
Submitted:
12 November 2024
Posted:
13 November 2024
You are already at the latest version
Abstract
In the Neotropical biogeographic region, there are about 290 known species and subspecies of fleas, which have as their main hosts in the Neotropics the subfamily Sigmodontinae. In Peru, more than 22 years have passed since the last list of known flea species was published. This study presents a review and update of the fleas recorded in Peru from 1908 to 2024. This list contains information on the flea species, their hosts, the localities in Peru where they were reported, the main references and some comments. In addition to the list, the specificity of flea-host interactions was assessed through the use of interaction networks. In the literature review, 86 flea species were found distributed among 9 families, 34 genera, and 19 subspecies. Regarding the hosts, these were made up of 72 species between mammals and birds (14 orders, 28 families, 54 genera) and seven undetermined ones. The flea-host interaction network exhibited a modular structure, indicating that these interactions tend to be specialized. A sigmodontine rodent (Akodon mollis) and a Rhopalopsyllidae flea (Polygenis litargus) were highly connected across modules and the overall network, thus were important to maintain the structure. This review contributes to the knowledge of fleas in Peru, synthesizing the scattered information and providing information on the interaction they have with their hosts.
Keywords:
Fleas
; Interaction networks
; Modularity
; Peru
; Siphonaptera
1. Introduction
The order Siphonaptera, which includes insects commonly known as fleas, represents a relatively small group of wingless, holometabolous, obligate hematophagous insects of higher vertebrates, especially mammals and birds, which they parasitize when the fleas reach the adult phase [1,2,3].
Fleas have a cosmopolitan distribution, including Antarctica, where Glaciopsyllus antarcticus Smit and Dunnet, 1962 has been reported parasitizing the silver petrel Fulmarus glacialoides (Smith, 1840)[4]. The geographical distribution of this group is mentioned as uneven since the greatest diversity is found in the Palearctic Region with 892 species, followed by Nearctic, Afrotropical and Neotropical respectively with 299, 275 and 289 species and finally the Eastern and Austral regions with 191 and 68 species, respectively [5,6,7].
Regarding the flea fauna of South America, it mainly comprises species in the families Malacopsyllidae and Rhopalopsyllidae and the subfamily Craneopsyllinae [7,8,9]. For the Neotropical biogeographic region, around 290 species are known [6] and 144 species for the Andean biogeographic region [10], with cricetid rodents of the subfamily Sigmodontinae as main hosts in the Neotropics [3].
The study of fleas in South America began in 1879 when Weyenbergh published the first descriptions of native species for this region. In Peru, one of the first efforts to quantify flea diversity was carried out by Macchiavello [11], who compiled the first list and zoogeographic distribution of all flea species recorded on the Southwestern Coast of America (Ecuador, Peru and Chile). In later years, the greatest contribution to the study of flea diversity at a taxonomic, systematic and zoogeographic level would be made by Johnson [12] in his monographs, where the South American flea fauna is described, in which 59 species are recognized for Peru. Later, the works of Hopkins and Rothschild [13], Smit [14,15,16,17], Schramm and Lewis [18, 19] and Hastriter et al. [20], would raise that figure to 77 species.
The association of fleas and their hosts represents an important ecological issue, since some flea species are strict parasites, while others show very low or no preference [21]. Marked host specificity is a trait seen in insect groups such as Phthiraptera that parasitize birds [22] and Nycteribiidae flies that parasitize bats [23]. This specificity can be counterproductive since it reduces the degree of their survival since it is dependent on the survival of a specific host species. In contrast, this characteristic of specificity is not prevalent in fleas. In this case, which presents a certain degree of specificity, it can be said that its inability to feed and reproduce on organisms other than its strict hosts can be interpreted as a negative factor in maintaining a high reproductive potential [21, 24, 25].
The specificity of parasitic interactions has been widely explored using species interaction network frameworks [26]. Through the structure of these networks, it is possible to examine community-level properties of ecological processes [27]. This approach also facilitates the identification of species' roles in maintaining the network, thus highlighting key species for the ecological processes at the community level [28]. In parasite ecology, interaction network methods have been applied to describe the temporal dynamics of parasitic associations [29], assess the effects of biological invasions [30], explore functional traits [31], and determine potential parasite transmission pathways [32], among other objectives (for more details, see [26, 33, 34]. Consequently, a network-based approach is often well-suited for describing these antagonistic interactions and can provide valuable insights to advance our understanding of parasite ecology.
Based on the above and due to the absence of updated compilations on the richness of fleas in Peru for more than 22 years and the dispersion of Peruvian scientific literature on this subject, it is important to consider that the synthesis of all this information, much of which is old and difficult to access, requires greater attention. Therefore, the objective of this review is to present an updated compilation of records of the flea species reported in Peru, their geographic distribution, their hosts, the degree of specificity and their interaction between species.
2. Materials and Methods
2.1. Parasite - Host Checklist
The list was compiled from an extensive bibliographic review in the main databases such as PubMed, Scopus, Science Direct, as well as physical and virtual documents from national and international libraries, collections and museums. Reports in all available languages published from 1908 to May 2024 were included. This list does not include records of abstracts from scientific meetings such as symposia, congresses, or undergraduate theses. The advanced search tools of Google Scholar, SciElo, PubMed, Scopus, Sciencedirect, Dialnet, JSTOR, Refseek were used, in which the terms and their combinations were used: “fleas”, “Siphonaptera”, “ectoparasites”, “Peru” and the names of the departments of the country. The list is presented in alphabetical order and is composed of two parts. The first is ordered according to the flea species, including their hosts, the localities where they were reported in Peru (including districts, provinces and/or departments), the type of specificity according to the number of hosts (including those recorded throughout America), the references and some comments if necessary (including the distribution in other countries, redescriptions, and taxonomic notes for diagnosis). The second part of the list is ordered according to the hosts and the fleas recorded on each of them and is presented as a supplementary table (Table S1).
The classification of fleas is based on Whiting et al., [35], and Hastriter and Bossard, [36]. The classification and validation of the mammalian host species follows that mentioned by Pacheco et al., [37]. The host specificity of the parasites was classified as: monoxenous when fleas were found on a single host species; oligoxenous when they were found on more than one host species, but of the same genus; pleioxenous found on hosts of the same family; polyxenous, found on hosts of different families but of the same order and euryxenous found on hosts of different orders [38, 39].
2.2. Flea-Host Interaction Network
A flea-host interaction network at the national scale was constructed using only records where species were identified to the species level, to minimize bias due to taxonomic uncertainty [40]. This network was binary, as the data sources were highly heterogeneous, making it a suitable representation of the fundamental niches of species at large scales [41]. The structure of the network was evaluated by assessing its modularity and nestedness, as well as the internal nestedness of modules. Modularity (Q) was determined using the DIRT-LPA algorithm [42], while nestedness was assessed with the NODF index. In line with Felix et al. [43], nestedness was calculated for the entire network (NODFtotal), as well as for pairs of species within the same module (NODFsm) and pairs from different modules (NODFdm). To determine the significance of modularity and total nestedness (NODFtotal), null models following Vázquez et al. [44] were employed. For NODFsm and NODFdm, however, the modular configuration needed to be considered, so the restricted null model of Felix et al. [45] was applied. The previously mentioned non-restricted null model [44] will be referred to as the "free" null model. For each test, 1,000 random matrices were generated using the respective null models, and Z-scores and p-values (derived from Monte Carlo procedures) were calculated.
Important species for entire network were identified using normalized degree centrality and betweenness centrality. The former measures the number of interactions a species has relative to the total number of possible interactions, while the latter reflects the frequency with which a species appears in the shortest paths between pairs of species in the network, indicating its role as a connector. Species that rank high in both centralities are well-connected both directly and indirectly within the network, thereby significantly contributing to its stability. Following an approach similar to Messeder et al. [46], the top 10 species in both metrics were considered crucial for maintaining network connectivity. Additionally, due to the network's modular structure (see results), species were further evaluated for among-module connectivity and within-module degree, as described by Guimerà and Amaral [47] and Olesen et al. [48]. The classification of species was based on thresholds established by Olesen et al. [48]: 0.62 for among-module connectivity and 2.5 for within-module degree. Species with high values in several of these metrics were identified as key to maintaining the network's structural integrity.
All analyses, including both network-level and species-level assessments, were conducted in R v.4.2.3 using the bipartite package [49].
3. Results
A total of 596 flea-host interactions were reported in the literature, of which 91 records did not contain information on the identity of the hosts. The interactions found were fleas belonging to 9 families, 34 genera, 86 species and 19 subspecies. Regarding the hosts, these were made up of 72 species among mammals and birds (14 orders, 28 families, 54 genera) and five undetermined ones.
Graph 1 shows the hierarchy of hosts among hosts recorded in Peru parasitizing some species of flea. It can be observed that the group of rodents were the hosts with the highest records of parasitization by fleas (63.9%) and within them, the Cricetidae family occupies 45.8%, followed by the Didelphimorphia group with 6%.

Graph 1. Hierarchy table of mammals and birds recorded as hosts cited in the flea literature in Peru.
The hosts with the highest number of reports of fleas associated with them were the species Akodon mollis Thomas, 1894 “Soft Grass Mouse” (n=17), Rattus rattus (Linnaeus, 1758) “House rat” (n=13), Rattus norvegicus (Berkenhout, 1769) “Brown Rat” (n=12), Phyllotis andium Thomas, 1912 “Andean Leaf-eared Mouse” (n=10), all belonging to the order Rodentia. The flea species with the highest host records in Peru were found to be Polygenis (Polygenis) litargus (Jordan & Rothschild. 1923) (n=23), Pulex irritans Linnaeus, 1758 (n=16), Ctenocephalides canis (Curtis, 1826) (n=12), Echidnophaga gallinacea (Westwood, 1875) (n=11). Regarding the geographic distribution of flea species in Peru, the four species with the greatest distribution were Ctenocephalides canis, Ctenocephalides felis, Tunga penetrans, Tiamastus cavicola, which were recorded in 19, 11, 10, and 9 departments, respectively.
Likewise, the departments that presented the highest flea richness were Piura (n=30), Ancash (n=26), Puno (n=25), La Libertad (n=17) and those that presented the highest diversity of flea-infested hosts were Piura (n=34), Puno (n=20), Lima (n=18), La Libertad (n=17). At least one flea-host association was reported in 22 departments except in Madre de Dios (Figure 1).
Fleas of the family Pulicidae were most widespread in across different families of hosts (15 families). Rhopalopsyllidae were the most prevalent fleas among mammal families (14 families). Only families Pulicidae and Tungidae were found in both birds and mammals. Some flea families were found only in rodent species (Hystrichopsyllidae, Ceratophyllidae, Pygiopsullidae and Leptosyllidae) or only bat species (Ischnopsyllidae) (Figure 2).
Regarding host specificity, 13 monoxenous species, 2 oligoxenous species, 13 pleioxenous species, 19 polyxenous species and 36 euryxenous species were found. The specificity of two species could not be determined because the hosts were not reported. Figure 3 and Figure 4 show some specimens of representative flea species of the 86 species reported in Peru deposited in the entomological collection of the Natural History Museum of the Faculty of Natural Sciences and Mathematics of the Federico Villarreal National University-MUFV.
3.1. Parasite - Host Checklist
Class Insecta
Order Siphonaptera
Family Ceratophyllidae
Subfamily Ceratophyllinae
1. Ceratophyllus titicacensis Smit, 1978
Host: Unknown.
Distribution in Peru: Capachica peninsula, Lago Titicaca, Puno.
Specificity: Unknown.
Reference: [16, 20]
This species was collected from an abandoned nest, possibly of the Big-billed Cormorant Phalacrocorax olivaceus (Humboldt). It is distributed in the Andean subregion, specifically in Puno, Peru. Its diagnosis is based on having a relatively longer pronotum and shorter pronotal spines, the male has a larger spiculose area, the female lacks a sinuosity on the posterior margin of sternum VII and a thicker spermathecal duct [16].
2. Dasypsyllus (Neornipsyllus) lewisi Hastriter & Schlatter, 2006
Host: Thomasomys sp.
Distribution in Peru: Huancabamba, Tambo, Piura.
Specificity: Monoxenous.
Reference: [50]
This species was described only from a male specimen collected in Piura, its distribution is restricted to this locality so far. It can be differentiated from the other male species of the subgenus by the presence of reduced and inconspicuous triangular lobes on the aedeagus, a wider ventral proctiger at the apex and a wide telomere with a thick and blunt spiniform on the dorsocaudal margin [50]. The female is unknown.
3. Dasypsyllus (Neornipsyllus) plumosissimus Smit, 1976
Host: Catamenia analis griseiventris (d'Orbigny & Lafresnaye, 1837).
Distribution in Peru: Cusco.
Specificity: Monoxenous.
Reference: [15, 50]
This species was only described from a male specimen collected in Cusco, its distribution is restricted to this locality so far. It can be distinguished from the other species of the subgenus by the extraordinary development of the intersegmental membrane between sternums VIII and IX [15]. The female is unknown.
4. Dasypsyllus (Neornipsyllus) stejnegeri (Jordan, 1937)
Host: Colaptes rupicola d'Orbigny, 1840.
Distribution in Peru: Recuay, Ancash.
Specificity: Euryxenous (Charadriiformes and Passeriformes).
Reference: [20, 50]
This species is distributed in the Canadian subregion, the Caribbean subregion, the Patagonian subregion and the Western American subregion. In South America it is reported in Argentina, Chile and central Peru. The male is distinguished from the rest of the species of the subgenus by having the telomere with a right angle on its ventrocaudal margin; narrow and tapered from the base to the apex with 2 setae on the ventral margin. The female has a pointed dorsal lobe of the sternum VII and a semiglobular spermatheca.
5. Nosopsyllus (Nosopsyllus) fasciatus (Bosc d’Antic, 1800)
Host: Unknown.
Distribution in Peru: Puno.
Specificity: Euryxenous (Rodentia and Didelphimorphia).
Reference: [20]
This species is native to the Palearctic region but has a cosmopolitan distribution. It has a predilection for synanthropic rodents and is a relevant species due to its association with disease transmission [51].
6. Nosopsyllus (Nosopsyllus) londiniensis londiniensis (Rothschild, 1903)
Host: Rattus rattus (Linnaeus, 1758), Rattus sp.
Distribution in Peru: Cajamarca.
Specificity: Polyxenous (Muridae and Cricetidae).
Reference: [11, 12, 20]
This species has been reported from the Eastern Mediterranean Province, the European Province, the Sahara-Arabian subregion and the Western Mediterranean Province. In America it has been reported from Argentina, Ecuador and Peru [9, 11].
7. Plusaetis dolens quitanus (Jordan, 1931)
Host: Akodon mollis Thomas, 1894; Ichthyomys stolzmanni Thomas, 1893; Mus musculus Linnaeus, 1758; Phyllotis amicus (Thomas, 1900); P. andium Thomas, 1912.
Distribution in Peru: Huaylas, Huayllan, Pomabamba, Pariacota, Huaraz, Parque Nacional del Huascarán, Huaylas, Recuay, Recuay, Ancash; Cicur, Huancabamba, Siclamache, Sondorillo, Piura.
Specificity: Polyxenous (Muridae and Cricetidae).
Reference: [12, 20, 52]
Reported as Pleochaetis dolens quitanus in Piura, Smit [53] later relocated this species to the genus Plusaetis Smit, 1983 [54]. Hastriter et al., [20], extends its geographic distribution further south, to the department of Ancash.
8. Plusaetis equatoris equatoris (Jordan, 1933)
Host: Akodon mollis; Cavia porcellus Linnaeus (1758); Oligoryzomys destructor (Tschudi, 1844); Sylvilagus sp.
Distribution in Peru: Huancabamba, Piura.
Specificity: Euryxenous (Rodentia and Lagomorpha).
Reference: [11, 12]
Reported as Pleochaetis equatoris equatoris by Macchiavello, [11]. This species would be relocated to the genus Plusaetis [54]. Hastriter et al., [20] consider that probably the specimens of P. equatoris equatoris recorded by Macchiavello are actually P. dolens quitanus. The host in Huancabamba was reported as Oligoryzomys longicaudatus (Bennett, 1832) but this species is not distributed in Peru [37].
Family Ischnopsyllidae
Subfamily Ischnopsyllinae
9. Hormopsylla egena Jordan, 1950
Host: Unknown.
Distribution in Peru: Trujillo, La Libertad.
Specificity: Unknown.
Reference: [12, 52]
This species was described from seven female specimens, collected from rat burrows in the roofs of houses inhabited by bats. The male is unknown. Its distribution is restricted to this locality [52].
10. Hormopsylla trux Jordan, 1950
Host: “Bat”
Distribution in Peru: Cementerio chino, Trujillo, La Libertad.
Specificity: Monoxenous.
Reference: [12, 52]
Species described from a male and female specimen. The host bat species have not been identified. Its distribution is currently restricted to this locality [52]. The species is similar to Hormopsylla fosteri Rothschild, 1903, but differs considerably in the sclerotization of certain parts of the skeleton and in the genitalia.
11. Myodopsylla setosa Johnson, 1956
Host: “Bat”
Distribution in Peru: Yucay, Cusco.
Specificity: Monoxenous.
Reference: [12, 55]
This species was first discovered and described in Peru by Johnson [55]. The specimens were collected from unidentified bats. It is distinguished from the rest of the species of the genus by the presence of a false comb on the third abdominal tergum and in the male it has a pair of caudal bristles longer than the width of the mobile finger and curved apically.
12. Myodopsylla wolffsohni wolffsohni (Rothschild, 1903)
Host: Myotis albescens (É. Geoffroy, 1806).
Distribution in Peru: Unknown
Specificity: Polyxenous (Chiroptera: Molossidae and Vespertilionidae).
Reference: [56]
In South America it has been reported in Argentina, Brazil, Chile, Paraguay, Peru and Uruguay. The type hosts are usually species of the genus Myotis. It is distinguished from the rest of the species of the genus by the presence of false combs only on tergum I - II and the presence of 5 - 6 setae at the apex of the eighth tergum in males.
13. Rothschildopsylla noctilionis (da Costa Lima, 1920)
Host: Eptesicus brasiliensis (Desmarest, 1819)
Distribution in Peru: Jenaro Herrera, Loreto.
Specificity: Polyxenous (Chiroptera: Noctilionidae and Vespertilionidae).
Reference: [57]
Remark: In South America it has been reported in Brazil and central eastern Peru. It is the only species of the genus, the diagnosis of the genus is the same for the species and is characterized by small thin bristles on front and combs present on abdominal terga I and II.
14. Sternopsylla distincta speciosa Johnson, 1957
Host: Tadarida brasiliensis (I. Geoffroy, 1824).
Distribution in Peru: Quince Mil, Cusco.
Specificity: Polyxenous (Chiroptera: Molossidae and Vespertilionidae).
Reference: [12]
In South America it is distributed in Argentina, Colombia, Paraguay, Peru, Panama and Venezuela. Its hosts are bats of the Vespertilionidae and Molossidae families.
Family Leptopsyllidae
Subfamily Leptopsyllinae
15. Leptopsylla segnis (Schonherr, 1811)
Host: Akodon sp; Cavia aperea Erxleben, 1777; Mus musculus; Rattus norvegicus (Berkenhout, 1769); Rattus rattus.
Distribution in Peru: Chimbote, Santa, Huayllan, Pomabamba, Valle del Río Santa, Choquechaca, Ancash; Chincha, Ica; Huacho, Lima; Moche, Otusco, Trujillo, La Libertad; Sapillica, Piura; Lambayeque; Tumbes.
Specificity: Euryxenous (Rodentia and Didelphimorphia).
Reference: [11, 12, 20, 52]
Cosmopolitan species of zoonotic importance as they transmit pathogens that cause bartonellosis, murine typhus (Rickettsia typhi), and flea-borne spotted fever (Rickettsia felis). Together with Xenopsylla cheopis, they are considered the largest flea species that effectively cause the transmission of murine typhus [58]. It is easily distinguishable from the rest of the species by the presence of a vertical genal comb with three to six spines and two or more short, thickened spiniform bristles on the anterodorsal angle of the head.
Family Ctenophthalmidae
Subfamily Ctenophthalminae
16. Agastopsylla hirsutior Traub, 1952
Host: Abrothrix jelskii (Thomas, 1894).
Distribution in Peru: Caccachara, Moquegua.
Specificity: Pleioxenous (Abrothrix and Akodon).
Reference: [12, 13, 59]
Distributed in Argentina and southern Peru. It was described only from a female holotype which was lost over time. The diagnosis of this species was modified by López-Berrizbeitia et al., [60] and the male of this species is described for the first time from specimens collected from Akodon spegazzinii Thomas 1897 from Tucumán, Argentina. It differs from the other species of the genus by having the lateral surface of the posterior tibia densely covered by 22-24 setae, the male telomere notably short; the lateral margins converge towards the apex.
17. Agastopsylla nylota nylota Traub, 1952
Host: Calomys Lepidus (Thomas, 1894).
Distribution in Peru: Carhuamayo, Junín.
Specificity: Pleioxenous (Cricetidae: Phyllotis and Calomys).
Reference: [12, 13, 59]
Distributed in Argentina and central Peru [60]. It was described from a single male specimen which is characterized from the other males of the genus by having a telomere with a strongly oblique apical margin. A subspecies A. nylota euneomys Lewis, 1984 was described from cricetids in Chile.
18. Agastopsylla pearsoni Traub, 1952
Host: Calomys lepidus; Chinchillula sahamae Thomas, 1898.
Distribution in Peru: Picotani, San Antonio de Putina, Puno.
Specificity: Pleioxenous (Cricetidae).
Reference: [13, 59]
In the Neotropics region distributed towards the south, including Argentina, Chile and southern Peru [9]. Described for the first time in Peru. Males are differentiated from the rest of the species of the genus by the absence of patches of bristles on tergum VII, the apical part of the truncated mobile finger forming two apical angles and sterno VIII with eleven setae. Females are differentiated by a ventral anal lobe with a defined submedial angle on the ventral margin.
19. Neotyphloceras chilensis Jordan, 1936
Host: Rattus norvegicus.
Distribution in Peru: San Damiano hacienda, Ancash.
Specificity: Euryxenous (Rodentia and Didelphimorphia).
Reference: [13]
It is distributed in Argentina, Chile and the central Andes in Peru. Initially it was listed as a subspecies of N. crassispina by Jordan, [61], later it was elevated to the status of species by Sanchez et al., [62] based on the shape of the apex of the fixed process of the clasper noticeably turned upwards and the chaetotaxy of the ventral margin of the distal arm of sternum IX with five curved spine-like bristles, the first one wider and strongly pigmented, located in the center above the others.
20. Neotyphloceras crassispina crassispina Rothschild, 1914
Host: Abrocoma cinerea Thomas, 1919; Abrothrix jelskii; Auliscomys boliviensis (Waterhouse, 1846); Calomys sorellus (Thomas, 1900); Chinchillula sahamae, Phyllotis amicus; P. osilae J.A. Allen, 1901; P. xanthopygus (Waterhouse, 1837), Rattus sp.
Distribution in Peru: Ancash; Hacienda Matara, Apurímac; Caylloma, Arequipa; San Ignacio, Cajamarca; Pachacayo, Junín; Matucana, Lima; Caccachara, Ilave, Huacullani, Pisacoma, Puno.
Specificity: Polyxenous (Cricetidae and Muridae).
Reference: [12, 13, 20, 52, 59, 63, 64]
The genus and species were described from murine mouse species captured in Peru. It is currently distributed in Argentina, Bolivia, Chile and Peru. Sanchez and Lareschi [65] developed the most current key for the taxonomic identification of the species and subspecies of the genus Neotyphloceras. It is distinguished from Neotyphloceras crassispina hemisus by the absence of dorsal spinelets on the abdominal tergites.
21. Neotyphloceras crassispina hemisus Jordan, 1936
Host: Aegialomys xanthaeolus (Thomas, 1894); Akodon mollis; Phyllotis andium; Rattus norvegicus; Thomasomys sp.
Distribution in Peru: Chiquian, Pariacota, Huaraz, Parque Nacional Huascarán, Huaylas, Recuay, Valle del Río Santa, Choquechaca, Hauzlas, Llampa, Caraz, Ancash.
Specificity: Polyxenous (Rodentia).
Reference: [13, 20]
It is distributed in Argentina, Bolivia, Chile and in Peru it has only been reported in the department of Ancash. It was redescribed by López-Berrizbeitia et al., [66] from male and female specimens collected in Catamarca, Argentina and they provide new diagnostic morphological characters for both sexes. It is distinguished from N. crassispina crassispina by the presence of dorsal spinelets in the abdominal tergites.
22. Neotyphloceras rosenbergi (Rothschild, 1914)
Host: Aegialomys xanthaeolus; Akodon mollis; Didelphis sp; Rhipidomys leucodactylus (Tschudi, 1845).
Distribution in Peru: Cascas, Cajamarca; Cascas, La Libertad; Huancabamba, La Laguna, Piura; Puno.
Specificity: Euryxenous (Rodentia and Didelphimorphia).
Reference: [12, 13, 20, 52, 61]
Geographic distribution in South America, including Bolivia, Colombia, Ecuador, Venezuela and Peru. Sanchez et al., [67] redescribes this species and extends its geographic range to Bolivia, previously it was only recorded as far north as Peru. It is easily distinguishable from the rest of the species of the genus by the first frontal row with eight or nine setae; chaetotaxy of the dorsal margin of the posterior tibia 2, 2, 2, 2, 2, 1, 2, 2; apical seta of the mobile process of the clasper slightly pigmented.
Subfamily Doratopsyllinae
23. Adoratopsylla (Tritopsylla) intermedia intermedia (Wagner, 1901)
Host: Didelphis marsupialis Linnaeus, 1758.
Distribution in Peru: Hacienda Limon, Balsas, Junín.
Specificity: Euryxenous (Rodentia and Didelphimorphia)
Reference: [13]
It is reported in didelphids and cricetid mice from Argentina, Brazil, Ecuador, Paraguay, Peru and Venezuela. In Peru it was reported in Didelphis (paraguayensis) andinus but this species is not found in Peru [37]. The males of this species are differentiated from the rest of the genus by having a posterodorsal lobe of the immobile process of the clasper that ends well below the subconical apical lobe, a sinus that is not as narrow.
24. Adoratopsylla (Tritopsylla) intermedia copha (Jordan, 1926)
Host: Didelphis marsupialis Lund, 1840.
Distribution in Peru: La Laguna, Piura.
Specificity: Euryxenous (Rodentia and Didelphimorphia).
Reference: [13]
Remark: It is reported in didelphids and cricetid mice from the northern part of South America, including Colombia, Costa Rica, Ecuador, French Guiana, Panama and northern Peru. In Peru, it was reported in Didelphis (paraguayensis) andinus, but this species is not found in Peru [37].
Family Stephanocircidae
Subfamily Craneopsyllinae
25. Cleopsylla townsendi Rothschild, 1914
Host: Abrothrix jelskii; Akodon albiventer Thomas, 1897; Akodon lutescens J.A.Allen, 1901; A. mollis; Auliscomys boliviensis; Auliscomys pictus (Thomas, 1884); Chinchillula sahamae; Oligoryzomys destructor (Tschudi, 1844); Phyllotis limatus Thomas, 1912; P. amicus; P. andium; Punomys lemminus Osgood, 1943.
Distribution in Peru: Chiquian, Pariacota, Huaraz, Huaylas, Parque Nacional Huascarán, Río Santos, Recuay, Collao, Pampa de Ancomarca, Pampa de Capazo, Ancash; Carhuamayo, Jauja, Pampa de Ancomarca, Junín; Caccachara, Mariscal Nieto, Moquegua; Pachacayo Pampa de Capazo Caccachara, Ilave, Limbani, Picotani, San Antonio de Putina, Puno; Tarata, Tacna.
Specificity: Polyxenous (Rodentia: Cricetidae and Octodontidae).
Reference: [12, 20, 63, 68]
The genus and species were described from specimens collected from mice in Pachacayo, Puno. It is currently distributed in Argentina, Chile, and south-central Peru [9, 68]. It is distinguished from other species of the genus by the absence of a false comb at the level of the fourth and fifth dorsal lateral notch on the metatibia and males by the length of the distal arm of the sterno IX, which is four times longer than its widest portion.
26. Craneopsylla minerva minerva (Rothschild, 1903)
Host: Akodon mollis; Akodon sp; Auliscomys sublimis (Thomas, 1900); Phyllotis amicus; Phyllotis andium; Phyllotis limatus; Rattus sp; Thomasomys sp.
Distribution in Peru: Valle del Rio Santa, Choquechaca, Hauzlas, Pariacota, Huaraz, Yungilla, Huari, Parque Nacional Huascarán, Huaylas, Recuay, Ancash; Arequipa; Sapillica, Ayabaca, Siclamanché, Piura; Limbani, Puno; Tarata, Tacna.
Specificity: Euryxenous (Rodentia and Didelphimorphia).
Reference: [11, 12, 20, 69]
This subspecies has a wide geographic distribution in South America, including Argentina, Brazil, Chile, Ecuador, Paraguay, Peru; Uruguay and Venezuela. In Peru, they have only been reported in cricetid and murid rodents; in Argentina and Paraguay, they are also reported in didelphids. It can be distinguished from the only other subspecies, Craneopsylla minerva wolffhuegeli (Rothschild, 1909), by the presence of five spines on each side of the genal comb.
27. Nonnapsylla rothschildi Wagner, 1938
Host: Galea musteloides Meyen, 1833
Distribution in Peru: Pisacoma, Pomata, Puno.
Specificity: Polyxenous (Rodentia).
Reference: [12]
The genus Nonnapsylla Wagner, 1938 includes only 1 species, Nonnapsylla rothschildi. Two subspecies were recognized by Johnson [12]. Moreover, Lareschi et al. [70] conclude that both subspecies must be considered synonymous. It is distributed in Argentina, Bolivia, Chile and southeastern Peru. The diagnosis of the genus is the same for the species, based on the angled anterior margin of the head, the helmet completely separated from the rest of the head, with striations, and the presence of an area of marginal and submarginal mesal setae on the procoxa.
28. Plocopsylla achilles (Rothschild, 1911)
Host: Akodon mollis; Nephelomys albigularis (Tomes, 1860); Phyllotis andium.
Distribution in Peru: Chiquian, Parque Nacional Huascarán, Huaylas, Recuay, Ancash; Chota, Cajamarca.
Specificity: Polyxenous (Cricetidae and Muridae)
Reference: [12, 20]
It is distributed in Chile, Ecuador and in central eastern and northeastern Peru. Schramm and Lewis [19], revised the genus Plocopsylla Jordan (1931), restructured it into two groups and six subgroups based on the mobile process, sterno IX and the clasper with the mesal process. The diagnosis of the species is based on the presence of five spines on the genal comb and a genal spine that is at least half as long as the longest adjacent one on the genal comb.
29. Plocopsylla enderleini Wagner, 1933
Host: Chinchillula sahamae.
Distribution in Peru: Cailloma, Arequipa; Limbani, Picotani, Puno; Tarata, Tacna.
Specificity: Polyxenous (Chinchillidae, Muridae and Cricetidae)
Reference: [12, 19]
It is geographically distributed in Bolivia, Chile, and southern Peru. The type host is “rat” from La Paz, Bolivia, but C. sahamae is mentioned in other localities and in undetermined cricetids from Puno [19]. The diagnosis of the species is based on the presence of six spines on the genal comb without basal notches, the prectenidial area of the helmet with a width 1.5 times the length of the longest spine of the adjacent comb.
30. Plocopsylla hector Jordan, 1931
Host: Akodon mollis; Oligoryzomys destructor; Rattus rattus; Thomasomys sp.
Distribution in Peru: Tambo, La Libertad; Huancabamba, Piura; Sauce, San Martín.
Specificity: Polyxenous (Cricetidae and Muridae).
Reference: [19]
It is distributed in Ecuador and northern Peru. Schramm and Lewis, [19], mention the locality of Tambo in Piura, but according to the coordinates (7.35S, 78.42W) it belongs to the department of La Libertad. The diagnosis is based on the setae on the dorsal margin of the metatibia forming a false comb and the width of the prectenidial hoof approximately equal to or greater than the length of the longest spine of the hoof.
31. Plocopsylla inti Johnson, 1957
Host: Phyllotis magister Thomas, 1912; P. limatus.
Distribution in Peru: Tarata, Tacna.
Specificity: Oligoxenous (Phyllotis)
Reference: [12]
Distributed only in southern Peru, it is easily distinguishable from the rest of the species of the genus by the presence of seven spines on the genal comb.
32. Plocopsylla kilya Schramm et Lewis, 1987
Host: Aegialomys xanthaeolus; Thomasomys sp; Thomasomys cinereus (Thomas, 1882).
Distribution in Peru: Tambo, La Libertad; Huancabamba, Piura.
Specificity: Pleioxenous (Cricetidae).
Reference: [19]
Distributed only in northern Peru. Schramm and Lewis [19] mention the locality of Tambo in Piura, but according to the coordinates (7.35S, 78.42W) it belongs to the department of La Libertad. This species has the same diagnosis as P. hector to separate it from the rest of the species of the genus. It differs from the latter in having the width of the prectenidial helmet much smaller than the length of the longest spine of the helmet.
33. Plocopsylla pallas (Rothschild, 1914)
Host: Akodon mollis; Calomys lepidus; Phyllotis amicus; Phyllotis andium.
Distribution in Peru: Río Santos, Recuay, Pariacota, Huaraz, Ancash; Carhuamayo, Pachacayo, Junín.
Specificity: Pleioxenous (Cricetidae).
Reference: [19, 20, 63]
Distributed only in the central Andes of Peru. Described from female specimens, later Hastriter et al., [20] described the male. The diagnosis is based on the presence of six spines on the genal comb without a basal notch and the width of the prectenidial helmet is twice the length of the longest spine on the helmet.
34. Plocopsylla viracocha Schramm et Lewis, 1987
Host: Phyllotis limatus; Phyllotis osilae; Phyllotis sp; Punomys lemminus.
Distribution in Peru: Heda, Arequipa; Huancavelica, Huancavelica; Limbani, Puno; Caccachara, Pampa de Ancomarca, Tarata, Tacna.
Specificity: Pleioxenous (Cricetidae).
Reference: [19]
Distributed in Chile and southern Peru. Schramm and Lewis [19] mention the coordinates 17.20S, 70.00W; 17.10S, 70.15W; 17.02S, 70.20W as belonging to the department of Puno, but in reality they belong to Tacna. Also for 16.10S, 71.00W it was wrongly attributed to Puno when in reality it belongs to Arequipa. The diagnosis is similar to P. pallas and P. enderleni, it is differentiated from both species by having the lowest of the 3 bristles on the fourth to sixth dorsal notch from the proximal end of the hind tibia, separated from the other 2 bristles by a space equal to or greater than the basal width of the lowest bristle.
35. Sphinctopsylla inca (Rothschild, 1914)
Host: Akodon mollis; Auliscomys pictus; Phyllotis andium; P. osilae.
Distribution in Peru: Chiquian, Parque Nacional Huascarán, Huaylas, Ancash; Carhuamayo, Pachacayo, Junín; Pomata, Puno.
Specificity: Pleioxenous (Cricetidae).
Reference: [12, 20, 63]
It is distributed in Bolivia, Ecuador, the central Andes and southern Peru [71]. Initially described as Craneopsylla inca and later changed to the genus Sphinctopsylla by Jordan [72]. Currently there are six species in this genus and S. inca is the only one recorded in Peru. It can be differentiated from the rest of the species of the genus by presenting only two rows of setae on the pronotum, the pronotal comb with the last dorsal spines with fine tips and a genal comb with five spines.
36. Tiarapsylla bella Johnson, 1956
Host: Leopardus garleppi (Matschie, 1912)
Distribution in Peru: Picotani, Puno.
Specificity: Monoxenous
Reference: [12, 55]
Distributed only in southeastern Peru. It was described from female specimens. It was discovered from wild cats in Peru. Johnson [55] suggests that this host is undoubtedly abnormal or accidental, the true host probably being vizcachas (as in the case of T. titschacki Wagner 1937) since the cat from which the specimens of T. bella were taken was observed hunting vizcachas. The genus currently has three species from which T. bella is distinguished by the lack of a membranous flap covering the second antennal segment, the base of the sinuous helmet comb and small bristles present posteriorly on the third or fourth dorsal helmet spine.
37. Tiarapsylla titschacki Wagner, 1937
Host: Lagidium viscacia (G. I. Molina, 1782); “felino”.
Distribution in Peru: Recuay, Ancash; Capillsa, Junín; Tayapampa, La Libertad; Picotani, Puno.
Specificity: Euryxenous (Rodentia and Carnivora).
Reference: [12, 20, 55]
Distributed in the Andean region of Peru. Hastriter et al., [20] mention that this species of flea has as host preference L. viscacia and that the reported feline is abnormal or accidental due to its association as a predator of the vizcacha. It differs from the other species of the genus by lacking a membranous flap covering the second antennal segment, convex base of the helmet comb and small bristles present behind the second dorsal spine of the helmet.
Family Pulicidae
38. Cediopsylla spillmanni Jordan, 1930
Host: Cavia porcellus; Sylvilagus brasiliensis (Linnaeus, 1758).
Distribution in Peru: Ayabaca, Huancabamba, Soccha, Siclamache, Piura.
Specificity: Euryxenous (Lagomorpha and Rodentia).
Reference: [11, 69]
Distributed in Ecuador and northwestern Peru. Diagnosis is based on the presence of a short vertical or subvertical genal comb with rounded spines.
39. Ctenocephalides canis (Curtis, 1826)
Host: Canis familiaris Linnaeus, 1758; Cavia sp; Didelphis paraguayensis andina; Didelphis sp; Felis catus Linnaeus, 1758; Gallus gallus (Linnaeus, 1758); Homo sapiens Linnaeus, 1758; Mus musculus; Neogale sp; Rattus norvegicus; Rattus rattus; Sus scrofa Linnaeus, 1758.
Distribution in Peru: Ancash; Cajamarca; Chincha, Ica; Junín; Moche, Trujillo, La Libertad; Ucupe, Lambayeque; Barranca, Comas, Independencia, San Juan de Lurigancho, San Martín de Porras, Lima; Piura; Tumbes.
Specificity: Euryxenous.
Reference: [11, 12, 20, 73, 74]
Cosmopolitan species, except at the poles. Macchiavello [11] mentions this species in “Canis” in all the departments that make up the Coast and Sierra of Peru.
40. Ctenocephalides felis felis (Bouche, 1835)
Host: Canis familiaris; Cavia porcellus; Didelphis sp; Felis catus; Gallus gallus; Homo sapiens; Rattus norvegicus; Rattus sp; Rattus rattus.
Distribution in Peru: Utcubamba, Amazonas; Chimbote, Valle del Río Santa, Choquechaca, Hauzlas, Ancash; Cajamarca; Ica; Trujillo, La Libertad; Lambayeque; Barranca, Comas, Huacho, Huaral, Independencia, San Juan de Lurigancho, San Martín de Porras, Lima; Piura; Tumbes.
Specificity: Euryxenous.
Reference: [11, 12, 20, 52, 74–77]
Cosmopolitan species, except at the poles. Macchiavello [11], mentions this species in all the departments that make up the Coast in Felis catus. It is probably found in all the departments of Peru in domestic animals.
41. Echidnophaga gallinacea (Westwood, 1875)
Host: Canis familiaris; Cavia aperea; Cavia porcellus; Columba livia J.F.Gmelin, 1789; Didelphis paraguayensis andina; Felis catus; Gallus gallus; Oryctolagus cuniculus (Linnaeus, 1758); Rattus norvegicus; Rattus rattus; Sigmodon peruanus J. A. Allen, 1897; Sus scrofa.
Distribution in Peru: Vinzos, Santa, Ancash; Cajamarca; Trujillo, La Libertad; Barranca, Carmen de la Legua, Comas, Huacho, Independencia, San Juan de Lurigancho, San Martín de Porras, Lima; Moquegua; Tumbes.
Specificity: Euryxenous.
Reference: [11, 12, 20, 52, 73, 74, 76, 78–80]
Cosmopolitan. This species is a major pest of poultry that can cause severe pathology or death if left untreated, and is rarely recorded in free-living bird species. Macchiavelli [11] reports it in multiple mammalian hosts although this species is more closely related to poultry, urban and, infrequently, wild birds.
42. Euhoplopsyllus andensis (Jordan, 1933)
Host: Sylvilagus sp.
Distribution in Peru: Soccha, Huancabamba; Piura.
Specificity: Euryxenous (Rodentia and Lagomorpha).
Reference: [11, 12]
It is distributed in Ecuador and northwestern Peru. This genus was elevated from its status as a subgenus of Hoplopsyllus Baker, 1905 to a genus by Ewing [81].
43. Euhoplopsyllus manconis (Jordan, 1950)
Host: Sylvilagus sp.
Distribution in Peru: Huancabamba, Siclamanche, Piura.
Specificity: Monoxenous.
Reference: [12, 20, 52, 82]
It is distributed in northwestern Peru. This species is closely related to E. andensis and is distinguished from it by having the anal stylet more than twice as long as it is wide.
44. Pulex irritans Linnaeus, 1758
Host: Caenolestes caniventer Anthony, 1921; Canis familiaris; Cavia porcellus; Conepatus sp; Felis catus; Gallus gallus; Homo sapiens; Lagidium viscacia; Lycalopex sp; Mus musculus; Oryctolagus cuniculus; Rattus norvegicus; R. rattus; Sigmodon peruanus; Sus scrofa; Sylvilagus sp.
Distribution in Peru: Valle del Río Santa, Choquechaca, Hauzlas, Ancash; Cajamarca; Ica; La Libertad; Lambayeque; Comas, Independencia, San Juan de Lurigancho, San Martín de Porras, Lima; Sapillica, Ayabaca, Montero, Suyo, Paimas, Lagunas, Piura; Tumbes.
Specificity: Euryxenous.
Reference: [11, 12, 20, 52, 69, 73]
Cosmopolitan species, of great relevance and importance in public health because it is one of the flea species most associated with humans, causing zoonoses by bacterial pathogens. Lareschi et al. [83] mention that this species was confused for many years with the closely related species Pulex simulans Baker, 1895 and discuss the diagnosis of both species in which they establish the differences: number of setae on each side of the sinus of the sternites VII in females (4-6 in P. irritans vs. 7-9 in P. simulans); shape of the aedegal sclerite (das) (thin in P. irritans vs. strong in P. simulans), length of clasper process 1 (P1) (long and completely covering processes 2 and 3 in P. irritans vs. short and not covering the other processes in P. simulans), and the presence (in P. irritans) or absence (in P. simulans) of an expansion at the clasper.
45. Pulex (Pulex) simulans Baker, 1895
Host: Cavia aperea; Canis familiaris.
Distribution in Peru: Chiribaya Baja, Valle de Moquegua, Moquegua.
Specificity: Euryxenous (Rodentia and Carnivora).
Reference: [78, 84, 85]
During a paleoparasitological study of several animal mummies (guinea pigs and dogs) from Chiribaya Baja, this species was found, which was recovered in good condition and DNA material was recovered [85].
46. Xenopsylla cheopis (Rothschild, 1903)
Host: Cavia porcellus; Didelphis sp; Gallus gallus; Homo sapiens; Mus musculus; Rattus norvegicus; R. rattus.
Distribution in Peru: Ancash; Cajamarca; Ica; La libertad; El Sauce, Barco, Algarrobo, Kerguer y La Ramada, Salas, Ucupe, Lambayeque; Huacho, Lima; Sapillica, Ayabaca, Suyo, Paimas, Lagunas, Piura; Tumbes.
Specificity: Euryxenous (Rodentia and Didelphimorphia).
Reference: [11, 12, 20, 52, 69, 86]
Considered a cosmopolitan species, it is the main vector of the bubonic plague bacterium, Yersinia pestis, and also of bacteria belonging to the genera Rickettsia and Bartonella [87]. It is also mentioned as the vector of protozoa such as Trypanosoma sp. and of cestodes such as Hymenolepis diminuta [88]. It differs from the other two species of the genus Xenopsylla Glinkiewicz, 1907 recorded in South America by having the distal arm of the ninth sternum in the male equally sclerotized throughout its length, and the female with the base of the tail of the spermatheca not strongly ventricose.
Family Rhopalopsyllidae
Subfamily Parapsyllinae
47. Delostichus phyllotis Johnson, 1957
Host: Cavia sp; Phyllotis xanthopygus.
Distribution in Peru: Yura, Arequipa.
Specificity: Euryxenous (Rodentia and Didelphimorphia).
Reference: [12, 17, 89]
It is distributed in southwestern South America, including Chile and southern Peru. The type host was mentioned as Phyllotis arenarius, which was later synonymized with P. xanthopygus. It differs from the rest of the species of the genus by having the procoxa without particularly large bristles than the others, the fifth protarsal segment longer and the row formed by its plantar bristles not oblique.
48. Delostichus xenurus (Rothschild, 1914)
Host: Lagidium viscacia.
Distribution in Peru: Llampa, Ancash.
Specificity: Monoxenous.
Reference: [17, 63]
Distributed in the central Andes of Peru at 4000 m above sea level. Initially described as Parapsyllus xenurus Rothschild, 1914; later modified by Jordan, [90]. The diagnosis to separate it from the other species of the genus is based on the presence of short bristles at the base of the first antennal segment, not reaching the base of the club; posterior margin of the genal process convex.
49. Ectinorus (Ectinorus) alejoi Hastriter, 2002
Host: Lagidium viscacia.
Distribution in Peru: Recuay, Ancash.
Specificity: Monoxenous.
Reference: [20, 91]
Distributed only in central western Peru. Hastriter and Sage [91] mention this species as being distributed in southern Peru, but do not specify the localities, districts or departments. The type host was Lagidium peruanum Meyen,1833, later synonymised with L. viscacia. It can be distinguished from the rest of the species in the genus by the absence of spurs on the first abdominal tergum, it is close to E. viscachae (Wagner, 1937) and it can be differentiated from it by the details of the clasper, St. VIII and st. IX in the male; and in the female by the modified abdominal segments and the shape of the spermatheca.
50. Ectinorus (Ectinorus) claviger (Rothschild, 1914)
Host: Calomys lepidus; Lagidium viscacia.
Distribution in Peru: Recuay, Ancash; Querobamba, Ayacucho; Carhuamayo, La Oroya, Junín; Querobamba, Puno.
Specificity: Euryxenous (Rodentia and Didelphimorphia).
Reference: [12, 17, 20, 63, 92]
It is distributed throughout the Peruvian Andes. Johnson, [12], mentions this species in two possible hosts, Akodon Chraeomys jelskii or Phyllotis P. limatus, but it is not defined to which it belongs. Hastriter et al., [20] complements the description of the females of this species. The diagnosis in both sexes is an antennal pedicel without a row of small lateral setae; tergum I without marginal spurs; labial palpus with 5 segments. Male with posterior margin of telomere not convex; process of basimeris ventralis not digitiform, not shorter than the telomere, with a rounded apex and covered with spiniform setae. Female with Sensilium with less than 20 sensitive pits on each side; protuberance of spermatheca not subspherical and perula of copulatory bursa spherical.
51. Ectinorus (Ectinorus) disjugis Jordan, 1942
Host: Auliscomys pictus.
Distribution in Peru: Cailloma, Arequipa.
Specificity: Polyxenous (Ctenomyidae and Cricetidae).
Reference: [17]
Distributed in Argentina and southern Peru. Initially described as female specimens, the male was first described by López-Berrizbeitia, et al. [93]. Both sexes are characterized by presenting a six-segmented labial palp; seven dorsal notches on the posterior tibia; middle abdominal terga with a seta of the main row below the level of the spiracle.
52. Ectinorus (Ectinorus) hecate Johnson, 1957
Host: Leopardus garleppi.
Distribution in Peru: Huaylarco, Arequipa.
Specificity: Monoxenous.
Reference: [12, 17]
Distributed only in southern Peru. Described only from female specimens, the male is unknown. The type host, L. garleppi, is considered an accidental host [12, 17]. The female is distinguished from the other species of the subgenus by presenting an antennal pedicel without a row of small lateral setae; tergum I with marginal spurs; labial palpus with 5 segments; pronotum with two rows of setae; caudal margin of S-VII strongly straight.
53. Ectinorus (Ectinorus) hertigi (Johnson, 1957)
Host: Ctenomys peruanus Sanborn & Pearson, 1947.
Distribution in Peru: Pampa Queullicota, Ilave, Pichupichuni, Puno.
Specificity: Oligoxenous (Ctenomys fulvus from Chile).
Reference: [12, 17]
Distributed in Chile and south-eastern Peru. First described in Peru as Dysmicus hertigi by Johnson [12], later modified by Smit [17]. Diagnosis similar to E. (Ectinorus) claviger, males differ in having smooth posterior margin of pronotum; metepimere with two rows of setae; arm of dorsoapical angle of S-IX more angular; straight ventral margin; manubrium with rounded apex; apex of basimere not bilobed. Females have sensilium with less than 20 sensory pits on each side; cribriform area at the level of the surface of the bulge; metepimere with two rows of setae.
54. Ectinorus (Ectinorus) hirsutus Hastriter, 2009
Host: Neotomys ebriosus.
Distribution in Peru: Yauli, Junín.
Specificity: Monoxenous.
Reference: [94]
Distributed only in Central Peru. The holotype host is Neotomys sp. captured in Junín, currently the only species in Peru of the genus Neotomys is N. ebriosus [37]. Males are distinguished from members of the subgenus Ectinorus by a combination of the following characters: apical spurs present on T-I; the process basimeris ventralis is apical and not attached ventrally to the basimere margin; the crochet is spoon-shaped and S-VIII has an accessory lobe with setae extending beyond the ventral apex of the caudal margin of S-VIII [94]
55. Ectinorus (Ectinorus) ineptus Johnson, 1957
Host: Auliscomys pictus.
Distribution in Peru: Picotani, Puno.
Specificity: Pleioxenous (Cricetidae).
Reference: [12, 17]
Distribution in Chile and south-eastern Peru. Described from male specimens by Johnson [12], the female was later described by Beaucournu and Gallardo [95] and redescribed by Beaucournu et al., [96].
56. Ectinorus (Ectinorus) pearsoni (Johnson, 1957)
Host: Phyllotis sp.
Distribution in Peru: Pomata, Puno.
Specificity: Pleioxenous (Akodon and Phyllotis).
Reference: [12, 17]
Distributed in Bolivia and south-eastern Peru. The female is unknown. The male is distinguished from the rest of the subgenus by having an antennal peduncle without a row of small lateral setae; tergum I without marginal spurs, basimere with a process basimeris ventralis, labial palpus with 5 segments, which does not extend beyond the middle of the anterior trochanter, posterior margin of the pronotum smooth, metepimer with a row of setae, manubrium basally quite narrow.
57. Ectinorus (Ectinorus) sentus (Rothschild, 1914)
Host: Lagidium viscacia.
Distribution in Peru: La Oroya, Junín.
Specificity: Monoxenous.
Reference: [12, 17, 20, 63]
Distributed only in central Peru. The female is unknown. The male is similar to E. (Ectinorus) claviger, but differs from the latter by having a ventral basimere process shorter than the telomere (not shorter than the telomere in E. (Ectinorus) claviger).
58. Ectinorus (Ectinorus) simonsi (Rothschild, 1904)
Host: Galea musteloides.
Distribution in Peru: Yura, Arequipa.
Specificity: Polyxenous (Octodontidae, Chinchillidae and Caviidae).
Reference: [17]
Distributed in Bolivia, Chile and south-western Peru. Species described as Pulex simonsi, later synonymised with Dysmicus acheronis (Johnson, 1957) and redescribed by Smit, [17]. Diagnosis similar to E. (Ectinorus) claviger and E. (Ectinorus) sentus, males are differentiated from other similar species by having a thin telomere, approximately five times longer than its maximum width; females are similar to Ectinorus (Ectinorus) hertigi and are differentiated only by the presence of a single row of setae on the metepimeron; posterior margin of sternum VI with a shallow sinus; non-quadrangular spermatheca.
59. Ectinorus (Ectinorus) viscachae (Wagner, 1937)
Host: Galea musteloides; Lagidium viscacia.
Distribution in Peru: Yura, Arequipa; Querobamba, Tayapampa, Ayacucho; La Oroya, Junín; Caccachara, Ilave, Santa Rosa, Pisacoma, Puno.
Specificity: Euryxenous (Rodentia and Lagomorpha).
Reference: [12, 17, 59]
Distributed in central and southern Peru. This species was reported with its synonyms Dysmicus viscachae Wagner, 1937; Dysmicus acheronis (Johnson, 1957) in Puno and Arequipa respectively. Males are differentiated from the others of the subgenus by having a basimere without a basimeris ventralis process, a club-shaped distal arm of sterno IX, and a wide telomere towards the apex. Females are differentiated from the others of the subgenus by having a labial palp with 5 segments, a sensilium with more than 20 pits on each side, an almost spherical spermatheca, and sterno II with a lateral patch with more than 6 setae.
60. Parapsyllus cedei Smit 1970
Host: Pelecanoides garnotii (Lesson, 1828).
Distribution in Peru: Islas de las viejas, Ica.
Specificity: Polyxenous (Procellariiformes: Procellariidae and Pelecanoides).
Reference: [17]
Distributed on Genovesa, Santa Cruz, Plaza Islands in the Galapagos and in Peru, only on the Viejas Islands. The originally described species was collected from nests of shearwater and petrels [17]. This species belongs to the longicornis-gwuip subgroup and is distinguished from all other members by the possession of 5 notches on the posterior margin of the meso- and metatibiae and 3 pairs of lateral plantar setae.
61. Tetrapsyllus (Phylliver) bleptus (Jordan et Rothschild, 1923)
Host: Abrocoma cinerea; Abrothrix jelskii; Akodon sp; Calomys lepidus; Ctenomys opimus Wagner, 1848; Chinchillula sahamae; Phyllotis andium; Phyllotis sp; P. xanthopygus; Punomys lemminus.
Distribution in Peru: Pariacota, Huaraz, Ancash; Carhuamayo, Junín; Caccachara, Puno.
Specificity: Pleioxenous (Cricetidae).
Reference: [12, 17, 20, 59]
This species has a distribution in the Andean and Patagonian subregions. In Argentina, it has been reported on Akodon simulator, A. spegazzinii and Phyllotis osilae [97]; also on Reithrodon caurinus [98]. Beaucournu and Gallardo [99] mention that T. (P.) bleptus is a recognized species in several points of western Argentina and in southern Peru; it was also reported in Chile on Phyllotis darwini. The male is distinguished from the rest of the subgenus Phylliver by having a telomere that extends beyond the apex of the basimere and 20 or fewer setae on the lateral surface of the hind tibia. The female has a non-angulated hilla, a sinus presents on the posterior margin of S-VII and the same characteristic of the tibia as the male [97].
62. Tetrapsyllus (Phylliver) elutus Johnson, 1957
Host: Akodon albiventer.
Distribution in Peru: Pampa de Capazo, Ilave, Puno.
Specificity: Pleioxenous (Cricetidae).
Reference: [12, 17]
It is distributed in the Andean subregion of Chile and southeastern Peru. This species is very similar to T. (P.) bleptus, the difference being only in the male specimens, in which in T. (P.) elutus the apex of the telomere does not extend beyond the basimere. The females of both species are indistinguishable.
63. Tetrapsyllus (Phylliver) tristis Johnson, 1957
Host: Abrothrix jelskii; Ctenomys peruanus; C. opimus.
Distribution in Peru: Lago Suches, Moquegua; Pampa de Ancomarca; Pampa Queullecota, Puno.
Specificity: Polyxenous (Cricetidae and Ctenomyidae).
Reference: [12, 17]
Distribution only in southern Peru. The morphological characterization of T. (P.) tristis stands out for the observation of more than 30 setae on the lateral surface of the metatibia, unlike the other species that consist of at least 20 setae in the same position.
64. Tetrapsyllus (Tetrapsyllus) comis Jordan, 1931
Host: Akodon mollis; Thomasomys cinereus.
Distribution in Peru: El Tambo. Huancabamba, Piura.
Specificity: Pleioxenous (Cricetiade).
Reference: [17]
Its distribution covers the Andean subregion. Berrizbeitia et al., [97] mentions a certain morphological characterization for this species through the study of its genitalia, the males of T. (T.) comis have a narrow telomere with few and tiny bristles on the mesal surface; as for the female, she has the spermatheca with a spherical protuberance.
Subfamily Rhopalopsyllinae
65. Polygenis (Ayeshaepsylla) thurmani Johnson, 1957
Host: Abrothrix jelskii; Oligoryzomys destructor; Phyllotis osilae.
Distribution in Peru: Limbani, Puno.
Specificity: Pleioxenous.
Reference: [12, 17]
This species is found in the Andean subregion. In male individuals of P. thurmani, the sclerotized internal tube of the aedeagus is reflected dorsally.
66. Polygenis (Gephyropsylla) klagesi klagesi (Rothschild, 1904)
Host: Akodon mollis; Neacomys sp; Neacomys spinosus (Thomas, 1882); Proechimys brevicauda (Gunther, 1877); Proechimys decumanus (Thomas, 1899); Proechimys simonsi Thomas, 1900; Proechimys sp.
Distribution in Peru: Jenaro Herrera, Loreto; Aguas Verdes, La Palma, Papayal, Tumbes.
Specificity: Polyxenous.
Reference: [17, 57]
It has a distribution in the Amazon, Brazilian and Caribbean subregions. Méndez [24] reports the presence of this species in Ecuador, Colombia, Venezuela, Trinidad and Costa Rica. P. klagesi is distinguished from the rest of the species of the genus by having a group of two setae on the subapical dorsal notch of the metatibia, the third segment of the protarsus is wider than it is long, and the fifth segment is one and a half times longer than it is wide.
67. Polygenis (Gephyropsylla) klagesi samuelis (Jordan & Rothschild 1923)
Host: Aegialomys xanthaeolus; Didelphis marsupialis; Myoprocta pratti Pocock, 1913; Proechimys simonsi.
Distribution in Peru: Isla Matapalo, Tumbes; Pucallpa, Ucayali.
Specificity: Euryxenous (Rodentia and Didelphimorphia).
Reference: [17, 57]
This species is distributed in the Caribbean, Andean and Amazonian subregions. In addition, it has been reported in Colombia [100] and Brazil, parasitizing marsupials [101].
68. Polygenis (Polygenis) bohlsi bohlsi (Wagner, 1901)
Host: Akodon mollis; Aegialomys xanthaeolus; Lycalopex sp; Phyllotis andium; Thomasomys taczanowskii (Thomas, 1882).
Distribution in Peru: Catacaos, Huancabamba, La Laguna, Suyo, Piura.
Specificity: Euryxenous.
Reference: [17]
Very wide distribution, found in Argentina, Bolivia, Brazil, Colombia, Ecuador, French Guaiana, Paraguay, Peru and Venezuela. Smit [17] mentions Phyllotis auritus (=Reithrodon auritus) (Fischer, 1814)) as a host of P. (Polygenis) bohlsi bohlsi, however, this mammal is not known from Peru [37].
69. Polygenis (Polygenis) brachinus Jordan, 1950
Host: Aegialomys xanthaeolus; Akodon mollis; Nectomys apicalis Peters, 1861; Rattus rattus; Sylvilagus sp; Thomasomys gracilis Thomas, 1917.
Distribution in Peru: Cascas, El Sauce, Pueblo Nuevo, Cajamarca; Chanchamayo, Junín; Cajas, Canchaque, Huancabamba, Huaylas, Pundín, Siclamache, Piura.
Specificity: Euryxenous.
Reference: [12, 17, 52]
Described from specimens collected from wild rabbits in Huancabamba, Piura. This species has very characteristic genitalia, which differs noticeably from closely related species, making it easier to recognize. The spermatheca of females has a very short tail, smaller than that of P. litargus; males have a ventral division close to the row of setae on sternites VIII.
70. Polygenis (Polygenis) impavidus Johnson, 1957
Host: Thomasomys sp.
Distribution in Peru: Limapunche, Cusco; Sandia, Tambopata, San Juan, Limbani, Puno.
Specificity: Monoxenous.
Reference: [12, 17]
This species has a geographical distribution in the Andean subregion. It is characterized by presenting 6 notches in the metatibia, including the apical notch, females are distinguished by having the anal stylet three and a half times longer than wide, and the males have the apical part of the coiled internal tube of the aedeagus very small and barely completes one turn.
71. Polygenis (Polygenis) litargus (Jordan & Rothschild 1923)
Host: Aegialomys sp; A. xanthaeolus; Akodon mollis; Akodon sp; Didelphis marsupialis; Didelphis sp; Euryoryzomys nitidus (Thomas, 1884); Neacomys spinosus; Oligoryzomys arenalis, O. destructor; Oxymycterus inca Thomas, 1900; Phyllotis amicus; P. andium; Proechimys decumanus; Proechimys sp; Rattus norvegicus; R. rattus; Rhipidomys leucodactylus; Sigmodon peruanus; Simosciurus nebouxii (I. Geoffroy St.- Hilaire, 1855); Sylvilagus sp; Thomasomys sp.
Distribution in Peru: Valle del Rio Santa, Choquechaca, Hauzlas, Llampa, Ancash; Cascas, El Sauce, El Mirador, Pueblo Nuevo, Cajamarca; Nazca, Ica; Verrugas Cañon, Junín; Campiña La Merced, Hacienda Laredo, Trujillo, La Libertad; Pimental, Lambayeque; Sapillica, Ayabaca, Montero, Suyo, Paimas, Catacaos, La Laguna, Cazaderos, Huancabamba, Huaylas, Jabonillos, Lancones, Quebrada Pasayo, Socchabamba, Piura; Aguas Verdes, Isla Matapalo, La Palma, Papayal, Tumbes.
Specificity: Euryxenous.
Reference: [11, 12, 17, 20, 52, 69, 102]
Macchiavello [11] mentions that, on the northern border of Peru, Polygenis litargus lives in open fields and is found as a unique species in various species of wild rodents, and highlights its potential as a vector of plague; it can also occasionally feed on humans.
72. Polygenis (Polygenis) litus (Jordan & Rothschild 1908)
Host: Simosciurus nebouxii; “Cricetinae”.
Distribution in Peru: Lancones, Piura.
Specificity: Polyxenous.
Reference: [11, 12, 17]
Macchiavello, [11]; Johnson, [12], mention this species in a mouse belonging to the subfamily Cricetinae. The host in Peru was reported as Sciurus stramineus nebouxii, but the only species of the genus Simosciurus in Peru is S. nebouxii [37].
73. Polygenis (Polygenis) roberti roberti (Rothschild, 1905)
Host: Unknown.
Distribution in Peru: Sivia, Cusco.
Specificity: Euryxenous.
Reference: [17]
Reported in the Amazon, Andean and Brazilian subregions. It has also been identified in Argentina [103]. In Brazil, P. r. roberti was found parasitizing marsupials, felines and rodents.
74. Polygenis (Polygenis) roberti beebei (I.Fox, 1947)
Host: Dasypus novemcinctus Linnaeus, 1758; Euryoryzomys nitidus; Oligoryzomys arenalis (Thomas, 1913).
Distribution in Peru: Sivia, Apurímac; Hacienda Cadena: Marcapata, Cusco.
Specificity: Euryxenous.
Reference: [12, 17]
Johnson, [12] mentions this species on O. longicaudatus in a locality mentioned as “Frontera con Ecuador” probably referring to some locality in the department of Tumbes and to another host, O. arenalis. It is distributed in the Amazon, Andean and Caribbean subregions, and is present in Argentina [104], Venezuela [105] and Costa Rica [106].
75. Polygenis (Polygenis) rimatus (Jordan, 1932)
Host: Aegialomys xanthaeolus; Akodon mollis.
Distribution in Peru: Llampa, Ancash; Huancabamba, Piura.
Specificity: Euryxenous.
Reference: [17]
This species has been reported in the Andean, Brazilian and Patagonian subregions. It has also been reported in northwestern Bolivia [107], and although rodents predominate among its hosts, in Brazil it has also been identified on marsupials [108].
76. Rhopalopsyllus australis tupinus (Jordan & Rothschild 1923)
Host: Myoprocta pratti; Speothos venaticus (Lund, 1842).
Distribution in Peru: Río Perené, Junín; Huancabamba, Piura; Pucallpa, Ucayali.
Specificity: Euryxenous.
Reference: [11, 12, 17, 57]
This species is distributed in the Brazilian subregion, and has also been reported in Colombia, Panama, Mexico, Trinidad, Venezuela and Guyana in a wide range of mammals such as marsupials, carnivores, lagomorphs, anteaters, ungulates and rodents [100, 109]. Jordan and Rothschild [110] report R. a. tupinus in Bolivia, parasitizing Speothos venaticus under its synonym Icticyon venaticus Lund 1842.
77. Rhopalopsyllus cacicus Jordan & Rothschild 1908
Host: Akodon mollis; Cavia porcellus; Dasypus novemcinctus; Phyllotis amicus; Sylvilagus brasiliensis.
Distribution in Peru: Río Perené, Junín; Matucana, Lima; Huancabamba, Soccha, Piura.
Specificity: Euryxenous.
Reference: [11, 12, 17, 111]
This species is found in the Andean subregion. It was first described in Peru from Tatusia novemcincta, synonym of Dasypus novemcinctus. It has been reported in Panama [112]; and Lewis [113] notes that subspecies of R. cacicus are distributed from Texas to Venezuela and Peru.
78. Rhopalopsyllus lugubris lugubris Jordan & Rothschild 1908
Host: Cuniculus paca (Linnaeus, 1766); Didelphis marsupialis; Felis catus.
Distribution in Peru: Oxapampa, Pasco; Piura; Pucallpa, Ucayali.
Specificity: Euryxenous.
Reference: [11, 12, 17, 20, 57]
This species has a wide distribution, being found in the Amazon, Andean, Brazilian and Caribbean subregions. In addition to Peru, it has been reported in Colombia, Argentina, Brazil, Bolivia, Venezuela, Costa Rica and Panama [100, 106, 114–116]. Although it is commonly associated with didelphids, it has been reported in cervids (Mazama gouazoubira) in Brazil [117, 118].
79. Tiamastus cavicola (Weyenbergh 1881)
Host: Aegialomys xanthaeolus; Canis familiaris; Cavia aperea; C. porcellus; Felis catus; Galea musteloides; Rattus norvegicus; R. rattus.
Distribution in Peru: Valle del Río Santa, Choquechaca, Hauzlas, Llampa, Ancash; Yura. Arequipa; Cutervo, El Páramo, Jaén, Cajamarca; La Raya, Cuzco; Hacienda San Pablo, Ica; Lago Junín, Junín; Otuzco, Trujillo, La Libertad; Villa Eten, Lambayeque; Moquegua; Huacho, Lima; Huancabamba, Sapillica, Ayabaca, Soccha, Piura: Tumbes.
Specificity: Euryxenous.
Reference: [11, 17, 20, 52, 69, 76, 77, 119]
This species is distributed in the Andean and Patagonian subregions. Dittmar [119] mentions that it is an indigenous species in South America, in the neotropical region, having a wide geographic range, found in Argentina, Peru, Chile and Bolivia.
Family Tungidae
80. Hectopsylla eskeyi Jordan, 1933
Host: Canis familiaris; Cavia aperea; Galea musteloides; Gallus gallus; Rattus norvegicus; R. rattus.
Distribution in Peru: Yura, Arequipa; Trujillo, La Libertad; Huacho, Lima.
Specificity: Euryxenous.
Reference: [11, 12]
It is distributed in the Andean subregion, and has also been reported in central, southern and western Ecuador and in La Paz, Bolivia [11, 52]. The males of most Hectopsylla species are differentiated by the presence of rows of 3 setae on tergites V and VI [120]. Likewise, in the male genitalia, H. eskeyi has the dorsal lobe (DA9) without lateral setae, unlike H. cypha, and four pairs of lateral setae on the fifth tarsal segment, unlike H. suarezi.
81. Hectopsylla psittaci Von Frauenfeld, 1860
Host: Didelphis sp., Eptesicus (Histiotus) montanus (Philippi & Landbeck, 1861).
Distribution in Peru: Trujillo, Huaura, La Libertad; Campiña, Huacho, Lima; Hacienda Las Vegas (Loreta, Barranca), Loreto.
Specificity: Euryxenous.
Reference: [12, 20, 52]
Although the genus Hectopsylla is common in mammals, this species is mainly associated with birds. In Peru it was reported in birds and also on a marsupial didelphid in Trujillo [52]. It has a distribution in the Andean and Brazilian subregion.
82. Hectopsylla suarezi C.Fox, 1929
Host: Aegialomys sp; Cavia aperea; C. porcellus; Rattus norvegicus, R. rattus; Sigmodon peruanus.
Distribution in Peru: Progreso, Huari, Chimbote, Ancash; Chincha, Ica; Trujillo, La Libertad; Ucupe, Lambayeque; Huacho, Lima; Huancabamba, Piura; Tumbes.
Specificity: Polyxenous.
Reference: [11, 12, 20, 52]
This species is distributed in the Andean subregion. It has also been reported in Ecuador, on Mus musculus and Cavia aperea [121, 122] and Chile [123]. It is distinguished from other Hectopsylla species by having five pairs of lateral setae on the fifth tarsal segment.
83. Hectopsylla pulex Haller, 1880
Host: Artibeus obscurus (Schinz, 1821); Eptesicus (Histiotus) sp; Molossus sp; Tadarida brasiliensis.
Distribution in Peru: Quince Mil, Cusco; Río Chinchao, Huánuco; Campiña, Huaura, Huacho, Lima; Santo Domingo, Puno.
Specificity: Polyxenous.
Reference: [11, 12, 20, 124, 125]
This species was recorded in bats, but under the name of its synonyms Rhynchopsyllus megastigmata Traub & Gammons, 1950 and Rhynchopsyllus pulex Haller, 1880. It has a distribution in the Amazon subregion, in the Brazilian subregion and the Patagonian subregion. It has been reported in Colombia, Panama, Argentina, Brazil, Bolivia, Chile, Ecuador and Venezuela [126,127,128].
84. Tunga penetrans Linnaeus, 1758
Host: Canis familiaris; Cavia sp; Felis catus; Gallus gallus; Homo sapiens; Rattus sp; Sus scrofa.
Distribution in Peru: Valle del Río Santa, Choquechaca, Hauzlas, Chimbote, Pomabamba, Pomabamba, Ancash; Cajamarca; Ica; Huacho, Lima; Piura.
Specificity: Euryxenous.
Reference: [11, 12, 20]
Macchiavello [11], reports this species throughout the Pacific coast in humans, dogs and pigs. It is also reported throughout the Amazon subregion, the Antilles, the Brazilian subregion, the Caribbean subregion, East America and West America.
85. Tunga trimamillata Pampiglione et al., 2002
Host: Cattles, sheeps and goats.
Distribution in Peru: Suyo, Ayabaca, Piura.
Specificity: Euryxenous.
Reference: [129]
This is a species that was reported only in Ecuador and northern Peru [130]; however, in 2013 it was reported for the first time in Brazil, in the state of São Paulo, Felixândia and Minas Gerais [131].
Family Pygiopsyllidae
86. Ctenidiosomus spillmanni Jordan, 1931
Host: Cavia sp.
Distribution in Peru: Huancabamba, Piura.
Specificity: Monoxenous.
Reference: [12, 52]
This species was described from a single female collected by Professor F. Spillmann in 1930 in Pichincha, Ecuador. Jordan [52] first described males of this species from fleas collected from rural dwellings and domestic animals in Huancabamba, Piura.
3.2. Flea-Host Interaction Network
The flea-host interaction network exhibited a modular structure, with a significant modularity (Q) and non-significant nestedness (NODF), and no internal nestedness within modules (non-significant NODFsm) (Table 1, Figure 5A). Modular structure grouped some phylogenetically related flea species, which can be more clearly observed for the genera Polygenis and Ectinorus (Figure S1). However, host are apparently more dispersed in in this regard (Figure S1).
Within this modular structure, two sigmodontine rodents (Calomys lepidus and Oligoryzomys destructor) and one rhopalopsyllid flea (Polygenis litargus) acted as connectors between modules; one sigmodontine rodent (Aegialomys xanthaeolus) and one rhopalopsyllid flea (Ectinorus viscachae) served as module hubs, while only one sigmodontine rodent functioned as a network hub (Akodon mollis) (Figure 5B). Despite the modular configuration, four flea species and three rodent species demonstrated high interaction counts and connectivity across the entire network: Polygenis litargus (Rhopalopsyllidae), Pulex irritans (Pulicidae), Neotyphloceras crassispina (Hystrichopsyllidae), Cleopsylla townsendi (Stephanocircidae), Akodon mollis (Cricetidae), Rattus norvegicus (Muridae), and Phyllotis andium (Cricetidae) (Table 2).
4. Discussion
4.1. Parasite - Host Checklist
The main result found in the literature about fleas in Peru is that mammals are the dominant group for these insects and within rodents the family of cricetidae is the most relevant. This is expected compared to what is reported in the rest of the world because a large percentage of the diversity of fleas in the world are associated with mammals (more than 94% of the species), while their relationship with birds is much less common. In an analysis of the associations between fleas and hosts carried out by Medvedev and Lobanov [132] and Medvedev [133] they showed that 70% of all associations between fleas and mammals involved rodents [39]. That is, rodents represent 82% of all specific and/or main hosts of fleas. This flea-rodent association is dominant in all parts of the world (except Australia where the main hosts are marsupials). Flea hosts in the Neotropics are usually sigmodontine rodents (Cricetidae) and caviomorphs (Caviidae, Chinchillidae, Capromyidae, Octodontidae) [7].
Flea species infesting birds in Peru only occupied 4.8% of the total number of hosts recorded. In the global diversity, about 214 flea species are known to infest birds, although only 60 species (approximately 3% of the total number of flea species) are considered specific parasites of birds [134, 135]. We found no references of fleas infesting other vertebrates such as reptiles. Flea infestations in reptiles are accidental, although they are capable of digesting the blood of these hosts [136]. Most flea species are relatively specific parasites, and may occasionally be found on hosts other than their preferred and/or primary hosts, and host selection appears to be influenced by environmental ecological factors, such as nests and burrows [137]. This could explain what happened with the species P. (Polygenis) litargus, P. irritans, C. canis and E. gallinacea, which were found parasitizing a larger number of hosts in Peru. Another reason why we might find fleas on non-specific hosts is because it is common to observe mammal species (usually mice) occupying the burrows of other animals, such as bird nests occupied by mice, mouse burrows occupied by bats, and vice versa. Finally, those occupying the site abandoned by another species often become infested with the fleas that were there, since the immature forms of the fleas remain in their habitats. In such circumstances, the presence of fleas on an animal may be purely accidental and the association may be seen as inauthentic, causing it to be a false correspondence [24, 25].
Regarding the spatial distribution of fleas, unlike most free-living species, the spatial distribution of parasites is not continuous, but consists of a set of “islands” or inhabited patches more or less uniformly represented by host organisms, while the environment between these patches is decidedly unfavorable [21]. In most fleas, a “habitat patch” also includes the host burrow, nest, or den. However, this does not negate the fragmented character of the spatial distribution of a conspecific flea assemblage. This assemblage is fragmented between (a) host individuals; (b) host species within a location; and (c) locations [21]. The main countries in South America that addressed this problem were Brazil [138,139,140], Argentina [141, 142] and Chile [143].
4.2. Flea-Host Interaction Network
The modular structure of the flea-host interaction network assembled for the Peruvian territory indicates the specificity in these parasitic interactions. The absence of nestedness at both the network and module scales supports this specificity, as there are no notable flea or host generalists. This modular structure aligns with patterns observed in various host-parasite interaction networks, which tend to exhibit modularity due to the selective behavior of parasites [26, 144]. Future studies could constitute a more diverse network which probably would maintain the modular configuration but with internally nested modules, as has been observed in other large-scale host-parasite networks [43]. In a more comprehensive network it would be possible to further explore these interactions considering phylogenetic relationships [43].
Five flea species emerged as central to the network's structure, with P. litargus being particularly significant in both the overall network and its modular configuration. Among the hosts, rodents were the primary key species, predominantly from the Sigmodontinae subfamily, with A. mollis being crucial in both the overall network and the modular structure. This suggests that only a few flea species have the potential to infect a wide range of hosts, while certain host species are more likely to disperse fleas among multiple hosts. These species warrant further ecological research as they may drive flea diversity within Peruvian ecosystems [145, 146]. Additionally, these species may play a significant role in the transmission of diseases that could eventually affect humans [147]. Therefore, this network-based information is essential for advancing parasite ecology studies in such a biodiverse region [148].
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Summary of hosts and their fleas recorded in Peru; Figure S1: Details on the modular structure of the national-scale interaction network between fleas and their hosts in the Peruvian territory.
Author Contributions
Conceptualization, D.M.; methodology, D.M.; C.F.; J.P.; J.I. and M.L.; investigation, D.M.; C.F. and J.P.; writing—original draft preparation, D.M.; C.F.; J.P.; J.I. and M.L.; writing—review and editing, D.M.; C.F.; J.P.; J.I. and M.L.; visualization, D.M.; C.F.; J.P.; J.I. and M.L.; supervision, J.I. and M.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data are contained within the manuscript and Supplementary Materials.
Acknowledgments
The authors would like to thank Dale, W. (Universidad Nacional Agraria La Molina) for his comments, recommendations and contribution in sharing bibliographic material; Vergara, C. (Universidad Nacional Agraria La Molina) for sharing bibliographic material; Pazos, J. (Universidad Nacional Federico Villarreal) and Espinoza, F. (Universidad Ricardo Palma) for their collaboration in transcribing the data into the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Lewis, R.E. Fleas (Siphonaptera). In: Lane RP, Crosskey RW (eds) Medical Insects and Arachnids.
- Linardi, P.M. Fleas and diseases. In: Marcondes C (ed) Arthropod Borne Diseases.
- Bossard, R.L.; Lareschi, M.; Urdapilleta, M.; Cutillas, C.; Zurita, A. Flea (Insecta: Siphonaptera) Family Diversity. Diversity (Basel) 2023, 15, 1096–10.3390. [Google Scholar] [CrossRef]
- Murray, M.D.; Orton, M.N.; Cameron, A.S. The Antarctic Flea Glaciopsyllus Antarcticus Smit and Dunnet. Antarct Res Ser 2013, 10, 393–395. [Google Scholar]
- Medvedev, S.G. Fauna and host-parasite associations of fleas (Siphonaptera) in different zoogeographical regions of the world. II. Entomologicheskoe Obozrenie 2000, 79, 640–655. [Google Scholar]
- Medvedev, S.G. Fauna and host-parasite associations of fleas (Siphonaptera) in different zoogeographical regions of the World. I. Entomologicheskoe Obozrenie 2000, 79, 409–435. [Google Scholar]
- Medvedev, S.G.; Krasnov, B.R. Fleas: Permanent satellites of small mammals. In: Morand S, Krasnov BR, Poulin R (eds) Micromammals and Macroparasites: From Evolutionary Ecology to Management.
- Beaucournu, J.C.; Moreno, L.; González-Acuña, D. Fleas (Insecta-Siphonaptera) of Chile: A review. Zootaxa 2014, 3900, 151–203. [Google Scholar] [CrossRef]
- Lareschi, M.; Sanchez, J.; Autino, A. A review of the fleas (Insecta: Siphonaptera) from Argentina. Zootaxa 2016, 4103, 239–258. [Google Scholar] [CrossRef]
- Sanchez, J.P.; Berrizbeitia, M.F.L.; Ezquiaga, M.C. Host specificity of flea parasites of mammals from the Andean Biogeographic Region. Med Vet Entomol 2023, 37, 511–522. [Google Scholar] [CrossRef]
- Macchiavello, A. Siphonaptera de la Costa Sur-Occidental de América: (Primera lista y distribución Zoo-Geográfica). Bol Of Sanit Panam (Engl) 1948, 27, 412–460. [Google Scholar]
- Johnson, P.T. A classification of the siphonaptera of South America: with descriptions of new species.
- Hopkins, G.H.E.; Rothschild, M. An illustrated catalogue of the Rothschild collection of fleas (Siphonaptera) in the British Museum (Natural History), Volume IV.
- Smit, F.G.A.M. A new species of flea from the Galapagos Islands. Entomolog Ber 1970, 30, 244–247. [Google Scholar]
- Smit, F.G.A.M. A New South American Ceratophyllid bird-flea. Entomolog Ber 1976, 36, 65–67. [Google Scholar]
- Smit, F.G.A.M. A new bird-flea from Peru. Zool J Linn Soc 1978, 62, 189–192. [Google Scholar] [CrossRef]
- Smit, F.G.A.M. An illustrated catalogue of the Rothschild collection of fleas in the British Museum (Natural History) Volume VII. T: Oxford University Press.
- Schramm, B.A.; Lewis, R.E. Four new species of Plocopsylla (Siphonaptera: Stephanocircidae) from South America. J Med Entomol 1987, 24, 399–407. [Google Scholar] [CrossRef]
- Schramm, B.A.; Lewis, R.E. A taxonomic revision of the Flea, genus Plocopsylla Jordan, 1931 (Siphonaptera: Stephanocircidae).
- Hastriter, M.W.; Zyzak, M.D.; Soto, R.; Fernandez, R.; Solorzano, N.; Whiting, M.F. Fleas (Siphonaptera) from Ancash Department, Peru with the description of a new species, Ectinorus Alejoi (Rhopalopsyllidae), and the description of the male of Plocopsylla pallas (Rothschild, 1914) (Stephanocircidae). Annals of Carnegie Museum 2002, 71, 87–106. [Google Scholar] [CrossRef]
- Krasnov, B.R. Functional and evolutionary ecology of fleas: A model for ecological parasitology. Functional and Evolutionary Ecology of Fleas: A Model for Ecological Parasitology. [CrossRef]
- Johnson, K.P.; Weckstein, J.D.; Bush, S.E.; Clayton, D.H. The evolution of host specificity in dove body lice. Parasitology 2011, 138, 1730–1736. [Google Scholar] [CrossRef] [PubMed]
- Dick, C.W.; Dittmar, K. Parasitic Bat Flies (Diptera: Streblidae and Nycteribiidae): Host Specificity and Potential as Vectors. In: Klimpel S., Melhorn H. (eds) Bats (Chiroptera) as vectors of diseases and parasites. Parasitology Research Monographs, Volumen 5. H: Springer.
- Mendez, E. Mammalian-siphonapteran associations, the environment, and biogeography of mammals of southwestern Colombia. Quaestiones Entomologicae 1977, 13, 91–182. [Google Scholar]
- Acosta, R. Relación huésped-parásito en pulgas (Insecta: Siphonaptera) y roedores (Mammalia: Rodentia) del estado de Querétaro, México. Folia Entomol Mex 2005, 44, 37–47. [Google Scholar]
- Runghen, R.; Poulin, R.; Monlleó-Borrull, C.; Llopis-Belenguer, C. Network Analysis: Ten Years Shining Light on Host–Parasite Interactions. Trends Parasitol 2021, 37, 445–455. [Google Scholar] [CrossRef]
- Vázquez, D.P.; Peralta, G.; Cagnolo, L.; Santos, M. Ecological interaction networks. What we know, what we don’t, and why it matters. Ecologia Austral 2022, 32, 670–697. [Google Scholar] [CrossRef]
- Delmas, E.; Besson, M.; Brice, M.-H.; et al. Analyzing ecological networks of species interactions. Biological Reviews 2019, 94, 16–36. [Google Scholar] [CrossRef]
- Pilosof, S.; Fortuna, M.A.; Vinarski, M. V.; Korallo-Vinarskaya, N.P.; Krasnov, B.R. Temporal dynamics of direct reciprocal and indirect effects in a host-parasite network. Journal of Animal Ecology 2013, 82, 987–996. [Google Scholar] [CrossRef]
- Llaberia-Robledillo, M.; Balbuena, J.A.; Sarabeev, V.; Llopis-Belenguer, C. Changes in native and introduced host–parasite networks. Biol Invasions 2022, 24, 543–555. [Google Scholar] [CrossRef]
- dos Santos Cardoso, T.; de Andreazzi, C.S.; Junior, A.M.; Gentile, R. Functional traits shape small mammal-helminth network: patterns and processes in species interactions. Parasitology 2021, 148, 947–955. [Google Scholar] [CrossRef] [PubMed]
- Pilosof, S.; Morand, S.; Krasnov, B.R.; Nunn, C.L. Potential parasite transmission in multi-host networks based on parasite sharing. PLoS One 2015, 10, e0117909–10.1371. [Google Scholar] [CrossRef] [PubMed]
- Bellay, S.; Oda, F.H.; Campião, K.M.; Yamada, F.H.; Takemoto, R.M.; de Oliveira, E.F. Host-Parasite Networks: An Integrative Overview with Tropical Examples. In: Dáttilo W, Rico-Gray V (eds) Ecological Networks in the Tropics:an integrative overview of species interactions from some of the most species-rich habitats on Earth.
- Poulin, R. Network analysis shining light on parasite ecology and diversity. Trends Parasitol 2010, 26, 492–498. [Google Scholar] [CrossRef] [PubMed]
- Whiting, M.F.; Whiting, A.S.; Hastriter, M.W.; Dittmar, K. A molecular phylogeny of fleas (Insecta: Siphonaptera): Origins and host associations. Cladistics 2008, 24, 677–707. [Google Scholar] [CrossRef]
- Hastriter, M.W.; Bossard, R.L. Robert E. Lewis World Species Flea (Siphonaptera) List. 20. [CrossRef]
- Pacheco, V.; Diaz, S.; Graham-Angeles, L.; Flores-Quispe, M.; Calizaya-Mamani, G.; Ruelas, D.; Sánchez-Vendizú, P. Updated list of the diversity of mammals from Peru and a proposal for its updating. Rev Peru Biol 2021, 28, e21019–10.15381. [Google Scholar]
- Jameson, E.W. Pleioxenous host-restriction in fleas. J Nat Hist 1985, 19, 861–876. [Google Scholar] [CrossRef]
- Medvedev, S. Specific features of the distribution and host associations of fleas (Siphonaptera). Entomol Rev 2002, 82, 1165–1177. [Google Scholar]
- Llopis-Belenguer, C.; Balbuena, J.A.; Blasco-Costa, I.; Karvonen, A.; Sarabeev, V.; Jokela, J. Sensitivity of bipartite network analyses to incomplete sampling and taxonomic uncertainty. Ecology 2023, 104, e3974–10.1002. [Google Scholar] [CrossRef]
- Fründ, J.; Mccann, K.S.; Williams, N.M. Sampling bias is a challenge for quantifying specialization and network structure: Lessons from a quantitative niche model. Oikos 2016, 125, 502–513. [Google Scholar] [CrossRef]
- Beckett, S.J. Improved community detection in weighted bipartite networks. R Soc Open Sci 2016, 3, 140536–10.1098. [Google Scholar] [CrossRef] [PubMed]
- Felix, G.M.; Pinheiro, R.B.P.; Poulin, R.; Krasnov, B.R.; Mello, M.A.R. The compound topology of host–parasite networks is explained by the integrative hypothesis of specialization. Oikos. [CrossRef]
- Vázquez, D.P.; Melián, C.J.; Williams, N.M.; Blüthgen, N.; Krasnov, B.R.; Poulin, R. Species abundance and asymmetric interaction strength in ecological networks. Oikos. [CrossRef]
- Felix, G.M.; Pinheiro, R.B.P.; Jorge, L.R.; Lewinsohn, T.M. A framework for hierarchical compound topologies in species interaction networks. Oikos 2022, 12, e09538–10.1111. [Google Scholar] [CrossRef]
- Messeder, J.V.S.; Guerra, T.J.; Dáttilo, W.; Silveira, F.A.O. Searching for keystone plant resources in fruit-frugivore interaction networks across the Neotropics. Biotropica 2020, 52, 857–870. [Google Scholar] [CrossRef]
- Guimerà, R.; Amaral, L.A.N. Functional cartography of complex metabolic networks. Nature 2005, 433, 895–900. [Google Scholar] [CrossRef]
- Olesen, J.M.; Bascompte, J.; Dupont, Y.L.; Jordano, P. The modularity of pollination networks. Proc Natl Acad Sci U S A 2007, 104, 19891–19896. [Google Scholar] [CrossRef]
- Dormann, C.F.; Gruber, B.; Fründ, J. Introducing the bipartite Package: Analysing Ecological Networks. R new 2008, 8, 8–11. [Google Scholar]
- Hastriter, M.W.; Schlatter, R.P. Revision of the fleas in the subgenus Dasypsyllus (Neornipsyllus) (Siphonaptera: Ceratophyllidae). Annals of Carnegie Museum 2006, 75, 247–257. [Google Scholar] [CrossRef]
- Zurita, A.; Callejón, R.; De Rojas, M.; Cutillas, C. Morphological and molecular study of the genus Nosopsyllus (Siphonaptera: Ceratophyllidae). Nosopsyllus barbarus (Jordan & Rothschild 1912) as a junior synonym of Nosopsyllus fasciatus (Bosc d’Antic, 1800). Insect Syst Evol 2018, 49, 81–101. [Google Scholar]
- Jordan, K. Notes on a collection of fleas from Peru. Bull World Hlth Org 1950, 2, 597–609. [Google Scholar]
- Smit, F.G.A.M. Key to the genera and subgenera of Ceratophyllidae. In: Traub R, Rothschild M, Haddow J (eds) The Rothschild collection of fleas. The Ceratophyllidae: key to the genera and host relationships.
- Johnson, P.T. Notes on Pleochaetis Jordan, 1933, from Colombia, with the description of a new species (Siphonaptera: Ceratophyllidae). Journal of the Washington Academy of Sciences 1954, 44, 289–296. [Google Scholar]
- Johnson, P.T. Myodopsylla setosa and Tiarapsylla bella, New Species of Fleas from Peru. Journal of the New York Entomological Society 1954, 62, 193–205. [Google Scholar]
- Autino, A.G.; Claps, G.L.; Barquez, R.M.; Díaz, M.M. Ectoparasitic insects (Diptera: Streblidae and siphonaptera: Ischnopsyllidae) of bats from iquitos and surrounding areas (Loreto, Peru). Mem Inst Oswaldo Cruz 2011, 106, 917–925. [Google Scholar] [CrossRef] [PubMed]
- Darskaya, N.F.; Malygin, V.M. O blokhakh mlekopitaiushchikh iz basseĭna reki Ukaiali (Peruanskaia Amazoniia) [On the fleas of mammals from the Ucayali river basin, Peruvian Amazonia]. Parazitologiya 1996, 30, 187–190. [Google Scholar]
- Zurita, A.; Rivero, J.; García-Sánchez, Á.M.; Callejón, R.; Cutillas, C. Morphological, molecular and phylogenetic characterization of Leptopsylla segnis and Leptopsylla taschenbergi (Siphonaptera). Zool Scr 2022, 51, 741–754. [Google Scholar] [CrossRef]
- Traub, R. Records and descriptions of fleas from Peru. Proc Entomol Soc Wash 1952, 54, 1–22. [Google Scholar]
- López-Berrizbeitia, M.F.; Sanchez, J.; Barquez, R.M.; Díaz, M. Taxonomic revision of the flea genus Agastopsylla Jordan & Rothschild 1923 (Siphonaptera: Ctenophthalmidae). An Acad Bras Cienc 2020, 92, e20181136–10.1590. [Google Scholar]
- Jordan, K. Some Siphonaptera from South America. Novitates zoologicae : a journal of zoology in connection with the Tring Museum 1936, 39, 305–310.
- Sanchez, J.; Amor, V.; Bazán-León, E.A.; Vásquez, R.A.; Lareschi, M. Redescription of Neotyphloceras chilensis Jordan, new status (Siphonaptera: Ctenophthalmidae: Neotyphloceratini). Zootaxa 2012, 3259, 51–57. [Google Scholar] [CrossRef]
- Rothschild, N.C. New Siphonaptera from Peru. Novitates zoologicae : a journal of zoology in connection with the Tring Museum 1914, 21, 239–251. [Google Scholar] [CrossRef]
- Fuller, H.S. Notes on Neotropical Siphonaptera. Revista de Entomología 1942, 13, 39–44. [Google Scholar]
- Sanchez, J.; Lareschi, M. Two new species of Neotyphloceras (Siphonaptera: Ctenophthalmidae) from Argentinean Patagonia. Zootaxa 2014, 3784, 159–170. [Google Scholar] [CrossRef]
- López Berrizbeitia, M.F.; Sanchez, J.; Díaz, M.M.; Barquez, R.M.; Lareschi, M. Redescription of Neotyphloceras crassispina hemisus Jordan (Siphonaptera: Ctenophthalmidae: Neotyphloceratini). Journal of Parasitology 2015, 101, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, J.; Lareschi, M.; Salazar-Bravo, J.; Gardner, S.L. Fleas of the genus Neotyphloceras associated with rodents from Bolivia: new host and distributional records, description of a new species and remarks on the morphology of Neotyphloceras rosenbergi. Med Vet Entomol 2018, 32, 462–472. [Google Scholar] [CrossRef] [PubMed]
- López Berrizbeitia, M.F.; Hastriter, M.W.; Díaz, M.M. A new flea species of the genus cleopsylla (Siphonaptera: Stephanocircidae) from Northwestern Argentina. Journal of Parasitology 2016, 102, 514–519. [Google Scholar] [CrossRef]
- Pozo, E.J.; Troncos, G.; Palacios F, A.; Arévalo, F.; Carrión, G.; Laguna-Torres, A. Distribución y hospederos de pulgas (Siphonaptera) en la provincia de Ayabaca, Piura-1999. Rev Peru Med Exp Salud Publica 2005, 22, 316–320. [Google Scholar]
- Lareschi, M.; Autino, A.G.; Díaz, M.M.; Barquez, R.M. Taxonomy and distribution of Nonnapsylla wagner, 1938 (Siphonaptera: Stephanocircidae: Craneopsyllinae). Journal of Parasitology 2011, 97, 954–955. [Google Scholar] [CrossRef] [PubMed]
- Pucu, E.; Lareschi, M.; Gardner, S.L. Bolivian ectoparasites: A survey of the fleas of ctenomys (Rodentia: Ctenomyidae). Comp Parasitol 2014, 81, 114–118. [Google Scholar] [CrossRef]
- Jordan, K. Three new South American fleas. Novitates zoologicae : a journal of zoology in connection with the Tring Museum 1931, 36, 311–316.
- Estares P., L.; Chávez V., A.; Casas A., E. Ectoparásitos en caninos de los distritos de la zona climática norte de Lima Metropolitana. Revista de Investigaciones Veterinarias del Perú 2000, 11, 72–76. [Google Scholar] [CrossRef]
- Nuntón, J.; Quintana, H.; Vivar, E. Prevalencia de ectoparásitos y endoparásitos en Canis Familiaris sacrificados en Tumbes; julio - diciembre, 2013. Manglar 2015, 10, 93–97. [Google Scholar] [CrossRef]
- Flores-Mendoza, C.; Florin, D.; Felices, V.; Pozo, E.J.; Graf, P.C.F.; Burrus, R.G.; Richards, A.L. Detection of Rickettsia parkeri from within Piura, Peru, and the first reported presence of candidatus rickettsia andeanae in the tick Rhipicephalus sanguineus. Vector-Borne and Zoonotic Diseases 2013, 13, 505–508. [Google Scholar] [CrossRef]
- Rizzo, M.F.; Billeter, S.A.; Osikowicz, L.; Luna-Caipo, D. V.; Cáceres, A.G.; Kosoy, M. Fleas and Flea-associated Bartonella species in dogs and cats from Peru. J Med Entomol 2015, 52, 1374–1377. [Google Scholar] [CrossRef]
- Rizzo, M.F.; Osikowicz, L.; Cáceres, A.G.; Luna-Caipo, V.D.; Suarez-Puyen, S.M.; Bai, Y.; Kosoy, M. Identification of Bartonella rochalimae in Guinea Pigs (Cavia porcellus) and fleas collected from rural peruvian households. American Journal of Tropical Medicine and Hygiene 2019, 101, 1276–10.4269. [Google Scholar] [CrossRef] [PubMed]
- Dittmar De La Cruz, K.; Ribbeck, R.; Daugschies, A. Vorkommen und verbreitung von ektoparasiten bei meerschweinchen (Cavia spp.) in Peru, Südamerika. Berl Munch Tierarztl Wochenschr 2003, 116, 102–107. [Google Scholar]
- Naupay I., A.; Castro H., J.; Caro C., J.; Sevilla D., L.; Hermosilla J., J.; Larraín L., K.; Quispe S., C.; Panana R., O. Ectoparásitos en palomas Columba livia comercializadas en un mercado del distrito de San Martín de Porres, Lima, Perú. Revista de Investigaciones Veterinarias del Perú 2015, 26, 259–265. [Google Scholar] [CrossRef]
- Castro, J.; Naupay, A.; Orozco, K.; Rodríguez, S.; Díaz, Y.; Navarro, J.; Purca, N. Ectoparásitos de Columba livia linnaeus, 1758 (aves columbiformes) del distrito de Carmen de la Legua, callao, Perú. The Biologist 2018, 15, 425–435. [Google Scholar] [CrossRef]
- Ewing, H.E. (Henry E. New North American Siphonaptera. Proceedings of the Biological Society of Washington 1940, 53, 35–37. [Google Scholar]
- Lewis, R.E. Notes on the Geographical Distribution and Host Preferences in the Order Siphonaptera: Part 1. Pulicidae1, 2. J Med Entomol 1972, 9, 511–520. [Google Scholar] [CrossRef]
- Lareschi, M.; Venzal, J.M.; Nava, S.; Mangold, A.J.; Portillo, A.; Palomar-Urbina, A.M.; Oteo-Revuelta, J.A. The human flea Pulex irritans (Siphonaptera: Pulicidae) in northwestern Argentina, with an investigation of Bartonella and Rickettsia spp. Rev Mex Biodivers 2018, 89, 375–381. [Google Scholar] [CrossRef]
- Dittmar, K. Evaluation of ectoparasites on the guinea pig mummies of El Yaral and Moquegua Valley, in Southern Peru. Chungara 2000, 32, 123–125. [Google Scholar] [CrossRef]
- Dittmar, K.; Mamat, U.; Whiting, M.; Goldmann, T.; Reinhard, K.; Guillen, S. Techniques of DNA-studies on Prehispanic Ectoparasites (Pulex sp., Pulicidae, Siphonaptera) from Animal Mummies of the Chiribaya Culture, Southern Peru. Mem Inst Oswaldo Cruz 2003, 98, 53–58. [Google Scholar] [CrossRef]
- Ortiz-Cusma, J.F.; Martínez-Bravo, K.M.; Iglesias-Osores, S. Prevalencia de Xenopsylla Cheopis en Rattus Rattus en el distrito de Salas, Lambayeque. Revista de la Facultad de Medicina Humana 2022, 22, 258–265. [Google Scholar] [CrossRef]
- Boyer, S.; Gillespie, T.R.; Miarinjara, A. Xenopsylla cheopis (rat flea). Trends Parasitol 2022, 38, 607–608. [Google Scholar] [CrossRef] [PubMed]
- Gárate, I.; Jiménez, P.; Flores, K.; Espinoza, B. Registro de Xenopsylla cheopis como hospedero intermediario natural de Hymenolepis diminuta en Lima, Perú. Rev Peru Biol 2011, 18, 249–252. [Google Scholar] [CrossRef]
- Smit, F.G.A.M. Siphonaptera (Fleas). In: Smit FGV (ed) Insect and other arthropods of medical importance.
- Jordan, K. On Parapsyllus and some closely related genera of Siphonaptera. Eos (Washington DC) 1942, 18, 7–29. [Google Scholar]
- Hastriter, M.W.; Sage, R.D. A description of two new species of Ectinorus (Siphonaptera: Rhopalopsyllidae) from Laguna Blanca National Park, Neuquén Province, Argentina. Proc Entomol Soc Wash 2009, 111, 581–597. [Google Scholar] [CrossRef]
- Wagner, J. Aphaniptera aus Süd-Peru sowie Bemerkungen über die Fam. Stephanocircidae Wagn. Zeitschrift für Parasitenkunde 1937, 9, 698–716. [Google Scholar] [CrossRef]
- López-Berrizbeitia, M.F.; Pérez, M.J.; Barquez, R.M. Description of two new species of Ectinorus (Ectinorus) (Siphonaptera, Rhopalopsyllidae) from Argentina, including a morphometric approach. Acta Trop 2020, 211, 105612–10.1016. [Google Scholar] [CrossRef]
- Hastriter, M. A description of four new species of fleas (Insecta, Siphonaptera) from Angola, Ethiopia, Papua New Guinea, and Peru. Zookeys 2009, 8, 39–61. [Google Scholar] [CrossRef]
- Beaucournu, J.-C.; Gallardo, M.H. Siphonaptères du Chili; description de quatre espèces nouvelles. Bull Soc Entomol France 1991, 96, 185–203. [Google Scholar] [CrossRef]
- Beaucournu, J.C.; Belaz, S.; Muñoz-Léal, S.; González-Acuña, D. A new flea, Ectinorus (Ectinorus) insignis n. sp. (Siphonaptera, Rhopalopsyllidae, Parapsyllinae), with notes on the subgenus Ectinorus in Chile and comments on unciform sclerotization in the superfamily Malacopsylloidea. Parasite 2013, 20, 1–12. [Google Scholar] [CrossRef]
- López Berrizbeitia, M.F.; Hastriter, M.W.; Barquez, R.M.; Díaz, M.M. Fleas of the genus Tetrapsyllus (Siphonaptera:Rhopalopsyllidae) associated with rodents from Northwestern Argentina. Int J Parasitol Parasites Wildl 2019, 9, 80–89. [Google Scholar] [CrossRef]
- Gimenez, D.F.; Ciccarelli, C.A.; de la Barrera, J.M. Siphonaptera de Mendoza. An Soc Cient Argent 1964, 178, 125–139. [Google Scholar]
- Beaucournu, J.-C.; Gallardo, M.H. Contribution à la faune du Chili. Puces nouvelles de la moitié nord [Siphonaptera]. Bull Soc Entomol France 1989, 94, 181–188. [Google Scholar] [CrossRef]
- Wells, E.A.; D’Alessandro, A.; Morales, G.A.; Angel, D. Mammalian wildlife diseases as hazards to man and livestock in an area of the Llanos Orientales of Colombia. J Wildl Dis 1981, 17, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Rafael, J.A. Notas & Comunicações Ocorrência de Polygenis Klagesi samuelis na Amazônia brasileira (Siphonaptera : Rhopaiopsyllidae). Acta Amazon 1982, 12, 231–232. [Google Scholar]
- Macchiavello, A. Estudios sobre peste selvetica en America del Sur: IV. Transmision experimental de la peste por polygenis litargus. Boletín de la Oficina Sanitaria Panamericana (OSP); 45 (2), ago 1958 1958, 45, 122–131. [Google Scholar]
- Zurita, A.; Lareschi, M.; Cutillas, C. New insights into the taxonomy of Malacopsylloidea superfamily (Siphonaptera) based on morphological, molecular and phylogenetic characterization of Phthiropsylla agenoris (Malacopsyllidae) and Polygenis (Polygenis) rimatus (Rhopalopsyllidae). Diversity (Basel) 2023, 15, 308–10.3390. [Google Scholar] [CrossRef]
- Lareschi, M.; Linardi, P.M.; Autino, A.G.; Barquez, R.M.; Díaz, M.M. First report of Polygenis (Polygenis) roberti beebei (Fox, 1947) (Siphonaptera: Rhopalopsyllidae) in Argentina, with a new host record and morphological data. Syst Parasitol 2003, 56, 183–187. [Google Scholar] [CrossRef]
- Traub, R.; Johnson, P.T. Fleas collected during a plague survey in Venezuela. Bol Oficina Sanit Panam 1952, 32, 111–135. [Google Scholar]
- Durden, L.A.; Campbell, D.C. Fleas, Lice, and Epifaunistic Pseudoscorpions of Some Native Mammals in Northwestern Costa Rica. Comp Parasitol 2016, 83, 240–244. [Google Scholar] [CrossRef]
- Ramos-Yana, V.S.; Mollericona-Quispe, J.L.; Arteaga, D.; Bernal-Hoverud, N. Distribución y caracterización morfológica y molecular de pulgas (Insecta, Siphonaptera), sus hospederos roedores e implicancia en salud humana, Parque Madidi. Bio Scientia 2021, 4, 4–5. [Google Scholar]
- Lareschi, M.; Linardi, P.M. New data on the morphology of Polygenis (Polygenis) rimatus (Jordan)(Siphonaptera: Rhopalopsyllidae). Neotrop Entomol 2005, 34, 121–125. [Google Scholar] [CrossRef]
- Durden, L.A.; Bermúdez, S.; Vargas, G.A.; Sanjur, B.E.; Gillen, L.; Brown, L.D.; Greiman, S.E.; Eremeeva, M.E. Fleas (Siphonaptera) Parasitizing Peridomestic and Indigenous Mammals in Panamá and Screening of Selected Fleas for Vector-Borne Bacterial Pathogens. J Med Entomol 2021, 58, 1316–1321. [Google Scholar] [CrossRef] [PubMed]
- Jordan, K.; Rothschild, N.C. On the genera Rhopalopsyllus and Parapsyllus. Ectoparasites 1923, 1, 320–370. [Google Scholar]
- Jordan, K.; Rothschild, N.C. Revision of the non-combed eyed siphonaptera. Parasitology 1908, 1, 1–100. [Google Scholar] [CrossRef]
- Bermúdez, S.C.; Miranda, R.C. Distribution of ectoparasites of Canis lupus familiaris L. (Carnivora: Canidae) from Panama. Rev MVZ Cordoba 2011, 16, 2274–2282. [Google Scholar]
- Lewis, R.E. New species of Palaeopsylla Wagner, 1903, from Nepal, with a discussion of the Remota species group (Siphonaptera: Hystrichopsyllidae). J Parasitol 1973, 59, 187–197. [Google Scholar] [CrossRef]
- Del Ponte, E. Notas sobre Suctoria argentinos. V. Nuevos datos sobre Rhopalopsyllidae. Rhopalopsyllinae. Rev Soc Entomol Argent 1963, 26, 1–4. [Google Scholar]
- Tipton, V.J.; Machado-Allison, C.E. Fleas of Venezuela. Brigham Young University Science Bulletin - Biological Series 1972, 17, 1–115. [Google Scholar]
- Botelho, J.R.; Linardi, P.M.; Williams, P.; Nagem, R.L. Alguns hospedeiros reais de ectoparasitos do município de Caratinga, Minas Gerais, Brasil. Mem Inst Oswaldo Cruz 1981, 76, 57–59. [Google Scholar] [CrossRef]
- Linardi, P.M. Checklist de Siphonaptera (Insecta) do Estado de São Paulo. Biota Neotrop 2011, 11, 607–617. [Google Scholar] [CrossRef]
- da Costa, A.L.M.; Teixeira, R.H.F.; Paschoalotti, M.H.; Gomes, R.P.; Felippi, D.A.; Franco, P.N. Controle de pulgas em cervo-do-pantanal (Blastocerus dichotomus) de cativeiro com uso de coleira com imidacloprida e flumetrina. Veterinária e Zootecnia 2019, 26, 1–7. [Google Scholar] [CrossRef]
- Dittmar, K. Arthropod and helminth parasites of the wild guinea pig, Cavia aperea, from the Andes and the Cordillera in Peru, South America. Journal of Parasitology 2002, 88, 409–411. [Google Scholar] [CrossRef] [PubMed]
- Blank, S.M.; Kutzscher, C.; Masello, J.F.; Pilgrim, R.L.C.; Quillfeldt, P. Stick-tight fleas in the nostrils and below the tongue: Evolution of an extraordinary infestation site in Hectopsylla (Siphonaptera: Pulicidae). Zool J Linn Soc 2007, 149, 117–137. [Google Scholar] [CrossRef]
- Macchiavello, A. Epidemiology of Plague in Ecuador. Am J Public Health Nations Health 1943, 33, 807–811. [Google Scholar] [CrossRef] [PubMed]
- Martinez, L.J. Plague in the City of Ambato, Ecuador. Proceedings 6th Pacific Sci Congr 1942, 5, 139–143. [Google Scholar]
- Macchiavello, A. A Note on Plague in Chile. Bol Oficina Sanit Panam 1933, 11, 909–915. [Google Scholar]
- Autino, A.G.; Claps, G.L.; Barquez, R.M. Insectos ectoparásitos de murciélagos de las Yungas de la Argentina. Acta Zool Mex 1999, 78, 119–169. [Google Scholar] [CrossRef]
- Autino, A.G.; Claps, G.E. Catalogue of the ectoparasitic insects of the bats of Argentina. Insecta mundi 2000, 14, 193–209. [Google Scholar]
- Linardi, P.M.; Guimarães, L.R. Sifonápteros do Brasil. Universidade De São Paulo.
- Luz, J.L.; de Moraes Costa, L.; Gomes, L.A.C.; Esbérard, C.E.L. The chiggerflea Hectopsylla pulex (Siphonaptera: Tungidae) as an ectoparasite of free-tailed bats (Chiroptera: Molossidae). Mem Inst Oswaldo Cruz 2009, 104, 567–569. [Google Scholar] [CrossRef]
- Ramírez-Chaves, H.E.; Tamayo-Zuluaga, A.F.; Henao-Osorio, J.J.; Cardona-Giraldo, A.; Ossa-López, P.A.; Rivera-Páez, F.A. The chiggerflea hectopsylla pulex (Siphonaptera: Tungidae): Infestation on molossus molossus (chiroptera: Molossidae) in the central andes of Colombia. Zoologia 2020, 37, e53092–10.3897. [Google Scholar] [CrossRef]
- Fioravanti, M.L.; Gustinelli, A.; Onore, G.; Pampiglione, S.; Trentini, M. Presence of Tunga trimamillata (Insecta, Siphonaptera) in Peru. Parasite 2006, 13, 85–86. [Google Scholar] [PubMed]
- Maco, V.; Maco, V.P.; Gotuzzo, E. Case report: An ectopic case of Tunga spp. infection in Peru. American Journal of Tropical Medicine and Hygiene 2010, 82, 1076–1078. [Google Scholar] [CrossRef] [PubMed]
- Linardi, P.M.; De Avelar, D.M.; Facury Filho, E.J. Establishment of Tunga trimamillata (Siphonaptera: Tungidae) in Brazil. Parasitol Res 2013, 112, 3239–3242. [Google Scholar] [CrossRef] [PubMed]
- Medvedev, S.G.; Lobanov, A.L. Information-Analytic System of the World Fauna of Fleas (Siphonaptera): Results and Prospects. Entomologicheskoe Obozrenie 1999, 78, 732–748. [Google Scholar]
- Medvedev, S.G. An Attempted system analysis of the evolution of the order of fleas (Siphonaptera). Lectures in memoriam N.A.
- Medvedev, S.G. Host-parasite relations in fleas (Siphonaptera). I. Entomol Rev 1997, 77, 200–215. [Google Scholar]
- Medvedev S., G. Host-parasite relations of fleas (Siphonaptera). II. Entomologicheskoe Obozrenie 1997, 76, 511–521. [Google Scholar]
- Vatschenok, V.S. Fleas–vectors of pathogens causing diseases in humans and animals. N: Leningrad, USSR.
- Whitaker, J.O.; Wren, Jr.W.J.; Lewis, R.E. Parasites. In: Genoways HH, Brown JH (eds) Biology of the Heteromyidae. Special Publication. N. 10. P: American Society of Mammalogists.
- Linardi, P.M.; Botelho, J.R.; Ximenez, A.; Padovani, C.R. Notes on ectoparasites of some small mammals from Santa Catarina State, Brazil. J Med Entomol 1991, 28, 183–185. [Google Scholar] [CrossRef]
- De Moraes, L.B.; Paolinetti Bossi, D.E.; Linhares, A.X. Siphonaptera Parasites of Wild Rodents and Marsupials Trapped in Three Mountain Ranges of the Atlantic Forest in Southeastern Brazil. Mem Inst Oswaldo Cruz 2003, 98, 1071–1076. [Google Scholar] [CrossRef]
- Pinto, I.D.S.; Botelho, J.R.; Costa, L.P.; Leite, Y.L.R.; Linardi, P.M. Siphonaptera associated with wild mammals from the central atlantic forest biodiversity corridor in southeastern Brazil. J Med Entomol 2009, 46, 1146–1151. [Google Scholar] [CrossRef]
- Lareschi, M. The relationship of sex and ectoparasite infestation in the water rat Scapteromys aquaticus (Rodentia: Cricetidae) in La Plata, Argentina. Rev Biol Trop 2006, 54, 673–679. [Google Scholar] [CrossRef]
- Lareschi, M.; Krasnov, B.R. Determinants of ectoparasite assemblage structure on rodent hosts from South American marshlands: The effect of host species, locality and season. Med Vet Entomol 2010, 24, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Landaeta-Aqueveque, C.; Moreno Salas, L.; Henríquez, A.L.; Silva-de la Fuente, M.C.; González-Acuña, D. Parasites of Native and Invasive Rodents in Chile: Ecological and Human Health Needs. Front Vet Sci 2021, 8, 643742–10.3389. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, R.B.P.; Felix, G.M.F.; Lewinsohn, T.M. Hierarchical compound topology uncovers complex structure of species interaction networks. Journal of Animal Ecology 2022, 91, 2248–2260. [Google Scholar] [CrossRef] [PubMed]
- Anderson, T.K.; Sukhdeo, M.V.K. Host centrality in food web networks determines parasite diversity. PLoS One 2011, 6, e26798–10.1371. [Google Scholar] [CrossRef] [PubMed]
- Dallas, T.; Cornelius, E. Co-extinction in a host-parasite network: Identifying key hosts for network stability. Sci Rep 2015, 5, 13185–10.1038. [Google Scholar] [CrossRef]
- Gómez, J.M.; Nunn, C.L.; Verdú, M. Centrality in primate-parasite networks reveals the potential for the transmission of emerging infectious diseases to humans. Proc Natl Acad Sci U S A 2013, 110, 7738–7741. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeler, R.A.; Mittermeler, C.G.; Da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
Figure 1.
Summary of the number of records of flea species and their hosts cited in each of the departments of Peru.
Figure 1.
Summary of the number of records of flea species and their hosts cited in each of the departments of Peru.
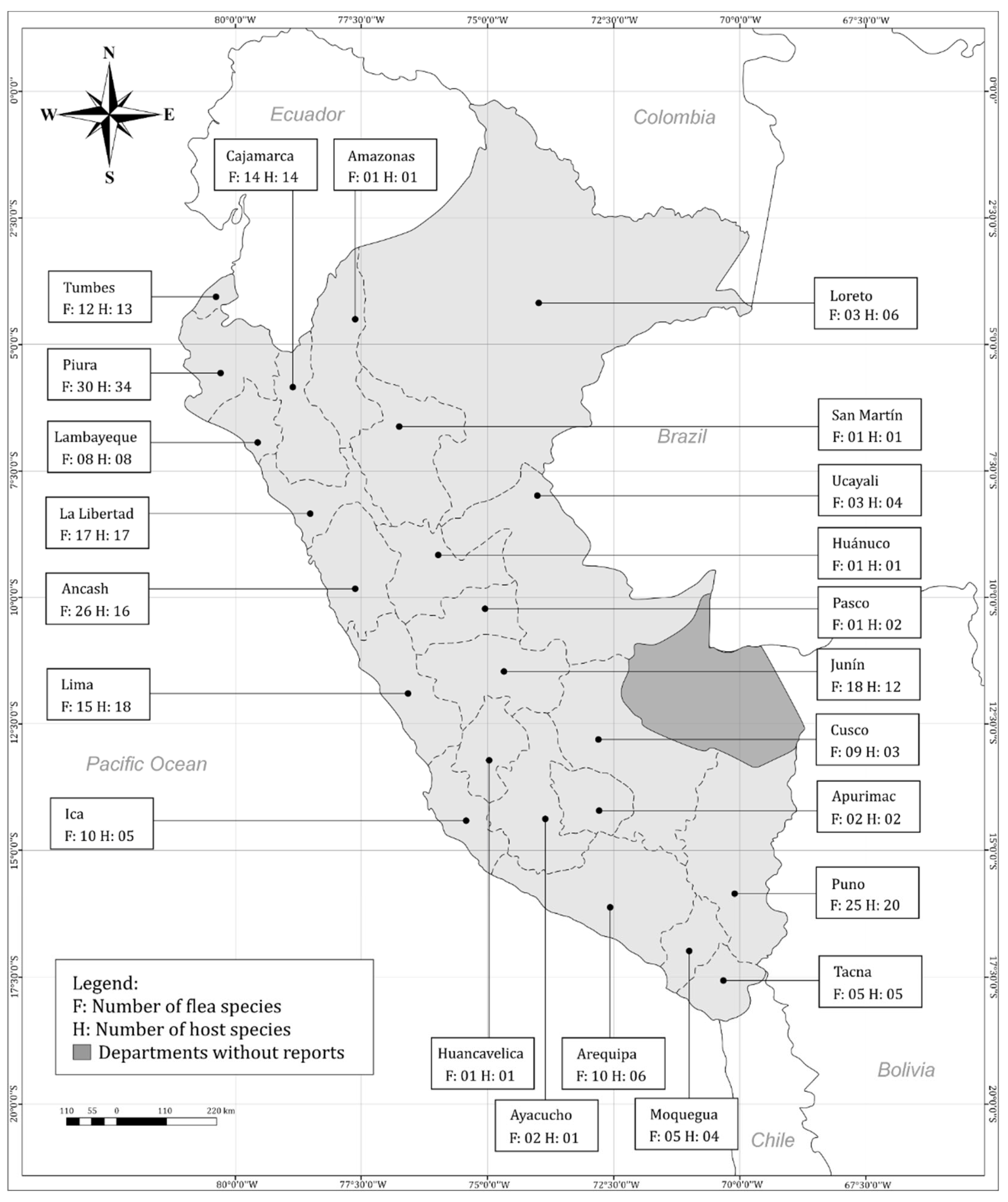
Figure 2.
Host-parasite bipartite interaction network between fleas vs birds and mammals recorded in Peru.
Figure 2.
Host-parasite bipartite interaction network between fleas vs birds and mammals recorded in Peru.
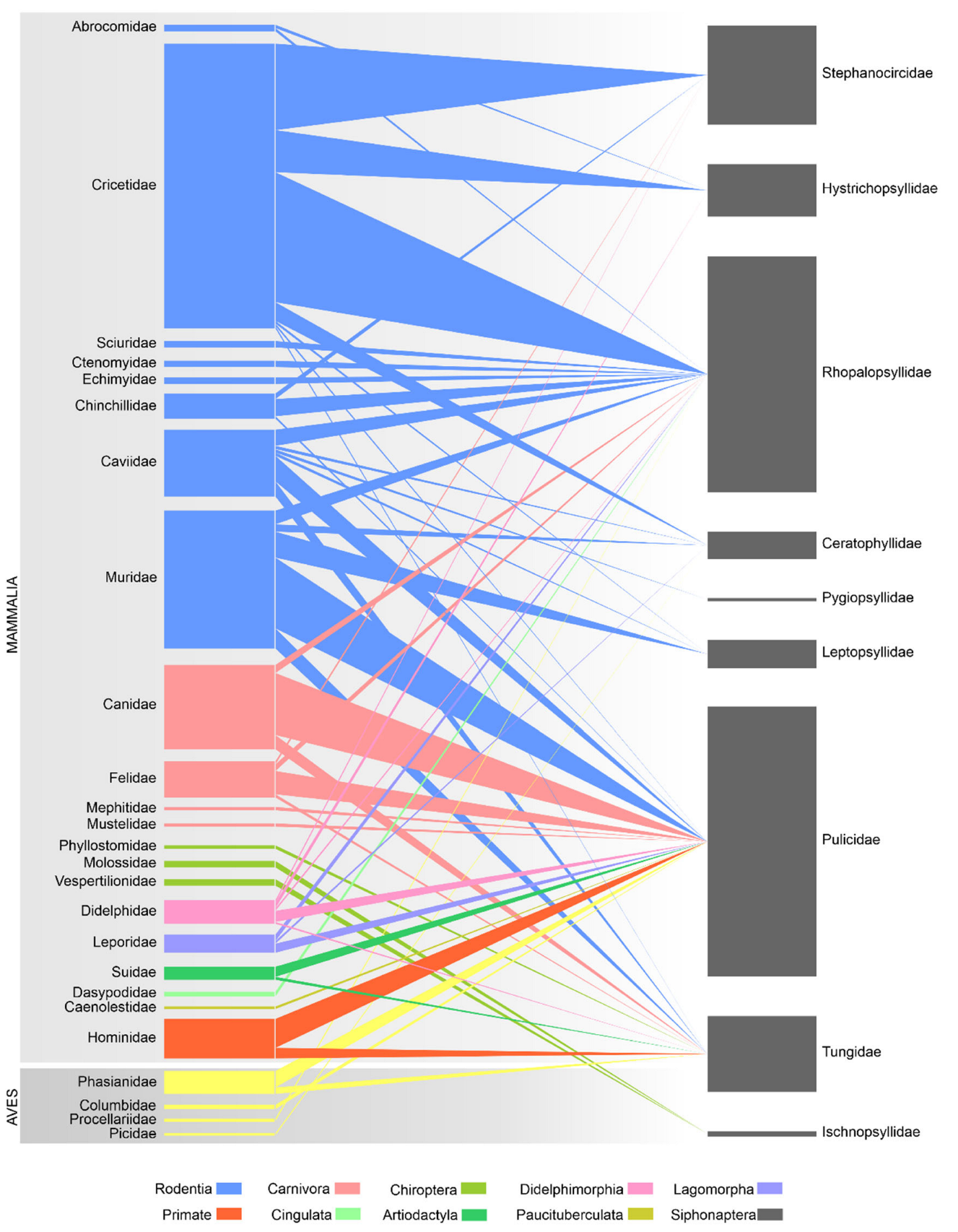
Figure 3.
A-B. Specimens of some representative fleas of the 85 species reported in Peru deposited in the entomological collection of the MUFV. A. Male of Plocopsylla achilles (Rothschild, 1911); B. Female of Sphinctopsylla inca (Rothschild, 1914); C. Male of Craneopsylla minerva minerva (Rothschild, 1903); D. Male of Cleopsylla townsendi Rothschild, 1914; E. Male of Neotyphloceras crassispina hemisus Jordan, 1936; F. Male of Plusaetis sp.
Figure 3.
A-B. Specimens of some representative fleas of the 85 species reported in Peru deposited in the entomological collection of the MUFV. A. Male of Plocopsylla achilles (Rothschild, 1911); B. Female of Sphinctopsylla inca (Rothschild, 1914); C. Male of Craneopsylla minerva minerva (Rothschild, 1903); D. Male of Cleopsylla townsendi Rothschild, 1914; E. Male of Neotyphloceras crassispina hemisus Jordan, 1936; F. Male of Plusaetis sp.
Figure 4.
A-B. Specimens of some fleas representative of the 85 species reported in Peru deposited in the MUFV entomological collection. A. Female of Polygenis (Polygenis) litargus; B. Male Ectinorus (Ectinorus) disjugis Jordan, 1942; C. Female of Tetrapsyllus (Phylliver) elutus Johnson, 1957; D. Female of Ctenocephalides felis felis (Bouche, 1835); E. Male of Pulex irritans Linnaeus, 1758; F. Female of Echidnophaga gallinacea (Westwood, 1875).
Figure 4.
A-B. Specimens of some fleas representative of the 85 species reported in Peru deposited in the MUFV entomological collection. A. Female of Polygenis (Polygenis) litargus; B. Male Ectinorus (Ectinorus) disjugis Jordan, 1942; C. Female of Tetrapsyllus (Phylliver) elutus Johnson, 1957; D. Female of Ctenocephalides felis felis (Bouche, 1835); E. Male of Pulex irritans Linnaeus, 1758; F. Female of Echidnophaga gallinacea (Westwood, 1875).

Figure 5.
(A) Modular structure of the national-scale interaction network between fleas and their hosts in the Peruvian territory and (B) the roles of the species in this structure. Interactions among species from different submodules in figure A are in gray..
Figure 5.
(A) Modular structure of the national-scale interaction network between fleas and their hosts in the Peruvian territory and (B) the roles of the species in this structure. Interactions among species from different submodules in figure A are in gray..
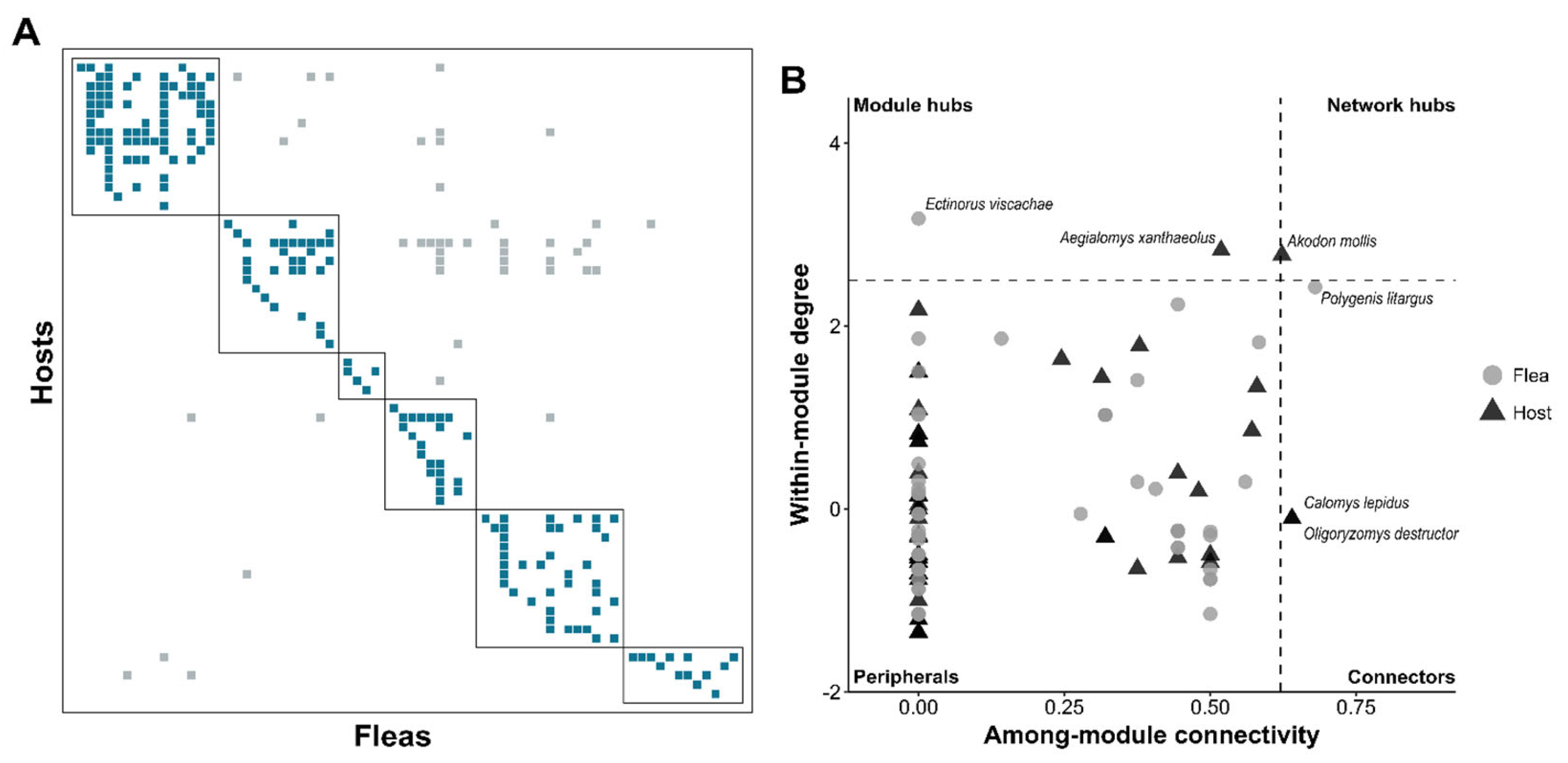

Table 1.
Structural properties of the national-scale interaction network between fleas and their hosts in the Peruvian territory. Metric include modularity (Q), nestedness for: the entire network (NODFtotal), pairs of species from the same module (NODFsm) and pairs of species from differente modules (NODFdm). Significance values (z-scores and p-values) are showed for both null models: free null model (free) and restricted null model (rest). Significant p-values are in bold.
Table 1.
Structural properties of the national-scale interaction network between fleas and their hosts in the Peruvian territory. Metric include modularity (Q), nestedness for: the entire network (NODFtotal), pairs of species from the same module (NODFsm) and pairs of species from differente modules (NODFdm). Significance values (z-scores and p-values) are showed for both null models: free null model (free) and restricted null model (rest). Significant p-values are in bold.
| Network metric |
Value | z-score (free) |
p-value (free) |
z-score (rest) |
p-value (rest) |
|---|---|---|---|---|---|
| Q | 0.57 | 9.52 | <0.001 | NA | NA |
| NODFtotal | 10.57 | -0.56 | 0.69 | 1.54 | 0.05 |
| NODFsm | 33.26 | 14.36 | <0.001 | 0.71 | 0.30 |
| NODFdm | 5.00 | -6.66 | <0.001 | 2.17 | <0.001 |
Table 2.
Normalized degree centrality and betweenness centrality of species at the entire national-scale interaction network between fleas and their hosts in the Peruvian territory. Only the 10 highest-ranked species in both metrics are displayed. Species present in both top 10 lists are in bold.
Table 2.
Normalized degree centrality and betweenness centrality of species at the entire national-scale interaction network between fleas and their hosts in the Peruvian territory. Only the 10 highest-ranked species in both metrics are displayed. Species present in both top 10 lists are in bold.
| Normalized degree centrality | Betweenness centrality | ||
|---|---|---|---|
| Species | Value | Species | Value |
| Akodon mollis | 0.254 | Akodon mollis | 0.200 |
| Polygenis litargus | 0.250 | Neotyphloceras crassispina | 0.153 |
| Pulex irritans | 0.203 | Rattus norvegicus | 0.128 |
| Rattus rattus | 0.194 | Phyllotis andium | 0.124 |
| Neotyphloceras crassispina | 0.188 | Polygenis litargus | 0.123 |
| Cleopsylla townsendi | 0.188 | Pulex irritans | 0.123 |
| Echidnophaga gallinacea | 0.188 | Aegialomys xanthaeolus | 0.107 |
| Rattus norvegicus | 0.164 | Cleopsylla townsendi | 0.099 |
| Phyllotis andium | 0.149 | Phyllotis amicus | 0.089 |
| Ctenocephalides canis | 0.141 | Tiamastus cavicola | 0.077 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



