
Submitted:
13 October 2024
Posted:
16 October 2024
You are already at the latest version
Abstract
Polyamines (PA), such as putrescine, spermine, and spermidine, are bioactive molecules that play a vital role in plant responses to stresses. Although they are frequently applied to achieve higher levels of stress tolerance in plants, their function in seed biology is still not fully under-stood. Only in a few papers PA were described as seed priming agents, but most of the data re-port only the physiological and biochemical PA effects, and only a few of them concern the molecular mechanisms. In this review, we summarized PA seed priming effects on germination, seedling establishment, and young plant response to abiotic stresses, and tried to draw a general scheme of PA action during early developmental plant stages.
Keywords:
abiotic stress
; chilling stress
; priming memory
; rice (Oryza sativa L.)
; salt stress
; seed biology
; seed germination
1. Seed Priming as a Strategy to Overcome the Negative Effect of Stresses on Plants
Seed biology, and in particular the process of germination, has been an area of interest to humanity since ancient times. This was not so much out of curiosity as a pragmatic need to cultivate plants to satisfy hunger. The earliest records indicating the need for discernment and the first hints describing the possibilities of regulating the process of seed germination can be found in the works of Theophrastus, who could be called the father of seed physiology [1]. In his extant botanical writings, History of Plants and Causes of Plants, Theophrastus presented his observations and conclusions, often indicating areas that require more in-depth knowledge to understand the nature of a given phenomenon. In the area of seed biology, he drew attention to, among other things, the state of being alive to dry seeds even without moisture and energy. He stated that the life span of a dry seed depends on both the plant species and the storage conditions and proposed that seed longevity (short or long) is an inherent characteristic of the plant species and can vary from less than one year to four years under common storage conditions [2]. Theophrastus also recommended the pre-soaking of cucumber seeds in milk or water to germinate earlier and vigorously [1,3], which can be considered the first written mention of pre-sowing seed treatment referred to as hardening or advancing and nowadays called seed priming. This approach was originally used to equalize emergence and mobilize poorly germinating seeds, and it was confirmed and advised along with others by Plinius Secundus and Charles Darwin [4]. The term “seed priming” was coined by Heydecker in 1973 [5].
Seed priming is understood as a pre-sowing treatment aimed at improving seed performance by regulating the germination process. It involves hydrating seeds to a point where germination processes begin, but radicle emergence does not occur. This controlled hydration is then followed by re-drying the seeds, so they can be handled and stored like dry seeds but with enhanced vigor and better germination performance [6,7,8,9]. Seed priming is distinguished from pre-soaking due to subsequent seed drying back into initial water content, which allows storing and transportation. Improving overall seed vigor and performance under suboptimal or stressful conditions quickly became a goal of seed priming, alongside synchronizing seed emergence and enhancing the germination of poorly performing seeds [10]. Primed seeds promote more consistent germination by activating enzymes, repairing cells, synthesizing proteins, and enhancing antioxidant defenses in comparison to seeds that have not been primed. Seed priming also enhances the accumulation of osmolytes (including proline (Pro), glycine-betaine (GB)) by modifying metabolic pathways' response for their biosynthesis. Various types of priming treatments, such as water-based, PGR-based (plant growth regulator), osmotic solution-based, and chemical-based methods, are commonly utilized to improve stress tolerance in crop plants. Despite sharing the common feature of partial pre-hydration and early activation of germination processes in seeds, the effectiveness of these techniques varies depending on the plant species being treated and the specific priming method employed [9,11]. Seed priming is now successfully used both in commercial vegetable production [12] and in basic research as a model of pre-germinative metabolism responding to seed priming and stress conditions [13] to increase the tolerance of a wide range of stress factors, including abiotic stresses like drought [11,14], salt [15,16], heat [17], as well as biotic stresses [18].
The priming process refers to a physiological and biochemical process where plants or seeds are exposed to a stimulus or treatment that enhances their ability to respond more effectively to subsequent stress or stimuli. This process involves exposing plants or seeds to a mild or sublethal stress or treatment, which activates defense mechanisms, metabolic pathways, and gene expression without causing significant damage. As a result, when a plant or seed encounters more severe stress later, it is "primed" to respond more effectively, leading to improved tolerance, resistance, or recovery [19,20,21]. The concept of priming memory as one of the mechanisms explaining the positive effect on further plant development and growth is commonly accepted [13,14,22]. Chen and Arora [23] proposed a hypothesis explaining the enhanced germination and stress tolerance observed in seeds that undergo priming. According to their hypothesis, priming-induced stress tolerance is achieved through two main strategies. The first strategy involves the activation of processes associated with germination, which facilitates the transition of dormant dry seeds into an active germinating state, thereby improving germination. The second strategy, referred to as stress imprinting or cross-tolerance, involves exposing seeds to pre-germination stress through imbibition-drying cycles and the use of various priming agents. The increased stress tolerance observed during the germination of primed seeds may result from a "priming memory". This priming-related stress response could be due to epigenetic mechanisms, such as DNA methylation and histone modification. Furthermore, the primed state can be passed on to the progeny, enhancing its stress resistance. This transgenerational priming is evident in plants grown from the seeds of primed parents, which retain the priming memory and are therefore better equipped to respond swiftly and effectively to stress [24]. Inheritance of epigenetic changes induced by stress factors leads to the need for more careful observation of visible changes, especially in the context of pronounced climatic changes and the resulting stress factors' impact on plants.
2. Polyamines as Regulatory Molecules That Impact Plant Stress Responses
Polyamines (PA) are small organic polycationic molecules that are known as protective molecules. Figure 1 schematically presents the key biochemical pathways associated with PA metabolism. The PA biosynthesis pathways described in plants derive their synthesis from the amino acids, mainly from arginine, ornithine and methionine. The degradation pathways are linked to the activity of diamine oxidase (DAO) and polyamine oxidase (PAO) [25,26]. PA are hubs promoting tolerance mechanisms by a complex signaling system and having a vital role in several physiological and metabolic processes, including antioxidative mechanisms, photosynthetic pigment protection, and hormonal interplay [25,27]. The chemical properties of PA make them likely to interact primarily with nucleic acids and phospholipids. The interaction with RNA, in particular, has led to the exploration of a potential link between PA and translation, which has emerged as one of the more established mechanisms by which PA affect cellular and developmental processes and involves various levels of control – from ensuring the proper assembly of ribosomes to broadly stimulating translation, enhancing translation accuracy, and promoting the translation of specific open reading frames (ORF) by increasing ribosomal frameshifting [28]. Based on this molecular mechanism of action, exogenously applied PA exert some physiological responses widely observed in plants, which are discussed as favorable under stress conditions. The most frequently used PA in seed priming are spermidine (Spd), putrescine (Put), and spermine (Spm).
While PA may not directly scavenge reactive oxygen species (ROS), they participate in the activation of the antioxidant machinery, both enzymatic and non-enzymatic, under a variety of stresses. Exogenous Spm enhanced the water deficit tolerance of orchid (Anoectochilus roxburghii) by increasing the activation of catalase (CAT) and decreasing hydrogen peroxide (H2O2) and malondialdehyde (MDA) levels of accumulation [29]. CAT is an antioxidative enzyme that catalyzes the decomposition of H2O2 to water and oxygen, whereas MDA can be considered a marker of membrane damage due to lipid peroxidation caused by ROS. Exogenous Put also enhanced drought stress tolerance in the seedlings of grapevine (Vitis vinifera L., Cabernet Sauvignon) by reducing H2O2 and O2ׄ− levels and increasing the activity of antioxidative enzymes, such as superoxide dismutase (SOD), peroxidase (POD), and CAT, as well as the contents of non-enzymatic antioxidants, like ascorbic acid (AsA) and glutathione (GSH) [30]. Similarly, pretreatment of barley (Hordeum vulgare L.) seeds with Spm improved their drought stress tolerance which was manifested by reduced MDA content and enhanced CAT activity [31]. Spm reduced H2O2 and MDA levels in barley leaves also under lead (Pb) stress [32]. In aluminum (Al) stress, the application of Spm lowered O2ׄ−, H2O2, and MDA content and stimulated SOD, POD, CAT, and ascorbate peroxidase (APX) activity in chloroplasts of rice (Oryza sativa L.) [33]. Moreover, in salt-sensitive wild type of tomato (Solanum lycopersicum L.) Spd improved salt tolerance which was manifested by reduced H2O2 and MDA content, and lower electrolyte leakage (EL), which could be discussed as a mechanism of membrane protection [34]. The tolerance to salt combined with paraquat (herbicide) was similarly improved in tomato plants also by Spm through lower H2O2 and MDA levels and decreased SOD activity [35].
The improved stress tolerance is also exerted by protecting photosynthetic pigments and structures. The photosynthetic rate was significantly higher in the Vitis vinifera seedlings treated with Put compared with the untreated control [30]. The study on Oryza sativa chloroplasts showed that exogenous Spd effectively reversed Al-induced chlorophyll losses and protected photosystem II (PSII) reaction centers and photosynthetic electron transport chain by stabilizing PSII thus improving photosynthetic performance and preserving the integrity and function of PSII by reducing the oxidative damage caused by Al toxicity [33]. Exogenous Spd alleviated the drought stress-induced inhibition of plant growth by improving photosynthetic performance in maize (Zea mays L.) seedlings, particularly by enhancing photochemical efficiency and the synthesis of ATP and maintaining the structural stability of PSII and stimulating photochemical quenching in light-harvesting complex II (LHCII) [36]. Spd and Spm also mitigate the destructive impacts of high temperature in lettuce (Lactuca sativa L.) seedlings [37] and salinity in rapeseed (Brassica napus L.) seedlings [38] by enhancing photosynthetic capacity.
Polyamines also influence the level of plant hormones modulating their signaling pathways, which may promote stress tolerance in particular species. The crosstalk between PA and phytohormones in plant response to abiotic stress is widely discussed and commonly accepted [39]. In maize seedlings exposed to drought, exogenous Spd increased indoleacetic acid (IAA), zeatin riboside (ZR), and gibberellin A3 (GA3) while decreased salicylic acid (SA) and jasmonate (JA) content [36]. Exogenous Spd significantly increased abscisic acid (ABA) and IAA content but reduced ethylene (ET) emission under hypoxia stress caused by flooding in bamboo (Phyllostachys praecox) seedlings [40]. Although PA affect many processes in plant response to stresses, only some of the regulatory mechanisms were confirmed on the gene expression level. Most of the confirmed evidence on regulation of gene expression concerns arginine decarboxylase (ADC), which catalyzes the conversion of arginine to agmatine (precursor of Put) in plants [41], archaea, and some bacteria [42]. ADC plays a crucial role in the biosynthesis of Pro and PA [41,42]. In Arabidopsis thaliana, there are two genes encoding ADC, ADC1 and ADC2. ADC1 shows a constitutive expression and is also responsive to cold and bacterial pathogen infection, while ADC2 is upregulated by abiotic stresses, such as salt, drought, cold, and wounding [43,44]. The ADC1, which is induced primarily in response to cold is located in the endoplasmic reticulum, where together with N-acetyltransferase1 provides a pathway for the synthesis of N-acetylputrescine in Arabidopsis thaliana [45]. The stimulation of ADC gene expression upon stress is widely noted in plants’ response and was proposed to serve as a hub connecting diverse functions of different plant hormones with Put [43].
3. Polyamine Seed Priming as a Way to Modify Plant Metabolism and Improve Stress Resistance
One of the seed priming methods is based on seed treatment with a solution composed of water and specific bioactive components. The most frequently used molecules in seed priming belong to compounds accumulated under stress exposure and phytohormones. Here, we focus on specific responses in plants grown from PA-primed seeds, which promote stress tolerance. The most often studied PA used for seed priming is Spd followed by Spm. Primed seeds were exposed to different stresses, however, in the study with PA, salt stress was the most frequently investigated followed by chilling stress. The species, which is the most frequently studied to observe the effect of seed priming with PA on stress resistance is Oryza sativa (Table 1). In this review, we have focused only on those studies in which the methodology explicitly states that seeds, after priming, were dried back to their initial water content. Most of the research focuses on seed germination behavior, seedling establishment, growth parameters, and basic biochemical features, such as changes in the content of photosynthetic pigments, carotenoids, and flavonoids, osmolyte accumulation, oxidative stress markers such as antioxidant enzyme activity, the presence of ROS, the accumulation of MDA, and the level of EL under stress conditions. Only a few studies focus on molecular changes, such as the accumulation of specific proteins or the activation and inhibition of particular gene expressions. Currently, there is no experimental data specifically concerning the molecular mechanisms by which the PA seed priming causes the observed effects.
The first and most significant outcome of PA seed priming is improved germination along with enhanced seedling morphology and growth parameters. All priming methods have been shown to increase both the rate and speed of germination under stress conditions. In the process of seedling establishment and stress response, one crucial factor is photosynthesis efficiency. Increased content of photosynthetic pigments, such as chlorophylls and carotenoids, was observed under salt stress in Brassica napus seedlings grown from Spd- and Spm-primed seeds [46], as well as in Cucurbita pepo seedlings from Put-primed seeds [47] and Spd-primed Oryza sativa seedlings [48]. In some studies also reduced chlorophyll degradation in Spd-primed Oryza sativa under chromium [49,50] and salt stress [51] was observed. Furthermore, increased expression of the gene encoding RuBisCO small subunit (RbcS) was noted in Spd- and Spm-primed Oryza sativa seeds under salt stress [52,53].
Abiotic stresses often disrupt plant growth and productivity by disturbing cellular homeostasis through the overproduction of ROS [54,55]. Seed priming induces antioxidant mechanisms in both seeds and seedlings, particularly under stress conditions [9,10]. Reduced ROS levels were observed in cabbage (Brassica oleracea L.) under chilling stress due to Spd seed priming [56], and in Oryza sativa exposed to heat [57] or chromium stress [49,50], as well as in Oryza sativa under salt stress following Spd priming [51]. Most studies report decreased ROS levels; however, an increase in H₂O₂ content was observed in silver maple (Acer saccharinum) exposed to desiccation after Spd seed priming [58]. The authors suggested that the moderate increase in H₂O₂ may play a role in stress signaling, contributing to enhanced stress tolerance. Additionally, an increase in CAT activity was observed. Enhanced activity of antioxidant enzymes, such as CAT, SOD, glutathione peroxidase (GPX), and APX, was reported in several studies [46,48,49,50,51,56,57,58,59,60,61,62,63,64]. In Oryza sativa, altered isozyme profiles of CAT, SOD, GPX, and APX were noted following Spd seed priming under salt stress [51]. In other studies performed on Oryza sativa, increased expression of genes encoding CAT, SOD, APX, and glutathione reductase (GR) was observed under salt stress in Spd- and Spm-primed seeds [52,53].
Oxidative stress also leads to lipid peroxidation in membranes, typically measured by MDA levels and membrane disruption, indicated by EL. Both parameters increase under a variety of stress conditions. However, in plants grown from primed seeds, the levels of damage were lower, as evidenced by reduced MDA accumulation and weaker EL. Lower MDA levels, indicating reduced lipid peroxidation, were observed after Spd seed priming in Acer saccharinum under desiccation [58] and in Cucurbita pepo [47], tomato (Solanum lycopersicum L.) [60], and tobacco (Nicotiana tabacum L.) [59] in response to salt and chilling stresses. Similarly, Spd-primed Oryza sativa seeds showed reduced MDA levels under salt stress [51,65], heat stress [57], chromium toxicity [49,50], chilling stress [61], and drought stress [63].
Under stresses that cause osmotic imbalance, such as salt, water (drought or flooding), temperature (chilling or heating), or heavy metal, the production of osmotically active substances is stimulated. Higher levels of soluble sugars were noted in Spd-primed Oryza sativa under various stress conditions [49,50,51,57,61,63], likely due to increased activity of α-amylase [57,62,63] and α-glucosidase [57], as well as enhanced expression of their respective genes [57]. Compounds, such as Pro and GB, also are accumulated under osmotic stress. Increased Pro levels were observed in Brassica napus [46], quinoa (Chenopodium quinoa) [66], Oryza sativa [61], and Solanum lycopersicum [60]. Also, GB was accumulated more strongly in Spd-primed Oryza. sativa under chilling stress [62], and the upregulation of genes encoding enzymes involved in Pro and GB biosynthesis was also noted under salt stress [52,53].
Polyamine seed priming also affects endogenous PA metabolism, leading to changes in PA content and the expression of PA metabolism genes [52,53,56,62,66]. For instance, in Brassica oleracea, seed treatment with Spd increased endogenous Spd levels, while Put levels decreased under chilling stress [56]. In Nicotiana tabacum seedlings, Put, Spd, and Spm levels were higher following Put seed priming in response to chilling [59]. Increased activity of enzymes involved in PA biosynthesis, such as ADC, ornithine decarboxylase (ODC), and S-adenosylmethionine decarboxylase (SAMDC) (Figure 1), was observed in Spd-primed Oryza sativa under chilling stress [62]. The Spd-priming promoted the accumulation of endogenous Spm content by upregulating spermine synthase genes SPMS1 and SPMS2 in Oryza sativa under chilling stress [61]. Modulation of gene expression involved in PA biosynthesis (SAMDC, SPDS, SPMS) and catabolism (DAO, PAO) was studied in Spd- and Spm-primed Oryza sativa under salt stress [52,53], showing increased endogenous Put (except in Spm-primed samples), Spd, and Spm in a salt-sensitive Oryza sativa cultivar, and decreased levels of these PA in a salt-tolerant cultivar [53]. Oryza sativa seed priming with Spm and Spd led to a decrease in Spm and Spd content in salt-stressed seedlings, which was linked to induced expression of the PAO gene responsible for PA degradation, even upon induction of genes involved in Spd and Spm biosynthesis, viz., SAMDC, SPDS, SPMS via PA priming [52].
The above-mentioned biochemical and molecular changes correspond with alterations in phytohormone levels, which may represent one mechanism for molecular control, triggering different types of stress responses [67]. Only a few studies have focused on the interaction between PA seed priming and plant hormones. Increased GA and ET levels, alongside decreased ABA content, were observed in Brassica oleracea under chilling stress after Spd seed priming [56]. In Oryza sativa under chromium stress, lower ABA, and higher SA levels were reported following Spd seed priming [49]. However, in response to salt stress, enhanced expression of genes involved in ABA biosynthesis and ABA-inducible transcription factors was observed in Oryza sativa, stimulated by Spd and Spm seed priming [52].
The modulation of the physiological and biochemical response of plants to abiotic stresses reflects alteration in gene expression. Priming with both Spm and Spd enhanced the expression of genes encoding antioxidants and enzymes involved in osmolytes biosynthesis pathways, ABA biosynthesis enzyme, and ABA-inducible, stress-responsive genes including transcription factors and genes of late embryogenesis abundant (LEA) proteins [53]. By accumulating higher levels of endogenous Spm and Spd, increasing the (Spm+Spd)/Put ratio, and boosting the expression of SAMDC, SPDS, SPMS, and DAO, the salt-sensitive Oryza sativa cultivar IR-64 demonstrated improved adaptability under salt stress after priming with both above-mentioned PA. However, Spm had a stronger effect [53]. Enhanced induction of transcription factors (TRAB-1 and WRKY-71), LEA genes, and ABA biosynthetic enzyme (NCED3) gene in Oryza sativa seedlings indicates that Spm and Spd seed priming regulated the adaptive mechanism by enhancing transcription of the regulatory genes [53]. Induction of the expression of genes from PA biosynthesis pathways together with induction of genes for enzymes of PA degradation pathways led to a decrease in PA accumulation under stress in Oryza sativa [52]. However, although some transcriptomic analyses have examined the impact of PA seed priming on gene expression under stress conditions, there is still a need to clarify the exact effect of PA in the seed priming process. For this purpose, in-depth analyses should be conducted to differentiate the gene expression levels between PA-priming and hydro-priming seeds. This becomes even more significant when considering studies that demonstrate the impact of priming on PA metabolism. Basra et al. [68] observed that osmopriming of onion (Allium cepa L.) seeds led to increased levels of Put and Spd, while Spm level remained unchanged compared to unprimed seeds. In Brassica napus, seed priming resulted in elevated Put accumulation, with no significant changes in the other PA content. Additionally, seed priming alters the balance between the free, bound, and conjugated forms of PA, which can influence their metabolic functions [69]. Studies have shown that the relative proportions of various PA are more crucial for regulating plant response to stress than the absolute concentrations of particular PA [70].

Table 1.
Physiological, biochemical, and molecular effects of seed priming with polyamines to enhance stress tolerance. Abbreviations are explained in the text above the Table.
Table 1.
Physiological, biochemical, and molecular effects of seed priming with polyamines to enhance stress tolerance. Abbreviations are explained in the text above the Table.
| Plant species | Stress | Priming agent | Effect | Reference |
|---|---|---|---|---|
| Acer saccharinum L. | Mild and severe desiccation | Spd |
|
[58] |
|
Brassica napus L. cv. SY Saveo (sensitive), Edimax CI (intermediate tolerant), and Dynastie (tolerant) |
Salinity | Spd Spm |
|
[46] |
|
Brassica oleracea L. var. acephala |
Chilling | Spd |
|
[56] |
|
Chenopodium quinoa Willd. |
Salinity | Spd Spm |
|
[66] |
|
Cucurbita pepo var. styriaka |
Salinity | Put |
|
[47] |
|
Nicotiana tabacum L. cv. MSk326 (sensitive) and Honghuadajinyuan (tolerant) |
Chilling | Put |
|
[59] |
|
Solanum lycopersicum L. cv. Principe Borghese |
Salinity | Put Spm Spd |
|
[60] |
|
Oryza sativa L. ssp. Japonica cv. Zhegeng 100 |
Heat | Spd |
|
[57] |
|
Oryza sativa L. cv. Chunyou 927 (sensitive), and Yliangyou 689 ( tolerant) |
Chromium | Spd |
|
[49] |
|
Oryza sativa L. cv. Chunyou 927 (sensitive) |
Chromium toxicity | Spd |
|
[50] |
|
Oryza sativa L. cv. Khao Dawk Mali 10 |
Salinity | Spd |
|
[65] |
| Oryza sativa L. | Chilling | Spd |
|
[61] |
| Oryza sativa L.cv. IR-64 | Salinity | Spd |
|
[51] |
|
Oryza sativa L. cv. Zhu Liang You 06 and Qian You No.1 |
Chilling | Spd |
|
[62] |
|
Oryza sativa L. cv. Gobindobhog |
Salinity | Spd Spm |
|
[52] |
|
Oryza sativa L. cv. IR-64 (sensitive) and Nonabokra (tolerant) |
Salinity | Spd Spm |
|
[53] |
|
Oryza sativa L. cv. viz., Huanghuazhan (HHZ, inbred) and Yangliangyou6 (YLY6, hybrid) |
Drought | Spd |
|
[63] |
|
Oryza sativa L. |
Chilling | Spd |
|
[64] |
|
Oryza sativa L. cv. Niewdam (tolerant) and KKU-LLR-039 (sensitive) |
Salinity | Spd |
|
[48] |
4. Conclusions and Perspectives
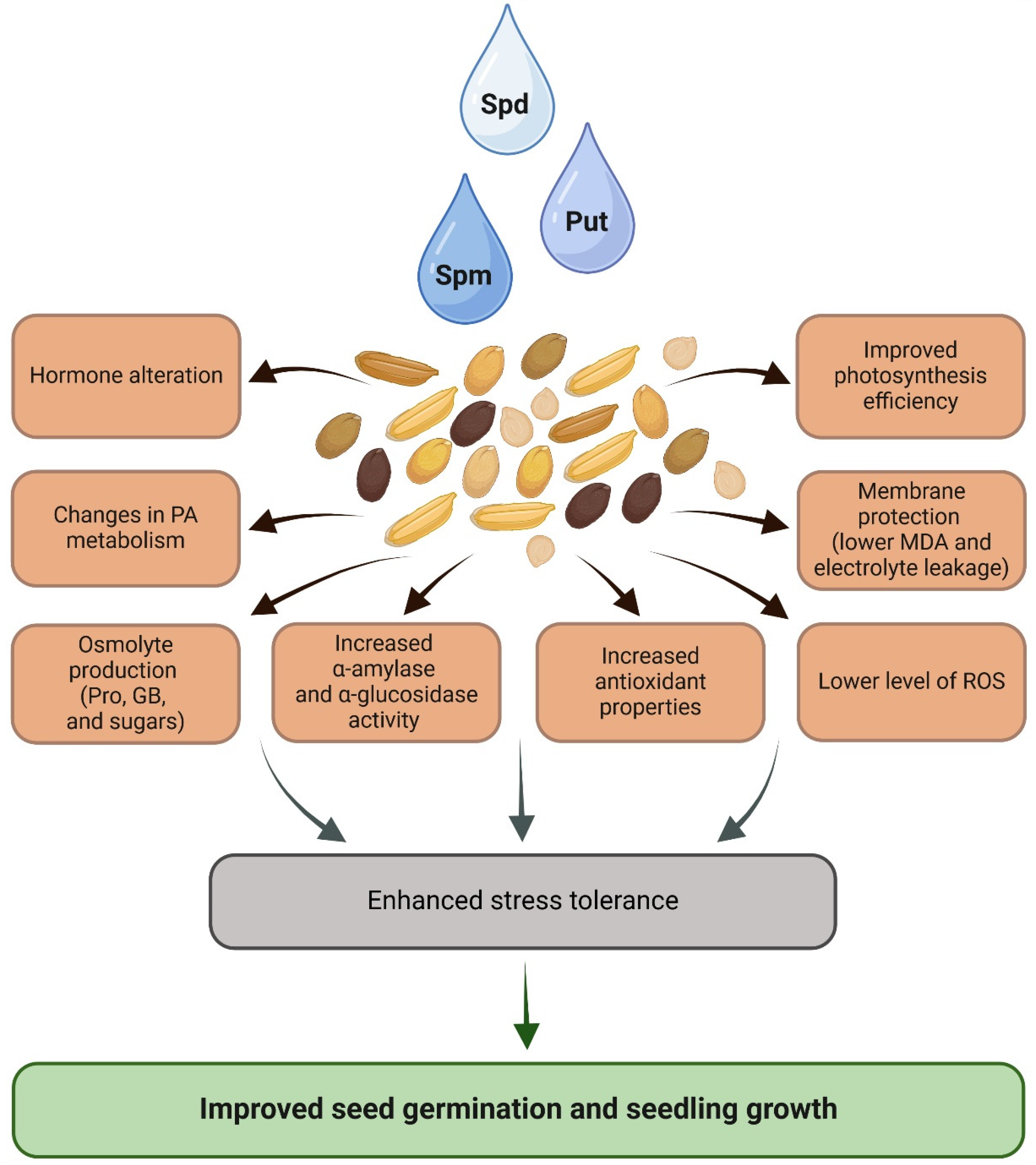
Polyamines play an important function in seed biology as they are involved in the processes of embryo development, seed ripening, and germination, and can be used as a priming agent to enhance seed performance. PA seed priming enhanced stress tolerance, which is manifested through improved seed germination and seedling growth. Numerous studies have shown that PA-primed seeds exhibit improved growth parameters, including increased photosynthetic pigment content, reduced oxidative damage, and enhanced antioxidant enzyme activity. These biochemical changes are accompanied by alterations in gene expression related to stress responses, antioxidant defense, osmolyte production, and PA metabolism. Some studies have highlighted the role of Spd and Spm priming in promoting photosynthesis, reducing lipid peroxidation, and regulating ROS levels (Figure 2). Molecular changes, such as the modulation in gene expression of PA biosynthesis and degradation enzym, further contribute to the enhanced stress tolerance observed in primed plants. However, despite the growing body of research, the precise molecular mechanisms underlying PA priming, particularly its effect on gene expression, remain unclear. Further in-depth analyses comparing PA-priming with water-priming are required to fully elucidate the role of PA in the complex gene expression regulatory network involved in plant stress responses.
Author Contributions
Conceptualization, Ł.W.; literature search, Ł.W., and K.W.; writing—original draft preparation, Ł.W.; table preparation, K.W.; figures, K.W. and S.B.; writing—review and editing, S.B., and M.G.; supervision, Ł.W., and M.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The authors thank Elżbieta Rudy for assistance with Figure 2 preparation. Science Suite Inc. dba BioRender ("BioRender") has granted Elżbieta Rudy to use this Completed Graphic in accordance with BioRender's Terms of Service and Academic License Terms.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Evenari, M. Seed Physiology: Its History from Antiquity to the Beginning of the 20th Century. Bot. Rev. 1984, 50, 119–142. [Google Scholar] [CrossRef]
- Thanos, C.A. Theophrastus the Founder of Seed Science - His Accounts on Seed Preservation. ENSCOnews 2007, 5. [Google Scholar]
- Raj, A.B.; Raj, S.K. Seed Priming: An Approach towards Agricultural Sustainability. JANS 2019, 11, 227–234. [Google Scholar] [CrossRef]
- Singh, V.K.; Singh, R.; Tripathi, S.; Devi, R.S.; Srivastava, P.; Singh, P.; Kumar, A.; Bhadouria, R. Seed Priming: State of the Art and New Perspectives in the Era of Climate Change. In Climate Change and Soil Interactions; Elsevier, 2020; pp. 143–170 ISBN 978-0-12-818032-7.
- Heydecker, W. Germination of an Idea: The Priming of Seeds. School of Agriculture Research, University of Nottingham 1973, 50–67.
- Di Girolamo, G.; Barbanti, L. Treatment Conditions and Biochemical Processes Influencing Seed Priming Effectiveness. Ital. J. Agronomy 2012, 7, 25. [Google Scholar] [CrossRef]
- Rajjou, L.; Duval, M.; Gallardo, K.; Catusse, J.; Bally, J.; Job, C.; Job, D. Seed Germination and Vigor. Annu. Rev. Plant Biol. 2012, 63, 507–533. [Google Scholar] [CrossRef]
- Paparella, S.; Araújo, S.S.; Rossi, G.; Wijayasinghe, M.; Carbonera, D.; Balestrazzi, A. Seed Priming: State of the Art and New Perspectives. Plant Cell Rep 2015, 34, 1281–1293. [Google Scholar] [CrossRef]
- Lutts, S.; Benincasa, P.; Wojtyla, L.; Kubala, S.; Pace, R.; Lechowska, K.; Quinet, M.; Garnczarska, M. Seed Priming: New Comprehensive Approaches for an Old Empirical Technique. In New Challenges in Seed Biology - Basic and Translational Research Driving Seed Technology; Araujo, S., Balestrazzi, A., Eds.; InTech, 2016. [CrossRef]
- Jisha, K.C.; Vijayakumari, K.; Puthur, J.T. Seed Priming for Abiotic Stress Tolerance: An Overview. Acta Physiol Plant 2013, 35, 1381–1396. [Google Scholar] [CrossRef]
- Marthandan, V.; Geetha, R.; Kumutha, K.; Renganathan, V.G.; Karthikeyan, A.; Ramalingam, J. Seed Priming: A Feasible Strategy to Enhance Drought Tolerance in Crop Plants. IJMS 2020, 21, 8258. [Google Scholar] [CrossRef]
- Thakur, M.; Tiwari, S.; Kataria, S.; Anand, A. Recent Advances in Seed Priming Strategies for Enhancing Planting Value of Vegetable Seeds. Scientia Horticulturae 2022, 305, 111355. [Google Scholar] [CrossRef]
- Wojtyla, Ł.; Lechowska, K.; Kubala, S.; Garnczarska, M. Molecular Processes Induced in Primed Seeds—Increasing the Potential to Stabilize Crop Yields under Drought Conditions. Journal of Plant Physiology 2016, 203, 116–126. [Google Scholar] [CrossRef]
- Pagano, A.; Macovei, A.; Balestrazzi, A. Molecular Dynamics of Seed Priming at the Crossroads between Basic and Applied Research. Plant Cell Rep. 2023, 42, 657–688. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, E.A. Seed Priming to Alleviate Salinity Stress in Germinating Seeds. Journal of Plant Physiology 2016, 192, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.; Puthur, J.T. Seed Priming as a Cost Effective Technique for Developing Plants with Cross Tolerance to Salinity Stress. Plant Physiology and Biochemistry 2021, 162, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, P.; Dwivedi, P. Seed Priming and Its Role in Mitigating Heat Stress Responses in Crop Plants. J Soil Sci Plant Nutr 2021, 21, 1718–1734. [Google Scholar] [CrossRef]
- Paul, S.; Dey, S.; Kundu, R. Seed Priming: An Emerging Tool towards Sustainable Agriculture. Plant Growth Regul. 2022, 97, 215–234. [Google Scholar] [CrossRef]
- Bruce, T.J.A.; Matthes, M.C.; Napier, J.A.; Pickett, J.A. Stressful “Memories” of Plants: Evidence and Possible Mechanisms. Plant Science 2007, 173, 603–608. [Google Scholar] [CrossRef]
- Conrath, U.; Beckers, G.J.M.; Langenbach, C.J.G.; Jaskiewicz, M.R. Priming for Enhanced Defense. Annu. Rev. Phytopathol. 2015, 53, 97–119. [Google Scholar] [CrossRef]
- Martinez-Medina, A.; Flors, V.; Heil, M.; Mauch-Mani, B.; Pieterse, C.M.J.; Pozo, M.J.; Ton, J.; Van Dam, N.M.; Conrath, U. Recognizing Plant Defense Priming. Trends in Plant Science 2016, 21, 818–822. [Google Scholar] [CrossRef]
- Yang, Z.; Zhi, P.; Chang, C. Priming Seeds for the Future: Plant Immune Memory and Application in Crop Protection. Front. Plant Sci. 2022, 13, 961840. [Google Scholar] [CrossRef]
- Chen, K.; Arora, R. Priming Memory Invokes Seed Stress-Tolerance. Environmental and Experimental Botany 2013, 94, 33–45. [Google Scholar] [CrossRef]
- Harris, C.J.; Amtmann, A.; Ton, J. Epigenetic Processes in Plant Stress Priming: Open Questions and New Approaches. Current Opinion in Plant Biology 2023, 75, 102432. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Huang, K.; Batool, M.; Idrees, F.; Afzal, R.; Haroon, M.; Noushahi, H.A.; Wu, W.; Hu, Q.; Lu, X.; et al. Versatile Roles of Polyamines in Improving Abiotic Stress Tolerance of Plants. Front. Plant Sci. 2022, 13, 1003155. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Shao, Q.; Yin, L.; Younis, A.; Zheng, B. Polyamine Function in Plants: Metabolism, Regulation on Development, and Roles in Abiotic Stress Responses. Front. Plant Sci. 2019, 9, 1945. [Google Scholar] [CrossRef] [PubMed]
- Alcázar, R.; Bueno, M.; Tiburcio, A.F. Polyamines: Small Amines with Large Effects on Plant Abiotic Stress Tolerance. Cells 2020, 9, 2373. [Google Scholar] [CrossRef]
- Blázquez, M.A. Polyamines: Their Role in Plant Development and Stress. Annual Review of Plant Biology 2024, 75, 95–117. [Google Scholar] [CrossRef]
- Sun, X.; Lv, A.; Chen, D.; Zhang, Z.; Wang, X.; Zhou, A.; Xu, X.; Shao, Q.; Zheng, Y. Exogenous Spermidine Enhanced the Water Deficit Tolerance of Anoectochilus Roxburghii by Modulating Plant Antioxidant Enzymes and Polyamine Metabolism. Agricultural Water Management 2023, 289, 108538. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, X.; Pan, X.; Jiang, Q.; Xi, Z. Exogenous Putrescine Alleviates Drought Stress by Altering Reactive Oxygen Species Scavenging and Biosynthesis of Polyamines in the Seedlings of Cabernet Sauvignon. Front. Plant Sci. 2021, 12, 767992. [Google Scholar] [CrossRef]
- Özmen, S.; Tabur, S.; Öney-Birol, S.; Özmen, S. The Effect of Exogenous Spermine Application on Some Biochemichal and Molecular Properties in Hordeum vulgare L. under Both Normal and Drought Stress. Biologia 2022, 77, 193–202. [Google Scholar] [CrossRef]
- Legocka, J.; Sobieszczuk-Nowicka, E.; Wojtyla, Ł.; Samardakiewicz, S. Lead-Stress Induced Changes in the Content of Free, Thylakoid- and Chromatin-Bound Polyamines, Photosynthetic Parameters and Ultrastructure in Greening Barley Leaves. Journal of Plant Physiology 2015, 186–187, 15–24. [Google Scholar] [CrossRef]
- Jiang, D.; Hou, J.; Gao, W.; Tong, X.; Li, M.; Chu, X.; Chen, G. Exogenous Spermidine Alleviates the Adverse Effects of Aluminum Toxicity on Photosystem II through Improved Antioxidant System and Endogenous Polyamine Contents. Ecotoxicology and Environmental Safety 2021, 207, 111265. [Google Scholar] [CrossRef]
- Raziq, A.; Mohi Ud Din, A.; Anwar, S.; Wang, Y.; Jahan, M.S.; He, M.; Ling, C.G.; Sun, J.; Shu, S.; Guo, S. Exogenous Spermidine Modulates Polyamine Metabolism and Improves Stress Responsive Mechanisms to Protect Tomato Seedlings against Salt Stress. Plant Physiology and Biochemistry 2022, 187, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Pascual, L.S.; López-Climent, M.F.; Segarra-Medina, C.; Gómez-Cadenas, A.; Zandalinas, S.I. Exogenous Spermine Alleviates the Negative Effects of Combined Salinity and Paraquat in Tomato Plants by Decreasing Stress-Induced Oxidative Damage. Front. Plant Sci. 2023, 14, 1193207. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Gu, W.; Li, J.; Li, C.; Xie, T.; Qu, D.; Meng, Y.; Li, C.; Wei, S. Exogenously Applied Spermidine Alleviates Photosynthetic Inhibition under Drought Stress in Maize (Zea mays L.) Seedlings Associated with Changes in Endogenous Polyamines and Phytohormones. Plant Physiology and Biochemistry 2018, 129, 35–55. [Google Scholar] [CrossRef]
- He, X.; Hao, J.; Fan, S.; Liu, C.; Han, Y. Role of Spermidine in Photosynthesis and Polyamine Metabolism in Lettuce Seedlings under High-Temperature Stress. Plants 2022, 11, 1385. [Google Scholar] [CrossRef] [PubMed]
- ElSayed, A.I.; Mohamed, A.H.; Rafudeen, M.S.; Omar, A.A.; Awad, M.F.; Mansour, E. Polyamines Mitigate the Destructive Impacts of Salinity Stress by Enhancing Photosynthetic Capacity, Antioxidant Defense System and Upregulation of Calvin Cycle-Related Genes in Rapeseed (Brassica napus L.). Saudi Journal of Biological Sciences 2022, 29, 3675–3686. [Google Scholar] [CrossRef] [PubMed]
- Napieraj, N.; Janicka, M.; Reda, M. Interactions of Polyamines and Phytohormones in Plant Response to Abiotic Stress. Plants 2023, 12, 1159. [Google Scholar] [CrossRef]
- Gao, J.; Zhuang, S.; Zhang, Y.; Qian, Z. Exogenously Applied Spermidine Alleviates Hypoxia Stress in Phyllostachys Praecox Seedlings via Changes in Endogenous Hormones and Gene Expression. BMC Plant Biol. 2022, 22, 200. [Google Scholar] [CrossRef]
- Borek, S.; Morkunas, I.; Ratajczak, W.; Ratajczak, L. Metabolism of Amino Acids in Germinating Yellow Lupin Seeds III. Breakdown of Arginine in Sugar-Starved Organs Cultivated in Vitro. Acta Physiol Plant 2001, 23, 141–148. [Google Scholar] [CrossRef]
- Burrell, M.; Hanfrey, C.C.; Murray, E.J.; Stanley-Wall, N.R.; Michael, A.J. Evolution and Multiplicity of Arginine Decarboxylases in Polyamine Biosynthesis and Essential Role in Bacillus subtilis Biofilm Formation. Journal of Biological Chemistry 2010, 285, 39224–39238. [Google Scholar] [CrossRef]
- Yariuchi, Y.; Okamoto, T.; Noutoshi, Y.; Takahashi, T. Responses of Polyamine-Metabolic Genes to Polyamines and Plant Stress Hormones in Arabidopsis Seedlings. Cells 2021, 10, 3283. [Google Scholar] [CrossRef]
- Hummel, I.; Bourdais, G.; Gouesbet, G.; Couée, I.; Malmberg, R.L.; El Amrani, A. Differential Gene Expression of ARGININE DECARBOXYLASE ADC1 and ADC2 in Arabidopsis thaliana : Characterization of Transcriptional Regulation during Seed Germination and Seedling Development. New Phytologist 2004, 163, 519–531. [Google Scholar] [CrossRef] [PubMed]
- Lou, Y.; Ahmed, S.; Yan, J.; Adio, A.M.; Powell, H.M.; Morris, P.F.; Jander, G. Arabidopsis ADC1 Functions as an N δ -acetylornithine Decarboxylase. JIPB 2020, 62, 601–613. [Google Scholar] [CrossRef] [PubMed]
- Stassinos, P.M.; Rossi, M.; Borromeo, I.; Capo, C.; Beninati, S.; Forni, C. Enhancement of Brassica Napus Tolerance to High Saline Conditions by Seed Priming. Plants 2021, 10, 403. [Google Scholar] [CrossRef] [PubMed]
- Farsaraei, S.; Mehdizadeh, L.; Moghaddam, M. Seed Priming with Putrescine Alleviated Salinity Stress During Germination and Seedling Growth of Medicinal Pumpkin. J Soil Sci Plant Nutr 2021, 21, 1782–1792. [Google Scholar] [CrossRef]
- Chunthaburee, S.; Sanitchon, J.; Pattanagul, W.; Theerakulpisut, P. Alleviation of Salt Stress in Seedlings of Black Glutinous Rice by Seed Priming with Spermidine and Gibberellic Acid. Not Bot Horti Agrobo 2014, 42, 405–413. [Google Scholar] [CrossRef]
- Basit, F.; Bhat, J.A.; Ulhassan, Z.; Noman, M.; Zhao, B.; Zhou, W.; Kaushik, P.; Ahmad, A.; Ahmad, P.; Guan, Y. Seed Priming with Spermine Mitigates Chromium Stress in Rice by Modifying the Ion Homeostasis, Cellular Ultrastructure and Phytohormones Balance. Antioxidants 2022, 11, 1704. [Google Scholar] [CrossRef]
- Basit, F.; Ulhassan, Z.; Mou, Q.; Nazir, M.M.; Hu, J.; Hu, W.; Song, W.; Sheteiwy, M.S.; Zhou, W.; Bhat, J.A.; et al. Seed Priming with Nitric Oxide and/or Spermine Mitigate the Chromium Toxicity in Rice (. Funct. Plant Biol. 2022, 50, 121–135. [Google Scholar] [CrossRef]
- Paul, S.; Roychoudhury, A.; Banerjee, A.; Chaudhuri, N.; Ghosh, P. Seed Pre-Treatment with Spermidine Alleviates Oxidative Damages to Different Extent in the Salt (NaCl)-Stressed Seedlings of Three Indica Rice Cultivars with Contrasting Level of Salt Tolerance. Plant Gene 2017, 11, 112–123. [Google Scholar] [CrossRef]
- Paul, S.; Roychoudhury, A. Effect of Seed Priming with Spermine/Spermidine on Transcriptional Regulation of Stress-Responsive Genes in Salt-Stressed Seedlings of an Aromatic Rice Cultivar. Plant Gene 2017, 11, 133–142. [Google Scholar] [CrossRef]
- Paul, S.; Roychoudhury, A. Seed Priming with Spermine and Spermidine Regulates the Expression of Diverse Groups of Abiotic Stress-Responsive Genes during Salinity Stress in the Seedlings of Indica Rice Varieties. Plant Gene 2017, 11, 124–132. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. REACTIVE OXYGEN SPECIES: Metabolism, Oxidative Stress, and Signal Transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed]
- Mishra, N.; Jiang, C.; Chen, L.; Paul, A.; Chatterjee, A.; Shen, G. Achieving Abiotic Stress Tolerance in Plants through Antioxidative Defense Mechanisms. Front. Plant Sci. 2023, 14, 1110622. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.; Huang, Y.; Mei, G.; Zhang, S.; Wu, H.; Zhao, T. Spermidine Enhances Chilling Tolerance of Kale Seeds by Modulating ROS and Phytohormone Metabolism. PLoS ONE 2023, 18, e0289563. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Mei, G.; Cao, D.; Qin, Y.; Yang, L.; Ruan, X. Spermidine Enhances Heat Tolerance of Rice Seeds during Mid-Filling Stage and Promote Subsequent Seed Germination. Front. Plant Sci. 2023, 14, 1230331. [Google Scholar] [CrossRef]
- Fuchs, H.; Plitta-Michalak, B.P.; Małecka, A.; Ciszewska, L.; Sikorski, Ł.; Staszak, A.M.; Michalak, M.; Ratajczak, E. The Chances in the Redox Priming of Nondormant Recalcitrant Seeds by Spermidine. Tree Physiology 2023, 43, 1142–1158. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Hu, J.; Li, Y.; Ma, W.; Zheng, Y.; Zhu, S. Chilling Tolerance in Nicotiana tabacum Induced by Seed Priming with Putrescine. Plant Growth Regul. 2011, 63, 279–290. [Google Scholar] [CrossRef]
- Borromeo, I.; Domenici, F.; Del Gallo, M.; Forni, C. Role of Polyamines in the Response to Salt Stress of Tomato. Plants 2023, 12, 1855. [Google Scholar] [CrossRef]
- Fu, Y.; Zhang, Z.; Liu, J.; Chen, M.; Pan, R.; Hu, W.; Guan, Y.; Hu, J. Seed Priming with Spermidine and Trehalose Enhances Chilling Tolerance of Rice via Different Mechanisms. J Plant Growth Regul 2020, 39, 669–679. [Google Scholar] [CrossRef]
- Sheteiwy, M.; Shen, H.; Xu, J.; Guan, Y.; Song, W.; Hu, J. Seed Polyamines Metabolism Induced by Seed Priming with Spermidine and 5-Aminolevulinic Acid for Chilling Tolerance Improvement in Rice (Oryza sativa L.) Seedlings. Environmental and Experimental Botany 2017, 137, 58–72. [Google Scholar] [CrossRef]
- Zheng, M.; Tao, Y.; Hussain, S.; Jiang, Q.; Peng, S.; Huang, J.; Cui, K.; Nie, L. Seed Priming in Dry Direct-Seeded Rice: Consequences for Emergence, Seedling Growth and Associated Metabolic Events under Drought Stress. Plant Growth Regul 2016, 78, 167–178. [Google Scholar] [CrossRef]
- Farzin, P.-D.; Mohammad, K.-H.; Masoud, E. Alleviating Harmful Effects of Chilling Stress on Rice Seedling via Application of Spermidine as Seed Priming Factor. Afr. J. Agric. Res. 2014, 9, 1412–1418. [Google Scholar] [CrossRef]
- Theerakulpisut, P.; Nounjan, N.; Kumon-Sa, N. Spermidine Priming Promotes Germination of Deteriorated Seeds and Reduced Salt Stressed Damage in Rice Seedlings. Not Bot Horti Agrobo 2021, 49, 12130. [Google Scholar] [CrossRef]
- Biondi, S.; Antognoni, F.; Marincich, L.; Lianza, M.; Tejos, R.; Ruiz, K.B. The Polyamine “Multiverse” and Stress Mitigation in Crops: A Case Study with Seed Priming in Quinoa. Scientia Horticulturae 2022, 304, 111292. [Google Scholar] [CrossRef]
- Vaishnav, D.; Chowdhury, P. Types and Function of Phytohormone and Their Role in Stress. In Plant Abiotic Stress Responses and Tolerance Mechanisms; Hussain, S., Hussain Awan, T., Ahmad Waraich, E., Iqbal Awan, M., Eds.; IntechOpen, 2023. [CrossRef]
- Basra, A.S.; Singh, B.; Malik, C.P. Amelioration of the Effects of Ageing in Onion Seeds by Osmotic Priming and Associated Changes in Oxidative Metabolism. Biologia Plant. 1994, 36. [Google Scholar] [CrossRef]
- Lechowska, K.; Wojtyla, Ł.; Quinet, M.; Kubala, S.; Lutts, S.; Garnczarska, M. Endogenous Polyamines and Ethylene Biosynthesis in Relation to Germination of Osmoprimed Brassica Napus Seeds under Salt Stress. IJMS 2021, 23, 349. [Google Scholar] [CrossRef]
- Lutts, S.; Hausman, J.-F.; Quinet, M.; Lefèvre, I. Polyamines and Their Roles in the Alleviation of Ion Toxicities in Plants. In Ecophysiology and Responses of Plants under Salt Stress; Ahmad, P., Azooz, M.M., Prasad, M.N.V., Eds.; Springer New York: New York, NY, 2013; pp. 315–353. [Google Scholar] [CrossRef]
Figure 1.
Simplified scheme of putrescine, spermidine, and spermine metabolism in plants.
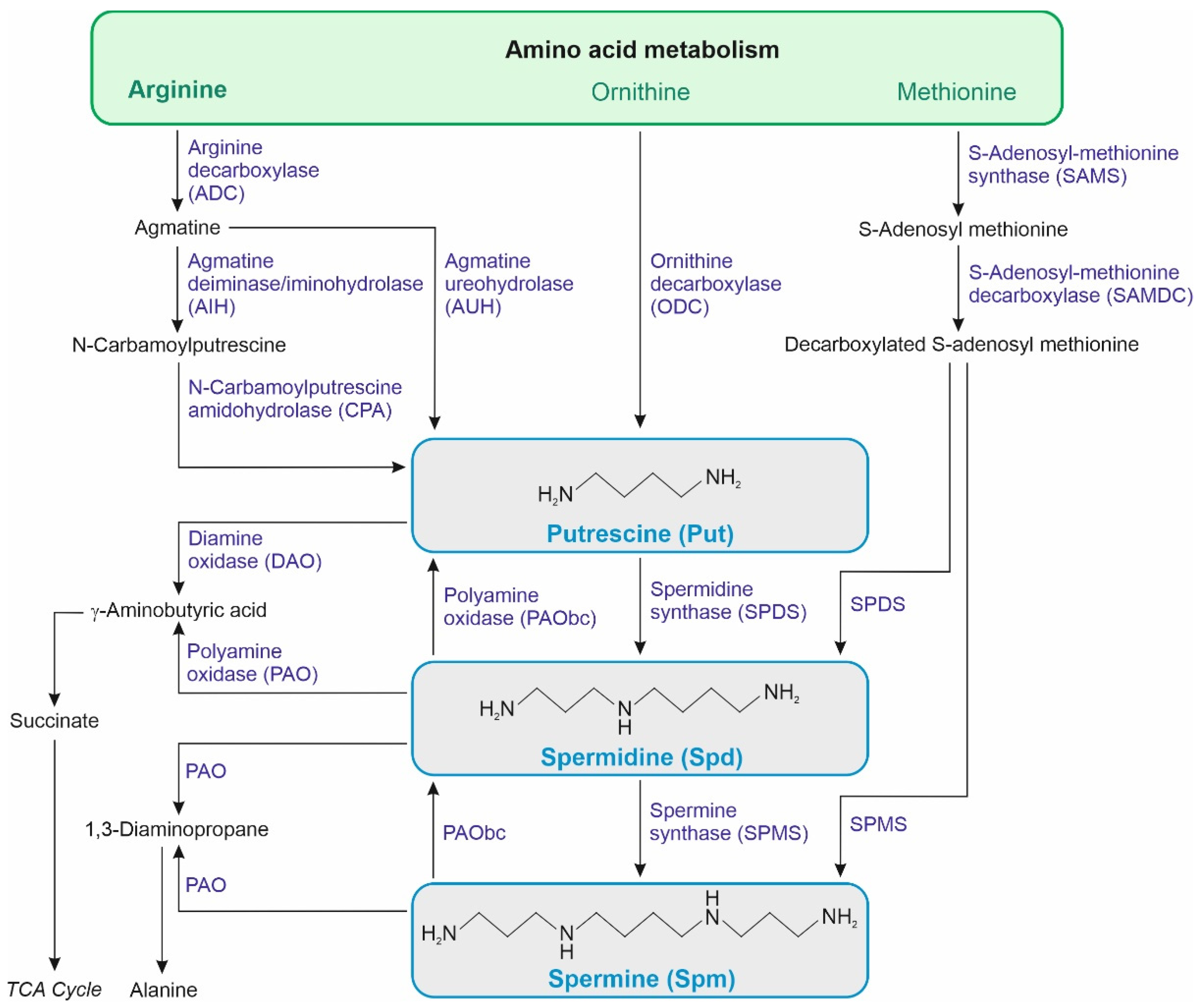
Figure 2.
The general overview of the mechanisms underlying induced stress response upon PA seed priming. Abbreviations are explained in the text. Created in BioRender.
Figure 2.
The general overview of the mechanisms underlying induced stress response upon PA seed priming. Abbreviations are explained in the text. Created in BioRender.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



