
Submitted:
13 January 2025
Posted:
13 January 2025
You are already at the latest version
Abstract
Objectives: The aim of this study was to evaluate the impact of a novel antioxidant formulation (RP-25) con-taining CoQ10, alpha-lipoic acid, and Chaga extract on mitochondrial dysfunction and oxidative stress. The formulation’s potential to modulate cellular energy metabolism and neurotransmission was explored using a combination of NMR-based metabolomics and UHPLC-HRMS analytical techniques. Methods: SH-SY5Y neu-roblastoma cells were treated with RP-25, and cell viability was assessed via CCK-8 assay. Metabolomic profiles of the treated and untreated cells were analyzed by 1D-NMR, providing insights into the endo- and esometab-olomes. Additionally, a UHPLC-HRMS method was developed for quality control and analysis of the RP-25 formulation. Multivariate statistical approaches, including PLS-DA and volcano plot analysis, were used to identify key metabolic changes. Results: RP-25 treatment did not affect cell viability but significantly modulat-ed metabolic pathways, including amino acid biosynthesis, oxidative phosphorylation, and glycolysis. Increased levels of ATP, glutamate, tyrosine, and proline were observed in treated cells, indicating enhanced cellular en-ergy production and neurotransmission activity. Pathway enrichment analysis highlighted the formulation's influence on mitochondrial energy metabolism and oxidative stress response. Conclusions: RP-25 demonstrated potential as a therapeutic strategy for mitigating mitochondrial dysfunction and oxidative stress, with implica-tions for conditions associated with aging and neurodegenerative diseases.
Keywords:
Coenzyme Q10
; Lipoic acid
; Inonotus obliquus
; SH-SY5Y cells
; UHPLC-HRMS/MS
; 1H NMR metabolomics
; Mitochondrial dysfunction
; Cellular metabolism
; Neurotransmission
; Fibromyalgia
1. Introduction
Mitochondrial dysfunction has emerged as a central feature of aging and a variety of age-related diseases, including neurodegenerative disorders, cardiovascular diseases, diabetes, cancer, and fibromyalgia [1,2]. Mitochondria, the powerhouse of the cell, are responsible for producing the majority of cellular ATP through oxidative phosphorylation, a process that is highly sensitive to stress and damage. In particular, mitochondrial dysfunction is often associated with inefficient respiration, leading to the overproduction of reactive oxygen species (ROS), which can damage cellular components such as lipids, proteins, and DNA [3,4]. This accumulation of oxidative damage is thought to contribute significantly to cellular senescence, chronic inflammation, and the pathogenesis of numerous diseases.
In recent years, research has increasingly implicated mitochondrial dysfunction in fibromyalgia [5] a chronic pain syndrome that affects millions of people worldwide. Patients with fibromyalgia exhibit altered mitochondrial function, which is believed to contribute to their symptoms, including fatigue, pain, and impaired energy production [6]. Studies have shown that mitochondrial dysfunction in fibromyalgia leads to impaired cellular respiration, exacerbating oxidative stress and inflammation, and contributing to the disease's hallmark features, including muscle pain and fatigue [7,8]. This has led to growing interest in targeting mitochondrial health as a potential therapeutic strategy for fibromyalgia and other chronic pain disorders.
A promising approach to combat mitochondrial dysfunction and its downstream effects is the use of bioactive compounds with antioxidant and mitochondrial-supportive properties. Among these, coenzyme Q10 (CoQ10), α-lipoic acid, and extracts from medicinal mushrooms have garnered significant attention due to their well-documented beneficial effects on cellular metabolism and mitochondrial health [9]. CoQ10, a vital component of the electron transport chain, has been shown to enhance mitochondrial function and reduce oxidative stress. Similarly, α-lipoic acid, a potent antioxidant, is known to improve mitochondrial efficiency and restore cellular redox balance [10,11]. Chaga mushroom (Inonotus obliquus), rich in bioactive compounds such as β-glucans, has also demonstrated promising effects on immune modulation, oxidative stress reduction, and mitochondrial health [12].
In this context, we developed a novel formulation, RP-25, which combines CoQ10, α-lipoic acid, and Chaga extract, along with additional micronutrients (vitamins E and B3, dihydroquercetin, zinc, manganese, copper, and selenium), designed to enhance mitochondrial function and combat oxidative stress. The aim of this study was to investigate the effects of RP-25 on the metabolism of SH-SY5Y neuroblastoma cells, focusing on its potential to modulate mitochondrial function and reduce oxidative stress. Using an untargeted 1H-NMR metabolomic approach, we aimed to gain insights into the broader metabolic shifts induced by RP-25 treatment, specifically in relation to mitochondrial respiration, energy production, and the regulation of metabolites linked to oxidative stress.
Metabolomics, particularly Nuclear Magnetic Resonance (NMR) spectroscopy, has emerged as a powerful tool for studying cellular metabolism and identifying disordered metabolic pathways that underlie various diseases. NMR-based metabolomics provides a non-invasive, comprehensive, and high-throughput method to monitor the entire metabolic profile of cells, revealing subtle changes in cellular metabolism that are often indicative of pathological states [13]. In the context of mitochondrial dysfunction, NMR allows the identification of key metabolic biomarkers and pathways disrupted by oxidative stress, energy dysregulation, or altered cellular function. This makes NMR-based metabolomics an invaluable tool for assessing the metabolic effects of therapeutic interventions and gaining insights into the molecular mechanisms of diseases linked to mitochondrial dysfunction, such as fibromyalgia, neurodegenerative diseases, and cancer.
Through this study, we sought to evaluate the potential of RP-25 as a therapeutic strategy for mitigating mitochondrial dysfunction and associated cellular damage, with implications not only for aging and related chronic diseases but also for conditions like fibromyalgia, where mitochondrial dysfunction plays a key role in disease pathophysiology.
2. Materials and Methods
2.1. Chemical and Standards
Analytical grade ethanol (EtOH), methanol (MeOH), chloroform, MS grade formic acid (HCOOH), and the reference standards of rutin, hesperidin, quercetin, were provided by Merck Chemicals (Milan, Italy). MS grade acetonitrile and water were purchased from Romil (Cambridge, UK). Ultrapure water (18 MΩ) was prepared using a Milli-Q purification system (Millipore, Bedford, TX, USA).
2.2. Formulation of RP-25
The formulation of RP-25 consisted of several active ingredients, including Coenzyme Q10 (CoQ10), α-lipoic acid, Chaga fruit extract (Inonotus obliquus) standardized to 25% beta-glucans, dihydroquercetin, and a selection of essential vitamins and minerals. Specifically, the formulation included CoQ10 at 200 mg, α-lipoic acid at 200 mg, Chaga fruit extract (Inonotus obliquus) with 25% beta-glucans at 200 mg, Vitamin E at 48 mg, dihydroquercetin at 50 mg, Vitamin B3 (niacin) at 16 mg, and essential trace elements including Zinc (10 mg), Manganese (4 mg), Copper (1 mg), and Selenium (0.075 mg). The Chaga extract used in the formulation was a water-based extract, along with all other components of the RP-25 formulation, kindly supplied by Laboratoarele Medica Srl (Bu-105 Charest, Romania). The preparation of RP-25 was carried out at the Department of Pharmacy of the University of Salerno, where all ingredients were measured using an analytical balance for precision. The active components were carefully weighed and combined in a mixing bowl, ensuring thorough mixing to achieve even distribution of the various ingredients. Special care was taken during the blending process to maintain the integrity and efficacy of each component, ensuring a homogenous mixture for optimal formulation. The final product was carefully prepared to meet the required specifications for each ingredient and to guarantee its therapeutic potential.
2.3. UHPLC-HRMS/MS Analysis of RP-25 Formulation
The RP-25 formulation was analyzed using a high-resolution UHPLC-HRMS/MS method to profile and characterize its chemical constituents. The samples were dissolved in a hydroalcoholic solution (70% v/v) with a matrix/solvent ratio of 1:20. Chromatographic analysis was performed using a Vanquish Flex UFPLC system (Thermo Fisher Scientific, Milan, Italy) interfaced with an Orbitrap Exploris 120 mass spectrometer equipped with a heated electrospray ionization source (HESI-II). The system utilized a Kinetex C18 column (100 × 2.1 mm I.D., 2.6 µm; Phenomex, Bologna, Italy) at a flow rate of 500 µL/min and a column temperature of 30°C. A 5 µL injection was used for analysis, with the mobile phase consisting of water (A) and MeCN (B), both containing 0.1% formic acid. A gradient elution method was employed, ranging from 2% to 98% MeCN, and each run was followed by a washing and re-equilibration step to ensure system stability. The mass spectrometer was operated in both negative and positive ionization modes, utilizing Full MS/dd-MS2 acquisition for data collection. The resolution was set to 60,000 FWHM for Full MS scans, with fragmentation via stepped HCD (20, 40, and 60 eV) for targeted MS/MS analysis. The detected compounds were identified based on accurate mass, fragmentation patterns, and retention times, with additional confirmation through reference standards or published chemo-taxonomic data. The analysis resulted in the identification of fifteen compounds in the RP-25 formulation, including seven flavonoids, three fatty acids, and four saponins, as detailed in Table 1.
2.4. Coenzyme Q10 Analysis
For the determination of CoQ10, a separate UHPLC-UV method was employed. The sample preparation involved extraction with n-hexane (matrix/solvent ratio of 1:20) followed by sonication for 30 minutes and centrifugation at 13,000 rpm. The supernatant was evaporated under a nitrogen stream, and the residues were reconstituted with the mobile phase (ACN:THF: Milli-Q water, 50:40:5, v/v/v). The chromatographic separation was achieved using a Kinetex C18 column (100 × 2.1 mm I.D., 2.6 µm; Phenomex, Bologna, Italy) at a flow rate of 400 µL/min, and the analysis was carried out under isocratic conditions with detection at 265 nm. CoQ10 was quantified by comparing the sample's UV profile to a reference standard (250 µg/mL).
2.5. Cell Culture
Undifferentiated SH-SY5Y neuroblastoma cells (ATCC, Rockville, MD, USA) were cultured in Dulbecco’s Modified Eagle Medium (DMEM, 4500 mg/mL glucose), supplemented with 10% fetal bovine serum, 2 mM L-glutamine, 100 U/mL penicillin, and 0.1 mg/mL streptomycin. Cells were maintained in a humidified incubator at 37°C with 5% CO2, and were subcultured every 2 days.
2.6. Cell Viability Assay
Cell viability was assessed using the Cell Counting Kit-8 (CCK-8; Dojindo Laboratories, Rockville, MD, USA), which measures the ability of viable cells to reduce the tetrazolium salt WST-8. SH-SY5Y cells were plated in a 96-well plate at a density of 8 × 10³ cells/well, allowed to adhere overnight, and then treated with various concentrations of RP-25 (0.80–200 μg/mL) for 24 hours. After treatment, CCK-8 solution (10 μL) was added to each well, and absorbance was measured at 450 nm. Cell viability was expressed as the percentage of viable cells relative to untreated controls. Data are presented as mean ± SD from three independent experiments. The statistical significancy was evaluated using the one-way analysis of variance (ANOVA), followed by Bonferroni’s test, with GraphPad Prism 8.0 software (San Diego, CA, USA) assuming significance at p < 0.05.
2.6. Exposure of Cells to RP-25
SH-SY5Y cells were plated in 100 mm culture dishes (Corning, Corning, NY, USA) and allowed to adhere overnight. The cells were then treated with RP-25 (25 μg/mL) for 24 hours. The control group was treated with vehicle only. At the end of the treatment, the cell medium was collected, and the dishes were washed with cold PBS (pH 7.4) before metabolite extraction.
2.7. Sample Extraction
The culture medium was collected, centrifuged at 1000× g for 5 minutes, and the cellular metabolites were extracted using a methanol:chloroform:water (1:1:1) mixture. After cell scraping and homogenization, samples were centrifuged at 6000 rpm for 10 minutes at 4°C to separate the phases [14]. The polar extracts were dried under vacuum using an SP-Genevac EZ-2 4.0 concentrator, and the resulting extracts were stored at -80°C prior to NMR analysis.
2.8. NMR Spectra Acquisition
Bruker Ascend™ 600 MHz spectrometer was used to acquire the spectra. The spectrometer was equipped with a 5 mm triple resonance Z gradient TXI probe (Bruker Co., Rheinstetten, Germany) at 298 K. TopSpin, version 3.2 was used for the spectrometer control and data processing (Bruker Biospin). All the experiments performed as Nuclear Overhauser Enhancement Spectroscopy (NOESY) 1D were acquired in triplicate. Spectra acquisition was made using 12 ppm spectral width, 20 k data points, presaturation during relaxation delay and mixing time for water suppression [15] and spoil gradient, 5 s relaxation delay, and mixing time of 10 ms. A weighted Fourier transform was applied to the time domain data with a line widening of 0.5 Hz followed by a manual step and baseline correction in preparation for targeted profiling analysis.
2.9. Statistical Analysis
NMR spectra of SH-SY-5Ycells cultures eso and endometabolome were analysed using an untargeted metabolomic approach. All the spectra were assigned using Chenomx NMR-Suite v8.0 (Chenomx Inc., Edmonton, 252 Canada) and quantified by NMRProcFlow as previously reported [16]. The quantification matrices reported the metabolites identifiedand quantified in the eso and endometabolome of SH-SY-5Y treated and untreated with ST-65 were analyzed using the open-source tool Metaboanalyst 6.0 [17]. The Volcano plot combined T-test and Fold Change performed the univariate approach [18,19]. After normalization by sum log and Pareto, we applied a supervised multivariate approach partial 1least-squares discriminant analysis (PLS-DA) method. The reliability of the supervised model was analyzed using a cross-validation approach, considering the accuracy and pa- rameters Q2 and R2. The metabolites responsible for clusters’ separation in the PLS-DA score plot were classified according to VIP, considering only the metabolites with VIP>1 [17]. Enrichment Pathways tools were applied to identify the dysregulated biochemical pathways by KEGG database. Only the KEGG paths that reported a rate of false discoveries (FDR) lower than 1, the p-value lower than 0.05 and the hits value related to the number of metabolites belonging to the pathway >1, were chosen [19].
3. Results
3.1. UHPLC−HRMS/MS Analysis
The UHPLC-HRMS/MS analysis of the RP-25 formulation facilitated the identification and characterization of several key bioactive compounds present in the product. Given that the Chaga extract (Inonotus obliquus) was incorporated into the formulation, the detected compounds can be attributed not only to the active ingredients added to the formulation (such as CoQ10, α-lipoic acid, and dihydroquercetin) but also to the bioactive constituents derived from the Chaga extract itself. As presented in Table 1 and the corresponding chromatograms (Figure 1), the high-resolution mass spectrometry analysis revealed the presence of several compound classes. Flavonoids: a series of flavonoid derivatives, including quercetin, kaempferol, isorhamnetin, and rutin, were identified in the RP-25 formulation. These compounds are likely derived from the Chaga extract, which is known for its high content of phenolic compounds exhibiting potent antioxidant activity [20]. The MS/MS fragmentation patterns provided clear evidence for the presence of quercetin and kaempferol derivatives, with product ions observed at m/z 301.0344, 285.0403 (quercetin derivatives) and m/z 285.0403, 271.0476 (kaempferol derivatives). These findings are consistent with previous reports attributing significant antioxidant activity to these flavonoids in Chaga [21]. The peaks observed in the UHPLC chromatogram at retention times of 9.7, 10.8, 12.6, and 13.0 minutes correspond to quercetin pentose-rutinoside (2), rutin (3), kaempferol rutinoside (4), and isorhamnetin rutinoside (5), as detailed in Table 1. Fatty Acids: alpha-lipoic acid (m/z 205.0362), a key ingredient in the formulation, was readily detected as expected. Additionally, isomers of dodecenedioic acid (compounds 10 and 11) were identified in the formulation. These compounds are likely derived from the Chaga extract, known to contain various fatty acids and their derivatives [22]. The mass spectra for these compounds (m/z 227.1289 for dodecenedioic acid isomers) are consistent with those reported in the literature for Chaga-derived fatty acids. The identification of alpha-lipoic acid at retention time 17.3 min and the dodecenedioic acid isomers at 22.8 min and 23.5 min further corroborates the formulation's composition as outlined in Table 1. Saponins: several saponin-like compounds (12-15) were detected, with the aglycone moiety identified through characteristic fragmentation patterns. The MS/MS spectra showed a prominent fragment ion at m/z 439 for compounds 12, 13, and 14, consistent with the presence of a C30H48O2 aglycone, identified as 3-β,22-dihydroxy-lanosta-7,9,24-triene. These compounds are likely derived from the Chaga extract, which is known to contain diverse saponins [23]. The observed fragmentation patterns confirm the presence of these compounds. These saponins were eluted between 26.9 min and 28.7 min, as indicated in Table 1. Since the analysis was conducted on the complete RP-25 formulation, the compounds detected, including flavonoids, fatty acids, and saponins, are attributed to the Chaga extract contribution within the formulation. UHPLC-UV analysis was employed to determine the concentration of CoQ10 in the RP-25 formulation. CoQ10 was detected at a concentration of 200 mg per dose, which corresponds to the expected amount in the formulation. The CoQ10 chromatogram shows a distinct peak at a retention time of 2.8 min, with a mass value of m/z 863.4467, corresponding to CoQ10 in its integrated form within the formulation matrix saponins [24]. The peak intensity and area under the curve are in full agreement with the expected CoQ10 concentration, thereby confirming that the CoQ10 content in the RP-25 formulation meets the intended specification. The CoQ10 analysis, therefore, validates the accuracy and precision of the formulation preparation, confirming the quality of the final product. The analytical data confirmed that the RP-25 formulation contains the expected active ingredients, CoQ10, α-lipoic acid, and dihydroquercetin, at their specified concentrations. The overall chromatographic and mass spectrometric profile aligns with the intended composition of the product, supporting its quality and consistency.
u

Table 1.
UHPLC-HRMS data of compounds detected in RP-25 formulation.
| N.a | RT [min]. | m/z | Formula | ppm | MS/MS | Name |
|---|---|---|---|---|---|---|
| 1 | 7.0 | 443.0696 | C32 H11 O3 | 1.424 | 221, 125, 80 | unknown |
| 2 | 9.7 | 741.188 | C32 H37 O20 | 1.053 | 300/301 | Quercetin pentose-rutinoside |
| 3 | 10.8 | 609.1457 | C27 H29 O16 | 1.213 | 300/301 | Rutinb |
| 4 | 12.6 | 593.1509 | C27 H29 O15 | 1.422 | 284/285, 255, 227 | Kaempferol rutinoside |
| 5 | 13.0 | 623.1613 | C28 H31 O16 | 1.057 | 315, 299, 271 | Isorhamnetin rutinoside |
| 6 | 15.0 | 253.0506 | C15 H9 O4 | 4.563 | Dadzeinb | |
| 7 | 16.3 | 301.0353 | C15 H9 O7 | 3.425 | 179, 151 | Quercetinb |
| 8 | 17.3 | 205.0362 | C8 H13 O2 S2 | 5.084 | 171, 127 | lipoic acid |
| 9 | 17.6 | 269.0456 | C15 H9 O5 | 4.349 | Apigeninb | |
| 10 | 22.8 | 227.1289 | C12 H19 O4 | 5.127 | 209,183,165 | Dodecenedioic acid isomer |
| 11 | 23.5 | 227.1288 | C12 H19 O4 | 4.686 | 209,183,165 | Dodecenedioic acid isomer |
| 12 | 26.9 | 941.5097 | C48 H77 O18 | 0.713 | 439 | lanostane-type saponin |
| 13 | 27.8 | 795.4530 | C42 H67 O14 | 0.587 | 439 | lanostane-type saponin |
| 14 | 27.9 | 911.5001 | C47 H75 O17 | 0.332 | 439, 421 | lanostane-type saponin |
| 15 | 28.7 | 925.5159 | C48 H77 O17 | 0.435 | 423 | lanostane-type saponin |
aCompounds are numbered according to their elution order. bCompared with reference standards.
3.2. Cell Viability
As a preliminary step, the effect of the RP-25 formulation on cell viability was evaluated using SH-SY5Y neuroblastoma cells. Figure 2 illustrates that the RP-25 formulation did not significantly affect the viability of SH-SY5Y cells after 24 hours of treatment, even at the highest concentration of 200 μg/mL.
3.3. Untargeted 1H-NMR Metabolomics Analysis
To further explore the metabolic effects of RP-25, we employed an untargeted 1H-NMR metabolomic approach to analyze the endo- and exometabolomes of SH-SY5Y cells, both treated and untreated with RP-25. Using Chenomix software, 42 metabolites were identified in the endometabolome and 39 in the exometabolome (Figure 3A, 3B). The metabolite quantification was performed relative to the internal standard (TSP).
The quantification matrices were analyzed using MetaboAnalyst 6.0, employing both univariate and multivariate approaches. After normalization (median scaling and log transformation), univariate analysis and multivariate analysis using Partial Least Squares Discriminant Analysis (PLS-DA) were performed [25].
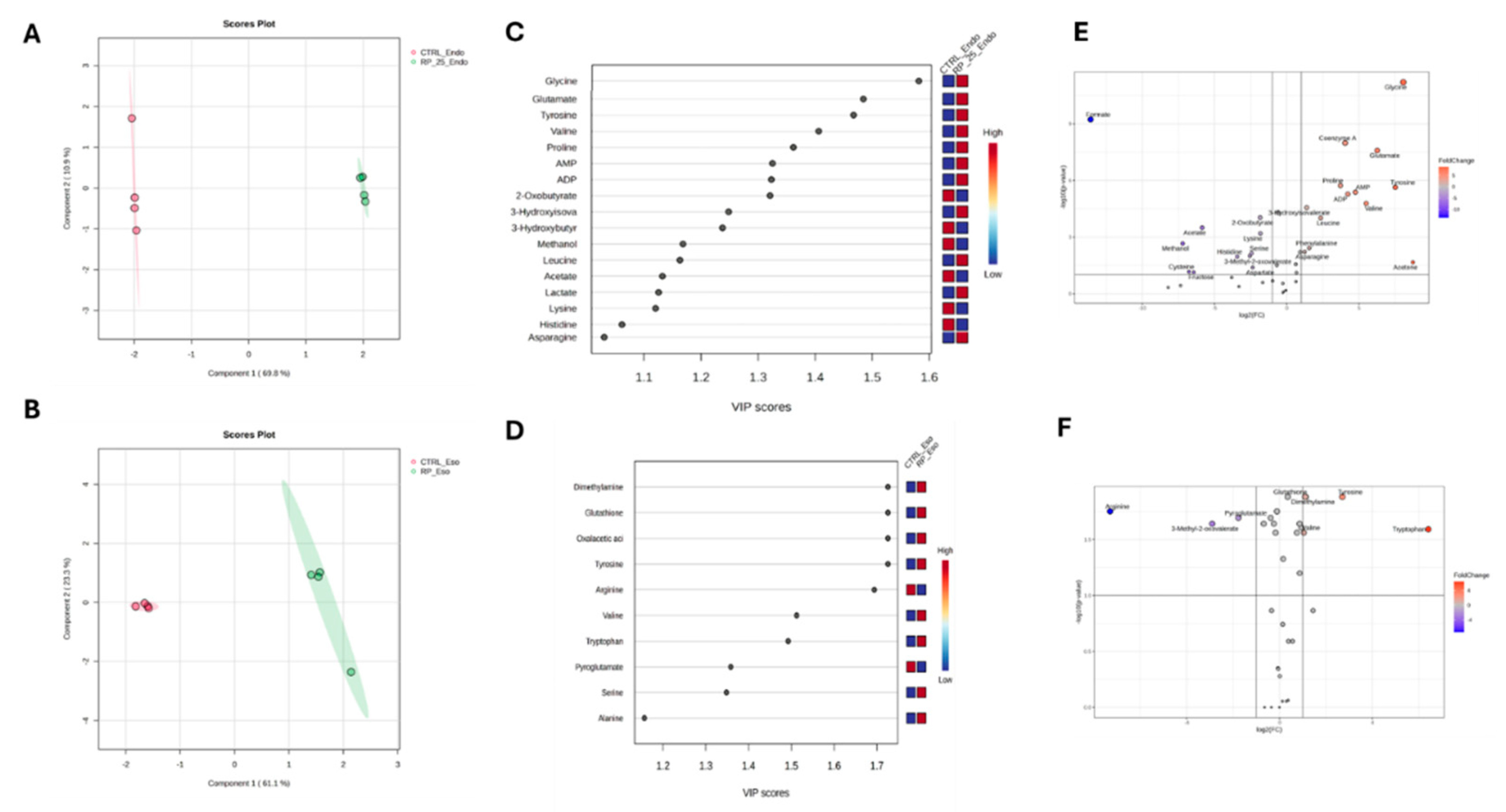
The PLS-DA score plots reveal distinct metabolic profiles between RP-25-treated and untreated cells in both the endo- and exometabolomes (Figure 4A, 4B). Cross-validation (CV) analysis confirmed the robustness of the model, with R2 values of 1.0 for both components and Q2 values of 0.98 and 0.86 for the endometabolome and exometabolome, respectively [26].
Volcano plot analysis showed a significant decrease in the concentration of Formate in the endometabolome of treated cells, along with increased levels of Glycine, Coenzyme A, Proline, Tyrosine, Valine, Acetone, AMP, and ATP. Elevated levels of Tyrosine were also observed in the exometabolome, along with increased concentrations of Tryptophan and decreased Arginine levels in the medium of treated cells (Figures 4C, F).
Variable Importance Projection (VIP) analysis identified key metabolites driving the separation between treated and untreated groups [27]. Among the metabolites with VIP > 1.0, Tyrosine, Glycine, Glutamate, Valine, and Proline were significantly upregulated in treated cells (Figure 4B). In the exometabolome, higher concentrations of Tyrosine, Serine, and Alanine were observed (Figure 4F).
3.4. Enrichment Pathway Analysis
Enrichment pathway analysis confirmed the impact of RP-25 on several metabolic pathways, particularly those related to neurotransmission and cellular energy metabolism. Notably, dysregulated pathways included Tyrosine metabolism, Phenylalanine metabolism, and Phenylalanine-Tyrosine-Tryptophan biosynthesis, as well as Glutamate, Alanine, Aspartate, and Glycolysis/Gluconeogenesis metabolism (Table 2). Increased AMP and ATP concentrations suggest potential effects on oxidative phosphorylation and cellular energy regulation.
Table 1.
Enrichment pathway analysis of significant biochemical pathways affected by RP-25 treatment. Pathways with more than two metabolites and p-value and False Discovery Rate (FDR) values below 0.05 are considered significant.
Table 1.
Enrichment pathway analysis of significant biochemical pathways affected by RP-25 treatment. Pathways with more than two metabolites and p-value and False Discovery Rate (FDR) values below 0.05 are considered significant.
| Hits | p.value | FDR | |
|---|---|---|---|
| Pantothenate and CoA biosynthesis | 4 | 9,56E-05 | 3,82E-03 |
| Glyoxylate and dicarboxylate metabolism | 6 | 4,60E-02 | 9,20E-01 |
| Tyrosine metabolism | 3 | 4,63E-01 | 0.00046293 |
| Lipoic acid metabolism | 3 | 0.00052912 | 0.0035275 |
| Histidine metabolism | 3 | 0.0015907 | 0.0079534 |
| Glutathione metabolism | 5 | 0.0018923 | 0.0084101 |
| Phenylalanine metabolism | 2 | 0.0028912 | 0.0096374 |
| Phenylalanine tyrosine and tryptophan biosynthesis | 2 | 0.0028912 | 0.0096374 |
| Glycolysis / Gluconeogenesis | 2 | 0.0060371 | 0.017249 |
| Pyruvate metabolism | 2 | 0.0060371 | 0.017249 |
| Taurine and hypotaurine metabolism | 2 | 0.015772 | 0.03943 |
| Alanine aspartate and glutamate metabolism | 4 | 0.020867 | 0.049098 |
| Arginine and proline metabolism | 3 | 0.026161 | 0.038135 |
4. Discussion
This study aimed to evaluate the effects of the RP-25 formulation, composed of CoQ10, α-lipoic acid, Chaga extract, and micronutrients, on the mitochondrial function and metabolic profiles of SH-SY5Y neuroblastoma cells, using untargeted 1H-NMR metabolomics. The results demonstrate that RP-25 treatment induces significant changes in cellular metabolism, providing valuable insights into its potential therapeutic effects, particularly in the context of mitochondrial dysfunction and diseases associated with oxidative stress, including neurodegenerative disorders and fibromyalgia.
4.1. RP-25 Formulation
The RP-25 formulation represents a complex blend of active, including CoQ10, alpha-lipoic acid, dihydroquercetin, Chaga extract, and essential micronutrients, each contributing to the overall cellular bioenergetic and antioxidant profile of the product. Analytical characterization via UHPLC-HRMS/MS and HPLC-UV confirmed the presence of all targeted compounds and further identified additional bioactive constituents derived from the Chaga extract, underscoring the complex nature of this formulation. The synergistic interaction between these components is aimed at improving mitochondrial efficiency, reducing oxidative stress, and supporting cellular integrity across multiple biological systems.
Coenzyme Q10 (CoQ10) is an integral component of the mitochondrial electron transport chain, specifically within complex I and III, where it functions as an electron carrier. CoQ10 facilitates the transfer of electrons from NADH and FADH2 to oxygen, driving the production of ATP through oxidative phosphorylation. Beyond its role in bioenergetics, CoQ10 is a potent lipophilic antioxidant that stabilizes cellular membranes and scavenges reactive oxygen species (ROS), mitigating oxidative damage at the cellular level [28]. The decline in CoQ10 levels with age is associated with mitochondrial dysfunction and increased oxidative stress, contributing to cellular senescence and age-related diseases[29]. In RP-25, CoQ10 is present at 200 mg per dose, as confirmed by UHPLC-UV analysis, ensuring that the formulation provides sufficient concentrations to support mitochondrial bioenergetics and antioxidant defenses. This dosage is intended to restore mitochondrial function, enhance ATP synthesis, and protect against ROS-induced cellular damage, particularly in energy-demanding tissues, such as muscle and nervous tissue [30,31].
Alpha-lipoic acid (ALA), a sulfur-containing fatty acid, is both a cofactor for mitochondrial enzymes and a multifaceted antioxidant. ALA participates in mitochondrial bioenergetics by acting as a coenzyme for pyruvate dehydrogenase, an enzyme complex crucial for the conversion of pyruvate into acetyl-CoA, thus facilitating the entry of carbon substrates into the citric acid cycle (Krebs cycle). This action optimizes the production of NADH and FADH2, which feed into the electron transport chain for ATP synthesis. ALA also enhances the regeneration of other antioxidants, including vitamins C and E, through the reduction of their oxidized forms [32,33]. This regenerative effect amplifies the antioxidant capacity of cells, particularly in high-oxidative stress environments. In RP-25, ALA is included at a concentration of 200 mg per dose, which has been shown in previous studies to restore mitochondrial function, enhance cellular redox balance, and reduce inflammatory markers associated with aging [34]. The UHPLC-HRMS analysis confirmed its incorporation at the expected retention time of 17.3 minutes, indicating its optimal concentration for cellular protection and energy metabolism.
A central feature of RP-25 is the inclusion of Chaga extract (Inonotus obliquus), which contains a diverse array of bioactive compounds, including flavonoids, polyphenols, polysaccharides and terpenoids, each contributing to the formulation’s antioxidant, anti-inflammatory, and immunomodulatory effects [35,36]. Flavonoid derivatives such as quercetin, kaempferol, and isorhamnetin, identified through UHPLC-HRMS/MS, are potent antioxidants known to scavenge free radicals and modulate intracellular signaling pathways that regulate oxidative stress. These flavonoids exert their effects by inhibiting the production of ROS and enhancing the activity of antioxidant enzymes, such as superoxide dismutase (SOD) and catalase, thereby reducing oxidative damage [37,38]. Polysaccharides from Chaga exhibit a wide range of health-promoting effects, one of which involves the activation of the Nrf2-Keap1 pathway, a pivotal cellular defense mechanism. This pathway orchestrates the transcriptional activation of phase II detoxifying enzymes, including quinone reductase 1 (NQO1) and glutathione S-transferase (GST). The upregulation of these enzymes enhances the cell’s antioxidant capacity, facilitating the neutralization and detoxification of reactive oxygen species (ROS) and electrophilic toxins, thereby maintaining cellular redox homeostasis and mitigating oxidative stress [39]. These mechanisms suggest the potential of RP-25 in protecting against oxidative damage, supporting cellular longevity and possibly reducing the risk of chronic degenerative diseases by enhancing cellular defense systems and maintaining redox balance.
Moreover, Chaga extract also contributes bioactive fatty acids, such as dodecenedioic acid, which have been implicated in the regulation of cellular membrane fluidity and stability. These fatty acids contribute to maintaining membrane integrity and enhancing the function of membrane-bound enzymes, which are essential for proper cell signaling and metabolic processes [40]. The identification of saponins (compounds 12–15) in RP-25 further supports its antioxidant and immune-modulating properties. Saponins are known for their ability to interact with cellular membranes and modulate immune responses by influencing the activity of macrophages and dendritic cells. These compounds can inhibit the activation of nuclear factor-kappa B (NF-κB), a transcription factor that regulates the expression of pro-inflammatory cytokines, thereby reducing systemic inflammation and promoting tissue homeostasis [41].
Micronutrients such as zinc, manganese, copper, and selenium are essential cofactors in key antioxidant enzyme systems. Zinc, for example, is a critical cofactor for the enzyme superoxide dismutase (SOD), which catalyzes the conversion of superoxide radicals to hydrogen peroxide, thereby preventing oxidative damage [42]. Selenium is incorporated into the active site of glutathione peroxidase, an enzyme responsible for detoxifying hydrogen peroxide and lipid peroxides, protecting cellular components such as lipids and proteins from oxidative modification [43]. Copper and manganese contribute to the activity of SOD by facilitating the conversion of superoxide radicals into less harmful molecules, reducing oxidative injury to cellular structures [44,45,46]. The inclusion of these micronutrients in RP-25 thus enhances the formulation's capacity to maintain cellular redox balance and mitigate the harmful effects of oxidative stress.
Vitamins such as vitamin E and vitamin B3 (niacinamide) further complement the formulation's antioxidant profile. Vitamin E, a fat-soluble antioxidant, integrates into cellular membranes, where it neutralizes lipid peroxyl radicals and protects membranes from oxidative damage. This action is particularly beneficial in reducing photoaging and UV-induced cellular damage by preventing lipid peroxidation in epidermal cells [47]. Niacinamide, or vitamin B3, plays an integral role in cellular respiration by participating in the synthesis of NAD+ (nicotinamide adenine dinucleotide), which is essential for mitochondrial function and energy production. Additionally, niacinamide has been shown to modulate the expression of matrix metalloproteinases (MMPs), enzymes involved in the breakdown of extracellular matrix components, thus improving tissue integrity and reducing collagen degradation [48]. The inclusion of these vitamins enhances the cellular resilience of tissues, supporting the overall longevity and health of cells exposed to chronic oxidative and environmental stress.
Finally, dihydroquercetin (taxifolin), a flavonoid with known vascular protective and antioxidant properties, contributes to the formulation’s efficacy by enhancing microcirculation and supporting collagen synthesis. Dihydroquercetin modulates the expression of key vascular markers, such as vascular endothelial growth factor (VEGF), thereby improving capillary function and promoting nutrient delivery to tissues. This action improves collagen cross-linking and maintains tissue elasticity, which is crucial for cellular function and the prevention of age-related tissue degeneration [49,50].
In summary, the RP-25 formulation combines multiple bioactive compounds that function synergistically to optimize mitochondrial function, enhance cellular bioenergetics, and provide robust antioxidant protection. The inclusion of both plant-derived bioactives and essential micronutrients supports the formulation’s broad-spectrum ability to combat oxidative stress, maintain redox balance, and improve cellular resilience. The detailed analysis through UHPLC-HRMS/MS and HPLC-UV has confirmed the identity and concentrations of all active ingredients, ensuring the consistency, quality, and therapeutic potential of the formulation.
4.2. Impact on Mitochondrial Function and Energy Metabolism
A key finding of this study is the modulation of cellular energy metabolism following RP-25 treatment, as revealed by 1H-NMR metabolomics. Notably, RP-25 treatment led to significant increases in the intracellular concentrations of AMP and ATP, indicating an enhancement in cellular energy production [51]. These changes suggest a potential improvement in mitochondrial bioenergetics, with RP-25 likely promoting oxidative phosphorylation and ATP synthesis. The increased ATP levels in particular point towards a more efficient mitochondrial energy production, which is essential for cellular function. This observation is consistent with the known roles of CoQ10 and α-lipoic acid in improving mitochondrial efficiency. CoQ10, a critical electron carrier in the mitochondrial electron transport chain (ETC), optimizes electron flow, thereby facilitating ATP production and maintaining redox balance, while α-lipoic acid serves as a cofactor in several key mitochondrial enzyme complexes, including pyruvate dehydrogenase, which is pivotal for efficient energy production.
In addition to the direct evidence of enhanced ATP production, the metabolomic analysis highlighted significant shifts in key metabolic pathways involved in energy metabolism, particularly in glycolysis, gluconeogenesis, and pyruvate metabolism. Specifically, RP-25 treatment was associated with a notable increase in oxaloacetate excretion and a reduction in ketone body levels such as 3-hydroxybutyrate, acetate, and 2-oxobutyrate. These metabolic changes suggest a reprogramming of cellular metabolism towards more efficient ATP production, potentially via enhanced glycolytic activity and reduced reliance on fatty acid oxidation pathways, including ketogenesis. The shift from ketogenesis to glycolysis aligns with a more efficient energy production mechanism, possibly facilitated by RP-25’s ability to optimize mitochondrial function [52].
The metabolic changes observed were further validated by enrichment pathway analysis, which identified significant dysregulation in energy-producing pathways, such as glycolysis, gluconeogenesis, and pyruvate metabolism. These alterations in key metabolic routes suggest that RP-25 may help restore metabolic homeostasis in cells affected by mitochondrial dysfunction, a hallmark of diseases like fibromyalgia. In fibromyalgia, mitochondrial inefficiency and disrupted cellular energy production contribute significantly to the pathophysiology of the disease, manifesting as fatigue, pain, and cognitive dysfunction [8,53,54]. The observed modulation of energy pathways in response to RP-25 treatment offers a promising avenue for therapeutic intervention in such conditions.
Taken together, these metabolomic findings support the hypothesis that RP-25 may help optimize cellular metabolism by improving mitochondrial function and energy production. These changes have significant therapeutic potential for conditions characterized by mitochondrial dysfunction and impaired energy metabolism, including fibromyalgia. Further studies are needed to confirm these effects and elucidate the specific molecular mechanisms by which RP-25 influences mitochondrial activity and overall metabolic regulation.
4.3. Neurotransmission and Amino Acid Metabolism
A second major observation from the NMR analysis was the modulation of amino acid metabolism, particularly in relation to neurotransmission. RP-25 treatment led to significant upregulation of several amino acids involved in neurotransmitter synthesis and synaptic function, including tyrosine, glutamate, glycine, proline, and valine. These metabolites are crucial for the synthesis of neurotransmitters such as dopamine, norepinephrine, and serotonin, all of which play key roles in maintaining normal brain function [55].
Tyrosine and phenylalanine metabolism were particularly affected, as evidenced by the Variable Importance in Projection (VIP) score analysis, which highlighted these pathways as major contributors to the differentiation between RP-25-treated and untreated cells. The increase in tyrosine and its metabolic products suggests that RP-25 may facilitate the biosynthesis of dopamine and norepinephrine, both of which are involved in mood regulation and cognitive function. Tyrosine is also a precursor of thyroid hormones, which are essential for metabolic regulation and neuroprotection [56].
Furthermore, the elevation of glutamate and glycine levels, key excitatory and inhibitory neurotransmitters respectively, suggests that RP-25 may enhance synaptic plasticity and neuronal activity. Glutamate is the principal excitatory neurotransmitter in the brain and plays a central role in synaptic transmission, learning, and memory, while glycine functions as an inhibitory neurotransmitter and modulates excitatory neurotransmission [57]. The increased presence of these metabolites may indicate improved neurotransmission and synaptic function, which are critical for maintaining cognitive and motor functions, particularly in the context of neurodegenerative conditions.
In addition, the observed changes in the exometabolome, specifically the elevation of alanine, serine, and tryptophan, provide further evidence that RP-25 influences brain metabolism. Alanine and serine are involved in the synthesis of proteins and neurotransmitters, while tryptophan is a precursor of serotonin, a neurotransmitter that regulates mood, sleep, and appetite [58]. The increase in these metabolites suggests that RP-25 may not only modulate neurotransmitter synthesis but also influence broader metabolic pathways linked to brain function.
The findings from this study are in line with existing literature on the neuroprotective effects of α-lipoic acid and CoQ10. Both compounds have been shown to exert beneficial effects on brain health through their antioxidant properties and by supporting mitochondrial function in neurons. CoQ10, in particular, is crucial for maintaining mitochondrial bioenergetics and has been shown to alleviate mitochondrial dysfunction in neurodegenerative diseases [59]. The observed upregulation of neurotransmitter-related metabolites such as glutamate, glycine, and tyrosine further supports the hypothesis that RP-25 may aid in restoring neurochemical balance and improving neuronal communication, making it a promising candidate for the treatment of conditions characterized by neurotransmission dysregulation, such as Alzheimer's disease, Parkinson's disease, and fibromyalgia [60,61].
4.4. Oxidative Stress and Redox Balance
The impact of RP-25 on oxidative stress was evident in our metabolomic analysis, which revealed significant alterations in key metabolites involved in cellular redox regulation. Specifically, RP-25 treatment led to a marked reduction in formate levels and an increase in coenzyme A and proline, metabolites that play critical roles in redox homeostasis and cellular function [13,62,63]. Formate is a product of one-carbon metabolism, generated as an intermediate in the folate cycle, and is frequently associated with oxidative stress and impaired mitochondrial function [62]. Elevated formate concentrations have been linked to mitochondrial dysfunction and increased production of reactive oxygen species (ROS), which can damage cellular macromolecules such as lipids, proteins, and DNA [64]. The observed decrease in formate concentration in RP-25-treated cells suggests a potential reduction in mitochondrial oxidative stress, indicating that RP-25 may help mitigate oxidative damage at the cellular level. This is consistent with the known antioxidant properties of α-lipoic acid and CoQ10, both of which have been shown to scavenge ROS and restore redox balance by regenerating other antioxidants [51]. The reduction in formate levels could, therefore, reflect a more favorable redox state, potentially contributing to the overall reduction in oxidative stress observed in RP-25-treated cells.
Additionally, the increase in coenzyme A levels suggests an enhancement of cellular energy metabolism and improved mitochondrial function. Coenzyme A plays a pivotal role in the Krebs cycle (citric acid cycle), which is central to cellular respiration and ATP production [65]. An increase in coenzyme A levels may indicate enhanced mitochondrial efficiency, allowing for better ROS detoxification and more efficient energy production [13]. This result aligns with previous studies demonstrating the role of α-lipoic acid and CoQ10 in supporting mitochondrial bioenergetics and reducing oxidative stress by facilitating proper electron transport in the mitochondrial respiratory chain [51].
Another important finding from the metabolomic data was the increase in proline levels, a key amino acid involved in the synthesis of collagen and other structural proteins, as well as a regulator of redox balance. Proline has antioxidant properties, as it can scavenge ROS and protect cellular structures from oxidative damage. Moreover, proline is involved in the activation of the proline dehydrogenase enzyme, which plays a role in the regulation of cellular oxidative stress responses [66]. The upregulation of proline suggests that RP-25 may enhance cellular antioxidant capacity, possibly by boosting the activity of antioxidant enzymes and promoting the repair of oxidative damage.
Furthermore, pathway enrichment analysis highlighted the involvement of glutathione metabolism in RP-25-treated cells. Glutathione is one of the most abundant and effective intracellular antioxidants, crucial for neutralizing ROS and maintaining cellular redox homeostasis [67]. Dysregulation of glutathione metabolism is often observed in various disease states characterized by oxidative stress, such as fibromyalgia, where chronic pain and fatigue are thought to result, at least in part, from persistent oxidative damage [8,68]. Our data suggest that RP-25 treatment may help restore glutathione homeostasis by modulating the expression of enzymes involved in its synthesis and recycling, thereby enhancing the cell’s capacity to combat oxidative stress. This is further supported by the observed changes in key metabolites such as formate, coenzyme A, and proline, which are interconnected in the cellular antioxidant network.
4.5. Potential Therapeutic Implications for Mitochondrial Diseases and Fibromyalgia
The results of our study suggest that RP-25 holds significant promise as a potential therapeutic agent for improving mitochondrial function, restoring metabolic balance, and reducing oxidative stress in cells affected by mitochondrial dysfunction. Given the crucial role of mitochondrial health in a variety of pathological conditions, including neurodegenerative diseases, cardiovascular disorders, and fibromyalgia, RP-25 may offer a novel approach for the treatment of these diseases. In particular, fibromyalgia has increasingly been recognized as a condition associated with mitochondrial dysfunction, where patients often exhibit impaired mitochondrial respiration and resultant energy deficits, which contribute to the chronic pain, fatigue, and cognitive dysfunction typical of this disorder.
Our recent clinical trials [8,54], which included a comprehensive metabolomic analysis of biological fluids from fibromyalgia patients, further underscores the relevance of mitochondrial dysfunction in fibromyalgia pathophysiology. In this study, we observed distinct alterations in the metabolome of patients with fibromyalgia, particularly in amino acid metabolism related to neurotransmission and energy production. For example, we found decreased serum phenylalanine and increased urinary isoleucine, indicating a disruption in amino acid balance that is integral to neurotransmitter synthesis. Furthermore, we identified changes in key pathways related to energy metabolism, including a reduced capacity for ATP production and the dysregulation of metabolic intermediates involved in mitochondrial function, such as proline and formate. These findings suggest that mitochondrial dysfunction in fibromyalgia may be coupled with an impaired metabolic response to stress, leading to the hallmark symptoms of fatigue and pain [8,54].
In light of these insights, the potential of RP-25 to modulate mitochondrial efficiency and enhance ATP production becomes particularly relevant. The combination of CoQ10, α-lipoic acid, chaga and other key components of RP-25, is known to support mitochondrial bioenergetics by improving electron transport and restoring redox balance, thereby promoting efficient oxidative phosphorylation. This mechanism may help address the mitochondrial dysfunction observed in fibromyalgia patients, as demonstrated by our clinical trial, where interventions aimed at improving energy metabolism showed promising results in alleviating symptoms such as fatigue and muscle pain [8,54]. Moreover, RP-25’s ability to modulate key amino acids, like glutamate and proline, which are involved in both neurotransmission and cellular metabolism, may help restore balance in these metabolic pathways, thus alleviating the neurological and muscular symptoms seen in fibromyalgia.
The integration of NMR-based metabolomics in our study provides a robust methodology for identifying subtle but clinically relevant metabolic changes in mitochondrial diseases and fibromyalgia. By capturing a broad spectrum of metabolic alterations, metabolomic profiling allows for a deeper understanding of the cellular mechanisms underlying these conditions, offering potential biomarkers and therapeutic targets for intervention. For example, the identification of dysregulated metabolites such as phenylalanine, tyrosine, and formate in fibromyalgia patients suggests specific metabolic pathways that may be targeted for therapeutic intervention with RP-25. Additionally, the modulation of redox balance and energy metabolism in RP-25-treated cells may offer a promising strategy for mitigating mitochondrial dysfunction and its associated symptoms.
5. Conclusions
Our study provides compelling evidence that RP-25 significantly influences cellular metabolism, particularly mitochondrial function, energy production, and oxidative stress regulation. By modulating key metabolic pathways involved in energy metabolism, neurotransmission, and redox balance, RP-25 shows potential as a therapeutic strategy for diseases linked to mitochondrial dysfunction, including fibromyalgia. The integration of 1H-NMR-based metabolomics has offered new insights into the complex metabolic changes induced by RP-25, highlighting its potential for identifying biomarkers and therapeutic targets.
A key strength of this study is its comprehensive use of advanced metabolomic techniques, which allowed us to capture a broad spectrum of metabolic alterations and gain valuable mechanistic insights. These findings contribute to the growing understanding of mitochondrial dysfunction in chronic diseases and underscore the potential of RP-25 to address these disturbances.
However, some limitations remain. While our in vitro results are promising, clinical studies are necessary to confirm RP-25's therapeutic effects in human populations. Additionally, while 1H-NMR profiling provided valuable information on metabolic shifts, further research incorporating additional omics approaches may offer a deeper understanding of its molecular impact.
In conclusion, RP-25 shows significant potential as a therapeutic agent for mitochondrial dysfunction-related conditions, including fibromyalgia. Further clinical trials are needed to validate these findings and fully explore its therapeutic potential.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1. Cross-validation performed with 10-fold methods reported the accuracy, R2 and Q2 value of three components.
Author Contributions
Conceptualization, A.M.D., M.D.E. and L.R.; methodology, C.M., R.C., E.N. and M.D.E.; software, C.M., R.C. and E.N.; validation, A.M.D., L.R., and M.D.E.; formal analysis, R.C., E.N. and C.M.; investigation, C.M., R.C., E.N. and M.D.E.; resources, L.R. data curation, L.R., and A.M.D.; writing—original draft preparation M.D.E. C.M. and L.R.; supervision, A.M.D. and L.R.; project administration L.R.; funding acquisition, L.R. All authors have read and agreed to the published version of the manuscript.
Funding
Project funded under the National Recovery and Resilience Plan (NRRP), Mission 4 Component 2 Investment 1.4—Call for tender No. 3138 of 16 December 2021, rectified by Decree n. 3175 of 18 December 2021 of Italian Ministry of University and Research funded by the European Union—NextGenerationEU; Award Number: Project code CN_00000033, Concession Decree No. 1034 of 17 June 2022 adopted by the Italian Ministry of University and Research, CUP: D43C22001260001, Project title “National Biodiversity Future Center—NBFC”.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article and Supplementary Materials.
Acknowledgments
The authors would like to thank the company Laboratoarele Medica srl (Bucharest, Romania) who kindly donated the chaga extract.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Klemmensen, M.M.; Borrowman, S.H.; Pearce, C.; Pyles, B.; & Chandra, B.; Chandra, B. . Mitochondrial dysfunction in neurodegenerative disorders. Neurotherapeutics, 2024, 21, e00292. [Google Scholar] [CrossRef] [PubMed]
- Somasundaram, I.; Jain, S.M.; Blot-Chabaud, M.; Pathak, S.; Banerjee, A.; Rawat, S.; Duttaroy, A.K. Mitochondrial dysfunction and its association with age-related disorders. Frontiers in Physiology, 2024, 15, 1384966. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Li, R.; Zhang, Y.; Zhao, Y.; Liu, Q.; Wang, J.; Su, J. Attenuating mitochondrial dysfunction-derived reactive oxygen species and reducing inflammation: the potential of Daphnetin in the viral pneumonia crisis. Frontiers in Pharmacology, 2024, 15, 1477680. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.N.; Mishra, V.K.; Munalisa, R.; Parveen, F.; Ali, S.F.; Akter, K.; Huang, C.Y. Mechanistic insight of mitochondrial dysfunctions in cardiovascular diseases with potential biomarkers. Molecular & Cellular Toxicology.
- Marino, Y.; Inferrera, F.; D'Amico, R.; Impellizzeri, D.; Cordaro, M.; Siracusa, R.; Di Paola, R. Role of mitochondrial dysfunction and biogenesis in fibromyalgia syndrome: Molecular mechanism in central nervous system. Biochimica et Biophysica Acta (BBA)-Molecular Basis of Disease, 1870. [Google Scholar]
- Inferrera, F.; Marino, Y.; D'Amico, R.; Impellizzeri, D.; Cordaro, M.; Siracusa, R.; Di Paola, R. Impaired mitochondrial quality control in fibromyalgia: Mechanisms involved in skeletal muscle alteration. Archives of Biochemistry and Bio physics, 2024, 758, 110083. [Google Scholar] [CrossRef]
- Macchi, C.; Giachi, A.; Fichtner, I.; Pedretti, S.; Puttini, P.S.; Mitro, N.; Gualtierotti, R. Mitochondrial function in patients affected with fibromyalgia syndrome is impaired and correlates with disease severity. Scientific Reports 2024, 14, 1–9. [Google Scholar] [CrossRef]
- Castaldo, G.; Marino, C.; Atteno, M.; D’Elia, M.; Pagano, I.; Grimaldi, M.; Rastrelli, L. Investigating the Effectiveness of a Carb-Free Oloproteic Diet in Fibromyalgia Treatment. Nutrients, 2024, 16, 1620. [Google Scholar] [CrossRef]
- Mantle, D.; Hargreaves, I.P.; Domingo, J.C.; & Castro-Marrero, J.; Castro-Marrero, J. Mitochondrial dysfunction and coenzyme Q10 supplementation in post-viral fatigue syndrome: an overview. International Journal of Molecular Sciences 2024, 25, 574. [Google Scholar] [CrossRef]
- Shanaida, M.; Lysiuk, R.; Mykhailenko, O.; Hudz, N.; Abdulsalam, A.; Gontova, T.; Bjørklund, G. Alpha-lipoic acid: An antioxidant with anti-aging properties for disease therapy. Current Medicinal Chemistry, 2025, 32, 23–54. [Google Scholar] [CrossRef]
- Tibullo, D.; Li Volti, G.; Giallongo, C.; Grasso, S.; Tomassoni, D.; Anfuso, C.D.; Bramanti, V. Biochemical and clinical relevance of alpha lipoic acid: antioxidant and anti-inflammatory activity, molecular pathways and therapeutic potential. Inflammation Research 2017, 66, 947–959. [Google Scholar] [CrossRef]
- Ern, P.T. Y. , Quan, T.Y.; Yee, F.S.; & Yin, A.C. Y. Therapeutic properties of Inonotus obliquus (Chaga mushroom): a review. Mycology 2024, 15, 144–161. [Google Scholar]
- D’Elia, M.; Marino, C.; Celano, R.; Napolitano, E.; D’Ursi, A.M.; Russo, M.; & Rastrelli, L.; Rastrelli, L. Impact of a Withania somnifera and Bacopa monnieri Formulation on SH-SY5Y Human Neuroblastoma Cells Metabolism Through NMR Metabolomic. Nutrients 2024, 16, 4096. [Google Scholar] [CrossRef] [PubMed]
- Beckonert, O.; Keun, H.C.; Ebbels, T.M.; Bundy, J.; Holmes, E.; Lindon, J.C.; & Nicholson, J.K.; Nicholson, J. K. Metabolic profiling, metabolomic and metabonomic procedures for NMR spectroscopy of urine, plasma, serum and tissue extracts. Nat. protoc. 2007, 2, 2692. [Google Scholar] [CrossRef] [PubMed]
- McKay, R.T. How the 1D-NOESY suppresses solvent signal in metabonomics NMR spectroscopy: An examination of the pulse sequence components and evolution. Concepts Magn. Reson. Part A 2011, 38A, 197–220. [Google Scholar] [CrossRef]
- Jacob, D.; Deborde, C.; Lefebvre, M.; Maucourt, M.; Moing, A. NMRProcFlow: A graphical and interactive tool dedicated 460 to 1D spectra processing for NMR-based metabolomics. Metabolomics 2017, 13, 1–5. [Google Scholar] [CrossRef]
- Xia, J.; Wishart, D.S. Using MetaboAnalyst 3.0 for comprehensive metabolomics data analysis. Current protocols in, 2016; 55. [Google Scholar]
- Kumar, N.; Hoque, M.A.; Sugimoto, M. Robust volcano plot: identification of differential metabolites in the presence of 464 outliers. BMC bioinformatics 2018, 19, 1–11. [Google Scholar] [CrossRef]
- Pang, Z.; Lu, Y.; Zhou, G.; Hui, F.; Xu, L.; Viau, C.; Xia, J. MetaboAnalyst 6.0: towards a unified platform for 466 metabolomics data processing, analysis and interpretation. Nucleic Acids Research.
- Peng, H.; & Shahidi, F.; Shahidi, F. Qualitative analysis of secondary metabolites of chaga mushroom (Inonotus Obliquus): phenolics, fatty acids, and terpenoids. Journal of Food Bioactives 2022, 17. [Google Scholar] [CrossRef]
- Abu-Reidah, I.M.; Critch, A.L.; Manful, C.F.; Rajakaruna, A.; Vidal, N.P.; Pham, T.H.; Thomas, R. Effects of pH and temperature on water under pressurized conditions in the extraction of nutraceuticals from chaga (Inonotus obliquus) mushroom. Antioxidants, 2021, 10, 1322. [Google Scholar] [CrossRef]
- Liao, Z.; Alrosan, M.; Alu'datt, M.H.; & Tan, T.; Tan, T. C 10-hydroxy decanoic acid, trans-10-hydroxy-2-decanoic acid, and sebacic acid: Source, metabolism, and potential health functionalities and nutraceutical applications. J. Food Sci. 2024, 89, 3878–3893. [Google Scholar] [CrossRef]
- Tanaka, R.; Toyoshima, M.; & Yamada, T. ; & Yamada, T. New lanostane-type triterpenoids, inonotsutriols D, and E, from Inonotus obliquus. Phytochemistry Letters 2011, 4, 328–332. [Google Scholar]
- Rakuša, Ž. T. , Kristl, A.; & Roškar, R. Quantification of reduced and oxidized coenzyme Q10 in supplements and medicines by HPLC-UV. Analytical methods, 2020, 12, 2580–2589. [Google Scholar]
- Worley, B.; Halouska, S.; & Powers, R.; Powers, R. Utilities for quantifying separation in PCA/PLS-DA scores plots. Analytical biochemistry, 2013, 433, 102–104. [Google Scholar] [CrossRef] [PubMed]
- Westerhuis, J.A.; Hoefsloot, H.C.; Smit, S.; Vis, D.J.; Smilde, A.K.; van Velzen, E.J.; van Dorsten, F.A. Assessment of PLSDA cross validation. Metabolomics, 2008, 4, 81–89. [Google Scholar] [CrossRef]
- Akarachantachote, N.; Chadcham, S.; & Saithanu, K.; Saithanu, K. Cutoff threshold of variable importance in projection for variable selection. Int J Pure Appl Math, 2014, 94, 307–322. [Google Scholar] [CrossRef]
- Gasmi, A.; Bjørklund, G.; Mujawdiya, P.K.; Semenova, Y.; Piscopo, S.; & Peana, M.; Peana, M. Coenzyme Q10 in aging and disease. Critical Reviews in Food Science and Nutrition, 2024, 64, 3907–3919. [Google Scholar] [CrossRef] [PubMed]
- Haas, R.H. (2019). Mitochondrial dysfunction in aging and diseases of aging. Biology.
- Hou, S.; Tian, Z.; Zhao, D.; Liang, Y.; Dai, S.; Ji, Q.; Yang, Y. Efficacy and Optimal Dose of Coenzyme Q10 Supplementation on Inflammation-Related Biomarkers: A GRADE-Assessed Systematic Review and Updated Meta-Analysis of Randomized Controlled Trials. Molecular Nutrition & Food Research, 2023, 67, 2200800. [Google Scholar]
- Campisi, L.; & La Motta, C.; La Motta, C. The use of the Coenzyme Q10 as a food supplement in the management of fibromyalgia: a critical review. Antioxidants, 2022, 11, 1969. [Google Scholar] [CrossRef]
- Podda, M.; Zollner, T.M.; Grundmann-Kollmann, M.; Thiele, J.J.; Packer, L.; & Kaufmann, R.; Kaufmann, R. Activity of alpha-lipoic acid in the protection against oxidative stress in skin. Oxidants and antioxidants in cutaneous biology, 2000, 29, 43–51. [Google Scholar]
- Tülüce, Y.; Osmanoğlu, D.; Rağbetli, M.Ç.; & Altındağ, F.; Altındağ, F. Protective action of curcumin and alpha-lipoic acid, against experimental ultraviolet-A/B induced dermal-injury in rats. Cell Biochemistry and Biophysics 2024, 1–12. [Google Scholar] [CrossRef]
- Superti, F.; & Russo, R. ; & Russo, R. Alpha-Lipoic Acid: Biological Mechanisms and Health Benefits. Antioxidants 2024, 13, 1228. [Google Scholar]
- Szychowski, K.A.; Rybczyńska-Tkaczyk, K.; Tobiasz, J.; Yelnytska-Stawasz, V.; Pomianek, T.; & Gmiński, J. ; & Gmiński, J. Biological and anticancer properties of Inonotus obliquus extracts. Process Biochemistry, 2018, 73, 180–187. [Google Scholar]
- Kou, R.W.; Han, R.; Gao, Y.Q.; Li, D.; Yin, X.; & Gao, J.M.; Gao, J. M. Anti-neuroinflammatory polyoxygenated lanostanoids from Chaga mushroom Inonotus obliquus. Phytochemistry 2021, 184, 112647. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Bai, M.; Xue, X.B.; Zou, C.X.; Huang, X.X.; & Song, S.J.; Song, S. J. Isolation of chemical compositions as dietary antioxidant supplements and neuroprotectants from Chaga mushroom (Inonotus obliquus). Food Bioscience 2022, 47, 101623. [Google Scholar] [CrossRef]
- Lee, I.K.; Kim, Y.S.; Jang, Y.W.; Jung, J.Y.; & Yun, B.S.; Yun, B. S. New antioxidant polyphenols from the medicinal mushroom Inonotus obliquus. Bioorganic & Medicinal Chemistry Letters, 2007, 17, 6678–6681. [Google Scholar]
- Luo, J.H.; Li, J.; Shen, Z.C.; Lin, X.F.; Chen, A.Q.; Wang, Y.F.; Wang, X.Y. Advances in health-promoting effects of natural polysaccharides: Regulation on Nrf2 antioxidant pathway. Frontiers in Nutrition, 2023, 10, 1102146. [Google Scholar] [CrossRef]
- Angelini, G.; Russo, S.; Carli, F.; Infelise, P.; Panunzi, S.; Bertuzzi, A.; Mingrone, G. Dodecanedioic acid prevents and reverses metabolic-associated liver disease and obesity and ameliorates liver fibrosis in a rodent model of diet-induced obesity. The FASEB Journal, 2024, 38, e70202. [Google Scholar] [CrossRef]
- Francis, G.; Kerem, Z.; Makkar, H.P.; & Becker, K.; Becker, K. The biological action of saponins in animal systems: a review. British journal of Nutrition, 2002, 88, 587–605. [Google Scholar] [CrossRef]
- Islam, M.N.; Rauf, A.; Fahad, F.I.; Emran, T.B.; Mitra, S.; Olatunde, A.; Mubarak, M.S. Superoxide dismutase: an updated review on its health benefits and industrial applications. Critical Reviews in Food Science and Nutrition, 2022, 62, 7282–7300. [Google Scholar] [CrossRef]
- Lee, K.H.; & Jeong, D.; Jeong, D. Bimodal actions of selenium essential for antioxidant and toxic pro-oxidant activities: the selenium paradox. Molecular medicine reports, 2012, 5, 299–304. [Google Scholar]
- An, Y.; Li, S.; Huang, X.; Chen, X.; Shan, H.; & Zhang, M.; Zhang, M. The role of copper homeostasis in brain disease. International journal of molecular sciences, 2022, 23, 13850. [Google Scholar] [CrossRef]
- Azadmanesh, J.; & Borgstahl, G.E.; Borgstahl, G. E. A review of the catalytic mechanism of human manganese superoxide dismutase. Antioxidants, 2018, 7, 25. [Google Scholar] [CrossRef]
- Bresciani, G.; da Cruz, I.B. M. , & González-Gallego, J. Manganese superoxide dismutase and oxidative stress modulation. Advances in clinical chemistry, 2015, 68, 87–130. [Google Scholar] [PubMed]
- Pincemail, J.; & Meziane, S.; Meziane, S. On the potential role of the antioxidant couple vitamin E/selenium taken by the oral route in skin and hair health. Antioxidants, 2022, 11, 2270. [Google Scholar] [CrossRef] [PubMed]
- Boo, Y.C. Mechanistic basis and clinical evidence for the applications of nicotinamide (niacinamide) to control skin aging and pigmentation. Antioxidants, 2021, 10, 1315. [Google Scholar] [CrossRef] [PubMed]
- Alalykina, E.S.; Sergeeva, T.N.; Ananyan, M.A.; Cherenkov, I.A.; & Sergeev, V.G.; Sergeev, V. G. A Water-soluble Form of Dihydroquercetin Reduces LPS-induced Astrogliosis, Vascular Remodeling, and mRNA VEGF-A Levels in the Substantia Nigra of Aged Rats. Journal of Neuroscience and Neurological Disorders, 2024, 8, 014–019. [Google Scholar]
- Liu, Z.; Qiu, D.; Yang, T.; Su, J.; Liu, C.; Su, X.; Zhang, S. Research progress of dihydroquercetin in the treatment of skin diseases. Molecules, 2023, 28, 6989. [Google Scholar] [CrossRef]
- Abadi, A.; Crane, J.D.; Ogborn, D.; Hettinga, B.; Akhtar, M.; Stokl, A.; Tarnopolsky, M. Supplementation with α-lipoic acid, CoQ10, and vitamin E augments running performance and mitochondrial function in female mice. PloS one, 6072. [Google Scholar]
- Muoio, D.M. Metabolic inflexibility: when mitochondrial indecision leads to metabolic gridlock. Cell, 2014, 159, 1253–1262. [Google Scholar] [CrossRef]
- Marino, Y.; Inferrera, F.; D'Amico, R.; Impellizzeri, D.; Cordaro, M.; Siracusa, R.; Di Paola, R. Role of mitochondrial dysfunction and biogenesis in fibromyalgia syndrome: Molecular mechanism in central nervous system. Biochimica et Biophysica Acta (BBA)-Molecular Basis of Disease, 1870. [Google Scholar]
- Castaldo, G.; Marino, C.; D’Elia, M.; Grimaldi, M.; Napolitano, E.; D’Ursi, A.M.; & Rastrelli, L.; Rastrelli, L. The Effectiveness of the Low-Glycemic and Insulinemic (LOGI) Regimen in Maintaining the Benefits of the VLCKD in Fibromyalgia Patients. Nutrients, 2024, 16, 4161. [Google Scholar] [CrossRef]
- Bon, L.I.; Maksimovich, N.Y.; & Burak, I.N. ; & Burak, I.N. Amino Acids that Play an Important Role in the Functioning of the Nervous System Review. Clinical Trails and Clinical Research.
- Schmitt, H.P. Neuro-modulation, aminergic neuro-disinhibition and neuro-degeneration: Draft of a comprehensive theory for Alzheimer disease. Medical hypotheses, 2005, 65, 1106–1119. [Google Scholar] [CrossRef]
- Gasmi, A.; Nasreen, A.; Menzel, A.; Gasmi Benahmed, A.; Pivina, L.; Noor, S.; Bjørklund, G. Neurotransmitters regulation and food intake: The role of dietary sources in neurotransmission. Molecules, 2022, 28, 210. [Google Scholar] [CrossRef]
- He, W.; & Wu, G. ; & Wu, G. Metabolism of amino acids in the brain and their roles in regulating food intake. Amino Acids in Nutrition and Health: Amino acids in systems function and health.
- Orsucci, D.; Mancuso, M.; Ienco, E.C.; LoGerfo, A.; & Siciliano, G.; Siciliano, G. Targeting mitochondrial dysfunction and neurodegeneration by means of coenzyme Q10 and its analogues. Current medicinal chemistry, 2011, 18, 4053–4064. [Google Scholar] [CrossRef]
- Miller, E.; Markiewicz, L.; Kabzinski, J.; Odrobina, D.; & Majsterek, I.; Majsterek, I. Potential of redox therapies in neurodegenerative disorders. Front Biosci, 2017, 9, 214–34. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Ahuja, A.; & Pathak, S.; Pathak, S. Potential Role of Oxidative Stress in the Pathophysiology of Neurodegenerative Disorders. Combinatorial Chemistry & High Throughput Screening 2024, 27, 2043–2061. [Google Scholar]
- Hughes, E.R.; Winter, M.G.; Duerkop, B.A.; Spiga, L.; de Carvalho, T.F.; Zhu, W.; Winter, S.E. Microbial respiration and formate oxidation as metabolic signatures of inflammation-associated dysbiosis. Cell host & microbe, 2017, 21, 208–219. [Google Scholar]
- Phang, J.M. Proline metabolism in cell regulation and cancer biology: recent advances and hypotheses. Antioxidants & redox signaling, 2019, 30, 635–649. [Google Scholar]
- Pietzke, M.; Meiser, J.; & Vazquez, A.; Vazquez, A. Formate metabolism in health and disease. Molecular metabolism, 2020, 33, 23–37. [Google Scholar] [CrossRef]
- Szrok-Jurga, S.; Turyn, J.; Hebanowska, A.; Swierczynski, J.; Czumaj, A.; Sledzinski, T.; & Stelmanska, E.; Stelmanska, E. The role of Acyl-CoA β-oxidation in brain metabolism and neurodegenerative diseases. International Journal of Molecular Sciences, 2023, 24, 13977. [Google Scholar] [CrossRef]
- Liang, X.; Zhang, L.; Natarajan, S.K.; & Becker, D.F.; Becker, D. F. Proline mechanisms of stress survival. Antioxidants & redox signaling, 2013, 19, 998–1011. [Google Scholar]
- Lushchak, V.I. Glutathione homeostasis and functions: potential targets for medical interventions. Journal of amino acids, 7368. [Google Scholar]
- Johnson, W.M.; Wilson-Delfosse, A.L.; & Mieyal, J.; Mieyal, J. JDysregulation of glutathione homeostasis in neurodegenerative diseases. Nutrients, 2012, 4, 1399–1440. [Google Scholar] [CrossRef]
Figure 1.
UHPLC−HRMS profile of ST-65 Formulation.
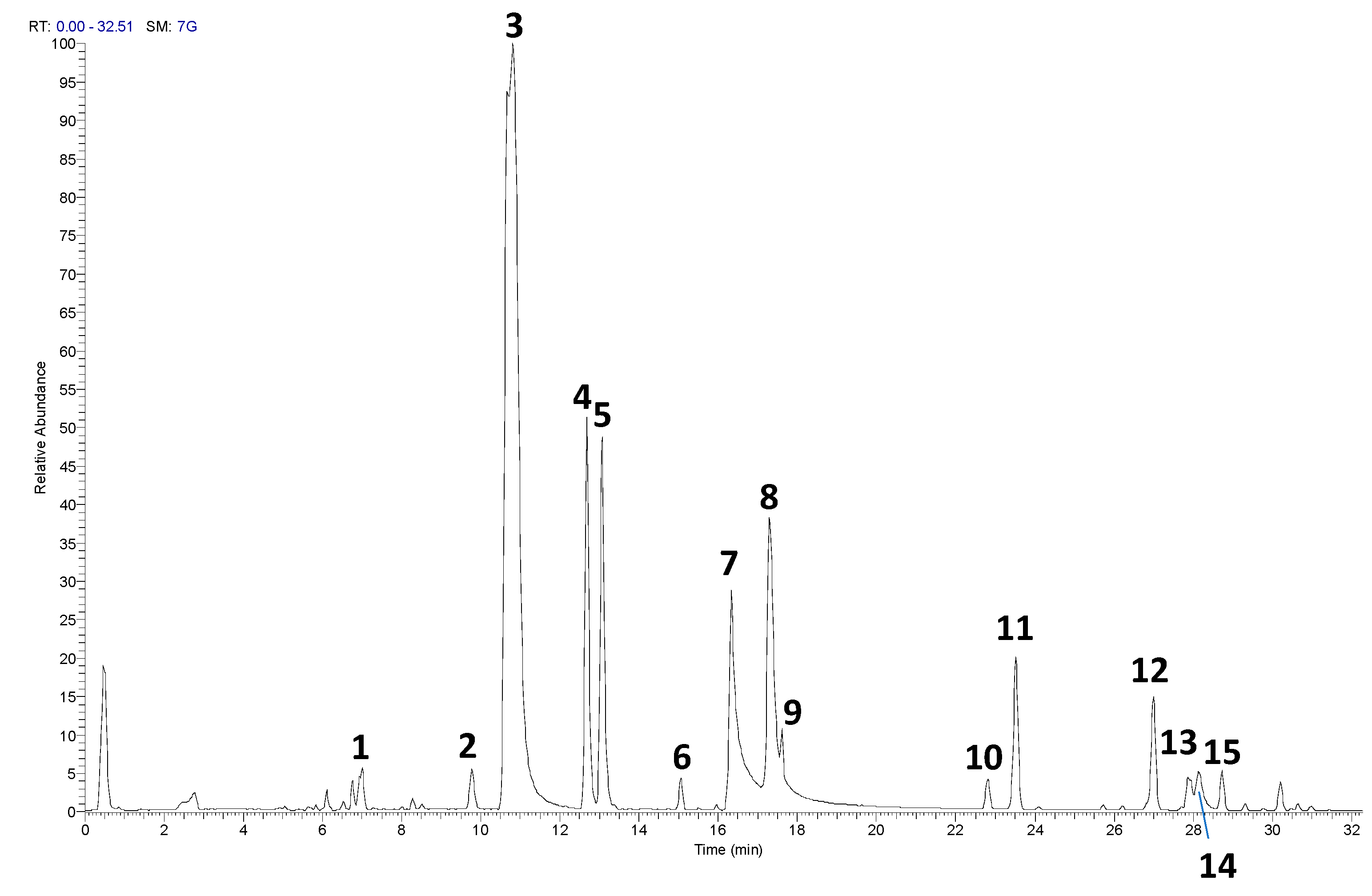
Figure 2.
Histogram representing the percentage of viable cells 24 hours after treatment with various concentrations of RP-25 (0.80–200 μg/mL). Cell viability was calculated as the percentage of viable cells in treated cultures relative to untreated controls. Data are presented as mean ± standard deviation (SD) from three independent experiments. The statistical significancy was evaluated using the one-way analysis of variance (ANOVA), followed by Bonferroni’s test, with GraphPad Prism 8.0 software (San Diego, CA, USA). Significance was assumed at p < 0.05. The statistical test showed the viability of cell exposed to each concentration of RP-25 is not significantly different from the control.
Figure 2.
Histogram representing the percentage of viable cells 24 hours after treatment with various concentrations of RP-25 (0.80–200 μg/mL). Cell viability was calculated as the percentage of viable cells in treated cultures relative to untreated controls. Data are presented as mean ± standard deviation (SD) from three independent experiments. The statistical significancy was evaluated using the one-way analysis of variance (ANOVA), followed by Bonferroni’s test, with GraphPad Prism 8.0 software (San Diego, CA, USA). Significance was assumed at p < 0.05. The statistical test showed the viability of cell exposed to each concentration of RP-25 is not significantly different from the control.
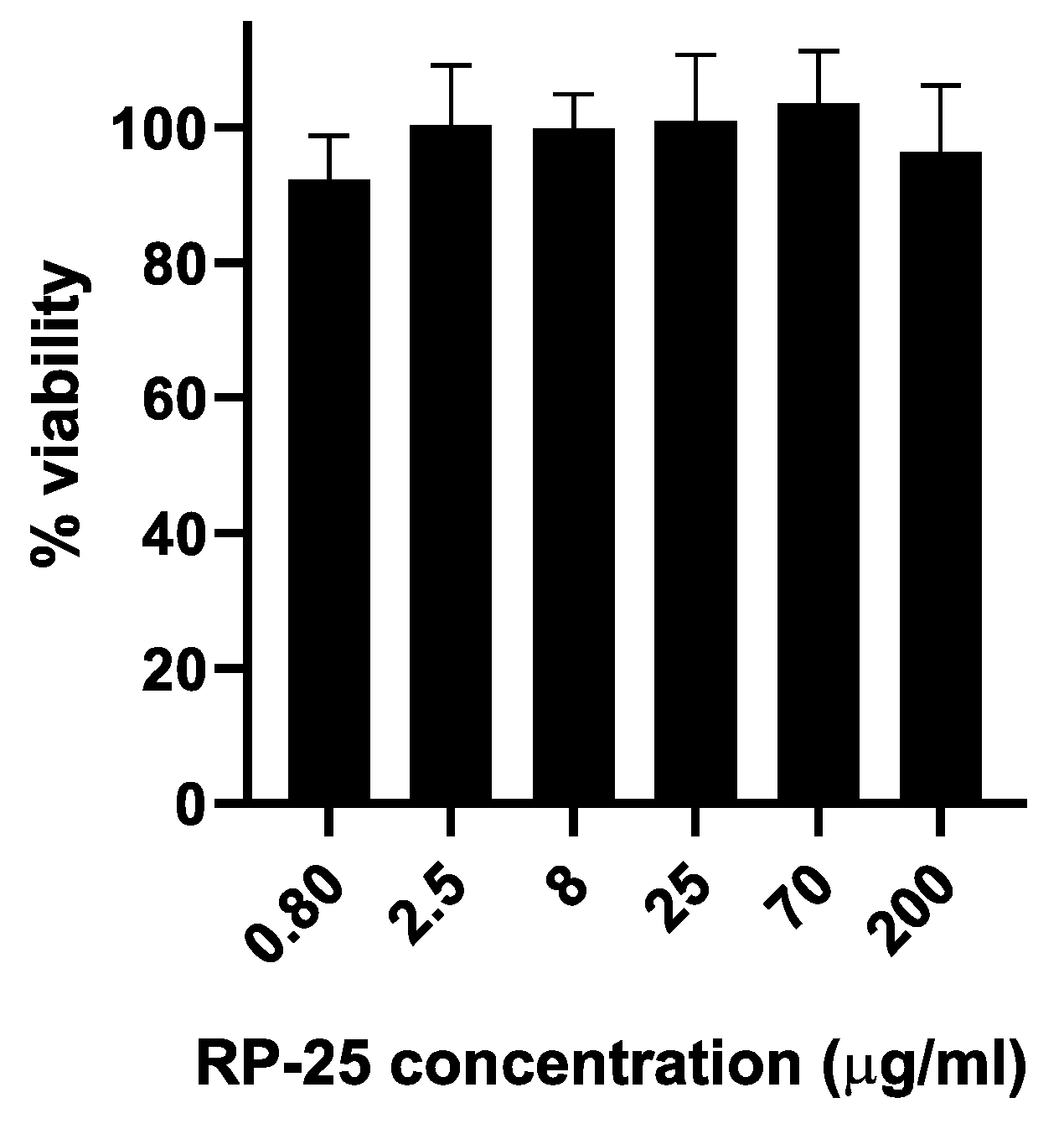
Figure 3.
1D-NMR spectra of the endometabolome (A) and exometabolome (B) of SH-SY5Y cell extracts, showing 42 and 39 metabolites, respectively.
Figure 3.
1D-NMR spectra of the endometabolome (A) and exometabolome (B) of SH-SY5Y cell extracts, showing 42 and 39 metabolites, respectively.
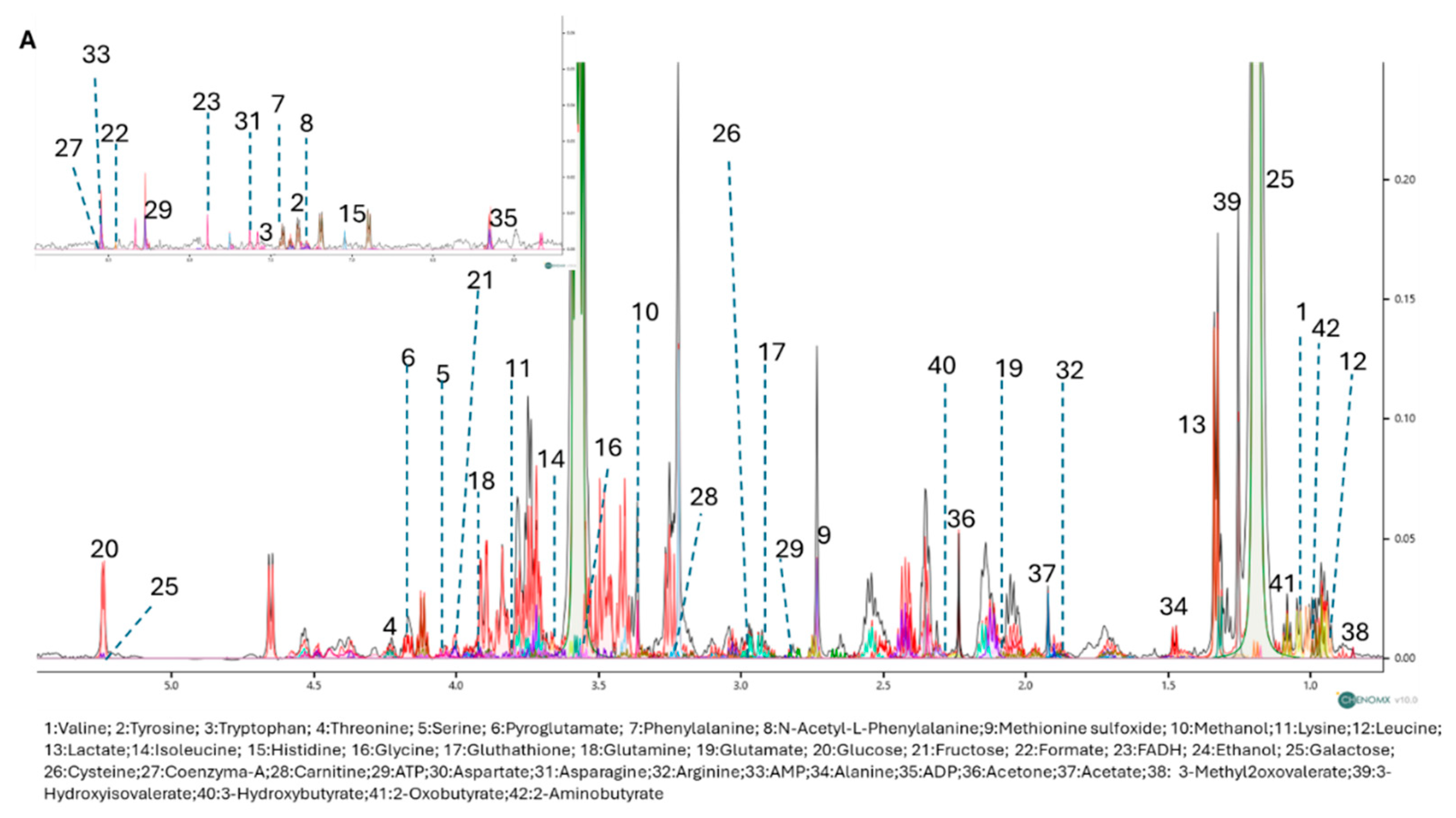
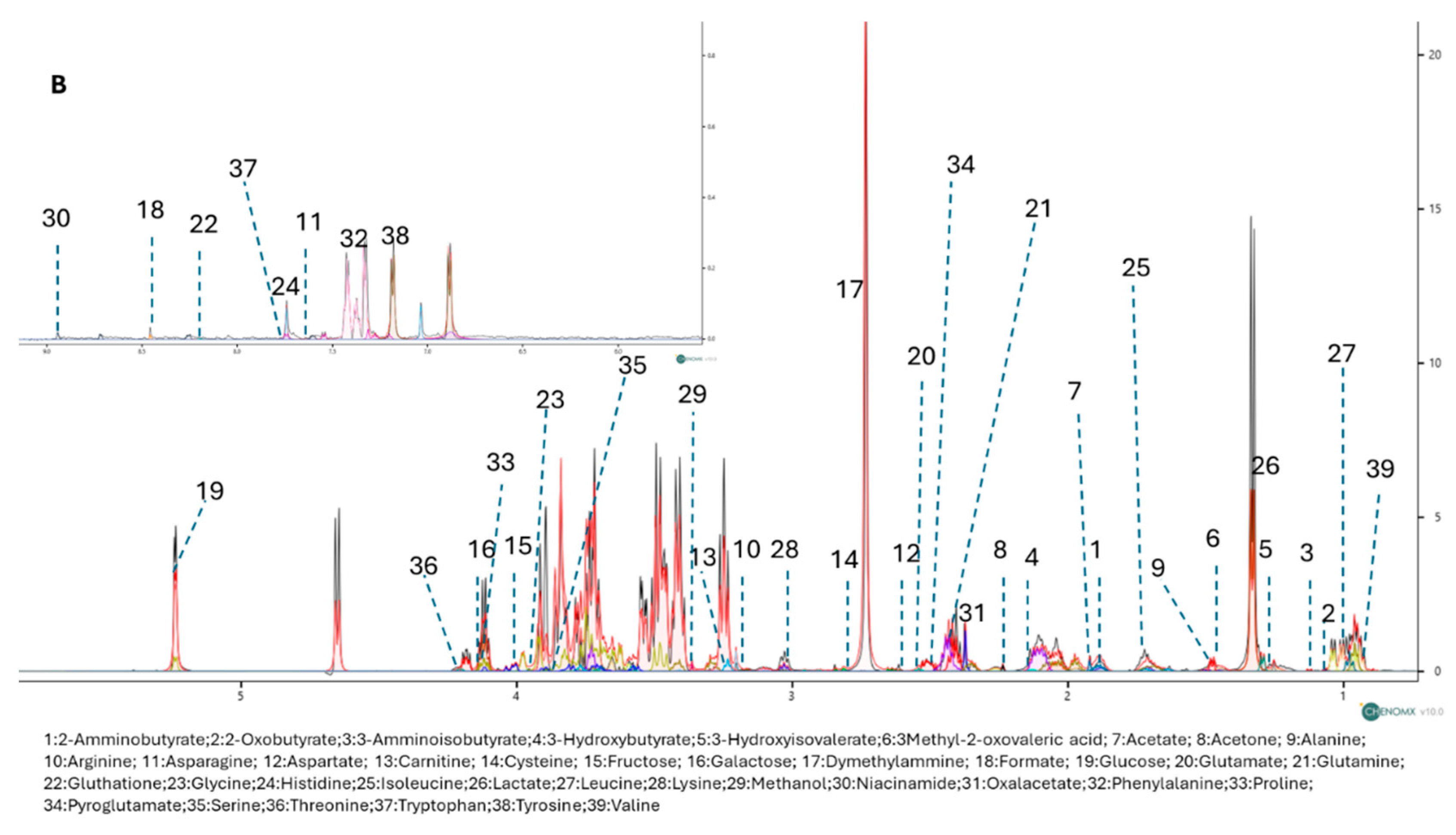
Figure 4.
PLS-DA score plots for the endometabolome (A) and exometabolome (B) of RP-25-treated (green) and untreated (red) cells. VIP scores (C, D) highlight the metabolites responsible for the clustering observed in both endometabolome and exometabolome. Volcano plots (E, F) show metabolic changes, with red points indicating upregulated metabolites and blue points indicating downregulated ones.
Figure 4.
PLS-DA score plots for the endometabolome (A) and exometabolome (B) of RP-25-treated (green) and untreated (red) cells. VIP scores (C, D) highlight the metabolites responsible for the clustering observed in both endometabolome and exometabolome. Volcano plots (E, F) show metabolic changes, with red points indicating upregulated metabolites and blue points indicating downregulated ones.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



