
Submitted:
14 October 2024
Posted:
15 October 2024
You are already at the latest version
Abstract
Plants are continuously exposed to environmental challenges, including pollutants, pesticides, and heavy metals, collectively termed xenobiotics. These substances induce oxidative stress by generating reactive oxygen species (ROS), which can damage cellular components such as lipids, proteins, and nucleic acids. To counteract this, plants have evolved complex metabolic pathways involving both enzymatic and non-enzymatic mechanisms to detoxify and process these harmful compounds. Oxidative stress in plants primarily arises from the overproduction of ROS, including hydrogen peroxide (H2O2), superoxide anions (O2•−), singlet oxygen (1O2), and hydroxyl radicals (•OH), by-products of metabolic activities such as photosynthesis and respiration. Exposure to xenobiotics significantly elevates ROS levels, causing cellular damage and disrupting metabolism. Plants employ a robust antioxidant defense system featuring enzymatic antioxidants like catalase (CAT), superoxide dismutase (SOD), and various peroxidases (POD). These enzymes collaborate to scavenge ROS, minimizing their detrimental effects. Additionally, plants produce numerous non-enzymatic antioxidants, such as flavonoids, phenolic acids, vitamins C and E, which neutralize ROS and regenerate other antioxidants, providing comprehensive protection against oxidative stress. Xenobiotic metabolism in plants involves three phases: Phase I (modification), Phase II (conjugation), and Phase III (compartmentalization), facilitated by enzymes like cytochrome P450 monooxygenases (CYPs), transferases such as glutathione S-transferases (GSTs), and ATP-binding cassette (ABC) transporters. Plant secondary metabolites, including alkaloids, terpenoids, and flavonoids, significantly contribute to detoxification and antioxidant defense. Understanding these oxidative processes and xenobiotic metabolism pathways in plants is crucial, as it offers insights into developing plant-based strategies to mitigate environmental pollution impacts and improve human health through enhanced detoxification and antioxidant mechanisms, as we discuss in this review.
Keywords:
plant metabolites
; xenobiotic interactions
; bioactive compounds
; metabolomics
; drug metabolism
; human health
1. Introduction
Understanding the interactions between plant metabolites and environmental xenobiotics, including pesticides, heavy metals, volatile organic compounds, and pharmaceuticals from treated wastewater and biosolids, is crucial due to their increasing exposure and potential impact [1]. A new approach in plant pharmacology aims to understand how pollutants interact with plants by studying plant pharmacokinetics (absorption, distribution, metabolism, and accumulation of xenobiotics) and pharmacodynamics (effects on plant enzymes and biochemical pathways). This dual framework enhances knowledge of xenobiotic behavior in plants, supports model development, and improves risk assessments for environmental and human health [1]. Plant metabolites, including alkaloids, polyphenols, flavonoids, saponins, and terpenes, are recognized for their bioactive properties and potential health benefits [2,3,4,5]. They play pivotal roles in plant defence, growth, and development, while recent research has highlighted also their significant interactions with xenobiotics, which encompass pharmaceutical drugs, environmental pollutants, and dietary compounds [6,7]. Contaminants that impact human health have become a significant concern, leading to numerous investigations into methods for mitigating their harmful effects [8]. Recent research has highlighted various biological roles of natural dietary compounds found in plants. These compounds have shown promise in disease prevention and in countering chemically-induced cancer. The toxicity and carcinogenic effects of contaminants are primarily due to the mutagenic properties of reactive metabolites and the disruption of normal biological processes. Thus, effectively managing the metabolism of these hazardous substances is crucial for minimizing their adverse health impacts. Additionally, enhancing the elimination of contaminants from the body through metabolic processes is a valuable approach for reducing toxicity [8]. Maintaining balanced levels of reactive oxygen species (ROS) and reactive nitrogen species (RNS) is crucial for normal cellular function, a process known as redox homeostasis [9]. Key ROS include superoxide radicals (O2•−), hydrogen peroxide (H2O2), hydroxyl radicals (•OH), and singlet oxygen (1O2), while nitric oxide (•NO) is a primary RNS, all of which emerge as by-products of cellular metabolism under physiological and pathological conditions [10]. Enzymes such as cytochrome P450 (CYP), nitric oxide synthase (NOS), xanthine oxidase (XO), NADPH oxidase (NOX), cyclooxygenase (COX), and lipoxygenase (LOX) are central to the production of intracellular ROS [11,12] with NOX, composed of membrane-bound proteins gp91phox and p22phox, generating superoxide radicals by transferring electrons from NADPH to oxygen [13]. On the other hand, XO produces substantial ROS during the catalytic conversion of hypoxanthine and xanthine to uric acid [14], whereas the CYP family is involved in phase I xenobiotic metabolism and can produce ROS through reaction uncoupling [15]. NOS catalyzes the production of nitric oxide radicals during the conversion of L-arginine to citrulline [16], while COX and LOX enzymes oxidize arachidonic acid (AA) to produce ROS, which in turn stimulate NOX [17]. This review delves into the oxidative processes and xenobiotic metabolism in plants, highlighting how plants manage environmental challenges such as pollutants, pesticides, and heavy metals.
1.1. Highlights
- Xenobiotic defence mechanisms in plants: the study emphasizes how plants are equipped with intricate enzymatic (e.g., cytochrome P450, peroxidases) and non-enzymatic (e.g., flavonoids, vitamins C and E) antioxidant systems to defend against oxidative stress induced by xenobiotics like pollutants, pesticides, and heavy metals.
- Oxidative stress pathways: xenobiotic exposure elevates ROS such as hydrogen peroxide, superoxide anions, and hydroxyl radicals, which damage cellular components. Plants counteract this with antioxidants like catalase, superoxide dismutase, and glutathione.
- Therapeutic potential: the metabolic pathways in plants offer valuable insights into developing plant-based detoxification strategies that can mitigate the impact of environmental pollution and improve human health by harnessing plant metabolites.
1.2. Significance of the Study
- Environmental health insights: this research contributes to understanding how plants can manage the adverse effects of xenobiotics, providing strategies to mitigate the impacts of environmental pollution on plant and human health.
- Therapeutic applications: by exploring the xenobiotic metabolism in plants, the study opens up possibilities for developing plant-based remedies or enhancing the dietary use of plants to boost human health through their detoxification and antioxidant properties.
- Redox homeostasis and disease prevention: understanding how plants maintain redox homeostasis offers insights into preventing diseases caused by oxidative stress, such as cancer, highlighting the role of plant-based compounds in counteracting chemically induced toxicity.
- Model for risk assessment: the dual focus on plant pharmacokinetics and pharmacodynamics enhances the ability to model and assess risks posed by environmental contaminants and develop new plant-based strategies for detoxification.
2. Oxidative Processes and Xenobiotic Metabolism by Plants
2.1. Environmental Challenges and Oxidative Stress in Plants
Plants face ongoing environmental challenges such as pollutants, pesticides, and heavy metals, collectively known as xenobiotics [18,19]. These substances induce oxidative stress by generating reactive oxygen species, which can harm cellular components such as lipids, proteins, and nucleic acids. To counteract this, plants have evolved intricate metabolic pathways that utilize both enzymatic and non-enzymatic mechanisms to detoxify and process these harmful compounds [20,21]. Oxidative stress in plants primarily arises from the overproduction of ROS, including H2O2, O2•−, 1O2, and •OH, which are by-products of metabolic activities like photosynthesis and respiration [22,23]. Exposure to xenobiotics can significantly elevate ROS levels, causing cellular damage and disrupting metabolism. Plants employ a robust antioxidant defense system to mitigate oxidative damage, prominently featuring enzymatic antioxidants such as catalase (CAT), superoxide dismutase (SOD), and several types of peroxidases (POD) [24]. These enzymes collaborate to scavenge ROS and minimize their detrimental effects: SOD converts superoxide anions into hydrogen peroxide and oxygen, while CAT breaks down hydrogen peroxide into water and oxygen, thus reducing oxidative stress. Peroxidases utilize hydrogen peroxide to oxidize diverse substrates, further detoxifying ROS [24]. Additionally, plants produce numerous non-enzymatic antioxidants, such as flavonoids, phenolic acids, vitamin C, and vitamin E, which directly neutralize ROS, chelate metal ions, and regenerate other antioxidants, thereby providing comprehensive protection against oxidative stress [25].
2.2. Xenobiotic Metabolism in Plants
Xenobiotics are metabolized in plants through a sequence of biochemical reactions known as xenobiotic metabolism, typically divided into three phases. Phase I involves oxidation, reduction, or hydrolysis of xenobiotics, often mediated by cytochrome P450 monooxygenases [26], which increase the compound’s reactivity and solubility. In Phase II, the reactive metabolites from Phase I are conjugated with endogenous molecules like glutathione, sugars, or amino acids by transferases such as glutathione S-transferases (GSTs), enhancing their solubility and facilitating detoxification and excretion. Phase III involves the transport of conjugated xenobiotics into vacuoles or extracellular spaces via ATP-binding cassette (ABC) transporters, preventing their interaction with cellular components and reducing toxicity [27]. Plant secondary metabolites, including alkaloids, terpenoids, and flavonoids, play crucial roles in xenobiotic metabolism and detoxification [28]. These compounds can modulate the activity of detoxification enzymes, bolster antioxidant defenses, and directly interact with xenobiotics to neutralize their toxic effects. For example, flavonoids can inhibit ROS production and enhance GST activity, while terpenoids, noted for their antioxidant properties, shield plant cells from oxidative damage induced by xenobiotics. Understanding the oxidative processes and xenobiotic metabolism in plants holds significant implications for human health. Many plants used in traditional medicine [29] contain bioactive metabolites that detoxify xenobiotics and alleviate oxidative stress (Table 1, Scheme 1) [30].
Integrating these plants into the diet or utilizing their compounds in pharmacology can enhance the body’s defenses against xenobiotics, potentially lowering the risk of diseases associated with oxidative stress and xenobiotic exposure. For example, the metabolites—especially flavonoids, saponins, and tannins—in leaf extracts of Solanum aethiopicum, whose fruits are widely used in ethnomedicine, are believed to be responsible for the ameliorative effects on phenylhydrazine-induced anemia in rats. This demonstrates the plant’s potential in mitigating the toxic effects of phenylhydrazine, a known xenobiotic compound (Figure 1) [50].
The results of the mentioned study indicated a significant increase in packed cell volume, haemoglobin, and red blood cell counts in both male and female rats compared to other groups. Furthermore, the S. aethiopicum extract reduced levels of aspartate aminotransferase (AST), alanine transaminase (ALT), alkaline phosphatase (ALP), urea, creatinine, and chloride, supporting the use of S. aethiopicum leaf extracts as an effective anti-anaemic tonic with a wide margin of safety and potential hepato/reno-protective properties [50]. Moreover, the same plant was found to have protective effects also against sodium fluoride that notably reduces sperm quality, hormone levels, antioxidant concentrations, and overall body weight, while significantly increasing the levels of markers of oxidative stress. It also leads to structural changes in the seminiferous tubules, including almost empty lumens and vascular hemorrhage. However, these adverse effects were mitigated by extracts from fruits of Solanum aethiopicum and Vitamin C [51]. Besides the specific effects of Solanum aethiopicum against xenobiotics, plants have evolved sophisticated mechanisms to manage oxidative stress and metabolize xenobiotics, employing a blend of enzymatic and non-enzymatic antioxidants alongside complex metabolic pathways [52,53]. These adaptations safeguard plants from environmental stresses and offer promising therapeutic avenues for improving human health by mitigating the adverse effects of xenobiotics.
3. The Effect of Oxidative Imbalance on Environmental Pollutants
Oxidative stress occurs when there is an imbalance between pro-oxidants and antioxidants, resulting in cellular damage. Essentially, it happens when the production of free radicals exceeds the body’s capacity to neutralize them with antioxidants [54]. Free radicals, such as reactive oxygen species and reactive nitrogen species, are generated at a rate of approximately fifteen thousand per cell per day in healthy humans. These include superoxide anions, hydroxyl radicals, hydrogen peroxide, singlet oxygen, and peroxyl radicals. While cellular ROS production is a well-documented phenomenon, their effects can be both beneficial and harmful under normal and pathological conditions. Moderate levels of ROS are crucial for various cellular functions like proliferation, immune responses, metabolism, and survival, as they regulate signaling pathways [55,56]. For instance, mitochondrial ROS (mROS) play pivotal roles in erythroid progenitor cell proliferation, red blood cell formation, human fibroblast cell proliferation, vascular smooth muscle cell activation, and T-lymphocyte proliferation and activation. ROS levels increase in response to platelet-derived growth factor (PDGF), essential for the growth of primary vascular smooth muscle cells (VSMCs) [57]. Inhibiting PDGF-induced ROS or enhancing catalase levels to scavenge ROS has been shown to reduce cell growth. Additionally, ROS regulate cell cycle progression, apoptosis, and cytokine and hormonal signaling through phosphoinositide-3-kinase (PI3K) [58] activation and downstream elements. Despite their critical roles in normal cellular functions, high ROS levels are cytotoxic. In particular, e levated ROS levels are associated with cancer cell proliferation, as cancer cells produce large amounts of ROS, particularly H2O2, required for mitogenic signaling. Loss of tumor suppressor genes and expression of oncogenes in a manner dependent on mROS have been observed in mouse embryonic fibroblast cells. Mutations in mitochondrial DNA (mDNA) encoding respiratory electron transport chain subunits lead to increased mROS levels, promoting tumorigenesis in mouse models and human and mouse cell lines. Heterozygous mice for mitochondrial transcription factor A (TFAM+/-), a stabilizing protein for mDNA, showed increased tumorigenesis in an mROS-dependent manner in intestinal carcinoma [59]. Free radicals act as signaling molecules at moderate concentrations, but their highly reactive nature necessitates strict regulation by the cellular antioxidant defense system to maintain redox balance. This balance is critical for normal cellular functioning and homeostasis. Consequently, biological processes generating ROS are coupled with various ROS scavenging mechanisms utilizing enzymatic and non-enzymatic antioxidants. Enzymatic antioxidants like SOD, CAT, and glutathione peroxidase [60,61,62] constitute a primary defense mechanism.
3.1. Environmental Pollutants and Their Impact on Oxidative Stress
Non-enzymatic antioxidants, including vitamins C, and A, GSH (Figure 2), carotenoids, and flavonoids, also play significant roles in maintaining redox homeostasis. Vitamin C, a hydrophilic antioxidant, acts alone or synergistically with vitamin E, carotenoids, and antioxidant enzymes to neutralize free radicals [59].
Vitamin E, a lipophilic antioxidant, effectively neutralizes lipid peroxyl radicals. GSH, a tripeptide found abundantly in the nucleus, mitochondria, and cytoplasm, actively participates in ROS neutralization and repairs ROS-induced cellular damage. Despite these antioxidant systems, exposure to environmental toxicants can disrupt redox balance, leading to elevated ROS levels. This accumulation of ROS is a primary consequence of environmental toxicant exposure, causing cellular damage by impairing biomolecules and disrupting cell survival pathways. Different environmental toxicants induce various biological effects by interfering with biological processes and targeting specific organs. Generally, ROS formation is a fundamental step in environmental toxicant-induced biological anomalies. The mechanisms of ROS generation vary depending on the toxicant’s chemical nature, dose, route, and duration of exposure. Environmental toxicants induce ROS formation by inhibiting the antioxidant defense system, initiating Fenton reactions (Figure 3a), causing mitochondrial dysfunction, and generating harmful chemical metabolites. For example, heavy metals such as arsenic, chromium, cadmium, and lead inhibit SOD, CAT, GPx, and GSH, leading to redox imbalance. The Fenton reaction (Figure 3a) [63,64,65], activated by heavy metals, generates free radicals through cascading reactions involving H2O2. Mitochondrial dysfunction, disturbed mitochondrial membrane potential (MMP), and production of harmful metabolites from volatile organic compounds also contribute to increased ROS levels. Exposure to environmental toxicants like cadmium and benzene can inhibit antioxidant enzymes, activate NADPH oxidase (which disrupts cellular homeostasis, promotes pathological conditions, and contributes to the development of various diseases), and produce harmful intermediates such as hydroquinone (Figure 3b), resulting in oxidative stress and cellular damage [59].
Given the diverse impact of environmental toxicants on oxidative stress and cellular damage, Table 2 presents a summary of some natural products that have been studied for their protective effects against such toxicity.
4. Phytochemicals as Free Radical Neutralizers in Combating Chemical-Induced Toxicity
Reproductive health, a vital aspect of overall well-being, intricately linked to both individual and population health, is influenced by a multitude of factors, including environmental exposures, which can significantly impact developmental and reproductive processes. In this context, understanding how to mitigate the negative effects of environmental stressors is crucial. One of the emerging areas of research focuses on the role of phytochemicals as free radical neutralizers in combating chemical-induced toxicity. These natural compounds, found in a variety of plants, offer promising protective effects against oxidative stress caused by free radicals. In particular, advances in understanding developmental and reproductive biology have been significant, yet unraveling their intricate interaction with the environment remains complex. Key factors such as genetics, nutrition, temperature, and environmental influences all play critical roles in ensuring healthy development and reproduction.
4.1. The Impact of Reactive Oxygen Species
Previous studies have elucidated the dual effects of reactive oxygen species levels on embryonic development, organismal growth, and reproductive systems. ROS are indispensable for early embryonic cell cycles in Xenopus embryos and facilitate rapid wound healing in embryos of Drosophila and zebrafish [79,80]. Additionally, ROS act as secondary messengers, modulating various signaling pathways during embryogenesis. However, heightened ROS levels have been linked to compromised embryo development and complications during pregnancy. In males, maintaining low ROS levels is essential for supporting crucial processes like sperm maturation, acrosome reaction, and sperm-oocyte fusion. Similarly, females require precisely regulated ROS levels for successful oocyte maturation and favorable pregnancy outcomes. Conversely, excessive ROS contributes to male infertility issues such as sperm mortality and female reproductive disorders such as polycystic ovary syndrome and endometriosis, often resulting in spontaneous abortions. Oxidative stress induced by environmental toxicants poses substantial risks to both development and reproduction, prompting increasing interest in plant-based interventions.
4.2. The Role of Boerhavia diffusa in Counteracting Toxic Effects in Drosophila
Traditional herbs like Boerhavia diffusa Linn show promise in reversing developmental and reproductive toxicity caused by substances like toluene in Drosophila larvae [81]. More in detail, a study including a preliminary phytochemical screening and HPTLC profiling of B. diffusa aqueous extract (BDAE) determined the LC50 of toluene, with a sublethal dose of 200 ppm being fixed for the investigation. Four doses of BDAE (25, 50, 100, and 200 mg/ml) were used, designated as low dose, medium dose 1, medium dose 2, and high dose. The parameters assessed included the larval period, pupal period, percentage of egg hatching, morphometric analysis of eggs, larvae, pupae, and adults, fertility, fecundity, lifespan, and levels of antioxidant enzymes such as catalase, glutathione-S-transferase, and superoxide dismutase. Overall, the results of this study showed that the phytochemical and HPTLC characteristics were consistent with pharmacopoeial standards. The LC50 of toluene was found to be 430 ppm, whereas BDAE at medium dose 2 and high dose significantly mitigated the adverse effects on reproductive and developmental parameters, including larval period, pupal period, percentage of egg hatching, morphometric characteristics of larvae, pupae, and adults, fertility, fecundity, and lifespan in Drosophila. Additionally, the extract significantly elevated the levels of antioxidant enzymes. Overall, B. diffusa, with its rich active ingredients, effectively prevented toluene-induced developmental and reproductive toxicity in Drosophila. This medicinal herb offers a promising solution for preventing reproductive and developmental toxicity induced by environmental toxins [81].
4.3. Additional Phytochemicals and Their Impacts
Isoliquiritigenin (ISL, Figure 4a) from Glycyrrhiza uralensis has demonstrated protective effects against developmental abnormalities induced by environmental chemicals in zebrafish embryos by enhancing antioxidant enzyme activities such as SOD and CAT [82].
Specifically, the study focused on 2,2’,4,4’-tetrabromodiphenyl ether (BDE47, Figure 4a), the most abundant polybrominated diphenyl ether (PBDE) found in biological samples. BDE47 has a strong tendency to bioaccumulate and potentially endangers mammalian development through oxidative stress. ISL [83], an emerging natural chalcone-type flavonoid, possesses various biological and pharmacological properties, including antioxidant, anti-allergic, anti-inflammatory, anti-tumor, and estrogenic activities. Interestingly, a recent study explored the antioxidant effect of ISL on ameliorating developmental anomalies induced by BDE47. Zebrafish (Danio rerio) [84] embryos were exposed to BDE47 (1 and 10 μM) and/or ISL (4 μM) for 4 to 120 hours post-fertilization, and the morphology, development, behavior, oxidative stress status, and related gene expressions were assessed. The results showed that BDE47 caused dose-dependent growth retardation and deformities, including delayed hatching, spinal curvature, reduced body length, increased death rate, aberrant behaviors, and impaired dark-adapted vision, which were significantly mitigated by ISL. Furthermore, ISL ameliorated excessive ROS accumulation and altered the expressions of apoptosis-related genes p53, Bcl-2, caspase 3, and caspase 9 induced by BDE47. This suggests that ISL protected zebrafish from the developmental toxicity of BDE47 by inactivating programmed apoptosis and activating antioxidant signaling pathways. Overall, developing ISL as a dietary supplement might be a promising preventive strategy for ameliorating developmental toxicity induced by environmental pollutants [82].
4.4. Mitigating Ovarian and Uterine Toxicity
Calliandra portoricensis (CP) has proven effective in alleviating ovarian and uterine toxicity in rats exposed to carcinogens due to its anti-inflammatory and antioxidant properties [85]. In fact, the study of Adefisan et al addressed the traumatic impact of infertility resulting from reproductive impairment within families. More in detail, the ovarian and uterine toxicity induced by benzo[a]pyrene (BaP) and N-methyl nitrosourea (NMU) was examined, along with the mitigating effects of Calliandra portoricensis [85]. The aim was to shed light on folk medical claims, uncover lost knowledge, advance natural chemopreventive agents, and present new evidence regarding CP’s effects. Although CP is known for its anticonvulsant, antidiarrheal, antipyretic, antirheumatic, and analgesic properties, the mentioned study demonstrated also its potential role in mitigating toxicity from chemical exposures. The findings confirmed and expanded on CP’s ability to reduce toxic responses resulting from oxidative damage and inflammatory reactions associated with NMU and BaP exposure. Therefore, the development of phytochemicals derived from CP may offer a natural therapy against chemical toxicities for individuals inadvertently exposed, thereby promoting human health and reproductive well-being [85].
4.5. Further Botanical Studies and Their Findings
Other botanical studies highlight sweet potato (Ipomoea batatas L. Lam.) extract, which has boosted antioxidant levels and reproductive health in male rats exposed to bisphenol A, and Mentha spicata extract, which has mitigated nicotine-induced oxidative stress in rat brains and testes by restoring antioxidant enzyme levels [86]. More in detail, the study focused on sweet potato [87], known as "Shakarqandi" in Pakistan, an important root vegetable traditionally used for its aphrodisiac, antiprostatic, anti-inflammatory, antidiabetic, cardiotonic, and anticancer properties, aimed to gauge the aphrodisiac potential of Ipomoea batatas ethyl acetate (IPT-EA, IPA-EA) and methanol (IPT-M, IPA-M) extracts from tuber and aerial parts, respectively, through behavioral and biochemical tests, and to assess their protective role in bisphenol A (BPA)-induced gonadotoxicity at a dose of 300 mg/kg in male Sprague Dawley rats. Phytochemical analysis was conducted both qualitatively and quantitatively, assessing total phenolic and flavonoid content (TPC and TFC) and using high-performance liquid chromatographic (HPLC-DAD) fingerprinting. Antioxidant profiling employed multimode in vitro assays, while biochemical validation included semen characteristics and levels of testosterone, follicle-stimulating hormone (FSH), luteinizing hormone (LH), and estradiol, while gonadoprotective ability was assessed using the comet assay and histomorphological examination of testes. Overall, qualitative analysis confirmed the presence of phenols, flavonoids, tannins, anthocyanins, saponins, coumarins, terpenoids, and betacyanins. Quantitative analysis revealed that the IPA-EA extract had the highest TPC (304.32±7.20 μg gallic acid equivalents/mg dry extract) and TFC (214.77±4.09 μg quercetin equivalents/mg dry extract). IPT-EA yielded maximum rutin (7.3±0.12 μg/mg dry extract) and myricetin (2.7±0.14 μg/mg dry extract), while IPA-EA and IPA-M extracts yielded the highest levels of caffeic acid (4.05±0.22 and 1.92±0.17 μg/mg dry extract, respectively) in HPLC-DAD analysis (Table 3).
The extracts were found to enhance sexual excitement, improve semen quality, and elevate levels of testosterone, FSH, LH, and estradiol, effectively mitigating the toxic effects of BPA. The levels of endogenous antioxidant enzymes (CAT, SOD, POD, and GSH) were restored, and nitric oxide (NO) abundance was minimized. Significant stimulation in sexual behavior, amelioration of toxicity symptoms, increased sperm production, enhanced viability, elevated levels of gonadal hormones, maintained endogenous enzyme activity, genoprotection, and improved testicular histology endorsed I. batatas as an effective aphrodisiac and gonadoprotective agent [86]. Apart from the above example, compounds like ellagic acid [88] and ferulic acid [89] have shown protective effects against testicular damage and oxidative stress caused by arsenic in mice, while taxifolin has protected fetal male rats from reproductive toxicity induced by di-n-butyl phthalate [90]. Treatment with the plant hormone 24-epibrassinolide [91] restored redox balance and enhanced health outcomes in ethanol-exposed zebrafish embryos, and the phytomolecules thymoquinone and curcumin have demonstrated beneficial effects in mitigating acrylamide-induced developmental issues in Drosophila by regulating ROS levels. Allicin derived from garlic [92] reversed lead-induced cognitive deficits in rats by managing oxidative stress [67]. These studies collectively underscore the potential of phytochemicals as safe and effective supplements for combating oxidative stress-related developmental and reproductive toxicity due to environmental pollutants. The liver, a vital organ, performs crucial functions such as xenobiotic metabolism, immune response facilitation, digestion, and vitamin storage. Hepatocytes, comprising 60–80% of liver cells, are primarily responsible for metabolic and detoxification activities. Oxidative stress is a pivotal pathological mechanism underlying liver damage, including conditions like alcoholic liver disease, steatosis, and cirrhosis. Within hepatocytes, ROS predominantly arise from cytochrome P450 enzymes in mitochondria and the endoplasmic reticulum. Environmental toxins induce hepatic injury through oxidative stress pathways, highlighting the importance of managing redox balance to mitigate liver toxicity. Recent research emphasizes the protective role of plant-derived compounds against chemical-induced liver damage. For instance, exposure to the above-mentioned BPA diminishes antioxidant enzyme activity while increasing lipid peroxidation, DNA damage, and markers of hepatotoxicity like AST, ALP, and ALT. In contrast, administration of Quercus dilatata Lindl. extract has shown potential in alleviating bisphenol A-induced liver injury [93]. Curcumin has been effective in ameliorating insulin resistance and oxidative stress induced by bisphenol A in HepG2 cells [59]. Ethanol consumption exacerbates liver damage by increasing ROS production during metabolism. Rats supplemented with Green Capsicum annum exhibited protective effects against ethanol-induced hepatotoxicity by enhancing the antioxidant defense system [94]. Purple potato (Solanum tuberosum) [95] extracts similarly protected against ethanol-induced liver damage by suppressing CYP2E1 (Figure 4b) expression. Additionally, a combination of curcumin and baicalin effectively ameliorated ethanol-induced liver damage by activating the Nrf2/HO-1 pathway [59]. Carbon tetrachloride (CCl4), a potent hepatotoxic agent [96,97,98], disrupts hepatic redox homeostasis by inducing CYP2E1. However, co-administration of Citrus aurantium ethyl acetate extract, Syzygium samarangense methanol extract, and Aerva javanica ethanolic extract effectively attenuated CCl4-induced hepatotoxicity by maintaining redox balance [99,100,101]. Coumarins such as umbelliferone and daphnetin protected against CCl4-induced liver damage by activating the Nrf2 pathway and enhancing cytoprotective antioxidant systems [102]. Commelina nudiflora supplementation also defended against CCl4-induced liver injury by modulating oxidative stress markers and inflammation, thereby preserving liver morphology [103]. Aluminum (Al) exposure increases ROS production in the liver, leading to oxidative stress-related liver injury. In this context, Nigella sativa oil [104,105,106], rich in phenolics, flavonoids, and tannins, has shown protective effects against aluminum chloride-induced liver injury in rats. Similarly, Monodora myristica and Terminalia arjuna extracts have mitigated oxidative stress and improved liver health in response to aluminum toxicity [59]. Cadmium (Cd), another toxic metal, induces oxidative stress in the liver, causing lipid peroxidation, mitochondrial dysfunction, and apoptosis. Supplementation with Vernonia amygdalina acetonic extract [107], Jessiaea nervosa aqueous extract [108], and Terminalia arjuna bark aqueous extract [109] attenuates Cd-induced liver injury by restoring antioxidant levels and reducing lipid peroxidation. Rosa persica supplementation enhances metallothionein content and reduces oxidative stress in Cd-exposed organisms, thereby improving liver function. Arsenic exposure induces oxidative stress and oxidative DNA damage in the liver, promoting hepatocarcinogenesis. In this frame, sulforaphane supplementation activates the Nrf2 pathway via PI3K/Akt signaling, exhibiting hepatoprotective effects against arsenic-induced liver injury [59]. insecticide
4.6. Phytochemicals in Renal Protection
Phytochemicals have emerged as promising preventive agents, enhancing antioxidant levels and supporting normal liver function in the face of toxic insults. Free radicals are critical in arsenic and cadmium-induced toxicity. Thymoquinone, found in Nigella sativa oil, counters As and Cd-related renal damage by restoring antioxidant enzymes like SOD, CAT, and GSH, curbing lipid peroxidation (LPO), and reducing apoptosis in renal tissue. Due to the lack of a specific excretory pathway, Cd accumulates in kidneys, particularly harming proximal tubules. Flavocoxid, a blend of natural plant-derived compounds rich in polyphenols, mitigates Cd-linked kidney lesions through its antioxidant properties. Ferulic acid, from curcumin, mitigates Cd renal damage in Wistar albino rats by inhibiting Cd accumulation, elevating antioxidant enzymes (TTH, TAC, SOD, CAT, GPx, TSH, and GSH), and controlling stress and inflammatory markers. The oxidative DNA damage monitored by the marker 8-OxodG, elevated in As-exposed C57BL/6 mice, was reduced with black raspberry. Beta vulgaris L. (beetroot), globally recognized for its minerals and vitamins, counters chlorpyrifos (CPF, Figure 5) nephrotoxicity by restoring antioxidants and limiting oxidative stress. Sodium dichromate and lithium carbonate increase renal markers (creatinine and blood urea nitrogen, BUN), LPO, reduce antioxidants, and cause DNA fragmentation [59]. Opuntia ficus indica [110,111,112,113] with its antioxidants, protects against renal damage from lithium carbonate and sodium dichromate [111,113]. Remarkably, the study of Ncigi et al. examined the effectiveness of Opuntia cladodes extract in mitigating liver damage caused by the organophosphorous insecticide chlorpyrifos in mice [112]. Liver damage was assessed by measuring liver weight and analysing various biochemical parameters, including alanine aminotransferase (ALT), aspartate aminotransferase (AST), alkaline phosphatase (PAL), lactate dehydrogenase, cholesterol, and albumin in serum using spectrophotometric methods. The experimental design spanned 48 hours and involved six different treatments with six mice each: (1) control, (2) 10 mg/kg (body weight, b.w) CPF, (3) 10 mg/kg (b.w) CPF with 100 mg/kg (b.w) cactus, (4) 150 mg/kg (b.w) CPF, (5) 150 mg/kg (b.w) CPF with 1.5 g/kg cactus, (6) 1.5 g/kg cactus. Both CPF and cactus were administered orally via gavage. The findings indicated that CPF significantly impacted all measured parameters. However, when CPF was administered with cactus extract, a recovery in all parameters was observed. Conversely, cactus alone did not alter the studied parameters. These results suggest that CPF is hepatotoxic and that Opuntia ficus indica stem extract offers protective effects against liver damage induced by this organophosphorous pesticide [112].
4.7. Neuroprotection and Environmental Toxins
Mercury triggers renal oxidative stress and acute failure. Juglans sinensis extract prevents mercury chloride damage in rabbits by bolstering antioxidants [114]. Eruca sativa seed extract counters Hg-induced renal toxicity [59,115]. Selenium-induced Wistar rat hepatic and renal injuries are resolved by curcumin; plant extracts and metabolites alleviate oxidative stress, inflammation, and renal dysfunction caused by environmental toxicants. Neurons are particularly susceptible to irreversible damage, profoundly affecting their structure and function, leading to the onset and progression of debilitating neurological disorders such as Alzheimer’s disease [116,117,118,119,120], Parkinson’s disease [121], and amyotrophic lateral sclerosis (ALS) [122,123]. These conditions pose significant global health challenges [124,125], ranking second only to cardiovascular diseases in terms of mortality, affecting countless individuals worldwide. The multifaceted nature of neurodegenerative diseases involves a complex interplay of genetic predispositions, lifestyle factors, and exposure to environmental agents. Key hallmarks of neurodegenerative diseases include cognitive impairment, dementia, motor dysfunction (such as ataxia), and speech difficulties. These symptoms manifest gradually over time and are exacerbated by increased oxidative stress, which plays a pivotal role in disease pathogenesis [126]. Neurons, being highly specialized cells with high oxygen consumption and relatively low levels of antioxidants, are particularly vulnerable to oxidative damage. This imbalance leads to impaired protein dynamics, aggregation of misfolded proteins, and ultimately, neuronal cell death. This imbalance leads to impaired protein dynamics, aggregation of misfolded proteins, and ultimately, neuronal cell death. Environmental chemicals can breach the protective blood-brain barrier, directly affecting brain health through various mechanisms. These include mitochondrial dysfunction, oxidative stress induction, disruption of neurotransmitter signaling, and epigenetic modifications, all of which contribute to the development and progression of neurological disorders. In response to these challenges, there is growing interest in exploring plant-based interventions as potential therapeutic strategies against neurotoxicity induced by environmental toxins. Plant-derived compounds have demonstrated promising neuroprotective effects by restoring redox homeostasis, enhancing antioxidant defenses, and mitigating oxidative damage to neural cells [59,127,128]. For instance, studies have highlighted the protective roles of various herbal extracts and phytochemicals such as kolaviron [129], Mentha spicata [130], Morus alba [131], and mangiferin [59,132]. These products have been shown to attenuate oxidative stress, reduce neuroinflammation, and preserve neuronal function in preclinical models. Thus, while oxidative stress remains a critical factor in the pathogenesis of various diseases, including neurodegenerative ones[122,125,126,127,133], plant-based interventions offer potential avenues for therapeutic development by targeting underlying mechanisms of toxicity and enhancing resilience against environmental insults (Table 4).
Continued research in this area holds promise for advancing novel treatments aimed at mitigating the burden of different socially-relevant disorders worldwide. Even natural substances intended to boost antioxidant levels should be used with caution, as excessive intake may pose risks; for example, research has indicated that high doses of beta-carotene can increase the risk of lung cancer, particularly in smokers or former smokers, despite its availability in multivitamins [103].
4.8. Variability in Plant Metabolite-Xenobiotic Interactions
Although limited research specifically addresses how interactions between plant metabolites and xenobiotics may vary across different plant species, environmental conditions, or types of xenobiotics, it is essential to critically analyse these factors for a comprehensive understanding of plant-xenobiotic interactions. The interaction between plant metabolites and xenobiotics encompasses various metabolic processes that modify xenobiotics to diminish their toxicity and facilitate their removal. To enhance their solubility and decrease toxicity, xenobiotics are often conjugated with more polar molecules, including sugars, amino acids, organic acids, or glutathione. Specifically, plants frequently incorporate sugar moieties into xenobiotics using enzymes like UDP-O-glucosyltransferases and UDP-N-glucosyltransferases, which results in the formation of glycoside derivatives [134]. These glycosylated compounds may then undergo further modifications, such as acylation with malonic acid via O- or N-malonyltransferases, which improves their stability and solubility. Additionally, glutathione S-transferases play a significant role in this interaction by conjugating xenobiotics with reduced glutathione. This process generates glutathione S-conjugates that contribute to detoxifying the xenobiotics, making them less harmful to the plant. Various isoforms of glutathione transferases possess different substrate specificities, leading to distinct metabolic outcomes based on the specific plant species and their enzyme profiles. In plants, the resulting conjugated xenobiotics are usually sequestered in vacuoles or associated with insoluble structures like lignin and polysaccharides, given that plants do not have an active excretory system. This sequestration helps reduce the mobility and potential toxicity of xenobiotics within the plant. The diversity in conjugated metabolites can vary widely among different plant species, illustrating the complexity of these interactions and the ways in which plants adapt to chemical stressors in their environment. Gaining insight into these interactions is essential for understanding the ecological consequences of xenobiotic exposure and how various plants manage chemical challenges [134]. In the study conducted by Kolb and Harms (2009), the authors focus on the metabolism of fluoranthene across various plant cell cultures, identifying significant differences in metabolic rates among the species examined. The findings indicate that specific plant species, such as Rosa sp. Paul’s Scarlet, exhibited remarkably high rates of fluoranthene metabolism compared to others. Additionally, the metabolic pathways activated in response to xenobiotics may vary with different chemical structures and functional groups. The study highlights the limited metabolic conversion of fluoranthene in comparison to other compounds like pentachlorophenol or 4-n-nonylphenol, suggesting that structural differences in these xenobiotics lead to varying enzymatic responses in plants [135]. The effectiveness of phytoremediation in removing or neutralizing xenobiotics from the environment depends heavily on various environmental conditions such as temperature, humidity, and sunlight, which can limit its application to specific areas [136]. Additionally, when xenobiotic concentrations are particularly high, they may become toxic to the plants themselves, affecting their capacity for remediation. Remarkably, the technique typically requires a large amount of land to accommodate the necessary plant growth for effective treatment [136]. Research has shown that different xenobiotic compounds can exhibit varying degrees of toxicity and metabolic complexity, affecting how plants respond to these stressors. Some xenobiotic compounds, such as acetaminophen, are more easily metabolized by plants. Acetaminophen, commonly found in agroecosystems due to the reuse of treated wastewater, can be quickly conjugated with molecules like glutathione, glucuronic acid, or sulphate, allowing plants to process it efficiently through established metabolic mechanisms [137]. On the other hand, heavy metals such as cadmium and lead are highly toxic and can generate reactive oxygen species that lead to oxidative stress in plants. Research shows that different xenobiotics like Cd and Pb can affect plants in varying ways, particularly in terms of toxicity and oxidative stress. In mulberry plants (Morus alba) exposed to Cd and Pb stress (100 and 200 μmol L⁻¹), both metals degraded chlorophyll and reduced photosynthesis, but Cd caused more severe damage overall. Cd stress led to higher production of ROS and greater oxidative damage than Pb, resulting in more pronounced reductions in photosynthetic activity. Pb, while still harmful, caused less disruption to electron transport and energy dissipation, making Cd the more challenging xenobiotic for the plant to handle [138]. Thlaspi caerulescens (alpine pennycress), a well-known hyperaccumulator of heavy metals, demonstrated a markedly different response compared to nonaccumulator species such as alfalfa (Medicago sativa), radish (Raphanus sativus), and lettuce (Lactuca sativa). Specifically, T. caerulescens showed superior resistance to the phytotoxic effects of Cd and Zn, and it was able to accumulate these metals at much higher concentrations within its tissues compared to the nonaccumulator plants. However, for Cu, T. caerulescens was less effective in accumulating it compared to alfalfa, indicating that its capacity to handle different types of heavy metals varies. This variation in metal accumulation abilities across species highlights that different plants possess distinct physiological and metabolic mechanisms to cope with xenobiotics, depending on their genetic traits and enzyme systems. These findings are supported by the significant differences in root elongation inhibition and metal translocation observed in the study, underscoring that species-specific traits greatly influence how plants deal with toxic compounds in the environment. Thus, this study confirms that plants vary significantly in their ability to handle different xenobiotics, with some species like T. caerulescens being particularly adept at accumulating certain metals like Cd and Zn, while struggling with others like Cu [139].
4.9. Differential Plant Responses to Xenobiotics
The xenobiotics can be organized into distinct categories based on their primary effects and the corresponding plant mechanisms involved in their amelioration. For neurotoxicity, examples include sodium azide, methyl mercury, and cadmium, which induce oxidative stress, leading to neuronal cell death and cognitive dysfunction. Phytochemicals such as kolaviron and mangiferin have shown protective effects through antioxidant activity and anti-inflammatory properties. In terms of hepatotoxicity, bisphenol A, carbon tetrachloride, and arsenic cause increased lipid peroxidation and decreased antioxidant enzyme levels, resulting in liver damage. Phytochemicals like curcumin, thymoquinone, and those extracted from Citrus aurantium exhibit hepatoprotective effects by enhancing antioxidant defenses and reducing oxidative stress markers. For nephrotoxicity, arsenic, cadmium, and lead disrupt renal function by inducing oxidative stress and inflammation. Protective compounds such as quercetin, taxifolin, and catechin help restore antioxidant balance and mitigate kidney damage. Regarding developmental and reproductive toxicity, xenobiotics such as toluene, ethanol, and nicotine can lead to developmental delays and reproductive issues via oxidative stress and hormonal disruption. Phytochemicals like those from Boerhavia diffusa and Mentha spicata help counteract these effects through their antioxidant properties. Finally, in the context of pulmonary toxicity, agents like paraquat and benzo(a)pyrene increase oxidative stress in lung tissues. Protective phytochemicals such as quercetin and procyanidin B2 have shown efficacy in reducing ROS levels and inflammation [59]. Understanding the impact of different dosages is crucial in evaluating plant responses, as the effects can vary significantly based on the concentration of the xenobiotic used. Although data on the relationship between varying xenobiotic doses and plant responses are limited, the study by Yang et al. (2016) provides valuable insights [140]. A bioassay was conducted to evaluate the effects of cycloxaprid on rice seedlings and third instar insects. To prepare cycloxaprid, it was initially dissolved in dimethylformamide (DMF) and then diluted in distilled water with 0.05% Triton-100 to create various concentrations. The bioassay determined the low dose (LC15) and high dose (LC85) for treatment, with the calculated LC15 being 0.44 μg ml⁻¹ (95% confidence interval: 0.31–0.61) and the LC85 measuring significantly higher at 19.86 μg ml⁻¹ (95% confidence interval: 13.63–28.95). In the experimental setup, four rice seedlings were immersed in each concentration for 15 seconds before being placed in a controlled environment, with mortality assessments conducted after 48 hours. Under the high-dose condition, 4,807 unigenes were differentially regulated, showing 1,743 upregulated and 3,063 downregulated. Conversely, the low-dose treatment affected 2,020 unigenes, with 918 upregulated and 1,122 downregulated. This variation in gene expression indicates that the insects activated different stress response pathways based on the levels of cycloxaprid exposure. Notably, 306 unigenes exhibited contrasting regulation patterns in response to high and low doses. The research also examined several gene families related to detoxification, including cytochrome P450s, glutathione S-transferases, and ATP-binding cassette transporters. In the case of P450s, ten unigenes showed increased expression under the high-dose treatment, with six belonging to the CYP6 family, highlighting their significant role in detoxification. A specific delta-GST unigene demonstrated a remarkable 66.64-fold increase in expression under low-dose treatment, in contrast to a modest 2.41-fold increase in a microsomal GST under high-dose conditions, indicating differing regulatory responses to the doses. Additionally, one acetylcholinesterase unigene was notably upregulated (9.46-fold) in the high-dose treatment, suggesting its potential involvement in the insect’s resistance mechanisms against cycloxaprid. Heat shock proteins (Hsps) also showed differential expression; the levels of two Hsp70 unigenes and two small Hsp unigenes rose over two-fold in the high-dose treatment relative to the control, while they decreased in response to the low dose. Overall, these results reveal that varying doses of cycloxaprid trigger distinct responses in both plants and insects, emphasizing the intricate regulatory mechanisms governing detoxification and stress responses in Sogatella furcifera [140]. The complexity of plant responses is further illustrated by research involving alfalfa plants (Medicago sativa) inoculated with Sinorhizobium meliloti, which demonstrated that cadmium and fluoranthene notably impacted nodulation and gene expression [140]. In this investigation, alfalfa was grown for 14 days in a mineral medium containing different concentrations of these xenobiotics. Key parameters measured included the number of root nodules, fresh weight of shoots and roots, and gene expression assessed through mRNA differential display techniques. The results showed that both cadmium and fluoranthene significantly decreased the number of root nodules before any visible signs of plant damage occurred. The effective concentration (EC50) for cadmium affecting nodulation was determined to be 5.8 µM, while fluoranthene exhibited an EC50 of 2.5 µg cm². Notably, nodulation was found to be ten times more sensitive to these pollutants compared to shoot and root biomass; for instance, the EC50 for root fresh weight due to cadmium was over 20 µM, and more than 35 µg cm² for fluoranthene. Regarding gene expression, the study identified 37 differentially displayed transcripts in response to the xenobiotics. One transcript, DDMsl, was confirmed to be down-regulated in the presence of both cadmium and fluoranthene. In control plants inoculated with rhizobia, DDMsl expression was 2.5–3 times higher than in non-inoculated plants. However, this expression was significantly diminished by fluoranthene and completely suppressed by cadmium, with an EC50 for DDMsl expression of 5.9 µM. Sequence analysis of DDMsl indicated a 50% similarity to a copper-transporting ATPase from Arabidopsis thaliana. Another transcript, DDMs2, also showed down-regulation in plants treated with xenobiotics, revealing a 50% decrease in expression and demonstrating a strong homology (82% similarity) to the cytoplasmic 60S ribosomal protein L18 from Arabidopsis thaliana [140]. These findings suggest that nodulation is more susceptible to environmental stressors than overall plant growth, indicating that the initial stages of root nodule formation may be especially vulnerable to contaminants, potentially compromising nitrogen fixation and plant health. The differential expression of genes like DDMsl and DDMs2 in response to xenobiotics implies that these transcripts may be vital for plant stress responses and adaptation to heavy metals and organic pollutants. Gaining insights into these molecular responses can aid in formulating strategies to enhance plant resilience against environmental contaminants. The calculated EC10, EC50, and EC90 values are crucial for understanding how varying concentrations of pollutants affect plant physiology, which is essential for environmental risk assessments and developing regulations concerning heavy metal and hydrocarbon pollution in agricultural soils. Overall, this study underscores the considerable negative effects of xenobiotics on alfalfa plants and highlights how different concentrations can distinctly impact nodulation and gene expression [140].
5. Conclusions
Human activities have significantly damaged the environment, resulting in widespread exposure to harmful xenobiotic chemicals. Both short-term and long-term contact with these substances adversely affects living organisms, including humans, leading to negative impacts on health and well-being. Research over the past century has primarily focused on how these chemicals influence various organisms, with a recent shift towards a more detailed understanding of toxicity aimed at reducing harmful effects. The clinical assessment of phytocompounds that ameliorate xenobiotic-induced diseases, particularly drug-induced liver toxicity, highlights the potential and efficacy of herbal antioxidants. Many natural products, especially those used in traditional medicine in regions like China and India, have garnered significant interest for their therapeutic properties against liver disorders. The compounds rich in triterpenes, flavonoids, and polyphenols are increasingly recognized for their hepatoprotective effects in experimental liver injury models. These natural products are believed to provide protection primarily through their antioxidant properties, which help eliminate free radicals and mitigate reactive oxygen species damage to cellular structures. The clinical evaluation of specific compounds such as silymarin, resveratrol, curcumin, and ginkgo biloba demonstrates their mechanisms of action, including cell regeneration, cytoprotection, and modulation of various cytochrome P450 isoforms. For instance, silymarin from milk thistle has been shown to protect against drug-induced hepatotoxicity in various animal models and exhibits antioxidant and cell-regenerative properties, with clinical trials suggesting that silymarin can significantly reduce liver damage in patients with chronic liver diseases, with minimal side effects reported. Resveratrol, a polyphenol found in grapes, has been linked to chemoprevention and protection against liver damage induced by substances like acetaminophen, while curcumin, derived from turmeric, exhibits potent antioxidant and anti-inflammatory activities, contributing to its protective effects against a range of hepatotoxins, including heavy metals and alcohol [30]. Despite the promising therapeutic potential of these herbal compounds, their clinical use is fraught with limitations and challenges. One significant challenge is the variability in phytochemical content among different plant sources, which can lead to inconsistencies in efficacy and safety. Factors such as geographical location, cultivation methods, and harvesting times can significantly influence the concentration of active ingredients. This variability complicates the standardization of herbal products and can affect the reproducibility of clinical outcomes. Additionally, the potential side effects of herbal medications are often underreported, and some compounds may interact negatively with conventional pharmaceuticals. This raises concerns about the safety profiles of these natural products, especially in patients with pre-existing conditions or those taking multiple medications. The limitations of animal studies further complicate the situation, as findings from these models do not always accurately predict human outcomes. Factors such as differences in metabolism, absorption, and individual biological variability can result in discrepancies between animal and human responses to herbal treatments. In summary, while herbal antioxidants present a promising avenue for ameliorating xenobiotic-induced diseases, particularly drug-induced liver toxicity, their clinical assessment reveals significant limitations. The variability in phytochemical content, potential side effects, and the limitations of animal studies in predicting human outcomes pose considerable challenges. A thorough understanding of these factors is crucial for the safe and effective incorporation of herbal medicines into clinical practice.
Perspectives
There is a growing interest in developing plant-based therapies due to the natural antioxidant properties of plants. Plant extracts and bioactive compounds are increasingly utilized in biomedicine to mitigate the harmful effects of chemicals, specifically targeting oxidative stress as a crucial therapeutic pathway. We believe future research should prioritize establishing consistent concentrations of active compounds and determining safe dosages, as this will minimize variability in therapeutic effects and prevent potential toxicity. Thorough assessments of safety and toxicity are essential to identify adverse effects associated with long-term use or high doses of these supplements, given that excessive intake can pose risks. Additionally, investigating potential interactions between plant-derived supplements and prescription medications is crucial for ensuring safe use and maintaining treatment efficacy. Ongoing research should also aim to elucidate the mechanisms of action of plant-derived ingredients, effective dosages, and long-term outcomes to build a robust evidence base supporting their efficacy and safety. Considering the widespread exposure to various toxic substances in industrial and agricultural settings, we feel that exploring the combined effects of these chemical mixtures on human health is vital. Investigating plant-based interventions against individual toxicants alone may not capture the full scope of their impacts, underscoring the need to isolate and evaluate specific bioactive components for their effectiveness as pharmaceutical agents. In our view, addressing these research gaps can inspire new directions in the field, enhancing our understanding and utilization of plant-derived antioxidants to combat chemical toxicity effectively. This comprehensive approach will pave the way for the development of safe, effective health supplements that leverage the protective effects of nature.
Author Contributions
All authors have contributed equally to this work. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No data available.
Conflicts of Interest
The authors declare no conflicts of interest.
List of Abbreviations
8-OxodG 8-hydroxy-2’-deoxyguanosine
ALT Alanine transaminase
ALP Alkaline phosphatase
AST Aspartate aminotransferase
ATPase Adenosine Triphosphatase
BDE47 2,2’,4,4’-Tetrabromodiphenyl Ether
BPA Bisphenol A
BUN Blood Urea Nitrogen
CAT Catalase
COX Cyclooxygenase
CP Calliandra portoricensis
CPF Chlorpyrifos
CYP Cytochrome P450
DMF Dimethylformamide
EC10 Effective Concentration 10%
EC50 Effective Concentration 50%
EC90 Effective Concentration 90%
GPx Glutathione Peroxidase
GSH Glutathione
GST Glutathione S-transferase
HPTLC High-Performance Thin Layer Chromatography
Hsp Heat Shock Protein
ISL Isoliquiritigenin
LC15 Lethal Concentration 15%
LC50 Lethal Concentration 50%
LC85 Lethal Concentration 85%
LOX Lipoxygenase
LPO Lipid Peroxidation
MDA Malondialdehyde
MMP Mitochondrial membrane potential
mROS Mitochondrial reactive oxygen species
mRNA Messenger RNA
Nfe2l2 Nuclear factor erythroid 2-related factor 2
NO Nitric oxide
NOX NADPH oxidase
Nrf2 Nuclear Factor Erythroid 2-Related Factor 2
PDGF Platelet-derived growth factor
PI3K Phosphoinositide-3-kinase
POD Peroxidase
RNS Reactive nitrogen species
ROS Reactive oxygen species
SOD Superoxide dismutase
StAR Steroidogenic acute regulatory protein
TAC Total Antioxidant Capacity
TFAM Mitochondrial transcription factor A
TTH Total Thiol Content
TSH Thyroid-Stimulating Hormone
XO Xanthine oxidase
References
- Malchi, T.; Eyal, S.; Czosnek, H.; Shenker, M.; Chefetz, B. Plant pharmacology: Insights into in-planta kinetic and dynamic processes of xenobiotics. Critical Reviews in Environmental Science and Technology 2021, 52(19), 3525–3546. [Google Scholar] [CrossRef]
- Kaushik, B.; Sharma, J.; Yadav, K.; Kumar, P.; Shourie, A. Phytochemical Properties and Pharmacological Role of Plants: Secondary Metabolites. Biosciences Biotechnology Research Asia 2021, 18(1), 23–35. [Google Scholar] [CrossRef]
- Fik-Jaskółka, M.; Mittova, V.; Motsonelidze, C.; Vakhania, M.; Vicidomini, C.; Roviello, G.N. Antimicrobial Metabolites of Caucasian Medicinal Plants as Alternatives to Antibiotics. Antibiotics 2024, 13(6), 487. [Google Scholar] [CrossRef] [PubMed]
- Roviello, V.; Gilhen-Baker, M.; Vicidomini, C.; Roviello, G.N. The Healing Power of Clean Rivers: In Silico Evaluation of the Antipsoriatic Potential of Apiin and Hyperoside Plant Metabolites Contained in River Waters. International Journal of Environmental Research and Public Health 2022, 19(5), 2502. [Google Scholar] [CrossRef] [PubMed]
- Roviello, V.; Gilhen-Baker, M.; Roviello, G.N.; Lichtfouse, E. River therapy. Environmental Chemistry Letters 2022, 20(5), 2729–2734. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Prasad, S.M.; Singh, R.P. 2016.
- Sauvêtre, A.; Eichhorn, P.; Pérez, S. Sauvêtre, A.; Eichhorn, P.; Pérez, S. Metabolism of Pharmaceuticals in Plants and Their Associated Microbiota. 2020, 103, 221–264.
- Wu, J.-C.; Lai, C.-S.; Tsai, M.-L.; Ho, C.-T.; Wang, Y.-J.; Pan, M.-H. Chemopreventive effect of natural dietary compounds on xenobiotic-induced toxicity. Journal of Food and Drug Analysis 2017, 25(1), 176–186. [Google Scholar] [CrossRef]
- Mandal, M.; Sarkar, M.; Khan, A.; Biswas, M.; Masi, A.; Rakwal, R.; Agrawal, G.K.; Srivastava, A.; Sarkar, A. Reactive Oxygen Species (ROS) and Reactive Nitrogen Species (RNS) in plants– maintenance of structural individuality and functional blend. Advances in Redox Research 2022, 5, 100039. [Google Scholar] [CrossRef]
- Di Meo, S.; Venditti, P. Evolution of the Knowledge of Free Radicals and Other Oxidants. Oxidative Medicine and Cellular Longevity 2020, 2020, 1–32. [Google Scholar] [CrossRef]
- Goncharov, N.; Avdonin, P.; Nadeev, A.; Zharkikh, I.; Jenkins, R. Reactive Oxygen Species in Pathogenesis of Atherosclerosis. Current Pharmaceutical Design 2015, 21(9), 1134–1146. [Google Scholar] [CrossRef]
- Chen, H.; Ma, A.; Qi, S. Antioxidant Therapy for Prevention of Inflammation, Ischemic Reperfusion Injuries and Allograft Rejection. Cardiovascular & Hematological Agents in Medicinal Chemistry 2008, 6(1), 20–43. [Google Scholar]
- Bedard, K.; Krause, K.-H. The NOX Family of ROS-Generating NADPH Oxidases: Physiology and Pathophysiology. Physiological Reviews 2007, 87(1), 245–313. [Google Scholar] [CrossRef] [PubMed]
- Giulia Battelli, M.; Polito, L.; Bortolotti, M.; Bolognesi, A. Xanthine Oxidoreductase in Drug Metabolism: Beyond a Role as a Detoxifying Enzyme. Current Medicinal Chemistry 2016, 23(35), 4027–4036. [Google Scholar] [CrossRef] [PubMed]
- Veith, A.; Moorthy, B. Role of cytochrome P450s in the generation and metabolism of reactive oxygen species. Current Opinion in Toxicology 2018, 7, 44–51. [Google Scholar] [CrossRef]
- Rosen, G.M.; Tsai, P.; Pou, S. Mechanism of Free-Radical Generation by Nitric Oxide Synthase. Chemical Reviews 2002, 102(4), 1191–1200. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.-J.; Seo, J.-M.; Kim, J.-H. Bioactive Lipoxygenase Metabolites Stimulation of NADPH Oxidases and Reactive Oxygen Species. Molecules and Cells 2011, 32(1), 1–6. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.; Kumar, P. Environmental threats posed by xenobiotics. 2024, 183–201.
- Wang, X.; Sial, M.U.; Bashir, M.A.; Bilal, M.; Raza, Q.-U.-A.; Ali Raza, H.M.; Rehim, A.; Geng, Y. Pesticides Xenobiotics in Soil Ecosystem and Their Remediation Approaches. Sustainability 2022, 14(6), 3353. [Google Scholar] [CrossRef]
- Singh, A.; Mehta, S.; Yadav, S.; Nagar, G.; Ghosh, R.; Roy, A.; Chakraborty, A.; Singh, I.K. How to Cope with the Challenges of Environmental Stresses in the Era of Global Climate Change: An Update on ROS Stave off in Plants. International Journal of Molecular Sciences 2022, 23(4), 1995. [Google Scholar] [CrossRef]
- Labudda, M.; Dziurka, K.; Fidler, J.; Gietler, M.; Rybarczyk-Płońska, A.; Nykiel, M.; Prabucka, B.; Morkunas, I.; Muszyńska, E. The Alleviation of Metal Stress Nuisance for Plants—A Review of Promising Solutions in the Face of Environmental Challenges. Plants 2022, 11(19), 2544. [Google Scholar] [CrossRef]
- Tiwari, S.; Tiwari, S.; Singh, M.; Singh, A.; Prasad, S.M. Generation Mechanisms of Reactive Oxygen Species in the Plant Cell. 2017, 1–22.
- Gill, S.S.; Peter Singh, L.; Gill, R.; Tuteja, N. Generation and Scavenging of Reactive Oxygen Species in Plants under Stress. 2012, 49–70.
- Hasanuzzaman, M.; Nahar, K.; Gill, S.S.; Fujita, M. Drought Stress Responses in Plants, Oxidative Stress, and Antioxidant Defense. 2013, 209–250.
- Marwicka, J.; Zięba, A. Antioxidants as a defence against reactive oxygen species. Aesthetic Cosmetology and Medicine 2021, 10(6), 271–276. [Google Scholar] [CrossRef]
- Fan, S.; Cong, Z. Emerging Strategies for Modifying Cytochrome P450 Monooxygenases into Peroxizymes. Accounts of Chemical Research 2024. [Google Scholar] [CrossRef]
- Döring, B.; Petzinger, E. Phase 0 and phase III transport in various organs: Combined concept of phases in xenobiotic transport and metabolism. Drug Metabolism Reviews 2014, 46(3), 261–282. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, P.; Biswas, A.; Dey, S.; Bhattacharjee, T.; Chakrabarty, S. Cytochrome P450 Gene Families: Role in Plant Secondary Metabolites Production and Plant Defense. Journal of Xenobiotics 2023, 13(3), 402–423. [Google Scholar] [CrossRef]
- Pirtskhalava, M.; Mittova, V.; Tsetskhladze, Z.R.; Palumbo, R.; Pastore, R.; Roviello, G.N. Georgian Medicinal Plants as Rich Natural Sources of Antioxidant Derivatives: A Review on the Current Knowledge and Future Perspectives. Current Medicinal Chemistry 2024, 31(28), 4407–4424. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Cho, W.C.; Upadhyay, G. Drug-Induced Liver Toxicity and Prevention by Herbal Antioxidants: An Overview. Frontiers in Physiology 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Duarte, A. Health Alternatives. Morton Grove, IL: Megasystems 1995.
- Markham, K.R.; Mitchell, K.A. The mis-identification of the major antioxidant flavonoids in young barley (Hordeum vulgare) leaves. Zeitschrift für Naturforschung C 2003, 58(1–2), 53–56. [Google Scholar] [CrossRef] [PubMed]
- Ross, A.C.; Caballero, B.; Cousins, R.J.; Tucker, K.L. Modern nutrition in health and disease. Jones & Bartlett Learning: 2020.
- Bol’shakova, I.; Lozovskaia, E.; Sapezhinskiĭ, I. Antioxidant properties of a series of extracts from medicinal plants. Biofizika 1997, 42(2), 480–483. [Google Scholar]
- Marderosion, A. The Review of Natural Products, Facts and Comparisons. St. Louis, MI, USA 2001, 630–632.s.
- Yu, C.; Kim, S.; Lim, J.; Kim, M.; Chung, I. Intraspecific relationship analysis by DNA markers and in vitro cytotoxic and antioxidant activity in Eleutherococcus senticosus. Toxicology in vitro 2003, 17(2), 229–236. [Google Scholar] [CrossRef]
- Halvorsen, B.L.; Holte, K.; Myhrstad, M.C.; Barikmo, I.; Hvattum, E.; Remberg, S.F.; Wold, A.-B.; Haffner, K.; Baugerød, H.; Andersen, L.F. A systematic screening of total antioxidants in dietary plants. The Journal of nutrition 2002, 132(3), 461–471. [Google Scholar] [CrossRef]
- Kim, D.S.; Kim, D.-S.; Oppel, M.N. Shogaols from Zingiber officinale protect IMR32 human neuroblastoma and normal human umbilical vein endothelial cells from β-amyloid (25–35) insult. Planta medica 2002, 68(04), 375–376. [Google Scholar] [CrossRef]
- Chen, J.-W.; Chen, Y.-H.; Lin, F.-Y.; Chen, Y.-L.; Lin, S.-J. Ginkgo biloba extract inhibits tumor necrosis factor-α–induced reactive oxygen species generation, transcription factor activation, and cell adhesion molecule expression in human aortic endothelial cells. Arteriosclerosis, thrombosis, and vascular biology 2003, 23(9), 1559–1566. [Google Scholar] [CrossRef]
- DeFeudis, F.V.; Papadopoulos, V.; Drieu, K. Ginkgo biloba extracts and cancer: A research area in its infancy. Fundamental & clinical pharmacology 2003, 17((4), 405–417. [Google Scholar]
- Guerra, M.; Speroni, E.; Broccoli, M.; Cangini, M.; Pasini, P.; Minghetti, A.; Crespi-Perellino, N.; Mirasoli, M.; Cantelli-Forti, G.; Paolini, M. Comparison between Chinese medical herb Pueraria lobata crude extract and its main isoflavone puerarin: Antioxidant properties and effects on rat liver CYP-catalysed drug metabolism. Life Sciences 2000, 67(24), 2997–3006. [Google Scholar] [CrossRef] [PubMed]
- Allen, S.J.; Gan, Q.; Matthews, R.; Johnson, P.A. Comparison of optimised isotherm models for basic dye adsorption by kudzu. Bioresource technology 2003, 88(2), 143–152. [Google Scholar] [CrossRef] [PubMed]
- Rezvani, A.H.; Overstreet, D.H.; Perfumi, M.; Massi, M. Plant derivatives in the treatment of alcohol dependency. Pharmacology Biochemistry and Behavior 2003, 75(3), 593–606. [Google Scholar] [CrossRef] [PubMed]
- Ben-Amotz, A.; Yatziv, S.; Sela, M.; Greenberg, S.; Rachmilevich, B.; Shwarzman, M.; Weshler, Z. e. Effect of natural b-carotene supplementation in children exposed to radiation from the Chernobyl accident. Radiation and Environmental Biophysics 1998, 37, 187–193. [Google Scholar] [CrossRef]
- Shimizu, I. Antifibrogenic therapies in chronic HCV infection. Current Drug Targets-Infectious Disorders 2001, 1, 227–240. [Google Scholar] [CrossRef]
- Kvasnička, F.; Bıba, B.; Ševčı́k, R.; Voldřich, M.; Kratka, J. Analysis of the active components of silymarin. Journal of Chromatography A 2003, 990(1–2), 239–245. [Google Scholar] [CrossRef]
- Upadhyay, G.; Kumar, A.; Singh, M.P. Effect of silymarin on pyrogallol-and rifampicin-induced hepatotoxicity in mouse. European journal of pharmacology 2007, 565(1–3), 190–201. [Google Scholar] [CrossRef]
- Gruenwald, J.; Brendler, T.; Jaenicke, C. PDR for herbal medicines. Thomson, Reuters: 2007.
- Lu, H.; Liu, G.-T. Anti-oxidant activity of dibenzocyclooctene lignans isolated from Schisandraceae. Planta Medica 1992, 58(04), 311–313. [Google Scholar] [CrossRef]
- Ekweogu, C.N.; Ude, V.C.; Nwankpa, P.; Emmanuel, O.; Ugbogu, E.A. Ameliorative effect of aqueous leaf extract of Solanum aethiopicum on phenylhydrazine-induced anaemia and toxicity in rats. Toxicological Research 2019, 36(3), 227–238. [Google Scholar] [CrossRef]
- Adelakun, S.A.; Ukwenya, V.O.; Ojewale, A.O.; Aniah, J.A.; Kolawole, B.P. Excessive exposure to sodium fluoride impaired spermatogenesis, induced hormonal and biochemical imbalance and testicular atrophy: Ameliorating potential of bioactive component of Solanum aethiopicum supplementation. Phytomedicine Plus 2023, 3(3), 100458. [Google Scholar] [CrossRef]
- Gawlik-Dziki, U. Modification of enzymatic and non-enzymatic in vitro oxidative defence system by bioaccessible phytonutrients of selected spices. LWT - Food Science and Technology 2014, 57(1), 434–441. [Google Scholar] [CrossRef]
- Piedrafita, G.; Keller, M.; Ralser, M. The Impact of Non-Enzymatic Reactions and Enzyme Promiscuity on Cellular Metabolism during (Oxidative) Stress Conditions. Biomolecules 2015, 5(3), 2101–2122. [Google Scholar] [CrossRef] [PubMed]
- Andre, C.; Larondelle, Y.; Evers, D. Dietary Antioxidants and Oxidative Stress from a Human and Plant Perspective: A Review. Current Nutrition & Food Science 2010, 6(1), 2–12. [Google Scholar]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS Sources in Physiological and Pathological Conditions. Oxidative Medicine and Cellular Longevity 2016, 2016, 1–44. [Google Scholar] [CrossRef]
- Zahra, K.F.; Lefter, R.; Ali, A.; Abdellah, E.-C.; Trus, C.; Ciobica, A.; Timofte, D.; Szewczyk-Golec, K. The Involvement of the Oxidative Stress Status in Cancer Pathology: A Double View on the Role of the Antioxidants. Oxidative Medicine and Cellular Longevity 2021, 2021, 1–25. [Google Scholar] [CrossRef]
- Cao, G.; Xuan, X.; Hu, J.; Zhang, R.; Jin, H.; Dong, H. How vascular smooth muscle cell phenotype switching contributes to vascular disease. Cell Communication and Signaling 2022, 20(1). [Google Scholar] [CrossRef]
- Safaroghli-Azar, A.; Sanaei, M.-J.; Pourbagheri-Sigaroodi, A.; Bashash, D. Phosphoinositide 3-kinase (PI3K) classes: From cell signaling to endocytic recycling and autophagy. European Journal of Pharmacology 2023, 953, 175827. [Google Scholar] [CrossRef]
- Dwivedi, S.; Kushalan, S.; Paithankar, J.G.; D’Souza, L.C.; Hegde, S.; Sharma, A. Environmental toxicants, oxidative stress and health adversities: Interventions of phytochemicals. Journal of Pharmacy and Pharmacology 2022, 74(4), 516–536. [Google Scholar] [CrossRef]
- Gusti, A.M.T.; Qusti, S.Y.; Alshammari, E.M.; Toraih, E.A.; Fawzy, M.S. Antioxidants-Related Superoxide Dismutase (SOD), Catalase (CAT), Glutathione Peroxidase (GPX), Glutathione-S-Transferase (GST), and Nitric Oxide Synthase (NOS) Gene Variants Analysis in an Obese Population: A Preliminary Case-Control Study. Antioxidants 2021, 10(4), 595. [Google Scholar] [CrossRef]
- Carmo de Carvalho e Martins, M.d.; Martins; da Silva Santos Oliveira, A.S.; da Silva, L.A.A.; Primo, M.G.S.; de Carvalho Lira, V.B. Biological Indicators of Oxidative Stress [Malondialdehyde, Catalase, Glutathione Peroxidase, and Superoxide Dismutase] and Their Application in Nutrition. 2022, 1–25.
- Pagano, J.S.; Dworzański, J.; Strycharz-Dudziak, M.; Kliszczewska, E.; Kiełczykowska, M.; Dworzańska, A.; Drop, B.; Polz-Dacewicz, M. Glutathione peroxidase (GPx) and superoxide dismutase (SOD) activity in patients with diabetes mellitus type 2 infected with Epstein-Barr virus. PLoS ONE 2020, 15(3), e0230374. [Google Scholar]
- Zhang, S.; Sun, M.; Hedtke, T.; Deshmukh, A.; Zhou, X.; Weon, S.; Elimelech, M.; Kim, J.-H. Mechanism of Heterogeneous Fenton Reaction Kinetics Enhancement under Nanoscale Spatial Confinement. Environmental Science & Technology 2020, 54(17), 10868–10875. [Google Scholar]
- Chen, S.; Zhu, M.; Guo, X.; Yang, B.; Zhuo, R. Coupling of Fenton reaction and white rot fungi for the degradation of organic pollutants. Ecotoxicology and Environmental Safety 2023, 254, 114697. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Zhang, X.; Liu, M.; Cai, B.; He, N.; Wang, Z. Fenton reaction-based nanomedicine in cancer chemodynamic and synergistic therapy. Applied Materials Today 2020, 21, 100864. [Google Scholar] [CrossRef]
- Vedi, M.; Rasool, M.; Sabina, E.P. Protective effect of administration of Withania somifera against bromobenzene induced nephrotoxicity and mitochondrial oxidative stress in rats. Renal failure 2014, 36(7), 1095–1103. [Google Scholar] [CrossRef]
- Cai, S.; Liu, J.; Shi, X.; Hu, S.; Zhao, L. Allicin alleviated learning and memory deficits caused by lead exposure at developmental stage. Life sciences 2019, 231, 116532. [Google Scholar] [CrossRef]
- Li, Z.; Yu, Y.; Li, Y.; Ma, F.; Fang, Y.; Ni, C.; Wu, K.; Pan, P.; Ge, R.-S. Taxifolin attenuates the developmental testicular toxicity induced by di-n-butyl phthalate in fetal male rats. Food and Chemical Toxicology 2020, 142, 111482. [Google Scholar] [CrossRef]
- Fernandes, P.; Monteiro, S.M.; Venâncio, C.; Félix, L. 24-Epibrassinolide protects against ethanol-induced behavioural teratogenesis in zebrafish embryo. Chemico-Biological Interactions 2020, 328, 109193. [Google Scholar] [CrossRef]
- Faddah, L.M.; Baky, N.A.A.; Al-Rasheed, N.M.; Al-Rasheed, N.M.; Fatani, A.J.; Atteya, M. Role of quercetin and arginine in ameliorating nano zinc oxide-induced nephrotoxicity in rats. BMC complementary and alternative medicine 2012, 12, 1–14. [Google Scholar] [CrossRef]
- Tu, P.; Xue, J.; Bian, X.; Chi, L.; Gao, B.; Leng, J.; Ru, H.; Knobloch, T.J.; Weghorst, C.M.; Lu, K. Dietary administration of black raspberries modulates arsenic biotransformation and reduces urinary 8-oxo-2′-deoxyguanosine in mice. Toxicology and Applied Pharmacology 2019, 377, 114633. [Google Scholar] [CrossRef]
- Sener, U.; Uygur, R.; Aktas, C.; Uygur, E.; Erboga, M.; Balkas, G.; Caglar, V.; Kumral, B.; Gurel, A.; Erdogan, H. Protective effects of thymoquinone against apoptosis and oxidative stress by arsenic in rat kidney. Renal Failure 2016, 38(1), 117–123. [Google Scholar] [CrossRef] [PubMed]
- Guvvala, P.R.; Ravindra, J.P.; Selvaraju, S.; Arangasamy, A.; Venkata, K.M. Ellagic and ferulic acids protect arsenic-induced male reproductive toxicity via regulating Nfe2l2, Ppargc1a and StAR expressions in testis. Toxicology 2019, 413, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Senthilkumar, S.; Raveendran, R.; Madhusoodanan, S.; Sundar, M.; Shankar, S.S.; Sharma, S.; Sundararajan, V.; Dan, P.; Mohideen, S.S. Developmental and behavioural toxicity induced by acrylamide exposure and amelioration using phytochemicals in Drosophila melanogaster. Journal of hazardous materials 2020, 394, 122533. [Google Scholar] [CrossRef] [PubMed]
- Crown, O.; Ogundele, O.; Akinmoladun, A.; Famusiwa, C.; Josiah, S.; Olaleye, M.; Akindahunsi, A. Effects of catechin, quercetin and taxifolin on redox parameters and metabolites linked with renal health in rotenone-toxified rats. Nigerian Journal of Physiological Sciences 2019, 34(1), 1–10. [Google Scholar] [PubMed]
- Erboga, M.; Kanter, M.; Aktas, C.; Sener, U.; Fidanol Erboga, Z.; Bozdemir Donmez, Y.; Gurel, A. Thymoquinone ameliorates cadmium-induced nephrotoxicity, apoptosis, and oxidative stress in rats is based on its anti-apoptotic and anti-oxidant properties. Biological trace element research 2016, 170, 165–172. [Google Scholar] [CrossRef]
- Micali, A.; Pallio, G.; Irrera, N.; Marini, H.; Trichilo, V.; Puzzolo, D.; Pisani, A.; Malta, C.; Santoro, G.; Laurà, R. Flavocoxid, a Natural Antioxidant, Protects Mouse Kidney from Cadmium-Induced Toxicity. Oxidative medicine and cellular longevity 2018, 2018(1), 9162946. [Google Scholar] [CrossRef]
- Saad, A.B.; Rjeibi, I.; Brahmi, N.; Elaloui, E.; Zouari, N. Nicotine-induced oxidative stress, testis injury, AChE inhibition and brain damage alleviated by Mentha spicata. Inflammopharmacology 2020, 28, 939–948. [Google Scholar] [CrossRef]
- Han, Y.; Ishibashi, S.; Iglesias-Gonzalez, J.; Chen, Y.; Love, N.R.; Amaya, E. Ca2+-Induced Mitochondrial ROS Regulate the Early Embryonic Cell Cycle. Cell Reports 2018, 22(1), 218–231. [Google Scholar] [CrossRef]
- Hunter, M.V.; Willoughby, P.M.; Bruce, A.E.E.; Fernandez-Gonzalez, R. Oxidative Stress Orchestrates Cell Polarity to Promote Embryonic Wound Healing. Developmental Cell 2018, 47(3), 377–387.e4. [Google Scholar] [CrossRef]
- Pb, B.; Rani, S.; Kim, Y.O.; Ahmed Al-Ghamdi, A.; Elshikh, M.S.; Al-Dosary, M.A.; Hatamleh, A.A.; Arokiyaraj, S.; Kim, H.-J. Prophylactic efficacy of Boerhavia diffusa L. aqueous extract in toluene induced reproductive and developmental toxicity in Drosophila melanogaster. Journal of Infection and Public Health 2020, 13(2), 177–185. [Google Scholar] [CrossRef]
- Wang, C.; Yang, L.; Hu, Y.; Zhu, J.; Xia, R.; Yu, Y.; Shen, J.; Zhang, Z.; Wang, S.-L. Isoliquiritigenin as an antioxidant phytochemical ameliorates the developmental anomalies of zebrafish induced by 2,2′,4,4′-tetrabromodiphenyl ether. Science of The Total Environment 2019, 666, 390–398. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.-L.; Yu, Y.-C.; Hsia, S.-M. Perspectives on the Role of Isoliquiritigenin in Cancer. Cancers 2021, 13(1), 115. [Google Scholar] [CrossRef] [PubMed]
- Gerlai, R. Zebrafish (Danio rerio): A newcomer with great promise in behavioral neuroscience. Neuroscience & Biobehavioral Reviews 2023, 144, 104978. [Google Scholar]
- Adefisan, A.O.; Madu, J.C.; Owumi, S.E.; Adaramoye, O.A. Calliandra portoricensisameliorates ovarian and uterine oxido-inflammatory responses inN-methyl-N-nitrosourea and benzo[a]pyrene-treated rats. Experimental Biology and Medicine 2020, 245(16), 1490–1503. [Google Scholar] [CrossRef] [PubMed]
- Majid, M.; Ijaz, F.; Baig, M.W.; Nasir, B.; Khan, M.R.; Haq, I.-u. Scientific Validation of Ethnomedicinal Use ofIpomoea batatasL. Lam. as Aphrodisiac and Gonadoprotective Agent against Bisphenol A Induced Testicular Toxicity in Male Sprague Dawley Rats. BioMed Research International 2019, 2019, 1–21. [Google Scholar] [CrossRef]
- Escobar-Puentes, A.A.; Palomo, I.; Rodríguez, L.; Fuentes, E.; Villegas-Ochoa, M.A.; González-Aguilar, G.A.; Olivas-Aguirre, F.J.; Wall-Medrano, A. Sweet Potato (Ipomoea batatas L.) Phenotypes: From Agroindustry to Health Effects. Foods 2022, 11(7), 1058. [Google Scholar] [CrossRef]
- Gupta, A.; Kumar, R.; Ganguly, R.; Singh, A.K.; Rana, H.K.; Pandey, A.K. Antioxidant, anti-inflammatory and hepatoprotective activities of Terminalia bellirica and its bioactive component ellagic acid against diclofenac induced oxidative stress and hepatotoxicity. Toxicology Reports 2021, 8, 44–52. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Hussein, O.E.; Hozayen, W.G.; Bin-Jumah, M.; Abd El-Twab, S.M. Ferulic acid prevents oxidative stress, inflammation, and liver injury via upregulation of Nrf2/HO-1 signaling in methotrexate-induced rats. Environmental Science and Pollution Research 2019, 27(8), 7910–7921. [Google Scholar] [CrossRef]
- Hu, J.; Jiang, K.; Tang, X.; Liu, H.; Zhang, H.; Yang, X.; Nie, X.; Luo, H. Chronic exposure to di-n-butyl phthalate causes reproductive toxicity in zebrafish. Journal of Applied Toxicology 2020, 40(12), 1694–1703. [Google Scholar] [CrossRef]
- Gupta, P.; Seth, C.S. 24-Epibrassinolide Regulates Functional Components of Nitric Oxide Signalling and Antioxidant Defense Pathways to Alleviate Salinity Stress in Brassica juncea L. cv. Varuna. Journal of Plant Growth Regulation 2022, 42(7), 4207–4222. [Google Scholar] [CrossRef]
- Nakamoto, M.; Kunimura, K.; Suzuki, J.I.; Kodera, Y. Antimicrobial properties of hydrophobic compounds in garlic: Allicin, vinyldithiin, ajoene and diallyl polysulfides (Review). Experimental and Therapeutic Medicine 2019. [Google Scholar] [CrossRef] [PubMed]
- Kazmi, S.T.B.; Majid, M.; Maryam, S.; Rahat, A.; Ahmed, M.; Khan, M.R.; Haq, I.u. Quercus dilatata Lindl. ex Royle ameliorates BPA induced hepatotoxicity in Sprague Dawley rats. Biomedicine & Pharmacotherapy 2018, 102, 728–738. [Google Scholar]
- Das, M.; Basu, S.; Banerjee, B.; Sen, A.; Jana, K.; Datta, G. Hepatoprotective effects of green Capsicum annum against ethanol induced oxidative stress, inflammation and apoptosis in rats. Journal of Ethnopharmacology 2018, 227, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Fossen, T.; Andersen, Ø.M. Anthocyanins from tubers and shoots of the purple potato, Solanum tuberosum. The Journal of Horticultural Science and Biotechnology 2015, 75(3), 360–363. [Google Scholar] [CrossRef]
- Fareed, M.M.; Khalid, H.; Khalid, S.; Shityakov, S. Deciphering Molecular Mechanisms of Carbon Tetrachloride-Induced Hepatotoxicity: A Brief Systematic Review. Current Molecular Medicine 2024, 24(9), 1124–1134. [Google Scholar] [CrossRef]
- Jeong, T.B.; Kwon, D.; Son, S.W.; Kim, S.H.; Lee, Y.-H.; Seo, M.-S.; Kim, K.S.; Jung, Y.-S. Weaning Mice and Adult Mice Exhibit Differential Carbon Tetrachloride-Induced Acute Hepatotoxicity. Antioxidants 2020, 9(3), 201. [Google Scholar] [CrossRef]
- Unsal, V.; Cicek, M.; Sabancilar, İ. Toxicity of carbon tetrachloride, free radicals and role of antioxidants. Reviews on Environmental Health 2021, 36(2), 279–295. [Google Scholar] [CrossRef]
- Arbab, A.H.; Parvez, M.K.; Al-Dosari, M.S.; Al-Rehaily, A.J.; Ibrahim, K.E.; Alam, P.; Alsaid, M.S.; Rafatullah, S. Therapeutic efficacy of ethanolic extract ofAerva javanicaaerial parts in the amelioration of CCl4-induced hepatotoxicity and oxidative damage in rats. Food & Nutrition Research 2016, 60(1), 30864. [Google Scholar]
- Ben Hsouna, A.; Gargouri, M.; Dhifi, W.; Saibi, W. Antioxidant and hepato-preventive effect ofCitrus aurantiumextract against carbon tetrachloride-induced hepatotoxicity in rats and characterisation of its bioactive compounds by HPLC-MS. Archives of Physiology and Biochemistry 2018, 125(4), 332–343. [Google Scholar] [CrossRef]
- Sobeh, M.; Youssef, F.S.; Esmat, A.; Petruk, G.; El-Khatib, A.H.; Monti, D.M.; Ashour, M.L.; Wink, M. High resolution UPLC-MS/MS profiling of polyphenolics in the methanol extract of Syzygium samarangense leaves and its hepatoprotective activity in rats with CCl4-induced hepatic damage. Food and Chemical Toxicology 2018, 113, 145–153. [Google Scholar] [CrossRef]
- Mohamed, M.R.; Emam, M.A.; Hassan, N.S.; Mogadem, A.I. Umbelliferone and daphnetin ameliorate carbon tetrachloride-induced hepatotoxicity in rats via nuclear factor erythroid 2-related factor 2-mediated heme oxygenase-1 expression. Environmental Toxicology and Pharmacology 2014, 38(2), 531–541. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.D.; D’Souza, U.J.A.; Iqbal, M. The potential protective effect of Commelina nudiflora L. against carbon tetrachloride (CCl4)-induced hepatotoxicity in rats, mediated by suppression of oxidative stress and inflammation. Environmental Health and Preventive Medicine 2017, 22(1). [Google Scholar] [CrossRef] [PubMed]
- Burits, M.; Bucar, F. Antioxidant activity of Nigella sativa essential oil. Phytotherapy Research 2000, 14(5), 323–328. [Google Scholar] [CrossRef] [PubMed]
- Mazaheri, Y.; Torbati, M.; Azadmard-Damirchi, S.; Savage, G.P. A comprehensive review of the physicochemical, quality and nutritional properties of Nigella sativa oil. Food Reviews International 2019, 35(4), 342–362. [Google Scholar] [CrossRef]
- Najmi, A.; Nasiruddin, M.; Khan, R.; Haque, S. Effect of <i> Nigella sativa</i> oil on various clinical and biochemical parameters of insulin resistance syndrome. International Journal of Diabetes in Developing Countries 2008, 28(1), 11. [Google Scholar]
- Imafidon, C.E.; Olukiran, O.S.; Ogundipe, D.J.; Eluwole, A.O.; Adekunle, I.A.; Oke, G.O. Acetonic extract of Vernonia amygdalina (Del.) attenuates Cd-induced liver injury: Potential application in adjuvant heavy metal therapy. Toxicology Reports 2018, 5, 324–332. [Google Scholar] [CrossRef]
- Ibiam, A.U.; Ugwuja, E.I.; Ejeogo, C.; Ugwu, O. Cadmium-induced toxicity and the hepatoprotective potentials of aqueous extract of jessiaea nervosa leaf. Adv Pharm Bull 2013, 3(2), 309–313. [Google Scholar]
- Bhattacharjee, B.; Pal, P.K.; Ghosh, A.K.; Mishra, S.; Chattopadhyay, A.; Bandyopadhyay, D. Aqueous bark extract of Terminalia arjuna protects against cadmium-induced hepatic and cardiac injuries in male Wistar rats through antioxidative mechanisms. Food and Chemical Toxicology 2019, 124, 249–264. [Google Scholar] [CrossRef]
- Vicidomini, C.; Roviello, V.; Roviello, G.N. In Silico Investigation on the Interaction of Chiral Phytochemicals from Opuntia ficus-indica with SARS-CoV-2 Mpro. Symmetry 2021, 13(6), 1041. [Google Scholar] [CrossRef]
- Hfaiedh, M.; Brahmi, D.; Zourgui, M.N.; Zourgui, L. Phytochemical analysis and nephroprotective effect of cactus (Opuntia ficus-indica) cladodes on sodium dichromate-induced kidney injury in rats. Applied Physiology, Nutrition, and Metabolism 2019, 44(3), 239–247. [Google Scholar] [CrossRef]
- Ncibi, S.; Ben Othman, M.; Akacha, A.; Krifi, M.N.; Zourgui, L. Opuntia ficus indica extract protects against chlorpyrifos-induced damage on mice liver. Food and Chemical Toxicology 2008, 46(2), 797–802. [Google Scholar] [CrossRef] [PubMed]
- Saad, A.b.; Rjeibi, I.; Ncib, S.; Zouari, N.; Zourgui, L. Ameliorative Effect of Cactus(Opuntia ficus indica)Extract on Lithium-Induced Nephrocardiotoxicity: A Biochemical and Histopathological Study. BioMed Research International 2017, 2017, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ahn, C.B.; Song, C.H.; Kim, W.H.; Kim, Y.K. Effects of Juglans sinensis Dode extract and antioxidant on mercury chloride-induced acute renal failure in rabbits. Journal of Ethnopharmacology 2002, 82(1), 45–49. [Google Scholar] [CrossRef] [PubMed]
- Sarwar Alam, M.; Kaur, G.; Jabbar, Z.; Javed, K.; Athar, M. Eruca sativa seeds possess antioxidant activity and exert a protective effect on mercuric chloride induced renal toxicity. Food and Chemical Toxicology 2007, 45(6), 910–920. [Google Scholar] [CrossRef]
- Ruiz, J.; Mahmud, M.; Modasshir, M.; Shamim Kaiser, M.; Alzheimer’s Disease Neuroimaging In, f. t. 3D DenseNet Ensemble in 4-Way Classification of Alzheimer’s Disease. 2020, 12241, 85–96.
- Bloom, G.S. Amyloid-β and tau: The trigger and bullet in Alzheimer disease pathogenesis. JAMA neurology 2014, 71(4), 505–508. [Google Scholar] [CrossRef] [PubMed]
- Castellani, R.J.; Rolston, R.K.; Smith, M.A. Alzheimer Disease. Disease-a-Month 2010, 56(9), 484–546. [Google Scholar] [CrossRef] [PubMed]
- Goedert, M.; Klug, A.; Crowther, R.A. Tau protein, the paired helical filament and Alzheimer’s disease. Journal of Alzheimer’s Disease 2006, 9(s3), 195–207. [Google Scholar] [CrossRef]
- Li, Y.; Liu, Y.; Wang, Z.; Jiang, Y. Clinical trials of amyloid-based immunotherapy for Alzheimer’s disease: End of beginning or beginning of end? Expert opinion on biological therapy 2013, 13(11), 1515–1522. [Google Scholar] [CrossRef]
- Cardoso, F.; Goetz, C.G.; Mestre, T.A.; Sampaio, C.; Adler, C.H.; Berg, D.; Bloem, B.R.; Burn, D.J.; Fitts, M.S.; Gasser, T.; Klein, C.; de Tijssen, M.A.J.; Lang, A.E.; Lim, S.Y.; Litvan, I.; Meissner, W.G.; Mollenhauer, B.; Okubadejo, N.; Okun, M.S.; Postuma, R.B.; Svenningsson, P.; Tan, L.C.S.; Tsunemi, T.; Wahlstrom-Helgren, S.; Gershanik, O.S.; Fung, V.S.C.; Trenkwalder, C. A Statement of the MDS on Biological Definition, Staging, and Classification of Parkinson’s Disease. Movement Disorders 2023, 39(2), 259–266. [Google Scholar] [CrossRef]
- Irwin, K.E.; Sheth, U.; Wong, P.C.; Gendron, T.F. Fluid biomarkers for amyotrophic lateral sclerosis: A review. Molecular Neurodegeneration 2024, 19(1). [Google Scholar] [CrossRef]
- Garofalo, M.; Pandini, C.; Bordoni, M.; Pansarasa, O.; Rey, F.; Costa, A.; Minafra, B.; Diamanti, L.; Zucca, S.; Carelli, S.; Cereda, C.; Gagliardi, S. Alzheimer’s, Parkinson’s Disease and Amyotrophic Lateral Sclerosis Gene Expression Patterns Divergence Reveals Different Grade of RNA Metabolism Involvement. International Journal of Molecular Sciences 2020, 21(24), 9500. [Google Scholar] [CrossRef] [PubMed]
- Zahra, W.; Rai, S.N.; Birla, H.; Singh, S.S.; Dilnashin, H.; Rathore, A.S.; Singh, S.P. The Global Economic Impact of Neurodegenerative Diseases: Opportunities and Challenges. 2020, 333–345.
- Pathak, N.; Vimal, S.K.; Tandon, I.; Agrawal, L.; Hongyi, C.; Bhattacharyya, S. Neurodegenerative Disorders of Alzheimer, Parkinsonism, Amyotrophic Lateral Sclerosis and Multiple Sclerosis: An Early Diagnostic Approach for Precision Treatment. Metabolic Brain Disease 2021, 37(1), 67–104. [Google Scholar] [CrossRef] [PubMed]
- Rekatsina, M.; Paladini, A.; Piroli, A.; Zis, P.; Pergolizzi, J.V.; Varrassi, G. Pathophysiology and Therapeutic Perspectives of Oxidative Stress and Neurodegenerative Diseases: A Narrative Review. Advances in Therapy 2019, 37(1), 113–139. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhu, M.; Kumar, M.; Ngo, F.Y.; Li, Y.; Lao, L.; Rong, J. A Pharmacological Appraisal of Neuroprotective and Neurorestorative Flavonoids Against Neurodegenerative Diseases. CNS & Neurological Disorders - Drug Targets 2019, 18(2), 103–114. [Google Scholar]
- Akter, R.; Chowdhury, M.A.R.; Rahman, M.H. Flavonoids and Polyphenolic Compounds as Potential Talented Agents for the Treatment of Alzheimer’s Disease and their Antioxidant Activities. Current Pharmaceutical Design 2021, 27(3), 345–356. [Google Scholar] [CrossRef]
- Olajide, O.J.; Enaibe, B.U.; Bankole, O.O.; Akinola, O.B.; Laoye, B.J.; Ogundele, O.M. Kolaviron was protective against sodium azide (NaN3) induced oxidative stress in the prefrontal cortex. Metabolic Brain Disease 2015, 31(1), 25–35. [Google Scholar] [CrossRef]
- Saad, A.B.; Rjeibi, I.; Brahmi, N.; Elaloui, E.; Zouari, N. Nicotine-induced oxidative stress, testis injury, AChE inhibition and brain damage alleviated by Mentha spicata. Inflammopharmacology 2019, 28(4), 939–948. [Google Scholar] [CrossRef]
- Rebai, O.; Belkhir, M.; Boujelben, A.; Fattouch, S.; Amri, M. Morus alba leaf extract mediates neuroprotection against glyphosate-induced toxicity and biochemical alterations in the brain. Environmental Science and Pollution Research 2017, 24(10), 9605–9613. [Google Scholar] [CrossRef]
- Abdul-Aziz Ahmed, K.; Jabbar, A.A.J.; Abdulla, M.A.; Zuhair Alamri, Z.; Ain Salehen, N.; Abdel Aziz Ibrahim, I.; Almaimani, G.; Bamagous, G.A.; Almaimani, R.A.; Almasmoum, H.A.; Ghaith, M.M.; Farrash, W.F. Mangiferin (mango) attenuates AOM-induced colorectal cancer in rat’s colon by augmentation of apoptotic proteins and antioxidant mechanisms. Scientific Reports 2024, 14(1). [Google Scholar] [CrossRef]
- Plavec, J. Quadruplex targets in neurodegenerative diseases. 2020, 54, 441–483.
- Wink, M. Compartmentation of Secondary Metabolites and Xenobiotics in Plant Vacuoles. In The Plant Vacuole, 1997; pp 141–169.
- Kolb, M.; Harms, H. Metabolism of fluoranthene in different plant cell cultures and intact plants. Environmental Toxicology and Chemistry 2009, 19(5), 1304–1310. [Google Scholar] [CrossRef]
- Sikandar, A.; Shehzadi, K.; Arshad, Q.; Munir, K. Phytoremediation: An analytical technique for the assessment of biodegradation of organic xenobiotic pollutants: A review. Int. J. Sci. Res 2013, 4(2), 2250–2253. [Google Scholar]
- Sun, C.; Dudley, S.; McGinnis, M.; Trumble, J.; Gan, J. Acetaminophen detoxification in cucumber plants via induction of glutathione S-transferases. Science of The Total Environment 2019, 649, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Huihui, Z.; Xin, L.; Zisong, X.; Yue, W.; Zhiyuan, T.; Meijun, A.; Yuehui, Z.; Wenxu, Z.; Nan, X.; Guangyu, S. Toxic effects of heavy metals Pb and Cd on mulberry (Morus alba L.) seedling leaves: Photosynthetic function and reactive oxygen species (ROS) metabolism responses. Ecotoxicology and Environmental Safety 2020, 195. [Google Scholar] [CrossRef] [PubMed]
- Benzarti, S.; Mohri, S.; Ono, Y. Plant response to heavy metal toxicity: Comparative study between the hyperaccumulator Thlaspi caerulescens (ecotype Ganges) and nonaccumulator plants: Lettuce, radish, and alfalfa. Environmental Toxicology 2008, 23(5), 607–616. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhang, Y.; Yang, B.; Fang, J.; Liu, Z. Transcriptomic responses to different doses of cycloxaprid involved in detoxification and stress response in the whitebacked planthopper, Sogatella furcifera. Entomologia Experimentalis et Applicata 2016, 158(3), 248–257. [Google Scholar] [CrossRef]
Scheme 1.
Flowcharts A, B, and C. Flowchart A describes the protective effects of a group of natural products, including Barley grass, carrot, and Citrus fruit, highlighting their antioxidant chemicals and mechanisms. Flowchart B reports on another group of natural products: Eleuthero root, Ginkgo leaf, and grape seed/skin, emphasizing their protective chemicals and actions against oxidative damage. Flowchart C shows the mechanisms of action for a further group of natural products, such as Kudzu root, milk thistle seed, rosemary leaf, Schisandra fruit, tomato, and Turmeric root, detailing their specific chemicals and how they inhibit inflammation and enhance antioxidant defenses.
Scheme 1.
Flowcharts A, B, and C. Flowchart A describes the protective effects of a group of natural products, including Barley grass, carrot, and Citrus fruit, highlighting their antioxidant chemicals and mechanisms. Flowchart B reports on another group of natural products: Eleuthero root, Ginkgo leaf, and grape seed/skin, emphasizing their protective chemicals and actions against oxidative damage. Flowchart C shows the mechanisms of action for a further group of natural products, such as Kudzu root, milk thistle seed, rosemary leaf, Schisandra fruit, tomato, and Turmeric root, detailing their specific chemicals and how they inhibit inflammation and enhance antioxidant defenses.
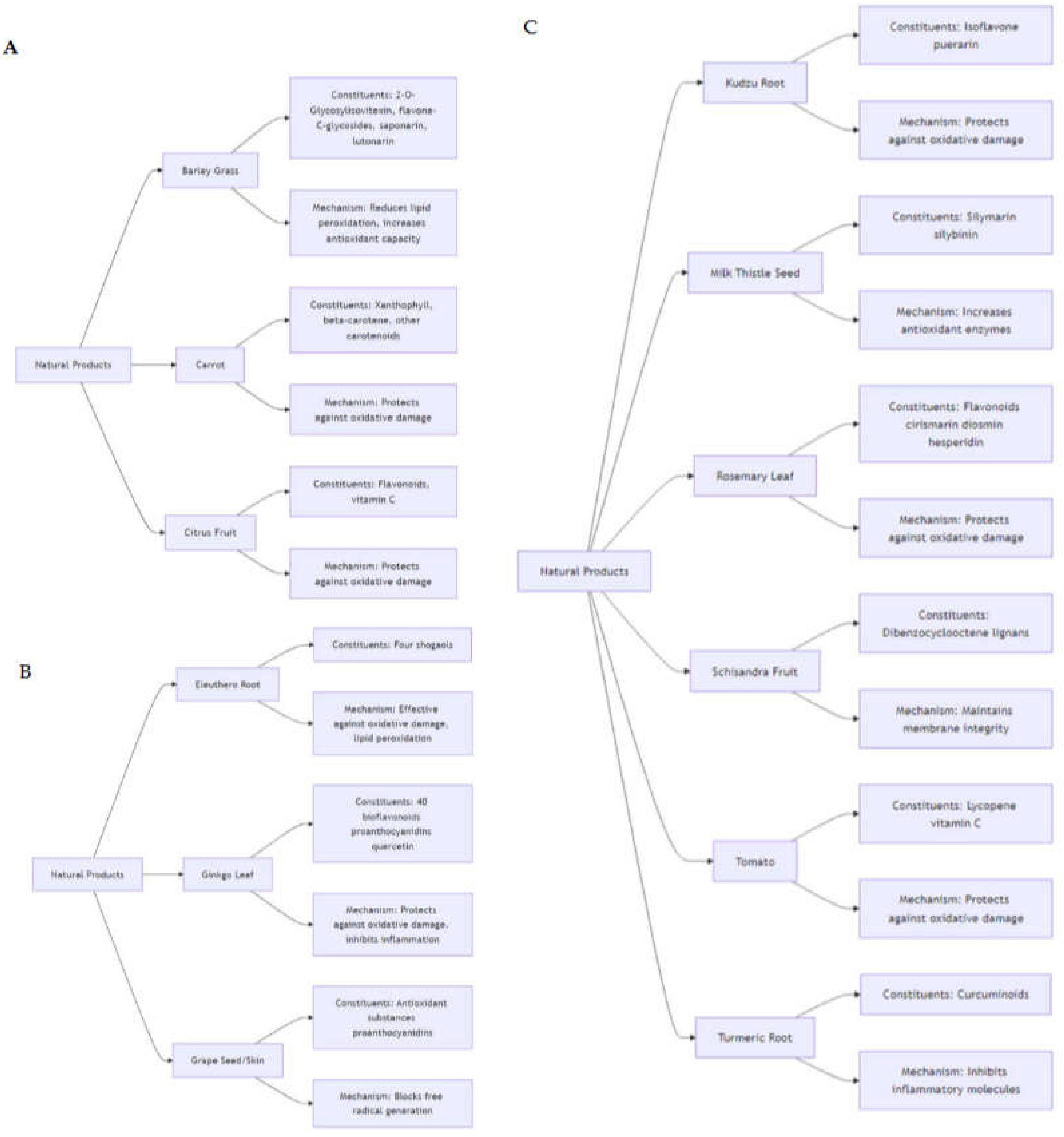
Figure 1.
Diagram showing the ameliorative effects of Solanum aethiopicum (photo courtesy of Giovanni N. Roviello) on anemia induced by the xenobiotic compound phenylhydrazine. The 3D structure of AST is freely available at the link https://www.rcsb.org/structure/1AAM (accessed on 8th August 2024).
Figure 1.
Diagram showing the ameliorative effects of Solanum aethiopicum (photo courtesy of Giovanni N. Roviello) on anemia induced by the xenobiotic compound phenylhydrazine. The 3D structure of AST is freely available at the link https://www.rcsb.org/structure/1AAM (accessed on 8th August 2024).
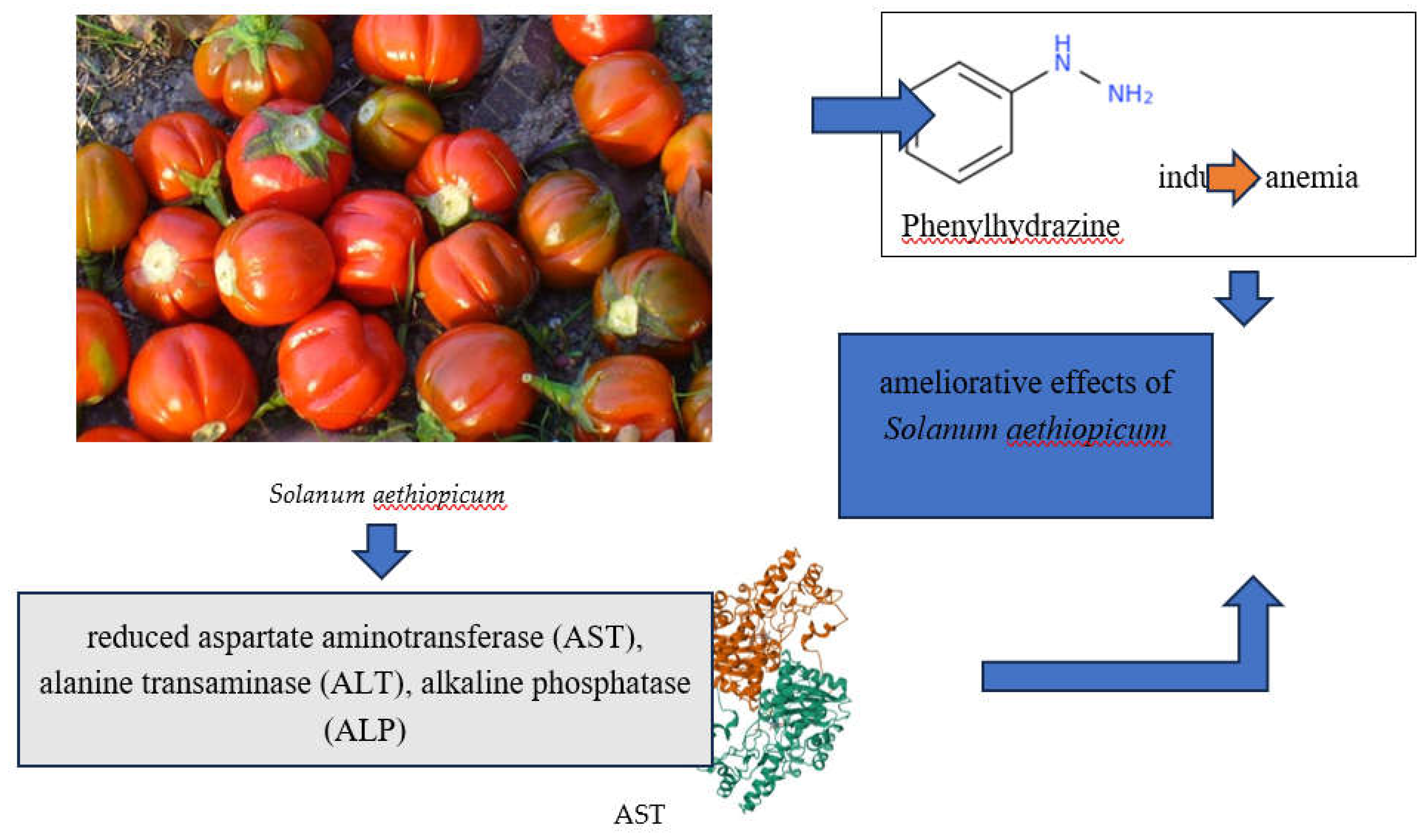
Figure 2.
(a) Vitamin C, Vitamin A, and Glutathione (GSH) are some of the main non-enzymatic antioxidants that play significant roles in maintaining redox homeostasis. 3D structure of GSH is freely available at the link https://pubchem.ncbi.nlm.nih.gov/compound/448041#section=3D-Conformer (accessed on 8th August 2024).
Figure 2.
(a) Vitamin C, Vitamin A, and Glutathione (GSH) are some of the main non-enzymatic antioxidants that play significant roles in maintaining redox homeostasis. 3D structure of GSH is freely available at the link https://pubchem.ncbi.nlm.nih.gov/compound/448041#section=3D-Conformer (accessed on 8th August 2024).

Figure 3.
(a) Representation of the Fenton reaction and the production of hydroxyl radicals (•OH), which play a crucial role in the development of oxidative stress-related diseases and conditions; (b) diagram showing hydroquinone, a harmful intermediate produced after xenobiotic exposure, which leads to cellular damage and oxidative stress.
Figure 3.
(a) Representation of the Fenton reaction and the production of hydroxyl radicals (•OH), which play a crucial role in the development of oxidative stress-related diseases and conditions; (b) diagram showing hydroquinone, a harmful intermediate produced after xenobiotic exposure, which leads to cellular damage and oxidative stress.
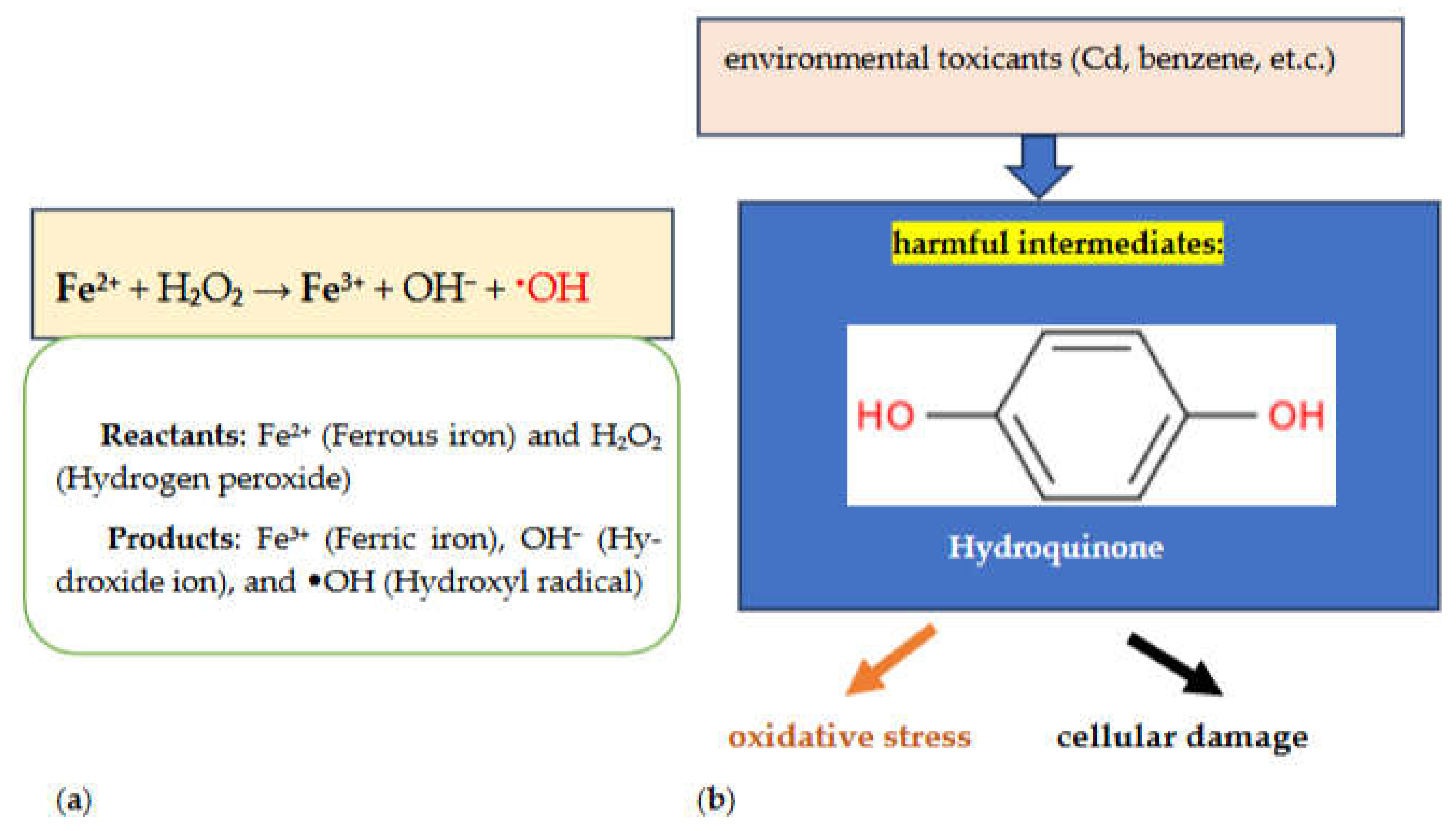
Figure 4.
(a) Diagram illustrating the ameliorating effects of ISL, a natural chalcone-type flavonoid with antioxidant, anti-allergic, anti-inflammatory, anti-tumor, and estrogenic properties, against the developmental toxicity of BDE47, an abundant polybrominated diphenyl ether; the 3D structure of caspase 9 is freely available at the link https://www.rcsb.org/structure/3V3K (accessed on 8th August 2024) (b) 3D views of the human microsomal cytochrome P450 2E1 (CYP2E1) structure in complex with the inhibitor 4-methylpyrazole. CYP2E1 is involved in the monooxygenation of over 70 low molecular weight xenobiotic compounds. The structure can be freely accessed at https://www.rcsb.org/3d-view/3E4E/1 (accessed on July 27, 2024).
Figure 4.
(a) Diagram illustrating the ameliorating effects of ISL, a natural chalcone-type flavonoid with antioxidant, anti-allergic, anti-inflammatory, anti-tumor, and estrogenic properties, against the developmental toxicity of BDE47, an abundant polybrominated diphenyl ether; the 3D structure of caspase 9 is freely available at the link https://www.rcsb.org/structure/3V3K (accessed on 8th August 2024) (b) 3D views of the human microsomal cytochrome P450 2E1 (CYP2E1) structure in complex with the inhibitor 4-methylpyrazole. CYP2E1 is involved in the monooxygenation of over 70 low molecular weight xenobiotic compounds. The structure can be freely accessed at https://www.rcsb.org/3d-view/3E4E/1 (accessed on July 27, 2024).
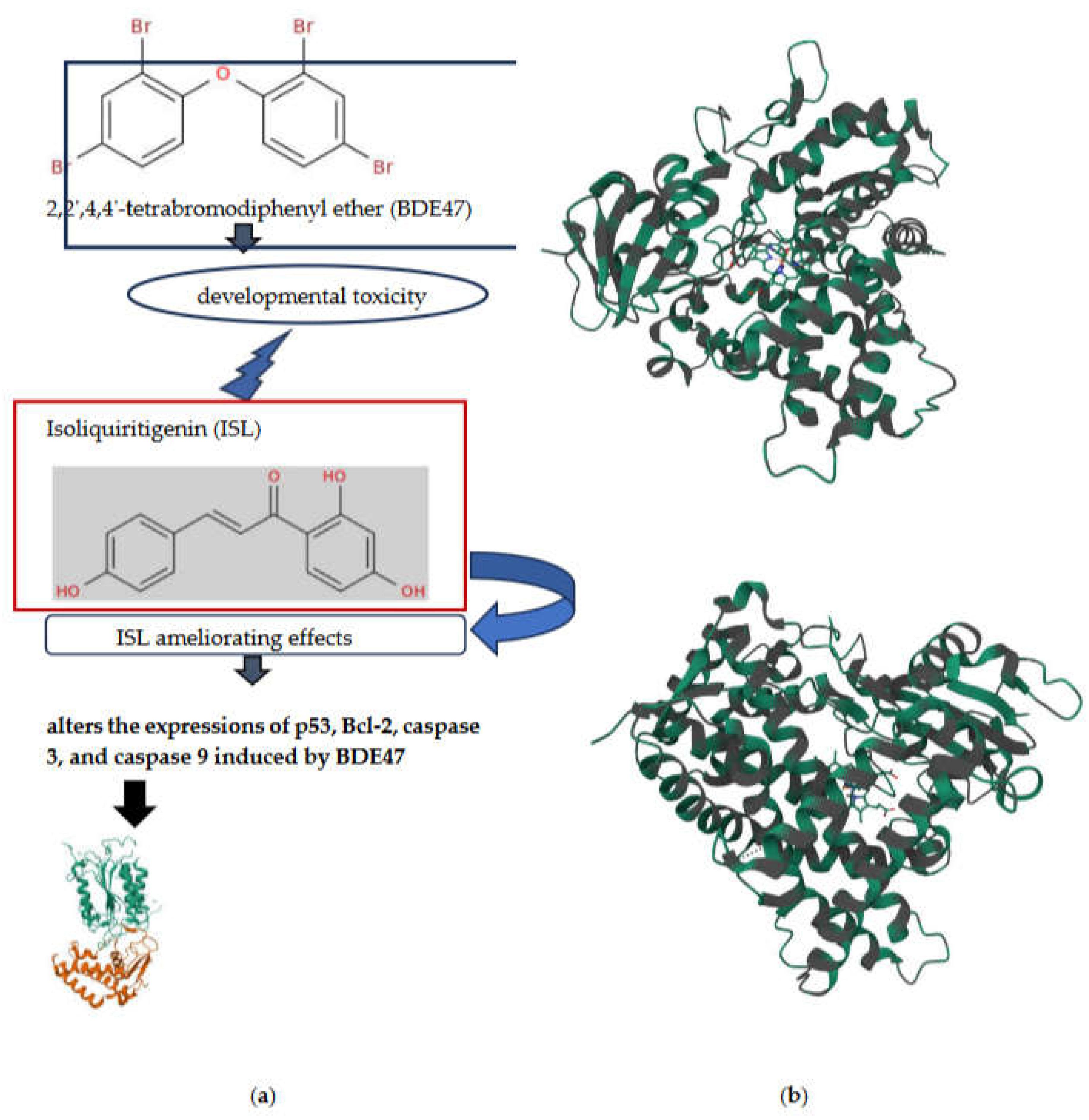
Figure 5.
Diagram showing how Beta vulgaris L. (beetroot), known for its minerals and vitamins, counteracts chlorpyrifos (CPF) nephrotoxicity by restoring antioxidants and limiting oxidative stress. Beetroot products have effects that increase nuclear factor erythroid 2-related factor 2 (Nrf2), which is crucial in promoting the expression of defensive enzymes
Figure 5.
Diagram showing how Beta vulgaris L. (beetroot), known for its minerals and vitamins, counteracts chlorpyrifos (CPF) nephrotoxicity by restoring antioxidants and limiting oxidative stress. Beetroot products have effects that increase nuclear factor erythroid 2-related factor 2 (Nrf2), which is crucial in promoting the expression of defensive enzymes
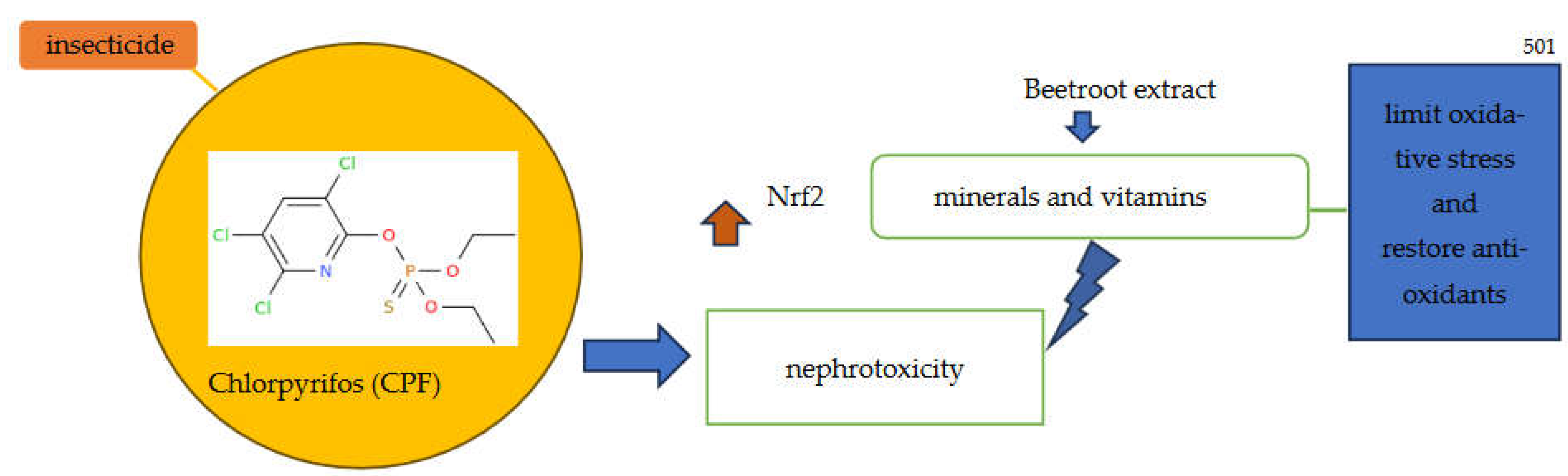

Table 1.
Overview of natural products, their key active ingredients, and their protective effects.
| Natural Product | Major Constituents | Effects | Ref. |
|---|---|---|---|
| Barley grass (Hordeum leporinum) | 2-O-glycosylisovitexin (2-O-GIV; C27H30O15), flavone-C-glycosides, saponarin (C27H30O15), and lutonarin (C27H30O16) | Decreased lipid peroxidation and enhanced antioxidant capacity. | [31,32] |
| Carrot (Daucus Carota) | Xanthophyll (C40H56O2), beta-carotene (C40H56), and other antioxidant carotenoids | Prevention of oxidative damage through strong antioxidant capacity. | [31] |
| Citrus fruit | Flavonoids and vitamin C (C6H8O6) | Highly effective in protecting against oxidative damage | [33] |
| Eleuthero root (Eleutherococcus senticosus) | Four shogaols (bioactive compounds derived from ginger) with antioxidant properties | Provides strong protection against oxidative damage and lipid peroxidation | [34,35,36] |
| Ginger root (Zingiber officinale) | About 40 bioflavonoids, including proanthocyanidins and quercetin (C15H10O7) | Offers protection against oxidative damage and lipid peroxidation | [35,37,38] |
| Ginkgo leaf (Ginkgo biloba) | About 40 bioflavonoids, including proanthocyanidins and quercetin (C15H10O7) | Provides protection against oxidative damage while inhibiting inflammation and atherosclerosis. | [33,35,39,40] |
| Grape (vitis sp.) seed/skin | Antioxidant substances, over 90% proanthocyanidins | Efficient at preventing free radical formation and decreasing oxidative stress. | [35] |
| Kudzu root (Pueraria lobata) | Isoflavone puerarin (C21H20O9, crude form more efficient) | Active against oxidative damage due to elevated antioxidant capacity. | [41,42,43] |
| Milk thistle seed (Silybum marianum) | Silymarin (C25H22O10, major constituent silybinin) | Provides protection against oxidative damage and lipid peroxidation; enhances antioxidant enzyme activity. | [44,45,46,47] |
| Rosemary leaf (Rosmarinus officinalis) | Flavonoids such as cirsimarin (C23H24O11), diosmin (C28H32O15), hesperidin (C28H34O15), and homoplantaginin (C22H22O11) | Effectively protects against oxidative damage and also possesses analgesic properties. | [31,48] |
| Schisandra fruit (Schisandra chinensis) | At least 9 dibenzocyclooctene lignans | Provides protection against oxidative damage and helps preserve membrane integrity. | [49] |
| Tomato (Solanum lycopersicum) | Lycopene (C15H10O7) and vitamin C (C6H8O6) | Offers strong protection against oxidative damage due to its high antioxidant content. | [33] |
| Turmeric root (Curcuma longa) | Curcuminoids | Protects against oxidative damage and inflammation; inhibits various inflammatory molecules | [35] |
Table 2.
Overview of the interactions between some natural products and their effects on organ toxicity induced by environmental toxicants. The table lists the specific toxicant involved, the oxidative stress parameters observed, the resulting effects on the organism, and the natural products that can ameliorate these toxic effects.
Table 2.
Overview of the interactions between some natural products and their effects on organ toxicity induced by environmental toxicants. The table lists the specific toxicant involved, the oxidative stress parameters observed, the resulting effects on the organism, and the natural products that can ameliorate these toxic effects.
| Organ toxicity | Environmental Toxicants | Oxidative stress Parameters | Effects | Natural Products | Ref. |
|---|---|---|---|---|---|
| nephrotoxicity | bromobenzene C₆H₅Br | increased LPO, reduced antioxidant enzyme activity | inflammation and irregularities in kidney function | withaferin A | [66] |
| reproductive and developmental toxicity | lead Pb | redox imbalance | learning and memory impairments in young rats | allicin | [67] |
| ” | di-n-butyl phthalate C₁6H₂₂O₄ |
increased MDA, decreased SOD and GPx levels | multinucleated gonocytes and reduced serum testosterone levels | taxifolin | [68] |
| ” | Ethanol C2H5OH | altered levels of reduced and oxidized glutathione | malformations and changes in embryonic behavior | 24-epibrassinolide | [69] |
| nephrotoxicity | zinc oxide (ZnO) nanoparticle | reduced non-enzymatic antioxidant levels | reduced renal GSH, elevated serum inflammatory markers, and glucose levels | quercetin and arginine | [70] |
| ” | Arsenic As | formation of 8-OxodG | raised methylation levels, urinary excretion | black raspberries | [71] |
| ” | ” | reduced antioxidant enzyme activity, increased MDA | renal damage causing cellular apoptosis | thymoquinone | [72] |
| reproductive and developmental toxicity | ” | regulation of Nfe2l2, StAR, Ppargc1a expressions | abnormalities in sperm and testis structure and function | ellagic and ferulic acid | [73] |
|
” |
acrylamide C3H5NO |
elevated ROS | mobility problems, egg development deficiencies | thymoquinone and curcumin | [74] |
| nephrotoxicity | rotenone C23H22O6 |
redox imbalance | renal dysfunction | catechin, quercetin, taxifolin | [75] |
| ” | cadmium Cd |
decreased antioxidant enzyme activity, increased LPO | injury to proximal tubules, dysfunction of the glomerulus | thymoquinone | [76] |
| ” | ” | decreased antioxidant enzyme activity, increased iNOS, MMP-9, pERK 1/2 | alterations in tubular epithelium and glomerular structure | flavocoxid | [77] |
| reproductive and developmental toxicity | nicotine C10H14N2 |
reduced antioxidant enzyme levels | structural and functional impairment in the testis and brain | Mentha spicata | [78] |
Table 3.
Phytochemical composition of sweet potato (Ipomoea batatas) extracts, showing the Total Phenolic Content (TPC), Total Flavonoid Content (TFC), and individual flavonoid contents (rutin and myricetin), as well as caffeic acid levels in both ethyl acetate and methanol extracts. Values are expressed in micrograms per milligram of dry extract, indicating the bioactive potential of the extracts.
Table 3.
Phytochemical composition of sweet potato (Ipomoea batatas) extracts, showing the Total Phenolic Content (TPC), Total Flavonoid Content (TFC), and individual flavonoid contents (rutin and myricetin), as well as caffeic acid levels in both ethyl acetate and methanol extracts. Values are expressed in micrograms per milligram of dry extract, indicating the bioactive potential of the extracts.
| Phytochemical | Content (Dry Extract) | ± |
|---|---|---|
| Total Phenolic Content (TPC) | 304.32 μg gallic acid equivalents/mg | 7.20 |
| Total Flavonoid Content (TFC) | 214.77 μg quercetin equivalents/mg | 4.09 |
| Rutin Content | 7.3 μg/mg | 0.12 |
| Myricetin Content | 2.7 μg/mg | 0.14 |
| Caffeic Acid Content (ethyl acetate extract) | 4.05 μg/mg | 0.22 |
| Caffeic Acid Content (methanol extract) | 1.92 μg/mg | 0.17 |
Table 4.
Examples of natural products mentioned in this section that were tested for their protective effects in various experimental models, detailing the experimental setup, observed results, and underlying mechanisms of action.
Table 4.
Examples of natural products mentioned in this section that were tested for their protective effects in various experimental models, detailing the experimental setup, observed results, and underlying mechanisms of action.
| Natural product | Experimental Model | Experimental Details & Results | Protective Effects & Mechanism | Ref. |
|---|---|---|---|---|
| Boerhavia diffusa | Drosophila larvae | B. diffusa aqueous extract (BDAE) was tested at four concentrations: 25, 50, 100, and 200 mg/ml. Results: BDAE mitigated toluene-induced developmental and reproductive toxicity, significantly improved reproductive and developmental parameters, and increased antioxidant enzyme levels. | Effectiveness: Reduced developmental and reproductive toxicity induced by toluene. Mechanism: Antioxidant activity, elevated antioxidant enzyme levels (catalase, glutathione-S-transferase, superoxide dismutase). | [81] |
| Isoliquiritigenin (ISL) | Zebrafish embryos | ISL was used at 4 μM with exposure to BDE47 (1 and 10 μM) for up to 120 hours post-fertilization. Results: ISL mitigated BDE47-induced developmental abnormalities, excessive ROS accumulation, and altered apoptosis-related gene expressions. | Effectiveness: Ameliorated developmental toxicity caused by BDE47. Mechanism: Enhanced antioxidant enzyme activities (SOD and CAT), modulated apoptosis-related gene expressions (p53, Bcl-2, caspase 3, caspase 9). | [82] |
| Calliandra portoricensis (CP) | Rats | CP was evaluated for its effects against ovarian and uterine toxicity induced by benzo[a]pyrene (BaP) and N-methyl nitrosourea (NMU). Results: CP reduced toxic responses from NMU and BaP, alleviating oxidative damage and inflammation. | Effectiveness: Mitigated ovarian and uterine toxicity from carcinogens. Mechanism: Antioxidant and anti-inflammatory properties. | [85] |
| Ipomoea batatas | Male Sprague Dawley rats | Ipomoea batatas extracts were tested for protective effects against BPA-induced gonadotoxicity at 300 mg/kg. Results: Enhanced sexual behavior, improved semen quality, increased levels of testosterone, FSH, LH, and estradiol, and restored antioxidant enzyme levels. | Effectiveness: Improved reproductive health and mitigated BPA-induced toxicity. Mechanism: Antioxidant activity, restoration of hormonal balance, improved semen quality. | [85] |
| Beetroot (Beta vulgaris L.) | Mice | Beetroot was evaluated for its protective effects against chlorpyrifos (CPF) nephrotoxicity. Results: Restored antioxidant levels and reduced oxidative stress markers. | Effectiveness: Mitigated CPF-induced nephrotoxicity. Mechanism: Enhanced antioxidant levels, reduced oxidative stress. Beetroot extracts enhance nuclear factor erythroid 2-related factor 2 (Nrf2), which is essential for promoting the expression of defensive enzymes. | [85] |
| Opuntia ficus indica | Mice | Opuntia cladodes extract was tested for its effects on liver damage caused by chlorpyrifos. Results: Improved liver function, reduced oxidative stress, and normalized biochemical parameters. | Effectiveness: Protected against liver damage from CPF. Mechanism: Restored antioxidant levels, reduced oxidative stress and liver damage. | [85] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



