Submitted:
03 November 2024
Posted:
05 November 2024
You are already at the latest version
Abstract
Background/Objectives: This study investigates the effectiveness of an herbal formulation, STRESSLESS (ST-65), which combines ashwagandha (Withania somnifera) and bacopa (Bacopa monnieri), on SH-SY5Y human neuroblastoma cells. Given the rising interest in natural compounds for neuroprotection and stress alleviation, we aimed to explore the cellular and molecular effects of this formulation. Methods: Utilizing a nuclear magnetic resonance (NMR) metabolomic approach and ultra-high-performance liquid chromatography-high-resolution mass spectrometry (UHPLC-HRMS), we identified key bioactive compounds in ST-65, including withanolides from ashwagandha and bacosides from bacopa. Results: Our findings indicate that ST-65 treatment significantly alters the metabolic profile of SH-SY5Y cells. Key changes included increased levels of metabolites linked to neuroprotection, energy metabolism, and antioxidant defense. Notable enhancements were observed in specific amino acids and neuroprotective compounds, suggesting activation of neuroprotective mechanisms and mitigation of stress-induced damage. Conclusions: The study reveals a complex phyto-chemical profile of ST-65 and underscores its potential as a natural active agent for addressing stress-related neurodegenerative conditions. These insights into neuronal mechanisms provide a foundation for further exploration of herbal formulations in neuroprotection.
Keywords:
Withania somnifera
; Bacopa monnieri
; stress
; SH-SY5Y cells
; UHPLC-HRMS/MS
; 1H NMR metabolomic
1. Introduction
Stress is a common experience that can have significant effects on both mental and physical health. Stress triggers the release of cortisol, a hormone produced by the adrenal glands. While cortisol helps regulate metabolism, immune response, and blood pressure, chronic stress can lead to prolonged high levels of cortisol, which may negatively affect physical and mental health [1]. Some effects of elevated cortisol include: increased anxiety and depression [2], weight gain [3], sleep issues [4], impaired immune function: chronic stress can weaken the immune response, making you more susceptible to illness [5]. To manage stress and cortisol levels, practices such as regular exercise, mindfulness meditation, adequate sleep and a balanced diet are increasingly considered[6]. Some natural compounds have been studied for their potential to help manage stress and promote relaxation, among them ashwagandha, an adaptogenic herb traditionally used in Ayurvedic medicine, has been reported in many studies for its ability to help the body manage stress, suggesting that it may help lower cortisol levels, reduce anxiety, and improve overall stress resilience [7]. Several clinical studies have investigated the effects of ashwagandha on stress, anxiety, and overall well-being. Ashwagandha may enhance sleep quality [8], which is often disrupted by stress and improve cognitive function and memory, which can be negatively impacted by stress [9]. A systematic review analysed human randomized controlled trials with a treatment arm that included Withania somnifera as a remedy for anxiety or stress and concluded that it may improve overall health and well-being, particularly in reducing stress and anxiety [10]. Guo & Rezaei [11]conducted a comprehensive review of the scientific literature focusing on the effects of ashwagandha supplementation, particularly its impact on antioxidant response and athletic performance. Their findings likely highlight how ashwagandha may enhance physical performance by reducing exercise-induced oxidative stress and improving recovery times. Withanolides are the biologically active compounds of Withania somnifera, a group of naturally occurring C28 steroids built on an ergostane skeleton functionalized at carbons 1, 22 and 26 [12]. Many studies have considered Bacopa monnieri L. to prevent or treat neurodegenerative conditions [13,14] and for its potential benefits in managing stress [15]. This plant is known as brahmi or “medhya rasayanas” in Ayurveda (meaning brain tonic). Bacopa has been shown to exert antioxidant effects, which may help protect neuronal cells from oxidative damage, a significant factor in neurodegenerative diseases like Alzheimer’s [16] and Parkinson’s [17]. A recent systematic review indicated that Bacopa can improve cognitive function, including memory, attention, and learning, which may be particularly beneficial for individuals at risk of neurodegenerative disorders [18]. Authors included clinical trials and preclinical studies that evaluated the efficacy of Bacopa in improving cognitive symptoms and overall brain health in populations at risk for or diagnosed with neurodegenerative conditions. The herb contains active, dammarane-type triterpenoidal saponins named bacosides, which are believed to promote synaptic communication, reduce inflammation, and enhance neurogenesis [19]. In our nutrition research laboratory (Nutriketo Lab) we have been involved in conducting dietary clinical trials to explores the ketogenic diet's applications in various medical contexts such as: metabolic disorders, examining its role in obesity management, insulin sensitivity [20], and type 2 diabetes [21], inflammation, as an important contributor to the pathogenesis of psoriasis [22,23], and oxidative stress involved in fibromyalgia a common chronic widespread pain disorder [24]. In our clinical experience, combining a ketogenic nutritional protocol with an Ashwagandha-Bacopa-based formulation offer a multifaceted approach to treating complex diseases and helping mitigate stress and anxiety that patients experience during restrictive dietary regimens. Ashwagandha-Bacopa supplementation improve adherence to the ketogenic diet and overall patient well-being. In this study, to confirm our clinical observations, we reported the impact on SH-SY5Y human neuroblastoma cells of a stress-reducing formulation consisting in 600 mg of ashwagandha and 50 mg of Bacopa in standardized extracts (named ST-65) using NMR metabolomic. We assessed the efficacy and safety of ST-65 providing valuable insights into metabolic alterations and potential therapeutic effects. We also developed a credible analytical method to identify the chemical components of ST-65. Eighteen chemical constituents, such as derivatives of flavonoids, withanolides bacosides and their derivatives, were identified or tentatively characterized.
2. Materials and Methods
2.1. Chemical and Standards
Analytical grade ethanol (EtOH), methanol (MeOH), chloroform, MS grade formic acid (HCOOH), and the reference standards of rutin, hesperidin, quercetin, were provided by Merck Chemicals (Milan, Italy). MS grade acetonitrile and water were purchased from Romil (Cambridge, UK). Ultrapure water (18 MΩ) was prepared using a Milli-Q purification system (Millipore, Bedford, TX, USA).
2.2. ST-65 formulation
STRESSLESS (ST-65) formulation consisted of the following extracts: KSM-66 ashwagandha (5% withanolides), a water-based extract of Withania somnifera (L.) Dunal roots made by Ixoreal Biomed of Los Angeles, California and a water-based extract of Bacopa monnieri (L.) Pennell aerial part (20% bacosides) kindly supplied by Laboratoarele Medica srl (Bucharest, Romania). The extracts were blended at Department of Pharmacy of University of Salerno in a 12 to 1 ratio accurately weigh 1200 mg of Ashwagandha E.S. and 100 mg of Bacopa using an analytical balance. The ingredients were combined and mixed in a mixing bowl thoroughly to ensure even distribution of the extracts.
2.3. UHPLC-HRMS/MS analysis
The samples were dissolved in a hydroalcoholic solution (70% v/v) with a matrix/ solvent ratio of 1:20. The analysis were performed using a Vanquish Flex UFPLC system interfaced to an Orbitrap Exploris 120 mass spectrometer, equipped with a heated electrospray ionization source, HESI-II (ThermoFisher Scientific, Milano, Italy). A Kinetex C18 column (100 x 2.1 mm I.D., 2.6 µm; Phenomex, Bologna, Italy) was used a flow rate of 500 µL min-1 at 30 °C. Volume of the injection was 5 µL. The mobile phase was a binary gradient of water (A) an MeCN (B), both containing 0.1%, v/v, formic acid. The gradient elution program is as follows: 0-3 min, 2 % B; 3-5 min, 2-13 % B; 5-9 min, 13 % B; 9-12 min, 13-18 % B; 12-13 min, 18 % B; 13-17 min, 18-30 % B; 17-20 min, 30 % B; 20-30 min, 30-40 % B; 30-38 min, 40-60 %B; 38-39 min, 60-98 % B; after each injection (5 µL), washing (98 % B, 4 min) and re-equilibration of the column (2 % B, 5 min) were performed. The mass spectrometer was operated in negative and positive mode. A Full MS data dependent MS/MS (Full MS/dd-MS2) acquisition mode was used. Full MS scan resolution (scan range m/z 100-1000), 60000 FWHM; dd-MS2 scan set, 30000 FWHM. A stepped collision energy HCD (20, 40 and 60) was applied for fragmentation. Detected compounds were identified to the corresponding spectral characteristics: mass spectra, accurate mass; fragmentation pathways and retention time. Xalibur software (version 4.6) was used for instrument control, data acquisition and data analysis.
2.4. Cell Culture
The human neuroblastoma cell line SH-SY5Y was obtained from American Type Culture Collection (ATCC, Rockville, MD, USA). Cells were cultured in Dulbecco’s Modified Eagle Medium (DMEM, 4500 mg/mL glucose) supplemented with 10% (v/v) fetal bovine serum, 2 mM L-glutamine, 100 U/mL penicillin, and 0.1 mg/mL streptomycin. Cells were grown in 100 mm culture dishes (Corning, Corning, NY, USA) in a humidified environment containing 5% CO2 at 37 °C and split every 2 days.
2.5. Cell Viability Assay
Cell viability was established by measuring mitochondrial metabolic activity to reduce the tetrazolium salt WST-8 (2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium, monosodic salt) through Cell Counting Kit-8 (CCK-8 Cat. CK04, Dojindo Laboratories, Rockville, MD, USA). Briefly, SH-SY5Y (8 × 103 cells/well) was plated into 96-well plates for 24 h, and then ST -65 in the range of 100-0.80 µg/mL was added for an additional 24 h. At the end of the treatment, CCK-8 solution (10 μL) was added to the cell media and incubated for 1 h in a humidified incubator. Absorbance was detected at a wavelength of 450 nm. Results are expressed as means ±SD of 3 independent experiments performed in triplicate and reported as the percentage of viable cells vs the untreated control.
2.6. Exposure of SH-SY5Y to ST-65
Cells were pleated in 100 mm culture dishes and allowed to adhere overnight. Afterward, ST-65 (25 'g/ml) was added and incubated for 24 h. For the control group, cells were treated only with a vehicle. At the end of treatments, cell medium was collected, and the dishes were washed with cold PBS (pH 7.4) to remove media residues just before the metabolite extraction procedure.
2.7. Sample Extraction
The culture medium was collected from each plate in microcentrifuge tubes and centrifuged at 1000× g for 5 min. For extracting cellular metabolites, a dual-phase extraction (methanol:chloroform:water; 1:1:1) was used after cell collection by scraping and homogenization [25]). The two phases were separated after centrifugation at 6000 rpm for 10 min at 4 °C. The resulting polar extracts were dried under vacuum in an SP-Genevac EZ-2 4.0 concentrator, and the lipophilic extracts were dried under a nitrogen flow for future analysis. Both growth media and cell extracts were stored at −80 °C before NMR analysis.
2.8. NMR spectra acquisition
Bruker Ascend™ 600 MHz spectrometer was used to acquire the spectra. The spectrometer was equipped with a 5 mm triple resonance Z gradient TXI probe (Bruker Co, Rheinstetten, Germany) at 298 K. TopSpin, version 3.2 was used for the spectrometer control and data processing (Bruker Biospin). All the experiments performed as Nuclear Overhauser Enhancement Spectroscopy (NOESY) 1D were acquired in triplicate. Spectra acquisition was made using 12 ppm spectral width, 20k data points, presaturation during relaxation delay and mixing time for water suppression [26] and spoil gradient, 5 s relaxation delay, and mixing time of 10 ms. A weighted Fourier transform was applied to the time domain data with a line widening of 0.5 Hz followed by a manual step and baseline correction in preparation for targeted profiling analysis.
2.9. Statistical analysis
NMR spectra of SH-SY-5Ycells cultures eso and endometabolome were analysed using an untargeted metabolomic approach. All the spectra were assigned using Chenomx NMR-Suite v8.0 (Chenomx Inc., Edmonton, 252 Canada) and quantified by NMRProcFlow as previously reported [27]. The quantification matrices reported the metabolites identified and quantified in the eso and endometabolome of SH-SY-5Y treated and untreated with ST-65 were analyzed using the open-source tool Metaboanalyst 6.0 [28]. The Volcano plot combined T-test and Fold Change performed the univariate approach [29,30]. After normalization by sum log and Pareto, we applied a supervised multivariate approach partial least-squares discriminant analysis (PLS-DA) method. The reliability of the supervised model was analyzed using a cross-validation approach, considering the accuracy and parameters Q2 and R2. The metabolites responsible for clusters’ separation in the PLS-DA score plot were classified according to VIP, considering only the metabolites with VIP>1 [28]. Enrichment Pathways tools were applied to identify the dysregulated biochemical pathways by KEGG database. Only the KEGG paths that reported a rate of false discoveries (FDR) lower than 1, the p-value lower than 0.05 and the hits value related to the number of metabolites belonging to the pathway >1, were chosen. [30].
3. Results
3.1. UHPLC-HRMS/MS analysis
In the present study, an UHPLC-HRMS/MS method for profiling and characterization of constituents present in the ST-65 formulation was developed. The UHPLC conditions were optimized to obtain maximal chromatographic resolution and MS signal. Figure 1 shows representative chromatogram of ST-65 under optimal conditions. HRMS/MS analysis were performed in positive and negative ionization mode to obtain complementary information needed to characterize ST-65 formulation. To avoid repetitions, data have been reported only in positive ionization mode (Table 1) as they show higher response in the detection. Compound assignments were made by HRMS spectra, tandem mass spectrometry experiments and when possible, by comparing MS spectra and retention time with authentic standards, or with chemo-taxonomic data reported in literature. Eighteen (1-18) major peaks were detected in ST-65 formulation, and their retention times and MS data are listed in Table 1. UHPLC-HRMS/MS analysis allowed to identify four flavonoids (3-6), ten withanolides (7-1) and one bacopa saponin (17). Compounds 3, 5 and 6 were ascertained to be rutin, hesperidin and quercetin, respectively, by reference standards. Compound 4 in MS2 spectra showed a base peak at m/z 287.0548 corresponding to the aglycon kaempferol and the characteristic losses of hexose and deoxyhexose residues. Thus, compound 4 could be tentatively identified as kaempferol-hexose-deoxyhexose. Compounds 7-16 were identified as withanolides, 28-carbon steroidal lactones, built up on an intact or rearranged ergostane framework [31].Their structural chemistry produce limited fragmentation in ESI mass spectra. Therefore, in MS2 spectra, only losses of the sugar units linked to the aglycone were detected. Thus, according MS analysis and literature data [31] the compounds 7-15 could be identified as withanolides reported in Table 1. Compound 17 showed ion at m/z 797.4688 with a molecular formula C42H69O14. It was identified as a Bacopaside N, as the MS2 spectra showed losses of two sugar units from the jujubogenin aglycon [32].
3.2. ST-65 viability
To study the biological effect of ST-65, we preliminary evaluated the extract's viability effect on SH-SY5Y neuroblastoma cells. Figure 2 clearly shows that the ST-65 extract did not significantly affect the viability of cells after 24 hours of treatment, even at the highest concentrations.
3.2. NMR Metabolomic
The cytoplasmatic metabolites, produced by cells (endometabolome) and the extracytoplasmatic metabolites (esometabolome) were investigated using 1H-NMR spectroscopy. We performed an untargeted metabolomic analysis on SH-SY-5Y cell cultures treated and untreated with the Bacopa and Ashwagandha formulations called ST-65. Figure 3A, Figure 2B show the representative 1D 1H NOESY NMR spectrum of cellular endo and esometabolome, respectively [33],. The spectra were assigned using Chenomx NMR-Suite v8.0 [34], highlighting 41 metabolites in the endometabolome and 36 in the esometabolome. NMRProcFlow was used to quantify the spectra, taking into account the internal standard trimethylsilyl propanoic acid (TSP), the number of H for each peak, and the metabolites’ molecular weight [27]. After quantification, metabolites matrices were analysed using MetaboAnalyst 6.0 by univariate and multivariate supervised approaches.
The dataset was first normalised by Log and Parate scaling to perform Partial Least Square determination analysis (PLS-DA) [35]. The score plots in Figure 4A,B showed a different metabolomic profile between the endo metabolome and the esometabolome of treated and untreated cells. The score plot was described in the Cartesian space by the main components: PC1: 68.6% and PC2:17.3% for endometabolome. On the other hand, the esometabolome score plot was described as PC1: 65.2% and PC2:30.1%. The clusters’ separation was confirmed by cross-validation. (Table S1) [36]. The metabolites responsible for clusters’ separation were identified according to the Variable Importance Projections (VIP) score analysis [37]. Accordingly, the metabolites with a VIP score > 1.00 are considered good classifiers between the clusters. (Figure 4C e 4D). VIP analysis showed a dysregulation of several energetic metabolites: in particular, cells treated with ST-65 produced lower concentrations of Formate, Acetate, 2-oxobutyrate, and Carnitine. The analysis identified the upregulation of several metabolites related to mitochondria, such as ADP, AMP, and Coenzyme A. (Figure 4B) ST-65 modulated aminoacidic metabolism; in particular, we detected a downregulated production of cysteine and aspartate. Conversely, the endometabolome aminoacidic concentration of proline, tyrosine, leucine and glutamate was higher in the cells treated with ST-65 and lower in the untreated cells. Moreover, VIP score analysis detected higher concentrations of tryptophan, tyrosine and valine in the esometabolome of treated cells and lower of arginine and carnitine (Figure 4D). The volcano plot confirmed the results of the VIP score analysis. Additionally, it showed a higher concentration of phenylalanine and a lower concentration of serine and glucose in the endomebolome of SH-SY-5Y treated cells (Figure 4E). Enrichment pathways analysis was performed to identify the dysregulated pathways according to the metabolites ranked by VIP score analysis.
The paths with hits>1 and p.value< 0.05 were considered discriminatory [38] (Figure 5; Table S2). Enrichment revealed that ST-65 impacts several energetic pathways, such as Pyruvate metabolism and the Citric acid Cycle. Furthermore, mitochondrial biochemical pathways related to fatty acids, such as beta-oxidation of short-chain fatty acids in mitochondria, beta-oxidation of long-chain fatty acids, and ketosis body metabolism, are also impacted. Moreover, ST-65 also modulates pathways related to hormone biosynthesis and neurotransmitter amino acids, such as glycine and serine metabolism.
4. Discussion
The combination of ashwagandha (Withania somnifera) and bacopa (Bacopa monnieri) has garnered attention in clinical settings due to their individual and potentially synergistic therapeutic effects. Both herbs have a long history of use in traditional medicine, and emerging clinical evidence supports their benefits for stress management, cognitive enhancement, and overall health and their combination may offer a synergistic effect due to their complementary pharmacological actions [7,8,9,10]. While ashwagandha is known for its adaptogenic properties, bacopa is recognized for cognitive enhancement. The optimization of UHPLC conditions achieved maximal chromatographic resolution and MS signal was crucial for accurately identify the bioactive compounds present in the in the ashwagandha-bacopa formulation. Withanolides are known for their adaptogenic properties, and their presence may explain some of the formulation's effects on stress and cognitive function, this compounds may help alleviate these conditions mainly by modulating the hypothalamic-pituitary-adrenal and sympathetic-adrenal medullary axes, as well as through the GABAergic and serotonergic pathways [10]. The detection of a bacopa saponin (peak 17) is another critical finding. Bacopa monnieri is traditionally used for enhancing cognitive function, and its saponins are believed to be responsible for its neuroprotective effects, the neuroprotective activity of bacosides is linked to their role in regulating mRNA translation and the surface expression of neuroreceptors, including AMPAR, NMDAR, and GABAR, in different regions of the brain. [16,19]. The metabolomic approach performed on cell cultures allowed us to identify a different metabolomic profile of ST-65 treated cells compared with untreated cells, showing a clear clustering of the two groups following a multivariate PLS-DA approach (Figure 4A). SH-SY5Y cells were treated with ST-65 to evaluate its action, mainly on neuronal energy. The metabolomics analysis validated this hypothesis and revealed further modulated mechanisms. SH-SY5Y cells treated with ST-65 showed modulation of aminoacidic pathways involved in neurotransmission. In detail, VIP analysis highlighted an increase in the concentrations of glutamate and a decrease in the intracellular concentrations of aspartate. At the same time, a lower tryptophan use was identified, accompanied by an increased production and expulsion of tyrosine (Figure 4C, 4D).These results suggest that ST-65 could influence the metabolism of aminoacids participating in neurotransmission and neuroplasticity processes [39]. Glutamate is also essential as it participates in cellular energy [40] and is the most abundant free amino acid in the brain [41]. The enrichment analysis identified among the most modulated pathways in cells treated with ST-65 pyruvate metabolism and citric acid cycle. (Figure 5) The increase in glutamate induced by treatment with ST-65 participates in the upregulation of the citric acid cycle. It also leads to a greater use of ATP, as demonstrated by the increase in its AMP and ADP products. The variations of ADP suggest that ST-65 could modulate neuronal energy positively. It is known that ADP stimulates respiration through oxidation processes, which in turn provide energy [42]. (Maldonado & Lemasters, 2014). The components of ST-65, especially Ashwagandha, are known to improve the production of sex hormones [43]. Our results confirmed this action. The treatment with ST-65 impacted the biosynthesis of steroids and sex hormones, as demonstrated by the enrichment analysis (Figure 5), which identified among the most dysregulated pathways: steroid biosynthesis and androgen and estrogen metabolism. It is known that the synthesis of sex hormones is closely related to cholesterol pathways [44]. Our study has shown a decrease in 2-oxobutyrate and intracellular formate (Figure 4C) in cells treated with ST-65. Formate is produced by the synthesis of cholesterol and by the degradation of folate deficiency [45]. A reduction proves an improvement of these biochemical pathways, indicating that ST-65 could also have a lipid-lowering action. Pathway analysis supports this hypothesis by identifying biochemical pathways related to the oxidation of short- and long-chain fatty acids among the dysregulated pathways. (Figure 5) As is known, the processes of beta-oxidation of fatty acids occur at the mitochondrial level. Our study identified a dysregulation of the biochemical pathways of mitochondrial oxidation in ST-65 -treated cells and a consequent increase of Coenzyme A concentration (Figure 3C), a metabolite essential for cellular bioenergetics, especially for processes related to lipid metabolism [46]. Metabolomics analysis also identified an increase in cell uptake and metabolism of carnitine by ST-65-treated cells (Figure 4 C,D). Carnitine is a central metabolite in the peroxidation process of fatty acids and in the biochemical processes of free radical scavengers [47]. In conclusion, the metabolomics analysis showed that ST-65 affects energy metabolism, particularly having a hypolipidemizing and hypoglycemic action. It also assists the synthesis of sex hormones and improves neuronal transmission and mitochondrial function. Further investigations are warranted to elucidate the specific pathways involved and to evaluate its clinical applicability.
5. Conclusions
The ashwagandha-bacopa formulation (ST-65 ) presents a compelling option for individuals seeking natural support for stress and cognitive function. By leveraging the unique properties of each herb, this formulation could provide a balanced approach to mental health. Continued clinical research will be essential in establishing its efficacy and safety, ultimately guiding its use in integrative health practices.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table 1. Cross-validation performed with 10-fold methods reported the accuracy, R2 and Q2 value of three components.; Table 2. Metabolic pathways corresponding to serum and urinary metabolomic profile of intervention group. The pathways are classified according to p-values, Holm adjustment, and FDR values. The pathways are selected based on Hits > 2, Raw p < 0.05, and Holm adjust, FDR > 1..
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, A.D., M.D.E., M.R., and L.R.; methodology, C.M., R.C., E.N. and M.D.E..; software, C.M., R.C., E.N; validation, A.D., L.R., M.R., and M.D.E.; formal analysis, R.C. E.N. and C.M.; investigation, C.M., R.C., E.N. and M.D.E.; resources, L.R. data curation, L.R., M.R. M.D.E. and A.D.; writing—original draft preparation M.D.E., L.R.; supervision, A.D., M.R., L.R.; project administration L.R.; funding acquisition, L.R. All authors have read and agreed to the published version of the manuscript.” Please turn to the CRediT taxonomy for the term explanation. Authorship must be limited to those who have contributed substantially to the work reported.
Funding
Project funded under the National Recovery and Resilience Plan (NRRP), Mission 4 Component 2 Investment 1.4 - Call for tender No. 3138 of December 16, 2021, rectified by Decree n.3175 of December 18, 2021 of Italian Ministry of University and Research funded by the European Union – NextGenerationEU; Award Number: Project code CN_00000033, Concession Decree No. 1034 of June 17, 2022 adopted by the Italian Ministry of University and Research, CUP: D43C22001260001, Project title “National Biodiversity Future Center - NBFC”.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to thank the company Laboratoarele Medica srl (Bucharest, Romania) who kindly donated the Withania somnifera and Bacopa monnieri extracts
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Knezevic, E.; Nenic, K.; Milanovic, V.; Knezevic, N.N. The role of cortisol in chronic stress, neurodegenerative diseases, and psychological disorders. Cells 2023, 12, 2726. [Google Scholar] [CrossRef] [PubMed]
- Chan, I.I.; Wu, A.M. Assessing the Role of Cortisol in Anxiety, Major Depression, and Neuroticism: A Mendelian Randomization Study Using SERPINA6/SERPINA1 Variants. Biological Psychiatry Global Open ScienceI 2024, 4, 100294. [Google Scholar] [CrossRef] [PubMed]
- Kuckuck, S.; van Der Valk, E.S.; Scheurink, A.J.; van Der Voorn, B.; Iyer, A.M.; Visser, J.A.; van Rossum, E.F.; et al. Glucocorticoids, stress and eating: The mediating role of appetite-regulating hormones. Obesity Reviews 2023, 24, e13539. [Google Scholar] [CrossRef] [PubMed]
- Passos, G.S.; Youngstedt, S.D.; Rozales, A.A.R.C.; Ferreira, W.S.; De-Assis, D.E.; De-Assis, B.P.; Santana, M.G. Insomnia severity is associated with morning cortisol and psychological health. Sleep Science 2023, 16, 092–096. [Google Scholar]
- Luo, J.; Zhou, C.; Wang, S.; Tao, S.; Liao, Y.; Shi, Z.; Yang, P.; et al. Cortisol synergizing with endoplasmic reticulum stress induces regulatory T-cell dysfunction. Immunology 2023, 170, 334–343. [Google Scholar] [CrossRef]
- Kip, E.; Parr-Brownlie, L.C. Healthy lifestyles and wellbeing reduce neuroinflammation and prevent neurodegenerative and psychiatric disorders. Frontiers in Neuroscience 2023, 17, 1092537. [Google Scholar] [CrossRef]
- Mikulska, P.; Malinowska, M.; Ignacyk, M.; Szustowski, P.; Nowak, J.; Pesta, K.; Cielecka-Piontek, J.; et al. Ashwagandha (Withania somnifera)—Current research on the health-promoting activities: A narrative review. Pharmaceutics 2023, 15, 1057. [Google Scholar] [CrossRef]
- Cheah, K.L.; Yaacob, L.H.; Rahman, R.A. Effect of Ashwagandha (Withania somnifera) extract on sleep: A systematic review and meta-analysis. PLoS ONE 2021, 16, e0257843. [Google Scholar] [CrossRef]
- Leonard, M.; Dickerson, B.; Estes, L.; Gonzalez, D.E.; Jenkins, V.; Johnson, S.; Kreider, R.B.; et al. Acute and Repeated Ashwagandha Supplementation Improves Markers of Cognitive Function and Mood. Nutrients 2024, 16, 1813. [Google Scholar] [CrossRef]
- Speers, A.B.; Cabey, K.A.; Soumyanath, A.; Wright, K.M. Effects of Withania somnifera (Ashwagandha) on stress and the stress-related neuropsychiatric disorders anxiety, depression, and insomnia. Current neuropharmacology 2021, 19, 1468. [Google Scholar] [CrossRef]
- Guo, S.; Rezaei, M.J. The benefits of ashwagandha (Withania somnifera) supplements on brain function and sports performance. Frontiers in Nutrition 2024, 11, 1439294. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, A.K.; Nagegowda, D.A. Biosynthesis of the triterpenoid withanolides in Withania somnifera. Current Opinion in Plant Biology 2024, 81, 102576. [Google Scholar] [CrossRef] [PubMed]
- Brimson, J.M.; Brimson, S.; Prasanth, M.I.; Thitilertdecha, P.; Malar, D.S.; Tencomnao, T. The effectiveness of Bacopa monnieri (Linn.) Wettst. as a nootropic, neuroprotective, or antidepressant supplement: Analysis of the available clinical data. Scientific reports 2021, 11, 596. [Google Scholar] [CrossRef]
- Sushma Sahu, M.R.; Murugan, N.A.; Mondal, A.C. Amelioration of Amyloid-β Induced Alzheimer's Disease by Bacopa monnieri through Modulation of Mitochondrial Dysfunction and GSK-3β/Wnt/β-Catenin Signaling. Molecular Nutrition & Food Research 2024, 68, 2300245. [Google Scholar]
- Lopresti, A.L.; Smith, S.J.; Ali, S.; Metse, A.P.; Kalns, J.; Drummond, P.D. Effects of a Bacopa monnieri extract (Bacognize®) on stress, fatigue, quality of life and sleep in adults with self-reported poor sleep: A randomised, double-blind, placebo-controlled study. Journal of Functional Foods 2024, 85, 104671. [Google Scholar] [CrossRef]
- Palollathil, A.; Najar, M.A.; Amrutha, S.; Pervaje, R.; Modi, P.K.; Prasad, T.S.K. Bacopa monnieri confers neuroprotection by influencing signaling pathways associated with interleukin 4, 13 and extracellular matrix organization in Alzheimer's disease: A proteomics-based perspective. Neurochemistry International 2024, 105864. [Google Scholar] [CrossRef]
- Gupta, V.; Prasad, S. Differential Alterations in the Expression of AMPA Receptor and Its Trafficking Proteins in the Hippocampus Are Associated with Recognition Memory Impairment in the Rotenone-Parkinson’s Disease Mouse Model: Neuroprotective Role of Bacopa monnieri Extract CDRI 08. Molecular neurobiology 2024, 1–19. [Google Scholar]
- Valotto Neto, L.J.; Reverete de Araujo, M.; Moretti Junior, R.C.; Mendes Machado, N.; Joshi, R.K.; dos Santos Buglio, D.; Barbalho, S.M.; et al. Investigating the Neuroprotective and Cognitive-Enhancing Effects of Bacopa monnieri: A Systematic Review Focused on Inflammation, Oxidative Stress, Mitochondrial Dysfunction, and Apoptosis. Antioxidants 2024, 13, 393. [Google Scholar] [CrossRef]
- Bhandari, P.; Sendri, N.; Devidas, S.B. Dammarane triterpenoid glycosides in Bacopa monnieri: A review on chemical diversity and bioactivity. Phytochemistry 2020, 172, 112276. [Google Scholar] [CrossRef]
- Castaldo, G.; Schiavo, L.; Pagano, I.; Molettieri, P.; Conte, A.; Sarno, G.; Rastrelli, L.; et al. Clinical impact of enteral protein nutritional therapy on patients with obesity scheduled for bariatric surgery: A focus on safety, efficacy, and pathophysiological changes. Nutrients 2023, 15, 1492. [Google Scholar] [CrossRef]
- Pellegrini, P.; Lemasson, P.; Rastrelli, L.; D’Elia, M. Effectiveness of ketogenic therapy in patients with obesity and diabetes: A narrative review. Exploration of Foods and Foodomics 2024, 2, 313–325. [Google Scholar] [CrossRef]
- Castaldo, G.; Rastrelli, L.; Galdo, G.; Molettieri, P.; Aufiero, F.R.; Cereda, E. Aggressive weight-loss program with a ketogenic induction phase for the treatment of chronic plaque psoriasis: A proof-of-concept, single-arm, open-label clinical trial. Nutrition 2020, 74, 110757. [Google Scholar] [CrossRef] [PubMed]
- Castaldo, G.; Pagano, I.; Grimaldi, M.; Marino, C.; Molettieri, P.; Santoro, A.; Rastrelli, L.; et al. Effect of very-low-calorie ketogenic diet on psoriasis patients: A nuclear magnetic resonance-based metabolomic study. Journal of proteome research 2020, 20, 1509–1521. [Google Scholar] [CrossRef]
- Castaldo, G.; Marino, C.; Atteno, M.; D’Elia, M.; Pagano, I.; Grimaldi, M.; Rastrelli, L.; et al. Investigating the Effectiveness of a Carb-Free Oloproteic Diet in Fibromyalgia Treatment. Nutrients 2024, 16, 1620. [Google Scholar] [CrossRef] [PubMed]
- Beckonert, O.; Keun, H.C.; Ebbels, T.M.D.; Bundy, J.; Holmes, E.; Lindon, J.C.; Nicholson, J.K. Metabolic profiling, metabolomic and metabonomic procedures for NMR spectroscopy of urine, plasma, serum and tissue extracts. Nature protocols 2007, 2, 2692. [Google Scholar] [CrossRef]
- Mckay, R.T. How the 1D-NOESY suppresses solvent signal in metabonomics NMR spectroscopy: An examination of the pulse sequence components and evolution. Concepts in Magnetic Resonance Part A 2011, 38, 197–220. [Google Scholar] [CrossRef]
- Jacob, D.; Deborde, C.; Lefebvre, M.; Maucourt, M.; Moing, A. NMRProcFlow: A graphical and interactive tool dedicated to 1D spectra processing for NMR-based metabolomics. Metabolomics 2017, 13, 1–5. [Google Scholar] [CrossRef]
- Xia, J.; Wishart, D.S. Using MetaboAnalyst 3.0 for comprehensive metabolomics data analysis. Current protocols in bioinformatics 2016, 55, 14.10. 1–14.10. 91. [Google Scholar] [CrossRef]
- Kumar, N.; Hoque, M.A.; Sugimoto, M. Robust volcano plot: Identification of differential metabolites in the presence of outliers. BMC bioinformatics 2018, 19, 1–11. [Google Scholar] [CrossRef]
- Pang, Z.; Lu, Y.; Zhou, G.; Hui, F.; Xu, L.; Viau, C.; Xia, J.; et al. MetaboAnalyst 6.0: Towards a unified platform for metabolomics data processing, analysis and interpretation. Nucleic Acids Research 2024, gkae253. [Google Scholar] [CrossRef]
- Abdelwahed, M.T.; Hegazy, M.A.; Mohamed, E.H. Major biochemical constituents of Withania somnifera (ashwagandha) extract: A review of chemical analysis. Reviews in Analytical Chemistry 2023, 42, 20220055. [Google Scholar] [CrossRef]
- Nuengchamnong, N.; Sookying, S.; Ingkaninan, K. LC-ESI-QTOF-MS based screening and identification of isomeric jujubogenin and pseudojujubogenin aglycones in Bacopa monnieri extract. Journal of pharmaceutical and biomedical analysis 2016, 129, 121–134. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.C.; Wang, S.Y.; Kuo, C.H.; Tseng, Y.J. Distribution-based classification method for baseline correction of metabolomic 1D proton nuclear magnetic resonance spectra. Analytical Chemistry 2013, 85, 1231–1239. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.Y.; Wu, H.; Tjeerdema, R.S.; Viant, M.R.J.M. Evaluation of metabolite extraction strategies from tissue samples using NMR metabolomics. Metabolomics 2007, 3, 55–67. [Google Scholar] [CrossRef]
- Worley, B.; Halouska, S.; Powers, R. Utilities for quantifying separation in PCA/PLS-DA scores plots. Analytical biochemistry 2013, 433, 102–104. [Google Scholar] [CrossRef]
- Westerhuis, J.A.; Hoefsloot, H.C.; Smit, S.; Vis, D.J.; Smilde, A.K.; van Velzen, E.J.; van Dorsten, F.A.J.M.; et al. Assessment of PLSDA cross validation. Metabolomics 2008, 4, 81–89. [Google Scholar] [CrossRef]
- Akarachantachote, N.; Chadcham, S.; Saithanu, K. Cutoff threshold of variable importance in projection for variable selection. Int J Pure Appl Math 2014, 94, 307–322. [Google Scholar] [CrossRef]
- Zhang, B.; Hu, S.; Baskin, E.; Patt, A.; Siddiqui, J.K.; Mathé, E.A. RaMP: A comprehensive relational database of metabolomics pathways for pathway enrichment analysis of genes and metabolites. Metabolites 2018, 8, 16. [Google Scholar] [CrossRef]
- Aquili, L. The Role of Tryptophan and Tyrosine in Executive Function and Reward Processing. Int J Tryptophan Res 2020, 13, 1178646920964825. [Google Scholar] [CrossRef]
- Fendt, S.M.; Verstreken, P. Neurons eat glutamate to stay alive. J Cell Biol 2017, 216, 863–865. [Google Scholar] [CrossRef]
- Zhou, Y.; Danbolt, N.C. Glutamate as a neurotransmitter in the healthy brain. J Neural Transm (Vienna) 2014, 121, 799. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, E.N.; Lemasters, J.J. ATP/ADP ratio, the missed connection between mitochondria and the Warburg effect. Mitochondrion 2014, 19 Pt A, 78–84. [Google Scholar] [CrossRef]
- Gómez Afonso, A.; Fernandez-Lazaro, D.; Adams, D.P.; Monserdà-Vilaró, A.; Fernandez-Lazaro, C.I. Effects of Withania somnifera (Ashwagandha) on Hematological and Biochemical Markers, Hormonal Behavior, and Oxidant Response in Healthy Adults: A Systematic Review. Current Nutrition Reports 2023, 12, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.Y.; Choi, M.H.; Kim, J. Metabolic profiling of cholesterol and sex steroid hormones to monitor urological diseases. Endocr Relat Cancer 2016, 23, R455–R467. [Google Scholar] [CrossRef] [PubMed]
- Lamarre, S.G.; Morrow, G.; Macmillan, L.; Brosnan, M.E.; Brosnan, J.T. Formate: An essential metabolite, a biomarker, or more? 2013, 51, 571–578. [Google Scholar] [CrossRef]
- Leonardi, R.; Zhang, Y.M.; Rock, C.O.; Jackowski, S. Coenzyme A: Back in action. Progress in lipid research 2005, 44, 125–153. [Google Scholar] [CrossRef]
- Kalaiselvi, T.; Panneerselvam, C. Effect of L-carnitine on the status of lipid peroxidation and antioxidants in aging rats. The Journal of Nutritional Biochemistry 1998, 9, 575–581. [Google Scholar] [CrossRef]
Figure 1.
UHPLC-HRMS profile of ST-65 Formulation.
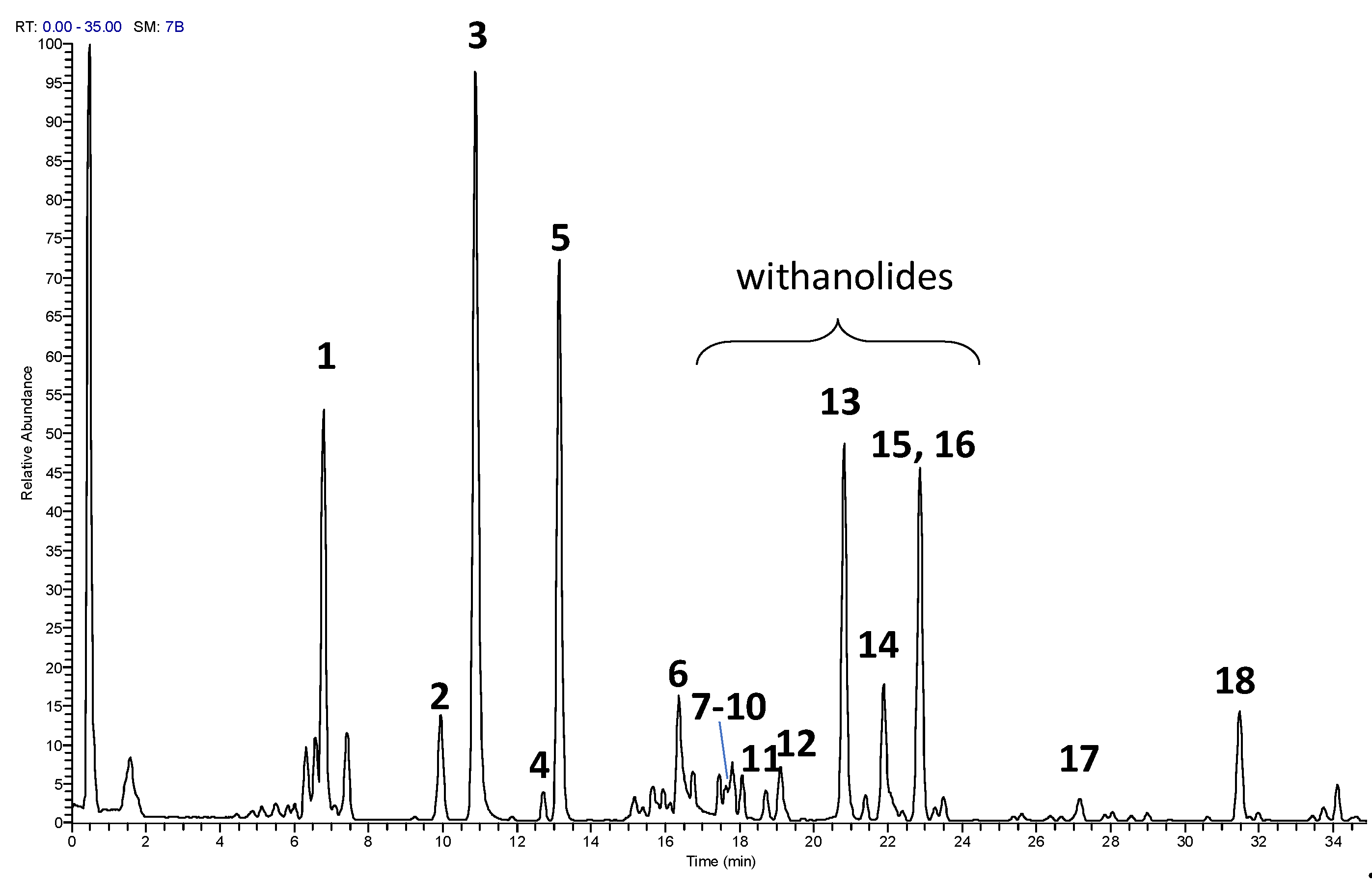
Figure 2.
Histogram graph showing the percentage of cell viability 24 h after the exposition of different concentrations of ST-65 extract (0.80-100 µg/mL). The viability variations were calculated as the percentage of viable cells in treated cultures compared to untreated ones. Results are shown as mean ± standard deviation (SD) from three independent experiments.
Figure 2.
Histogram graph showing the percentage of cell viability 24 h after the exposition of different concentrations of ST-65 extract (0.80-100 µg/mL). The viability variations were calculated as the percentage of viable cells in treated cultures compared to untreated ones. Results are shown as mean ± standard deviation (SD) from three independent experiments.
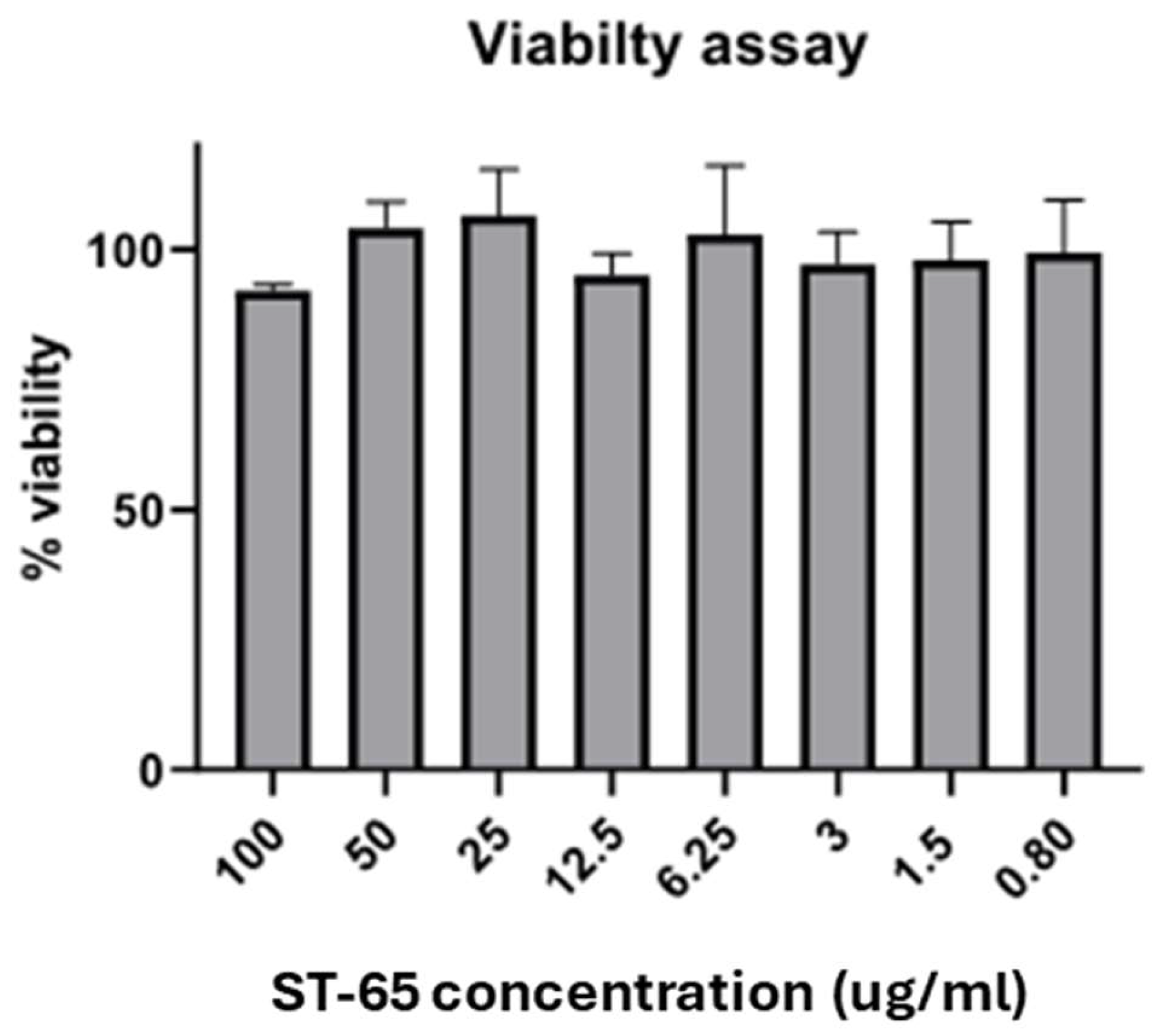
Figure 3.
1D shows the NOESY spectrum of the endometabolome (A) and esometabolome (B) of SH-SY-5Y polar extracts. The spectrum was acquired, as mentioned before. Forty-one and Thirty-six metabolites were detected in the endometabolome and esometabolome, respectively.
Figure 3.
1D shows the NOESY spectrum of the endometabolome (A) and esometabolome (B) of SH-SY-5Y polar extracts. The spectrum was acquired, as mentioned before. Forty-one and Thirty-six metabolites were detected in the endometabolome and esometabolome, respectively.
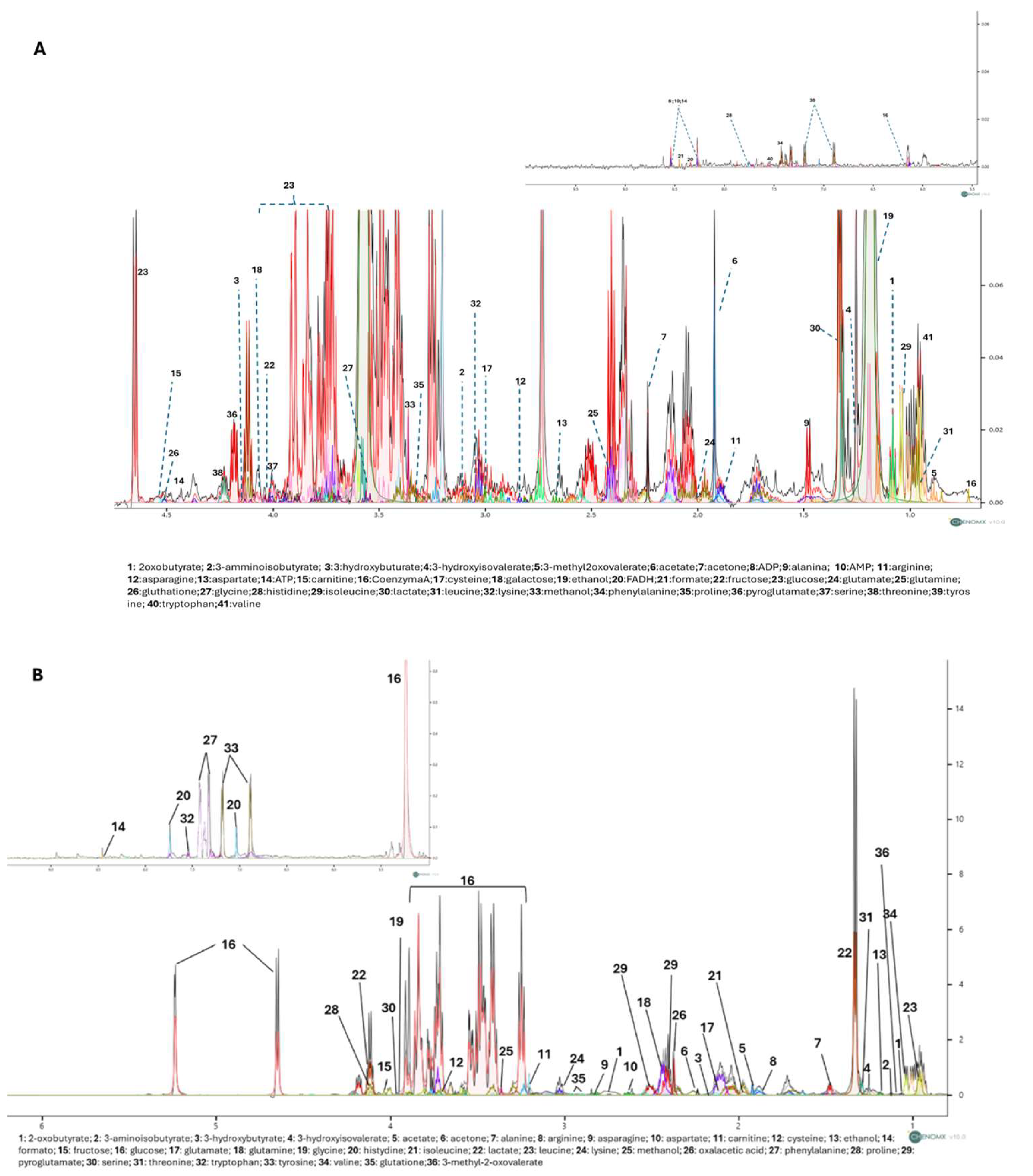
Figure 4.
PLS-DA score plots related to the endometabolome (A) and the esometabolome (B) of treated (green) and untreated (red) cells with ST-65 formulation. VIP score graph reported the metabolites responsible for clusters endometabolome (C) and esometabolome cluster separation (D). Volcano plot analysis of metabolic changes in the cellular polar extract of endo (E) and esometabolome (F) of treated and untreated cells. Each point on the volcano plot was based on p-value and fold-change values, set at 0.05 and 2.0, respectively. Red points identify upregulated metabolites, whereas blue points identify down-regulated metabolites.
Figure 4.
PLS-DA score plots related to the endometabolome (A) and the esometabolome (B) of treated (green) and untreated (red) cells with ST-65 formulation. VIP score graph reported the metabolites responsible for clusters endometabolome (C) and esometabolome cluster separation (D). Volcano plot analysis of metabolic changes in the cellular polar extract of endo (E) and esometabolome (F) of treated and untreated cells. Each point on the volcano plot was based on p-value and fold-change values, set at 0.05 and 2.0, respectively. Red points identify upregulated metabolites, whereas blue points identify down-regulated metabolites.
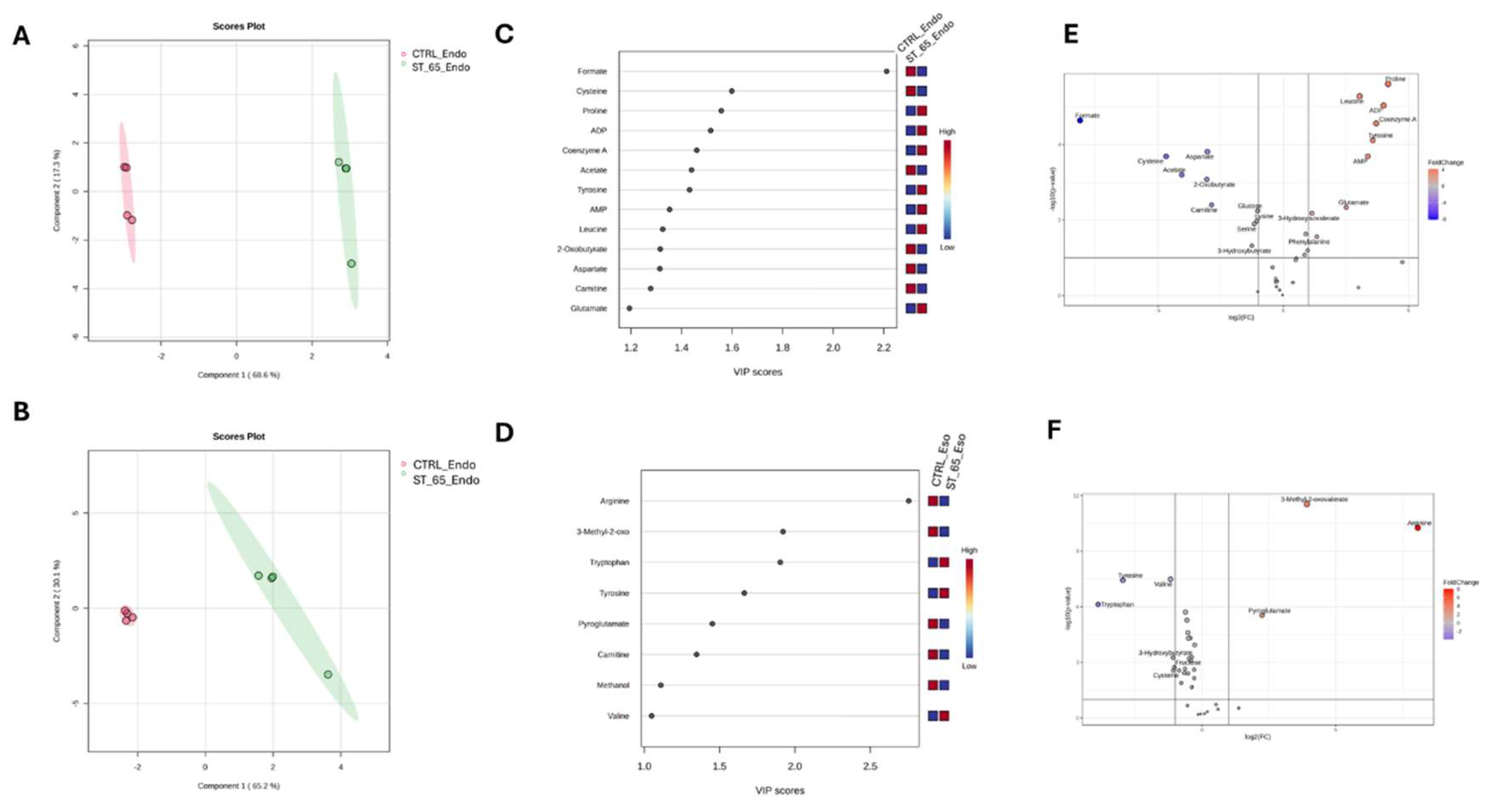
Figure 5.
Enrichment pathways analysis. The y-axis shows the discriminating metabolic pathways in order of p-value (expressed on the x-axis as negative of logarithm) increasing. The size of the circles represents the number of hits given as an enrichment ratio.
Figure 5.
Enrichment pathways analysis. The y-axis shows the discriminating metabolic pathways in order of p-value (expressed on the x-axis as negative of logarithm) increasing. The size of the circles represents the number of hits given as an enrichment ratio.
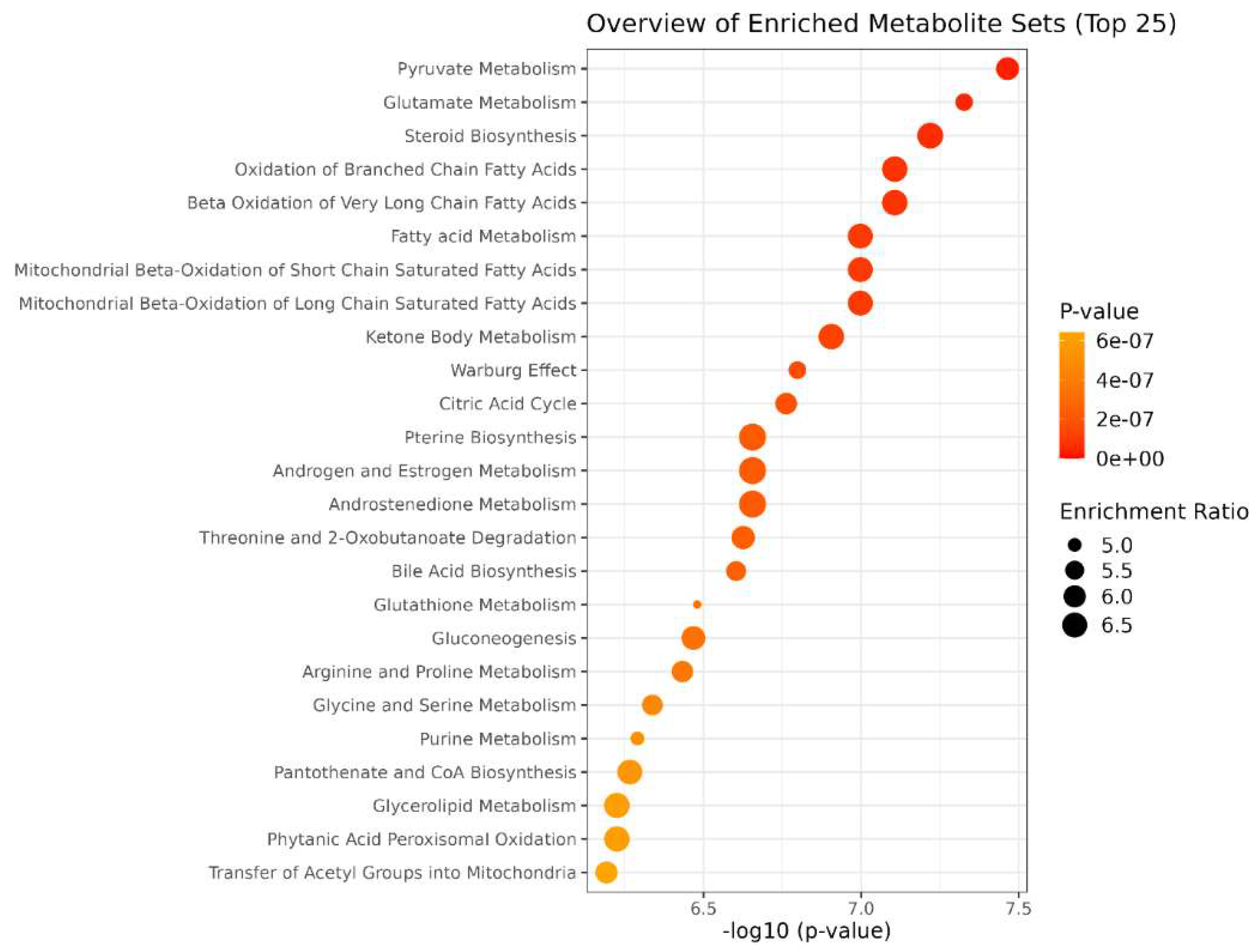

Table 1.
UHPLC-HRMS data of compounds detected in ST-65 formulation.
| N.a | RT [min] | m/z | Formula | ppm | MS/MS | Name |
|---|---|---|---|---|---|---|
| 1 | 6.8 | 224.1644 | C13 H22 O2 N | -0.47 | 124, 93 | unknown |
| 2 | 9.9 | 226.1801 | C13 H24 O2 N | -0.334 | 144, 126, 84 | unknown |
| 3 | 10.9 | 611.1597 | C27 H31 O16 | -1.524 | 465, 303 | Rutinb |
| 4 | 12.71 | 595.1658 | C27 H31 O15 | 0.241 | 449, 287 | Kaempferol hexose-deoxyhexose |
| 5 | 13.1 | 611.1959 | C28 H35 O15 | -1.745 | 447, 303 | Hesperidinb |
| 6 | 16.3 | 303.0497 | C15 H11 O7 | -0.723 | 153 | Quercetinb |
| 7 | 17.4 | 783.4157 | C40 H63 O15 | -0.482 | 459, 441, 423 | Withanoside IV/Withanoside X |
| 8 | 17.6 | 489.2847 | C28 H41 O7 | 0.225 | 317, 299, 281 | Vicosalactone B isomer |
| 9 | 17.8 | 489.2847 | C28 H41 O7 | 0.163 | 317, 299, 281 | Vicosalactone B isomer |
| 10 | 17.8 | 783.4156 | C40 H63 O15 | -0.635 | 459, 441, 423 | Withanoside IV/Withanoside X |
| 11 | 18.1 | 621.363 | C34 H53 O10 | -0.457 | 459, 441, 423 | Coagulin Q |
| 12 | 18.7 | 621.3632 | C34 H53 O10 | -0.264 | 459, 441, 423 | Coagulin Q isomer |
| 13 | 20.8 | 471.2737 | C28 H39 O6 | -0.733 | 435, 341, 299, 281 | Whitanolide A isomer |
| 14 | 21.8 | 471.2739 | C28 H39 O6 | -0.457 | 435, 341, 299, 281 | Whitanolide A isomer |
| 15 | 22.8 | 471.2737 | C28 H39 O6 | -0.797 | 435, 341, 299, 281 | Whitanolide A isomer |
| 16 | 22.8 | 767.4209 | C40 H63 O14 | -0.434 | 443, 425, 407, 389 | Withanoside V |
| 17 | 28.0 | 797.4688 | C42 H69 O14 | 0.861 | 599, 441, 423 | Bacopaside N |
| 18 | 31.4 | 931.5315 | C40 H83 O23 | -0.51 | 477 | unknown |
aCompounds are numbered according to their elution order. bCompared with reference standards.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



