
Submitted:
12 November 2024
Posted:
14 November 2024
You are already at the latest version
Abstract
Inflammatory Bowel Diseas21e (IBD) is a multifactorial gastrointestinal condition that encompasses two major forms of intestinal inflammation: Crohn's disease (CD) and ulcerative colitis (UC). Both conditions are linked to auto-inflammatory reactions and genetic predispositions, Various drug therapies and biological treatments proposed to reduce IBD associated inflammation. We induced IBD in a mouse model by stimulating bowel inflammation with oral dextran sodium sulfate (DSS) beverage. Our novel cell therapy approach for IBD involves intramuscular (IM) and intraperitoneal (IP) delivery of non-matched, expanded, potent xenogeneic fetal human mesenchymal stromal cells (f-hPSCs) in 2 × 10⁶ cell injections. This cell therapy has already shown previously to induce pro-regenerative and anti-inflammatory effects in different systemic and local disorders. In the current study, IP-injected f-hPSCs, following DSS-induced IBD, enhanced the regenerative processes of the damaged bowel and reduced inflammation, leading to rapid weight regain and decreased inflammation-associated parameters, such as colon edema, bowel shortening, and a threefold increase in bowel mass. Although both IP and IM delivery of f-hPSCs were effective, IP injections produced a superior response. The f-hPSCs injected mice lost only ~15% of their weight at the peak of the IBD damage, compared to ~25% in untreated mice. A reduction in the inflammatory response of the gut was observed, evidenced by a decrease in neutrophil infiltration as measured by a myeloperoxidase (MPO) assay. Additionally, a significant improvement in the histological score of the gut and faster recovery to 90% of its original size were recorded. These findings suggest that f-hPSCs treatments could serve as an effective and safe anti-inflammatory and pro-regenerative treatment for IBD.
Keywords:
Expanded Fetal Human Placental Stromal Cells (f-hPSCs)
; inflammatory bowel disease (IBD)
; adult cell therapy
; dextran sodium sulfate (DSS)
1. Introduction
Ulcerative colitis and Crohn's disease (CD) are distinct manifestations of Inflammatory Bowel Disease (IBD), both of which involve inflammation along the lining of the gastrointestinal tract, particularly the large intestine (colon) and rectum. These multifactorial disorders are commonly associated with a disrupted interaction between the host immune system and the commensal microflora. This disruption may include an overreaction to non-virulent gut viruses, bacteria, certain foods, and other luminal antigens, ultimately leading to mucosal inflammation[1,2].
Many of the treatments of CD are based on new biological therapies, such as monoclonal antibodies, which target pro-inflammatory cytokines to the affected organ, as well the release of factors that may modulate cellular immune responses[3,4]. In recent years other new cell therapy approaches of for treatment of IBD have been proposed, such as intravenous administration of adult mesenchymal stromal bone-marrow-derived stem cells (BM-MSC)[5,6,7]. But the exact mechanism behind the proposed benefits of these cell therapies remains unclear and the observed effects of the f-hPSCs injections seem to be indirect. There is no evidence indicating that locally injected stem cells directly differentiate into the building blocks of damaged tissues in the gastrointestinal tract[8,9,10]. Therefore, the repair of the damaged tissues by integration of implanted cells is ruled out[11]. Futheremore, studies have also demonstrated that in systemic intravenous administration the cells tend to be trapped in micro-vessels in the first pass, especially in the lungs, immediately after their injection, rather than migrate to the targeted damaged tissues[12,13].
MSCs delivery in human studies for alleviation of CD are generally based on the expected indirect modulating effect of inflammatory processes, resulting in a significant disease remission, including notable mucosal healing[14]. This effect appeared to be associated with the pro-regenerative secretome of the MSCs[15,16,17,18]. Local MSCs treatments in human studies of CD seemed to control indirectly the inflammatory processes, leading to modest remission of the disease, including notable mucosal healing. This suggests an indirect influence, possibly through the induction of the secretion of anti-inflammatory cytokines and pro-regenerative growth factors by these cells, both in a soluble form or as exosomes[12,13,15,16,19,20,21].
Human placenta tissues provide a readily accessible source of large quantities of expanded mesenchymal stromal cells (MSCs)[22]. Commercial preparations of placental stromal cells were derived from fetal placental tissues have demonstrated a greater ability to induce pro-regenerative effects[12,16,22,23,24]. We isolated and expanded allogeneic fetal only human stromal placental cells (f-hPSCs) from the connective tissues of the chorionic plate of full-term fetus and further expanded them by 6-8 passages before their injection[22,25]. During normal pregnancy, the allogeneic non-matched fetal part of the placentae which reside side by side with the allogneic maternal placental tissues are well tolerated and minimally rejected by the maternal immune system throughout pregnancy[26]. f-hPSCs expanded in culture also offer a significant advantage over BM-MSCs due to their higher phenotype stability and limited trans-differentiation potential. Therefore, they are not expected to undesired trans-differentiation to other cell phenotypes following their injection. Their potency appears to be linked to their secretome comprising of cytokines and chemokines which also prevent their rejection as allogeneic of xenogeneic cells, while sensing stress and with oneset of a secretome for inducing regeneration of the affected tissues in the target organ before the slow clearance of the cells[9,23]. Upon local or systemic delivery, the f-hPSCs appear to enhance the healing of injured tissues, by the continuous induction of the secretion of pro-regenerative and anti-inflammatory secretome[23,27], thereby promoting indirectly pro-regenerative processes. This effect occur most likely with commercially manufactured cells, which have already show promise in pro-regenerative therapy of various disorders [12,28].
The major advantage of therapy with the cell injection into vascularized tissues, specifically when non-matched f-hPSCs are used, in contrast to the injection of their secretome, is straightforward. As we have previously shown, the f-hPSCs can reside for prolonged period of more than 6 days in the injection site before their clearance. During this time interval they can respond to the systemically circulated host’s stress signals and subsequently secret continuously the factors needed to promote the tissue recovery[23]. We have also demonstrated the advantage of the stromal cells that derive form the fetal placenta tissues, as they seem to be more responsive to such stresses than the maternal placenta cells[12].
As it is not desirable that the allogeneic/ xenogeneic cells reside for longer time intervals in the injection site, though they are relatively immune-tolerated, the cells eventually are cleared away within a few days{Gaberman, 2013 #7180}. Nevertheless, if further treatment is needed, additional boosts of injected cells can be given, as done here and in our previous studies based on f-hPSCs injection[12,25,29].
Flow cytometry (FACS) of the cell surface profile of f-hPSCs demonstrate that these cells have typical mesenchymal stromal cell (MSC) surface markers, with positive expression of CD29 and CD90, and lower expression of CD105. Due to their mesodermal origin, the f-hPSCs were found negative for typical surface markers of hematopoietic cells, such as CD45, CD19, and CD11b and the endothelial marker CD34, as well as HLA-DR and HLA-G[22,23,25]. f-hPSCs were also reported to express CD146 and CD166[30], which are found in the placental stromal cells and are not typical to BM-MSCs. The positive expression of CD276 on the membrane of the f-hPSCs may hint on its possible role in the immune tolerance towards the xenogeneic f-hPSCs implants[27].
2. Methods and Materials
2.1. The Mice Model
C57BL/6JOla mice were used as a model for induced CD. This mouse model was chosen since cyptitis and crypt depletion, as a common feature of IBD in human, were also evident in these mice following DSS-induced colitis [31]. Though both males and females develop robust DSS induced colitis and IBD by this procedure, the males seem to develop more rapidly an aggressive disease than females [32]. Therefore, male mice only were chosen to test the effect of the f-hPSCs therapy. The mice were randomized, housed in 6-8 mice per box, and kept in the strictly clean Specific Pathogen Free (SPF) regulated animal colony. Each mouse was identified by a tag and monitored 5 days a week for its weight and its general condition. The mice were sacrificed at the end of the experiment by IP overdose injection of 0.9 mL of Ketamine (1g/ml) and 0.1ml Xylazine (20mg/ml)+1ml normal saline, followed by immediate invivo blood collection from the heart. Then the colon was dissected, rinsed and sampled for general histology by H&E staining and immunohistology, as now described in detail.
Four experimental groups were tested. IBD was induced in 3 of the groups by DSS exposures, one group of 12 mice were not treated for IBD, 11 mice exposed to DSS were treated IP with f-hPSCs and additional small control group of 6 mice were treated IM.
2.2. f-hPSCs Preparation and Their Cell Surface Phenotype
The f-hPSCs were isolated from donated placentae of planned caesarean section with male offspring with informed consent of the parents. The tissue sample of the fetal tissue were dissected and tested for their specific MSCs phenotype. (Figure 1A). The cell surface markers of the f-hPSCs used were confirmed as previously described in detail [22,25]. The subsequent f-hPSCs population used were proven to be from pure fetal source. Since placentae from only male offspring were used, the source of the expanded cells were 100% fetal, as verified by testing their male X-Y fetus karyotype by FISH assay[22,29].
2.3. DSS Induced IBD
The mice with induced IBD were supplied with drinking water with 3.5% DSS. To examine the impact of f-hPSCs on the inflammatory and regenerative processes in the intestine, four experimental groups were studied: three groups received 3.5% DSS in water for 4 days, while one group served as the control group. On days 4 and 6, f-hPSCs were administered either intramuscularly (IM) or intraperitoneally (IP) to the groups exposed to DSS, while no cells were injected into the non-cell-treated control group. The mice were monitored daily, and in case of apparent signs of dehydration, they were given subcutaneous injections of normal saline, while wetted food was placed in their cages. All mice were sacrificed on day 16. The donated placenta from which the tissues for f-hPSC were harvested were donated with informed consent with the approval of the institutional Ethical committee for Human Experimet #0361-14HMO. The mice experiment were planned and prerformed with the approval of the Institutional animal welfare committee of Hebrew-University-Hadassah Medical Center #MD-15-14388-5.
2.4. f-hPSCs Injections
The f-hPSCs (2*106 cells) were prepared for injection by suspending them in 0.2ml Plasmalite buffered solution for injection on day 4 and 6: one small ontrol group was injected IM and the other by IP administration. The selection of the optimal cells number chosen for each injection is based on previous experimental work in which the cells were successfully tested for their effect for other pro-regenerative and anti-inflammatory indications in mice[12,23,29].
2.5. Blood Cell Count (CBC)
The mice were euthanized by the IP injection of an overdose of 0.9 mL of Ketamin (1g/ml) and 0.1ml Xylazine (20mg/ml)+1ml Salin IV. Then, immediately under their overdose anaesthesia, blood for CBC analyses was drawn from the living mice directly from the hearts by a fine syringe into dedicated blood collection micro-tubes containing EDTA. CBC analysis was performed using BC-2800Vet Mindray Hematology Analyzer (GuangZhou City, PRC), calibrated specifically for differential blood counts in mice. A short colon sample adjacent to the caecum was dissected and sampled for histology of H&E staning and immunohistology, as described below.
2.6. Assessment of Inflammatory Process by Mesenteric Lymph Nodes (MLN) WBC Cellularity
The MLN were collected from the sacrificed mice, transferred to low glucose DMEM and fully smashed by a plastic syringe through a mesh. The collected cells were subsequently re-suspended in DMEM and counted by Countess II Cell Counter (Life technologies, Thermo-Fisher, USA).
2.7. Assessment of the Severity of Inflammatory Processes by Neutrophil Activity by Myeloperoxisdase (MPO) Assay in Colon Samples
The colons collected from the euthenized mice were washed and their content was thoroughly rinsed with saline solution using a Venflon tubing. The colons were weighed . Then a short colon sample of ~2 mm, adjacent to the caecum, was cut off and reserved for histological analysis. The rest of the colons were homogenized for 5 minutes on ice in 1 ml of 0.5% hexadecyltrimethylammonium bromide (HATB, Sigma #SLBK3798V) using a blade homogenizer. The homogenized tissue was collected into small tubes and underwent twice 2 freeze and thaw cycles. The single cells in the test-tubes were further dissociated by high power sonication for 15 sec. Then the samples were centrifuged (13,000g) for 15 min in ~4°C and the supernatant collected. 10µl of the supernatant from each sample were taken into 96 wells plate. Calibration solutions and samples were added to 96-well plate. 290µl of O-dianisidine (Sigma Lot #MKBP7560V) were then added to each well for initiation of the MPO assay. The OD of the samples were read immediately with Sunrise Tecan, plate reader (USA) at 460 nm. The results (in unit/mg) were normalized to the initial weight of the samples.
2.8. Colon Histology
The colon tissue samples for histology were kept at room temperature for 24 hours in 4% formaldehyde. The samples were then preserved in 70% ethanol before their further processing. The fixed samples were embedded in paraffin, cut to 4µM sections and stained by H&E.
Immuno-histochemical CD68 staining to count tissue macrophages (Mɸ) density was performed on 4µm sections using the Leica Bond III system (Leica Biosystems Newcastle Ltd, UK). The slides were pretreated with epitope-retrieval solution for 10 minutes (ER2, Leica Biosystems Newcastle Ltd, UK), followed by 30 minutes incubation with a rabbit anti CD68 primary antibody (Abcam, ab125212, 1:1500). Leica Refine-HRP kit (Leica Biosystems Newcastle Ltd, UK) was used before counter-staining with Hematoxylin. Histological samples were photographed sequentially at various magnifications. These full section photographs were subsequently utilized for quantitative assessment of Mɸ prevalence and for objective, blinded pathological expert scoring.
The histological samples were photographed in sequential pictures at different magnifications for quantitative evaluation of Mɸ prevalence and for objective blind pathological expert scoring. For the presentation of a conclusive view of the full area of the gut cross-section, series of the sequential photographs of medium magnification (mag x 100-200) where taken. The series of such pictures of the same magnification were combined and stitched together by photo-editing program to yield a high resolution large photographic record as an overview of a whole area of the gut section, from which specific regions of interest are presented with different magnifications.
The prevalence of immune HRP CD68 Mɸ was evaluated by counting the brown stained cells with anti-CD68 conjucated with HRP in the tissue section with Nikon’s Element software (NIS) which detected the atined cells of interest. The dark brown stained cells were identified in the areas of interest of the sections. The results and are presented then as total number of cells/total combined area of the crypts in the section.
2.9. Statistics
All graphs designs and statistical analyses were conducted with the aid of Microsoft Excel's statistical software package. Statistical significance assessments were carried out with th use of standard paired Student's t-test to compare the results among different experimental arms.
3. Results
3.1. Characterization of the f-hPSCs
The f-hPSCs used were of the fetal placenta tissue of male offspring and their source from only the fetal tissue was verified by their X-Y katyotype[22,29]. FACS analysis of cell surface markers of the expanded f-hPSCs confirmed their typical mesenchymal stromal cells (MSC) markers. As previously reported, the cells were found positive for mesenchymal markers CD29 and CD90, and partially positive for CD105 expression, a marker prominently expressed in many other mesenchymal stromal cells[12,22,25] (Figure 1A). Additional markers positively expressed on f-hPSCs included CD146 and CD166, typically reported to be present in chorionic hPSCs[33]. Similar to stromal cells isolated from other mesenchymal tissues, the f-hPSCs were found negative for hematopoietic markers CD45, CD19, and CD11b. The expression of HLA-DR and HLA-G in f-hPSCs was also negative, as was the expression of the endothelial cell marker CD34[12]. (Figure 1A).
3.2. DSS Induced of IBD
The experimental setup for IBD induction in mice is presented graphically in Figure 1B. Three different groups tested were exposed to 3.5% DSS in their drinking water for four days. The f-hPSCs for injection were trypsinized from nearly confluent cultures washed once and counted, as previously described[25]. 2*106 f-hPSCs were delivered on days 4 and 6, either intramuscularly (IM) or intra-peritoneally (IP). In the untreated group, no cells were injected. During the experiment, mice exposed to DSS experienced a significant weight loss (Figure 1C), but none of them in any of the experimental groups died during the exposure period. All DSS-exposed groups showed continuous weight loss between days 0-8 after the onset of IBD. The non-treated group and the group treated with IM f-hPSCs lost approximately 25% of their body weight, with no significant weight regain by the end of the 16 days follow-up. In contrast, mice injected IP with f-hPSCs experienced significantly less weight loss by day 8 (approximately 15%, p<0.01). These mice also showed significant weight recovery towards their initial weight by the end of the 16-day follow-up.
3.3. The Influence of f-hPSCs Injection on the Mice Colon
At the conclusion of the follow-up on day 16, the mice were euthanized and their colons were dissected, cleansed, and measured. An extensive inflammatory process, colon shortening, which are characterized inflammatory colitis, and an increase of colon mass. A significant colon shortening was observed in the non-treated DSS-exposed group, whereas the colon length in the DSS-exposed f-hPSCs-treated groups did not differ significantly from that of the naïve controls (Figure 1C). However, the most notable changes in colon mass were demonstrated by the colon weight-to-length ratio (Figure 1D). The colon mass per unit length increased significantly in all mice exposed to DSS, with the greatest increase observed in the non-treated group. Treatment with f-hPSCs notably reduced the mass increase in the inflamed DSS-induced colons. In the IP-treated mice, the average colon mass per length was significantly lower by approximately 30%, compared to the non-cell-treated group, though it remained slightly higher, albeit not significantly, compared to the controls (Figure 1D).
3.4. The Influence of f-hPSCs Treatments on the Total MLN Cell Counts
The cell numbers harvested from the MLN, consist primarily of lymphocytes, showed a substantial increase in all experimental groups following DSS consumption, compared to naïve controls. The highest values were observed in f-hPSCs treated mice via intramuscular (IM) administration, reaching levels comparable to those in untreated controls (Fig.1E).
3.5. Effect of the DSS and f-hPSCs Treatments on the Peripheral Blood Cells Counts
A small non-significant decrease in RBC counts was observed at the end of the experiment across all experimental arms of the DSS-exposed mice (Figure 2A), most likely due to bleeding in the inflamed GI tract caused by DSS exposure. However, no significant differences between the f-hPSCs-treated and non-treated mice were noted. Platelet counts were moderately elevated in all DSS-exposed mice, with no significant differences between the f-hPSCs-treated and non-treated groups (Figure 2B). Total white blood cell (WBC) counts, indicative of significant systemic inflammatory processes, increased significantly in all DSS-exposed groups, with slightly higher counts in the non-treated group, compared to the f-hPSCs-injected groups (Figure 2D). Similarly, elevated lymphocyte counts were observed in both the non-treated and IM-f-hPSCs-treated DSS-exposed mice, while counts in the IP f-hPSCs-treated group were comparable to the naïve controls and similar to the non-DSS-treated group. Monocyte counts were elevated in all DSS-exposed mice relative to the naïve controls (Figure 2E). The differences in granulocyte counts between the groups was more apparent, with the non-treated DSS-exposed group showing the highest monocyte counts, compared to less elevated counts in DSS-exposed mice treated with f-hPSCs via IM or IP injection (Figure 2F).
3.6. Colon Histology
Histological sections of the colon performed on H&E colon paraffin sections of the different arms tested are shown in Figure 3A-D with further magnification of a representative region in each arm, marked in the detailed low magnification microphotographs with a black rectangle. A representative section of the colon of a naïve mouse is shown in Figure 3A.
In the section of non-treated mice (Figure 3B), the crypts are seen severely damaged or depleted, and were replaced with inflammatory reaction. The histological colon sections of IM and IP f-hPSCs treated mice are shown in Figure 3C&D, respectively. Major regeneration processes are seen in the DSS-exposed mice treated with f-hPSCs injections, with apparently best effect in those treated IP.
Based on the histological evaluation of colon sections, the main histological parameters presented in the table in Figure 4A were scored, using a scale of 0-4 through an objective, blinded assessment by a pathology expert. The parameters evaluated included the extent of regenerating tissue, the degree of crypt damage, and the percentage of tissue involved in the degeneration process. The score for inflammatory processes was found to be higher in all DSS exposed group, with the highest values in non-treated mice. The f-hPSCs-treated groups showed somewhat lower scores, although the differences were not statistically significant.
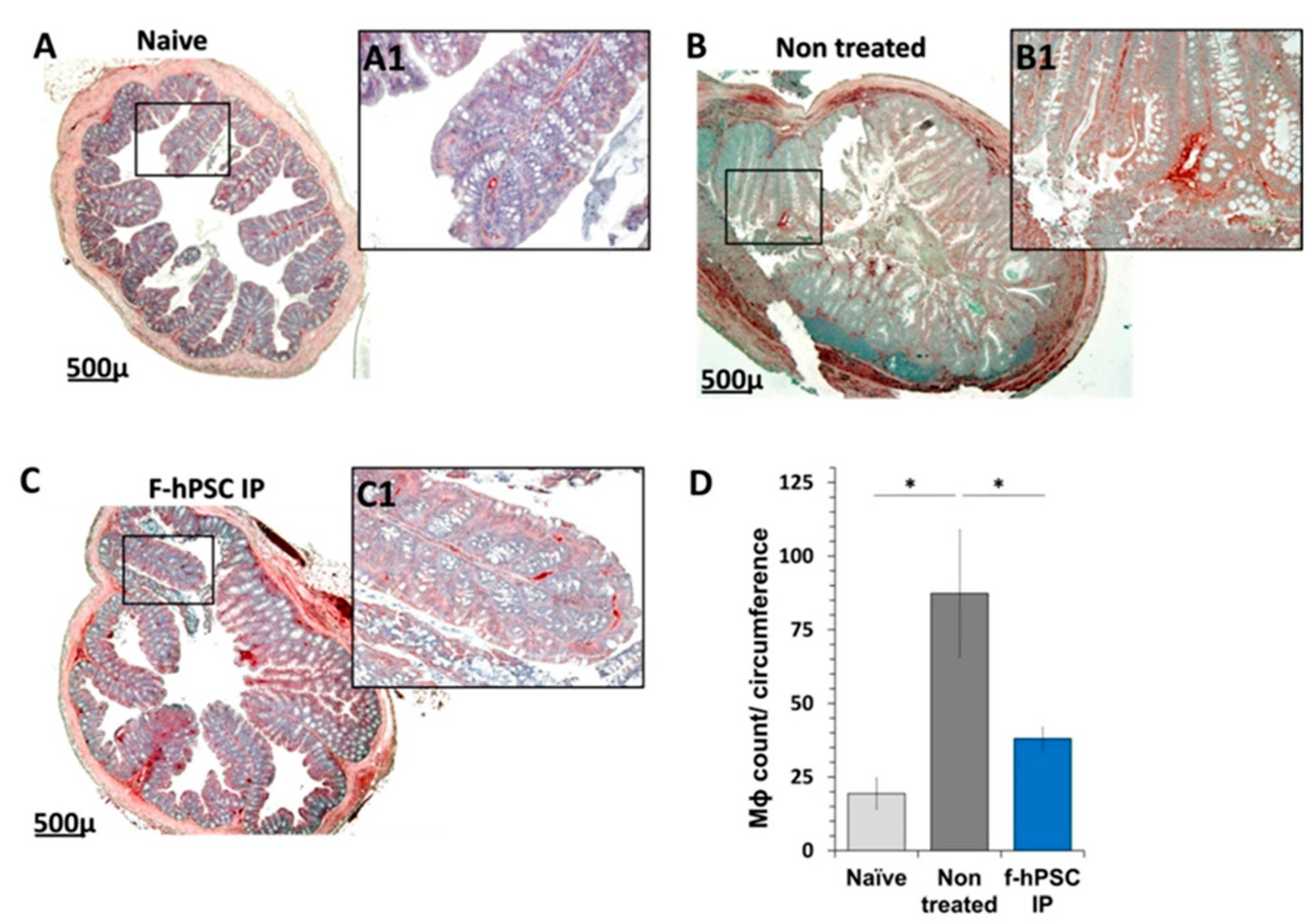
The counting of the CD68 immune-stained Mɸ in the area of the crypts in the sections is presented in Figure 5. A significant elevation in the prevalence of the Mɸ in the crypts area was recorded in the DSS exposed and f-hPSCs injected and mice animals, as well as untreated, while their number was significantly reduced towards normal values in the f-hPSCs treated DSS exposed mice.
3.7. MPO Assay for Neutrophils Density
Another parameter associated with inflammation is the density of neutrophils in the colon as evaluated by the MPO enzymatic reaction activity assay. The maximum reading was reached within 30 minutes. In all groups of mice exposed to DSS, the level of MPO in the colon at the end of the follow-up was increased significantly relative to the control group, indicating the recruitment of neutrophils to the inflamed colon tissues (Fig 4C). The MPO activity in the group IP injected with f-hPSCs was slightly but not statistically decreased (Figure 4C). But a high correlation (f=0.98) was observed between the MPO levels in the tissues and the histological score in the different groups tested (Figure 4D).
The inflammatory processes in the colon involved an increased Mɸ infiltration of into the affected crypt tissues. To assess the increase in the number of CD68-immunostained Mɸ and their distribution in the tissues, their numbers was measured per the total crypts in gut section circumference for each mouse (Figure 5A-C, with magnifications of the regions of interest (A1, B1 & C1, respectively). In the control group (Figure 5A and A1), fewer Mɸ are observed in the crypts, primarily located their periphery. In contrast, in non-cell treated mice with inflamed colon, a high number of Mɸ are present in both the crypts and the submucosa (see Figure 4B and B1). Following IP f-hPSCs treatment (Figure 5C and 5C1), a significant reduction in Mɸ density per gut circumference relative to untreated mice was observed, reaching almost the normal cell counts (Figure 5D).
4. Discussion
The over-reaction of the immune system in the GI tract in CD, possibly mediated by activated CD4(+)-T cells lymphocytes under Th17 differentiating conditions, has been proposed as the mechanism of induction of the undesired inflammatory reaction associated with IBD [34,35].
Cell therapies with different types of potent/stem cells have already been proposed as effective pro-regenerative therapy. In most of the related studies, the effect of the cells was found to be predominantly indirectly mediated through a vast release of different pro-regenerative and anti-inflammatory factors[23]. In many of previously reported studies on PSCs-based cell therapy the cells were injected systemically by IV delivery[6,36,37] and not by IP injections, as we propose here. Only in a few of these studies, the critical issue of the exact fetal tissue source of the placenta, from where our cells were isolated was referred to[6,36,37,38]. It is expected that most of previous studies, where no clear indication was given on the exact tissue source used, the stromal cells from the maternal placental tissues were probably used, which is the predominant stromal cells population upon their isolation from whole placenta. This may be a critical issue as we previously demonstrated the therapeutic advantage and the higher potency of f-hPSCs isolated from only the fetal placental tissues[12,39]. This may derive from the role of such cells to support the growing fetus and supply its needs during its development, while the role of the maternal placenta, which is not connected directly to the blood supply of the fetus, is to feed the fetal placental tissues.
As adult fully differentiated cells, the f-hPSCs are not “stem cells” by definition. Due to their negligible potential for trans-differentiation they are not expected to serve as potential building blocks of the regenerating tissues[12,23,40]. Furthermore, no specific homing is expected in the systemic injection of these cells. When injected into tissues or body spaces, the cells reside temporarily in the injection niche until their clearance with minimal complications. During that time interval they respond to blood born stress factors by releasing pro-regenerative and anti-inflammatory secretome[12,23,41,42]. The option of f-hPSCs injection via IV administration was not explored since IV-injected cells are expected to become trapped in the lung microvascular system during their first pass until their clearance, with possible related complications[13]. The f-hPSCs derived exclusively from fetal placenta tissue and are expected to mediate highly efficient anti-inflammatory and pro-regenerative effects. The f-hPSCs, were found to have negligible rejection rate for longer time intervals in the injected tissues, allowing anextended release of their secretome, before being cleared away[27,43,44,45],
The f-hPSCs from only the fetal placenta were shown to express low levels of MHC class I antigens. They lack MHC class II on top of their CD276 expression, which may contribute to the reduction of their immune rejection[27,46,47,48,49]. This unique expression profile allows these placental cells confront immune detection and maternal-fetal tolerance during pregnancy, with their reported secretion of immune-regulatory factors, such as transforming growth factor-beta (TGF-β), indoleamine 2,3-dioxygenase (IDO), and interleukin-10 (IL-10)[44,50] that may help to suppress inflammatory responses and promote tissue repair.
We showed here that IP injection (and in a lesser degree IM injection) of f-hPSCs alleviated the inflammatory IBD induced by DSS, as expressed by lesser colon shortening and reduction of the number of circulating granulocytes and significant improvement in most other inflammation related parameters tested. These include the reduction of inflammation-associated increase of the colon mass, regain of weight and reduced MPO activity in the gut, reduction in the number of infiltrating Mɸ counted in the colon histological section leading to improved histological scoring in the regenerative process of the affected gut.
The results of our exploratory study, which examined the use of xenogeneic f-hPSCs, injected IM or IP, as an indirect treatment of experimental induced IBD in a mouse model. suggest that this non-invasive safe treatment could be considered as a cell therapy which could be applied for human IBD. The results further confirm the previously described findings on the indirect effect of f-hPSCs treatments in the induction of significant anti-inflammatory and pro-regenerative effects in other similar conditions[12,23,25,29,45].
In conclusion, our findings suggest that the intraperitoneal (IP) administration of f-hPSCs is significantly more effective as an anti-inflammatory and pro-regenerative treatment compared to intramuscular (IM) administration. Based on these results, we propose that xenogeneic or allogeneic f-hPSCs, cultured under properly regulated conditions, could be a serve as a safe and effective cell therapy for a treatment for alleviating IBD.
Further studies should establish and prove that mesenchymal cells isolated from the fetal placental tissues may be more immuno-competent[51,52,53]. The mechanism of the indirect pro-regenerative effect of-fhPSC in alleviating inflammatory disorders may be complex and may deserve further investigation, from growth factors and anti-inflammatory agents secretion to indirect effects of secreted exosomes[23,54,55]. These studies may apply to the outcome of allogeneic f-hPSCs injections in the reduction of their rejection and their effect in different other disease models.
Clinical studies have already examined safety and tolerability on different PSCs preparations isolated from postpartum placenta and injected twice IV with different numbers of allogeneic cells. Generally, As previously shown[6], the cells seemed to be well tolerated, with minor adverse effects. Therefore, the need for corticosteroids to prevent rejection during the short interval activity of the cells following their injection seemed to be redundant, in view of their low allogeneic or xenogeneic rejection and the short time interval needed to exert their effect. Phase1b/ 2a study already established the safety and tolerability of 1.5-6x108 injected placental cells, associated with clinical improvement, reduction of inflammatory markers, such as CRP, with no significant adverse event[9].
In conclusion, our study showed the potency of IP f-hPSCs treatment in alleviating induced IBD in a mouse model. We found that though the effect of the cells seems to be indirect, IP cells injection adjacent to the damaged organ seems to have some therapeutic advantage over IM injections.
Funding
The project was supported in part by a research grant of the Israeli Science Foundation (ISF# 1639/14) to RG Hadassah-Hebrew University Research Funds
Acknowledgments
We wish to thank Dr. Mamoun Basheer, Nerel Cohen and Claudia Moressi for their technical experimental laboratory work help in some of the experiments. We also thank Zohar Gavish for his help in part of the immunohistological preparations.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hold, G. L., Smith, M., Grange, C., Watt, E. R., El-Omar, E. M., and Mukhopadhya, I. (2014) Role of the gut microbiota in inflammatory bowel disease pathogenesis: what have we learnt in the past 10 years?, World journal of gastroenterology 20, 1192-12.10.
- Strober, W. (2013) Impact of the gut microbiome on mucosal inflammation, Trends Immunol 34, 423-430.
- Singh, S., Andersen, N. N., Andersson, M., Loftus, E. V., Jr., and Jess, T. (2018) Comparison of infliximab with adalimumab in 827 biologic-naive patients with Crohn's disease: a population-based Danish cohort study, Alimentary pharmacology & therapeutics 47, 596-604.
- Feagan, B. G., Rutgeerts, P., Sands, B. E., Hanauer, S., Colombel, J. F., Sandborn, W. J., Van Assche, G., Axler, J., Kim, H. J., Danese, S., Fox, I., Milch, C., Sankoh, S., Wyant, T., Xu, J., Parikh, A., and Group, G. S. (2013) Vedolizumab as induction and maintenance therapy for ulcerative colitis, The New England journal of medicine 369, 699-710.
- Mishra, R., Dhawan, P., Srivastava, A. S., and Singh, A. B. (2020) Inflammatory bowel disease: Therapeutic limitations and prospective of the stem cell therapy, World journal of stem cells 12, 1050-1066.
- Mayer, L., Pandak, W. M., Melmed, G. Y., Hanauer, S. B., Johnson, K., Payne, D., Faleck, H., Hariri, R. J., and Fischkoff, S. A. (2013) Safety and tolerability of human placenta-derived cells (PDA001) in treatment-resistant crohn's disease: a phase 1 study, Inflammatory bowel diseases 19, 754-760.
- Pak, S., Hwang, S. W., Shim, I. K., Bae, S. M., Ryu, Y. M., Kim, H. B., Do, E. J., Son, H. N., Choi, E. J., Park, S. H., Kim, S. Y., Park, S. H., Ye, B. D., Yang, S. K., Kanai, N., Maeda, M., Okano, T., Yang, D. H., Byeon, J. S., and Myung, S. J. (2018) Endoscopic Transplantation of Mesenchymal Stem Cell Sheets in Experimental Colitis in Rats, Scientific reports 8, 11314.
- Liao, Y., Lei, J., Liu, M., Lin, W., Hong, D., Tuo, Y., Jiang, M. H., Xia, H., Wang, M., Huang, W., and Xiang, A. P. (2016) Mesenchymal Stromal Cells Mitigate Experimental Colitis via Insulin-like Growth Factor Binding Protein 7-mediated Immunosuppression, Molecular therapy : the journal of the American Society of Gene Therapy 24, 1860-1872.
- Melmed, G. Y., Pandak, W. M., Casey, K., Abraham, B., Valentine, J., Schwartz, D., Awais, D., Bassan, I., Lichtiger, S., Sands, B., Hanauer, S., Richards, R., Oikonomou, I., Parekh, N., Targan, S., Johnson, K., Hariri, R., and Fischkoff, S. (2015) Human Placenta-derived Cells (PDA-001) for the Treatment of Moderate-to-severe Crohn's Disease: A Phase 1b/2a Study, Inflamm Bowel Dis 21, 1809-1816.
- Trebol, J., Georgiev-Hristov, T., Pascual-Miguelanez, I., Guadalajara, H., Garcia-Arranz, M., and Garcia-Olmo, D. (2022) Stem cell therapy applied for digestive anastomosis: Current state and future perspectives, World journal of stem cells 14, 117-141.
- Caplan, A. I. (2017) Mesenchymal Stem Cells: Time to Change the Name!, Stem cells translational medicine 6, 1445-1451.
- Gaberman, E., Pinzur, L., Levdansky, L., Tsirlin, M., Netzer, N., Aberman, Z., and Gorodetsky, R. (2013) Mitigation of Lethal Radiation Syndrome in Mice by Intramuscular Injection of 3D Cultured Adherent Human Placental Stromal Cells, PloS one 8, e66549.
- Lee, R. H., Pulin, A. A., Seo, M. J., Kota, D. J., Ylostalo, J., Larson, B. L., Semprun-Prieto, L., Delafontaine, P., and Prockop, D. J. (2009) Intravenous hMSCs improve myocardial infarction in mice because cells embolized in lung are activated to secrete the anti-inflammatory protein TSG-6, Cell stem cell 5, 54-63.
- Pampalone, M., Corrao, S., Amico, G., Vitale, G., Alduino, R., Conaldi, P. G., and Pietrosi, G. (2021) Human Amnion-Derived Mesenchymal Stromal Cells in Cirrhotic Patients with Refractory Ascites: A Possible Anti-Inflammatory Therapy for Preventing Spontaneous Bacterial Peritonitis, Stem cell reviews and reports 17, 981-998.
- Duan, L., Huang, H., Zhao, X., Zhou, M., Chen, S., Wang, C., Han, Z., Han, Z. C., Guo, Z., Li, Z., and Cao, X. (2020) Extracellular vesicles derived from human placental mesenchymal stem cells alleviate experimental colitis in mice by inhibiting inflammation and oxidative stress, International journal of molecular medicine 46, 1551-1561.
- Eiro, N., Fraile, M., Gonzalez-Jubete, A., Gonzalez, L. O., and Vizoso, F. J. (2022) Mesenchymal (Stem) Stromal Cells Based as New Therapeutic Alternative in Inflammatory Bowel Disease: Basic Mechanisms, Experimental and Clinical Evidence, and Challenges, International journal of molecular sciences 23.
- Fu, Y., Zhang, C., Xie, H., Wu, Z., Tao, Y., Wang, Z., Gu, M., Wei, P., Lin, S., Li, R., He, Y., Sheng, J., Xu, J., Wang, J., and Pan, Y. (2023) Human umbilical cord mesenchymal stem cells alleviated TNBS-induced colitis in mice by restoring the balance of intestinal microbes and immunoregulation, Life sciences 334, 122189.
- Ke, C., Biao, H., Qianqian, L., Yunwei, S., and Xiaohua, J. (2015) Mesenchymal stem cell therapy for inflammatory bowel diseases: promise and challenge, Current stem cell research & therapy 10, 499-508.
- Lightner, A. L., Irving, P. M., Lord, G. M., and Betancourt, A. (2024) Stem Cells and Stem Cell-Derived Factors for the Treatment of Inflammatory Bowel Disease with a Particular Focus on Perianal Fistulizing Disease: A Minireview on Future Perspectives, BioDrugs 38, 527-539.
- Bandzar, S., Gupta, S., and Platt, M. O. (2013) Crohn's disease: a review of treatment options and current research, Cellular immunology 286, 45-52.
- Cao, X., Duan, L., Hou, H., Liu, Y., Chen, S., Zhang, S., Liu, Y., Wang, C., Qi, X., Liu, N., Han, Z., Zhang, D., Han, Z. C., Guo, Z., Zhao, Q., and Li, Z. (2020) IGF-1C hydrogel improves the therapeutic effects of MSCs on colitis in mice through PGE(2)-mediated M2 macrophage polarization, Theranostics 10, 7697-7709.
- Adani, B., Basheer, M., Hailu, A. L., Fogel, T., Israeli, E., Volinsky, E., and Gorodetsky, R. (2019) Isolation and expansion of high yield of pure mesenchymal stromal cells from fresh and cryopreserved placental tissues, Cryobiology 89, 100-103.
- Pinzur, L., Akyuez, L., Levdansky, L., Blumenfeld, M., Volinsky, E., Aberman, Z., Reinke, P., Ofir, R., Volk, H. D., and Gorodetsky, R. (2018) Rescue from lethal acute radiation syndrome (ARS) with severe weight loss by secretome of intramuscularly injected human placental stromal cells, J Cachexia Sarcopenia Muscle 9, 1079-1092.
- El-Nakeep, S. (2022) Stem Cell Therapy for the Treatment of Crohn's Disease; Current Obstacles and Future Hopes, Current stem cell research & therapy 17, 727-733.
- Volinsky, E., Lazmi-Hailu, A., Cohen, N., Adani, B., Faroja, M., Grunewald, M., and Gorodetsky, R. (2020) Alleviation of acute radiation-induced bone marrow failure in mice with human fetal placental stromal cell therapy, Stem Cell Res Ther 11, 337.
- Bai, K., Li, X., Zhong, J., Ng, E. H. Y., Yeung, W. S. B., Lee, C. L., and Chiu, P. C. N. (2021) Placenta-Derived Exosomes as a Modulator in Maternal Immune Tolerance During Pregnancy, Frontiers in immunology 12, 671093.
- Amend, B., Buttgereit, L., Abruzzese, T., Harland, N., Abele, H., Jakubowski, P., Stenzl, A., Gorodetsky, R., and Aicher, W. K. (2023) Regulation of Immune Checkpoint Antigen CD276 (B7-H3) on Human Placenta-Derived Mesenchymal Stromal Cells in GMP-Compliant Cell Culture Media, International journal of molecular sciences 24.
- Shapira, I., Fainstein, N., Tsirlin, M., Stav, I., Volinsky, E., Moresi, C., Ben-Hur, T., and Gorodetsky, R. (2016) Placental Stromal Cell Therapy for Experimental Autoimmune Encephalomyelitis: The Role of Route of Cell Delivery, Stem cells translational medicine.
- Adani, B., Sapir, E., Volinsky, E., Lazmi-Hailu, A., and Gorodetsky, R. (2022) Alleviation of Severe Skin Insults Following High-Dose Irradiation with Isolated Human Fetal Placental Stromal Cells, International journal of molecular sciences 23.
- Abumaree, M. H., Al Jumah, M. A., Kalionis, B., Jawdat, D., Al Khaldi, A., AlTalabani, A. A., and Knawy, B. A. (2013) Phenotypic and functional characterization of mesenchymal stem cells from chorionic villi of human term placenta, Stem cell reviews 9, 16-31.
- Perse, M., and Cerar, A. (2012) Dextran sodium sulphate colitis mouse model: traps and tricks, Journal of biomedicine & biotechnology 2012, 718617.
- Chassaing, B., Aitken, J. D., Malleshappa, M., and Vijay-Kumar, M. (2014) Dextran sulfate sodium (DSS)-induced colitis in mice, Current protocols in immunology 104, Unit 15 25.
- Abomaray, F. M., Al Jumah, M. A., Alsaad, K. O., Jawdat, D., Al Khaldi, A., AlAskar, A. S., Al Harthy, S., Al Subayyil, A. M., Khatlani, T., Alawad, A. O., Alkushi, A., Kalionis, B., and Abumaree, M. H. (2016) Phenotypic and Functional Characterization of Mesenchymal Stem/Multipotent Stromal Cells from Decidua Basalis of Human Term Placenta, Stem cells international 2016, 5184601.
- Galvez, J. (2014) Role of Th17 Cells in the Pathogenesis of Human IBD, ISRN inflammation 2014, 928461.
- Yi, Q., Wang, J., Song, Y., Guo, Z., Lei, S., Yang, X., Li, L., Gao, C., and Zhou, Z. (2019) Ascl2 facilitates IL-10 production in Th17 cells to restrain their pathogenicity in inflammatory bowel disease, Biochemical and biophysical research communications 510, 435-441.
- Riddell, M. R., Winkler-Lowen, B., Chakrabarti, S., Dunk, C., Davidge, S. T., and Guilbert, L. J. (2012) The characterization of fibrocyte-like cells: a novel fibroblastic cell of the placenta, Placenta 33, 143-150.
- Zhao, Y., Gillen, J. R., Harris, D. A., Kron, I. L., Murphy, M. P., and Lau, C. L. (2014) Treatment with placenta-derived mesenchymal stem cells mitigates development of bronchiolitis obliterans in a murine model, The Journal of thoracic and cardiovascular surgery 147, 1668-1677 e1665.
- Shin, S., Lee, J., Kwon, Y., Park, K. S., Jeong, J. H., Choi, S. J., Bang, S. I., Chang, J. W., and Lee, C. (2021) Comparative Proteomic Analysis of the Mesenchymal Stem Cells Secretome from Adipose, Bone Marrow, Placenta and Wharton's Jelly, International journal of molecular sciences 22.
- Pinzur, L., Akyuez, L, Levdansky L.,Blumenfeld,M, Volinsky,E, Aberman,Z, Reinke,P, Ofir,R, Volk,HD, Gorodetsky,R. (2018) Rescue from lethal acute radiation syndrome (ARS) with severe weight loss by secretome of intramuscularly injected human placental stromal cells, Journal of Cachexia, Sarcopenia and Muscle (in press).
- Lahiani, A., Zahavi, E., Netzer, N., Ofir, R., Pinzur, L., Raveh, S., Arien-Zakay, H., Yavin, E., and Lazarovici, P. (2015) Human placental eXpanded (PLX) mesenchymal-like adherent stromal cells confer neuroprotection to nerve growth factor (NGF)-differentiated PC12 cells exposed to ischemia by secretion of IL-6 and VEGF, Biochimica et biophysica acta 1853, 422-430.
- Petrou, P., Gothelf, Y., Argov, Z., Gotkine, M., Levy, Y. S., Kassis, I., Vaknin-Dembinsky, A., Ben-Hur, T., Offen, D., Abramsky, O., Melamed, E., and Karussis, D. (2016) Safety and Clinical Effects of Mesenchymal Stem Cells Secreting Neurotrophic Factor Transplantation in Patients With Amyotrophic Lateral Sclerosis: Results of Phase 1/2 and 2a Clinical Trials, JAMA neurology 73, 337-344.
- Shapira, I., Fainstein, N., Tsirlin, M., Stav, I., Volinsky, E., Moresi, C., Ben-Hur, T., and Gorodetsky, R. (2017) Placental Stromal Cell Therapy for Experimental Autoimmune Encephalomyelitis: The Role of Route of Cell Delivery, Stem cells translational medicine 6, 1286-1294.
- Adams, K. M., Yan, Z., Stevens, A. M., and Nelson, J. L. (2007) The changing maternal "self" hypothesis: a mechanism for maternal tolerance of the fetus, Placenta 28, 378-382.
- Chang, C. J., Yen, M. L., Chen, Y. C., Chien, C. C., Huang, H. I., Bai, C. H., and Yen, B. L. (2006) Placenta-derived multipotent cells exhibit immunosuppressive properties that are enhanced in the presence of interferon-gamma, Stem cells 24, 2466-2477.
- Gorodetsky, R., and Aicher, W. K. (2021) Allogenic Use of Human Placenta-Derived Stromal Cells as a Highly Active Subtype of Mesenchymal Stromal Cells for Cell-Based Therapies, International journal of molecular sciences 22.
- Maier, C. L., and Pober, J. S. (2011) Human placental pericytes poorly stimulate and actively regulate allogeneic CD4 T cell responses, Arteriosclerosis, thrombosis, and vascular biology 31, 183-189.
- Forbes, G. M. (2017) Mesenchymal Stromal Cell Therapy in Crohn's Disease, Dig Dis 35, 115-122.
- Macholdova, K., Machackova, E., Proskova, V., Hromadnikova, I., and Klubal, R. (2019) Latest findings on the placenta from the point of view of immunology, tolerance and mesenchymal stem cells, Ceska gynekologie / Ceska lekarska spolecnost J. Ev. Purkyne 84, 154-160.
- Magatti, M., De Munari, S., Vertua, E., Gibelli, L., Wengler, G. S., and Parolini, O. (2008) Human amnion mesenchyme harbors cells with allogeneic T-cell suppression and stimulation capabilities, Stem cells 26, 182-192.
- Wang, Y., Zhao, X., Li, Z., Wang, W., Jiang, Y., Zhang, H., Liu, X., Ren, Y., Xu, X., and Hu, X. (2024) Decidual natural killer cells dysfunction is caused by IDO downregulation in dMDSCs with Toxoplasma gondii infection, Commun Biol 7, 669.
- Li, C., Zhang, W., Jiang, X., and Mao, N. (2007) Human-placenta-derived mesenchymal stem cells inhibit proliferation and function of allogeneic immune cells, Cell and tissue research 330, 437-446.
- Liu, W., Morschauser, A., Zhang, X., Lu, X., Gleason, J., He, S., Chen, H. J., Jankovic, V., Ye, Q., Labazzo, K., Herzberg, U., Albert, V. R., Abbot, S. E., Liang, B., and Hariri, R. (2014) Human placenta-derived adherent cells induce tolerogenic immune responses, Clinical & translational immunology 3, e14.
- Wang, Q., Liu, T., Zhang, Y., Chen, D., Wang, L., Li, Y., and Wei, J. (2013) [Immunomodulatory effects of human placental-derived mesenchymal stem cells on immune rejection in mouse allogeneic skin transplantation], Zhongguo xiu fu chong jian wai ke za zhi = Zhongguo xiufu chongjian waike zazhi = Chinese journal of reparative and reconstructive surgery 27, 775-780.
- Kshirsagar, S. K., Alam, S. M., Jasti, S., Hodes, H., Nauser, T., Gilliam, M., Billstrand, C., Hunt, J. S., and Petroff, M. G. (2012). Immunomodulatory molecules are released from the first trimester and term placenta via exosomes, Placenta 33, 982-990.
- Ouyang, Y., Mouillet, J. F., Coyne, C. B., and Sadovsky, Y. (2014) Review: placenta-specific microRNAs in exosomes - good things come in nano-packages, Placenta 35 Suppl, S69-73.
Figure 1.
DSS induced colitis experiment anf f-hPSCs treatment. FACS analysis of the cell surface markers of the IP injected f-hPSCs (A). A follow up of the mice weight in all tested experimental groups (B). Four experimental arms were tested: 3 groups were exposed to 3.5% DSS in drinking water for 4 days. f-hPSCs were injected IM or IP on days 4 and 6. At the termination of the experiment, the colon was excised, weighted and its length measured. The records of the colon mass/length ratio weight/length at the end of the experiment (C). Lymphocytes counts in the MLN. The bars above the columns indicate the significance of the difference between the tested groups (*=P<0.05, **=p<0.01).
Figure 1.
DSS induced colitis experiment anf f-hPSCs treatment. FACS analysis of the cell surface markers of the IP injected f-hPSCs (A). A follow up of the mice weight in all tested experimental groups (B). Four experimental arms were tested: 3 groups were exposed to 3.5% DSS in drinking water for 4 days. f-hPSCs were injected IM or IP on days 4 and 6. At the termination of the experiment, the colon was excised, weighted and its length measured. The records of the colon mass/length ratio weight/length at the end of the experiment (C). Lymphocytes counts in the MLN. The bars above the columns indicate the significance of the difference between the tested groups (*=P<0.05, **=p<0.01).
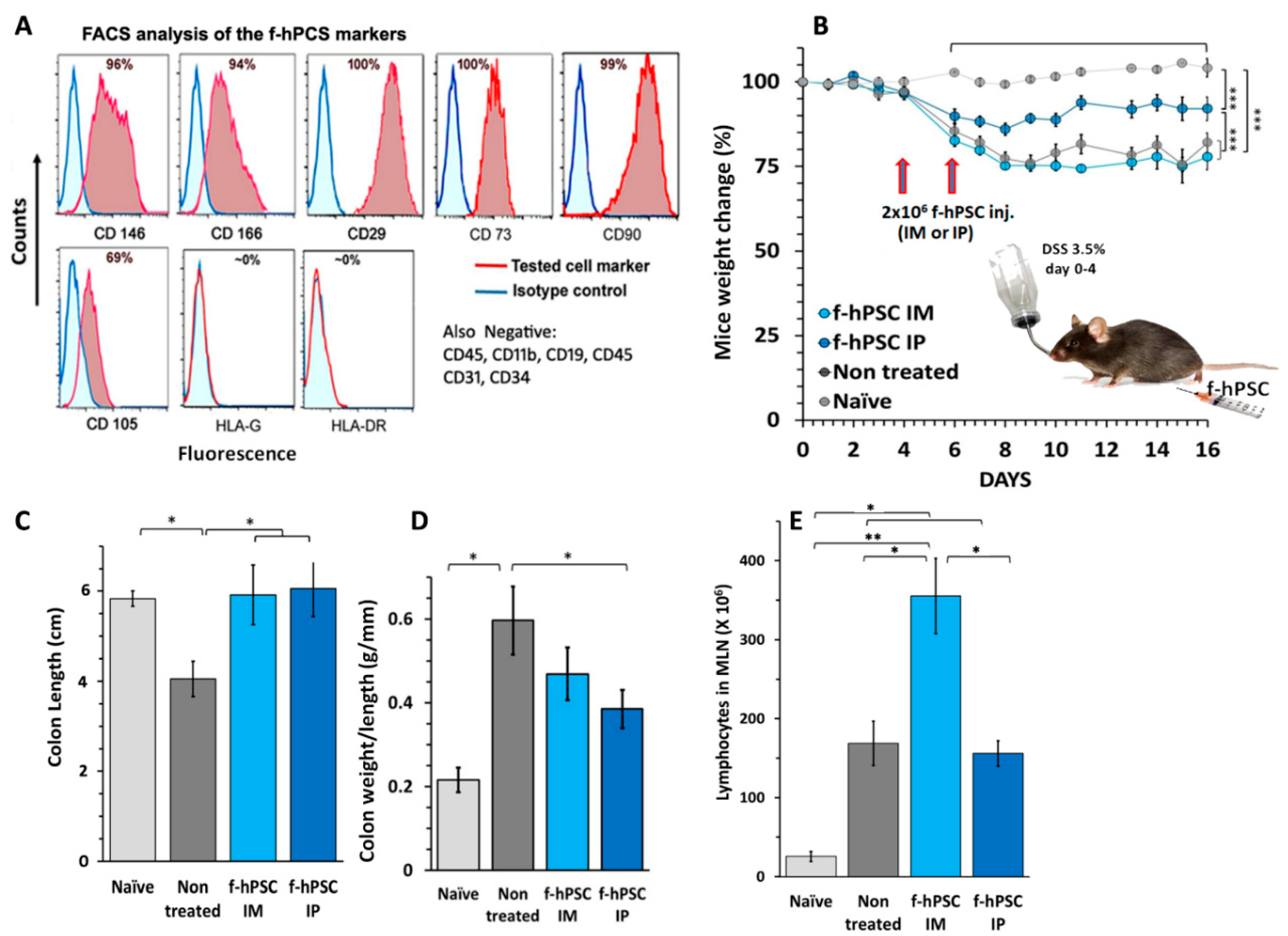
Figure 2.
CBC at the en of the experiment. CBC values performed at the termination of the experiment for the different experimental groups tested are presented. The results of RBC (A) PLT (B) and WBC (C) are presented. The peripheral WBC were found significantly elevated in all the DSS exposed mice. Differential leukocytes counting showed a major decrease of lymphocytes number in f-hPSCs DSS exposed IP treated mice (D) similar significant elevation of monocytes counts were recorded in all DSS exposed mice (E). The most apparent effect of the f-hPSCs treatment is indicated by very significant reduction of the granulocytes, especially in the IP treated mice (F). The bars above columns indicate the significance of the difference between groups tested (* P<0.05, ** P<0.01).
Figure 2.
CBC at the en of the experiment. CBC values performed at the termination of the experiment for the different experimental groups tested are presented. The results of RBC (A) PLT (B) and WBC (C) are presented. The peripheral WBC were found significantly elevated in all the DSS exposed mice. Differential leukocytes counting showed a major decrease of lymphocytes number in f-hPSCs DSS exposed IP treated mice (D) similar significant elevation of monocytes counts were recorded in all DSS exposed mice (E). The most apparent effect of the f-hPSCs treatment is indicated by very significant reduction of the granulocytes, especially in the IP treated mice (F). The bars above columns indicate the significance of the difference between groups tested (* P<0.05, ** P<0.01).
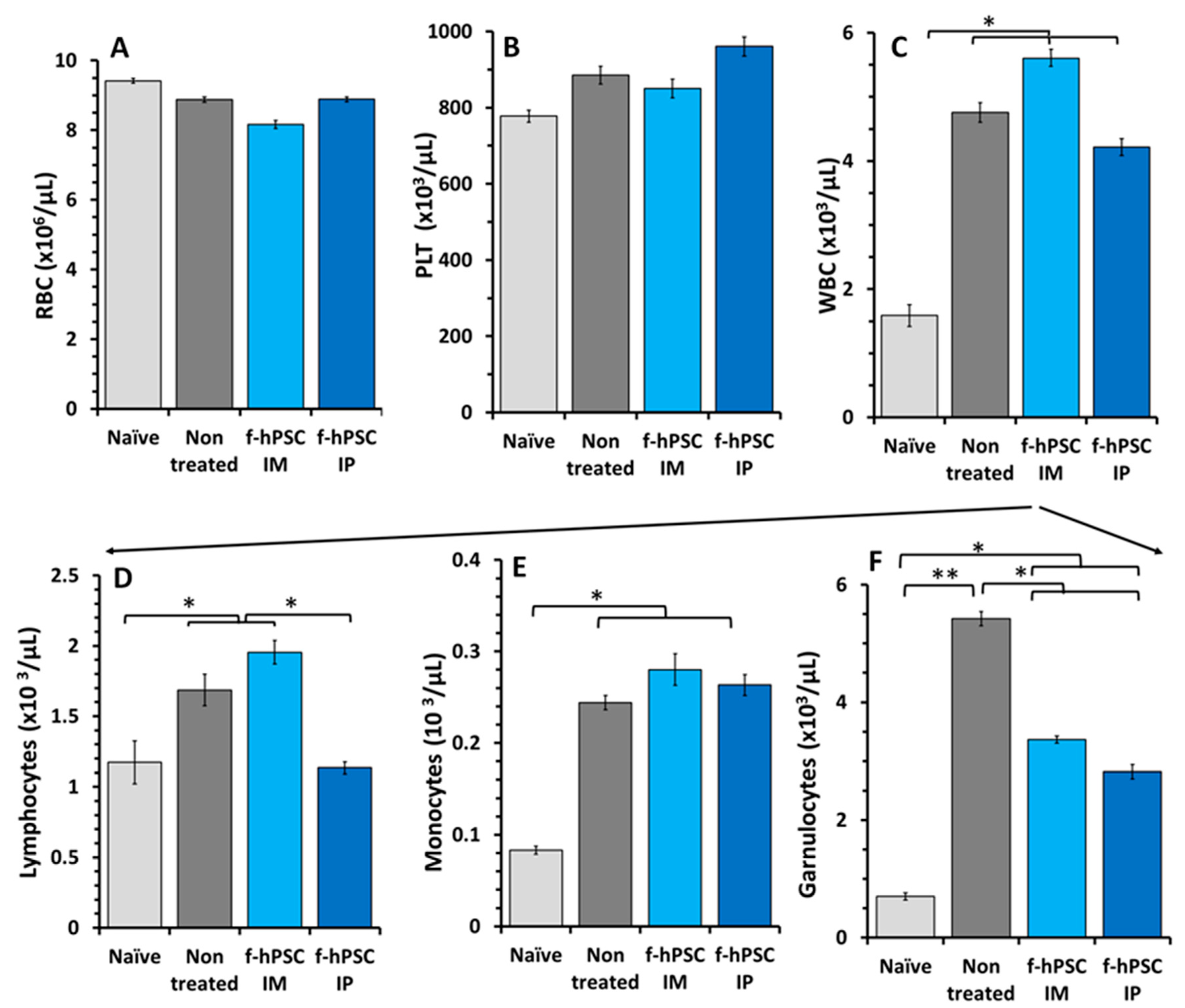
Figure 3.
H&E stained Histology of the colon. Small colon samples were collected from the sacrificed mice and were processed in paraffin for histology. Representative H&E stained slides were photographed for each group tested (A-D) with magnification of a selected area (A1, B1, C1, D1). The histological sections were used for scoring the severity of the inflammatory response (4A).
Figure 3.
H&E stained Histology of the colon. Small colon samples were collected from the sacrificed mice and were processed in paraffin for histology. Representative H&E stained slides were photographed for each group tested (A-D) with magnification of a selected area (A1, B1, C1, D1). The histological sections were used for scoring the severity of the inflammatory response (4A).
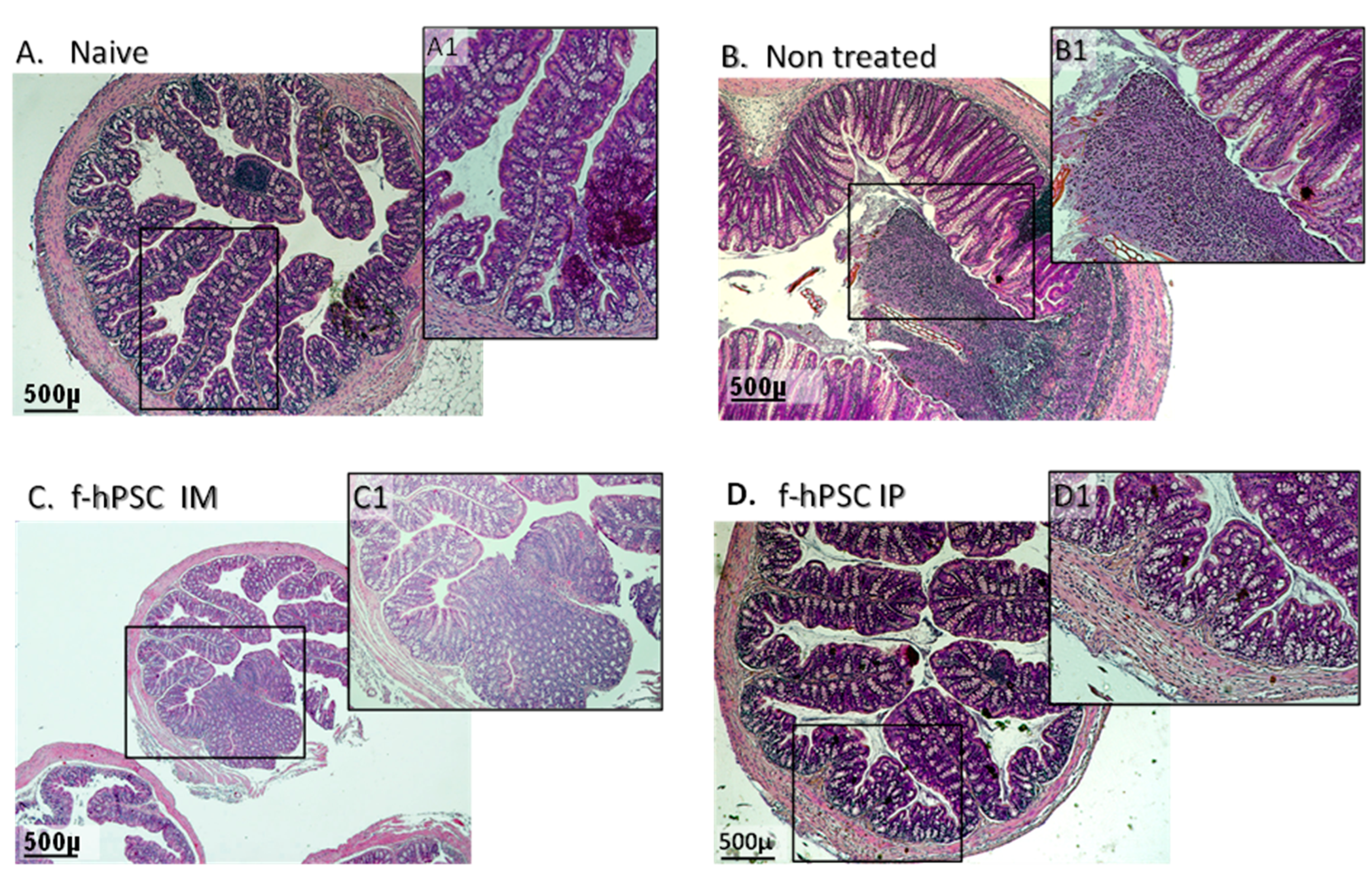
Figure 4.
Tissue inflammation and regeneration. The summary of the records of blind histological score for evaluation of tissue inflammation and regeneration are presented (A). processes by a pathological expert by parameters. The correlation between the severity of the inflammatory response (B), as evaluated by MPO assay for infiltrated neutrophils is presented. The bars above columns indicate significant different between groups where applicable (**=P<0.001).
Figure 4.
Tissue inflammation and regeneration. The summary of the records of blind histological score for evaluation of tissue inflammation and regeneration are presented (A). processes by a pathological expert by parameters. The correlation between the severity of the inflammatory response (B), as evaluated by MPO assay for infiltrated neutrophils is presented. The bars above columns indicate significant different between groups where applicable (**=P<0.001).
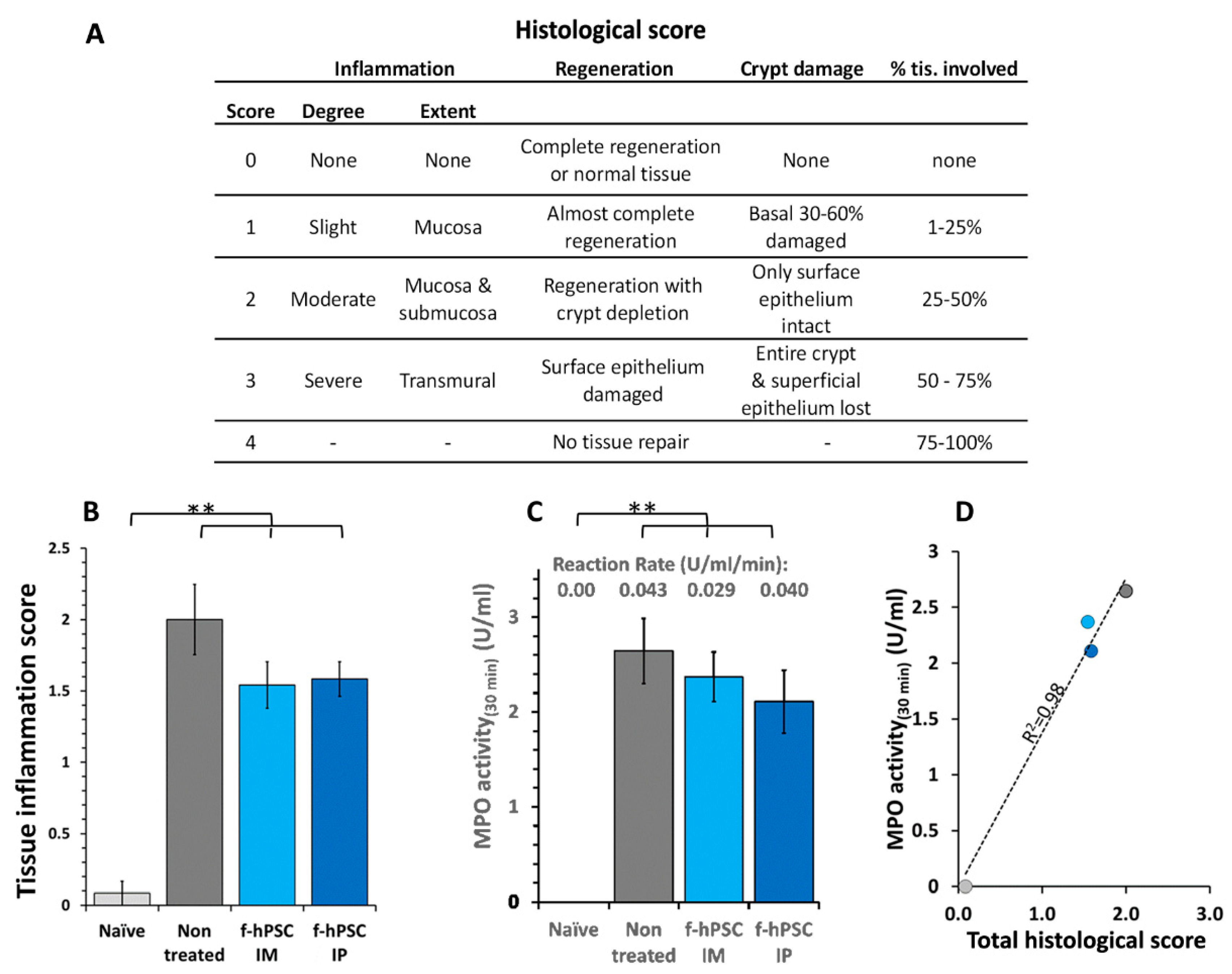
Figure 5.
Mɸ infiltration to the crypts of the colon. Immuno-stained histology of Mɸ infiltration in the colon histological sections of the different groups was tested. The sections of the colon of all mice in different arms were immune-stained with CD68 (with hematoxylin background). (A-C). Counts of Mɸ number per total crypts area in the immune-stained sections for naïve, non-treated and IP f-hPSCs groups are shown (D). The bars above the columns indicate the significant differences between the groups tested (*=P<0.05).
Figure 5.
Mɸ infiltration to the crypts of the colon. Immuno-stained histology of Mɸ infiltration in the colon histological sections of the different groups was tested. The sections of the colon of all mice in different arms were immune-stained with CD68 (with hematoxylin background). (A-C). Counts of Mɸ number per total crypts area in the immune-stained sections for naïve, non-treated and IP f-hPSCs groups are shown (D). The bars above the columns indicate the significant differences between the groups tested (*=P<0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



