Submitted:
15 October 2024
Posted:
16 October 2024
You are already at the latest version
Abstract
Adventitious root (AR) formation in plants origins from non-root organs such as leaves and hy-pocotyls. Auxin signaling is essential for AR formation, but the role of other phytohormones is less clear. In Arabidopsis, at least two distinct mechanisms can produce ARs either from hypocotyls as part of the general root architecture, or from wounded organs during de novo root regeneration (DNRR). In previous reports, gibberellin acid (GA) seems to play reverse roles in both types of AR formation, since GA treatment blocks AR formation from hypocotyls, whereas depletion of GA signaling results in reduced DNRR from detached leaves. Here, we report the positive effects of GA biosynthesis inhibitor paclobutrazol (PBZ/PAC) treatment on both types of AR formation in Arabidopsis. Consistently, loss of GA synthesis and signaling promoted DNRR in our conditions. Furthermore, PBZ treatment can rescue declined AR formation from leaf explants in erecta recep-tor mutants. Transcriptional profiling revealed that PBZ treatment altered GA, brassinosteroids and auxin responses, which included up-regulation of LBD16 that plays a pivotal role in AR initi-ation. This study reveals that depletion of GA biosynthesis promotes DNRR in Arabidopsis, and thus will contribute to further clarification of the role of GA during AR formation in plants.
Keywords:
Adventitious roots (ARs)
; de novo root regeneration (DNRR)
; Paclobutrazol (PBZ/PAC)
; Blocking of gibberellin (GA) biosynthesis
; LBD16
; LRP1
1. Introduction
Adventitious roots (ARs) origin by definition from non-root tissue and organs. There are different types of ARs, and their overall function, form, and placement vary depending on the species and their individual needs [1]. In the grass family, AR formation often builds up the main part of the root system architecture, whereby the crown roots of maize and rice, which origin secondarily from the stem, are the best investigated examples of ARs in monocots [2]. Members of the plant-specific LATERAL ORGAN BOUNDARIES DOMAIN (LBD) transcription factor family are key regulators of the post-embryonic shoot-borne crown root initiation in rice and maize [3,4,5,6]. Beside grafting [7], AR formation from stem cuttings is the most common method for asexual plant propagation in forestry and agriculture. In the model plant Arabidopsis, there are at least two types of AR formation, wounding-induced de novo root regeneration (DNRR) from detached organs such as leaves and etiolation-induced AR formation in intact hypocotyls [8]. LBD16 is a key regulator of auxin-induced callus induction, lateral root (LR) initiation, and both types of AR formation, and works downstream of AUXIN RESPONSE FACTORs (ARFs) such as ARF7/19 and ARF6/8 [9,10,11,12,13,14]. Loss of LBD16 results in reduced DNRR, initiation of LRs, and AR formation from hypocotyls, demonstrating the general role of LBD16 in initiating secondary roots [12].
Plant development relies on stem cells harbored in meristems and the correct patterning of cell fates during organogenesis [15]. Adventitious rooting requires the accumulation of auxin in competent vasculature-associated pluripotent cells (VPCs) to activate the ARFs leading to the fate transition of these cells to AR founder cells that establish de novo root apical meristems (RAMs) including root stem cells [16,17,18]. Therefore, auxin biosynthesis and transport play a key role in DNRR in Arabidopsis leaf explants [19,20]. In plants, indole-3-acetic acid (IAA) is the main auxin that is primarily synthesized via two chemical reactions [21]. First, the TRYPTOPHAN AMINOTRANSFERASE OF ARABIDOPSIS (TAA) family of amino transferases converts the amino acid tryptophan to indole-3-pyruvate (IPA), which is followed by the conversion of IPA to IAA by the catalysis of flavin-containing monooxygenases encoded by YUCCA (YUC) genes [22,23]. Several YUC genes contribute to AR formation from Arabidopsis leaf explants, while YUC9 expression is primarily induced in response to darkness, and yuc9 single mutants have a rooting defect in the dark but not in the light [19]. To trigger earlier steps of auxin biosynthesis in detached Arabidopsis leaves, wounding-induced jasmonate signaling activates the ETHYLENE RESPONSE FACTORs ERF109 and ERF111, which in turn upregulates the auxin biosynthesis gene ANTHRANILATE SYNTHASE α1 (ASA1) [24,25]. To prevent hypersensitivity to the wounding signaling, SQUAMOSA PROMOTER BINDING PROTEIN-LIKE (SPL) transcription factors, SPL10 and SPL11, and the epigenetic regulator ULTRAPETALA1 (ULT1) attenuate the transcriptional induction of ERF109, and partially other ERFs, while JASMONATE-ZIM-DOMAIN (JAZ) proteins inhibit ERF109 activity by direct protein-protein interaction two hours after detachment of the leaf [24,25,26]. Therefore, jasmonate serves as a transient wound signal that triggers auxin biosynthesis that is required for forming the auxin maximum that stimulates the fate transition of VPCs to become AR founder cells. In contrast, jasmonate serves as a negative regulator of etiolation-induced AR formation from Arabidopsis hypocotyls by crosstalk with cytokinin [27]. The jasmonate signalling pathway inhibits AR initiation from hypocotyls through repression of CYTOKININ OXIDASE/DEHYDROGENASE 1 (CKX1) that encodes the CK-degrading enzyme that decreases the cytokinin content during adventitious rooting [1,27]. Accordingly, jasmonate plays opposite regulatory roles in both types of AR formation at different stages and from different sources through cross-talk with other hormones [1].
The phytohormone gibberellic acid (GA) regulates several key processes in plants that are of significant agricultural importance, such as root and shoot development, flowering, seed germination, and general plant growth [28]. During GA biosynthesis, various precursors and non-bioactive GAs act as intermediates in the conversion to bioactive GAs catalyzed by a group of enzymes including ENT-COPALYL DIPHOSPHATE SYNTHETASE 1/GA REQUIRING 1 (CPS/GA1) and GA 20-oxidases such as GA20ox1/GA5 [29]. GA binds to the soluble receptor protein GIBBERELLIN INSENSITIVE DWARF 1 (GID1), and functions as a molecular glue to promote the formation of the GID1–DELLA complexes, which subsequently recruits the ubiquitin E3 ligase SLEEPY1 to trigger polyubiquitination and degradation of DELLA proteins [30]. As primary target of GA signaling, DELLA proteins modulate gene expression by interacting with other transcription factors such as PHYTOCHROME INTERACTING FACTORs (PIFs) [31,32], BRASSINAZOLE RESISTANT 1 (BZR1) [33,34], JAZs causing the release of MYC2 [35], ARFs [36], and SPLs [37,38], and other chromatin-associated proteins such as histone H2A [39].
GA is widely recognized to inhibit AR formation from tissue cultures, stem cuttings, and leaf explants in a large variety of plant species [40,41,42,43,44,45,46,47]. The few studies that reported increased AR formation by GA treatment described likely indirect effects such as promotion of preexisting root anlagen or GA stimulates AR formation via a mechanism that requires the presence of ethylene [40,48]. Nevertheless, a recent study indicated the requirement of GA and tight regulation of GA signaling through DELLA proteins such as GIBBERELLIC ACID INSENSITIVE (GAI) to promote wounding-induced AR formation in Arabidopsis leaf explants [49]. This conclusion based on the phenotype analysis of several classical and newly identified GA synthesis and signaling mutants in DNRR assays, which did not include GA or other phytohormone treatments [49]. In contrast, GA treatment strongly inhibits etiolation-induced AR formation from hypocotyl [42]. These contrary outcomes might be caused by different mechanisms involved in both types of AR formation or resulted from the different methods that were used in both studies.
The plant growth retardant paclobutrazol (PBZ, PAC, PP333) [(2RS,3RS)-1-(4-chlorophenyl)-4,4-dimethyl-2-(1,2,4-triazol-1-yl)pentan-3-ol] inhibits specifically the three steps in the oxidation of the GA-precursor ent-kaurene to ent-kaurenoic acid catalyzed by ENT-KAURENE OXIDASE/GA REQUIRING 3 (KO/GA3) [50,51,52]. In Arabidopsis, PBZ treatment was frequently employed as GA biosynthesis inhibitor to investigate the role of GA signaling during plant development [53,54,55,56,57,58,59], but PBZ has not been previously used for Arabidopsis AR formation studies. Nevertheless, studies in other plant species demonstrated that PBZ treatment has the opposite effect of GA and promotes AR formation either in the presence of exogenous auxin or during exclusive treatment with PBZ [42,60,61,62,63].
In this study, we report that treatment with the GA biosynthesis inhibitor PBZ promotes etiolation-induced AR formation from hypocotyl and wounding-induced AR formation from leaf explants (DNRR) in Arabidopsis. Accordingly, we found that GA treatment blocked AR formation in both type of adventitious rooting. This seems to be in conflict with a previous report that GA is required for DNRR in Arabidopsis leaf explants [49]. By reanalyzing the classical GA biosynthesis and signaling mutants ga5-1 [64], ga1-4 [65], and gai-1 [66] in Ler-0 background, we found that (i) the receptor-kinase ERECTA (ER) is required for DNRR and (ii) depletion of GA biosynthesis and signaling promotes DNRR rescuing AR formation that is declined in Ler-0 leaf explants. We also found similar reduction of DNRR in er-105 mutants in Col-0 background, which we could rescue by PBZ treatment. Since we found substantial variation in the rate of AR formation in wild-type background among experiments, we employed er-105 mutants as genetic tool to find putative effectors of DNRR by whole-genome transcription profiling. Among the differential expressed genes, we found a significant overlap of PBZ-regulated genes with genes directly bound by the DELLA-targets PIF4, BZR1 and ARF6 [36] and/or regulated by GA [67,68,69] or auxin [67,70]. Among them, we focused on transcription factor genes that are transcriptionally regulated during DNRR and identified several candidate genes including LBD16, a known regulator of DNRR [12]. DNRR assays with an LBD16::GUS plant line confirmed the positive response to PBZ treatment, which was abolished by adding GA or blocking polar auxin transport. We discuss how depleted GA signaling can increase LBD16 expression considering auxin-dependent and auxin-independent pathways, but taking also in account that other candidate genes such as LATERAL ROOT PRIMORDIUM 1 (LRP1) could contribute to the positive effects of PBZ on DNRR.
2. Results
2.1. Blocking of GA Synthesis by PBZ Accelerates AR Formation
In Arabidopsis, PBZ treatment was frequently employed as inhibitor of GA biosynthesis to investigate the role of GA signaling in plant development [53,54,55,56,57,58,59]. However, PBZ treatment was neither used during etiolation-induced AR formation from hypocotyl nor wounding-induced DNRR from leaf explants. In contrast, GA is known to inhibit adventitious rooting from hypocotyl in Arabidopsis [42]. To test the effects of PBZ on AR formation from Arabidopsis hypocotyl, we performed AR formation assays [71]. Wild-type (Col-0) plants were germinated by incubating the seeds in light for 3 - 5 hours before being grown in dark for 3 days to induce hypocotyl elongation. Well-elongated and etiolated seedlings were transferred to various treatments under long day (LD) light conditions for 7 days, and then, the number of ARs were determined (Figure 1A-B). As expected, treatment with 1 µM GA significantly reduced the AR number in wild-type, while PBZ treatment (5 µM) increased the number of ARs, similar as described previously in other plant species [42,61,63]. In line with previous studies [72,73,74,75], we found that PBZ treatment had overall negative effect on plant growth causing smaller leaves and shorter roots (Figure 1A). To exam the effects of PBZ on the main root system, we grew wild-type plants on media supplemented with different concentrations of PBZ (Supplementary Figure S1). We found that the main root length decreased proportionally with increased PBZ concentrations confirming the negative effects of PBZ for main root growth, while the LR number were unaffected. Therefore, only the etiolation-induced AR formation from hypocotyl was positively affected by PBZ in Arabidopsis wild-type plants.
Etiolation-induced AR formation from hypocotyl and wounding-induced DNRR from leaf explants share many features, but they partially show opposite responses to hormone treatments such as with jasmonate [1]. Therefore, we also tested the effects of GA and PBZ on DNRR from leaf explants (Figure 1C-E,D). The response pattern was very similar to that of AR formation from hypocotyl; the rooting rate from leaf explants was significantly decreased by GA treatment and increased by treatment with PBZ. Notably, PBZ treatment did not only promote DNRR but also resulted in thicker ARs (Figure 1D,H). This is in line with previous observations in newly developed roots of other plant species [45,76,77,78,79].
Next, we tested the effects of PBZ on DNRR in time courses and found that PBZ treatments can accelerate AR formation in leaf explants reaching rooting rates of 100 percent after 10 DAC (days after culturing started), while untreated plants display about 45 percent rooting rate (Figure 1F). During DNRR in the dark, auxin biosynthesis by YUC proteins is increased in comparison to light conditions accelerating the rooting rate in wild-type, which can reach almost 100 percent [19]. We therefore performed DNRR assays in darkness and found that AR formation was significantly increased in untreated leaf explants, while additional PBZ treatment can enhance the rooting rates in comparison to the mock treated plants (Figure 1G). Nevertheless, the accelerating effect of PBZ in the dark on AR formation was not always reflected in the rooting rates but in the increased root length caused by earlier root initiation (Figure 2A-B).
2.2. The Positive Effects of Auxin and PBZ Treatment on DNRR Are Distinguishable
Previously, it has been suggest that GA inhibits AR formation by perturbing polar auxin transport [42]. Naphthylphthalamic acid (NPA) is an inhibitor of auxin transport by PIN proteins, which can prevent the formation of the auxin maximum and ARs in Arabidopsis leaf explants during DNRR [20]. To compare the effects of GA and PBZ treatment with the effects of exogenously applied auxin and NPA on AR formation from Arabidopsis leaf explants, we performed DNRR assays under dark conditions (Figure 2).
Culturing in darkness up-regulates PIF genes that triggers dark-induced senescence in Arabidopsis leaf explants, which is inhibited in whole darkened plants [80,81]. We observed that GA treatment could accelerate dark-induced senescence in our DNRR assays (Figure 2A). This is in line with GA promoting PIF function by degrading DELLA proteins [82]. Therefore, senescence might play a minor role in reduced AR formation by GA under dark conditions. At 7 DAC, the rooting rate of IAA-treated leaves reached over 20%, while the rooting rate of other treatments remained under 10%. The IAA driven rooting rate remained significantly stronger than the other treatments and reached over 90% at 11 DAC. In view of the rooting capacity, the enhanced DNRR activity of IAA treated leaves was even more eminent (Figure 2C). PBZ-treated leaf explants had significantly longer roots than the mock treated leaf explants, which might reflect earlier AR initiation (Figure 2A). However, PBZ had negative effects on the root capacity (Figure 2A-C). Therefore, IAA and PBZ treatment resulted in clearly distinguishable phenotypes. IAA strongly increased the root number, while PBZ promoted an early emergence of a single AR (Figure 2A). This is in line with the assumption that reduced GA signaling by PBZ accelerates auxin transport, but do not significantly increase the IAA content that would result in more ARs per leaf explant. In contrast, the AR phenotypes of NPA and GA treatment were similar but not identical. GA treatment caused significantly decreased rooting rates and capacities, while NPA treatment blocked all DNRR activity (Figure 2B-C). Therefore, GA might decrease auxin transport, but does not block the transport entirely.
We nevertheless hypnotized that polar auxin transport is essential for the acceleration of AR formation by PBZ during DNRR. To test this hypothesis, NPA+PBZ treatment was performed. As expect, the rooting rate of PBZ-treated leaf explants increased faster than in wild-type, while NPA-treatment blocked all rooting, which was also the case during NPA+PBZ treatment indicating that polar auxin transport remains essential for DNRR in the presence of PBZ (Figure 2D-E). We also tested the effects of IAA+PBZ double treatment on DNRR. Although IAA and PBZ treatment resulted in clearly distinguishable rooting phenotypes (Figure 2A), both treatments can accelerate the rooting rate from leaf explants (Figure 2F). However, the acceleration of the rooting rate by IAA is stronger and epistatic to the weaker effects by PBZ, while PBZ decreased the rooting capacity also in the presents of IAA (Figure 2F-G). Thus, IAA+PBZ double treatment did not show additive effects on the rooting rates, which is in line with the hypothesis that the accelerating effects by PBZ on rooting rates is mainly based on increased auxin transport and/or signaling. On the other hand, the negative effect of PBZ on rooting capacity can only partially be compensated by exogenous auxin treatment (Figure 2G).
2.3. Loss of GA Biosynthesis Enzymes and GA Signaling Overcome Eradication of AR Formation in Erecta Mutant Leaf Explants
Recently, it has been reported that Arabidopsis leaf explants of the classical GA biosynthesis mutant ga5-1 and GA signaling mutant gai-1, which are both in Landsberg erecta (Ler-0) background, had an lower rooting rate and capacity than the ecotype Col-0 that served as wild-type control [49]. The ga5-1 mutant contains a G to A point mutation that inserts a translational stop codon in the protein-coding sequence of GA20ox1, which encodes an enzyme that catalyzes the final step of GA biosynthesis [64], while the gai-1 mutant expresses a dominant version of a DELLA protein that is a negative regulator of GA signaling [66,83]. Therefore, both mutants have strongly depleted GA signaling similar to plants treated with PBZ. Nonetheless, the GA mutants and PBZ treatment seemed to have opposite effects on AR formation from Arabidopsis leaf explants [49]. In order to clarify these contrary findings, we performed DNRR assays with gai-1, ga5-1, and another GA biosynthesis mutant ga requiring 1-4 (ga1-4) in Ler-0 background [65], while Ler-0 served as wild-type control (Figure 3A). Surprisingly, we found that almost all activity of AR formation was eliminated in Ler-0 leaf explants suggesting that the ER receptor is a positive regulator of DNRR. In contrast, ga1-4, ga5-1, and gai-1 mutants, which all carry the same er mutation, displayed a strong DNRR activity than Ler-0. These findings are in line with the assumption that dampening of GA signaling, either achieved genetically or by PBZ treatment, can increase DNRR activity in Arabidopsis leaf explants.
2.4. PBZ Treatment Can Rescue Root Regeneration in er Mutants
Since we found almost all rooting activity abolished in Ler-0 leaf explants, we hypothesized that ER signaling is a positive regulator of DNRR. To confirm this hypothesis, we employed a second er allele in our DNRR assays. ER encodes a member of the receptor-like protein kinases (RLK) gene family containing an extracellular receptor domain with 20 leucine-rich repeats (LRR), a transmembrane domain, and a cytoplasmic serine/threonine protein kinase domain [84]. The importance of the protein kinase domain for ER function is illustrated by the Ler-0 allele that carries a missense mutation of isoleucine 750 to lysine, which is a conserved residue in most RLKs [85]. This allele is equivalent in phenotypic strength to er-105 that has no mRNA signal on a Northern blots [84]. Thus, both er alleles, er-1 (Ler-0) and er-105 (Col-0) are likely to be null alleles [84,85]. In DNRR time courses, we found no significant differences between Ler-0 and er-105, while both er alleles displayed significantly reduced rooting rates in comparison to Col-0 (Figure 3B).
Since mutants with loss of GA synthesis or GA signaling can overcome the depletion of AR formation in Ler-0 leaf explants (Figure 2A), we assumed that PBZ treatment can rescue the declined rooting rate in both er alleles. To test this assumption, we performed DNRR assays with both er alleles and Col-0 (Figure 3C). PBZ treatment increased the rooting rate in all three lines. Although both er mutants did not reached the same rooting rate like the Col-0 wild-type after PBZ treatment, both er alleles displayed a much higher ratio of PBZ- to mock-treated leaf explants than wild-type. We conclude that this significantly higher difference in rooting between PBZ- to mock-treatment in er mutants could be a big advantage in whole-genome transcription profiling during DNRR.
2.5. Transcriptomic Analysis, Using er Leaf Explants, Revealed That PBZ Treatment Suppresses GA and Brassinosteroids Responses, While Inducing the Expression of the Rooting Factor LBD16
To identify genes that regulate DNRR and are differentially expressed by PBZ treatment, we performed whole-genome transcription profiling. Under LD condition, average rooting rates of wild-type (Col-0) leaf explants strongly variate in independent experiments ranging from 30 to 100 percent [25,49], while under dark condition, the rooting-promoting effects of PBZ are occasionally covered, and mock- and PBZ-treated Col-0 leaf explants can reach similar high rooting rates (Figure 1G and 2B). In contrast, the rooting rate ratios of PBZ- to mock-treated leaves were significantly higher and less variable in er mutants compared with Col-0 (Figure 3C). We therefore employed the er-105 mutant as genetic tool in our whole-genome expression analysis by RNA Seq and cultured er-105 leaf explants in darkness with and without PBZ for 24 hours (24 HAC; hours after cultivation started). Comparing PBZ- to mock-treated er-105 mutants, genes with a ≥1.5-fold expression change and a p-value ≤0.05 were considered to have significantly different expression levels. Differentially expressed genes (DEGs) are listed in the Supplementary Tables S1-S2. A total of 772 genes were regulated by PBZ (336 up- and 436 down-regulated) in leaf explants, 24 HAC (Figure 4A). This corresponds to 2.8% of the 27,655 protein-coding loci in the Arabidopsis genome [86].
In order to verify decreased GA response by PBZ, we compared our RNA Seq data with GA-regulated genes identified in previous studies [67,68,69]. As expected, we found a significant overlap of PBZ-repressed genes with GA-induced genes (p < 3.9e-09) and PBZ-induced genes with GA-repressed genes (p < 1.2e-06) indicating a reduced GA response in PBZ-treated er-105 leaf explants (Supplementary Figure S2A). These findings are in line with the assumption that PBZ disrupted the GA biosynthesis, which ultimately results in reduced GA signaling and GA response. This conclusion was supported by increased expression of the GA synthesis genes GA3ox1, GA20ox1/GA5, GA20ox2, and GA20ox3 (Figure 4E and Supplementary Figure S3A-B), which are negative feedback regulated by GA [87,88].
The hormone signaling pathways of GA and brassinosteroids (BR) are directly linked through the DELLA-PIF4- BZR1 module [89]. We therefore compared the PBZ-regulated genes with BR-regulated genes identified in previous studies [67,90]. As expected, we found evidence for reduced BR response with significant overlaps of PBZ-repressed genes with BR-induced genes (41 genes, p < 4.7e-08) and PBZ-induced genes with BR-repressed genes (22 genes, p < 1.6e-03; Supplementary Figure S2B). Notably, five genes, DWF4/CYP90B1, CPD/CYP90A1, CYP90D1, ROT3/CYP90C1, and BR6OX2/CYP85A2, encoding enzymes of the BR biosynthesis pathway [91], were up-regulated by PBZ treatment (Figure 4E and Supplementary Figure S3A-B). All these BR biosynthesis genes are direct targets of BZR1 [90]. More than 60% of the PBZ-repressed genes and more than 40% of the PBZ-induced genes are known to be also regulated by GA and/or BR (Figure 4B). This suggests that about half of all transcriptional changes by PBZ were caused by directly interfering with GA and/or BR signaling.
To narrow down which of the PBZ-regulated genes are likely involved in AR formation, we compared them with wounding-regulated genes in leaf explants during DNRR [25]. We found a large overlap of genes that were regulated by PBZ and wounding (Supplementary Figure S2C). The overlap of genes up-regulated by PBZ and wounding was most significant (71 genes p < 1.1e-19), followed by the overlap of genes downregulated by PBZ and wounding (44 genes p < 3.0e-09), and the overlap of genes up-regulated by wounding but down-regulated by PBZ (47 genes p < 4.1e-04). A total of 174 genes were regulated by both, PBZ and wounding, which is 25% of the 694 PBZ-regulated genes. This finding suggests that a large number of the PBZ-regulated genes are specifically involved in DNRR.
Since auxin signaling is essential for AR formation during DNRR [19,20,92], we compared our RNA Seq data with IAA-regulated genes identified in previous studies [67,70]. Although we found large overlaps between PBZ- and auxin-regulated genes, their expression changes correlated positively or negatively (Supplementary Figure S2D). However, genes regulated by PBZ- and auxin largely overlapped with genes regulated by PBZ- and GA/BR (Figure 4B). Next, we identified PBZ-regulated genes known to participate in auxin metabolism, transport, and signal transduction (Figure 4C-D and Supplementary Figure S3B). Among the genes up-regulated by PBZ treatment, we found two IAA-synthesis genes NIT2 [93] and YUC9 [19], the MeIAA hydrolase gene METHYL ESTERASE 16 (MES16) [94], and the UDP-Glucosyltransferase UGT74E2 that indirectly increases IBA content [95], which together can increase the content of biological active auxins. Likely in response to increased auxin levels, we found three GRETCHEN HAGEN 3 (GH3) genes GH3.5/WES1, GH3.15 and GH3.17 up-regulated by PBZ. These GH3 genes encodes IAA/IBA-amido synthetases that can decrease the amount of free auxin [96,97,98]. Although the PBZ treatment resulted in expression changes of auxin metabolism genes, they are likely not related to expression changes in the jasmonate-SPL10/11-ERF109/111-JAZ module [24,25], since ABR1/ERF111, SPL10, SPL11, and especially, its key target, the auxin biosynthesis gene ASA1 were not significant up-regulated (Supplementary Figure S3B) although the expression of ERF109 was increased and JAZ8 expression was abolished by PBZ (Figure 4D and Supplementary Figure S3C). We found the intracellular auxin transporter gene PILS5 [99] and PIN1, encoding an essential auxin efflux carrier [100], induced by PBZ (Figure 4C-E and Supplementary Figure S3A-B)), while negative regulators of polar auxin transport of PIN1 were repressed (Supplementary Figure S3C). Increased expression of MAP KINASE KINASE7 (MKK7) causes via MPK6 sustained phosphorylation of the PIN1 S337 site, which disturbs PIN1 polarity and auxin gradients [101,102]. Thus, reduced MKK7 expression by PBZ treatment should lead to stable auxin transport. Similarly, PATL5, another significant down-regulated gene by PBZ, is also involved in PIN1 location at the cell membrane by repolarization [103]. Taken together, our transcriptional profiling suggest that PBZ at least partially promotes DNRR by increased auxin biosynthesis and transport, and ultimately, auxin signalling and response.
Therefore, we extent our search for potential positive regulators of DNRR by PBZ to genes that are not GA/BR response genes but inducible by wounding and auxin (Figure 4E). Focusing on transcription factors, we identified LBD16 as gene that is inducible by auxin treatment and wounding (during DNRR) [12], while its promoter can be bound by the DELLA-targets PIF4, BZR1, and ARF6 [36]. Remarkably, HOMEOBOX PROTEIN (HB23), encoding a Zinc finger homeobox gene and known transcriptional repressor of LBD16 [104], were down-regulated, which could contribute to the higher LBD16 expression by PBZ treatment (Figure 4C-E).
To confirm that PBZ is a positive regulator of LBD16 expression, and consequently, GA represses LBD16, we employed LBD16::GUS plants [105] in DNRR assays (Figure 5). As expected, PBZ treatment increased LBD16::GUS expression, while additional GA treatment significantly decreased LBD16::GUS expression, and NPA treatment abolished all expression. These results suggest that LBD16, a known positive regulator of DNRR [12,106], is directly or indirectly repressed by GA signalling, which can be overcome by PBZ treatment that functions at least partially through changes in auxin transport and/or auxin signalling.
3. Discussion
In this study, we examined the effects of GA and its biosynthesis inhibitor PBZ on wounding-induced AR formation from Arabidopsis leaf explants (DNRR). Our interest in GA signaling and adventitious rooting was triggered by two contradictory studies in Arabidopsis stating either that GA inhibits etiolation-induced AR formation from hypocotyl [42] or GA is required to promote DNRR by tight regulation of GA signaling through DELLA repressors [49]. We wondered whether GA could play reverse roles in both types of AR formation since jasmonate has such contrary effects [1,24,27]. Since the effects of GA and PBZ treatment on AR formation in Arabidopsis leaf explants were not studied yet, we conducted these DNRR assays and compare the results with the GA and PBZ response of etiolation-induced AR formation in hypocotyl. Under our conditions, GA treatment inhibited AR formation in both types of AR, while PBZ treatment resulted in accelerating effects on adventitious rooting. These findings are in line with the large canon of studies in other plant species [40,41,42,43,44,45,46,47,60,61,62,63]. Accordantly, we found that loss of GA synthesis and signaling in classical GA mutants promotes DNRR. The discrepancy of our results with [49] and the novel role of ER in DNRR might be clarified in future studies. Nevertheless, our study suggests that PBZ treatment has a direct effect on GA and BR signaling during DNRR. Our transcriptional profiling at 24 HAC can only give a glimpse in the molecular mechanisms of PBZ controlled AR formation in leaf explants. Further studies of the early events and genetic validation will be necessary to get a full picture. Nevertheless, our transcription profiling and the literature provided enough information for conceptional models (Figure 4C and Supplementary figure S3A).
3.1. Probable Direct Effects of PBZ Treatment on GA and BR Signaling During DNRR
This study revealed that GA inhibits DNRR, while the GA synthesis inhibitor PBZ promotes AR but not LR formation in Arabidopsis (Figure 1 and Supplementary Figure S1B-C). Our hormone and hormone inhibitor DNRR assays as well as expression data gave a brief glimpse into the putative GA pathways during DNRR. PBZ treatment in darkness results in accumulation of DELLA proteins that block GA signaling [57,107]. The hormone signaling pathways of GA and brassinosteroids (BR) are directly linked through the DELLA-PIF4-BZR1 module [89]. The BR signaling cascade activates the transcription factor BZR1 that directly regulates BR-responsive gene expression and development [108]. BZR1 and PIF4 form heterodimers that directly bind and synergistically activate common target genes, which is inhibited by DELLA proteins. Nevertheless, BZR1 and PIF4 have partially opposite effects on the expression of common target genes. In our conceptional model, the accumulation of DELLAs by PBZ treatment blocks BZR1/PIF4-dependent GA and BR signaling in leaf explants (Figure 4C). Reduced BZR1 function by DELLA could directly result in up-regulation of the auxin biosynthesis genes NIT2 and MES16 and down-regulation of IAA6/SHY, IAA14/SLR and HB23, which can promote LBD16 expression.
Many of the PBZ-regulated genes are bound by the DELLA-targets PIF4, BZR1 and ARF6 (Figure 4E) [36]. This is in line with the hypothesis that GA signaling controls directly DNRR. On the other hand, the huge number of PIF4, BZR1 and/or ARF6 targets among the PBZ-regulated genes makes it impossible to investigate the relevance of all of them for PBZ-promoted DNRR. Many GA and BR biosynthesis genes are feed-back regulated. Accordantly, our expression data confirmed that the PBZ treatment dampened GA and BR responses indicated by the increased expression of the GA biosynthesis genes GA3ox1, GA20ox2, and GA20ox3 (Figure 4E and Supplementary Figure S3A-B), which are negative feedback regulated by GA [87,88]. The BR biosynthesis genes DWF4/CYP90B1, CPD/CYP90A1, CYP90D1, ROT3/CYP90C1, and BR6OX2/CYP85A2, which are direct targets of BZR1 [90,91], were also up-regulated by PBZ treatment (Figure 4E and Supplementary Figure S3A-B). Furthermore, we compared the expression of GAI, RGA1, and INDOLE-3-ACETIC ACID INDUCIBLE 7/AUXIN RESISTANT 2 (IAA7/AXR2), which are well-known marker genes of GA response [68], in er-105 mutant leaf explants with and without PBZ treatment in comparison with Col-0 (Supplementary Figure S3C). As expected, GAI, RGA1, and IAA7/AXR2 were down-regulated by PBZ.
The down-regulation of several bHLH transcription factor genes such as IBL1, PRE1, HBL1, BEE2, and PAR1 can be a direct consequence of reduced BZR1 and PIF function though DELLA accumulation by PBZ, which could be enhanced by the up-regulation of their repressor IBH1 (Supplementary Figure 3). BZR1, PIF4 and ARF6 interact and activate PRE1, which is inhibited by DELLA [36]. The full-name of PRE1, PACLOBUTRAZOL RESISTANCE1, indicated the importance of this gene for PBZ response and it is therefore a good candidate for DNRR studies. Furthermore, the PBZ-repressed transcription factor genes bZIP61, bZIP34, GASA4 and the auxin receptor gene GRH1 are direct targets of BZR1 (Supplementary Figure 3A) and therefore interesting for future DNRR studies. In conclusion, many of the PBZ-regulated transcription factor genes are direct targets of PIF4, BZR1 and/or ARF6 suggesting a direct role of PBZ and, consequently, GA signaling in AR formation during DNRR.
3.2. Probable Direct and Indirect Effects of PBZ Treatment on the Expression of the Rooting Factor LBD16 During DNRR
There are at least four probable pathways of how depletion of GA signaling by PBZ can increase the expression of LBD16 (Figure 4C): (i) The higher expression of auxin biosynthesis such as NIT2 and YUC9 and auxin transport genes such as PIN1 may increase the auxin response that should be also enhanced by (ii) the down-regulation of IAA6/SHY and IAA14/SLR (Figure 4D), but also other Aux/IAAs such as IAA7/AXR2 (Supplementary Figure 3C). (iii) The down-regulation of HB23, encoding a repressor of LBD16 [104], should have a more direct impact of LBD16 expression. Since the promoter of LBD16 is directly bound by PIF4 and BZR1, we could speculate that (iv) they are repressors of LBD16 and their reduced function through DELLA accumulation would be the most direct regulation of LBD16 expression by PBZ. The precise contribution of these pathways on the transcriptional up-regulation of LBD16 might be subject in future GA/DNRR studies.
3.3. Probable GA/BR-Related DNRR Factors Beyond and in Parallel to LBD16
Transcription factor genes, which are transcriptional regulated by wounding signaling during DNRR [25], are naturally good candidates for being regulators of AR formation in leaf explants. Our expression analysis revealed that seven of the wounding-repressed transcription factor genes [25], which are beside IAA14/SLR and HB23, HB21, GRXS13, IBL1, PAR1, and GASA4, were stronger down-regulated by PBZ treatment. On the other hand, the wounding-activated transcription factor genes [25], which are beside LBD16, SPEEDY HYPONASTIC GROWTH (SHG/NAC47), DEHYDRATION RESPONSE ELEMENT-BINDING PROTEIN 19 (DREB19), and OVATE FAMILY PROTEIN 1 (OFP1), and LRP1, were stronger up-regulated by PBZ in our DNRR expression analysis (Figure 4E). The correlation of up-regulation by wounding signaling [25] and even stronger up-regulation by PBZ treatment (this study) is a strong indication that these transcription factors are (i) positive regulators of DNRR and that (ii) they negatively regulated by GA signaling, and vice versa. Notably, only the up-regulation of IDD16 (Figure 4E and Supplementary Figure S3B), encoding a transcription factor that promotes auxin biosynthesis [109], by PBZ broke the pattern of induction by wounding, while repressed by GA, since IDD16 is down-regulated by wounding signaling [25].
Like HB23, HB21 and its paralogue HB31 were down-regulated in our expression data (Supplementary Figure 3B). HB21 controls shoot branching; and together with HB31 control floral architecture through the miR396/GRF module [110]. Notably, the miR396/GROWTH-REGULATING FACTOR (GRF) module is known to form a negative feedback loop with PLETHORA (PLT) genes in the RAM [111]: The GRFs repress PLTs, regulating their spatial expression gradient. On the contrary, PLTs activate MIR396 in the root stem cells to repress the GRFs. Thus, HB21 and HB31 are involved in shoot and root development and could play a role in AR formation via the miR396/GRF module. GRXS13, encoding a member of the CC-type glutaredoxin (ROXY) family, can interact with the transcription factor TGA2 and suppress ORA59 promoter activity that can be induced by jasmonate signaling [112]. However, ORA59 is down-regulated as well (Supplementary Figure 3B), and therefore, the relevance of repression of GRXS13 by PBZ for DNRR remains elusive. GASA4 promotes GA responses [113] and therefore the down-regulation of GASA4 by PBZ should promote DNRR. The bHLH transcription factor genes IBL1 and PAR1 are direct targets of PIF4 and BZR1, and PAR1 is also a target of IBL1, while IBL1 antagonize PIF4 but not the PIF4–BZR1 dimer [89]. Their deep interconnection with the GA signaling pathway and down-regulation during DNRR [25] make IBL1 and PAR1 good candidates for future DNRR studies.
SHG/NAC47 promotes hyponastic growth that involves the volatile phytohormone ethylene [114,115] that might switch the AR-inhibiting effect of GA into an AR-promoting pathway as observed in rice [48], but whether SHG/NAC47 promotes AR formation is unclear. DREB19 is involved in response to drought stress [116] that naturally appear in detached leaves, but also it remains for DREB19 unclear whether it promotes AR formation. OFP1 is a member of the plant specific ovate protein family that can bind to KNOX and BELL-like TALE class homeodomain proteins to relocate these TALE homeodomain from the nucleus to the cytoplasm. Thus, OFP1 functions as a transcriptional repressor, while one of its known target genes is GA20ox1/GA5 [117]. From there, OFP1 might suppresses GA biosynthesis by repression of GA20ox1/GA5 during DNRR to promote AR formation. Nevertheless, LRP1 is beside LBD16 the most probable candidate to affect significantly GA-regulated DNRR, since LRP1 is an early wounding response gene during DNRR [25], responsible to auxin, and is directly bound by BZR1 and ARF6 [36].Furthermore, LRP1 is expressed in LR and AR primordia [118], while overexpression of LRP1, resulting in reduced emerged LR density, which might be related to the reduced rooting capacity by PBZ treatment (Figure 2). Although LRP1 expression is induced by auxin during AR and LR development [118,119], it belongs to neither the fast IAA-response genes in tissue culture [70] nor in seedlings [67]. Differential sensitivity of developmental responses to auxin is well-known. Unlike arf7 arf19 double mutants, LRP1 OE plants showed normal auxin sensitivity. In future studies, it would be interesting to explore how LRP1, OFP1, and the other candidates contribute to AR formation during DNRR.
4. Materials and Methods
Plant Materials and Culture Conditions
Most experiments were performed with the Arabidopsis ecotype Columbia (Col-0). The receptor mutant er-105 [84,85] and LBD16::GUS plants [105], both in Col-0 background were also used. Landsberg erecta (Ler-0) served as wild-type control for the GA mutants gai-1 [66], ga1-4 [65], and ga5-1 [64]. After surface-sterilization with 70% ethanol (twice), the seeds were sowed on half-strength Murashige and Skoog (½ MS) plates with 10 g/L Sucrose, 0.5 g/L MES and 9 g/L Agar, pH=5.8, and stratified at 4°C for 3 days. Then, the plates were transferred to a plant growth cabinet with long-day (LD) condition (16/8h light/dark cycle) at 22℃.
De Novo Root Regeneration (DNRR) and Hormone Treatment
The first pair of rosette leaves (primary leaves) from 14-days-old Arabidopsis plants were used for DNRR assays. Growth conditions of plants and culture conditions of leaf explants followed the protocols described earlier [120]. The leaf explants were cultured on B5 medium (Gamborg B5 medium with 0.5g/L MES and 8g/L Agar, pH=5.7) with 30g/L Sucrose under dark conditions or LD without Sucrose. The rooting rate is represented by the percentage of explants with regenerated roots at a given time point [120], while the regenerative capacity is represented by the percentage of leaf explants with different number of regenerated roots [25]. For each experiment more than six plates of each treatment with more than 10 explants per genotype were used. For hormone and hormone inhibitor treatments, we used 0.1 µM IAA (Sigma), 1 µM NPA (Sigma), 1 µM GA3 (Phytotech) and/or 0.1/1.0 µM Paclobutrazol (PBZ/PAC) (Solarbio/ Phytotech) in the B5 medium. One-tailed Student’s t tests, resulting in p values, were employed to assess statistical significance between value pairs of different treatments [121]. For RNA-seq materials, leaf explants were cultured on B5 medium plates with and without 5 µM PBZ for 24h, harvested in liquid nitrogen, and stored in -80℃.
Adventitious Root Formation from Etiolated Hypocotyls
Col-0 plants were grown in vitro. The AR induction conditions were previously described [71]. After sterilization, the seeds were sown in lines on plates and left for 3 days at 4°C for stratification, exposed for 3 h to light to induce germination, transferred to the dark for 3 days (until hypocotyls were around 6 mm in length) and then exposed to LD conditions in a plant growth cabinet for 7 days before counting the ARs formed on the hypocotyls. To apply the treatment with GA and PBZ, the etiolated seedlings were carefully transferred to new plates of ½ MS medium with or without supplementary 1.0 µM GA3 or 1.0 µM PBZ and then exposed to light for 7 days.
Root Length and Lateral Root (LR) Number Assays
Root growth assays were performed with minor modifications as described before [122]. In brief, primary root length was measured under different concentration of PBZ. Seeds were sowed on ½ MS plates, kept at 4℃ for 3 days and then incubated in a plant growth chamber at 22℃ for 5 days. Then transferred the plants to the ½ MS with 1 µM, 5 µM, 25 µM PBZ for 7 days and measured the length by Image J. The number of LRs was counted by mature LRs greater than 0.5 mm in length. At least 23 plants were used for each treatment.
RNA-Seq and Expression Analysis
Total RNA was isolated and used for the sample preparations. After RNA extraction, mRNA was purified from total RNA by poly-T oligo-attached magnetic beads and cut into small fragments of preferentially 370-420 bp for cDNA synthesis. The libraries were purified by the AMPure XP system (Beverly, USA). The qualified libraries were pooled and sequenced on IIIumina platforms. We used fastqc to evaluate the raw data and trim-galore to filter low-quality bases, N bases, and adapters to obtain clean data for further analysis. In order to obtain gene quantitative information, we used hisat2 software to align the clean data to the reference genome. Then, StringTie software was used for gene transcript quantification analysis. R package DESeq2 was used to perform analysis of differentially expressed genes. Further analyses and comparisons of differentially expressed genes (DEGs, with a ≥ 1.5-fold expression change and a p-value p ≤ 0.05), expression fold changes were calculated with a regular spreadsheet program (Excel, Microsoft) and the DEGs are listed in Supplementary Tables S2-5. In addition, significant different expression was verified by Student’s t-test. Venny 2.1 (https://bioinfogp.cnb.csic.es/tools/venny/index.html) was used for comparisons of gene lists.
Detection of β-glucuronidase (GUS) activity in leaf explants were performed as described, with minor modifications [123,124] (Li et al., 2020). In brief, leaf explants were harvested at specific time points (0 HAC, 24 HAC, and 48 HAC) and immersed in the GUS assay solution (50 mM NaHPO4, 0.5 mM ferrocyanide, 0.5 mM ferricyanide, and 1% Triton X-100, pH 7.2) containing 1 mM X-Gluc. The leaves in the GUS solution were vacuum infiltrated for 30 min, and then incubated at 37 °C overnight. To remove the chlorophyll, stained leaves were carried through ethanol series and then photographed with a stereomicroscope. The digital photographs a were collated with Adobe Photoshop and adjusted as described before [125].
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
T.J. performed all experiments; R.M.-X. created the concept and designed the research; T.J. and R.M.-X. analyzed the rooting data; Y.S. performed the basic RNA-Seq analysis that was further analyzed and graphed by T.J. and R.M.-X.; X. L., Q.X., and R.M.-X. co-supervised T.J.; R.M.-X. wrote the manuscript with the help of T.J. and Q.X..
Funding
This research was funded by the National Natural Science Foundation of China (project Nos. 31640054 and 31771602), two Starting Grants of the Lushan Botanical Garden, Chinese Academy of Sciences, Jiujiang (Grant Nos. 2021ZWZX24 and 2021ZWZX25) to Q.X and R.M.-X., and the High-Level Foreign Experts Project (G2021022004) from the Chinese Ministry of Science and Technology to R.M.-X.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All relevant data are included within this article.
Acknowledgments
We thank all members of our research groups for their support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pan, X.; Yang, Z.; Xu, L. Dual roles of jasmonate in adventitious rooting. EXBOTJ 2021, 72, 6808–6810. [Google Scholar] [CrossRef] [PubMed]
- Hochholdinger, F.; Park, W.J.; Sauer, M.; Woll, K. From weeds to crops: Genetic analysis of root development in cereals. Trends Plant Sci. 2004, 9, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Taramino, G.; Sauer, M.; Stauffer, J.L.; Multani, D.; Niu, X.; Sakai, H.; Hochholdinger, F. The maize (Zea mays L.) RTCS gene encodes a LOB domain protein that is a key regulator of embryonic seminal and post-embryonic shoot-borne root initiation. Plant J 2007, 50, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Tai, H.; Saleem, M.; Ludwig, Y.; Majer, C.; Berendzen, K.W.; Nagel, K.A.; Wojciechowski, T.; Meeley, R.B.; Taramino, G.; et al. Cooperative action of the paralogous maize lateral organ boundaries (LOB) domain proteins RTCS and RTCL in shoot-borne root formation. New Phytol 2015, 207, 1123–1133. [Google Scholar] [CrossRef]
- Liu, H.; Wang, S.; Yu, X.; Yu, J.; He, X.; Zhang, S.; Shou, H.; Wu, P. ARL1, a LOB-domain protein required for adventitious root formation in rice. Plant J. 2005, 43, 47–56. [Google Scholar] [CrossRef]
- Inukai, Y.; Sakamoto, T.; Ueguchi-Tanaka, M.; Shibata, Y.; Gomi, K.; Umemura, I.; Hasegawa, Y.; Ashikari, M.; Kitano, H.; Matsuoka, M. Crown rootless1, Which Is Essential for Crown Root Formation in Rice, Is a Target of an AUXIN RESPONSE FACTOR in Auxin Signaling. Plant Cell 2005, 17, 1387–1396. [Google Scholar] [CrossRef]
- Gill, R.A.; Li, X.; Duan, S.; Xing, Q.; Müller-Xing, R. Citrus threat huanglongbing (HLB) - Could the rootstock provide the cure? Front. Plant Sci. 2024, 15, 355. [Google Scholar] [CrossRef]
- Druege, U.; Hilo, A.; Pérez-Pérez, J.M.; Klopotek, Y.; Acosta, M.; Shahinnia, F.; Zerche, S.; Franken, P.; Hajirezaei, M.R. Molecular and physiological control of adventitious rooting in cuttings: Phytohormone action meets resource allocation. Ann. Bot. 2019, 123, 929–949. [Google Scholar] [CrossRef] [PubMed]
- Fan, M.; Xu, C.; Xu, K.; Hu, Y. LATERAL ORGAN BOUNDARIES DOMAIN transcription factors direct callus formation in Arabidopsis regeneration. Cell Res. 2012, 22, 1169–1180. [Google Scholar] [CrossRef]
- Okushima, Y.; Fukaki, H.; Onoda, M.; Theologis, A.; Tasaka, M. ARF7 and ARF19 regulate lateral root formation via direct activation of LBD/ASL genes in Arabidopsis. Plant Cell 2007, 19, 118–130. [Google Scholar] [CrossRef]
- Lee, H.W.; Cho, C.; Pandey, S.K.; Park, Y.; Kim, M.-J.; Kim, J. LBD16 and LBD18 acting downstream of ARF7 and ARF19 are involved in adventitious root formation in Arabidopsis. BMC Plant Biol 2019, 19, 639. [Google Scholar] [CrossRef] [PubMed]
- Sheng, L.; Hu, X.; Du, Y.; Zhang, G.; Huang, H.; Scheres, B.; Xu, L. Non-canonical WOX11-mediated root branching contributes to plasticity in Arabidopsis root system architecture. Development 2017, 144, 3126–3133. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Ge, Y.; Cai, G.; Pan, X.; Xu, L. WOX-ARF modules initiate different types of roots. Cell Rep. 2023, 42, 112966. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Hu, X.; Qin, P.; Prasad, K.; Hu, Y.; Xu, L. The WOX11-LBD16 Pathway Promotes Pluripotency Acquisition in Callus Cells During De Novo Shoot Regeneration in Tissue Culture. Plant Cell Physiol. 2018, 59, 734–743. [Google Scholar] [CrossRef]
- Müller-Xing, R.; Xing, Q. In da club: The cytoplasmic kinase MAZZA joins CLAVATA signaling and dances with CLV1-like receptors. J. Exp. Bot. 2021, 72, 4596–4599. [Google Scholar] [CrossRef]
- Ge, Y.; Fang, X.; Liu, W.; Sheng, L.; Xu, L. Adventitious lateral rooting: The plasticity of root system architecture. Physiol Plant 2019, 165, 39–43. [Google Scholar] [CrossRef]
- Müller-Xing, R.; Xing, Q. The plant stem-cell niche and pluripotency: 15 years of an epigenetic perspective. Front. Plant Sci. 2022, 13, 1047. [Google Scholar] [CrossRef]
- Jing, T.; Ardiansyah, R.; Xu, Q.; Xing, Q.; Müller-Xing, R. Reprogramming of Cell Fate During Root Regeneration by Transcriptional and Epigenetic Networks. Front. Plant Sci. 2020, 11, 317. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Tong, J.; Xiao, L.; Ruan, Y.; Liu, J.; Zeng, M.; Huang, H.; Wang, J.-W.; Xu, L. YUCCA-mediated auxin biogenesis is required for cell fate transition occurring during de novo root organogenesis in Arabidopsis. J. Exp. Bot. 2016, 67, 4273–4284. [Google Scholar] [CrossRef]
- Liu, J.; Sheng, L.; Xu, Y.; Li, J.; Yang, Z.; Huang, H.; Xu, L. WOX11 and 12 are involved in the first-step cell fate transition during de novo root organogenesis in Arabidopsis. Plant Cell 2014, 26, 1081–1093. [Google Scholar] [CrossRef]
- Zhao, Y. Auxin Biosynthesis: A Simple Two-Step Pathway Converts Tryptophan to Indole-3-Acetic Acid in Plants. Molecular Plant 2012, 5, 334–338. [Google Scholar] [CrossRef]
- Mashiguchi, K.; Tanaka, K.; Sakai, T.; Sugawara, S.; Kawaide, H.; Natsume, M.; Hanada, A.; Yaeno, T.; Shirasu, K.; Yao, H.; et al. The main auxin biosynthesis pathway in Arabidopsis. Proc. Natl. Acad. Sci. U. S. A. 2011, 108, 18512–18517. [Google Scholar] [CrossRef] [PubMed]
- Won, C.; Shen, X.; Mashiguchi, K.; Zheng, Z.; Dai, X.; Cheng, Y.; Kasahara, H.; Kamiya, Y.; Chory, J.; Zhao, Y. Conversion of tryptophan to indole-3-acetic acid by TRYPTOPHAN AMINOTRANSFERASES OF ARABIDOPSIS and YUCCAs in Arabidopsis. Proc. Natl. Acad. Sci. U. S. A. 2011, 108, 18518–18523. [Google Scholar] [CrossRef]
- Zhang, G.; Zhao, F.; Chen, L.; Pan, Y.; Sun, L.; Bao, N.; Zhang, T.; Cui, C.-X.; Qiu, Z.; Zhang, Y.; et al. Jasmonate-mediated wound signalling promotes plant regeneration. Nat. Plants 2019. [CrossRef] [PubMed]
- Ye, B.-B.; Shang, G.-D.; Pan, Y.; Xu, Z.-G.; Zhou, C.-M.; Mao, Y.-B.; Bao, N.; Sun, L.; Xu, T.; Wang, J.-W. AP2/ERF Transcription Factors Integrate Age and Wound Signals for Root Regeneration. Plant Cell 2020, 32, 226–241. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Xing, Q.; Jing, T.; Fan, X.; Zhang, Q.; Müller-Xing, R. The epigenetic regulator ULTRAPETALA1 suppresses de novo root regeneration from Arabidopsis leaf explants. Plant Signal. Behav. 2022, 11, 118. [Google Scholar] [CrossRef]
- Dob, A.; Lakehal, A.; Novak, O.; Bellini, C.; Murphy, A. Jasmonate inhibits adventitious root initiation through repression of CKX1 and activation of RAP2.6L transcription factor in Arabidopsis. EXBOTJ 2021, 72, 7107–7118. [Google Scholar] [CrossRef]
- Binenbaum, J.; Weinstain, R.; Shani, E. Gibberellin Localization and Transport in Plants. Trends Plant Sci. 2018, 23, 410–421. [Google Scholar] [CrossRef]
- Lange, T. Molecular biology of gibberellin synthesis. Planta 1998, 204, 409–419. [Google Scholar] [CrossRef]
- Shani, E.; Hedden, P.; Sun, T.-p. Highlights in gibberellin research: A tale of the dwarf and the slender. Plant Physiol. 2024, 195, 111–134. [Google Scholar] [CrossRef]
- Lucas, M. de; Davière, J.-M.; Rodríguez-Falcón, M.; Pontin, M.; Iglesias-Pedraz, J.M.; Lorrain, S.; Fankhauser, C.; Blázquez, M.A.; Titarenko, E.; Prat, S. A molecular framework for light and gibberellin control of cell elongation. Nature 2008, 451, 480–484. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Martinez, C.; Gusmaroli, G.; Wang, Y.; Zhou, J.; Wang, F.; Chen, L.; Yu, L.; Iglesias-Pedraz, J.M.; Kircher, S.; et al. Coordinated regulation of Arabidopsis thaliana development by light and gibberellins. Nature 2008, 451, 475–479. [Google Scholar] [CrossRef] [PubMed]
- Bai, M.-Y.; Shang, J.-X.; Oh, E.; Fan, M.; Bai, Y.; Zentella, R.; Sun, T.-p.; Wang, Z.-Y. Brassinosteroid, gibberellin and phytochrome impinge on a common transcription module in Arabidopsis. Nat Cell Biol 2012, 14, 810–817. [Google Scholar] [CrossRef] [PubMed]
- Gallego-Bartolomé, J.; Minguet, E.G.; Grau-Enguix, F.; Abbas, M.; Locascio, A.; Thomas, S.G.; Alabadí, D.; Blázquez, M.A. Molecular mechanism for the interaction between gibberellin and brassinosteroid signaling pathways in Arabidopsis. Proceedings of the National Academy of Sciences 2012, 109, 13446–13451. [Google Scholar] [CrossRef]
- Wild, M.; Davière, J.-M.; Cheminant, S.; Regnault, T.; Baumberger, N.; Heintz, D.; Baltz, R.; Genschik, P.; Achard, P. The Arabidopsis DELLA RGA - LIKE3 Is a Direct Target of MYC2 and Modulates Jasmonate Signaling Responses. Plant Cell 2012, 24, 3307–3319. [Google Scholar] [CrossRef]
- Oh, E.; Zhu, J.-Y.; Bai, M.-Y.; Arenhart, R.A.; Sun, Y.; Wang, Z.-Y. Cell elongation is regulated through a central circuit of interacting transcription factors in the Arabidopsis hypocotyl. Elife 2014, 3, 601. [Google Scholar] [CrossRef]
- Yu, S.; Galvão, V.C.; Zhang, Y.-C.; Horrer, D.; Zhang, T.-Q.; Hao, Y.-H.; Feng, Y.-Q.; Wang, S.; Schmid, M.; Wang, J.-W. Gibberellin Regulates the Arabidopsis Floral Transition through miR156-Targeted SQUAMOSA PROMOTER BINDING–LIKE Transcription Factors. Plant Cell 2012, 24, 3320–3332. [Google Scholar] [CrossRef]
- Hyun, Y.; Richter, R.; Vincent, C.; Martinez-Gallegos, R.; Porri, A.; Coupland, G. Multi-layered Regulation of SPL15 and Cooperation with SOC1 Integrate Endogenous Flowering Pathways at the Arabidopsis Shoot Meristem. Dev. Cell 2016, 37, 254–266. [Google Scholar] [CrossRef]
- Huang, X.; Tian, H.; Park, J.; Oh, D.-H.; Hu, J.; Zentella, R.; Qiao, H.; Dassanayake, M.; Sun, T.-p. The master growth regulator DELLA binding to histone H2A is essential for DELLA-mediated global transcription regulation. Nat. Plants 2023, 9, 1291–1305. [Google Scholar] [CrossRef]
- Urban, I.; Libbert, E. Adventivwurzelbildung bei der Windepflanze Calystegia sepium (L.) R. Br: Stimulation durch Auxine, Vitamine und den Gihherellinantagonisten 2-(Chloräthyl)trimethylammoniumchlorid 1 1)Zweiter Teil einer Dissertation der Mathematisch-Naturwissenschaftlichen Fakultät der Universität Rostock (URBAN 1965). Der dritte Teil wird anschließend publiziert. ). Flora oder Allgemeine botanische Zeitung. Abt. A, Physiologie und Biochemie 1967, 157, 373–388. [Google Scholar] [CrossRef]
- BRIAN, P.W.; HEMMING, H.G.; LOWE, D. Inhibition of Rooting of Cuttings by Gibberellic Acid: With one Figure in the Text. Ann. Bot. 1960, 24, 407–419. [Google Scholar] [CrossRef]
- Mauriat, M.; Petterle, A.; Bellini, C.; Moritz, T. Gibberellins inhibit adventitious rooting in hybrid aspen and Arabidopsis by affecting auxin transport. Plant J. 2014, 78, 372–384. [Google Scholar] [CrossRef] [PubMed]
- Busov, V.; Meilan, R.; Pearce, D.W.; Rood, S.B.; Ma, C.; Tschaplinski, T.J.; Strauss, S.H. Transgenic modification of gai or rgl1 causes dwarfing and alters gibberellins, root growth, and metabolite profiles in Populus. Planta 2006, 224, 288–299. [Google Scholar] [CrossRef]
- Jansen, H. Die Wirkung von Gibberellinsure und Indolylessigsure auf die Wurzelbildung von Tomatenstecklingen. Planta 1967, 74, 371–378. [Google Scholar] [CrossRef]
- Zhang, J.; Zhou, T.; Zhang, C.; Zheng, W.; Li, J.; Jiang, W.; Xiao, C.; Wei, D.; Yang, C.; Xu, R.; et al. Gibberellin disturbs the balance of endogenesis hormones and inhibits adventitious root development of Pseudostellaria heterophylla through regulating gene expression related to hormone synthesis. Saudi Journal of Biological Sciences 2021, 28, 135–147. [Google Scholar] [CrossRef] [PubMed]
- BRIAN, P.W.; HEMMING, H.G.; Radley, M. A Physiological Comparison of Gibberellic Acid with Some Auxins. Physiol Plant 1955, 8, 899–912. [Google Scholar] [CrossRef]
- Kato, J. Studies on the Physiological Effect of Gibberellin II. Physiol Plant 1958, 11, 10–15. [Google Scholar] [CrossRef]
- Steffens, B.; Wang, J.; Sauter, M. Interactions between ethylene, gibberellin and abscisic acid regulate emergence and growth rate of adventitious roots in deepwater rice. Planta 2006, 223, 604–612. [Google Scholar] [CrossRef]
- Ibáñez, S.; Ruiz-Cano, H.; Fernández, M.Á.; Sánchez-García, A.B.; Villanova, J.; Micol, J.L.; Pérez-Pérez, J.M. A Network-Guided Genetic Approach to Identify Novel Regulators of Adventitious Root Formation in Arabidopsis thaliana. Front. Plant Sci. 2019, 10, 461. [Google Scholar] [CrossRef]
- Hedden, P.; Graebe, J.E. Inhibition of gibberellin biosynthesis by paclobutrazol in cell-free homogenates ofCucurbita maxima endosperm andMalus pumila embryos. J Plant Growth Regul 1985, 4, 111–122. [Google Scholar] [CrossRef]
- Helliwell, C.A.; Sheldon, C.C.; Olive, M.R.; Walker, A.R.; Zeevaart, J.A.D.; Peacock, W.J.; Dennis, E.S. Cloning of the Arabidopsisent -kaurene oxidase gene GA 3. Proceedings of the National Academy of Sciences 1998, 95, 9019–9024. [Google Scholar] [CrossRef] [PubMed]
- Helliwell, C.A.; Poole, A.; James Peacock, W.; Dennis, E.S. Arabidopsis ent -Kaurene Oxidase Catalyzes Three Steps of Gibberellin Biosynthesis. Plant Physiol. 1999, 119, 507–510. [Google Scholar] [CrossRef] [PubMed]
- Alabadí, D.; Gil, J.; Blázquez, M.A.; García-Martínez, J.L. Gibberellins Repress Photomorphogenesis in Darkness. Plant Physiol. 2004, 134, 1050–1057. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.-L.; Ogawa, M.; Fleet, C.M.; Zentella, R.; Hu, J.; Heo, J.-o.; Lim, J.; Kamiya, Y.; Yamaguchi, S.; Sun, T.-p. SCARECROW-LIKE 3 promotes gibberellin signaling by antagonizing master growth repressor DELLA in Arabidopsis. Proceedings of the National Academy of Sciences 2011, 108, 2160–2165. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Schwarz, S.; Saedler, H.; Huijser, P. SPL8, a local regulator in a subset of gibberellin-mediated developmental processes in Arabidopsis. Plant Mol Biol 2007, 63, 429–439. [Google Scholar] [CrossRef]
- An, F.; Zhang, X.; Zhu, Z.; Ji, Y.; He, W.; Jiang, Z.; Li, M.; Guo, H. Coordinated regulation of apical hook development by gibberellins and ethylene in etiolated Arabidopsis seedlings. Cell Res 2012, 22, 915–927. [Google Scholar] [CrossRef]
- Li, K.; Yu, R.; Fan, L.-M.; Wei, N.; Chen, H.; Deng, X.W. DELLA-mediated PIF degradation contributes to coordination of light and gibberellin signalling in Arabidopsis. Nat Commun 2016, 7, 215. [Google Scholar] [CrossRef]
- Cao, H.; Han, Y.; Li, J.; Ding, M.; Li, Y.; Li, X.; Chen, F.; Soppe, W.J.; Liu, Y.; Leubner, G. Arabidopsis thaliana SEED DORMANCY 4-LIKE regulates dormancy and germination by mediating the gibberellin pathway. EXBOTJ 2020, 71, 919–933. [Google Scholar] [CrossRef]
- Wang, Z.; Cao, H.; Zhang, C.; Chen, F.; Liu, Y. The SNF5-type protein BUSHY regulates seed germination via the gibberellin pathway and is dependent on HUB1 in Arabidopsis. Planta 2022, 255, 653. [Google Scholar] [CrossRef]
- Henrique, A.; Campinhos, E.N.; Ono, E.O.; Pinho, S.Z.d. Effect of plant growth regulators in the rooting of Pinus cuttings. Braz. arch. biol. technol. 2006, 49, 189–196. [Google Scholar] [CrossRef]
- Marino, G. The effect of paclobutrazol on in vitro rooting, transplant establishment and growth of fruit plants. Plant Growth Regulation 1988, 7, 237–247. [Google Scholar] [CrossRef]
- Križan, B.; Ondrušiková, E.; Dradi, G.; Rocasaglia, R. The effect of paclobutrazol on in vitro rooting and growth of GF-677 hybrid peach rootstock. Acta Physiol Plant 2006, 28, 21–26. [Google Scholar] [CrossRef]
- Wang, Y.; Yao, R. Optimization of rhizogenesis for in vitro shoot culture of Pinus massoniana Lamb. J. For. Res. 2021, 32, 203–209. [Google Scholar] [CrossRef]
- Xu, Y.L.; Li, L.; Wu, K.; Peeters, A.J.; Gage, D.A.; Zeevaart, J.A. The GA5 locus of Arabidopsis thaliana encodes a multifunctional gibberellin 20-oxidase: Molecular cloning and functional expression. Proceedings of the National Academy of Sciences 1995, 92, 6640–6644. [Google Scholar] [CrossRef]
- Sun, T.; Goodman, H.M.; Ausubel, F.M. Cloning the Arabidopsis GA1 Locus by Genomic Subtraction. Plant Cell 1992, 119–128. [Google Scholar] [CrossRef]
- Alabadí, D.; Gallego-Bartolomé, J.; Orlando, L.; García-Cárcel, L.; Rubio, V.; Martínez, C.; Frigerio, M.; Iglesias-Pedraz, J.M.; Espinosa, A.; Deng, X.W.; et al. Gibberellins modulate light signaling pathways to prevent Arabidopsis seedling de-etiolation in darkness. Plant J 2008, 53, 324–335. [Google Scholar] [CrossRef]
- Goda, H.; Sasaki, E.; Akiyama, K.; Maruyama-Nakashita, A.; Nakabayashi, K.; Li, W.; Ogawa, M.; Yamauchi, Y.; Preston, J.; Aoki, K.; et al. The AtGenExpress hormone and chemical treatment data set: Experimental design, data evaluation, model data analysis and data access. Plant J. 2008, 55. [Google Scholar] [CrossRef]
- Ogawa, M.; Hanada, A.; Yamauchi, Y.; Kuwahara, A.; Kamiya, Y.; Yamaguchi, S. Gibberellin Biosynthesis and Response during Arabidopsis Seed Germination[W]. Plant Cell 2003, 15, 1591–1604. [Google Scholar] [CrossRef]
- Ribeiro, D.M.; Araújo, W.L.; Fernie, A.R.; Schippers, J.H.M.; Mueller-Roeber, B. Translatome and metabolome effects triggered by gibberellins during rosette growth in Arabidopsis. EXBOTJ 2012, 63, 2769–2786. [Google Scholar] [CrossRef]
- Vain, T.; Raggi, S.; Ferro, N.; Barange, D.K.; Kieffer, M.; Ma, Q.; Doyle, S.M.; Thelander, M.; Pařízková, B.; Novák, O.; et al. Selective auxin agonists induce specific AUX/IAA protein degradation to modulate plant development. Proceedings of the National Academy of Sciences 2019, 116, 6463–6472. [Google Scholar] [CrossRef]
- Sorin, C.; Bussell, J.D.; Camus, I.; Ljung, K.; Kowalczyk, M.; Geiss, G.; McKhann, H.; Garcion, C.; Vaucheret, H.; Sandberg, G.; et al. Auxin and Light Control of Adventitious Rooting in Arabidopsis Require ARGONAUTE1. Plant Cell 2005, 17, 1343–1359. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Chai, M.; Li, Y.; Sun, M.; Zhang, J.; Sun, G.; Jiang, C.; Shi, L. Global Transcriptome Profiling Analysis of Inhibitory Effects of Paclobutrazol on Leaf Growth in Lily (Lilium Longiflorum-Asiatic Hybrid). Front. Plant Sci. 2016, 7, 1188. [Google Scholar] [CrossRef] [PubMed]
- Divya Nair, V.; Jaleel, C.A.; Gopi, R.; Panneerselvam, R. Changes in growth and photosynthetic characteristics of Ocimum sanctum under growth regulator treatments. Front. Biol. China 2009, 4, 192–199. [Google Scholar] [CrossRef]
- Karidas, P.; Challa, K.R.; Nath, U. The tarani mutation alters surface curvature in Arabidopsis leaves by perturbing the patterns of surface expansion and cell division. EXBOTJ 2015, 66, 2107–2122. [Google Scholar] [CrossRef]
- Cohen, Y.; Aloni, D.D.; Adur, U.; Hazon, H.; Klein, J.D. Characterization of Growth-Retardant Effects on Vegetative Growth of Date Palm Seedlings. J Plant Growth Regul 2013, 32, 533–541. [Google Scholar] [CrossRef]
- Sarkar, J.; K. , A.; Kumar, R.K.; Jacob, J. MORPHOLOGICAL CHANGES IN YOUNG PLANTS OF HEVEA BRASILIENSIS INDUCED BY PACLOBUTRAZOL. Rubber Science 2015, 28, 22–30. [Google Scholar]
- Burrows, G.E.; Boag, T.S.; Stewart, W.P. Changes in leaf, stem, and root anatomy of Chrysanthemum cv. Lillian Hoek following paclobutrazol application. J Plant Growth Regul 1992, 11, 189–194. [Google Scholar] [CrossRef]
- Teixeira, E.C.; Matsumoto, S.N.; Ribeiro, A.F.F.; Viana, A.E.S.; Tagliaferre, C.; Carvalho, F.D.; Pereira, L.F.; Silva, V.A. Morphophysiology and quality of yellow passion fruit seedlings submitted to inhibition of gibberellin biosynthesis. Acta Sci. Agron. 2020, 43, e51541. [Google Scholar] [CrossRef]
- Tsegaw, T.; Hammes, S.; Robbertse, J. Paclobutrazol-induced Leaf, Stem, and Root Anatomical Modifications in Potato. HortScience 2005, 40, 1343–1346. [Google Scholar] [CrossRef]
- Weaver, L.M.; Amasino, R.M. Senescence Is Induced in Individually Darkened Arabidopsis Leaves, but Inhibited in Whole Darkened Plants. Plant Physiol. 2001, 127, 876–886. [Google Scholar] [CrossRef]
- Song, Y.; Yang, C.; Gao, S.; Zhang, W.; Li, L.; Kuai, B. Age-Triggered and Dark-Induced Leaf Senescence Require the bHLH Transcription Factors PIF3, 4, and 5. Molecular Plant 2014, 7, 1776–1787. [Google Scholar] [CrossRef] [PubMed]
- Leivar, P.; Monte, E. PIFs: Systems Integrators in Plant Development. Plant Cell 2014, 26, 56–78. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Carol, P.; Richards, D.E.; King, K.E.; Cowling, R.J.; Murphy, G.P.; Harberd, N.P. The Arabidopsis GAI gene defines a signaling pathway that negatively regulates gibberellin responses. Genes Dev. 1997, 11, 3194–3205. [Google Scholar] [CrossRef] [PubMed]
- Torii, K.U.; Mitsukawa, N.; Oosumi, T.; Matsuura, Y.; Yokoyama, R.; Whittier, R.F.; Komeda, Y. The Arabidopsis ERECTA gene encodes a putative receptor protein kinase with extracellular leucine-rich repeats. Plant Cell 1996, 8, 735–746. [Google Scholar] [CrossRef]
- Lease, K.A.; Lau, N.Y.; Schuster, R.A.; Torii, K.U.; Walker, J.C. Receptor serine/threonine protein kinases in signalling: Analysis of the erecta receptor-like kinase of Arabidopsis thaliana. New Phytol 2001, 151, 133–143. [Google Scholar] [CrossRef]
- Cheng, C.-Y.; Krishnakumar, V.; Chan, A.P.; Thibaud-Nissen, F.; Schobel, S.; Town, C.D. Araport11: A complete reannotation of the Arabidopsis thaliana reference genome. Plant J 2017, 89, 789–804. [Google Scholar] [CrossRef] [PubMed]
- Rieu, I.; Ruiz-Rivero, O.; Fernandez-Garcia, N.; Griffiths, J.; Powers, S.J.; Gong, F.; Linhartova, T.; Eriksson, S.; Nilsson, O.; Thomas, S.G.; et al. The gibberellin biosynthetic genes AtGA20ox1 and AtGA20ox2 act, partially redundantly, to promote growth and development throughout the Arabidopsis life cycle. Plant J 2008, 53, 488–504. [Google Scholar] [CrossRef]
- Matsushita, A.; Furumoto, T.; Ishida, S.; Takahashi, Y. AGF1, an AT-Hook Protein, Is Necessary for the Negative Feedback of AtGA3ox1 Encoding GA 3-Oxidase. Plant Physiol. 2007, 143, 1152–1162. [Google Scholar] [CrossRef]
- Zhiponova, M.K.; Morohashi, K.; Vanhoutte, I.; Machemer-Noonan, K.; Revalska, M.; van Montagu, M.; Grotewold, E.; Russinova, E. Helix–loop–helix/basic helix–loop–helix transcription factor network represses cell elongation in Arabidopsis through an apparent incoherent feed-forward loop. Proceedings of the National Academy of Sciences 2014, 111, 2824–2829. [Google Scholar] [CrossRef]
- Sun, Y.; Fan, X.-Y.; Cao, D.-M.; Tang, W.; He, K.; Zhu, J.-Y.; He, J.-X.; Bai, M.-Y.; Zhu, S.; Oh, E.; et al. Integration of Brassinosteroid Signal Transduction with the Transcription Network for Plant Growth Regulation in Arabidopsis. Dev. Cell 2010, 19, 765–777. [Google Scholar] [CrossRef]
- Chung, Y.; Choe, S. The Regulation of Brassinosteroid Biosynthesis in Arabidopsis. Critical Reviews in Plant Sciences 2013, 32, 396–410. [Google Scholar] [CrossRef]
- Zeng, Y.; Verstraeten, I.; Trinh, H.K.; Lardon, R.; Schotte, S.; Olatunji, D.; Heugebaert, T.; Stevens, C.; Quareshy, M.; Napier, R.; et al. Chemical induction of hypocotyl rooting reveals extensive conservation of auxin signalling controlling lateral and adventitious root formation. New Phytologist 2023, 240, 1883–1899. [Google Scholar] [CrossRef] [PubMed]
- Vorwerk, S.; Biernacki, S.; Hillebrand, H.; Janzik, I.; Müller, A.; Weiler, E.W.; Piotrowski, M. Enzymatic characterization of the recombinant Arabidopsis thaliana nitrilase subfamily encoded by the NIT 2/ NIT 1/ NIT 3-gene cluster. Planta 2001, 212, 508–516. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Pu, H. Crystal structure of methylesterase family member 16 (MES16) from Arabidopsis thaliana. Biochemical and Biophysical Research Communications 2016, 474, 226–231. [Google Scholar] [CrossRef]
- Tognetti, V.B.; van Aken, O.; Morreel, K.; Vandenbroucke, K.; van de Cotte, B.; Clercq, I. de; Chiwocha, S.; Fenske, R.; Prinsen, E.; Boerjan, W.; et al. Perturbation of Indole-3-Butyric Acid Homeostasis by the UDP-Glucosyltransferase UGT74E2 Modulates Arabidopsis Architecture and Water Stress Tolerance. Plant Cell 2010, 22, 2660–2679. [Google Scholar] [CrossRef]
- Guo, R.; Hu, Y.; Aoi, Y.; Hira, H.; Ge, C.; Dai, X.; Kasahara, H.; Zhao, Y. Local conjugation of auxin by the GH3 amido synthetases is required for normal development of roots and flowers in Arabidopsis. Biochemical and Biophysical Research Communications 2022, 589, 16–22. [Google Scholar] [CrossRef]
- Sherp, A.M.; Westfall, C.S.; Alvarez, S.; Jez, J.M. Arabidopsis thaliana GH3.15 acyl acid amido synthetase has a highly specific substrate preference for the auxin precursor indole-3-butyric acid. Journal of Biological Chemistry 2018, 293, 4277–4288. [Google Scholar] [CrossRef]
- Takase, T.; Nakazawa, M.; Ishikawa, A.; Manabe, K.; Matsui, M. DFL2, a New Member of the Arabidopsis GH3 Gene Family, is Involved in Red Light-Specific Hypocotyl Elongation. Plant and Cell Physiology 2003, 44, 1071–1080. [Google Scholar] [CrossRef]
- Barbez, E.; Kubeš, M.; Rolčík, J.; Béziat, C.; Pěnčík, A.; Wang, B.; Rosquete, M.R.; Zhu, J.; Dobrev, P.I.; Lee, Y.; et al. A novel putative auxin carrier family regulates intracellular auxin homeostasis in plants. Nature 2012, 485, 119–122. [Google Scholar] [CrossRef]
- Gälweiler, L.; Guan, C.; Müller, A.; Wisman, E.; Mendgen, K.; Yephremov, A.; Palme, K. Regulation of Polar Auxin Transport by AtPIN1 in Arabidopsis Vascular Tissue. Science 1998, 282, 2226–2230. [Google Scholar] [CrossRef]
- Dai, Y.; Wang, H.; Li, B.; Huang, J.; Liu, X.; Zhou, Y.; Mou, Z.; Li, J. Increased Expression of MAP KINASE KINASE7 Causes Deficiency in Polar Auxin Transport and Leads to Plant Architectural Abnormality in Arabidopsis. Plant Cell 2006, 18, 308. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Li, B.; Li, S.; Liang, Y.; Wu, X.; Ma, M.; Wang, J.; Gao, J.; Cai, Y.; Zhang, Y.; et al. Mitogen-Activated Protein Kinase Cascade MKK7-MPK6 Plays Important Roles in Plant Development and Regulates Shoot Branching by Phosphorylating PIN1 in Arabidopsis. PLoS Biol 2016, 14, e1002550. [Google Scholar] [CrossRef] [PubMed]
- Tejos, R.; Rodriguez-Furlán, C.; Adamowski, M.; Sauer, M.; Norambuena, L.; Friml, J.; Russinova, J. PATELLINS are regulators of auxin-mediated PIN1 relocation and plant development in Arabidopsis thaliana. Journal of Cell Science 2018, 131, 20. [Google Scholar] [CrossRef] [PubMed]
- Perotti, M.F.; Ribone, P.A.; Cabello, J.V.; Ariel, F.D.; Chan, R.L. AtHB23 participates in the gene regulatory network controlling root branching, and reveals differences between secondary and tertiary roots. Plant J 2019, 100, 1224–1236. [Google Scholar] [CrossRef]
- Bargmann, B.O.R.; Birnbaum, K.D.; Brenner, E.D. An undergraduate study of two transcription factors that promote lateral root formation. Biochem Molecular Bio Educ 2014, 42, 237–245. [Google Scholar] [CrossRef]
- Liu, W.; Yu, J.; Ge, Y.; Qin, P.; Xu, L. Pivotal role of LBD16 in root and root-like organ initiation. Cell. Mol. Life Sci. 2018, 75, 3329–3338. [Google Scholar] [CrossRef]
- Zheng, Y.; Gao, Z.; Zhu, Z. DELLA–PIF Modules: Old Dogs Learn New Tricks. Trends Plant Sci. 2016, 21, 813–815. [Google Scholar] [CrossRef]
- Zhu, J.-Y.; Sae-Seaw, J.; Wang, Z.-Y. Brassinosteroid signalling. Development 2013, 140, 1615–1620. [Google Scholar] [CrossRef] [PubMed]
- Cui, D.; Zhao, J.; Jing, Y.; Fan, M.; Liu, J.; Wang, Z.; Xin, W.; Hu, Y.; Yu, H. The Arabidopsis IDD14, IDD15, and IDD16 Cooperatively Regulate Lateral Organ Morphogenesis and Gravitropism by Promoting Auxin Biosynthesis and Transport. PLoS Genet 2013, 9, e1003759. [Google Scholar] [CrossRef]
- Lee, Y.K.; Olson, A.; Kim, K.; Ohme-Takagi, M.; Ware, D. HB31 and HB21 regulate floral architecture through miRNA396/GRF modules in Arabidopsis. Plant Biotechnol Rep 2023, 149, 1713. [Google Scholar] [CrossRef]
- Rodriguez, R.E.; Ercoli, M.F.; Debernardi, J.M.; Breakfield, N.W.; Mecchia, M.A.; Sabatini, M.; Cools, T.; Veylder, L. de; Benfey, P.N.; Palatnik, J.F. MicroRNA miR396 Regulates the Switch between Stem Cells and Transit-Amplifying Cells in Arabidopsis Roots. Plant Cell 2016, 27, 3354–3366. [Google Scholar] [CrossRef]
- Zander, M.; Thurow, C.; Gatz, C. TGA Transcription Factors Activate the Salicylic Acid-Suppressible Branch of the Ethylene-Induced Defense Program by Regulating ORA59 Expression. Plant Physiol. 2014, 165, 1671–1683. [Google Scholar] [CrossRef]
- Rubinovich, L.; Weiss, D. The Arabidopsis cysteine-rich protein GASA4 promotes GA responses and exhibits redox activity in bacteria and in planta. Plant J 2010, 64, 1018–1027. [Google Scholar] [CrossRef]
- Hofmann, N.R. A NAC Transcription Factor for Flooding: SHYG Helps Plants Keep Their Leaves in the Air. Plant Cell 2014, 25, 4771. [Google Scholar] [CrossRef]
- Rauf, M.; Arif, M.; Fisahn, J.; Xue, G.-P.; Balazadeh, S.; Mueller-Roeber, B. NAC Transcription Factor SPEEDY HYPONASTIC GROWTH Regulates Flooding-Induced Leaf Movement in Arabidopsis. Plant Cell 2014, 25, 4941–4955. [Google Scholar] [CrossRef] [PubMed]
- Krishnaswamy, S.; Verma, S.; Rahman, M.H.; Kav, N.N.V. Functional characterization of four APETALA2-family genes (RAP2.6, RAP2.6L, DREB19 and DREB26) in Arabidopsis. Plant Mol Biol 2011, 75, 107–127. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Chang, Y.; Guo, J.; Chen, J.-G. Arabidopsis Ovate Family Protein 1 is a transcriptional repressor that suppresses cell elongation. Plant J 2007, 50, 858–872. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.L.; Fedoroff, N.V. LRP1, a gene expressed in lateral and adventitious root primordia of arabidopsis. Plant Cell 1995, 7, 735–745. [Google Scholar] [CrossRef]
- Singh, S.; Yadav, S.; Singh, A.; Mahima, M.; Singh, A.; Gautam, V.; Sarkar, A.K. Auxin signaling modulates LATERAL ROOT PRIMORDIUM 1 (LRP 1 ) expression during lateral root development in Arabidopsis. Plant J 2020, 101, 87–100. [Google Scholar] [CrossRef]
- Chen, X.; Qu, Y.; Sheng, L.; Liu, J.; Huang, H.; Xu, L. A simple method suitable to study de novo root organogenesis. Front. Plant Sci. 2014, 5, 208. [Google Scholar] [CrossRef]
- Xiong, Y.; Xing, Q.; Müller-Xing, R. A novel UV-B priming system reveals an UVR8-depedent memory, which provides resistance against UV-B stress in Arabidopsis leaves. Plant Signal. Behav. 2021, 16, 1879533. [Google Scholar] [CrossRef]
- Liu, X.; Xing, Q.; Liu, X.; Müller-Xing, R. Expression of the Populus Orthologues of AtYY1, YIN and YANG Activates the Floral Identity Genes AGAMOUS and SEPALLATA3 Accelerating Floral Transition in Arabidopsis thaliana. IJMS 2023, 24, 7639. [Google Scholar] [CrossRef]
- Müller-Xing, R.; Ardiansyah, R.; Xing, Q.; Faivre, L.; Tian, J.; Wang, G.; Zheng, Y.; Wang, X.; Jing, T.; Leau, E. de; et al. Polycomb proteins control floral determinacy by H3K27me3-mediated repression of pluripotency genes in Arabidopsis thaliana. J. Exp. Bot. 2022, 8, e0127. [Google Scholar] [CrossRef]
- Li, X.; Zheng, Y.; Xing, Q.; Ardiansyah, R.; Zhou, H.; Ali, S.; Jing, T.; Tian, J.; Song, X.S.; Li, Y.; et al. Ectopic expression of the transcription factor CUC2 restricts growth by cell cycle inhibition in Arabidopsis leaves. Plant Signal. Behav. 2020, 15, 1706024. [Google Scholar] [CrossRef]
- Müller-Xing, R.; Clarenz, O.; Pokorny, L.; Goodrich, J.; Schubert, D. Polycomb-Group Proteins and FLOWERING LOCUS T Maintain Commitment to Flowering in Arabidopsis thaliana. Plant Cell 2014, 26, 2457–2471. [Google Scholar] [CrossRef]
Figure 1.
Effects of GA and PBZ treatment on rooting rate of AR formation from hypocotyls and DNRR. (A-B) AR formation from hypocotyls. (A) Rooting phenotype of Col-0 seedlings under mock, GA, and PBZ treatment. Scale bar = 10 mm. (B) ARs per hypocotyls (Col-0) under mock, GA, and PBZ treatment 7 days after shift from dark to LD, 10 DAG. (C-H) DNRR from Col-0 explants under mock, GA and PBZ treatment (LD and Dark conditions). (C) The rooting rate of Col-0 leaf explants under mock, GA, and PBZ treatment, 9 DAC (LD). (D-E,H) Phenotype of Col-0 leaf explants under mock, GA, and PBZ treatment, 9 DAC (LD). The arrow indicates the absence of an AR (E). Note that the roots are thicker after PBZ treatment in comparison to mock (arrow heads in D and H). Scale bar = 500 µm. (G) Dark conditions accelerates rooting additive to PBZ treatment. Asterisks indicate significant changes (Student’s t-test: *, p < 0.05; **, p < 0.01; ***, p < 0.001).
Figure 1.
Effects of GA and PBZ treatment on rooting rate of AR formation from hypocotyls and DNRR. (A-B) AR formation from hypocotyls. (A) Rooting phenotype of Col-0 seedlings under mock, GA, and PBZ treatment. Scale bar = 10 mm. (B) ARs per hypocotyls (Col-0) under mock, GA, and PBZ treatment 7 days after shift from dark to LD, 10 DAG. (C-H) DNRR from Col-0 explants under mock, GA and PBZ treatment (LD and Dark conditions). (C) The rooting rate of Col-0 leaf explants under mock, GA, and PBZ treatment, 9 DAC (LD). (D-E,H) Phenotype of Col-0 leaf explants under mock, GA, and PBZ treatment, 9 DAC (LD). The arrow indicates the absence of an AR (E). Note that the roots are thicker after PBZ treatment in comparison to mock (arrow heads in D and H). Scale bar = 500 µm. (G) Dark conditions accelerates rooting additive to PBZ treatment. Asterisks indicate significant changes (Student’s t-test: *, p < 0.05; **, p < 0.01; ***, p < 0.001).
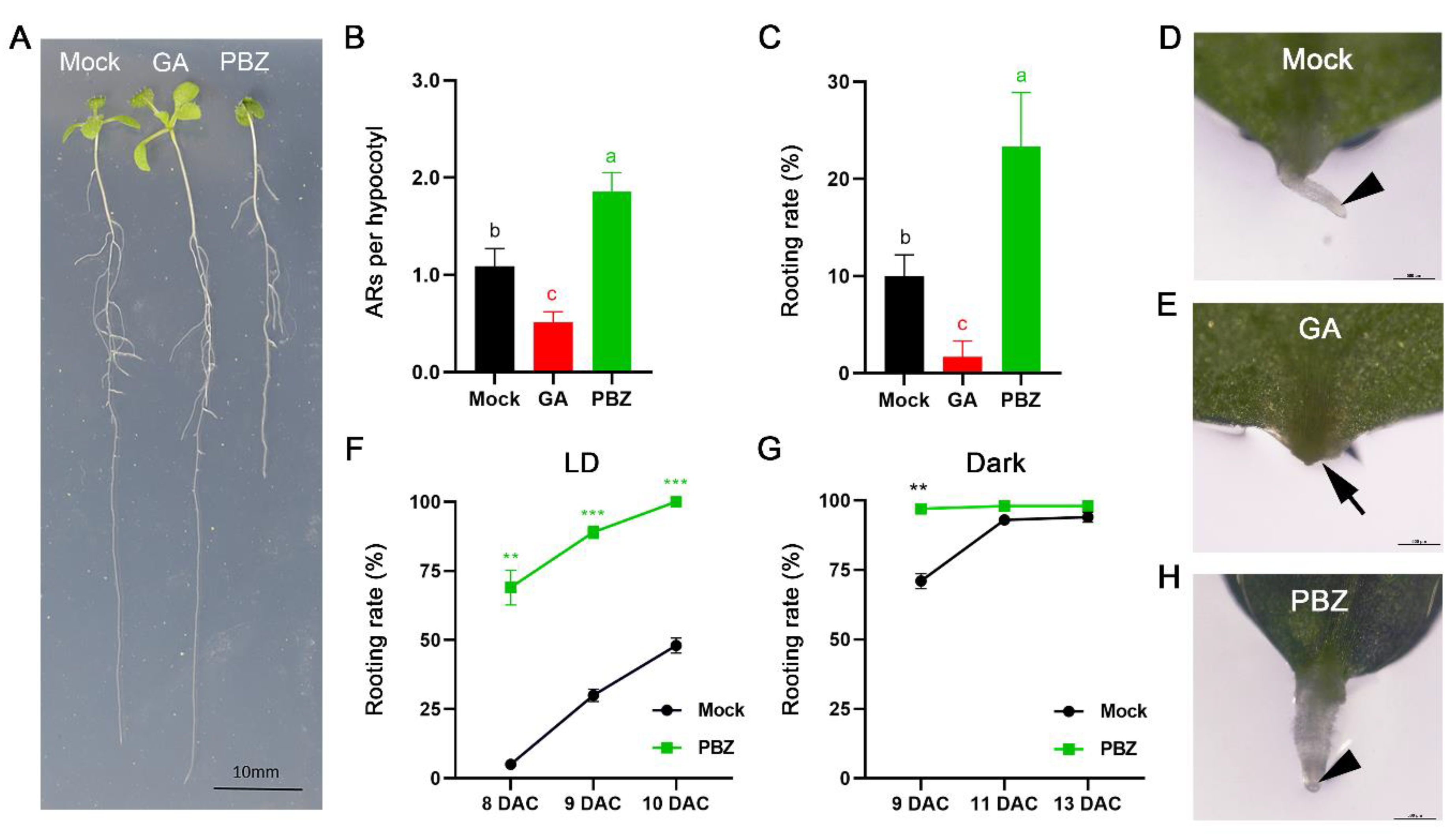
Figure 2.
Effects of differs hormone treatments on rooting rate (time courses) and rooting capacity in wild-type (Col-0) leaf explants in darkness. (A-C) Single treatments with IAA, NPA, GA and PBZ. (A) Rooting phenotype of Col-0 leaf explants. Scale bar = 10 mm. (B) Rooting rate. (C) The rooting capacity. Note that the rooting capacity was decreased by PBZ, while IAA treatment increased rooting rate and rooting capacity. (D-E) Treatments with NPA, PBZ and NPA+PBZ. Note that rooting is complete abolished by NPA treatment even in the presence of PBZ indicating that polar auxin transport is essential for the accelerating effects of PBZ on DNRR. (F-G) Treatments with IAA, PBZ and IAA+PBZ. Note that the accelerating effects on rooting rates by IAA is epistatic to PBZ treatment (no additive effect), but PBZ significantly reduced the root capacity even in the presents of IAA. Treatments: 0.1 µM IAA, 1 µM NPA, 1 µM GA and 5 µM, PBZ. Asterisks indicate significant changes (Student’s t-test: *, p < 0.05; **, p < 0.01; ***, p < 0.001).
Figure 2.
Effects of differs hormone treatments on rooting rate (time courses) and rooting capacity in wild-type (Col-0) leaf explants in darkness. (A-C) Single treatments with IAA, NPA, GA and PBZ. (A) Rooting phenotype of Col-0 leaf explants. Scale bar = 10 mm. (B) Rooting rate. (C) The rooting capacity. Note that the rooting capacity was decreased by PBZ, while IAA treatment increased rooting rate and rooting capacity. (D-E) Treatments with NPA, PBZ and NPA+PBZ. Note that rooting is complete abolished by NPA treatment even in the presence of PBZ indicating that polar auxin transport is essential for the accelerating effects of PBZ on DNRR. (F-G) Treatments with IAA, PBZ and IAA+PBZ. Note that the accelerating effects on rooting rates by IAA is epistatic to PBZ treatment (no additive effect), but PBZ significantly reduced the root capacity even in the presents of IAA. Treatments: 0.1 µM IAA, 1 µM NPA, 1 µM GA and 5 µM, PBZ. Asterisks indicate significant changes (Student’s t-test: *, p < 0.05; **, p < 0.01; ***, p < 0.001).
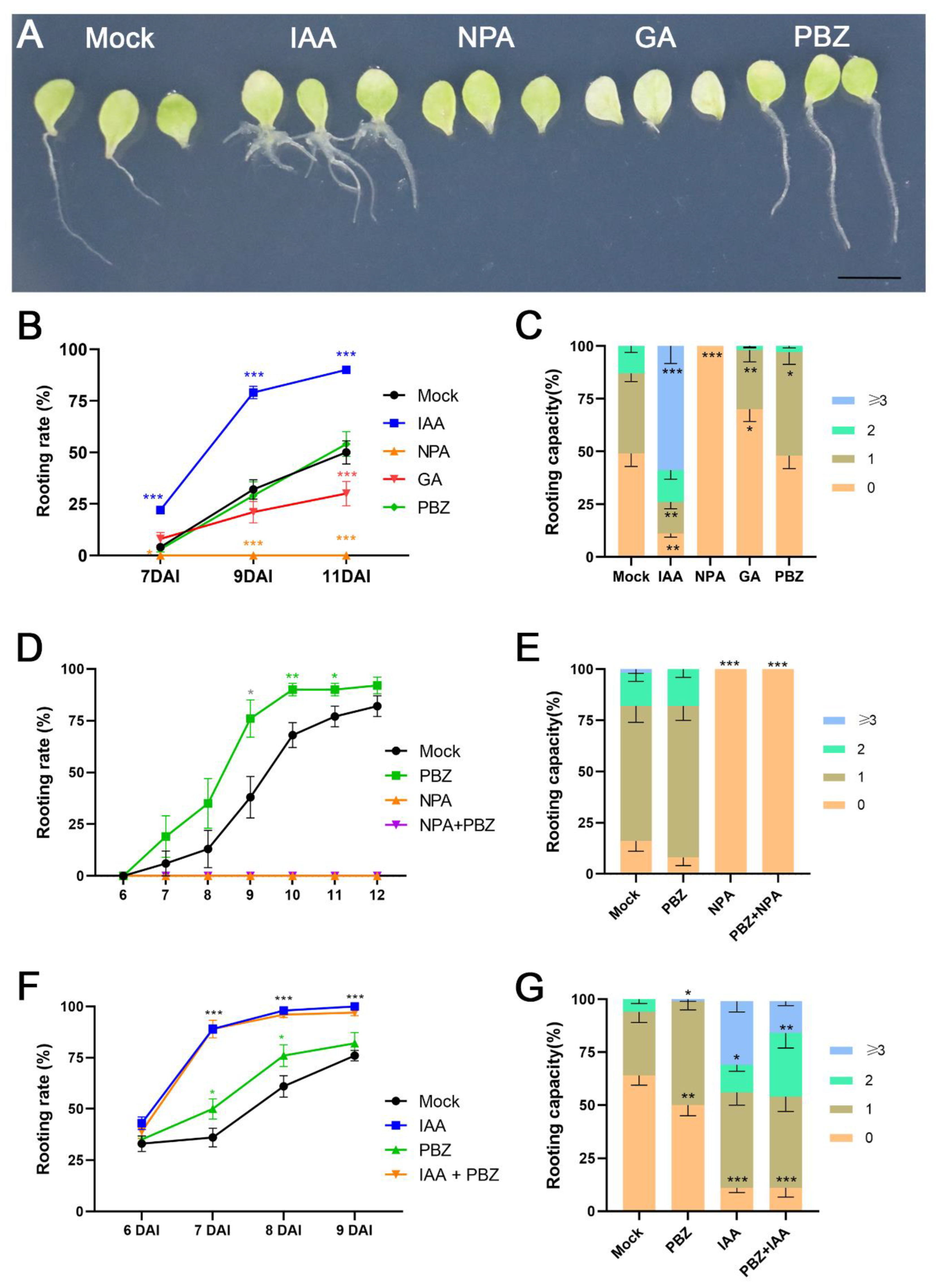
Figure 3.
Reanalysis of GA biosynthesis and GA signaling mutants reveals (i) the negative effects of GA and (ii) the positive effects of ER signaling on DNRR in darkness. (A) Rooting rate time course: The rooting rate of the three GA mutants ga1-4, ga5-1 and gai-1 are significant higher than in wild-type (Ler-0). (B) Rooting rate time course: In comparison to the wild-type Col-0, rooting was almost abolished in the leaf explants of both er-1 (Ler-0) and er-105 (Col-0) mutants. (C) Treatment with 5 µM PBZ can recue the low rooting rate of both er-1 (Ler-0) and er-105 (Col-0) mutants (9 DAC). Asterisks indicate significant changes (Student’s t-test: *, p < 0.05; **, p < 0.01; ***, p < 0.001).
Figure 3.
Reanalysis of GA biosynthesis and GA signaling mutants reveals (i) the negative effects of GA and (ii) the positive effects of ER signaling on DNRR in darkness. (A) Rooting rate time course: The rooting rate of the three GA mutants ga1-4, ga5-1 and gai-1 are significant higher than in wild-type (Ler-0). (B) Rooting rate time course: In comparison to the wild-type Col-0, rooting was almost abolished in the leaf explants of both er-1 (Ler-0) and er-105 (Col-0) mutants. (C) Treatment with 5 µM PBZ can recue the low rooting rate of both er-1 (Ler-0) and er-105 (Col-0) mutants (9 DAC). Asterisks indicate significant changes (Student’s t-test: *, p < 0.05; **, p < 0.01; ***, p < 0.001).
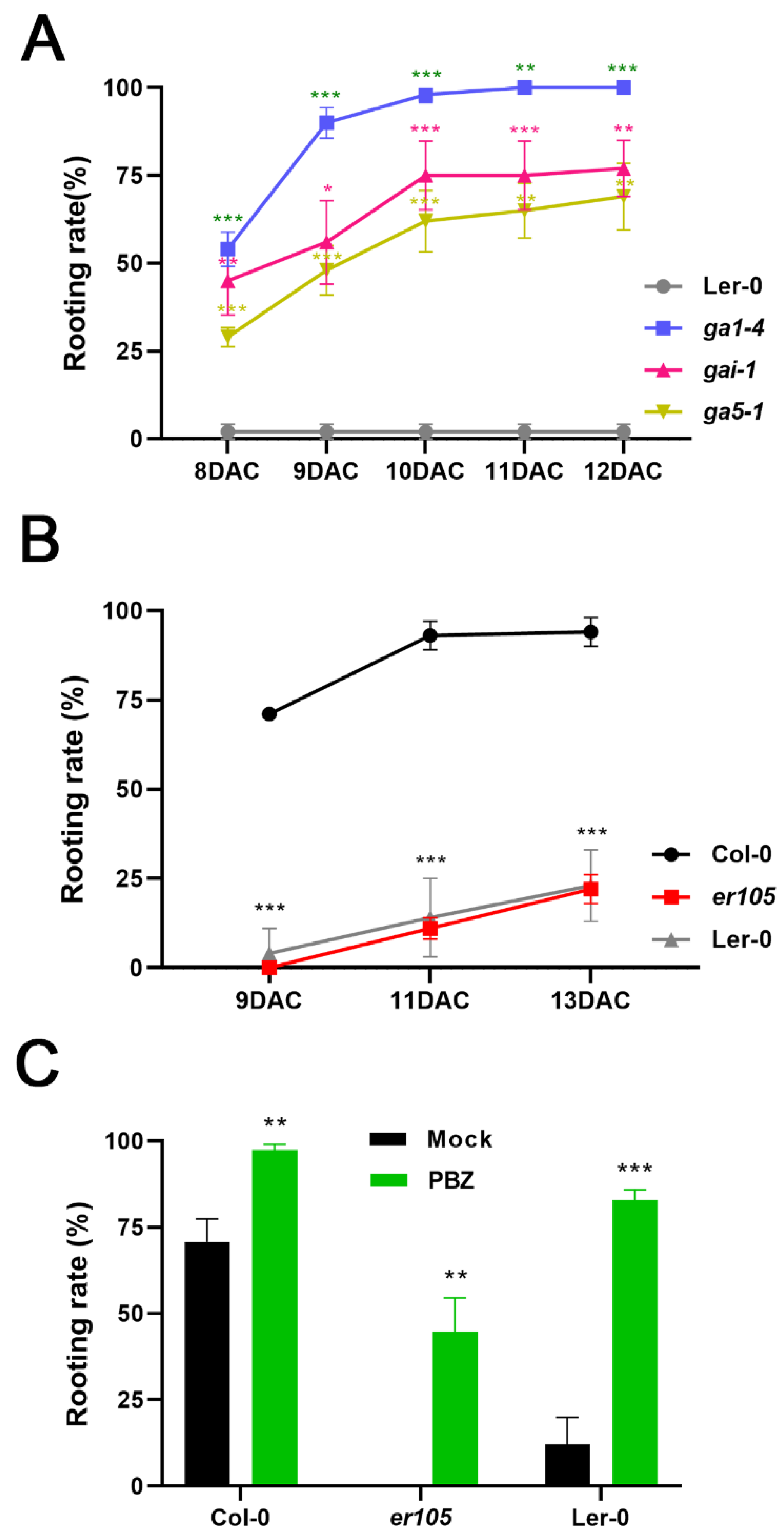
Figure 4.
Expression changes in response to PBZ treatment, 24 HAC. (A) Number of genes with significantly decreased (down) and increased (up) expression rates. (B) Percentage of GA/BR- and auxin-regulated genes and their overlaps. (C) Conceptional model of potential pathways how PBZ could increase LBD16 expression. Dotted lines, known interactions that seems to be covered during DNRR with PBZ. Broken lines, putative, probable indirect interactions. Note that most arrows indicate transcriptional regulation but DELLA proteins bind directly to BZR1, PIF and ARF6 proteins, PBZ blocks GA synthesis, and PIN1 is an auxin (IAA) transporter. (D) Expression changes in response to PBZ treatment (24 HAC) of genes that can theoretically increase LBD16 and LRP1 expression levels. Asterisks indicate significant changes (Student’s t-test: *, p < 0.05; **, p < 0.01). (E) Selection of genes with significantly decreased (down) and increased (up) expression rates. Regulation by GA/BR, IAA and/or wounding as well as promoter binding by PIF4, BZR1 and ARF6 is indicated.
Figure 4.
Expression changes in response to PBZ treatment, 24 HAC. (A) Number of genes with significantly decreased (down) and increased (up) expression rates. (B) Percentage of GA/BR- and auxin-regulated genes and their overlaps. (C) Conceptional model of potential pathways how PBZ could increase LBD16 expression. Dotted lines, known interactions that seems to be covered during DNRR with PBZ. Broken lines, putative, probable indirect interactions. Note that most arrows indicate transcriptional regulation but DELLA proteins bind directly to BZR1, PIF and ARF6 proteins, PBZ blocks GA synthesis, and PIN1 is an auxin (IAA) transporter. (D) Expression changes in response to PBZ treatment (24 HAC) of genes that can theoretically increase LBD16 and LRP1 expression levels. Asterisks indicate significant changes (Student’s t-test: *, p < 0.05; **, p < 0.01). (E) Selection of genes with significantly decreased (down) and increased (up) expression rates. Regulation by GA/BR, IAA and/or wounding as well as promoter binding by PIF4, BZR1 and ARF6 is indicated.
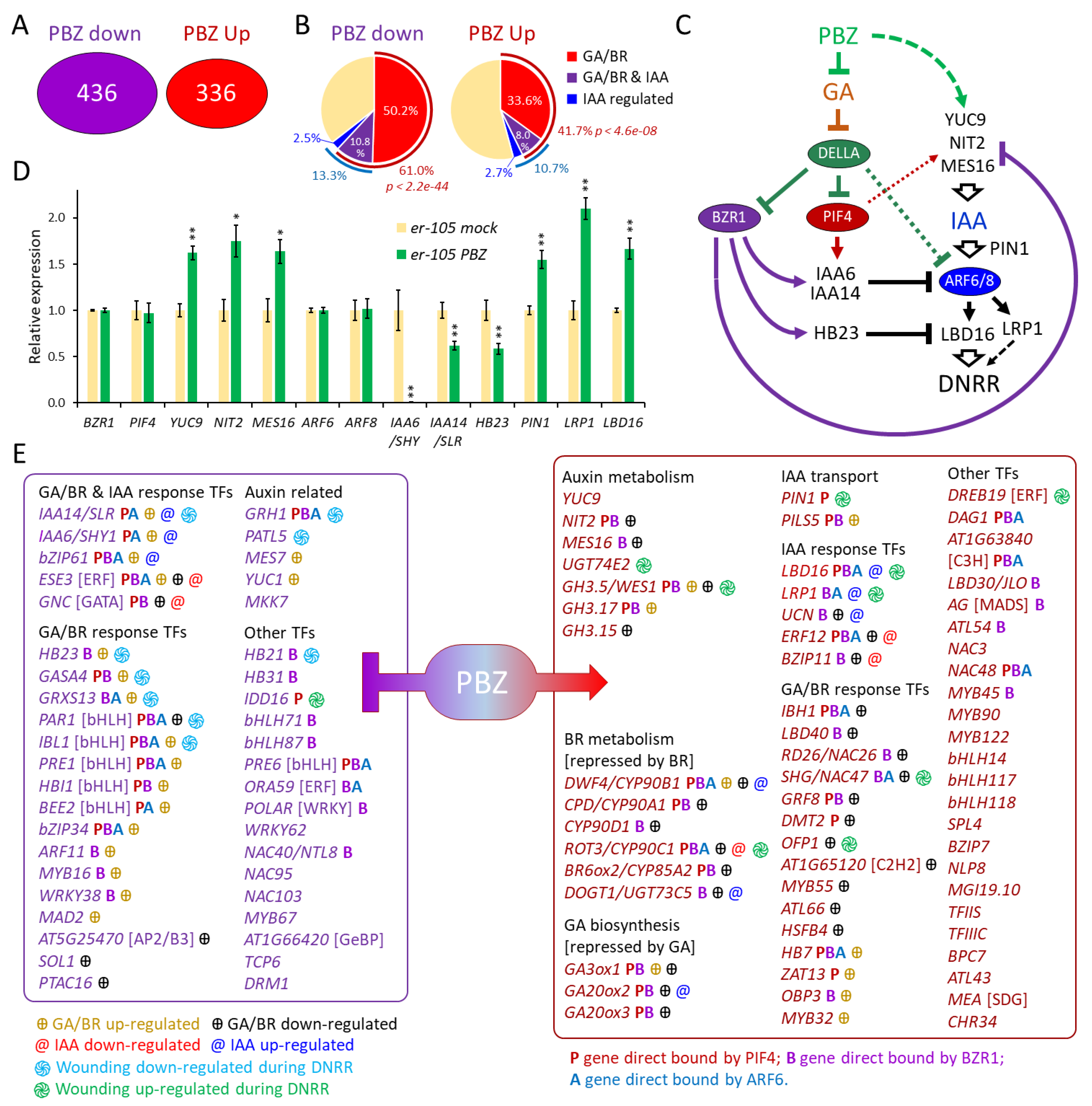
Figure 5.
Time courses of LBD16::GUS expression in response to mock, PBZ, PBZ+GA and PBZ+NPA treatment. HAC, hours after cultivation (induction). (A-C) Mock treatment. Note that LBD16::GUS expression is evidently visible at the future rooting site after 48 HAC (C and C'). (D-E) PBZ treatment. Note that LBD16::GUS expression is stronger after PBZ (48 HAC, E and E'). (D-E) PBZ+GA treatment. Note that LBD16::GUS expression is strongly reduced by GA (48 HAC, G and G'). (F-G) PBZ+GA treatment. Note that LBD16::GUS expression is strongly reduced by GA (48 HAC, G and G'). (H-I) PBZ+NPA treatment. Note that LBD16::GUS expression is fully quenched by NPA (48 HAC, I and I'). Scale bar = 250 µm.
Figure 5.
Time courses of LBD16::GUS expression in response to mock, PBZ, PBZ+GA and PBZ+NPA treatment. HAC, hours after cultivation (induction). (A-C) Mock treatment. Note that LBD16::GUS expression is evidently visible at the future rooting site after 48 HAC (C and C'). (D-E) PBZ treatment. Note that LBD16::GUS expression is stronger after PBZ (48 HAC, E and E'). (D-E) PBZ+GA treatment. Note that LBD16::GUS expression is strongly reduced by GA (48 HAC, G and G'). (F-G) PBZ+GA treatment. Note that LBD16::GUS expression is strongly reduced by GA (48 HAC, G and G'). (H-I) PBZ+NPA treatment. Note that LBD16::GUS expression is fully quenched by NPA (48 HAC, I and I'). Scale bar = 250 µm.
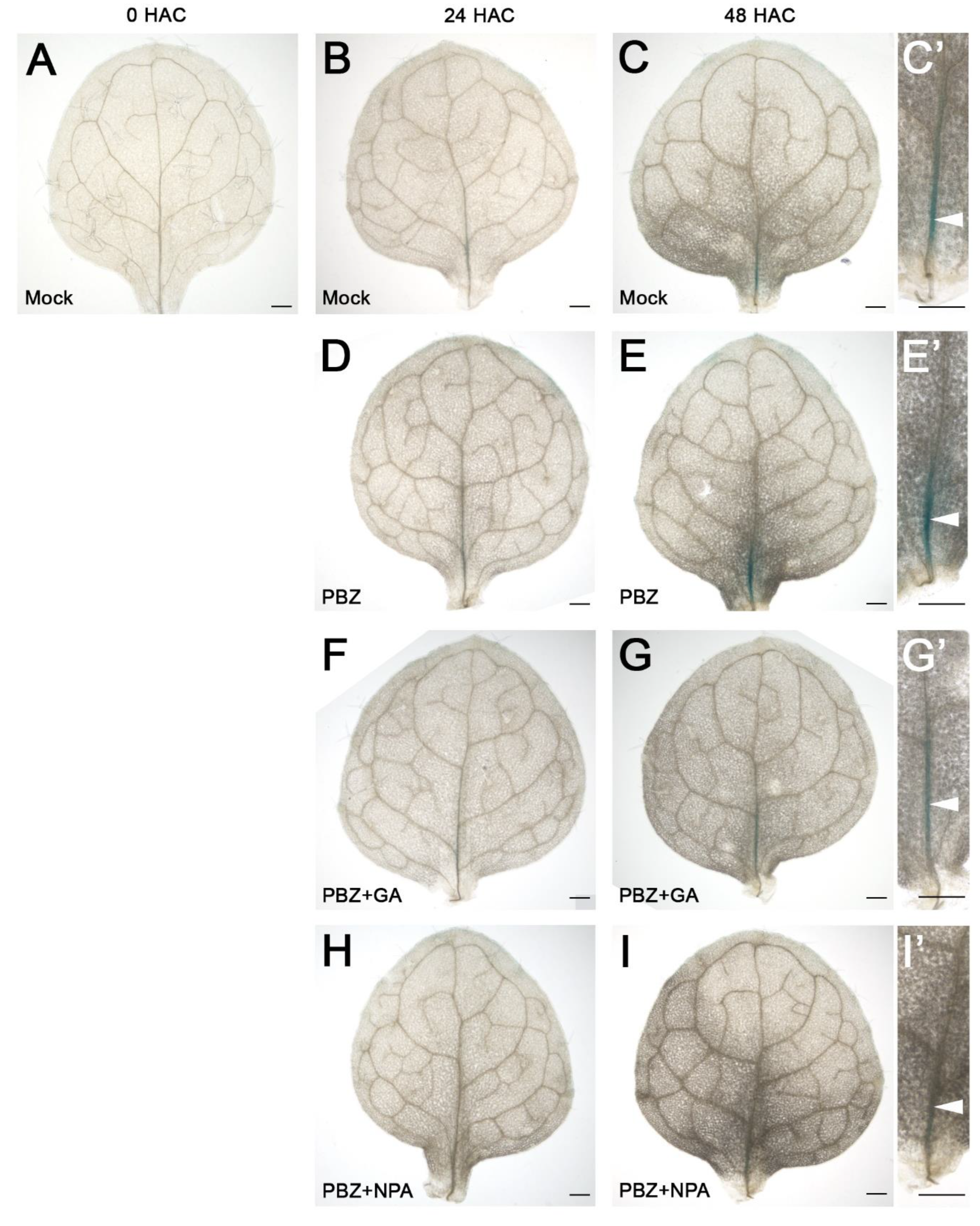
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



