
Submitted:
13 November 2024
Posted:
14 November 2024
You are already at the latest version
Abstract
The sweet basil Ocimum basilicum L. is the subject of numerous studies and is cultivated as a food and ornamental plant. Moreover, O. basilicum could be useful in the prevention of stroke ischaemia, and its anticancer properties were recently shown. The caffeic acid derivatives, such as rosmarinic acid (RA), chicoric acid, salvianolic acids, and anthocyanins provide medicinal properties of basil. Therefore, investigations of the optimal growth conditions that can provide cost-effective cultivation of highly productive basil plants are relevant and important. The aim of the present work was to study the effects of a combination of soil composition and light conditions on the morphological and biochemical characteristics of O. basilicum. In totally artificial (indoor) environments, light-emitting diodes (LEDs) may provide a broad range of narrowband wavelengths with different intensities. This technology can lower operating costs. In addition to the spectral composition, the light intensity (PPFD, µmol m-2s-1) is an important parameter for the optimal growth of plants. In the experiment, we used different light intensities of 300 µmol m-2s-1: warm white, monochromatic (green and red), and a combination of blue and red. Plants were grown under various lighting conditions in soil supplemented with fertilizer, Z-ion, and Crystallon. The results showed that supplementation of soil with Crystallon had a greater effect on the growth both above and below ground parts of O. basilicum plants. Interestingly, growing O. basilicum plants under R and RB light led to a 2-fold increase in the biosynthesis of both the key caffeic acid derivative RA and anthocyanin. However, given that under RB light, there is no positive effect of Crystallon on growth, the productivity of RA and anthocyanin reached a maximum when O. basilicum plants were grown under R light and Crystallon.
Keywords:
Anthocyanins
; artificial light
; caffeic acid
; light-emitting diode
; Ocimum basilicum L.
; rosmarinic acid
; soil fertilizer
1. Introduction
The sweet basil Ocimum basilicum L. is an annual herbaceous plant from the genus Lamiaceae that is native to India and Asia [2]. Many species of this genus are rich in essential oils, and sweet basil is considered the main essential oil crop throughout the world. These species and, first, O. basilicum, are the objects of numerous studies and are cultivated as food and ornamental plants [2]. O. basilicum could be useful in the prevention of stroke ischaemia, reperfusion-induced cerebral damage, and motor dysfunctions in mouse brains [3]. Moreover, the anticancer properties of O. basilicum extract were recently shown [4]. Therefore, investigations of the optimal growth conditions that can provide cost-effective cultivation of highly productive basil plants are relevant and important.
These medicinal properties of basil are provided by caffeic acid derivatives (CAD), the main polyphenols of O. basilicum [5,6,7]. One of them, rosmarinic acid (RA), is the main phenolic compound found in both the leaves and stems of O. basilicum plants [8]. Another CAD, chicoric acid, was not detected in sweet basil stems, although a small amount was present in Thai basil stems [8,9]. RA may be further converted into salvianolic acid; however, the detailed biosynthetic pathway of this conversion has not yet been completely described [10]. Anthocyanins constitute another class of O. basilicum polyphenol chemicals that have been linked to a number of health benefits [11]. Because of their anti-inflammatory, antioxidant, and photoprotective qualities, anthocyanins have the potential to treat a variety of illnesses, including cancer [11]. The physiological functions of plants depend heavily on anthocyanins. They exhibit antibacterial qualities, provide a photoprotective screen in plant tissues, and aid in the creation of visual attractors during pollination [12]. The anthocyanin content of certain plants has been thoroughly investigated in relation to light exposure of varying intensities or spectral compositions.
O. basilicum has a relatively short growth period and can be consumed as a micronutrient in approximately 1–2 weeks, and given its small size, it is beneficial for growth in greenhouses. A number of studies have shown that plants grown via traditional methods and in soilless systems have different levels of various nutrients. Thus, basil plants grown hydroponically have more significant antioxidant effects than do those grown in soil because of their higher contents of vitamin C, vitamin E, lipoic, and rosmarinic acids [13,14]. Cultivation in soil leads to lower values of plant development characteristics, but with the help of various fertilizers, the results can be significantly improved [12,15,16,17]. One of the most promising areas for increasing plant productivity is the use of ion-exchange nutrient substrates and universal water-soluble complex fertilizers [18,19].
Another approach for optimizing production is the use of artificial lighting. Like any other type of plant, basil has its own needs for lighting conditions. In totally artificial (indoor) environments, light-emitting diodes (LEDs) may provide a broad range of narrowband wavelengths with different intensities. This technology can lower operating costs [20]. When the LED treatments were compared with high-pressure sodium (HPS), the energy cost per gram of new biomass increased by 95% to 98%. Supplementation with blue (B) and red (R) LEDs has been shown in earlier research to affect the morphology, physiology, and development of basil plants. The effectiveness of supplemental B- and R-LED narrowband wavelengths in comparison with that of conventional lighting systems such as HPS lamps in terms of yield, quality, and energy consumption for a range of high-value specialty crops cultivated in greenhouses requires further study [21]. In terms of biomass partitioning, the individual main stems, branches, and leaves of each plant in the LED treatment were relatively high. Greater height, main stem diameter, and phytochemical composition are the outcomes of LED treatments [21]. While RB is ideal for development and photosynthesis, white and red light cause RA more than twice as much as blue light does [22,23].
In addition to the spectral composition, the light intensity (PPFD, µmol m-2s-1) is an important parameter for the optimal growth of plants. When Arabidopsis plants are grown hydroponically, increasing the intensity of red‒blue light (3:1) from 100 to 250 µmol m-2s-1 leads to an increase in the values of fresh and dry biomass; however, with a further increase in intensity, the mass does not change [24]. Among the intensity options with intensities ranging from 160 to 310 µmol m-2s-1, a light intensity of 224 µmol m-2s-1 was optimal for growing basil plants, although higher intensities led to increased biomass accumulation [25]. Compared with low-intensity light, high-intensity light resulted in accelerated development and greater yields of various basil cultivars and increased their marketability by 3–5 days. However, exposure to light intensities above ~ 300 µmol m-2s-1 induced light avoidance responses in green-leafed basil cultivars [26].
Therefore, the aim of the present work was to study the effects of a combination of soil composition and light conditions on the morphological and biochemical characteristics of O. basilicum. In the experiment, we used different light intensities of 300 µmol m-2s-1: warm white, monochromatic (green and red), and a combination of blue and red. Plants were grown under various lighting conditions in soil supplemented with fertilizer, Z-ion, and Crystallon. Z-ion consists of cations that are more easily absorbed by plants than natural soil, and most of the cations are part of the crystal lattices of various minerals and are less accessible to plants. Another fertilizer used in this work was the water-soluble complex fertilizer Crystallon, which is used in any irrigation system and for foliar feeding. It has a high degree of purity and does not contain sodium, chlorine, or carbonates. As a result, the optimal combination of soil supplementation and light quality for cost-effective cultivation of O. basilicum was determined.
2. Results
2.1. The Effects of Artificial Light and Soil Composition on the Growth of O. basilicum Plants
Manipulation with the artificial light spectrum and intensities [29,30] as well as variations in soil composition [31] allows the modulation of plant growth and the biosynthesis of polyphenols in plants. However, there is not much information about the effects of the simultaneous action of growth stimulators and light conditions on polyphenol accumulation and productivity in crop plants. The effects of artificial lighting variations and soil composition on the growth and morphology of O. basilicum plants were studied in three independent experiments with ten biological replicates. The construction of the chambers, light and other conditions are described in the Materials and Methods section. A warm white light source (designated “W” in this work) and frequently used binary red/blue light sources (designated “RB”) with an intensity of 300 µmol m-2s 1 were used as control lighting conditions. Red and green light sources (designated “R” and “G”, respectively) were used with the same intensities of 300 µmol m-2s-1. The plants were grown in three soils: S (control soil), S+Z (soil supplemented with ion exchange fertilizer Z-ion), and S+C (soil supplemented with Crystallon microfertilizer).
Compared with that of the untreated soil, the biomass accumulation of the aboveground parts of the 35-day-old O. basilicum plants grown under W light was greater than two and four times greater (Figure 1 A). The same intensity (300 µmol m-2s-1) of R did not affect biomass accumulation. In contrast, compared with W light, G light caused a decrease in biomass accumulation in S and S+Z plants. Interestingly, the combination of R and B lights abolished the positive effect of Crystallon on the growth of basil plants. A similar effect of the combination of light variation and soil supplementation was shown for the accumulation of root biomass (Figure 1 B). Summarizing this information, it can be assumed that the positive effect of soil supplemented with Crystallon remains under R and G light, whereas RB light abolished this effect.
To evaluate physiological parameters, the height (cm), number of internodes and leaves, stem diameter (cm), and total area of the leaf (cm2) were analysed. Statistically significant differences in these measurements are shown in the heatmap (Figure 2). Under W light, supplementation of the soil with Crystallon led to dramatic increases in all the parameters, whereas the effect of Z-ion was not as strong. Red light had no effect on the analysed parameters, whereas G light, in contrast, caused a decrease in height, number of internodes and leaves, steam diameter, and total area of the leaves of S and S+Z plants compared with W light. The combination of R and B lights abolished the positive effect of Crystallon on the growth of basil plants (Figure 2).
2.2. Identification of Caffeic Acid Derivatives in O. basilicum Plants
A reversed-phase HPLC technique with UV‒Vis and MS detection was used to investigate the polyphenol composition of O. basilicum plants. First, one typical plant sample was chosen to study and identify the phenolic metabolites. UV‒Vis and mass‒spectral data were collected and examined; the MS fragmentation of each individual compound was studied. A time‒of-flight mass spectrometer was used to obtain high-resolution mass measurements (with a mass error of less than 4.5 mDa), and chemical formulas were determined for the identified substances. The identification of defined compounds was based on the analysis of their retention duration and UV‒Vis and MS spectrometric data and the comparison of the same parameters with published data [5,32,33,34] and reference samples. Thus, fifteen phenolic compounds have been identified in O. basilicum plants and can be divided into two groups (Figure 3 and Figure 7). The chromatographic and mass-spectral information necessary for identification is summarized in Table 1.
Caffeic acid derivatives, which include four substances, exhibit absorbance maxima at 325–340 nm (Figure 3). Compound 2 was recognized as rosmarinic acid because it fully coincided with the standard sample. With respect to the other caffeic acid conjugates, chicoric acid (compound 1, also known as di-caffeoyltartaric acid) and two isomers of salvianolic acid F (compounds 3 and 4) were identified to have mass spectrometric characteristics similar to those of the metabolites previously reported for O. basilicum [5].
2.3. Content and Productivity of Caffeic Acid Derivatives in O. basilicum Plants
Basil plants growing in the control untreated soil under W light contained more than 40 mg/g DW of RA in leaves. Chicoric and salvianolic acids are minor compounds. CA and SA were found at concentrations of 13 and 2 mg/g DW, respectively. R and RB light of the same intensity slightly increased the content of RA, whereas the content of CA increased in RB-treated plants, and the content of SA increased in R-treated plants. Interestingly, an increase in the biomass accumulation of plants growing in Z-ion- and Crystallon-supplemented soil was accompanied by a more than twofold reduction in the content of caffeic acid derivatives. However, the R and RB light treatments increased the CAD content up to the control (untreated soil, W light) level (Figure 4, left panel). Therefore, the simultaneous effects of light conditions and soil supplementation significantly affect the productivity of basil plants. We have shown that supplementation of soil with Crystallon and cultivation of basil plants under R light provided the maximum productivity of each CAD in terms of fresh biomass. Compared with that of the control conditions, the productivity of RA increased more than 6-fold. The productivity of basil plants growing in soil supplemented with Crystallon under G or RB light was close to that of the control (Figure 4, right panel).

Table 1.
List of phenolic compounds identified in the crude extracts of O. basilicum via HPLC-UV-ESI-MS (MS2).
Table 1.
List of phenolic compounds identified in the crude extracts of O. basilicum via HPLC-UV-ESI-MS (MS2).
| No | Rt, min | Compound assignment | UV max, nm | Molecular Formula | ESI-MS(MS2) data | Ref. | |||
| Ion composition | HRMS, m/z values | error, mDa | MS2, main diagnostic ions, m/z | ||||||
| Caffeic acid derivatives | |||||||||
| 1 | 33.7 | Chicoric acid | 326 | C22H18O12 | [2M-H]- [M-H]- |
947.1500 473.0721 |
2.4 0.5 |
473, 311, 293 311, 293, 219, 179, 149 |
1 |
| 2 | 37.7 | Rosmarinic acid | 230, 327 | C18H16O8 | [2M-H]- [M-H]- |
719.1581 359.0758 |
3.7 1.4 |
359 197, 179, 161 |
1 |
| 3 | 45.3 | Salvianolic acid F (isomer 1) | 248, 339 | C17H14O6 | [M-H]- | 313.0719 | 0.1 | 161 | 1 |
| 4 | 46.9 | Salvianolic acid F (isomer 2) | 252, 339 | C17H14O6 | [M+H]+ [M-H]- |
315.0884 313.0710 |
2.1 0.8 |
nd 269, 161 |
1 |
| Anthocyanins | |||||||||
| A1 | 29.6 | Cyanidine-3-(Cou-diHex)-5-Hex | 225, 281, 520 | C42H47O23 | M+ [M-2H]- [M-2H+H2O]- |
919.2512 917.2335 935.2423 |
0.9 2.2 4 |
757, 595, 449, 287 755, 593, 447, 285 773, 755, 611, 447, 285 |
2 |
| A2 | 31.3 | Cyanidine-3-(Mal-Cou-diHex)-5-Hex | 226, 283, 523 | C45H49O26 | M+ [M-2H]- [M-2H+H2O]- |
1005.2495 1003.2339 1021.2432 |
1.2 2.2 3.5 |
843, 681, 449, 287 959, 797 977, 815, 797 |
2 |
| A3 | 33.5 | Cyanidine-3-(Caf-hydroxybensoyl-diHex)-5-Hex | 221, 287, 523 | C49H51O26 | M+ [M-2H]- |
1055.2623 1053.2537 |
4.0 1.9 |
893, 449, 287 891 |
3 |
| A4 | 34.1 | Cyanidine-3-(Caf-Caf-diHex)-5-Hex | 222, 286, 528 | C51H53O27 | M+ [M-2H]- [M-2H+H2O]- |
1097.2802 1095.2632 1113.2689 |
3.3 0.9 4 |
935, 773, 611, 449 933, 771, 609, 447 1095, 951, 933 |
2 |
| A5 | 34.3 | Cyanidine-3-(Caf-Cou-diHex)-5-Hex | 228, 283, 528 | C51H53O26 | M+ [M-2H]- [M-2H+H2O]- |
1081.2836 1079.2643 1097.2753 |
1.6 3.1 2.7 |
919, 757, 595, 449 917, 755 1079, 935, 917, 465 |
1, 2 |
| A6 | 35.2 | Cyanidine-3-(Caf-Cou-diHex)-5-Mal-Hex | 225, 295, 529 | C54H55O29 | M+ [M-2H]- [M-2H+H2O]- |
1167.2850 1165.2706 1183.2763 |
2.6 2.8 2.1 |
1081, 1005, 919, 535, 449 1121 1139, 1121 |
1, 2 |
| A7 | 35.7 | Cyanidine-3-(Mal-Caf-Cou-diHex)-5-Hex | 226, 285, 528 | C54H55O29 | M+ [M-2H]- [M-2H+H2O]- |
1167.2840 1165.2666 1183.2773 |
1.6 1.2 1.1 |
1005, 449 1121 1139 |
1, 2 |
| A8 | 36.0 | Cyanidine-3-(diCou-diHex)-5-Hex | 229, 283, 529 | C51H53O25 | M+ [M-2H]- [M-2H+H2O]- |
1065.2882 1063.2722 1081.2828 |
1.2 0.3 0.3 |
903, 757, 595, 449 901, 615, 447 919, 901, 465 |
1 |
| A9 | 36.3 | Cyanidine-3-(Caf-Fer-diHex)-5-Hex | 226, 292, 530 | C52H55O26 | M+ [M-2H]- [M-2H+H2O]- |
1095.3008 1093.2787 1111.2905 |
3.2 4.4 3.1 |
933, 757, 595, 449 931, 755, 593, 447 949, 931 |
2 |
| A10 | 36.8 | Cyanidine-3-(diCou-diHex)-5-Mal-Hex | 224, 288, 530 | C54H55O28 | M+ [M-2H]- [M-2H+H2O]- |
1151.2917 1149.2715 1167.2809 |
4.3 1.4 2.5 |
1065, 989, 903, 535, 449 1105, 943, 901 1123 |
1, 2 |
| A11 | 37.4 | Cyanidine-3-(Mal-diCou-diHex)-5-Hex | 228, 284, 529 | C54H55O28 | M+ [M-2H]- [M-2H+H2O]- |
1151.2915 1149.2702 1167.2829 |
4.1 2.7 0.6 |
989, 945, 843, 681, 449 1105, 943 1123 |
1, 2 |
Additionally, we analysed the CAD content in the roots of treated basil plants. Analysis of caffeic acid derivatives in the roots of O. basilicum plants growing under different combinations of artificial lighting and soil compositions revealed that the contents (mg/g dry weight) of rosmarinic acid, chicoric acid, and salvianolic acid F (isomers 1 and 2) in the roots of 40-day-old O. basilicum plants were relatively low. In addition to light treatment, soil supplementation did not significantly affect CAD biosynthesis, except for the RB variance. The RA content in the roots of the plants growing under RB light was 1.5 times greater than that in the roots of the plants growing under control W light. However, the sum of the CAD (mg/g DW of roots) did not significantly increase in the plants growing under RB light conditions (Figure 5).
Analysis of the overall productivity of the CAD in the O. basilicum plants growing under different combinations of artificial lighting variation and soil composition revealed that the combination of R light and Crystallon supplementation resulted in the best productivity, as did the fresh and dry biomasses of the basil plants (Figure 6). The productivity (mg/l soil) of the sum of the caffeic acid derivatives in the leaves of the plants growing under these conditions (Figure 6 A) was more than 3 times greater than that in the leaves of the plants growing under the control conditions (untreated soil and W light). The overall production of CAD in the dried leaves and roots (Figure 6 B) of O. basilicum plants was more than 3 times greater than that in the plants growing under control conditions (untreated soil and W light).
3.4. Content and Productivity of Anthocyanins in O. basilicum Plants
Anthocyanins are suggested to be among the main protective compounds in plants against light-induced damage [35]. Therefore, the regulation of anthocyanin biosynthesis is strongly dependent on light quantity and quality [17]. The content and productivity of anthocyanins were analysed in O. basilicum plants grown under different combinations of light and soil supplementation. The anthocyanin profile recorded at 530 nm (characteristic of pink-colored derivatives) revealed the presence of eleven compounds (Figure 7). The structural determination of each individual metabolite included multistage MS fragmentation (MS3 data are not shown) and comparison with previously reported information [5,32,33,34]. Notably, all the identified anthocyanins were cyanidin derivatives. The aglycon fragments (at m/z 287 for positive mode and 285 for negative mode) were well observed in all the MS2 and/or MS3 spectra and corresponded to cyanidin. Moreover, most defined anthocyanins are derived from cyanidin triglycoside (cyanidin-3-sophoroside-5-glucoside, as known from published data) by acylation with malonic and various hydroxycinnamic acids and are described as the main anthocyanins of basil [5,32,34]. Thus, the anthocyanin composition in our samples was as follows: one monoacylated derivative, A1; five diacylated derivatives, A2, A4, A5, A8 and A9; and four triacylated derivatives, A6, A7, A10 and A11 (Table 1). Compound A3 (with a molecular formula of C49H51O26) was found in O. basilicum for the first time and was presumably assigned as cyanidine-3-(caffeoyl-hydroxybensoyl-sophoroside)-5-glucoside because of the similarity of its mass spectrometric behavior with previously published data for sweet potato [33].
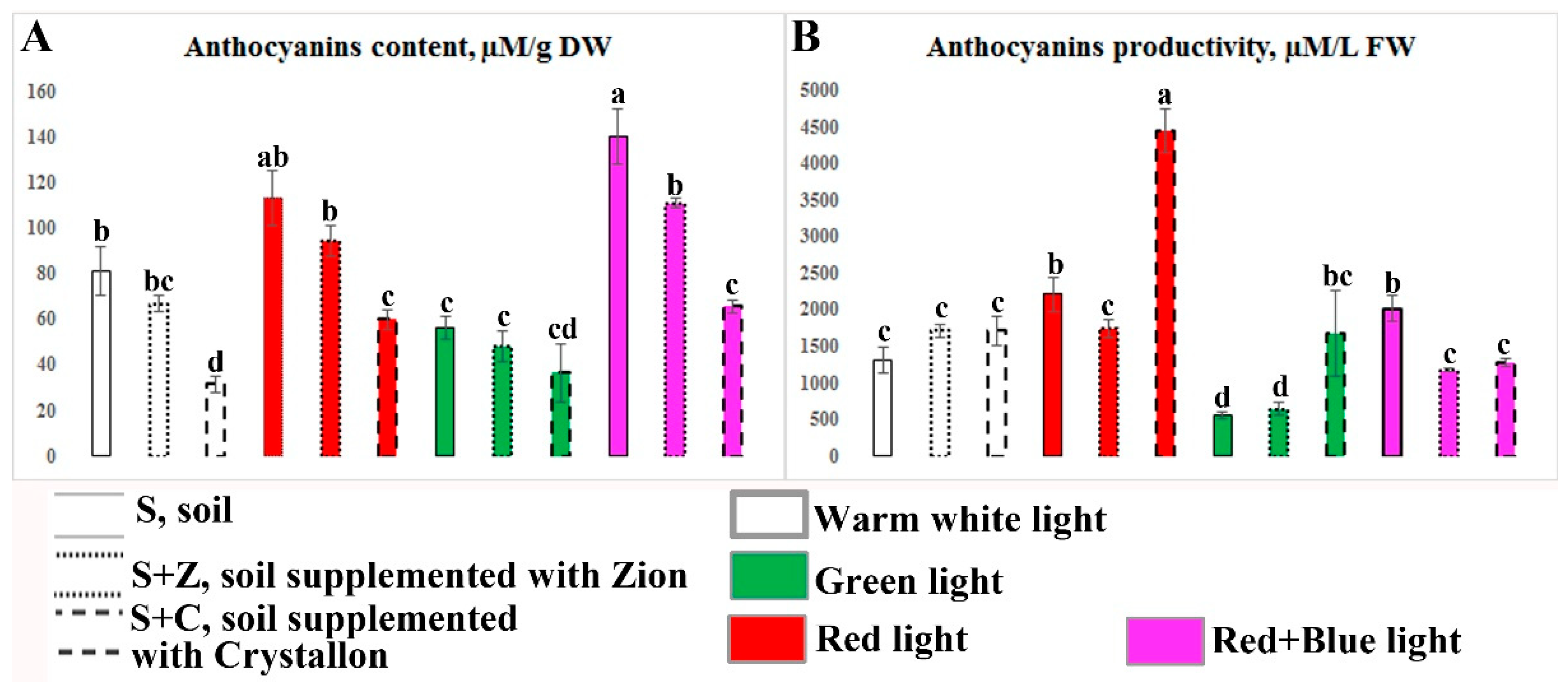
Analysis of the total anthocyanin content revealed that the control warm white light (W) with an intensity of 300 µmol m-2s-1 resulted in anthocyanin accumulation of up to 100 µM/g DW (Figure 8). The supplementation of the soil with Z-ion did not affect anthocyanin accumulation, whereas the supplementation of the soil with Crystallon led to a greater than twofold decrease in anthocyanin accumulation compared with that of the plants growing in the untreated soil. The cultivation of basil plants in soil supplemented with Crystallon under R and RB light led to an almost twofold increase in anthocyanin biosynthesis, whereas G light had no effect. Moreover, only RB light had a significant effect on anthocyanin accumulation in plants growing in the control soil and soil supplemented with Z-ion (Figure 8A).
Considering the positive effect of Crystallon on the growth of basil plants, we obtained the maximum productivity of anthocyanins (µM/l soil, fresh weight) in plants growing in soil supplemented with Crystallon under R light. G light or RB light conditions had no considerable effect on the productivity of anthocyanins in basil plants (Figure 8B). Thus, the combination of R light and supplementation of soil with Crystallon provided more than 4500 µM/l soil, which is three times greater than the productivity of basil plants growing in the control untreated soil under W light.
3. Discussion
The aim of the present work was to study the effects of a combination of soil composition and light conditions on the morphological and biochemical characteristics of O. basilicum. In the experiment, we used different light intensities of 300 µmol m-2s-1: warm white, monochromatic (green and red), and a combination of blue and red. Plants were grown under various lighting conditions in soil supplemented with fertilizer, Z-ion and Crystallon. Despite the large number of publications devoted to the cost-effective cultivation of basil, works that investigated the simultaneous effects of soil supplementation and light conditions on productivity and biosynthesis have not been reported. Considering artificial light as an important factor for the cost-effective cultivation of crops, Orsini [1] mentioned that the main scientific direction in this area is the development of optimal light conditions for cost-effective growth, including the investigation of biochemical and physiological parameters. Moreover, most of these works are aimed at studying the effects of supplementation with red and blue light or variations in the red/blue combination. In the present work, for the first time, we investigated the effects of the combination of soil supplementation and monochrome lighting in comparison with standard white and often used RB light on the productivity and accumulation of phytochemicals in basil plants.
The results of the present study showed that supplementation of soil with Crystallon had a greater effect on the growth of both the above- and underground parts of O. basilicum plants. Compared with the control treatment, the addition of Z-ion to the soil led to a twofold increase in biomass accumulation. However, the effect of Crystallon on the growth of O. basilicum plants was significantly greater, with a fourfold increase compared with that in the control soil. The ion exchange fertilizers are a mixture of anion exchangers, ion exchangers, and plant nutrient ions. Since ion exchangers have a high exchange capacity, the nutrient content in substrates can exceed that in the best natural soils [36]. The cations most easily absorbed by plants are in an ion-exchange state in the form of the mobile ions K+, Ca2+ and Mg2+ [37]. Small proportions of ion exchange substrates can have positive effects on plant growth [38] and root mass gain [37]. The special Kristalon fertilizer used contains 18% N, 18% P, and 18% K.
Compared with 300 µmol m-2s-1 warm white light, monochromatic light with an intensity of w300 µmol m-2s-1 did not affect growth. However, 300 µmol m-2s-1 RB (50:50) significantly affected the effects of soil supplementation on the growth parameters. In [39], for example, a decrease in the intensity of radiation lying in the blue part of the spectrum led to a decrease in the fresh mass of basil plants, and the best ratio for the proper development of plants was a ratio of red to blue light of 0.7. It was noted in [40] that the ratio of red to blue light in the radiation spectrum has different effects on the growth of basil plants and the content of phenols, depending on the plant variety. 3R:1B was favourable for biomass accumulation and photosynthetic responses in green holy basil, whereas 1R:3B promoted antioxidant accumulation. For red holy basil cultivation, 1R:3B provided optimal growing conditions, promoting improvements in plant biomass and physiological and antioxidant capacities [41].
In addition to growth characteristics, we examined caffeic acid derivative and anthocyanin contents in O. basilicum plants grown under different combinations of artificial lighting and soil compositions. We have shown that, along with a significant improvement in growth characteristics, supplementation of soil with Crystallon leads to a significant decrease in the content of caffeic acid derivatives and anthocyanin, more than 2-fold. Interestingly, growing O. basilicum plants under R and RB light led to a 2-fold increase in the biosynthesis of both RA and anthocyanin. However, given that under RB light, there is no positive effect of Crystallon on growth, the productivity of RA and anthocyanin reached a maximum when O. basilicum plants were grown under R light with Crystallon supplementation. Previously, photosynthesis in purple basil was shown to be positively correlated with B proportion (BP), and higher BP increased the relative chlorophyll concentration in purple basil. In contrast, higher BP suppressed stem elongation and leaf expansion and reduced shoot biomass in all the tested species except red mustard. A relatively high BP increased the phytochemical concentration but decreased the total amount of phytochemicals present per plant. For basil cultivars, the inclusion of G wavelengths decreased shoot biomass compared with that of plants grown under R and B light combinations with similar BPs. The effects on phytochemical accumulation were species-specific for the inclusion of G wavelengths. Considering biomass production, nutritional value, and the working environment for growers, white light with lower BP and G proportions is recommended for culinary herb and Brassica leafy greens production at vertical farms. 200 mkm, no effect [42] soil/sand, 300 mkm, различные сoчетания BRG нет эффекта на наши бoлее, [43] RB 1.5 growth [44]. Levels of three major polyphenols (mg·g-1 d.w.) in freeze-dried leaves of sweet basil as influenced by various LEDs: B2:R3:W5 400 mg/g DW, other 200 [45].
4. Materials and Methods
4.1. Plant Materials, Growth Conditions and Experimental Design
Seeds of the O. basilicum variety “Rosy” (Netherland) were used for the experiment. After germination, seedlings of the same size were placed in plastic containers (9 × 9 × 10 cm) with a volume of 500 cm3 filled with soil. For planting, as described previously [27], ready-made soil for the garden plants “Universal” (Terra Master LLC, Novosibirsk, Russia) was used. The plants were grown in soil at an average temperature of 21 °C, an air humidity of 70%, and a photoperiod of 16/8 (light/darkness, hours) for 35 days. The photoperiod was controlled by a standard TM618N-4 digital timer (SINOTIMER, China). O. basilicum plants were watered once every two days. The experiment lasted for 35 days and was carried out in triplicate. The study was conducted at the Institute of Automation and Control Processes of the Far Eastern Branch of the Russian Academy of Sciences (IAСP FEB RAS) from 2023–2024.
The plants were grown in three different types of soil. The control soil without any supplementation was designated “S”. Two variants of soil supplemented with growth stimulators were used as experimental conditions. The soil supplemented (up to 5%) with Z-ion (Ecokhimprom, Republic of Belarus) was designated “S+Z”. Z-ion contents N1:P1:K1, mg/kg: 4960:4730:11280, pH 6.9. The third variant was soil supplemented with complex fertilizer from Crystallon (Fertica, Russia), designated “S+C”. The contents of the microelements in the Crystallon samples are as follows: N:P:K – 18:18:18, Mg – 3, S – 5, Fe – 0.07, Mn – 0.04, B – 0.025, Cu – 0.01, Mo – 0.004, and Zn – 0.025. Complex fertilizer (0.002 g/ml) was applied in dissolved form during irrigation.
The plants were grown in three-section chambers with light sources designed and manufactured at the IAСP FEB RAS as described previously [27]. The matrices of the light sources were composed of three-watt (CHANZON, China) LEDs of different colours. In this study, the following varieties of light were used: warm white (W) as a control; monochrome light sources, red (R) and green (R); and binary light sources, red/blue (RB). All light variants were used with an intensity of 300 µmol m-2s-1. The dimensions of the chamber were 100 × 50 × 50 cm; other parameters were described previously [27]. The temperature regime was maintained by an exhaust fan FFB1212SH 12025 (power: 14.8 W, speed: 3700 rpm, air volume: 140.16 cfm, China). The spectra were measured using a PG200N spectrophotometer (UPRtek, Taiwan). A UT61A digital multimeter (Uni-T, China) controlled the driver supply currents.
4.2. Measurements of Morphometric Characteristics
Measurements of the morphometric characteristics (area, length, width of the leaves) of the 35-day-old O. basilicum plants were carried out using a scanner Epson Perfection V850 Pro (Epson, Japan) with WinFolia Pro 2020 software (Regent Instruments, UK). The weights of the aerial parts of the plants and roots were measured via Ohaus EX225/AD automatic scales (Ohaus Corporation, USA).
The dry weights were calculated using the following formula:
4.3. Anthocyanin and Caffeic Acid Derivative Analysis
Chemicals
An analytical standard of cyanidine chloride was obtained from Sigma‒Aldrich (St. Louis, MO, USA). We used a standard sample of research-grade rosmarinic acid, which was previously obtained [28]. All extraction solutions and eluents were prepared with ultrapure water (Millipore, Bedford, MA, USA). All solvents were of analytical grade.
Sample preparation for analytical chromatograph
The sample preparation procedure was carried out as described in our earlier publication [27]. To summarize, 80% v/v methanol was used for sonicated extraction of dried and powdered plant tissue. The resulting supernatant was then filtered through a 0.45-μm membrane (Millipore, Bedford, MA, USA), and a 1 µl aliquot was used for analysis.
Analytical chromatography and mass spectrometry
An Agilent 1260 Infinity analytical HPLC instrument (Agilent Technologies, Santa Clara, California, USA) equipped with a PDA detector was used for metabolite profiling. The separation of the determined compounds was carried out on a Zorbax C18 column (150 mm, 2.1 mm i.d., 3.5 μm, Agilent Technologies, USA) at a column temperature of 40°C. The gradient elution with a flow rate of 0.2 ml/min was realized with two eluents, A (0.1% aqueous formic acid) and B (acetonitrile with 0.1% formic acid addition). The following linear gradient was used: 0–10 min – 0% B; 45 min – 40% B; and 55 min – 100% B. UV‒Vis spectral measurements were recorded in the range of 200–800 nm using a PDA detector, and chromatograms for quantification were acquired at wavelengths of 325 and 530 nm. An ion trap mass spectrometer Bruker HCT ultra PTM Discovery System (Bruker Daltonik GmbH, Bremen, Germany) interfaced with an HPLC system was used for the MS experiments. The MS instrument was operated in electrospray ionization (ESI) mode and in both positive and negative ionization modes. The following settings were used: mass range, 100-1.400 Da; drying gas (N2) flow rate, 8.0 l/min; nebulizer gas (N2) pressure, 25 psi; and drying gas temperature, 325 °C. The auto-MSn mode (smart fragmentation) was used for the MS2 and MS3 experiments. A Shimadzu LCMS-IT-TOF instrument (Shimadzu, Japan) including a tandem ion trap/time-of-flight mass spectrometer was utilized for the high-resolution MS studies. The following operating conditions were used: the drying gas (N2) pressure was 100 kPa, the nebulizer gas flow rate was 1.5 l/min, the ion source potential was changed from -3.8 to 4.5 kV, and the interface temperature was 200°C. The external standard method was employed for quantification of the detected compounds. An analytical standard of cyanidine chloride was chosen for quantifying the discovered anthocyanins, and a rosmarinic acid methanol solution was used for evaluating caffeic acid derivatives.
Since the plants were grown in separate containers and not by continuous sowing of the area, the productivity was calculated per liter of soil; in one container, there was 0.5 liters of soil. Productivity was calculated as follows:
where Content is the content of substances in a plant (µM/g FW) and FW is the fresh weight of the above-ground part of the O. basilicum plants per liter of soil (g/l).
Productivity (µM/l) = Content × FW,
4.4. Statistical Analysis
An Excel add-on called XFolia 2020 was used to handle the statistical data (morphometric parameters). The STATISTICA software package (StatSoft, Inc., USA) was used for statistical analysis. Every value is presented as the mean ± standard error (SE). Student’s t-test was employed for the statistical assessment to compare two independent groups. Analysis of variance (ANOVA) and a multiple comparison process were used to compare multiple datasets. A post hoc test called Fisher’s protected least significant difference (PLSD) was employed. At P < 0.05, the threshold for statistical significance was established. The correlations between the two variables were determined via Pearson correlation analysis.
5. Conclusions
In recent years, research on lighting conditions that can improve the profitability and sustainability of PFALs has been essential and relevant [1]. In the present work, in addition to growth characteristics, we examined the content of caffeic acid derivatives and anthocyanin in the leaves of O. basilicum plants growing under different combinations of artificial lighting and different soil compositions. We have shown that, along with a significant improvement in growth characteristics, supplementation of soil with Crystallon leads to a significant decrease in the content of caffeic acid derivatives and anthocyanin, more than 2-fold. Interestingly, growing O. basilicum plants under R and RB light led to a 2-fold increase in the biosynthesis of both the key caffeic acid derivative RA and anthocyanin. However, given that under RB light, there is no positive effect of Crystallon on growth, the productivity of RA and anthocyanin reached a maximum when O. basilicum plants were grown under R light and Crystallon.
Author Contributions
Veremeichik G.N., Y.N. Kulchin, Subbotin E.P.: Conceptualization, Data curation, Project administration, Supervision. G.N. Veremeichik, Subbotin E.P.: Validation, Visualization, Writing – original draft, Writing – review & editing. Veremeichik G.N., Grigorchuk V.P., Tikhonova O.A., Brodovskaya E.V., Silantieva S.A., Kozhanov S.O., Subbotina N.I., Yaroshenko Y.L.: Investigation, Methodology, Formal Analysis. V.P. Bulgakov, Y.N. Kulchin: Resources.
Funding
This research was carried out within the state assignment of the IACP FEB RAS (Theme FWFW-2024-0004).
Data Availability Statement
The datasets generated during and/or analysed during the current study are available from the corresponding author upon reasonable request.
Acknowledgments
The analyses described in this work were performed using equipment from the Instrumental Centre for Biotechnology and Gene Engineering at the Federal Scientific Centre of East Asia Terrestrial Biodiversity of the Far East Branch of the Russian Academy of Sciences.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Orsini, F.; Pennisi, G.; Gianquinto, F.Z.G. Sustainable use of resources in plant factories with artificial lighting (PFALs). Europ J Hortic Sci 2020, 85, 297–309. [Google Scholar] [CrossRef]
- Makri, O.; Kintzios, S. Ocimum sp. (Basil): Botany, Cultivation, Pharmaceutical Properties, and Biotechnology. J Herb Spices Med P 2008, 13, 123–150. [Google Scholar] [CrossRef]
- Bora, K.S.; Arora, S.; Shri, R. Role of Ocimum basilicum L. in prevention of ischemia and reperfusion-induced cerebral damage, and motor dysfunctions in mice brain. J of Ethnopharmac 2011, 137, 1360–1365. [Google Scholar] [CrossRef] [PubMed]
- Qin, T.; Rasul, A.; Sarfraz, A.; Sarfraz, I.; Hussain, G.; Anwar, H.; Riaz, A.; Liu, S.; Wei, W.; Li, J.; Li, X. Salvianolic acid A & B: potential cytotoxic polyphenols in battle against cancer via targeting multiple signaling pathways. Int J Biol Sci 2019, 15, 2256–2264. [Google Scholar] [CrossRef] [PubMed]
- Prinsi, B.; Morgutti, S.; Negrini, N.; Faoro, F.; Espen, L. Insight into composition of bioactive phenolic compounds in leaves and flowers of green and purple Basil. Plants 2019, 9, 22. [Google Scholar] [CrossRef]
- Yu, J.; Xie, J.; Sun, M.; Xiong, S.; Xu, C.; Zhang, Z.; Li, M.; Li, C.; Lin, L. Plant-derived caffeic acid and its derivatives: an overview of their NMR data and biosynthetic pathways. Molecules 2024, 29, 1625. [Google Scholar] [CrossRef]
- Khan, F.; Bamunuarachchi, N.I.; Tabassum, N.; Kim, Y.-M. Caffeic acid and its derivatives: antimicrobial drugs toward microbial pathogens. J Agric Food Chem 2021, 69, 2979–3004. [Google Scholar] [CrossRef]
- Lee, J.; Scagel, C.F. Chicoric acid found in basil (Ocimum basilicum L.) leaves. Food Chem 2009, 115, 650–656. [Google Scholar] [CrossRef]
- Romano, R.; De Luca, L.; Aiello, A.; Pagano, R.; Di Pierro, P.; Pizzolongo, F.; Masi, P. Basil (Ocimum basilicum L.) leaves as a source of bioactive compounds. Foods 2022, 11, 3212. [Google Scholar] [CrossRef]
- Trócsányi, E.; György, Z.; Zámboriné-Németh, É. New insights into rosmarinic acid biosynthesis based on molecular studies. Cur Plant Biol 2020, 23, 100162. [Google Scholar] [CrossRef]
- Bulgakov, V.P.; Vereshchagina, Y.V.; Veremeichik, G.N. Anticancer polyphenols from cultured plant cells: production and new bioengineering strategies. CMC 2018, 25, 4671–4692. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.-S.; Liu, A. Light signaling induces anthocyanin biosynthesis via AN3 mediated COP1 expression. Plant Signal Behav 2015, 10, e1001223. [Google Scholar] [CrossRef] [PubMed]
- Taulavuori, K.; Pyysalo, A.; Taulavuori, E.; Julkunen-Tiitto, R. Responses of phenolic acid and flavonoid synthesis to blue and blue-violet light depends on plant species. Environ Exp Bot 2018, 150, 183–187. [Google Scholar] [CrossRef]
- Wang, W.; Su, M.; Li, H.; Zeng, B.; Chang, Q.; Lai, Z. Effects of supplemental lighting with different light qualities on growth and secondary metabolite content of Anoectochilus roxburghii. PeerJ 2018, 6, e5274. [Google Scholar] [CrossRef]
- Aghaye Noroozlo, Y.; Souri, M.K.; Delshad, M. Effects of soil application of amino acids, ammonium, and nitrate on nutrient accumulation and growth characteristics of sweet basil. Com Soil Sci Plant Anal 2019, 50, 2864–2872. [Google Scholar] [CrossRef]
- Lim, P.O.; Kim, H.J.; Gil Nam, H. leaf senescence. Annu Rev Plant Biol 2007, 58, 115–136. [Google Scholar] [CrossRef]
- Ma, Y.; Ma, X.; Gao, X.; Wu, W.; Zhou, B. Light induced regulation pathway of anthocyanin biosynthesis in plants. IJMS 2021, 22, 11116. [Google Scholar] [CrossRef]
- Pasch, J.; Appelbaum, S.; Palm, H.W.; Knaus, U. Growth of basil (Ocimum basilicum) in aeroponics, DRF, and raft systems with effluents of african catfish (Clarias gariepinus) in decoupled aquaponics (s.s.). AgriEngineering 2021, 3, 559–574. [Google Scholar] [CrossRef]
- Fussy, A.; Papenbrock, J. An overview of soil and soilless cultivation techniques—chances, challenges and the neglected question of sustainability. Plants 2022, 11, 1153. [Google Scholar] [CrossRef]
- Maran, B.; Silvestre, W.P.; Pauletti, G.F. Preliminary study on the effect of artificial lighting on the production of basil, mustard, and red cabbage seedlings. AgriEngineering 2024, 6, 1043–1054. [Google Scholar] [CrossRef]
- Hammock, H.A.; Kopsell, D.A.; Sams, C.E. Supplementary blue and red LED narrowband wavelengths improve biomass yield and nutrient uptake in hydroponically grown basil. horts 2020, 55, 1888–1897. [Google Scholar] [CrossRef]
- Shiga, T.; Shoji, K.; Shimada, H.; Hashida, S.; Goto, F.; Yoshihara, T. Effect of light quality on rosmarinic acid content and antioxidant activity of sweet basil, Ocimum basilicum L. Plant Biotechnol 2009, 26, 255–259. [Google Scholar] [CrossRef]
- Matysiak, B.; Kowalski, A. The growth, photosynthetic parameters and nitrogen status of basil, coriander and oregano grown under different led light spectra. Acta Sci Pol Hortorum Cultus 2021, 20(2), 13–22. [Google Scholar] [CrossRef]
- Inoue, S.; Kinoshita, T.; Takemiya, A.; Doi, M.; Shimazaki, K. Leaf positioning of Arabidopsis in response to blue light. Mol Plant 2008, 1, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Jones-Baumgardt, C.; Llewellyn, D.; Zheng, Y. Different microgreen genotypes have unique growth and yield responses to intensity of supplemental PAR from light-emitting diodes during winter greenhouse production in southern ontario, Canada. horts 2020, 55, 156–163. [Google Scholar] [CrossRef]
- Tabbert, J.M.; Riewe, D.; Schulz, H.; Krähmer, A. Facing energy limitations – approaches to increase basil (Ocimum basilicum L.) growth and quality by different increasing light intensities emitted by a broadband LED light spectrum (400-780 nm). Front Plant Sci 2022, 13, 1055352. [Google Scholar] [CrossRef] [PubMed]
- Veremeichik, G.N.; Grigorchuk, V.P.; Makhazen, D.S.; Subbotin, E.P.; Kholin, A.S.; Subbotina, N.I.; Bulgakov, D.V.; Kulchin, Y.N.; Bulgakov, V.P. High production of flavonols and anthocyanins in Eruca sativa (Mill) Thell plants at high artificial LED light intensities. Food Chem 2023, 408, 135216. [Google Scholar] [CrossRef]
- Fedoreyev, S.A.; Veselova, M.V.; Krivoschekova, O.E.; Mischenko, N.P.; Denisenko, V.A.; Dmitrenok, P.S.; Glazunov, V.P.; Bulgakov, V.P.; Tchernoded, G.K.; Zhuravlev, Y.N. Caffeic acid metabolites from Eritrichium sericeum cell cultures. Planta med 2005, 71, 446–451. [Google Scholar] [CrossRef]
- Parrine, D.; Greco, T.M.; Muhammad, B.; Wu, B.-S.; Zhao, X.; Lefsrud, M. Color-specific recovery to extreme high-light stress in plants. Life 2021, 11, 812. [Google Scholar] [CrossRef]
- Yavari, N.; Tripathi, R.; Wu, B.-S.; MacPherson, S.; Singh, J.; Lefsrud, M. The effect of light quality on plant physiology, photosynthetic, and stress response in Arabidopsis thaliana leaves. PLoS ONE 2021, 16, e0247380. [Google Scholar] [CrossRef]
- Van Delden, S.H.; SharathKumar, M.; Butturini, M.; Graamans, L.J.A.; Heuvelink, E.; Kacira, M.; Kaiser, E.; Klamer, R.S.; Klerkx, L.; Kootstra, G.; Loeber, A.; Schouten, R.E.; Stanghellini, C.; Van Ieperen, W.; Verdonk, J.C.; Vialet-Chabrand, S.; Woltering, E.J.; Van De Zedde, R.; Zhang, Y.; Marcelis, L.F.M. Current status and future challenges in implementing and upscaling vertical farming systems. Nat Food 2021, 2, 944–956. [Google Scholar] [CrossRef] [PubMed]
- Deineka, V.I.; Kul’chenko, Ya.Yu.; Blinova, I.P.; Chulkov, A.N.; Deineka, L. A Anthocyanins of basil leaves: determination and preparation of dried encapsulated forms. Rus J Bioorg Chem 2019, 45, 895–899. [Google Scholar] [CrossRef]
- Saga Kaboré, D.; Héma, A.; Kabré, E.; Bazié, R.; Sakira, A.K.; Koala, M.; Koussao Somé, P.-A.; Palé, E.; Issa Somé, T.; Duez, P. Identification of five acylated anthocyanins and determination of antioxidant contents of total extracts of a purple-fleshed Ipomoea batatas L. variety grown in Burkina Faso. IRJPAC 2021, 48–66. [Google Scholar] [CrossRef]
- Luna, M.C.; Bekhradi, F.; Ferreres, F.; Jordán, M.J.; Delshad, M.; Gil, M.I. Effect of water stress and storage time on anthocyanins and other phenolics of different genotypes of fresh sweet basil. J Agric Food Chem 2015, 63, 9223–9231. [Google Scholar] [CrossRef]
- Castañeda-Ovando, A.; Pacheco-Hernández, Ma.D.L.; Páez-Hernández, Ma.E.; Rodríguez, J.A.; Galán-Vidal, C.A. Chemical studies of anthocyanins: A review. Food Chem 2009, 113, 859–871. [Google Scholar] [CrossRef]
- Chomczyńska, M.; Soldatov, V.; Wasąg, H. Effect of different variants of the ion exchange substrate on vegetation of Dactylis glomerata L on the degraded soil. Proc ECOpole 2010, 4. [Google Scholar]
- Chomczyńska, M.; Soldatov, V.; Wasąg, H.; Turski, M. Effect of ion exchange substrate on grass root development and cohesion of sandy soil. Int Agrophysics 2016, 30, 293–300. [Google Scholar] [CrossRef]
- Chomczyńska, M.; Zdeb, M. The effect of Z-ion zeolite substrate on growth of Zea mays L. as energy crop growing on marginal soil. J Ecol Eng 2019, 20, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Piovene, C.; Orsini, F.; Bosi, S.; Sanoubar, R.; Bregola, V.; Dinelli, G.; Gianquinto, G. Optimal red:blue ratio in led lighting for nutraceutical indoor horticulture. Sci Horticult 2015, 193, 202–208. [Google Scholar] [CrossRef]
- Lobiuc, A.; Vasilache, V.; Oroian, M.; Stoleru, T.; Burducea, M.; Pintilie, O.; Zamfirache, M.-M. Blue and red LED illumination improves growth and bioactive compounds contents in acyanic and cyanic Ocimum basilicum L. microgreens. Molecules 2017, 22, 2111. [Google Scholar] [CrossRef]
- Chutimanukul, P.; Wanichananan, P.; Janta, S.; Toojinda, T.; Darwell, C.T.; Mosaleeyanon, K. The influence of different light spectra on physiological responses, antioxidant capacity and chemical compositions in two holy basil cultivars. Sci Rep 2022, 12, 588. [Google Scholar] [CrossRef] [PubMed]
- Dou, H.; Niu, G.; Gu, M.; Masabni, J. Morphological and physiological responses in basil and brassica species to different proportions of red, blue, and green wavelengths in indoor vertical farming. J Amer Soc Hort Sci 2020, 145, 267–278. [Google Scholar] [CrossRef]
- Kivimäenpä, M.; Mofikoya, A.; Abd El-Raheem, A.M.; Riikonen, J.; Julkunen-Tiitto, R.; Holopainen, J.K. Alteration in light spectra causes opposite responses in volatile phenylpropanoids and terpenoids compared with phenolic acids in sweet basil (Ocimum basilicum) leaves. J Agric Food Chem 2022, 70, 12287–12296. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Vasiliev, M.; Alameh, K. LED illumination spectrum manipulation for increasing the yield of sweet basil (Ocimum basilicum L.). Plants 2021, 10, 344. [Google Scholar] [CrossRef]
- Song, T.-E.; Moon, J.-K.; Lee, C.H. Polyphenol content and essential oil composition of sweet basil cultured in a plant factory with light-emitting diodes. HST 2020, 38, 620–630. [Google Scholar] [CrossRef]
- Author 1, A.B. (University, City, State, Country); Author 2, C. (Institute, City, State, Country). Personal communication, 2012.
Figure 1.
Impact of combinations of artificial lighting and soil composition on the growth and morphology of O. basilicum plants. A, biomass accumulation (g) of 40-day-old plants (aboveground part only); B, biomass accumulation (g) of 40-day-old plants (roots); FW, fresh weight (left panel), DW, dry weight (right panel). C, Morphology of 40-day-old plants. The plants were grown in soil (S), soil supplemented with Zion (S+Z), or soil supplemented with Crystallon (S+C). W, R, G, and RB represent different light variations with intensities of 300 µmol m-2s-1. The detailed specifications of the cultivation conditions are given in the Materials and Methods section. Data obtained from three independent experiments with ten biological replicates are presented as the mean ± standard error of the mean, and different letters above the error bars indicate statistical significance (ANOVA, p ˂0.05).
Figure 1.
Impact of combinations of artificial lighting and soil composition on the growth and morphology of O. basilicum plants. A, biomass accumulation (g) of 40-day-old plants (aboveground part only); B, biomass accumulation (g) of 40-day-old plants (roots); FW, fresh weight (left panel), DW, dry weight (right panel). C, Morphology of 40-day-old plants. The plants were grown in soil (S), soil supplemented with Zion (S+Z), or soil supplemented with Crystallon (S+C). W, R, G, and RB represent different light variations with intensities of 300 µmol m-2s-1. The detailed specifications of the cultivation conditions are given in the Materials and Methods section. Data obtained from three independent experiments with ten biological replicates are presented as the mean ± standard error of the mean, and different letters above the error bars indicate statistical significance (ANOVA, p ˂0.05).
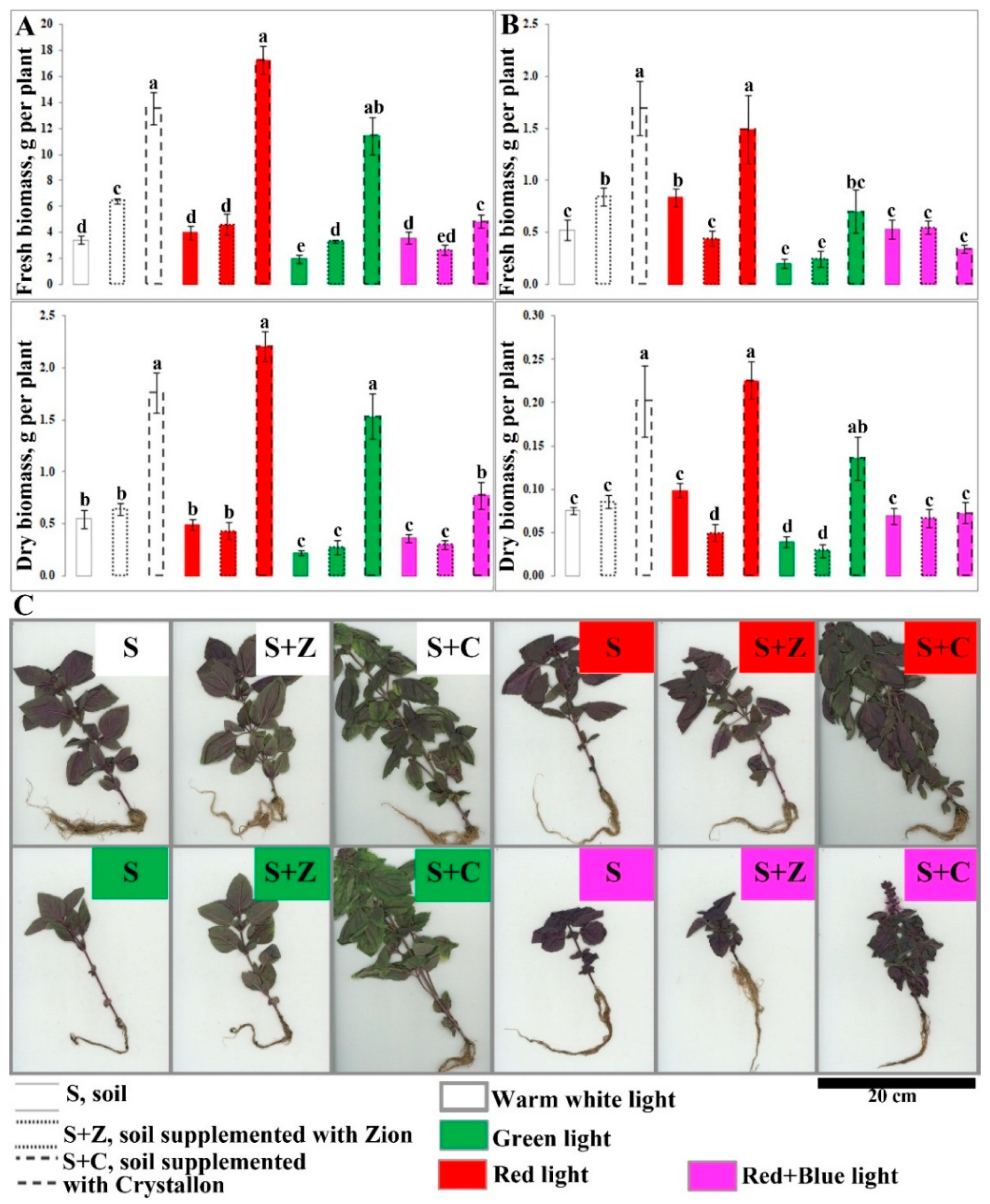
Figure 2.
Morphological parameters of O. basilicum plants growing under different combinations of artificial lighting and soil composition. The heatmap shows statistically significant differences in the measurements of height (cm), number of internodes and leaves, steam diameter (cm), and total area of the leaf (cm2). The plants were grown in soil (S), soil supplemented with Zion (S+Z), or soil supplemented with Crystallon (S+C). W, R, G, and RB represent different light variations with intensities of 300 µmol m-2s-1. Data obtained from three independent experiments with ten biological replicates are presented as the mean ± standard error of the mean, and different letters above the error bars indicate statistical significance (ANOVA, p ˂0.05).
Figure 2.
Morphological parameters of O. basilicum plants growing under different combinations of artificial lighting and soil composition. The heatmap shows statistically significant differences in the measurements of height (cm), number of internodes and leaves, steam diameter (cm), and total area of the leaf (cm2). The plants were grown in soil (S), soil supplemented with Zion (S+Z), or soil supplemented with Crystallon (S+C). W, R, G, and RB represent different light variations with intensities of 300 µmol m-2s-1. Data obtained from three independent experiments with ten biological replicates are presented as the mean ± standard error of the mean, and different letters above the error bars indicate statistical significance (ANOVA, p ˂0.05).
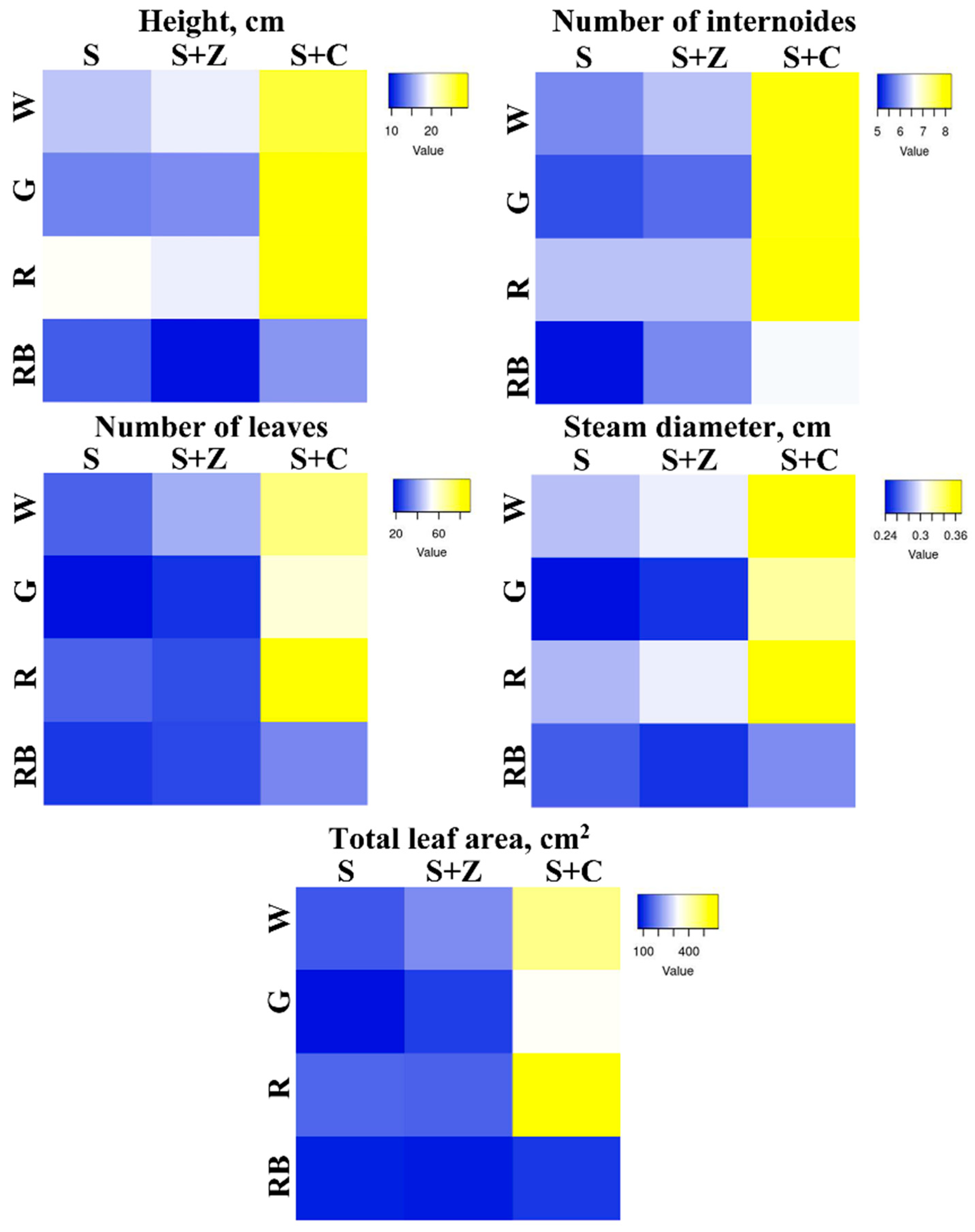
Figure 3.
HPLC‒UV‒MS determination of caffeic acid derivatives identified in crude extracts of O. basilicum. The UV profile of a typical studied sample recorded at 325 nm is shown in comparison with that of a standard sample of rosmarinic acid (A). The extracted ion chromatograms (EICs) acquired in negative ion mode correspond to the deprotonated molecules of the individual compounds with a restricted window of ± 0.5 m/z units and are shown with an overlay (B). The peak numbers correspond to the identified components and are listed in Table 1.
Figure 3.
HPLC‒UV‒MS determination of caffeic acid derivatives identified in crude extracts of O. basilicum. The UV profile of a typical studied sample recorded at 325 nm is shown in comparison with that of a standard sample of rosmarinic acid (A). The extracted ion chromatograms (EICs) acquired in negative ion mode correspond to the deprotonated molecules of the individual compounds with a restricted window of ± 0.5 m/z units and are shown with an overlay (B). The peak numbers correspond to the identified components and are listed in Table 1.
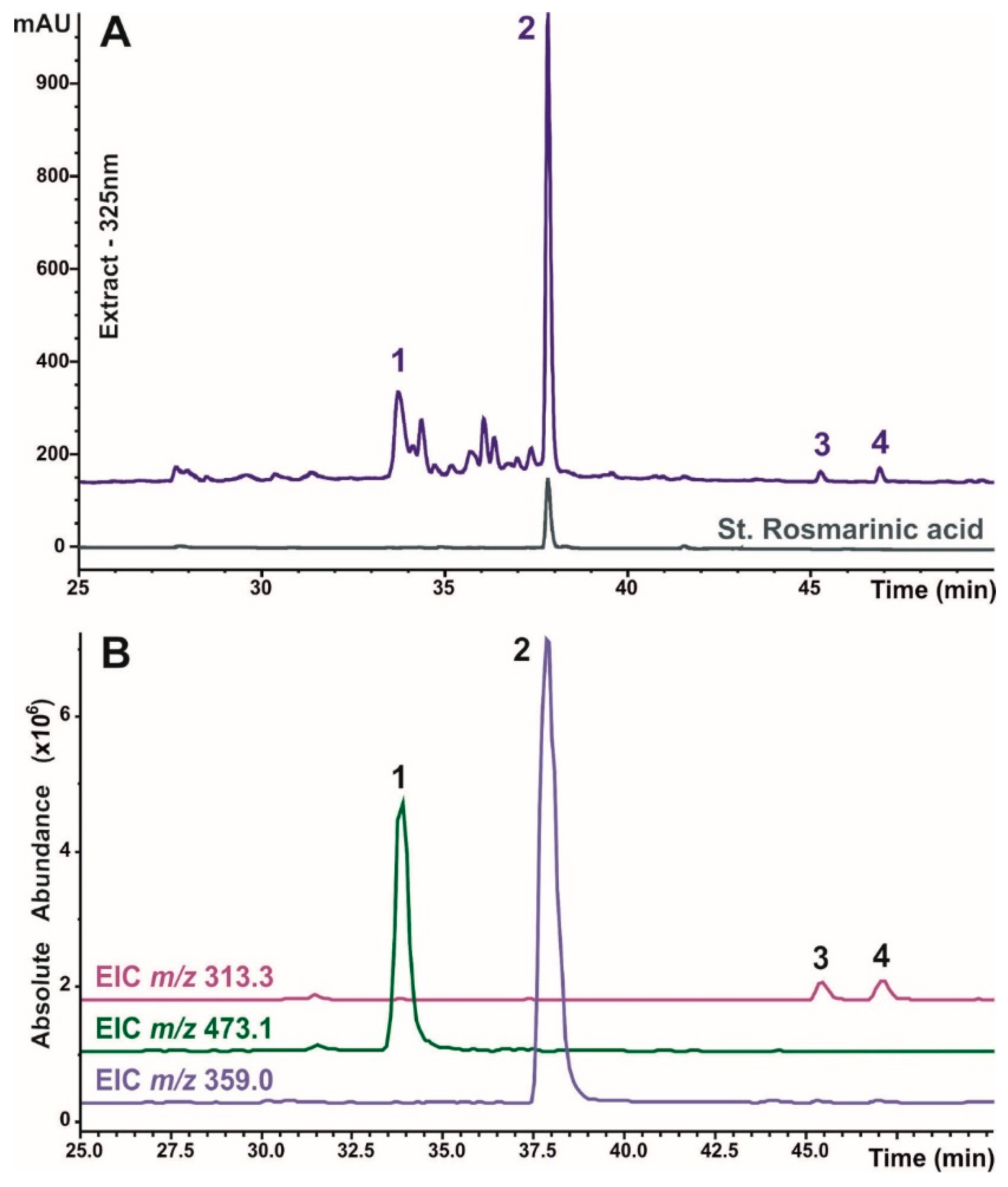
Figure 4.
Analysis of caffeic acid derivatives in the leaves of O. basilicum plants growing under different combinations of artificial lighting and soil composition. Content (left diagrams, mg/g dry weight) and productivity (right diagrams, mg/l soil) of rosmarinic acid, chicoric acid, and salvianolic acid F (isomers 1 and 2) in 40-day-old O. basilicum plants. The plants were grown in soil (S), soil supplemented with Zion (S+Z), or soil supplemented with Crystallon (S+C). W, R, G, and RB represent different light variations with intensities of 300 µmol m-2s-1. Data obtained from three independent experiments with ten biological replicates are presented as the mean ± standard error of the mean, and different letters above the error bars indicate statistical significance (ANOVA, p ˂0.05).
Figure 4.
Analysis of caffeic acid derivatives in the leaves of O. basilicum plants growing under different combinations of artificial lighting and soil composition. Content (left diagrams, mg/g dry weight) and productivity (right diagrams, mg/l soil) of rosmarinic acid, chicoric acid, and salvianolic acid F (isomers 1 and 2) in 40-day-old O. basilicum plants. The plants were grown in soil (S), soil supplemented with Zion (S+Z), or soil supplemented with Crystallon (S+C). W, R, G, and RB represent different light variations with intensities of 300 µmol m-2s-1. Data obtained from three independent experiments with ten biological replicates are presented as the mean ± standard error of the mean, and different letters above the error bars indicate statistical significance (ANOVA, p ˂0.05).
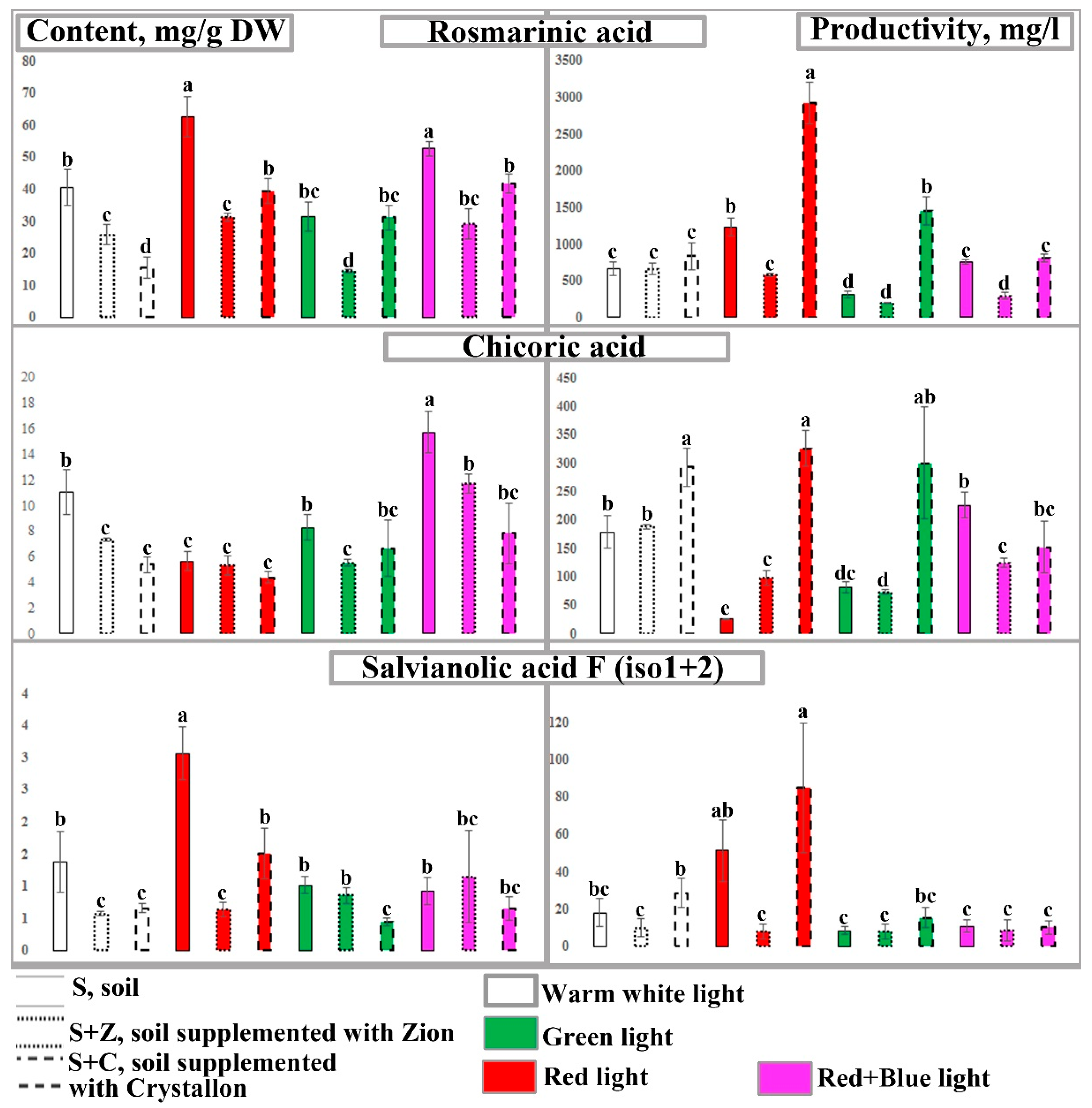
Figure 5.
Analysis of caffeic acid derivatives in the roots of O. basilicum plants growing under different combinations of artificial lighting and soil composition. Contents (mg/g dry weight) of rosmarinic acid, chicoric acid, and salvianolic acid F (isomers 1 and 2) in the roots of 40-day-old O. basilicum plants. The plants were grown in soil (S), soil supplemented with Zion (S+Z), or soil supplemented with Crystallon (S+C). W, R, G, and RB represent different light variations with intensities of 300 µmol m-2s-1. Data obtained from three independent experiments with ten biological replicates are presented as the mean ± standard error of the mean, and different letters above the error bars indicate statistical significance (ANOVA, p ˂0.05).
Figure 5.
Analysis of caffeic acid derivatives in the roots of O. basilicum plants growing under different combinations of artificial lighting and soil composition. Contents (mg/g dry weight) of rosmarinic acid, chicoric acid, and salvianolic acid F (isomers 1 and 2) in the roots of 40-day-old O. basilicum plants. The plants were grown in soil (S), soil supplemented with Zion (S+Z), or soil supplemented with Crystallon (S+C). W, R, G, and RB represent different light variations with intensities of 300 µmol m-2s-1. Data obtained from three independent experiments with ten biological replicates are presented as the mean ± standard error of the mean, and different letters above the error bars indicate statistical significance (ANOVA, p ˂0.05).
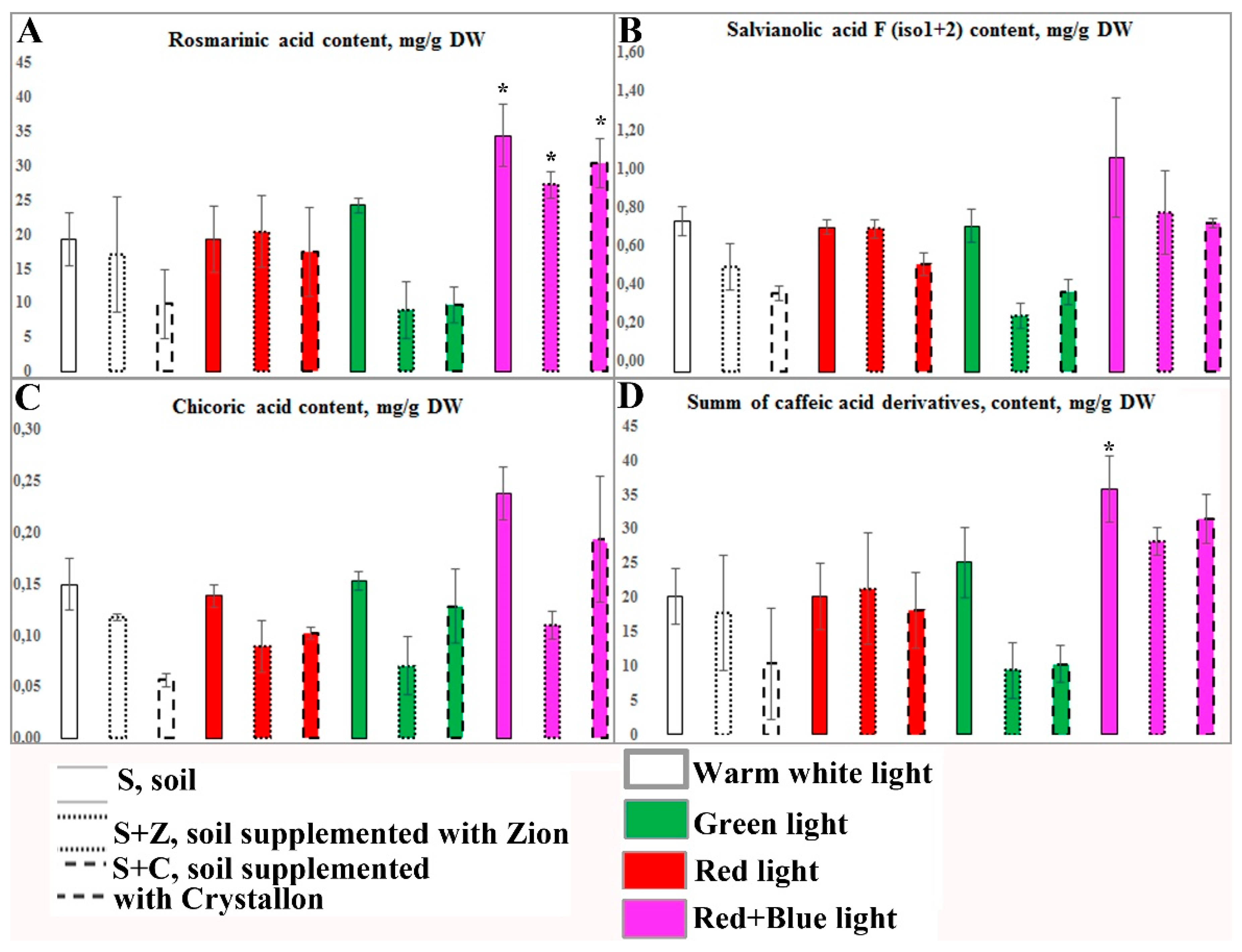
Figure 6.
Analysis of the productivity of caffeic acid derivatives in O. basilicum plants growing under different combinations of artificial lighting and soil composition. Productivity (mg/l soil) of the sum of the caffeic acid derivatives in the leaves (A, fresh weight) and leaves and roots (B, dry weight) of 40-day-old O. basilicum plants. The plants were grown in soil (S), soil supplemented with Zion (S+Z), or soil supplemented with Crystallon (S+C). W, R, G, and RB represent different light variations with intensities of 300 µmol m-2s-1. Data obtained from three independent experiments with ten biological replicates are presented as the mean ± standard error of the mean, and different letters above the error bars indicate statistical significance (ANOVA, p ˂0.05).
Figure 6.
Analysis of the productivity of caffeic acid derivatives in O. basilicum plants growing under different combinations of artificial lighting and soil composition. Productivity (mg/l soil) of the sum of the caffeic acid derivatives in the leaves (A, fresh weight) and leaves and roots (B, dry weight) of 40-day-old O. basilicum plants. The plants were grown in soil (S), soil supplemented with Zion (S+Z), or soil supplemented with Crystallon (S+C). W, R, G, and RB represent different light variations with intensities of 300 µmol m-2s-1. Data obtained from three independent experiments with ten biological replicates are presented as the mean ± standard error of the mean, and different letters above the error bars indicate statistical significance (ANOVA, p ˂0.05).
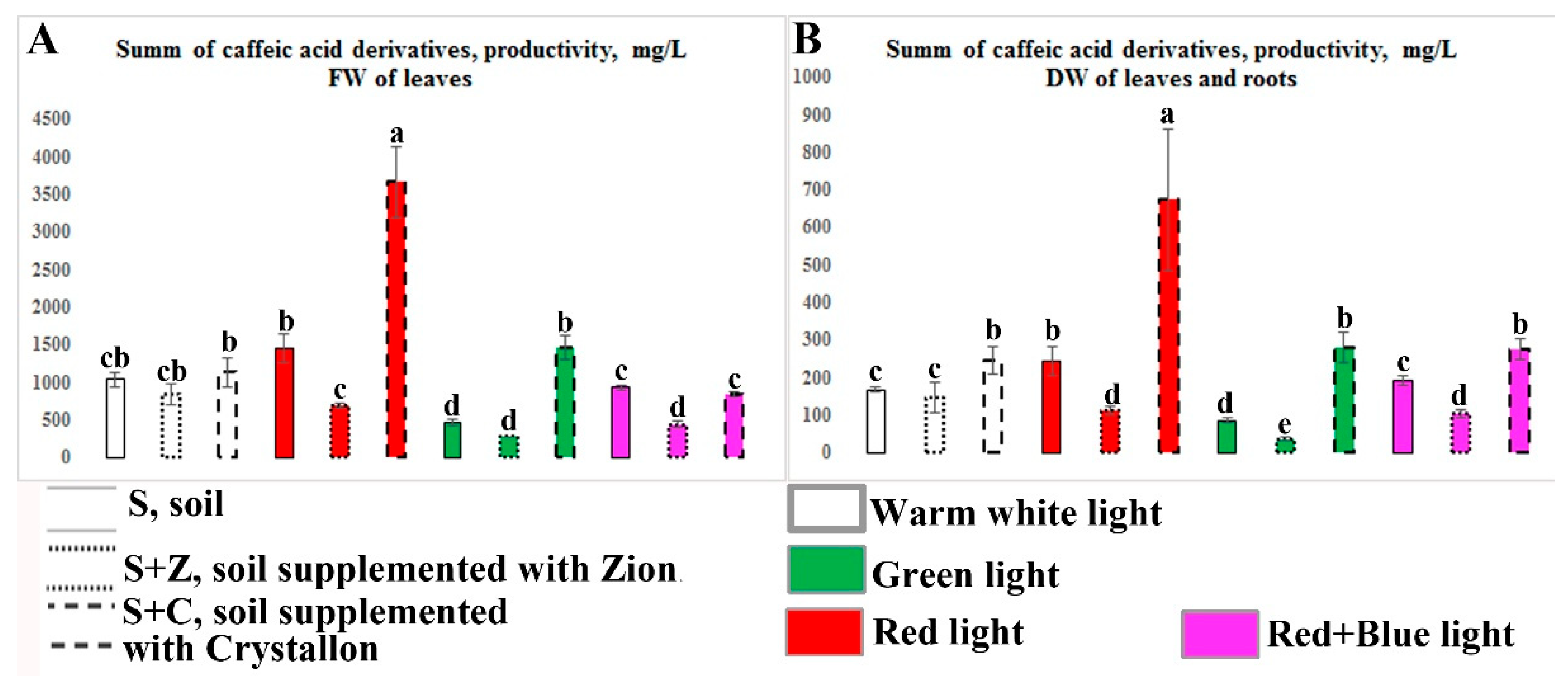
Figure 7.
HPLC-UV/Vis-MS determination of anthocyanin compounds identified in crude extracts of O. basilicum. Vis profile of a typical studied sample recorded at 520 nm (A). Extracted ion chromatograms (EICs) of the studied anthocyanins acquired in positive ion mode with a restricted window of ± 0.5 m/z and are displayed in each trace (B). The peak numbers correspond to the identified components and are listed in Table 1.
Figure 7.
HPLC-UV/Vis-MS determination of anthocyanin compounds identified in crude extracts of O. basilicum. Vis profile of a typical studied sample recorded at 520 nm (A). Extracted ion chromatograms (EICs) of the studied anthocyanins acquired in positive ion mode with a restricted window of ± 0.5 m/z and are displayed in each trace (B). The peak numbers correspond to the identified components and are listed in Table 1.
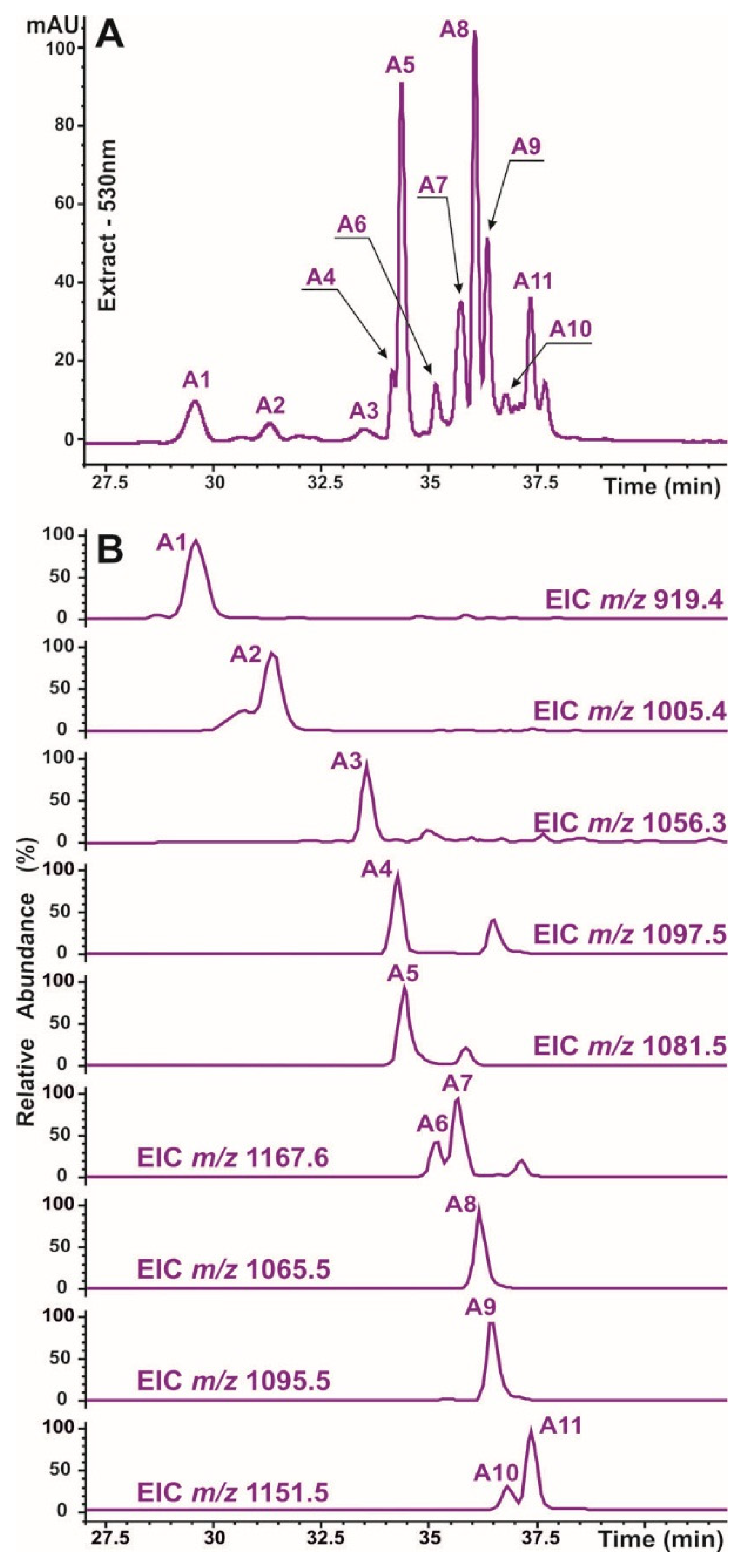
Figure 8.
Content and productivity of anthocyanins in O. basilicum plants growing under different combinations of artificial lighting and soil composition. Total anthocyanin content (A, µM/g DW) and productivity (B, µM/l soil, fresh weight) in 40-day-old O. basilicum plants. The plants were grown in soil (S), soil supplemented with Zion (S+Z), or soil supplemented with Crystallon (S+C). W, R, G, and RB represent different light variations with intensities of 300 µmol m-2s-1. Data obtained from three independent experiments with ten biological replicates are presented as the mean ± standard error of the mean, and different letters above the error bars indicate statistical significance (ANOVA, p ˂0.05).
Figure 8.
Content and productivity of anthocyanins in O. basilicum plants growing under different combinations of artificial lighting and soil composition. Total anthocyanin content (A, µM/g DW) and productivity (B, µM/l soil, fresh weight) in 40-day-old O. basilicum plants. The plants were grown in soil (S), soil supplemented with Zion (S+Z), or soil supplemented with Crystallon (S+C). W, R, G, and RB represent different light variations with intensities of 300 µmol m-2s-1. Data obtained from three independent experiments with ten biological replicates are presented as the mean ± standard error of the mean, and different letters above the error bars indicate statistical significance (ANOVA, p ˂0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



