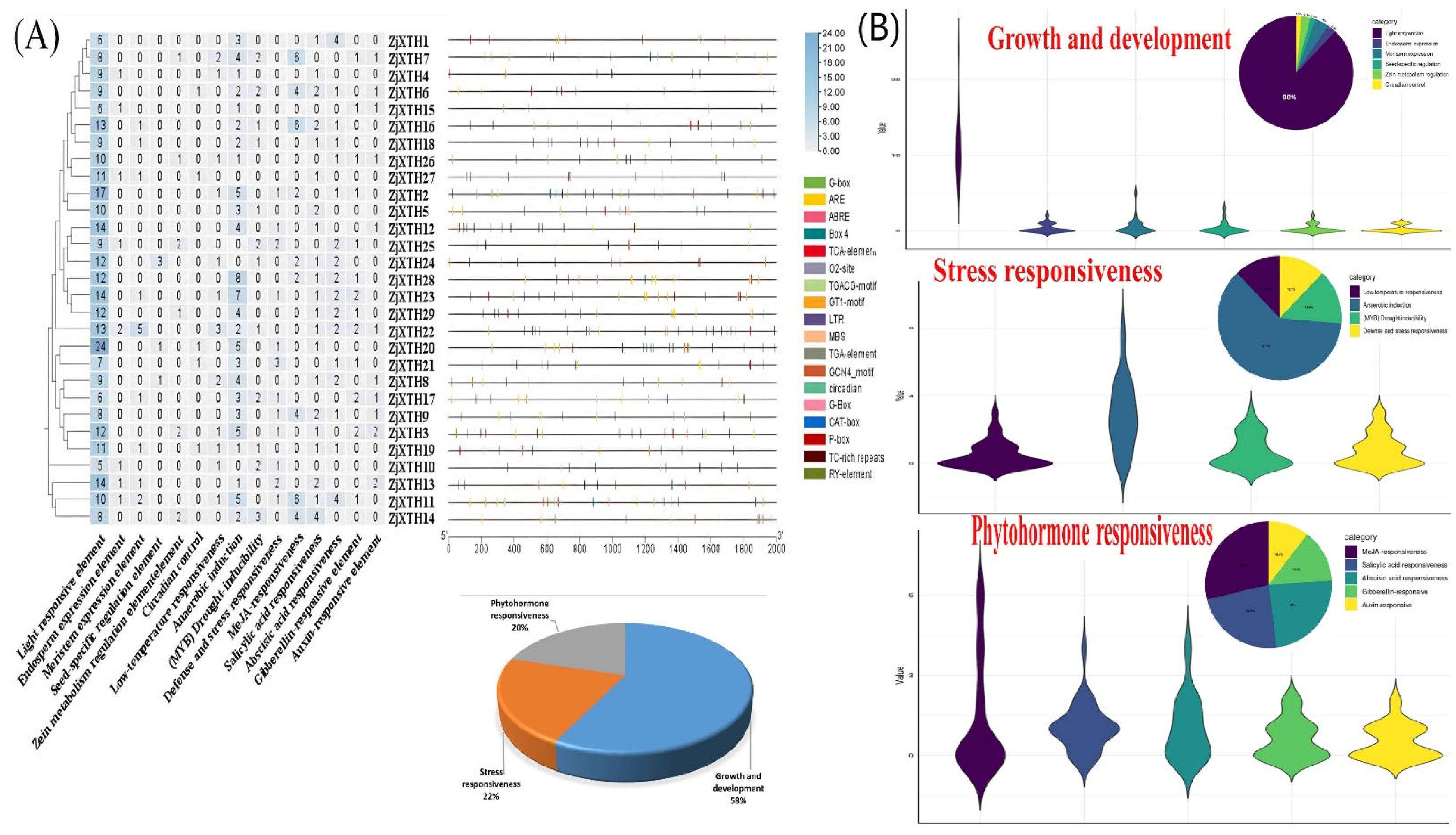
Figure 5.
Gene Ontology (GO) analysis was conducted on the ZjXTH gene family to assess its functional distribution across the genome. GO annotations were assigned to the ZjXTH gene sequences, categorizing them into three primary domains: molecular function, cellular component, and biological process. The resulting bar graph illustrates the proportional distribution of ZjXTH genes across these categories, providing insights into their potential roles in various biological pathways and cellular functions.
Figure 5.
Gene Ontology (GO) analysis was conducted on the ZjXTH gene family to assess its functional distribution across the genome. GO annotations were assigned to the ZjXTH gene sequences, categorizing them into three primary domains: molecular function, cellular component, and biological process. The resulting bar graph illustrates the proportional distribution of ZjXTH genes across these categories, providing insights into their potential roles in various biological pathways and cellular functions.
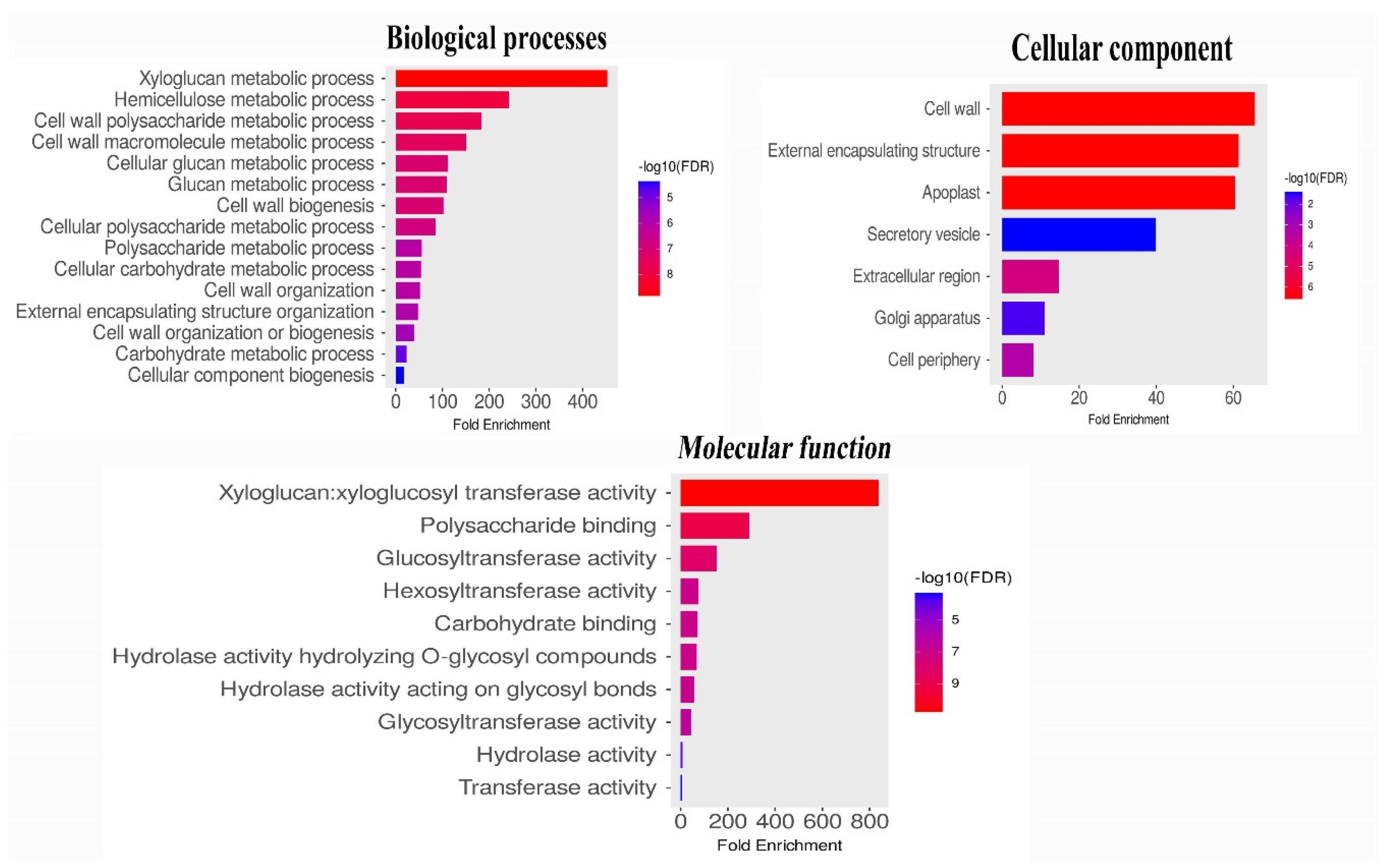
Figure 7.
Expression patterns of ZjXTHs of 29 differentially expressed genes in Z. jujuba. var. spinosa diploid and tetraploid seedlings, representing sensitive and tolerant types respectively, were used in a salinity treatment, gradually applied at 50, 100, and 150 mM NaCl. A deeper red indicates higher expression levels, while a deeper green represents lower expression levels.
Figure 7.
Expression patterns of ZjXTHs of 29 differentially expressed genes in Z. jujuba. var. spinosa diploid and tetraploid seedlings, representing sensitive and tolerant types respectively, were used in a salinity treatment, gradually applied at 50, 100, and 150 mM NaCl. A deeper red indicates higher expression levels, while a deeper green represents lower expression levels.

Figure 8.
Expression patterns of 29 differentially expressed ZjXTH genes were analyzed in diploid and tetraploid Z. jujuba. var. spinosa seedlings, representing sensitive and tolerant types, respectively, under PEG6000 concentrations of 5%, 10%, 15%, and 20% applied over 1-day intervals. The heatmaps represent the average FPKM values of the genes. A deeper red indicates higher expression levels, while a deeper green represents lower expression levels.
Figure 8.
Expression patterns of 29 differentially expressed ZjXTH genes were analyzed in diploid and tetraploid Z. jujuba. var. spinosa seedlings, representing sensitive and tolerant types, respectively, under PEG6000 concentrations of 5%, 10%, 15%, and 20% applied over 1-day intervals. The heatmaps represent the average FPKM values of the genes. A deeper red indicates higher expression levels, while a deeper green represents lower expression levels.

Figure 9.
Expression patterns of 29 differentially expressed ZjXTH genes were analyzed in the cold-sensitive cultivar ‘Dongzao’ and the cold-tolerant cultivar ‘Jinsixiaozao.’ A deeper red indicates higher expression levels, while a deeper green represents lower expression levels.
Figure 9.
Expression patterns of 29 differentially expressed ZjXTH genes were analyzed in the cold-sensitive cultivar ‘Dongzao’ and the cold-tolerant cultivar ‘Jinsixiaozao.’ A deeper red indicates higher expression levels, while a deeper green represents lower expression levels.

Figure 10.
Heatmap of 29 differentially expressed genes in Chinese jujube under biotic stress caused by jujube witches' broom phytoplasma (JWB). Cuimi (Z. mauritiana Lam.) and ‘Huping’ (Z. jujuba Mill.) scions were grafted onto the diseased ‘Jinsixiaozao’ (Z. jujuba Mill.) cultivar. Phenotypic observations were performed 21 weeks after grafting. The heatmaps represent the average FPKM values of the genes. A deeper red indicates higher expression levels, while a deeper green represents lower expression levels.
Figure 10.
Heatmap of 29 differentially expressed genes in Chinese jujube under biotic stress caused by jujube witches' broom phytoplasma (JWB). Cuimi (Z. mauritiana Lam.) and ‘Huping’ (Z. jujuba Mill.) scions were grafted onto the diseased ‘Jinsixiaozao’ (Z. jujuba Mill.) cultivar. Phenotypic observations were performed 21 weeks after grafting. The heatmaps represent the average FPKM values of the genes. A deeper red indicates higher expression levels, while a deeper green represents lower expression levels.


Table 1.
Characterization of XTH proteins.
| Gene Locations | Deduced Protein | ||||||||||||||||
| Gene Name | Protein ID | AA | bp | Chr | Signal peptide (SP) | LOC | Start | End | MW(KDa) | PI | II | AI | GRAVY | ||||
| ZjXTH1 | XP_015899780.1 | 288 | 865 | 1 | 22 | LOC107433023 | 2142963 | 2144333 | 32,104.7 | 5.24 | 38.45 | 60.31 | -0.439 | ||||
| ZjXTH2 | XP_015875377.2 | 297 | 899 | 1 | 34 | LOC107412146 | 994499 | 995798 | 33,451.7 | 5.4 | 34.94 | 77.41 | -0.292 | ||||
| ZjXTH3 | XP_015868033.1 | 336 | 1014 | 12 | 25 | LOC107405483 | 17889849 | 17892964 | 38,242.9 | 6.23 | 42.04 | 66.67 | -0.475 | ||||
| ZjXTH4 | XP_060672082.1 | 271 | 815 | 1 | _ | LOC107432095 | 2146026 | 2147442 | 30,453.67 | 5.26 | 36.72 | 55.79 | -0.577 | ||||
| ZjXTH5 | XP_015874851.4 | 296 | 877 | 1 | 30 | LOC107411692 | 1002556 | 1004014 | 33,595.72 | 6.2 | 40.42 | 69.49 | -0.401 | ||||
| ZjXTH6 | XP_015899683.3 | 291 | 878 | 1 | 25 | LOC107432937 | 2139826 | 2141413 | 32,396.09 | 5.42 | 38.32 | 64.71 | -0.374 | ||||
| ZjXTH7 | XP_015899503.3 | 292 | 867 | 1 | 25 | LOC107432789 | 2137263 | 2138660 | 32,320.89 | 5.42 | 35.55 | 62.71 | -0.398 | ||||
| ZjXTH8 | XP_015898435.1 | 293 | 881 | 11 | 19 | LOC107431919 | 7545976 | 7548758 | 33,835.31 | 9.38 | 43.23 | 62.25 | -0.499 | ||||
| ZjXTH9 | XP_048322840.2 | 310 | 928 | 12 | 27 | LOC107428620 | 6457682 | 6460783 | 34,870.81 | 8.13 | 50.18 | 77.97 | -0.137 | ||||
| ZjXTH10 | XP_015892932.3 | 295 | 883 | 9 | 26 | LOC107427100 | 15865101 | 15867060 | 34,701.11 | 8.56 | 37.48 | 66.1 | -0.513 | ||||
| ZjXTH11 | XP_015890447.3 | 293 | 882 | 7 | 30 | LOC107425034 | 14371805 | 14373572 | 33,370.77 | 6.94 | 40.81 | 72.9 | -0.31 | ||||
| ZjXTH12 | XP_015887252.2 | 281 | 842 | 3 | 29 | LOC107422327 | 13280316 | 13282451 | 32,220.62 | 9.17 | 36.22 | 74.95 | -0.3 | ||||
| ZjXTH13 | XP_015884034.2 | 304 | 911 | 1 | 33 | LOC107419753 | 16657548 | 16659222 | 35,202.27 | 4.75 | 39.29 | 64.74 | -0.518 | ||||
| ZjXTH14 | XP_048328785.2 | 292 | 875 | 4 | 24 | LOC107416878 | 25908816 | 25911364 | 33,164.31 | 5.41 | 32.69 | 69.42 | -0.275 | ||||
| ZjXTH15 | XP_015875279.3 | 292 | 882 | 1 | 27 | LOC107412056 | 979197 | 980607 | 32,874.68 | 5.82 | 38.58 | 64.49 | -0.418 | ||||
| ZjXTH16 | XP_060671190.1 | 297 | 891 | 1 | 30 | LOC107411876 | 969728 | 976681 | 33,09.22 | 6.81 | 41.14 | 68.32 | -0.367 | ||||
| ZjXTH17 | XP_015874702.3 | 283 | 851 | 10 | 19 | LOC107411602 | 17245584 | 17247133 | 31,912.58 | 6.79 | 48.08 | 64.13 | -0.494 | ||||
| ZjXTH18 | XP_048332201.1 | 294 | 884 | 1 | 29 | LOC107406936 | 2149644 | 2151057 | 33,056.15 | 8.56 | 35.37 | 69.69 | -0.296 | ||||
| ZjXTH19 | XP_048320582.2 | 352 | 1045 | 6 | 35 | LOC107403271 | 8140587 | 8143653 | 40,646.18 | 8.66 | 49.22 | 71.22 | -0.422 | ||||
| ZjXTH20 | XP_060668086.1 | 281 | 846 | 10 | 26 | LOC132799666 | 6971363 | 6973149 | 32,513.47 | 5.48 | 35.04 | 73.49 | -0.513 | ||||
| ZjXTH21 | XP_060676394.1 | 279 | 837 | 9 | _ | LOC125424114 | 6979194 | 6981594 | 32,552.23 | 6.19 | 35.98 | 60.07 | -0.713 | ||||
| ZjXTH22 | XP_015898303.1 | 295 | 887 | 12 | 24 | LOC107431811 | 25837449 | 25840155 | 34,115.81 | 8.62 | 36.67 | 67.76 | -0.451 | ||||
| ZjXTH23 | XP_015893429.2 | 300 | 903 | 4 | 32 | LOC107427557 | 6965305 | 6966770 | 33,878.08 | 6.95 | 42.65 | 72.5 | -0.288 | ||||
| ZjXTH24 | XP_024928685.3 | 293 | 882 | 4 | 25 | LOC107416347 | 6947614 | 6949140 | 33,775.19 | 9.47 | 36.74 | 68.26 | -0.465 | ||||
| ZjXTH25 | XP_048328992.1 | 311 | 936 | 1 | _ | LOC107412897 | 6046322 | 6048175 | 34,908 | 8.97 | 43.25 | 65.79 | -0.406 | ||||
| ZjXTH26 | XP_015875174.3 | 305 | 918 | 1 | _ | LOC107411968 | 1008911 | 1010755 | 34,241.17 | 4.98 | 43.09 | 67.51 | -0.283 | ||||
| ZjXTH27 | XP_060671181.1 | 288 | 867 | 1 | 26 | LOC107411514 | 982063 | 988879 | 32,731.87 | 5.97 | 34.92 | 74.51 | -0.29 | ||||
| ZjXTH28 | XP_048336175.2 | 300 | 903 | 9 | 32 | LOC125424038 | 23312053 | 23313630 | 34,157.58 | 8.54 | 39.98 | 73.13 | -0.293 | ||||
| ZjXTH29 | XP_048327183.1 | 300 | 903 | 4 | 32 | LOC125421763 | 6952934 | 6954513 | 33,92.92 | 8.54 | 42.4 | 71.5 | -0.305 | ||||
Table 2.
Inter-specific gene duplication analysis of ZjXTHs.
| Gene1 | Gene2 | Identity (%) | Ks | Ka | Ka/Ks | MAY |
|---|---|---|---|---|---|---|
| ZjXTH1 | ZjXTH7 | 94.77 | 2.0593 | 1.6380 | 0.79 | 1.68 |
| ZjXTH2 | ZjXTH5 | 68.75 | 2.1000 | 1.8905 | 0.9 | 1.72 |
| ZjXTH4 | ZjXTH6 | 91.54 | 0.6033 | 0.0675 | 0.11 | 4.94 |
| ZjXTH17 | ZjXTH8 | 71.84 | 0.7556 | 0.8956 | 1.18 | 6.19 |
| ZjXTH23 | ZjXTH28 | 93.67 | 0.0545 | 0.0605 | 1.11 | 4.46 |
| ZjXTH12 | ZjXTH29 | 66.9 | 2.4965 | 1.2844 | 0.51 | 2.04 |
| ZjXTH15 | ZjXTH26 | 80.51 | 0.2084 | 0.3212 | 1.54 | 1.7 |
| ZjXTH16 | ZjXTH18 | 86.81 | 0.1081 | 0.1524 | 1.41 | 8.85 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



