
Submitted:
16 October 2024
Posted:
16 October 2024
You are already at the latest version
Abstract
Heavy infestations of weedy rice can reduce rice yields and affect farmers' incomes. So far, the control of weedy rice is still unsolved and depends more on the use of herbicides, which can harm the environment and human health. The phenomenon of allelopathy can be used to develop a new strategy to control weedy rice that does not depend on the use of herbicides. The objective of this study was to evaluate the allelopathic ability of T. procumbens against the growth of weedy rice with different morphological characteristics. Although morphologically different, seedling growth of all weedy rice was found to be inhibited by the presence of leaf leaching and root exudates of T. procumbens, as determined by sandwich and plant box methods. GC-MS analysis identified 21 allelochemicals present in the leaves of T. procumbens, mainly consisting of various fatty acids such as hexadecanoic acid. Further investigation revealed that the aqueous extract of T. procumbens at higher concentration (100%) reduced the germination rate of weed variants WR3 and WR7 by 24% and 19%, respectively. Seedling growth of the weedy variants was inhibited with increasing concentration of leaf extract, with radicle growth of both WR3 and WR7 reduced by 100% and 98%, respectively, at the highest concentration. The leaf debris incorporated into the soil similarly inhibited the growth of the seedlings and the chlorophyll content of the weedy rice. However, the cultivar MR220 CL2 showed lower sensitivity to T. procumbens, and its total chlorophyll content increased with increasing concentration of leaf debris. Our study revealed that T. procumbens is suitable as a bio-herbicide for future sustainable control of weedy rice, especially in the granaries of MR220 CL2
Keywords:
allelochemicals
; allelopathy
; oryza sativa
; rice
; tridax procumbens
; weedy rice management
1. Introduction
The competitive behaviour of weedy rice makes it difficult to control and become a significant threat to rice cultivation activities worldwide [1]. Weedy rice, in particular, Oryza sativa or locally known as 'padi angin'. Its local name derives from its easily shattering characteristic [2]. Competition between cultivated rice and weedy rice thrives in the granary areas mainly because of the similarities in characteristics, specifically during the early growth [3]. These relative attributes make the weedy rice undetectable in the rice fields and continuously competes for space and nutrients from the surrounding [2]. Since the 2000s, weedy rice has been a widespread nationwide problem [4]. The infestation of weedy rice occurred due to the shifting of agricultural practice from hand-transplanting method to direct-seeding approach [2,5,6]. According to Shrestha [7], weedy rice possessed several characteristics such as taller habit, higher tillering and great nutrient user to compete with the cultivated rice.
Various preventive measures need to be taken to reduce weedy rice infestation such as use of weed-free rice seed, proper soil preparation, selection of competitive rice varieties and intensive weed control [8,9,10]. Originally, the use of chemicals for weedy rice control was very limited and usually included 2,4-D phenoxy compounds, which were commonly used to control broadleaf weeds [11]. Some types of herbicides such as oxadiazon and metalactors can control the growth of weedy rice, but these herbicides can also affect the growth of rice varieties [12]. Herbicide resistant rice varieties can be a solution for managing weedy rice [13]. So far, three types of resistant rice varieties have been developed, namely those resistant to imidazolinone, gluphosinate and glyphosate [13]. In Malaysia, two imidazolinone-resistant rice varieties were introduced in 2010 to control weedy rice infestations [13]. However, resistant rice varieties can lead to the emergence of herbicide-resistant weedy rice through gene flow and make it increasingly difficult to control weedy rice [13]. A study conducted by Dilipkumar et al. [14] found that there were cases of crosses where weedy rice resistant to the herbicides Imazapic and Imazapyr occurred in fields of Clearfield® rice varieties.
Allelopathy is a natural phenomenon that can be defined as the inhibitory or stimulatory effect of one plant towards another plant, which involves releasing the chemical compounds to the surrounding environment [15]. The exploration of allelopathy and its potential in the agricultural ecosystem have been prominent among researchers around the globe since 40 years ago [15,16]. The biochemical produced by the donor plant, also known as allelochemical usually found on several parts of plants such as leaves, stems, seeds, rhizomes, buds, fruits and roots [17,18,19]. The toxicity of allelochemicals is more diminutive than chemical pesticides [17]. Therefore, these natural compounds in plants harbour a broad biological effect and can contribute functional benefit to the agricultural weed management system [20,21].
T. procumbens is a perennial herbaceous plant in the family Asteraceae [22]. According to Holm et al. [23], T. procumbens is native to Central America and has now spread to most tropical and subtropical regions. In Malaysia, Holm et al. [24] listed T. procumbens as a common weed found mainly in agricultural areas and orchards. Recently, studies have been conducted on the potential allelopathy of T. procumbens. T. procumbens was found to have a growth inhibitory effect on radish [25] and some legumes such as Vigna radiata and V. unguiculata [26]. However, studies on the potential of allelopathy of T. procumbens on rice crops and major weeds are not yet widely available. The study of allelopathic activities of rice usually refers to cultivated rice varieties. The study of weedy rice has not been extensively researched. According to various findings, the incorporation of allelopathy in the management and control of weedy rice is still in its initial stage. Karim et al. [27] stated that detailed studies on biology and ecology of rice weeds including weedy rice (O. sativa) should be carried out to formulate successful weed control. Therefore, this study was conducted to answer several questions: 1.) Does T. procumbens have the potential to inhibit morphologically diverse weedy rice without affecting the growth of modern rice varieties? 2.) What possible allelochemicals are present in T. procumbens that are responsible for its allelopathic potential? and 3.) Can aqueous extract and debris of T. procumbens suppress the growth of weedy rice so that it can be used as a bio-herbicide to reduce dependence on herbicide use? In addition, the results of this study can provide researchers and farmers with helpful information for managing weedy rice while protecting the environment.
2. Materials and Methods
2.1. Collection of Plant Materials
Leaves of T. procumbens were collected from the vicinity of Kolej Rahim Kajai, UKM (2.936°N 101.782°E) and stored in a large container before being transferred to the laboratory at Plant House Complex, Faculty of Science and Technology, UKM, Bangi, Selangor. The sampled leaves were then washed with distilled water and oven dried at 60°C for 3 days. After drying, the leaf samples were ground using a commercial grinder and stored in an airtight container. The airtight container containing the samples of dried T. procumbens leaves was then stored in a cold room (4°C) until use. Seeds of weedy rice variants were collected from different areas at Peninsular Malaysia and classified into six groups according to their morphological characteristics (Table 1). Meanwhile, modern rice cultivars consisting of MR 220 CL2, Sebernas 307 and MR 297 were obtained from the Gene and Seed Bank Center, MARDI Seberang Perai .
2.2. Sandwich Method
The sandwich method was applied to evaluate allelopathic activity due to leachate from donor plant foliage on bioassay plant growth, using agar as the growth medium [28]. First, the dried leaves of T. procumbens were divided into two total weights, namely 10 mg and 50 mg. Both weights of the dried leaves were placed in a multidish with six wells (Nalge Nunc International, Rochester, New York), each well measuring 35 mm x 18 mm. The multidish without the dried leaves of T. procumbens served as a control (0 mg). Each experiment was repeated three times. The agar (Nacalai Tesque, Kyoto, Japan) used as growth medium was first autoclaved at a temperature of 121 °C and a pressure of 121 kPa for 15 min. The autoclaved agar was then cooled to 45 °C before being poured into each multidish. Approximately 5 ml of agar was poured into the multidish to form the first agar layer, and after the first layer was gelled, another 5 ml of agar was poured to form the second layer. The agar layer was to form a physical barrier between the leaves of the donor plant and the seeds of the bioassay plant [29]. Meanwhile, 10 ml of the autoclaved agar was poured onto the control dish. Then, three seeds of weedy rice variants and modern rice varieties were placed on the surface of the second gelatinized agar layer. The multidish was labeled, sealed with tape, and incubated at 28°C for seven days. After seven days, shoot and radicle elongation of the bioassay plants were measured and recorded.
2.3. Plant Box Method
The plant box method was developed by Fujii et al. [30] to evaluate the allelopathic activity of the donor plant on the bioassay plant by root exudation. T. procumbens seedlings that had germinated in a greenhouse for two months were harvested and the roots were cleaned with distilled water. One T. procumbens seedling was placed in a PVC tube with a nylon net (which served as a root separation zone) attached to one edge of the magenta box. The agar (Nacalai Tesque, Kyoto, Japan) (0.75% w/v) used as the germination medium was autoclaved at 121°C and 121 kPa pressure for 15 min and then cooled to 40-45°C. The cooled agar was then poured into a magenta box containing seedlings to a height of 6 cm and gelatinized. A total of 33 bioassay plant seeds were placed on the gelatinized agar with the distance to the seedling determined in the corner of the magenta box. The lower part of the magenta box was wrapped with aluminum foil, while the upper part was covered with a clear plastic film. The magenta boxes without donor plants were designated as control. All magenta boxes were placed in a growth chamber at a temperature of 27-28°C and allowed to germinate for seven days. Two growth parameters, radicle and shoot elongation, were measured and recorded after seven days.
2.4. Gas Chromatography – Mass Spectrometry
The extraction of T. procumbens plants was carried out and modified following Kushwaha et al.[31]. A total of 100 g of the dried leaves of T. procumbens were placed in an Erlenmeyer flask. Then, the Erlenmeyer flask was filled with 500 ml of methanol and heated on a hot plate at 64 °C for 15 min. After heating, the Erlenmeyer flask was shaken with an orbital shaker for 24 hours. The resulting extract was filtered using Whatman filter paper No. 1 and then placed in a centrifuge tube (Thermo Fisher Scientific, Waltham, USA). The extract was centrifuged in a centrifuge (Allegra X-22R Centrifuge, Beckman Coulter, Inc., Switzerland) for 15 minutes to separate the solid particles present in the extract. The resulting supernatant was filtered using a 0.22-µm filter syringe (Merck KgaA, Darmstadt, Germany) and transferred to a 1.5-ml glass vial with PTFE plastic septum (Shimadzu Europa GmbH, Duisburg, Germany). The vials were then stored in a cold room (-20°C) before being injected into the GC-MS instrument for analysis.
GC- MS analysis (Agilent Technologies, Inc., Santa Clara, USA) was performed using a gas chromatograph (GC) (Agilent 7890A) connected to an inert MSD mass spectrometry system (MS) (Agilent 5975C) with a three-axis detector. The column conditions are shown in Table 2. Helium (99.999%) was used as the carrier gas with a constant flow of 1.0 ml/min. An injection of 0.5 EI was used with a 10:1 ratio between the injector temperature (250°C) and the ion source temperature (280°C). The oven temperature was programmed at 110°C (isothermal for 2 minutes). The temperature was increased from 10°C - 200°C and then from 5°C - 280°C, followed by isotherm for 9 minutes at a temperature of 280°C. GC-MS analysis was performed for 30 minutes and the mass spectrum (MS) was recorded at 70 eV, a scan interval of 0.5 and a fraction of 40-550 Da. All peaks in the raw GC chromatogram data were identified using MSD Chemstation software. A library search was then performed for all identified peaks using NIST /EPA/NIH version 2.0 software, and the results were summarized in a peak table. The GC-MS instrument used in this experiment was located at Molecular Structure Characterization Laboratory (MPSM), Research Management and Instrumentation Center (CRIM), Universiti Kebangsaan Malaysia (UKM), Bangi, Selangor.
2.5. Aqueous Extract Preparation
The preparation of extract from the leaves of T. procumbens was modified following the study of Femina et al. [26] and Anwar et al. [32]. A total of 100 g of dried leaves of T. procumbens were placed in an Erlenmeyer flask and then made up with 100 ml of distilled water. The dried leaves were immersed in the distilled water and shaken with an orbital shaker at 160 rpm for 24 hours. Then the resulting extract was filtered using a commercial filter and filtered using Whatman filter paper No. 2 to remove solid particles. The filtered extract was then centrifuged at a speed of 6000 rpm for 15 minutes. The resulting supernatant was then filtered through a water pump using cellulose nitrate filter paper (0.22 µm). The resulting extract was prepared as stock solution with concentration of 100%. Then concentrations of 10%, 25%, 50% and 75% were prepared by serial dilution of the stock solution. All extract solutions were then filled into separate bottles, labeled and stored in a cold room (4°C) until use. To break dormancy of bioassay plant seeds, all bioassay seeds were heated in distilled water at 90°C. The seeds were pre-germinated for two days to test their viability. After two days, the petri dishes were disinfected with 70% ethanol to prevent fungal growth and filled with Whatman filter paper No. 2 as growth medium. A total of seven bioassay seeds were added to the Petri dish, separated according to the concentration of T. procumbens leaf extract solution. Each Petri dish was then moistened with 5 ml of the extract solution at different concentrations while the Petri dish was given 5 ml of distilled water as control (0%). All Petri dishes were then sealed, labeled and incubated in a germination chamber at 27 ± 3 °C for seven days. Germination was recorded daily, and all petri dishes were checked to maintain the humidity of the filter paper containing the extract solution. After seven days, some germination and growth parameters such as germination percentage (GP), germination rate/ index (GI), seed vigor index (SVI), radicle and hypocotyl length, radicle to shoot elongation ratio, fresh and dry weight were recorded.
2.6. Pot Experiment
The dried leaves of T. procumbens were weighed to three total weights (2.5 g, 5 g and 10 g) using an electronic balance. Each weight was then mixed with 500 g of soil to obtain three concentrations: 2.5 g / 500 g of soil, 5 g / 500 g of soil and 10 g / 500 g of soil. A total of seven seeds per bioassay plant were heated to 90°C and pre-germinated for two days to break dormancy and test seed viability. Seeds were then sown in soil mixed with three concentrations of T. procumbens leaf debris in small pots without holes (15 cm x 12 cm). Pots without T. procumbens leaf debris were designated as control (0 g / 500 g soil). Then each pot was filled with 250 ml of distilled water. This experiment was conducted in a greenhouse at the Plant House Complex, Faculty of Science and Technology, UKM, Bangi, Selangor. Each pot was controlled and 250 ml of water was replenished daily to avoid water loss. Seed germination (GP and GI) was recorded on the seventh day. After seven days, seedlings were lined to three and allowed to grow for 30 days. Growth parameters such as seedling height, fresh weight and dry weight of leaves, and chlorophyll content were recorded on day 30.
2.7. Statistical Analysis
In the sandwich method, the mean ± standard error of elongation and inhibition of shoot and radicle were determined and compared with the control using one-way analysis of variance (ANOVA) with Duncan's Multiple Range Test (DMRT) at p 0.05 level of significance. In plant box method, two growth parameters namely, radicle and shoot elongation were recorded and the means ± standard errors were determined and compared with the controls using independent samples t-test. The relationship between the two parameters in relation to distance from donor plants was analyzed using Pearson's correlation at a significant p< 0.01. Data on allelopathic effects of aqueous leaves and leaf debris depend on one-way analysis of variance (ANOVA) and mean ± standard error for each concentration compared to control using Duncan's Multiple Range Test (DMRT) at 95% level of significance (p <0.05). All analyzes were performed using SPSS version 23 software. Experiments were conducted using a randomized complete block design (RCBD), with each concentration and control having three replicates.
3. Results
3.1. Allelopathic Potential of T. procumbens Foliar Leachate
The leachate from the dried leaves of T. procumbens showed allelopathic potential on seedling and shoot growth in the bioassay (Table 3 and Table 4). Despite the differences in morphological characteristics, the radicle and hypocotyl elongation of weedy variants were significantly inhibited (p< 0.05) by the presence of foliar leachate from T. procumbens. As measured by the mean percentage of inhibition, radicle and hypocotyl growth of the weedy variants were reduced by 30-63% and 15-41%, respectively. It was found that radicle growth was more sensitive to the foliar leachate of T. procumbens than that of the hypocotyl. The strongest inhibition effect on radicle growth was observed in WR2 and WR3 with 60-69% followed by WR4, WR5 and WR6 with inhibition effect of 50-59%. The least inhibition of seedling growth was observed in weedy variant WR7 (30.23%). In the modern cultivars, the inhibition effect on seedling growth was less than 21% except MR297 (43.01%). The hypocotyl growth of the modern cultivars was probably stimulated and ranged from 2 to 7%, except for MR220 CL2 which was inhibited at 13.30 %. Based on the ranking, it was observed that the foliar leachate of T. procumbens had less inhibitory effect on the radicle growth of the modern cultivars, with Sebernas 307 and MR220 CL2 ranked 8th and 9th respectively.
The values in each column that had the same letter were not significant (p<0.05) based on the Duncan's Multiple Range Test. The percentage of radicle inhibition is based on comparison with the percentage of elongation of the control. The ranking is based on the leaching inhibitory effect of T. procumbens leaves on bioassay plants.
The values in each column that had the same letter were not significant (p<0.05) based on the Duncan's Multiple Range Test. The percentage of hypocotyl inhibition is based on comparison with the percentage of elongation of the control. The ranking is based on the leaching inhibitory effect of T. procumbens leaves on bioassay plants.
In general, foliage leaching of T. procumbens was found to affect hypocotyl and radicle growth of bioassay species by the sandwich method. Both hypocotyl and radicle growth of the bioassay were reduced with increasing concentration of dried leaves. This result is in agreement with a study by Nurul Ain et al. [33], which found that T. procumbens leachate inhibited hypocotyl and root elongation of lettuce seedlings with increasing concentration from 10 mg to 50 mg. The leachate had a stronger inhibitory effect on the weedy variants rather than on the modern cultivars. The leaf potential of T. procumbens was dependent on the species or cultivar. A study by Popoola et al. [34] found that an extract of T. procumbens leaves inhibited the growth of Vigna unguiculata cultivar IT07K-292-10 more than that of cultivar IT99K-573-1-1. The sensitivity of radicle growth to foliar leachate of T. procumbens could be due to the absorption and concentration of phytotoxins in root tissues upon direct contact with the substrate [35]. Morikawa et al. [36] discovered this phenomenon and found that leaf leachate from 170 plant species in Peru affected lettuce root growth rather than hypocotyl growth.
3.2. Allelopathic Potential of T. procumbens Root Exudates
Based on Table 5, the exudates from T. procumbens roots displayed suppressing activities on the radicle growth of the bioassay plants. Significantly, distance influenced the radicle elongation of the weedy variants as the inhibition occurred mainly on the seedlings located at proximity to the roots of T. procumbens (< 10 mm) (Figure 1). A more significant inhibitory effect was recorded in the radicle elongation of WR3 with 36.8% elongation and high correlation coefficient value, R2 (0.88) of linear regression followed by WR2 (44.1%) and WR6 (49.1%). In addition, radicle elongation of 60–69% was recorded in WR4, WR5, WR7 weedy variants and MR297 cultivar. However, a less inhibitory effect was observed in the radicle of MR220 CL2 with 79.2% elongation and a small R2 value (0.35) (Figure 2). Small R2 values indicate that the radicle elongation of MR220 CL2 was less likely to be affected by the distances between their seedlings and T. procumbens roots.
Radicle growth of bioassay plants was found to be inhibited according to their distance from T. procumbens. This finding coincides with the study conducted by Itani et al. [37], who found some plants from the Asteraceae family such as Cynara scolymus, Matricaria reticutita, and Helianthus tuberosum suppressed the radicle growth of Lactuca sativa depending on their distance with those plants. High inhibition of the radicle elongation was observed on the bioassay plants, especially at the closest distance to the recipient plant. Similarly, a plant box study conducted by Shiraishi et al. [38] found that Phlox subulata showed the highest inhibitory activity on radicle elongation of the nearest (<20 mm) L. sativa seeds with 11% elongation compared to controls with high correlation coefficient, R2 value (0.93). The slopes from the linear regression analysis showed the movement of allelochemicals presence in T. procumbens roots. On the other hand, the allelopathic activities exhibited by T. procumbens roots occurred only at a constricted area. Less inhibitory effects on MR220 CL2 variety assumed that the potential of T. procumbens root exudates was variety-dependent and proved that its hold specific activities.
3.3. Allelochemicals Presence in T. procumbens Leaves
Based on the GC-MS analysis, 21 chemical compounds were identified from the leaves methanolic extract of T. procumbens. As shown in Table 6, some of the major compounds consisted in T. procumbens leaves are hexadecanoic acid, trimethylsilyl ester (RT, 14.89; peak area, 12.46%), 2-[(4-(4-fluorocinnamoyl) anilino]-3-piperidino-1,4-naphthoquinone (RT, 26.02; peak area, 11.55%), squalene (RT, 25.88; peak area, 10.63%), cis-pinane (RT, 12.24; peak area, 8.36%), phytol (RT, 15.87; peak area, 6.65%), 9,12,15-octadecatrienoic acid, (Z, Z, Z)-(RT, 16.29; peak area, 6.22%), β-sitosterol (RT, 34.99; peak area, 6.09%) and acid n-hexadecanoic (RT, 13.85; peak area, 5.02%). Other compounds identified were stigmasterol and α-linolenic acid, trimethylsilyl ester with peak areas below 5% and several compounds with 1-3% peak areas.
The three significant allelochemicals identified in T. procumbens leaves were hexadecanoic acid, trimethylsilyl ester followed by 2-[(4-(4-fluorocinnamoyl)anilino]-3-piperidino-1,4-naphthoquinone and squalene. In addition, the leaves were also dominated by several compounds from the carboxylic acid group (fatty acids) such as n-hexadecanoic acid, 9,12,15-octadecatrienoic acid, (Z,Z,Z)-, 9,12-octadecadienoic acid (Z,Z)- and α-linolenic acid, trimethylsilyl ester. Similarly, extract of T. procumbens leaves collected in India was identified to contain various fatty acid compounds that possessed pharmacological potentials, such as octadecanoic acid, n-hexadecanoic acid and phthalic acid [31]. A study conducted by Andriana et al. [25] revealed that the ethyl acetate extract of T. procumbens leaves sampled in Indonesia contained several fatty acids, namely palmitic acid, butanoic acid and hexanoic acid, besides having a high amount of phenolic compounds. Although there are similarities in the identified chemical compounds, some compounds have never been identified from previous studies. This occurrence explained that the diversity of plants in one area could influence ecosystem characteristics and allelochemical content in those plants [39,40]. Plant physiology is also influenced by its environment [41].
3.4. Allelopathic Influzences of T. procumbens Leaf Aqueous Extract
- A Germination
Based on the sandwich and plant box screening method, four bioassay plants were selected to investigate T. procumbens allelopathic abilities further. Therefore, two weedy variants (WR3 and WR7) and two modern cultivars (Sebernas 307 and MR220 CL2) were determined to assess their germination and growth responses towards T. procumbens leaf aqueous extract. Based on Figure 3, there is no significant reduction in the germination percentage of the bioassay plants, especially at 10-50% aqueous extract concentrations. However, at the highest concentration (100%), the GP of WR3 and WR7 weedy variants were reduced by 24% and 19%, whilst the highest inhibition was recorded on Sebernas 307 (reduced by 67% compared to control). The lowest was observed on MR220 CL2 (reduced by 5% compared to control). On the other hand, the rate of germination (RG) of the bioassay plants was significantly decreased (p<0.05) with increasing concentration of the leaf aqueous extract except for MR220 CL2 cultivar. At 100% concentration, the RG of WR3, WR7 and Sebernas 307 were highly inhibited, reducing 62%, 58%, and 85% compared to control, respectively. On the contrary, only 15% inhibition was observed on the RG of MR220 CL2 at the same concentration compared to control. The seedling vigor index (SVI) of the bioassay plants was also affected by T. procumbens leaf aqueous extract with different concentrations (Figure 4). More significant inhibition on the SVI of the bioassay plants was observed to have occurred at the maximum concentration (100%). At the concentration mentioned earlier, the SVI of Sebernas 307 significantly reduced by 98% compared to control, followed by WR3, WR7 and MR220 CL2, reducing 92%, 89%, and 80% to control, respectively. However, the decreasing pattern of SVI per the increasing concentration was only observed on the WR3 significantly (p<0.05). Meanwhile, the SVI of WR7 and Sebernas 307 were started to reduce significantly at 50% concentration of the aqueous leaf extract, whilst the SVI of MR220 CL2 displayed a less significant reduction at 10-75% concentration. A low concentration (10%) of T. procumbens leaf aqueous extract was observed to stimulate the SVI of both weedy variants.
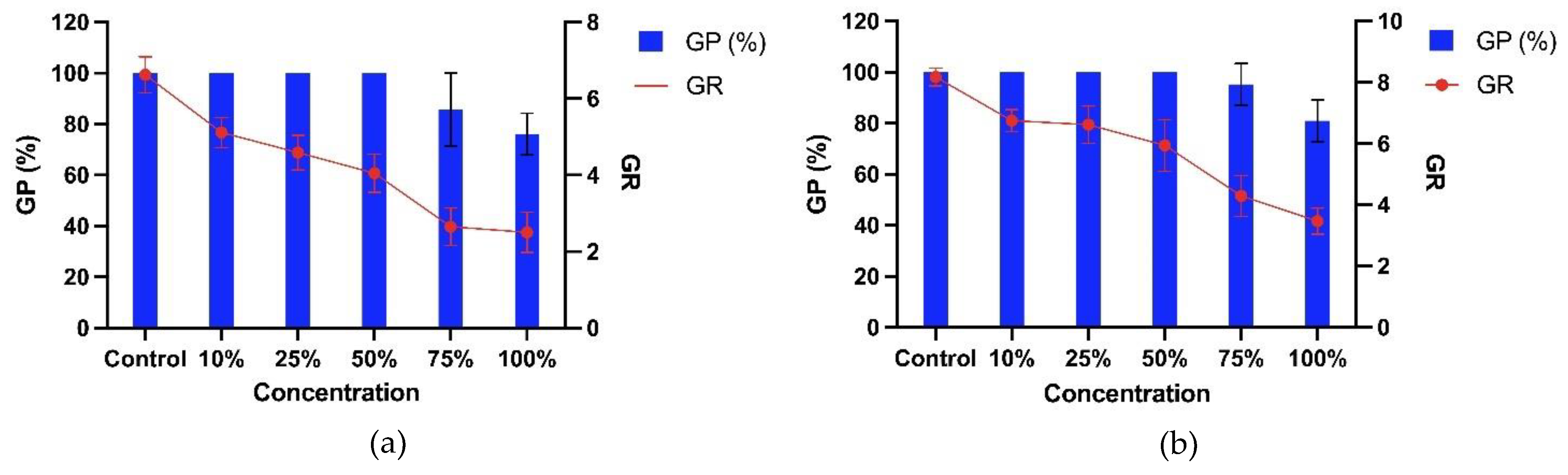
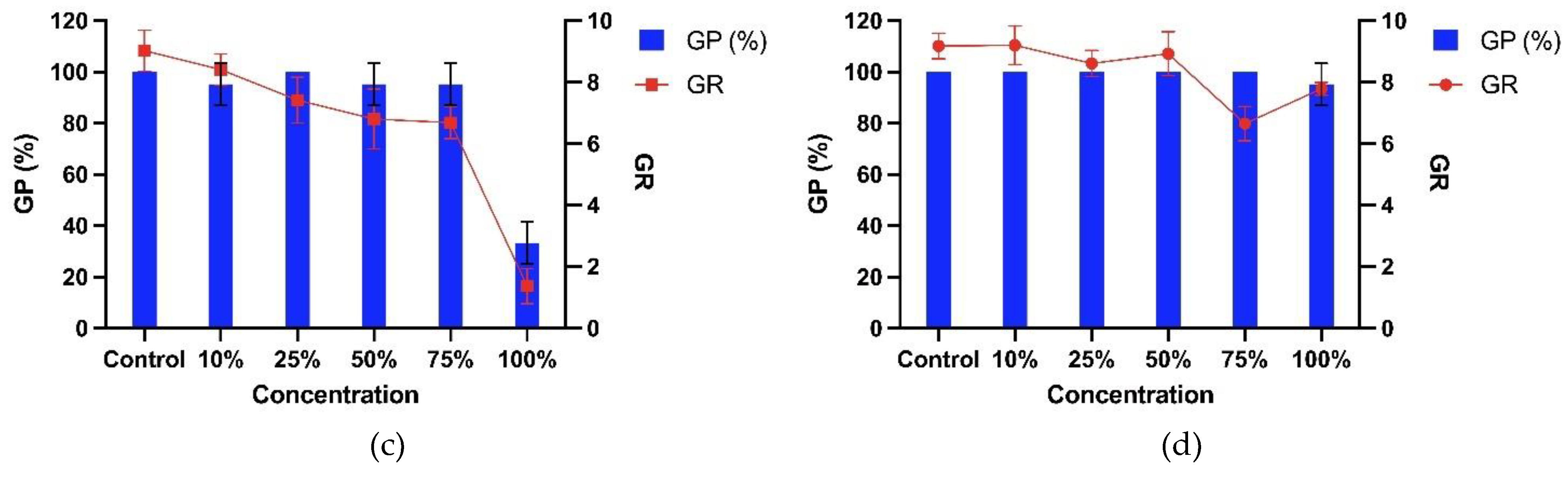
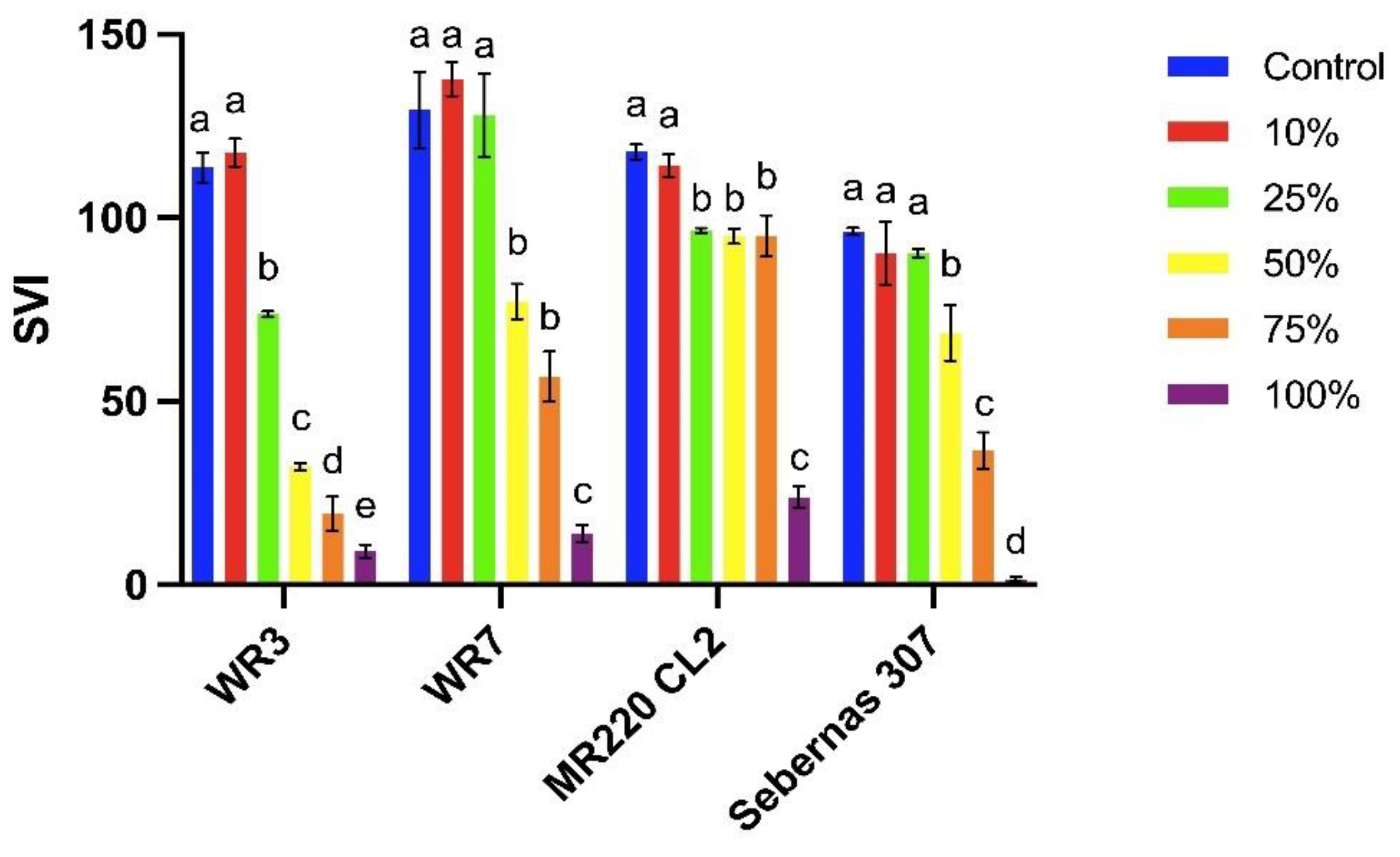
Figure 1.
Allelopathic effects of the aqueous extract of T. procumbens leaves at different concentrations on the GP and GR of (a) WR3, (b) WR7, (c) Sebernas 307 and (d) MR220 CL2. Abbreviations: GP, percentage of germination and GR, germination rate.
Figure 1.
Allelopathic effects of the aqueous extract of T. procumbens leaves at different concentrations on the GP and GR of (a) WR3, (b) WR7, (c) Sebernas 307 and (d) MR220 CL2. Abbreviations: GP, percentage of germination and GR, germination rate.
Figure 2.
Allelopathic effects of aqueous extract of T. procumbens leaves at different concentrations on SVI of bioassay plants. Bars (mean ± standard error) with same alphabet are not significantly different (p< 0.05) according to Duncan's Multiple Range Test. Abbreviations: SVI, seedling vigor index.
Figure 2.
Allelopathic effects of aqueous extract of T. procumbens leaves at different concentrations on SVI of bioassay plants. Bars (mean ± standard error) with same alphabet are not significantly different (p< 0.05) according to Duncan's Multiple Range Test. Abbreviations: SVI, seedling vigor index.
The leaf extract of T. procumbens displayed different allelopathic effects on the germination of the bioassay plants depending on the concentration. Lower concentration caused slight inhibition or showed zero inhibitory effects on the GP of the bioassay. Meanwhile, more significant inhibition on the GP occurred at the highest concentration of the leaf extract. The inhibition influences of T. procumbens leaf extract were in agreement with Femina et al. [26], who discovered that the aqueous extract of T. procumbens at the highest concentration (75%) suppressed the germination percentage of Vigna radiata, Dolichos biflorus and V. anguiculata whilst vaguely affected the germination percentage at low concentration (25 and 50%). Aqueous extract of certain weeds, such as the greater club rush (Scirpus grossus), has no adverse effects on the germination of rice and weedy rice [42]. A study by Ghahari & Miransari [43] found that the germination of different rice cultivars in Iran responded differently with each other aqueous extracts. Similarly, the GP of MR220 CL2 experienced no inhibition at 10-75% concentration. A comparative study between two weeds, namely T. procumbens and Parthenium hysterophorus, indicated that the aqueous extract of T. procumbens at lower concentration displayed less inhibitory effects on the germination percentage of black gram (V. mungo) crop [44]. The results proved that the allelopathic effects of T. procumbens leaf aqueous extract towards the GP of the bioassay were depended on concentration and the variety involved.
In addition, the presence of aqueous extract from T. procumbens leaves was found to delay the seed germination rate (GR) of the bioassay plants. Moreover, the GR of the weedy variants was found to be inhibited with increasing concentration of T. procumbens leaf aqueous extract. A similar discovery by Mubeen et al. [45] found that the root extracts slowed the germination of rice seedlings from the weed Trianthema portulacastrum. The study also found that rice seeds soaked with distilled water (depicted as control) had a maximum germination rate than seeds soaked with leaf extract, root, and whole parts of the T. portulacastrum tree. Meanwhile, leaf extracts from two species of broadleaf aquatic weeds, namely Althenanthera philoxeroides and A. sessilis were found to inhibit the seed germination rate of rice seedlings with increasing concentration of extracts of both weeds [46]. On the other hand, the leaf aqueous extract of T. procumbens was less likely to disrupt the GR of MR220 CL2 at any concentration. According to Ghahari and Miransari (2009) [43], most rice cultivars that undergo genetic modification tend to have a greater seed germination rate to increase the efficiency of the cultivars.
A recent study revealed that T. procumbens leaf aqueous extract also affected the seedling vigor index (SVI) for both weedy rice variants. The SVI for weedy variants and Sebernas 307 was found to be inhibited with increasing concentration of T. procumbens leaf extract. The allelochemicals from the T. procumbens leaf extract may have diminished the surrounding environment's quality and subsequently affected the fertility of the seedlings. This finding was in agreement with a study conducted by Joshi & Joshi [47], who discovered that extracts from several medicinal plants such as Hyptis sauveolens, Ricinus communis, A. sessilis, Ipomoea carnea, Malachra capitata and Cymbopogon citratus at two concentrations (3% and 5%) inhibited the SVI of wheat. However, the SVI for MR220 CL2 variety was less affected by the presence of T. procumbens leaf extract. MADA revealed that several rice varieties introduced by MARDI between 2000-2010 possessed a short maturation period and high seed rate to produce a high yield [48]. This finding coincides with Krishna et al. [49] who found that Parthenium and Lantana camara weed extracts inhibited the SVI of some rice cultivars in India, such as Abhilash varieties, 9926 and BPT.5204. Still, extracts from Cassia tora and Chromolaena odorata weeds stimulated the SVI of those varieties. The results proved that the allelopathic ability of T. procumbens leaf aqueous extract on the SVI of the bioassay is variety-dependent.
- B Seedling growth
The hypocotyl and radicle growth of the bioassay plants was affected when treated with different concentrations of the aqueous leaf extract of T. procumbens (Figure 5). At the highest concentration (100%), the radicle growth of WR3 and Sebernas 307 were completely inhibited whilst the WR7 and MR220 CL2 were reduced by 98% and 95% compared to control, respectively. The aqueous leaf extract started to stunt the radicle growth of the bioassay at 50% concentration except for MR220 CL2. The inhibitory effect of T. procumbens leaf aqueous extract least affected the MR220 CL2 variety in which there was no significant reduction in radicle elongation at a concentration of 25–75%. At 10% concentration, the extract was observed to stimulate the radicle elongation of both weedy rice variants with 18% and 3% increments compared to control. A significant reduction was also recorded in the hypocotyl growth of the bioassay plants at the highest concentration (100%) of the aqueous leaf extract. The hypocotyl elongation of Sebernas 307, WR3, WR7 were significantly reduced by 92%, 86% and 78% compared to control at the concentration mentioned earlier. Meanwhile, the lowest inhibition was recorded by MR220 CL2 with a 65% reduction compared to control. Stimulation of the hypocotyl growth was recorded on WR7 at the lowest concentration (10%). The inhibitory effects of T. procumbens aqueous leaf extract were more likely to inhibit the radicle growth than the hypocotyl growth.
Subsequently, the aqueous extract of T. procumbens leaf at different concentrations was observed to affect the fresh dan dry weight of the bioassay plants (Figure 6). The fresh weight of WR3, WR7, Sebernas 307 and MR220 CL2 was significantly reduced by 83%, 84%, 91% and 73% compared to control at the highest concentration (100%) of the aqueous extract. Similarly, the dry weight of the bioassay plants was significantly inhibited when treated with the same concentration. The highest inhibition of dry weight was recorded on Sebernas 307 with a 91% reduction compared to control, followed by WR3 and WR7, which reduced by 88% and 83% compared to control, respectively. The less inhibitory effect was recorded on the dry weight of MR220 CL2 (reduced by 61% compared to control). Meanwhile, at the lowest concentration (10%) of the aqueous leaf extract, the fresh weight of the bioassay plants experienced stimulation. The fresh weight of WR3, WR7, Sebernas 307 and MR220 CL2 was increased by 6%, 16%, 10% and 6% compared to control, respectively. The aqueous leaf extract of T. procumbens started to inhibit the fresh weight of the bioassay plants at 25% concentration except for Sebernas 307. The dry weight of both weedy variants began to decrease at the same concentration. There is no significant reduction observed on MR220 CL2 dry weight when treated with concentration below 100
Aqueous extract of T. procumbens leaves has an allelopathic influence on seedling growth for all bioassay plants. The inhibitory efficacy on the radicle and hypocotyl growth of the bioassay plants depended on the increasing concentration of the aqueous extract, especially at higher concentrations (> 50%). This finding was in agreement with Popoola et al. [34], who found that aqueous extract from several broadleaf weeds such as Chromolaena odorata, Euphorbia heterophylla and T. procumbens inhibited the plumule and radicle elongation of two Vigna unguiculata varieties with increasing concentration. In addition, aqueous extracts of several Asteraceae plants such as Helianthus annuus, C. odorata, and Tithonia diversifolia with different concentrations suppressed the radicle growth of V. unguiculata crop [50]. The radicle growth was more sensitive towards the T. procumbens leaf aqueous extract than the hypocotyl growth. According to Nishida et al. [51], more significant inhibition on the radicle is likely due to substantial allelochemicals with permeability properties that permeate through the radicle tissues. This phenomenon proved that the radicle is in direct contact with allelochemicals secreted by the T. procumbens leaf aqueous extract that can inhibit cell division in the meristematic tissue at the tip of the radicle [52]. A study conducted by Sahu & Devkota [53] found that the aqueous extract from the invasive Mikania micrantha significantly inhibits Raphanus sativus and Oryza sativa radicle than its hypocotyl elongation.
Concurrently, the leaf aqueous extract of T. procumbens displayed allelopathic activities towards the fresh and dry weight of the bioassay plants. The fresh and dry weight inhibition occurred with the increasing concentration of the aqueous extract. A similar finding by Mahmoodzadeh & Mahmoodzadeh [54] revealed that four concentrations from Cynodon dactylon weed restrained the fresh and dry weight of Triticum aestivum. Meanwhile, Khaliq et al. [55] reported that aqueous extract of Eucalyptus camaldulensis inhibited the total dry weight of Trianthema portulacastrum and Echinochloa colona weeds while aqueous extract of Withania somnifera significantly inhibited (over 60%) both fresh and dry weight of E. crus-galli weeds. The inhibitory effect of T. procumbens leaf aqueous extract was variety-dependent. The growth of MR220 CL2 was more tolerant towards the presence of leaf extract than weedy rice variants and Sebernas 307. Verma & Rao [56] found the effect of aqueous extracts from four weed species (Ageratum conyzoides, C. dactylon, P. hysterophorus and Solanum nigrum) on several soybean crop varieties was dependent on the selected soybean variety. For instance, maximum inhibition occurred on plumule elongation of Bhatt soy variety. Still, a stimulating effect occurred on the plumule growth of PS - 1042 variety in the presence of C. dactylon weed leaf extract. On the other hand, Macias et al. [57] suggested that the susceptibility of the rice varieties towards the leaf aqueous extract may be due to the indifferent physiological and morphological characteristics of the genotype. Most rice varieties are durable towards the herbicides and possessed a short maturation period, including MR220 CL2, which is resistant to the OnDutyTM herbicide [58,59].
3.5. Allelopathic Influences of T. procumbens Leaf Debris
- A Germination
Different concentrations of T. procumbens leaf debris inhibited the germination percentage (GP) of the bioassay plants, especially at the highest concentration of the debris (Figure 7). All bioassay plant's GP was found to be inhibited between 5-29% compared to control when sown together with T. procumbens leaf debris at the highest concentration (10 g / 500 g soil). The highest inhibition was recorded on the GP of WR7 with a 29% reduction compared to control, followed by WR3, MR220 CL2, and Sebernas 307, reduced by 14%, 10%, and 5% compared to control, respectively. A low concentration of the leaf debris (2.5 g / 500 g soil) was not affecting the GP of both modern cultivars. On the other hand, no significant reduction was recorded by the GP of WR3, MR220 CL2, and Sebernas 307 with increasing concentration of the leaf debris.
Meanwhile, the debris was found to inhibit the germination rate (GR) of the bioassay plants with increasing concentration (Figure 7). The highest concentration (10 g / 500 g soil), the GR of WR7 and WR3 was reduced by 62% and 48% compared to control, while the GR of MR220 CL2 and Sebernas 307 was only decreased by 19% and 11% compared to control, respectively. The lowest concentration of leaf debris still suppresses the GR of the bioassay plants. The highest inhibition was recorded on the GR of WR7 (reduced by 12% compared to control), followed by WR3 with a 10% reduction. The GR of both modern cultivars was inhibited by 6% compared to control at the same concentration. The influence of T. procumbens leaf litter was found to have a more negligible effect on the GR of MR220 CL2 and Sebernas 307. No significant inhibition was recorded by both varieties, especially at the leaf debris concentration of 2.5-10 g.
T. procumbens debris incorporated with soil was found to have allelopathic effects on the germination of the bioassay plants. The germination of the weedy variants was inhibited, especially with the presence of the debris at high concentrations. At the same time, no significant inhibition was recorded to occur in the germination of the bioassay plants at the lowest leaf litter concentration. Katoch et al. [60] found residues of three weed species, namely Eupatorium adenophorum, Ageratum conyzoides and Lantana camara incorporated with soil, inhibited wheat, rice and maize germination at high concentrations (10 g / 100 g of soil). However, the germination of rice crops was found not to experience significant inhibition with the presence of residues of all three weeds at lower concentrations (5 g / 100 g of soil). Usuah et al. [61] found mulch from several weed species such as Apilia africana, Emilia sonchifolia, Chromolaena odorata, and Cyperus esculentus inhibited the germination percentage of some crop species such as corn, wheat and soybeans according to different residue concentrations. Meanwhile, T. procumbens leaf litter was also found to inhibit the seed germination rate for all bioassay plants. The inhibitory effect of T. procumbens leaf litter is dependent on the increase in its total concentration. This occurrence coincides with a study conducted by Fatunbi et al. [62], who found that the seed germination index of Brassica oleracea, Eragrotis curvula and Zea mays underwent inhibition to the increasing concentration of Acacia mearnsii leaf litter.
- B Seedling growth
The presence of T. procumbens leaf debris influenced the seedling height of both weedy rice variants. Leaf debris with increasing concentration significantly inhibited the seedling height of both variants (Figure 8). At the highest debris concentration (10 g / 500 g soil), WR3 variant seedling height decreased by 46% compared to control, followed by WR7 with a 20% reduction compared to control. However, the seedling height of MR220 CL2 was only inhibited by 1% compared to control, whilst Sebernas 307 experienced 2% stimulation at the same concentration. T. procumbens debris at different concentrations did not affect the seedling height of the modern cultivars compared to the weedy variants. Low concentration (2.5 g / 500 g soil) of the leaf debris stimulated the seedling height of WR7 and Sebernas 307 with increments of 4% and 3% compared to control, respectively.
The fresh leaf weight of both weedy variants was influenced by the presence of T. procumbens leaf debris. In general, the total fresh leaf weight of the weedy variants underwent significant inhibition according to the increase in T. procumbens leaf debris concentration, as shown in Figure 9. Significantly, the fresh leaf weight of WR7 and WR3 was reduced by 66% and 54% compared to control when treated with the highest concentration (10 g / 500 g soil) of the debris. On the other hand, the same concentration stimulated the fresh leaf weight of MR220 CL2 (increased by 4% compared to control). At the lowest concentration (2.5 g / 500 g soil), the debris was found to inhibit the fresh leaf weight of all the bioassay plants. The T. procumbens leaf debris also affects the dry leaf weight of the bioassay plants. The dry leaf weight of the weedy variants was significantly reduced with the increasing concentration of the debris. Compared to the control, the dry leaf weight of WR3 and WR7 was reduced by 66% and 56% at the highest concentration of the debris. Meanwhile, dry leaf weight of MR220 CL2 was only inhibited by 10% compared to control, while stimulation of 7% was recorded by Sebernas 307. The dry leaf weight of WR3, WR7, MR220 CL2 and Sebernas 307 was reduced by 30%, 7%, 12% and 24% compared to control at the lowest concentration. No significant reduction was noted by the dry leaf weight of MR220 CL2, especially at 2.5-10 g debris concentration.
T. procumbens debris incorporated into soil was found to have allelopathic effects on all bioassay plants. However, the growth of both modern rice cultivars is less sensitive to the presence of the debris. The debris' inhibitory effect on the seedling height of the weedy variants is in line with the study by Begum et al. [63] who reported that seedling height and dry weight of rice, mustard and peanut crops were found to experience inhibition by the presence of C. odorata debris at different concentrations. The tolerancy of modern cultivars to T. procumbens debris is coincident with a study conducted by Koodkaew & Rottasa [64], who found that Mimosa pigra leaf debris at different concentrations inhibited shoot elongation and fresh weight of Ruellia tuberosa species but stimulated shoot growth and fresh weight of Portulaca oleracea. Stimulation of seedling height of the bioassay plants was noted to occur in the presence of T. procumbens debris at low concentrations. Ismail et al. [65] discovered that root and shoot growth of Eleusine indica was found to be stimulated by the presence of Pennisetum purpureum leaf litter incorporated into soil at low concentrations (1%). According to Ismail & Kumar [66], the stimulus effect occurring at low concentrations is due to the quantity of allelochemicals produced in plant tissues and allelopathic interactions are highly dependent on the chemical stability of the bioactive material as well as the amount of decomposed residues in the soil. Farooq et al. [67,68] reported that the presence of plant residues in the soil may produce nutrients, increasing organic matter, in turn, improving soil quality leading to growth stimulation of recipient plants.
- C Chlorophyll content
In general, the presence of T. procumbens leaf debris affects the chlorophyll content of all bioassay plants. The total chlorophyll content of the weedy variants was significantly decreased with increasing concentration of leaf debris (Figure 10). At the highest concentration of leaf debris, the total chlorophyll content of WR7 and WR3 had decreased by 42% and 25%, respectively, compared to the control. It was observed that leaf debris decreased the chlorophyll a and b content of both weedy variants at the same concentration. Thus, chlorophyll a content of WR3 and WR7 decreased by 25% and 40%, respectively, compared to the control, while chlorophyll b content decreased by 27% and 46%, respectively. At the same time, total chlorophyll content increased with increasing concentration of leaf debris in both modern cultivars. The total chlorophyll content of Sebernas 307 and MR220 CL2 was greatly increased by 59% and 12%, respectively, at the highest concentration of leaf debris compared to the control. Chlorophyll a content of MR220 CL2 and Sebernas 307 was increased by 58% and 12%, respectively, at the same concentration compared to the control. The same observation was discovered in chlorophyll b content of the two cultivars which increased by 64% and 12% respectively.
T. procumbens leaf debris incorporated into soil had an allelopathic effect on total chlorophyll content of bioassay plants. It was found that the chlorophyll content of weedy variants was inhibited by the presence of leaf debris with increasing concentration. However, only the chlorophyll content of MR220 CL2 and Sebernas 307 varieties did not decrease with increasing concentration of T. procumbens leaf litter. The decrease in chlorophyll content a and b in weedy variants is in agreement with a study by Yaseen & Mutwali [69] who found that residues of Datura stramonium species mixed with soil inhibited chlorophyll content a and b in Beta vulgaris cultivars with increasing concentration of D. stramonium species residues. In turn, Hegab et al. [70] found that leaf residues of Eucalyptus rostrata had an inhibitory effect on the chlorophyll content of a maize crop at high residue concentrations. Meanwhile, a study by Siddique & Ismail [71] reported that Fimbristylis miliacea weed litter mixed with soil inhibited the chlorophyll content of some Malaysian rice cultivars such as MR211, MRQ74, MR84 and MR232 above a concentration of one-fourth (5 g/kg soil) of weed litter. However, the chlorophyll content of variety MR220 was not inhibited at this concentration and even an increase in chlorophyll content was observed in this variety.
Chlorophyll content is closely related to dry matter production, and a decrease in chlorophyll content can impair photosynthetic activity and thus inhibit overall plant growth [72]. The decrease in chlorophyll content is likely due to disturbances in the process of chlorophyll biosynthesis and an increase in chemicals that cause the degradation of chlorophyll [73]. Hussain & Reigosa [74] reported that the decrease in chlorophyll is related to the water content in leaves, which leads to the closure of stomata, which in turn reduces CO2 production in mesophyll cells and the rate of photosynthesis. The increase in total chlorophyll content in MR220 CL2 and Sebernas 307 cultivars is probably due to the interaction between allelochemicals and microbial content in the soil and other soil factors. Ilori & Otusanya [75] reported that shoots of Tithonia rotundifolia mixed with soil stimulate the chlorophyll content of some plant species such as soybean, bush bean, sorghum and maize and that the results are related to the interaction between the allelochemicals of T. rotundifolia and the presence of microbes in the soil. The increase in chlorophyll content in cultivar MR220 CL2 is probably due to the fact that allelochemicals previously detected on T. procumbens leaves, such as palmatic acid, did not affect the cultivar. Most of the allelochemicals found in T. procumbens leaves are fatty acids from the carboxylic acid group. MR220 CL2 is a cultivar resistant to the herbicides imazapic and imazapyr. The herbicide is a synthetic compound from the imidazolinone group, with an imidazolinone ring and a carboxylic acid group attached to its molecular structure [76]. The tolerance of MR220 CL2 may be due to the fact that the herbicide has similar properties to the allelochemicals produced by the leaf residues of T. procumbens.
4. Conclusion
The results of this study are the first to demonstrate the allelopathic potential of T. procumbens on weedy rice germination and growth. Through the initial screening by sandwich and plant box method, it was found that leaf litter leachate and root exudation of T. procumbens reduced seedling growth of weedy rice with different morphological characteristics. Further investigations revealed that several allelochemicals such as palmitic acid and other fatty acids were detected in the leaves of T. procumbens. The aqueous leaf extract of T. procumbens suppressed seedling growth of weedy rice with increasing concentration of the extract, while germination was significantly affected at the highest concentration (100%). The results of the pot culture experiment showed a similar trend where it was observed that the leaf debris of T. procumbens incorporated into the soil had an inhibitory effect on the germination and seedling growth of weedy rice. In contrast, the cultivar MR220 CL2 is tolerant to the aqueous leaf extract and leaf debris of T. procumbens, which may be due to an abundance of allelochemicals that have similar properties to the herbicides that MR220 CL2 resisted. The ability of T. procumbens to suppress weed rice can obviously be investigated to reduce the persistent use of herbicides. The potential allelochemical can be isolated and used in rice growing areas especially MR220 CL2 granaries for future sustainable management of weedy rice.
Author Contributions
Initial ideas and conceptualization, M.S.I,; methodology and sampling, M.S.I., K.M.J. and N.A.A.S; laboratory and greenhouse study, M.S.I.; data analysis, M.S.I.; resources, K.M.J.; first draft preparation, M.S.I.; draft review and editing, K.M.J.; supervision, K.M.J and N.A.A.S.
Acknowledgments
The authors would like to thank all the staff and students of Plant Biotechnology Lab, Greenhouse Complex, Faculty of Science and Technology, Universiti Kebangsaan Malaysia for their support and ideas on this study. Our thanks also go to Gene and Seed Bank Center, MARDI Seberang Perai for providing research materials. We would also like to thank the officials and staff of Muda Agricultural Development Area (MADA), Kemubu Agricultural Development Area (KADA), Integrated Agricultural Development Area (IADA) Seberang Perak and IADA Barat Laut Selangor for their support in field sampling.
Conflicts of Interest
The authors declare no conflicts of interest for this study.
References
- Singh, K.; Kumar, V.; Saharawat, Y.; Gathala, M. Weedy rice: An emerging threat for direct-seeded rice production systems in India. Journal of Rice Research, 2013; 1, 106–111. [Google Scholar]
- Sudianto, E.; Neik, T.-X.; Tam, S.M.; Chuah, T-S.; Idris, A.A.; Olsen, K.M.; Song, B-K. Morphology of Malaysian weedy rice (Oryza sativa): Diversity, origin and implications for weed management. Weed Science 2016, 64, 501–512. [Google Scholar] [CrossRef]
- Hussain, Z.P.M.D.; Azmi, M.; Othman, A.S. Morphological study of the relationships between weedy rice accessions (Oryza sativa complex) and commercial rice varieties in Pulau Pinang rice granary area. Tropical Life Sciences Research 2010, 21, 27–40. [Google Scholar] [PubMed]
- Karim, R.S.M.; Zainal, M.; Mansor, M.; Azmi, M. Weedy rice: a cancerous threat for rice growers in Malaysia. In. Proceedings of the National Rice Conference 2010. Perak, Malaysia: Malaysian Agricultural Research and Development Institute, 2010; pp. 327–329.
- Azmi, M.; Watanabe, H.; Abdullah, M.Z.; Zainal, A.H. Padi angin, an emerging threat to direct-seeded rice. In. Proceedings of the Malaysian Congress of Science and Technology, Kuala Lumpur. Confederation of Scientific and Technological Association in Malaysia, 1994.
- Ziska, L.H.; Gealy, D.R.; Burgos, N.; Caicedo, A.L.; Gressel, J.; Lawton-Rauh, A.L.; Avila, L.A.; Theisen, G.; Norsworthy, J.; Ferrero, A.; Vidotto, F. 2015. Weedy (red) rice: an emerging constraint to global rice production. Advances in Agronomy. 2015, 129, 181–228. [Google Scholar]
- Shrestha, S.; Stallworth, S. ; Tseng, T-M. 2018. Weedy rice: Competitive ability, evolution and diversity [Online First]. Available online: https://www.intechopen.com/online-first/weedy-rice-competitive-ability-evolution-and-diversity (accessed on 29 November 2018).
- Azmi, M.; Abdullah, M.Z.; Muhammad, H. Pengenalan dan Kawalan Padi Angin di Malaysia. Serdang: MARDI Publication, 2001.
- Nakayama, S.; Ghani, R.A.; Azmi, M. Chemical control and a case study on some ecological characteristics. In. Ecology of major weeds and their control in direct seeded rice culture of Malaysia.Malaysian Agricultural Research and Development Institute (MARDI): Penang, 1996; pp. 98–105. P.
- Watanabe, H.; Vaughan, D.A.; Tomooka, N. 2000. Weedy rice complexes: case studies from Malaysia, Vietnam, and Surinam. In. Baki, B.B. et al. (eds.). Wild and Weedy Rice in Rice Ecosystems in Asia: A Review. IRRI: Philippines, 2000; pp. 25.
- Saharan, H.A. Rice Weed Control in Malaysia. MARDI Report No. 66. MARDI: Serdang, 1979.
- Estorninos Jr., L. E.; Gealy, D.R.; Talbert, R.E.; Gbur, E.E. Rice and red rice interference. Response of red rice (Oryza sativa) to sowing rates of tropical japonica and indica rice cultivars. Weed Science 2005, 53, 676–682. [Google Scholar] [CrossRef]
- Chauhan, B.S. Strategies to manage weedy rice in Asia. Crop Protection 2013, 48, 51–56. [Google Scholar] [CrossRef]
- Dilipkumar, M.; Burgos, N.R.; Chuah, T.S.; Ismail, S. Cross-resistance to imazapic and imazapyr in a weedy rice (Oryza sativa) biotype found in Malaysia. Planta Daninha. 2018, 36, e018182239. [Google Scholar] [CrossRef]
- Rice, E.L. Allelopathy, 2nd ed.; Academic Press: New York, 1984. [Google Scholar]
- Singh, H.P.; Batish, D.R.; Kohli, R.K. Allelopathy in agroecosystems: An overview. Journal of Crop Production 2001, 4920, 1–41. [Google Scholar] [CrossRef]
- Khan, M.A.; Kulsoom, M.I.; Khan, R.; Khan, S.A. Screening the allelopathic potential of various weeds. Pakistan Journal of Weed Science Research 2011, 17, 73–81. [Google Scholar]
- Nornasuha, Y.; Ismail, B.S. Sustainable weed management using allelopathic approach. Malaysian Applied Biology 2017, 46, 1–10. [Google Scholar]
- Srivastava, J.N.; Ghatak, A.; Singh, A.K. Allelopathy: How plants suppress other plants. Rashtriya Krishi 2017, 12, 103–106. [Google Scholar]
- Duke, S.O.; Lydon, J. Herbicides from natural compounds. Weed Technology 1987, 1, 122–128. [Google Scholar] [CrossRef]
- Macias, F.A.; Marin, D.; Oliveros-Bastidas, A.; Castellano, D.; Simonet, A.M.; Molinillo, J.M.G. Structure-activity relationship (SAR) studies of benzoxazinones, their degradation products, and analogues. Phytotoxicity on problematic weeds Avena fatua L. and Lolium rigidum Gaud. Journal of Agricultural and Food Chemistry 2006, 54, 1040–1048. [Google Scholar] [CrossRef] [PubMed]
- Barnes, D.E.; Chan, L.G. Common Weeds of Malaysia and Their Control. Ancom Berhad: Kuala Lumpur, 1990.
- Holm, L.; Plucknett, D.; Pancho, J.; Herberger, J. The World's Worst Weeds: Distribution and Biology. University of Hawaii Press, Honolulu, 1977.
- Holm, L.; Pancho, J.V.; Herberger, J.P.; Plucknett, D.L. A Geographical Atlas of World Weeds. Wiley and Sons Inc: New York, 1979.
- Andriana, Y.; Xuan, T.D.; Quan, N.V.; Quy, T.N. Allelopathic potential of Tridax procumbens L. on radish and identification of allelochemicals. Allelopathy Journal 2018, 43, 223–238. [Google Scholar] [CrossRef]
- Femina, D.; Lakshmipriya, P.; Subha, S.; Manonmani, R. Allelopathic effect of the weed (Tridax procumbens L.) on seed germination and seedling growth of some leguminous plants. International Research Journal of Pharmacy 2012, 3, 90–95. [Google Scholar]
- Karim, R.S.M.; Man, A.B.; Sahid, I.B. Weed problems and their management in rice fields of Malaysia: an overview. Weed Biology and Management 2004, 4, 177–186. [Google Scholar] [CrossRef]
- Fujii, Y.; Parvez, S.; Parvez, M.M.; Ohmae, Y.; Iida, O. Screening of 239 medicinal plant species for allelopathic activity using the sandwich method. Weed Biology and Management 2003, 3, 233–241. [Google Scholar] [CrossRef]
- Mardani, H.; Kazantseva, E.; Onipchenko, V.; Fujii, Y. Evaluation of allelopathic activity of 178 Caucassian plant species. International Journal of Basic and Applied Science 2016, 5, 75–81. [Google Scholar] [CrossRef]
- Fujii, Y.; Pariasca, D.; Shibuya, T.; Yasuda, T.; Kahn, B.; Waller, G.R. Plant-box method: a specific bioassay to evaluate allelopathy through root exudates. In. Fujii, Y.; Hiradate, S. (eds.). Allelopathy: New Concepts and Methodology. Science Publishers: Enfield, 2007.
- Kushwaha, P.; Yadav, S.S.; Singh, V.; Dwivedi, L.K. Phytochemical screening and GC-MS studies of the methanolic extract of Tridax procumbens. International Journal of Pharmaceutical Sciences and Research 2019, 10, 2492–2496. [Google Scholar]
- Anwar, T.; Khalid, S.; Saeed, M.; Mazhar, R.; Qureshi, H.; Rashid, M. Allelopathic interference of leaf powder and aqueous extracts of hostile weed: Parthenium hysterophorus (Asteraceae). Science International 2016, 4, 86–93. [Google Scholar] [CrossRef]
- Nurul Ain, M.B.; Nornasuha, Y.; Ismail, B.S. Allelopathic assessment of selected common weeds in Malaysia. AIP Conference Proceedings 2016, 1784, 060039. [Google Scholar]
- Popoola, K.M.; Akinwale, R.O.; Adelusi, A.A. Allelopathic effect of extracts from selected weeds on germination and seedling growth of cowpea (Vigna unguiculata (L.) Walp.) varieties. African Journal of Plant Science 2020, 14, 338–349. [Google Scholar] [CrossRef]
- Correia, N.M.; Centurion, M.A.P.D.C.; Alves, P.L.D.C.A. Influência de extratos aquosos de sorgo sobre a germinação e o desenvolvimento de plântulas de soja. Ciência Rural 2005, 35, 498–503. [Google Scholar] [CrossRef]
- Morikawa, C.I.O.; Miyaura, R.; Tapia Y Figueroa, M.D.L.; Rengifo Salgado, E.L.; Fujii, Y. Screening of 170 Peruvian plant species for allelopathic activity by using the Sandwich Method. Weed Biology and Management 2012, 12, 1–11. [Google Scholar] [CrossRef]
- Itani, T.; Nakahata, Y.; Kato-Noguchi, H. Allelopathic activity of some herb plant species. International Journal of Agriculture and Biology 2013, 15, 1359–1362. [Google Scholar]
- Shiraishi, S.; Watanabe, I.; Kuno, K.; Fujii, Y. Allelopathic activity of leaching from dry leaves and exudate from roots of ground cover plants assayed on agar. Weed Biology and Management 2002, 2, 133–142. [Google Scholar] [CrossRef]
- El Moujahid, L.; Le Roux, X.; Michalet, S.; Bellvert, F.; Weigelt, A.; Poly, F. Effect of plant diversity on the diversity of soil organic compounds. PloS One 2017, 12, e0170494. [Google Scholar] [CrossRef]
- Uddin, M.R.; Li, X.; Won, O.J.; Park, S.U.; Pyon, J.Y. Herbicidal activity of phenolic compounds from hairy root cultures of Fagopyrum tataricum. Weed Research 2011, 52, 25–33. [Google Scholar] [CrossRef]
- Mierziak, J.; Kostyn, K.; Kulma, A. Flavonoids as important molecules of plant interactions with the environment. Molecules 2014, 19, 16240–16265. [Google Scholar] [CrossRef]
- Karthikeyan, P.G.; George, S. Allelopathic effect of greater club rush on rice and weedy rice. Trends in Biosciences 2017, 10, 1099–1102. [Google Scholar]
- Ghahari, S.; Miransari, M. Allelopathic effects of rice cultivars on the growth parameters of different rice cultivars. International Journal of Biological Chemistry 2009, 3, 56–70. [Google Scholar] [CrossRef]
- Babu, G. P,; Vinita, H,; Audiseshamma, K.; Paramageetham, C. Allelopathic effects of some weeds on germination and growth of Vigna mungo (L). Hepper. Int J Curr Microbiol App Sci 2014, 3, 122–128. [Google Scholar]
- Mubeen, K.; Nadeem, M.A.; Tanveer, A.; Zahir, Z.A. Allelopathic effect of aqueous extracts of weeds on the germination and seedling growth of rice (Oryza sativa L.). Pakistan Journal of Life and Social Sciences. 2011, 9, 7–12. [Google Scholar]
- Mehmood, A.; Tanveer, A.; Nadeem, M.A.; Zahir, Z.A. Comparative allelopathic potential of metabolites of two Alternanthera species against germination and seedling growth of rice. Planta Daninha 2014, 32, 1–10. [Google Scholar] [CrossRef]
- Joshi, N.; Joshi, A. Allelopathic effects of weed extracts on germination of wheat. Annals of Plant Sciences 2016, 5, 1330–1334. [Google Scholar] [CrossRef]
- MADA. Rice R&D @ MARDI. 2019. Available online: http://www.mada.gov.my/wp- content/uploads/2019/09/lawatan-delegasi-filipina2-edited-140119-1.pdf (accessed on 21 January 2020).
- Krishna, A.; Ramana, P.; Talekar, M. Allelopathic effect of weed extracts on seed germination of paddy cultivars. Karnataka Journal of Agricultural Science 2006, 20, 671–673. [Google Scholar]
- Ilori, O.J.; Otusanya, O.O.; Adelusi, A.A.; Sanni, R.O. Allelopathic activities of some weeds in the Asteraceae family. International Journal of Botany 2010, 6, 161–163. [Google Scholar] [CrossRef]
- Nishida, N.; Tamotsu, S.; Nagata, N.; Saito, C.; Sakai, A. Allelopathic effects of volatile monoterpenoids produced by Salvia leucophylla: inhibition of cell proliferation and DNA synthesis in the root apical meristem of Brassica campestris seedlings. Journal of Chemical Ecology 2005, 31, 1187–1203. [Google Scholar] [CrossRef]
- Rietjens, I.M.; Alink, G.M. Nutrition and health-toxic substances in food. Ned. Tijdschr Geneeskd 2003, 147, 2365–2370. [Google Scholar]
- Sahu, A.; Devkota, A. Allelopathic effects of aqueous extract of leaves of Mikania micrantha HBK on seed germination and seedling growth of Oryza sativa L. and Raphanus sativus L. Scientific World 2013, 11, 91–93. [Google Scholar] [CrossRef]
- Mahmoodzadeh, H.; Mahmoodzadeh, M. Allelopathic effects of Cynodon dactylon L. on germination and growth of Triticum aestivum. Annals of Biological Research 2013, 5, 118–123. [Google Scholar]
- Khaliq, A.; Matloob, A.; Khan, M.B.; Tanveer, A. Differential suppression of rice weeds by allelopathic plant aqueous extracts. Planta Daninha 2013, 31, 21–28. [Google Scholar] [CrossRef]
- Verma, M.; Rao, P.B. Allelopathic effect of four weed species extracts on germination, growth and protein in different varieties of Glycine max (L.) Merrill. Journal of Environmental Biology 2006, 27, 571–577. [Google Scholar]
- Macías, F.A.; Galindo, J.C.G.; Massanet, G.M. Potential allelopathic activity of several sesquiterpene lactone models. Phytochemistry 1992, 31, 1969–1977. [Google Scholar] [CrossRef]
- Harun, R.; Serin, T.; A. Man. Economic evaluation of Clearfield® Paddy Production System. Economic and Technology Management Review 2013, 8, 47–57. [Google Scholar]
- Ruzmi, R.; Ahmad-Hamdani, M.S.; Abidin, M.Z.Z.; Burgos, N.R. Evolution of imidazolinone-resistant weedy rice in Malaysia: The current status. Weed Science 2021, 1–11. [Google Scholar] [CrossRef]
- Katoch, R.; Thakur, M.; Paul, Y.S. Antifungal activity of the essential oils of Chromolaena adenophorum, Ageratum conyzoides and Lantana camara. Indian Phytopathology 2012, 65, 409–411. [Google Scholar]
- Usuah, P.E.; Udom, G.N.; Edem, I.D. Allelopathic effect of some weeds on the germination of seeds of selected crops grown in Akwa Ibom State, Nigeria. World Journal of Agricultural Research 2013, 1, 59–64. [Google Scholar]
- Fatunbi, A.O.; Dube, S.; Yakubu, M.T.; Tshabalala, T. Allelopathic potential of Acacia mearnsii De wild. World Applied Sciences Journal 2009, 7, 1488–1493. [Google Scholar]
- Begum, M.; Salam, M.A.; Zaman, F. Allelopathic effect of siam weed debris on seed germination and seedling growth of three test crop species. Acta Scientifica Malaysia (ASM) 2021, 5, 1–4. [Google Scholar] [CrossRef]
- Koodkaew, I.; Rottasa, R. Allelopathic effects of giant sensitive plant (Mimosa pigra) leaf powder on germination and growth of popping pod and purslane. International Journal of Agricultural Biology 2017, 18, 1113–1118. [Google Scholar] [CrossRef]
- Ismail, B.S.; Tan, P.W.; Chuah, T.S. Assessment of the potential allelopathic effects of Pennisetum purpureum Schumach. on the germination and growth of Eleusine indica (L.) gaertn. Sains Malaysiana 2015, 44, 269–274. [Google Scholar] [CrossRef]
- Ismail, B.S.; Kumar, A. Effects of aqueous extracts and residues decomposition of Mikania micrantha H.B.K. on selected crops. Allelopathy Journal 1996, 3, 195–206. [Google Scholar]
- Farooq, M.; Jabran, K.; Cheema, Z.A.; Wahid, A.; Siddique, K.H. The role of allelopathy in agricultural pest management. Pest Management Science 2011, 67, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Bajwa, A.A.; Cheema, S.A.; Cheema, Z.A. Application of allelopathy in crop production. International Journal of Agriculture and Biology 2013, 15, 1367–1378. [Google Scholar]
- Yaseen, S.E.; Mutwali, E.M. Allelopathic effect of Datura stramonium on germination and some growth parameters of swiss chard (Beta vulgaris var. cicla). International Journal of Innovative Science and Research Technology 2018, 3, 726–730. [Google Scholar]
- Hegab, M.M.; Gabr, M.A.; Al-Wakeel, S.A.M.; Hamed, B.A. Allelopathic potential of Eucalyptus rostrata leaf residue on some metabolic activities of Zea mays L. Universal Journal of Plant Science 2016, 4, 11–21. [Google Scholar]
- Siddique, M.A.B.; Ismail, B.S. Allelopathic effects of Fimbristylis miliacea on the physiological activities of five Malaysian rice varieties. Australian Journal of Crop Science 2013, 7, 2062–2067. [Google Scholar]
- Bottery, B.R.; Bozzell, R.I. 1977 The relationship between chlorophyll content and rate of photosynthesis in soybean. Canadian Journal of Plant Science 1977, 57, 1–5. [Google Scholar] [CrossRef]
- Yang, D.L.; Jing, R.L.; Chang, X.P.; Li, W. Quantitative trait loci mapping for chlorophyll fluorescence and associated traits in wheat (Triticum aestivum). Journal of Integrative Plant Biology 2007, 49, 646–654. [Google Scholar] [CrossRef]
- Hussain, M.I.; Reigosa, M.J. A chlorophyll fluorescence analysis of photosynthetic efficiency, quantum yield and photon energy dissipation in PSII antennae of Lactuca sativa L. leaves exposed to cinnamic acid. Plant Physiology and Biochemistry 2011, 49, 1290–1298. [Google Scholar] [CrossRef]
- Ilori, O.J.; Otusanya, O.O. Changes in chlorophyll and protein contents of Vigna unguiculata, Glycine max, Zea mays and Sorghum bicolor raised in soil incorporated with the shoots of Tithonia rotundifolia. Journal of Advances in Biology & Biotechnology 2020, 23, 33–39. [Google Scholar]
- Krieger, R. Handbook of Pesticide Toxicology: Principles and Agents. Academic Press: Oxford, 2001.
Figure 1.
Evaluation of allelopathic potential of T. procumbens root exudates on the radicle growth of WR3 weedy variant (b), compared to control (a).
Figure 1.
Evaluation of allelopathic potential of T. procumbens root exudates on the radicle growth of WR3 weedy variant (b), compared to control (a).
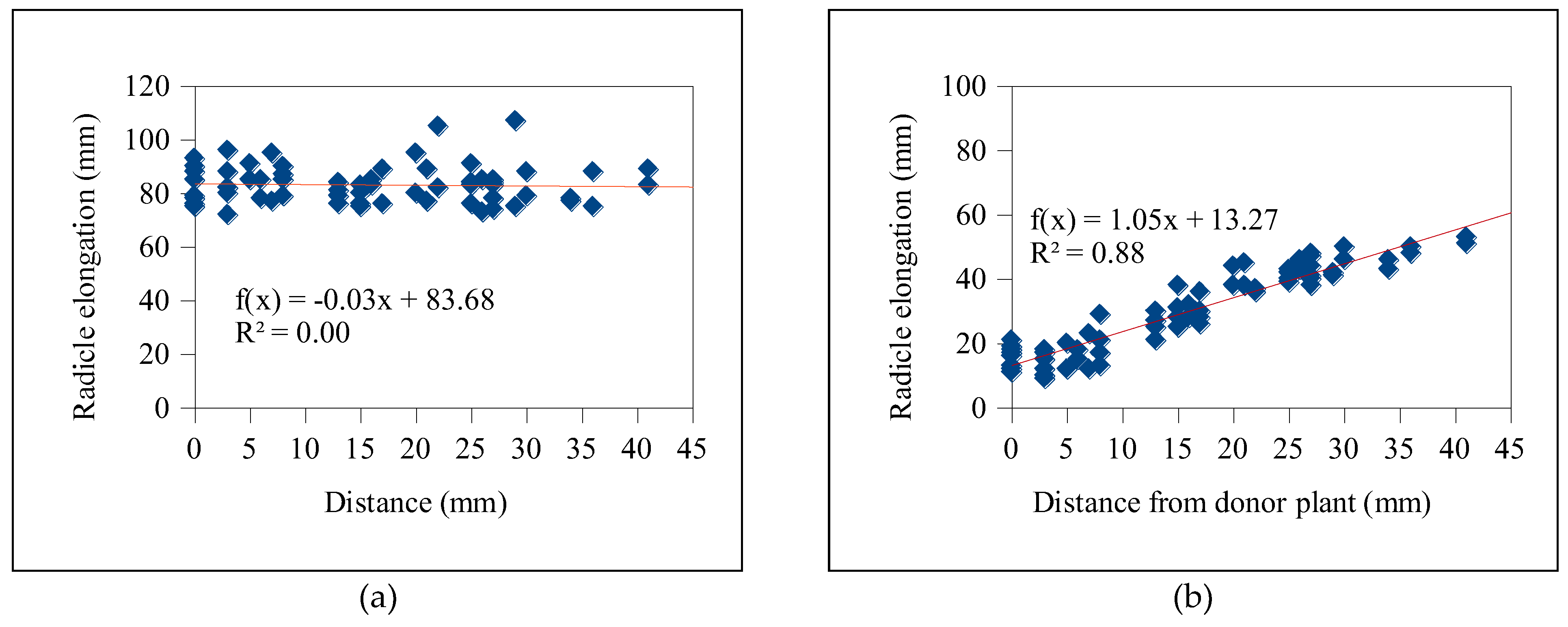
Figure 1.
Evaluation of allelopathic potential of T. procumbens root exudates on the radicle growth of MR220 CL2 (b), compared to control (a).
Figure 1.
Evaluation of allelopathic potential of T. procumbens root exudates on the radicle growth of MR220 CL2 (b), compared to control (a).
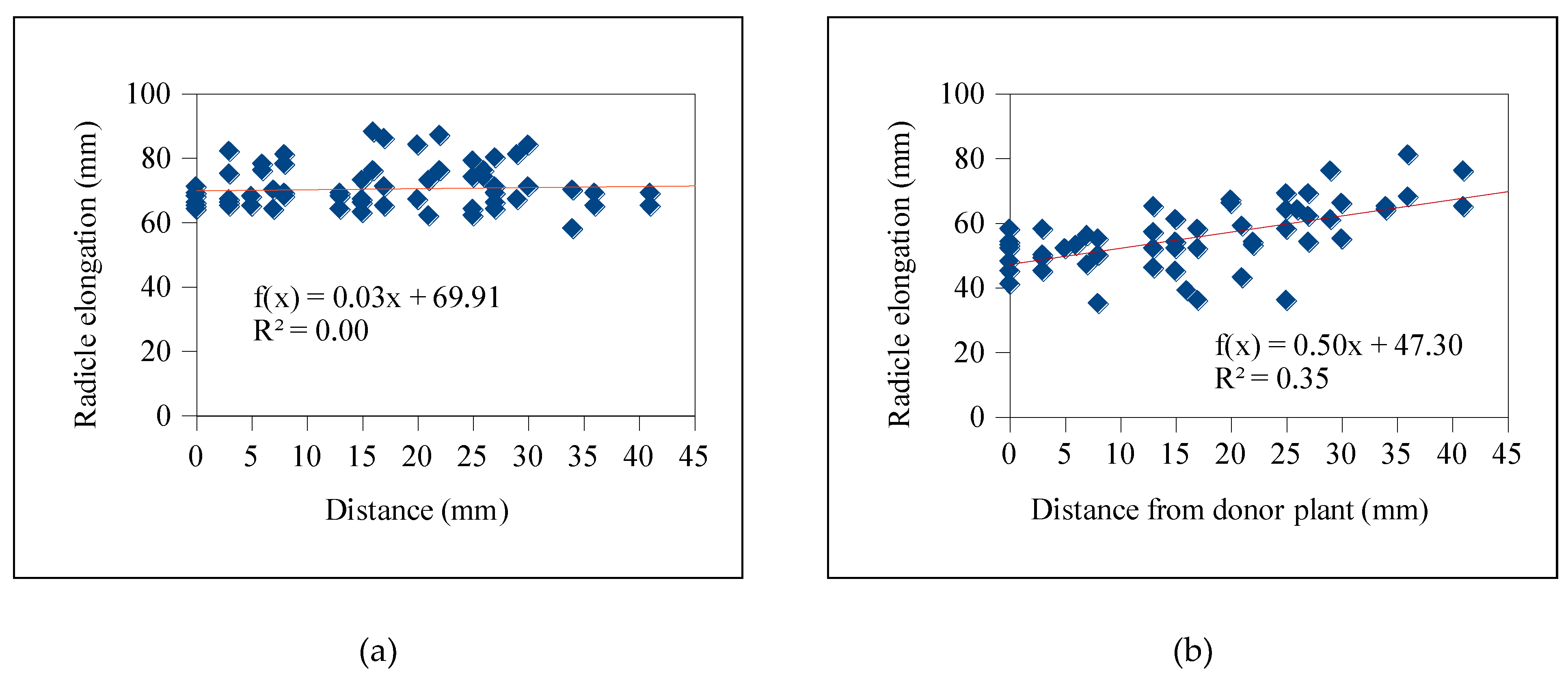
Figure 3.
Allelopathic effects of the aqueous extract of T. procumbens leaves at different concentrations on the hypocotyl and radicle growth of (a) WR3, (b) WR7, (c) Sebernas 307 and (d) MR220 CL2. Bars (mean ± standard error) with same alphabet are not significantly different (p< 0.05) according to Duncan's Multiple Range Test.
Figure 3.
Allelopathic effects of the aqueous extract of T. procumbens leaves at different concentrations on the hypocotyl and radicle growth of (a) WR3, (b) WR7, (c) Sebernas 307 and (d) MR220 CL2. Bars (mean ± standard error) with same alphabet are not significantly different (p< 0.05) according to Duncan's Multiple Range Test.
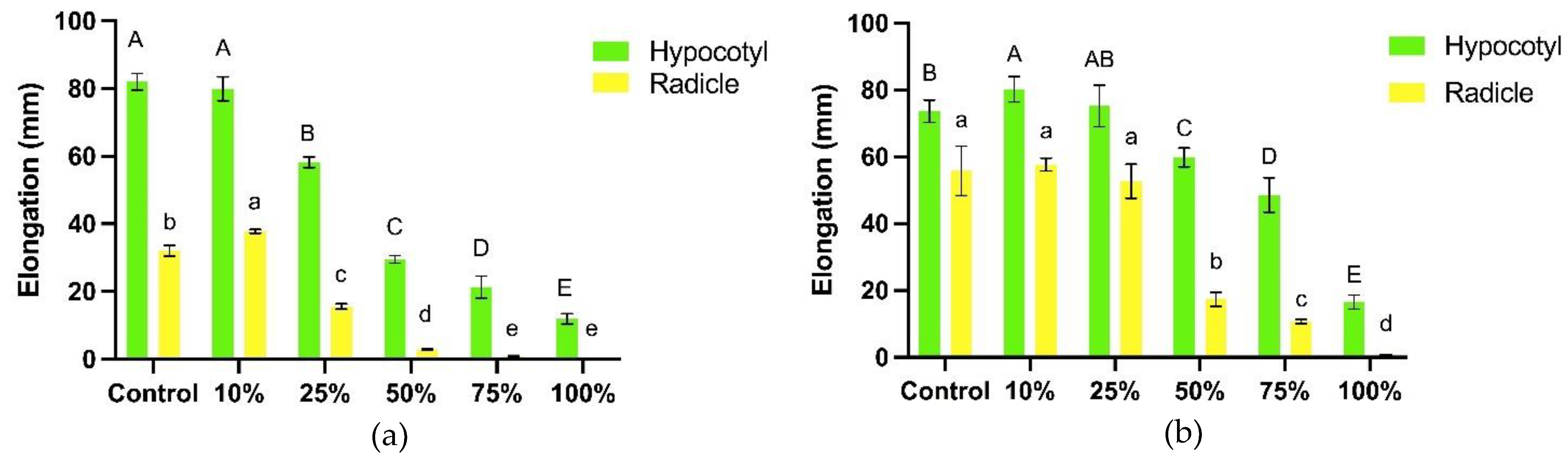
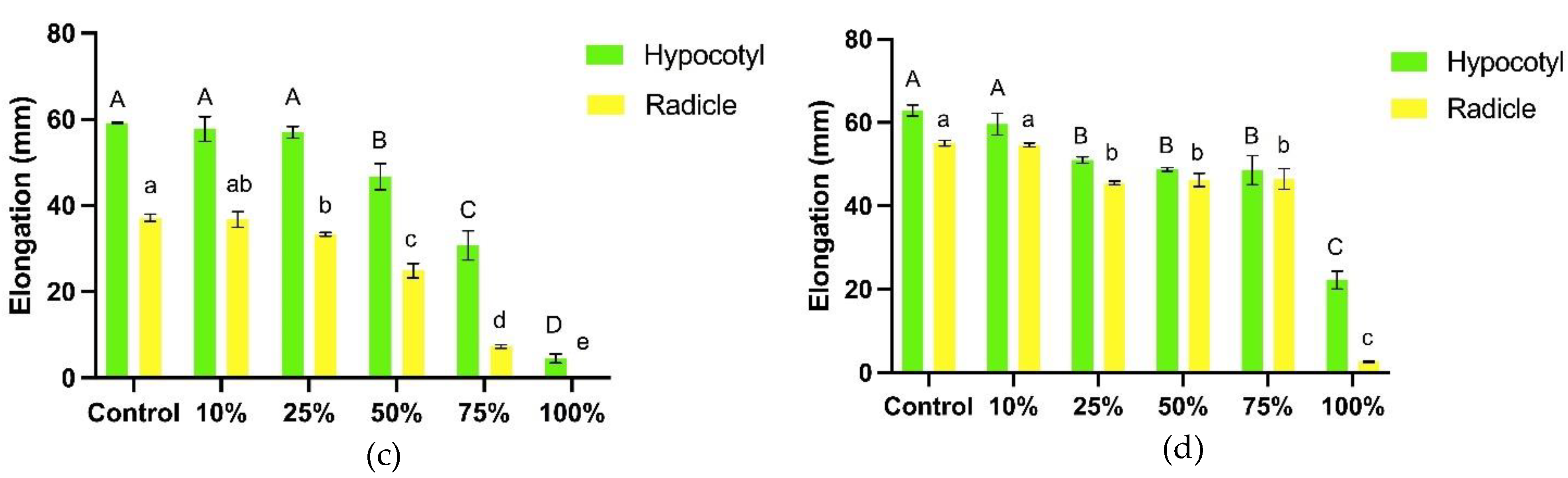
Figure 4.
Allelopathic effects of the aqueous extract of T. procumbens leaves at different concentrations on the fresh and dry weight of (a) WR3, (b) WR7, (c) Sebernas 307 and (d) MR220 CL2. Bars (mean ± standard error) with same alphabet are not significantly different (p< 0.05) according to Duncan's Multiple Range Test.
Figure 4.
Allelopathic effects of the aqueous extract of T. procumbens leaves at different concentrations on the fresh and dry weight of (a) WR3, (b) WR7, (c) Sebernas 307 and (d) MR220 CL2. Bars (mean ± standard error) with same alphabet are not significantly different (p< 0.05) according to Duncan's Multiple Range Test.
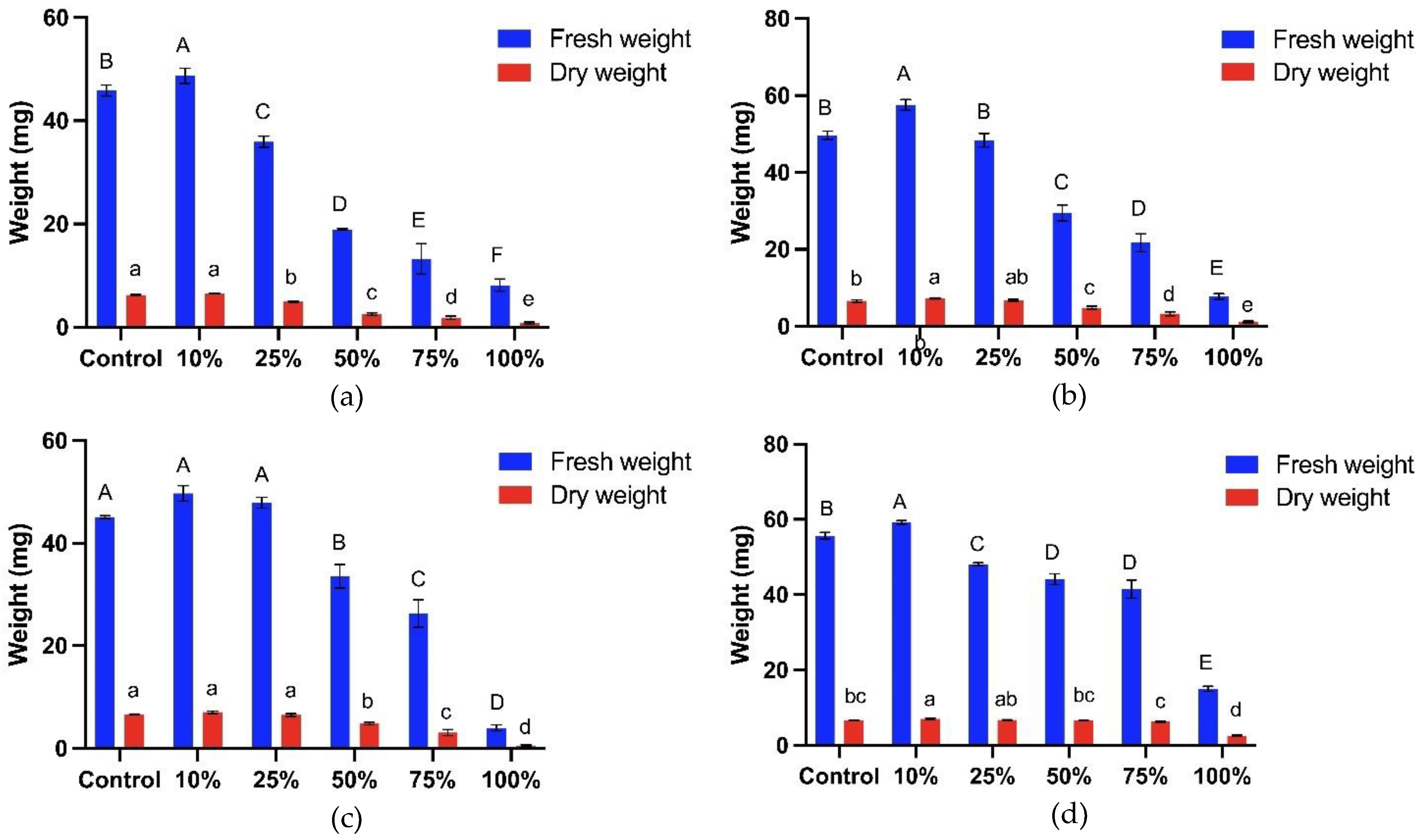
Figure 7.
Allelopathic effects of the debris of T. procumbens leaves incorporated into soil at different concentrations on the GP and GR of (a) WR3, (b) WR7, (c) Sebernas 307 and (d) MR220 CL2. Abbreviations: GP, percentage of germination and GR, germination rate.
Figure 7.
Allelopathic effects of the debris of T. procumbens leaves incorporated into soil at different concentrations on the GP and GR of (a) WR3, (b) WR7, (c) Sebernas 307 and (d) MR220 CL2. Abbreviations: GP, percentage of germination and GR, germination rate.
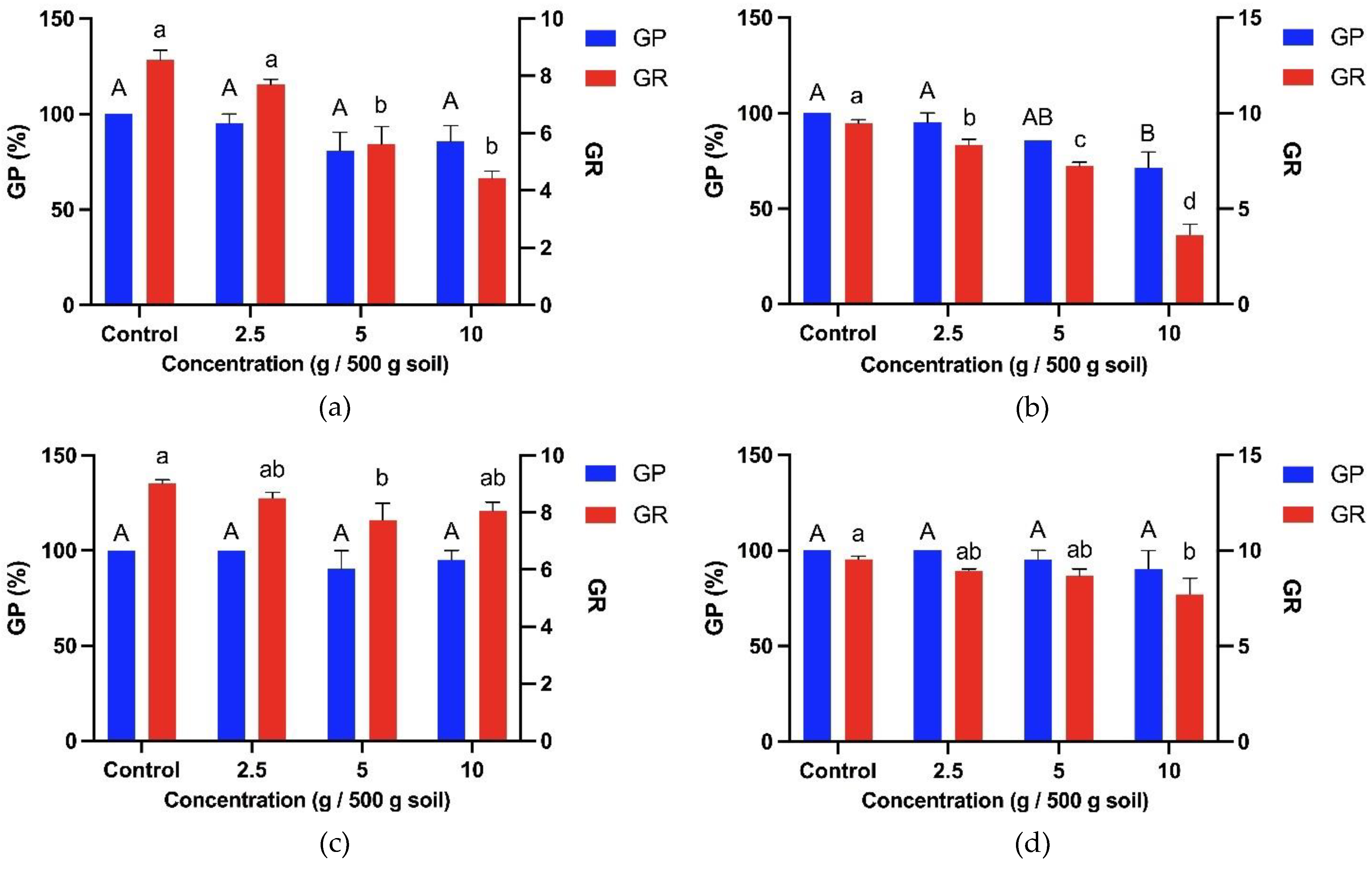
Figure 8.
Allelopathic effects of debris of T. procumbens leaves incorporated into soil at different concentrations on the seedling height of bioassay plants. Bars (mean ± standard error) with same alphabet are not significantly different (p< 0.05) according to Duncan's Multiple Range Test.
Figure 8.
Allelopathic effects of debris of T. procumbens leaves incorporated into soil at different concentrations on the seedling height of bioassay plants. Bars (mean ± standard error) with same alphabet are not significantly different (p< 0.05) according to Duncan's Multiple Range Test.
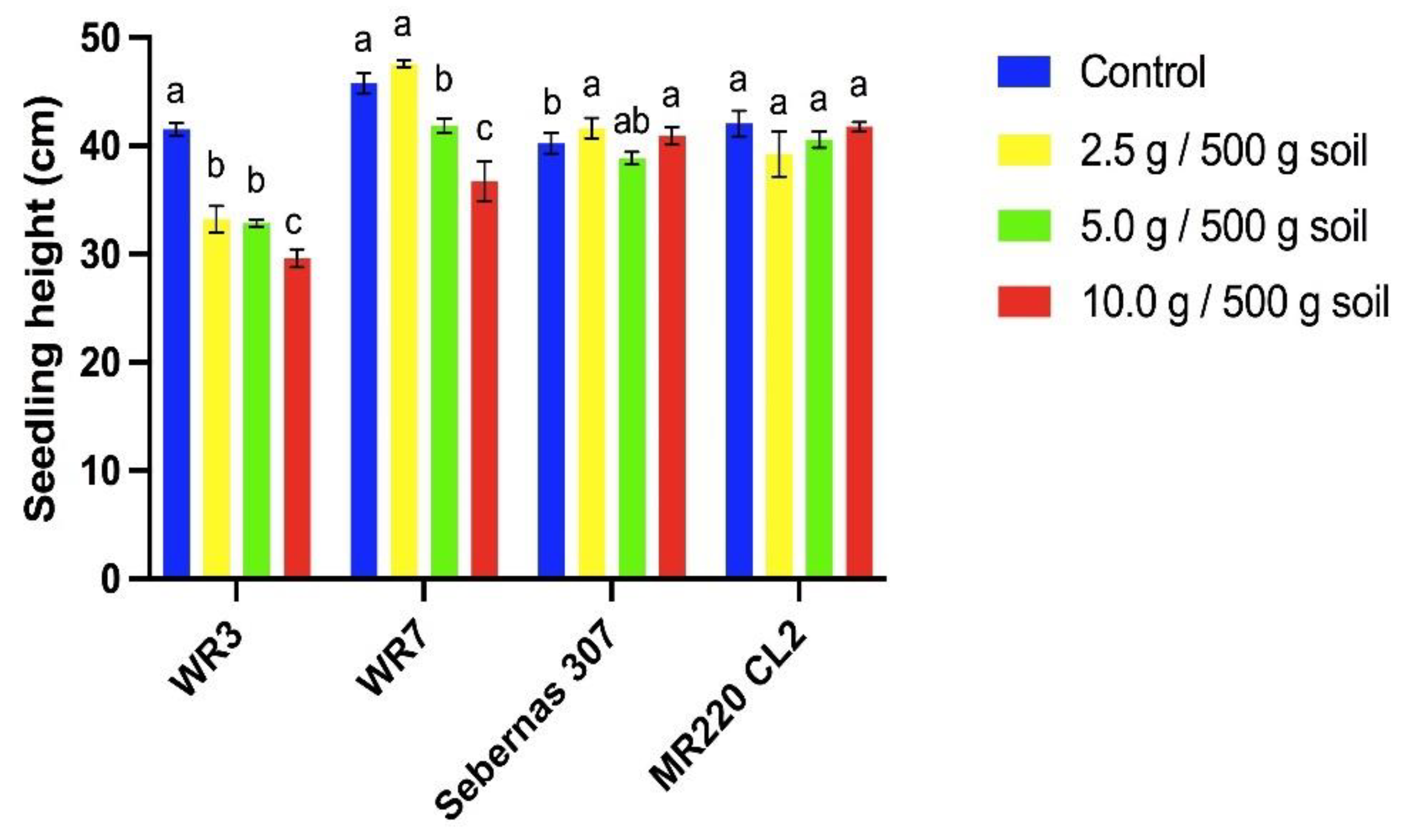
Figure 9.
Allelopathic effects of the debris of T. procumbens leaves incorporated into soil at different concentrations on the fresh and dry weight of (a) WR3, (b) WR7, (c) Sebernas 307 and (d) MR220 CL2. Bars (mean ± standard error) with same alphabet are not significantly different (p< 0.05) according to Duncan's Multiple Range Test.
Figure 9.
Allelopathic effects of the debris of T. procumbens leaves incorporated into soil at different concentrations on the fresh and dry weight of (a) WR3, (b) WR7, (c) Sebernas 307 and (d) MR220 CL2. Bars (mean ± standard error) with same alphabet are not significantly different (p< 0.05) according to Duncan's Multiple Range Test.
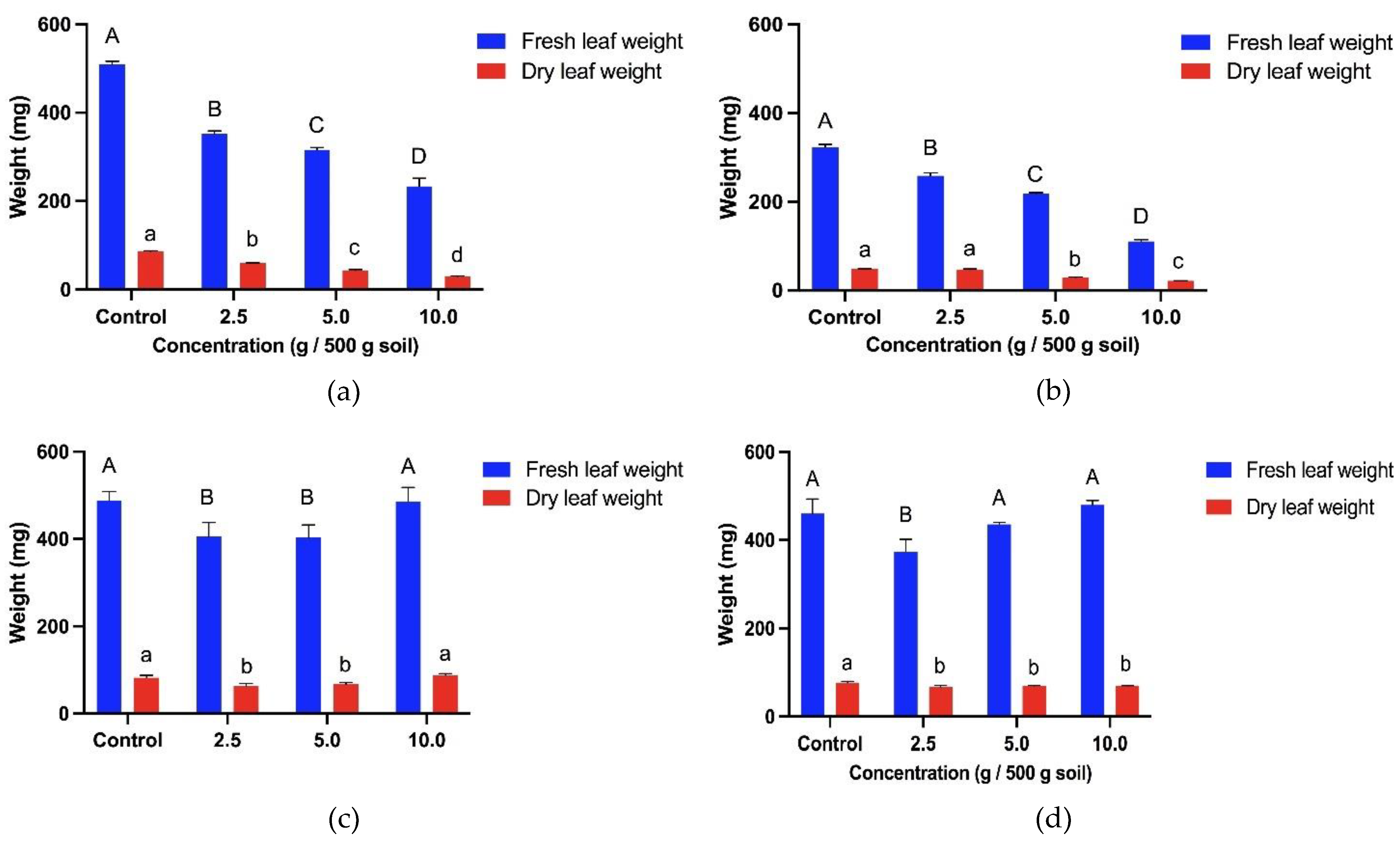
Figure 10.
Allelopathic effects of the debris of T. procumbens leaves incorporated into soil at different concentrations on the chlorophyll content of (a) WR3, (b) WR7, (c) Sebernas 307 and (d) MR220 CL2. Bars (mean ± standard error) with same alphabet are not significantly different (p< 0.05) according to Duncan's Multiple Range Test.
Figure 10.
Allelopathic effects of the debris of T. procumbens leaves incorporated into soil at different concentrations on the chlorophyll content of (a) WR3, (b) WR7, (c) Sebernas 307 and (d) MR220 CL2. Bars (mean ± standard error) with same alphabet are not significantly different (p< 0.05) according to Duncan's Multiple Range Test.
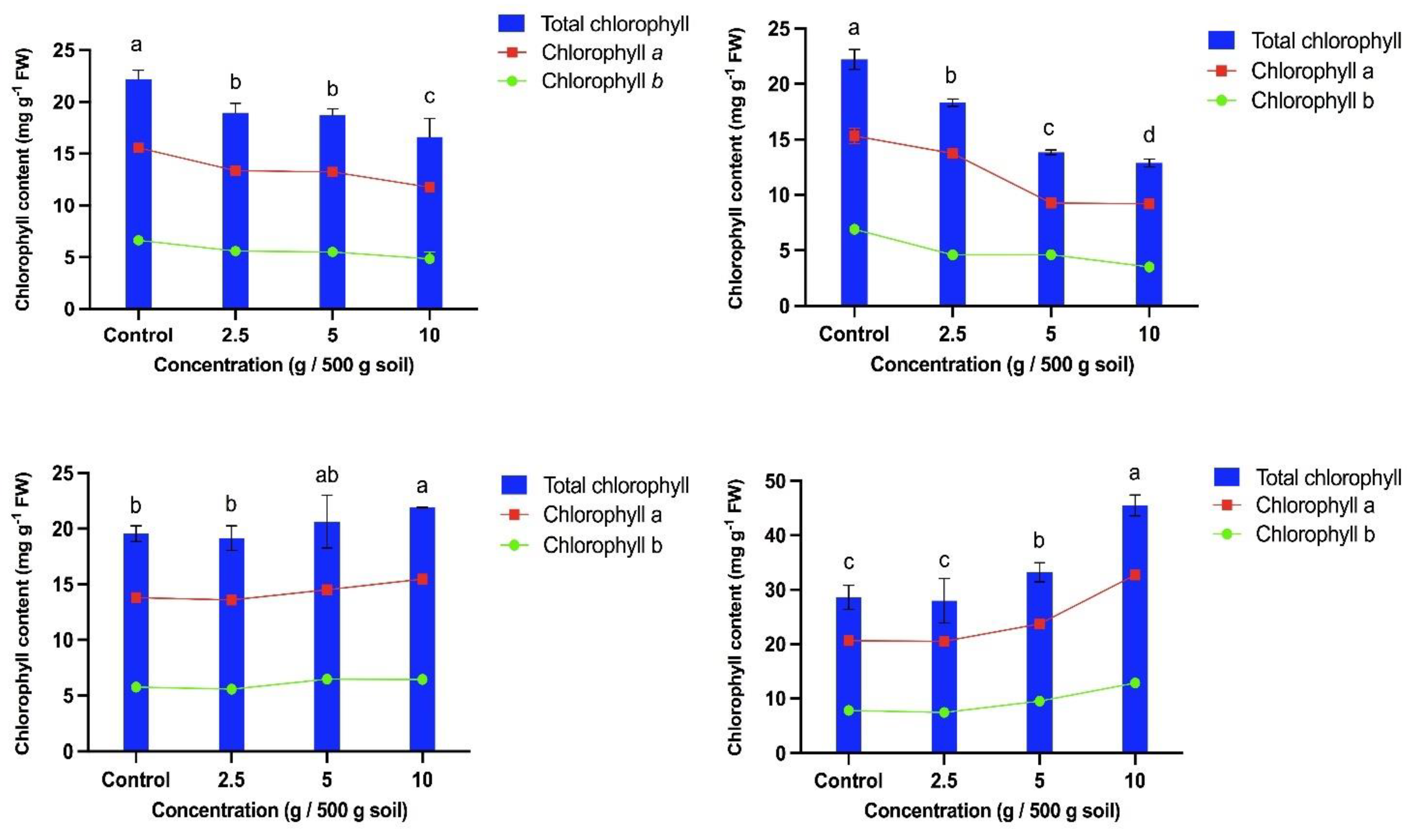

Table 1.
Designation, sampling locations and the morphological characteristics of bioassay plants.
| Designation | Rice type | Sampling location | Coordinates | Morphological characteristics |
|---|---|---|---|---|
| WR2 | Weedy variant | Kg. Sawah Sempadan | 3.453°N 101.214°E | Awnless; straw-hulled; open panicle |
| WR3 | Weedy variant | Kg. Pasir Panjang | 3.567°N 101.067°E | Awnless; straw-hulled; closed panicle |
| WR4 | Weedy variant | Telok Mesira, Bachok. | 6.143°N 102.343°E | Awnless; brown-hulled; open panicle |
| WR5 | Weedy variant | Jln. Tanjong, Tawang, Bachok | 6.102°N 102.321°E | Awless; brown-hulled; closed panicle |
| WR6 | Weedy variant | Kg. Tok Ajam, Pasir Puteh | 5.9038°N 102.374°E | Awned; straw-hulled; open panicle |
| WR7 | Weedy variant | Kg. Titi Serong, Parit Buntar | 5.099°N 100.471°E | Awned; brown-hulled; closed panicle |
| MR 220 CL2 | Modern cultivar | Gene and Seed Bank Center, MARDI Seberang Perai | - | - |
| Sebernas 307 | Modern cultivar | Gene and Seed Bank Center, MARDI Seberang Perai | - | - |
| MR 297 | Modern cultivar | Gene and Seed Bank Center, MARDI Seberang Perai | - | - |
Table 2.
Parameters for GC-MS analysis.
| Parameter | Description |
|---|---|
| Model | DB-5MS-UI |
| Column length (m) | 30 |
| Diameter (mm) | 0.25 |
| Stationary phase | 5% phenyl methylpolysiloxane |
| Film width (µm) | 0.25 |
| Company | Agilent Technologies |
Table 3.
Inhibitory effect of foliar leachate of T. procumbens on the radicle growth of bioassay plants.
Table 3.
Inhibitory effect of foliar leachate of T. procumbens on the radicle growth of bioassay plants.
| Rice type | Bioassay | Radicle inhibition percentage (%) | Mean inhibition percentage (%) | Rank | |
|---|---|---|---|---|---|
| 10 mg | 50 mg | ||||
| Weedy rice | WR2 | 30.93 ab | 93.24 a | 62.09 | 2 |
| WR3 | 36.09 a | 97.31 a | 66.70 | 1 | |
| WR4 | 29.68 ab | 76.65 b | 53.17 | 4 | |
| WR5 | 26.29 bc | 91.63 a | 58.96 | 3 | |
| WR6 | 9.58 de | 94.67 a | 52.13 | 5 | |
| WR7 | 6.89 e | 53.57 d | 30.23 | 7 | |
| Modern cultivars |
MR220 CL2 | 8.67 de | 25.43 e | 17.05 | 9 |
| Sebernas 307 | 14.17 cde | 27.34 e | 20.76 | 8 | |
| MR297 | 21.34 bcd | 64.68 c | 43.01 | 6 | |
Table 4.
Inhibitory effect of foliar leachate of T. procumbens on the hypocotyl growth of bioassay plants.
Table 4.
Inhibitory effect of foliar leachate of T. procumbens on the hypocotyl growth of bioassay plants.
| Rice type | Bioassay | Hypocotyl inhibition percentage (%) | Mean inhibition percentage (%) | Rank | |
|---|---|---|---|---|---|
| 10 mg | 50 mg | ||||
| Weedy rice | WR2 | 19.83 a | 33.10 bc | 26.47 | 4 |
| WR3 | 18.93 a | 61.44 a | 40.19 | 1 | |
| WR4 | 19.83 bc | 33.10 bc | 26.47 | 3 | |
| WR5 | 8.87 ab | 22.75 cd | 15.81 | 6 | |
| WR6 | 10.76 a | 61.02 a | 35.89 | 2 | |
| WR7 | 13.01 a | 22.22 cd | 17.62 | 5 | |
| Modern cultivars |
MR220 CL2 | 14.16 a | 12.43 de | 13.30 | 7 |
| Sebernas 307 | -9.55 c | -5.20 f | -7.38 | 9 | |
| MR297 | -9.89 c | 5.37 ef | -2.26 | 8 | |
Table 5.
Inhibitory effect of root exudates of T. procumbens on the radicle elongation of bioassay plants.
Table 5.
Inhibitory effect of root exudates of T. procumbens on the radicle elongation of bioassay plants.
| Bioassay | Radicle elongation(mm) | Radicle elongation (%) | Correlation Pearson | Correlation coefficient, R2 | |
|---|---|---|---|---|---|
| Control | Exudates | ||||
| WR2 | 65.8 ± 0.46 | 29.0 ± 1.50 | 44.1 ** | 0.80 ** | 0.64 |
| WR3 | 82.8 ± 0.91 | 30.5 ± 1.60 | 36.8 ** | 0.94 ** | 0.88 |
| WR4 | 82.9 ± 0.83 | 56.3 ± 2.16 | 67.9 ** | 0.88 ** | 0.78 |
| WR5 | 73.9 ± 0.47 | 46.4 ± 2.14 | 62.8 ** | 0.93 ** | 0.87 |
| WR6 | 85.0 ± 0.78 | 41.7 ± 3.07 | 49.1 ** | 0.94 ** | 0.86 |
| WR7 | 68.8 ± 0.54 | 42.9 ± 2.20 | 62.3 ** | 0.90 ** | 0.81 |
| MR220 CL2 | 70.5 ± 0.85 | 55.7 ± 1.17 | 79.2 ** | 0.53 ** | 0.35 |
| MR297 | 69.3 ± 0.58 | 44.8 ± 1.82 | 64.8 ** | 0.90 ** | 0.81 |
| Sebernas 307 | 85.9 ± 0.89 | 49.8 ± 0.92 | 58.0 ** | 0.77 ** | 0.60 |
Radicle elongation is the mean ± standard error of the three replicates. The percentage value was based on the difference from the elongation value of the control, and the difference was tested using an independent sample t-test. The value of percent elongation of radicles marked with *, ** was significant at the p<0.05, 0.01 level based on independent sample t-test. The correlation values marked *, ** were significant based on Pearson correlation analysis at a significant level of p <0.05, 0.01.
Table 6.
Allelochemicals identified in T. procumbens leaves by GC-MS.
| No. | Retention time (min) | Peak area (%) | Chemical compounds | Molecular formula | Molecular weight |
|---|---|---|---|---|---|
| 1 | 12.24 | 8.36 | cis-Pinane | C10H8 | 138.25 |
| 2 | 12.53 | 1.37 | 1,8-Nonadiene, 2,8-dimethyl- | C11H20 | 152.28 |
| 3 | 12.66 | 2.59 | 7-Tridecanone | C13H26O | 198.34 |
| 4 | 12.77 | 2.39 | 3,7,11,15-Tetramethyl-2-hexadecen-1-ol | C20H40O | 296.50 |
| 5 | 13.35 | 2.04 | Hexadecanoic acid, methyl ester | C17H34O2 | 270.50 |
| 6 | 13.85 | 5.02 | n-Hexadecanoic acid | C16H32O2 | 256.42 |
| 7 | 14.67 | 1.98 | β-D-Mannofuranoside, farnesil- | C21H36O6 | 384.50 |
| 8 | 14.89 | 12.46 | Hexadecanoic acid, trimethylsilyl ester | C19H40O2Si | 328.60 |
| 9 | 15.69 | 2.47 | 9,12,15-Octadecatrienoic acid, methyl ester, (Z,Z,Z)- | C19H32O2 | 292.50 |
| 10 | 15.86 | 6.65 | Phytol | C20H40O | 296.50 |
| 11 | 16.20 | 2.41 | 9,12-Octadecadienoic acid (Z,Z)- | C18H30O | 278.43 |
| 12 | 16.29 | 6.22 | 9,12,15-Octadecatrienoic acid (Z,Z,Z)- | C18H30O2 | 278.40 |
| 13 | 16.63 | 1.51 | Silane, [(3,7,11,15-tetramethyl-2-hexadecenyl)oxi]trimethyl- | C23H48OSi | 368.70 |
| 14 | 17.20 | 2.59 | 9,12-Octadecadienoic acid (Z,Z)-, trimethylsilyl ester | C21H40O2Si | 352.60 |
| 15 | 17.30 | 4.42 | α-Linolenic acid, trimethylsilyl ester | C21H38O2Si | 350.61 |
| 16 | 18.09 | 2.78 | Dispiro[2.2.2.0]Octane, 4,5-trans-diphenyl- | C20H20 | 260.40 |
| 17 | 20.93 | 1.48 | Bis(2-(Dimethyllamino)ethyl) ether | C8H20N20 | 160.26 |
| 18 | 25.88 | 10.63 | Squalene | C30H50 | 410.70 |
| 19 | 26.02 | 11.55 | 2-[4-(4-Fluorocinnamoyl)anilino]-3-piperidino-1,4-naphthoquinon | C30H25FN2O3 | 480.50 |
| 20 | 33.42 | 4.99 | Stigmasterol | C29H48O | 412.70 |
| 21 | 34.99 | 6.09 | β-sitosterol | C29H50O | 414.70 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



