
Submitted:
29 September 2024
Posted:
30 September 2024
You are already at the latest version
Abstract
The interplay between wildfires and drought stress has significantly limited forest recovery in the Mediterranean ecosystem of central Chile. Since 2010, the region has faced a prolonged 'Mega Drought,' which intensified into a 'Hyper Drought' in 2019, with record-low precipitation and high temperatures, further constraining forest recovery amidst increasing wildfire frequency. This study evaluates short-term vegetation recovery across drought gradients in three sclerophyllous evergreen forests, analyzing Landsat time series (1987–2022) for 42 wildfires. Using the LandTrendr algorithm, we assessed post-fire forest recovery based on NDVI changes between pre-fire values and subsequent years. The results revealed significant differences in recovery across drought gradients and fire severity interactions. After five years, the xeric forest dominated by Quillaja saponaria and Lithrea caustica showed 89% recovery during the Mega Drought and 34% during the Hyper Drought. In contrast, the mesic forest dominated by Cryptocarya alba and Peumus boldus exhibited 28% recovery during the Mega Drought and 43% during the Hyper Drought. Degraded thorny shrublands showed the lowest recovery, averaging 23% under Hyper Drought and medium fire severity. These findings underscore the critical role of prolonged, severe drought in shaping forest recovery dynamics and highlight the need to understand these patterns to improve future forest resilience under increasingly arid conditions.
Keywords:
Chile
; Mediterranean
; NDVI
; forest recovery
; drought
; wildfire
; fire severity
; remote sensing
; disturbance
; stressors interaction
1. Introduction
Natural disturbances, such as wildfires and droughts, play a crucial role in shaping the ecological dynamics of forest ecosystems by altering their structure, composition, and function [1,2]. These disturbances leave long-lasting imprints on forest trajectories, influencing their future states [3,4]. However, anthropogenic climate change and land-use practices have significantly altered these disturbances' frequency, intensity, and interactions [5,6,7]. As disturbances related to climate grow in duration and severity, they may cease to be isolated events and become chronic stressors, such as prolonged drought [8]. These persistent stressors can severely limit vegetation regrowth and elevate the risk of surpassing ecological thresholds, potentially leading to irreversible changes in forest structure and function [6,9]. This is particularly evident in Mediterranean regions, which may be vulnerable, where the interaction between wildfires and droughts can create tipping points[10], reducing the ability of ecosystems to provide essential services like carbon sequestration and hydrological regulation [3,11]. Comprehensive assessments of post-fire vegetation recovery under these stressors are essential for understanding how these ecosystems recover. Such evaluations also help to anticipate the broader impacts of environmental changes, ensuring the continued provision of ecosystem functions, including resource supply, biodiversity conservation, and ecosystem service maintenance.
Among Mediterranean ecosystems, central Chile faces intensified environmental stressors [12,13]. Located along the southern Pacific coast of South America, central Chile presents a unique opportunity to study the effects of changing disturbance regimes and the interactions between wildfires and drought. The region is recognized for its high level of endemism, making it one of the world's biodiversity hotspots [14]. Central Chile has been undergoing a significant dry period since 2010, characterized by rising temperatures and a long-term decline in rainfall since 1915 [15,16,17], with a precipitation deficit ranging from 25% to 45% for over a decade [18]. This prolonged drought, known as the "Mega Drought," reached extreme levels in 2019, with rainfall dropping below 30% of historical averages, alongside record-high temperatures during the spring/summer season [19]. This year was defined as a 'Hyper Drought', and the region experienced a noticeable decline in vegetation productivity compared to 2000–2010 [20].
These severe drought conditions have coincided with an increase in wildfire frequency and severity, exacerbated by climatic conditions favorable to the spread of fires, posing substantial risks to native vegetation [21,22,23]. Similar to other Mediterranean ecosystems, the historical transformation of land use in central Chile has significantly reduced the extent of original forests, leaving most remaining forest stands confined to steeper slopes with limited access to groundwater. Covering 3,970 hectares, these forests, mainly found in mesic, south-facing areas, consist of three primary forests of different species compositions (forest subtypes): degraded thorny shrublands dominated by Vachellia caven & Maytenus boaria located in xeric areas; Coastal Mediterranean Forests primarily covered by Cryptocarya alba & Peumus boldus located in mesic condition; and Andean Mediterranean Forests dominated by Quillaja Saponaria & Lithrea caustica established in drier, xeric location [24,25,26]. However, dense canopy coverage areas are mostly restricted to topographically water-accumulating sites or microclimates that favor their persistence. Evergreen sclerophyllous forests in central Chile are adapted to withstand low soil water potential and fire [27], natural ignitions are rare in central Chile due to infrequent thunderstorms without rain [28]. Many tree and shrub species in central Chile's Mediterranean forests exhibit resilience by regenerating primarily through resprouting, with some species maintaining viable seeds even after low-intensity fires [29,30,31]. Unlike other regions where fire stimulates seed production, only a few Chilean species display this trait [32,33].
Post-fire and drought recovery in these forests frequently rely on stored resources within survival structures and the varying ability of basal buds, such as lignotubers, roots, rhizomes, or nodes, to sprout [31]. While resprouting from underground structures is possible if sufficient carbon reserves are available, it can fail due to hydraulic limitations, making forest recovery uncertain if drought conditions persist and water balance declines [34]. Moreover, seedling recruitment after disturbances in Chile's sclerophyllous forests is limited by the rapid establishment of exotic species and the high abundance of herbivores. It remains unclear how different forests, and their dominant species respond to post-fire recovery under drought-driven declines in productivity.
Remote sensing is a practical tool for monitoring post-disturbance vegetation recovery. Analysis of time-series vegetation indices derived from remote sensing, such as the Normalized Difference Vegetation Index (NDVI), helps track vegetation recovery and photosynthetic capacity after events like wildfires. However, the effects of the interaction of long-term drought, Hyper Drought, and wildfire have been poorly documented because they rarely occur in extensive areas, high intensity, and decadal long periods. The Landsat program has been instrumental in offering high-resolution imagery, supporting advanced research using techniques like LandTrendr [35], that allow for in-depth analysis of vegetation changes following disturbances [36,37,38]. Notably, many studies have underscored the effectiveness of the Normalized Difference Vegetation Index (NDVI) as a proxy in assessing vegetation productivity recovery and more accurately than other indices [39,40]. NDVI is closely associated with photosynthetic capacity, as its greenness primarily indicates chlorophyll content [41,42]. By integrating advanced remote sensing techniques with analytical models, we enhanced our ability to characterize, assess, and monitor forest responses to fire, providing robust insights to inform conservation and management strategies effectively.
Given the expected increase in aridity for this region and the higher frequency of extreme events [43,44], comprehensive assessments at both regional and local scales were crucial for enhancing our understanding of vegetation recovery following fire disturbances. While recent studies have begun to explore the complex relationships between forests, drought, and wildfires in central Chile [13,21,45,46], few have examined the specific effects of post-fire recovery in these ecosystems. Mediterranean forests in the Southern Hemisphere, particularly in central Chile, remain underrepresented in global analyses despite their significant ecological importance [42]. As a result, there is a need for targeted research to better assess the dynamics of vegetation recovery in response to increasing climate stressors and fire severity
Considering the above context, this research aims to characterize and compare the short-term (5-year) spectral forest recovery following wildfire, as measured by NDVI, focusing on variations across drought levels and fire severity between forests of different species compositions (forest subtypes). We tried to respond to the following questions: Are there significant differences in forest recovery, across levels of drought and fire severity by forest subtypes? Are there interaction effects between drought and fire severity? Which forest subtype exhibited the lowest recovery, depending on drought level and fire severity?
2. Materials and Methods
2.1. Study Area
The study area is situated in central Chile, spanning from the Pacific coast to the Andean foothills, between longitudes 71.5° W and 70.0° W and latitudes 32.0° S to 36.0° S, covering four administrative regions: Valparaíso, Metropolitana, O'Higgins, and Maule (Figure 1). This region encompasses the core of Chile's Mediterranean sclerophyllous shrublands and forests, stretching from coastal zones to the pre-Andean mountains. The area experiences a Mediterranean climate, characterized by winter precipitation and a prolonged dry summer season, heavily influenced by the El Niño-Southern Oscillation (ENSO) [16]. Annual precipitation varies from approximately 220 mm in the central valley to around 700 mm along the coast, while mean annual temperatures range between 12°C and 15.1°C (data obtained from http://explorador.cr2.cl/). Although this ecosystem typically endures one to two years of drought, from 2010 to 2022, it experienced an unprecedented, prolonged drought, leading to a marked decline in vegetation productivity [20].
2.2. Selection of Burned Forest Areas
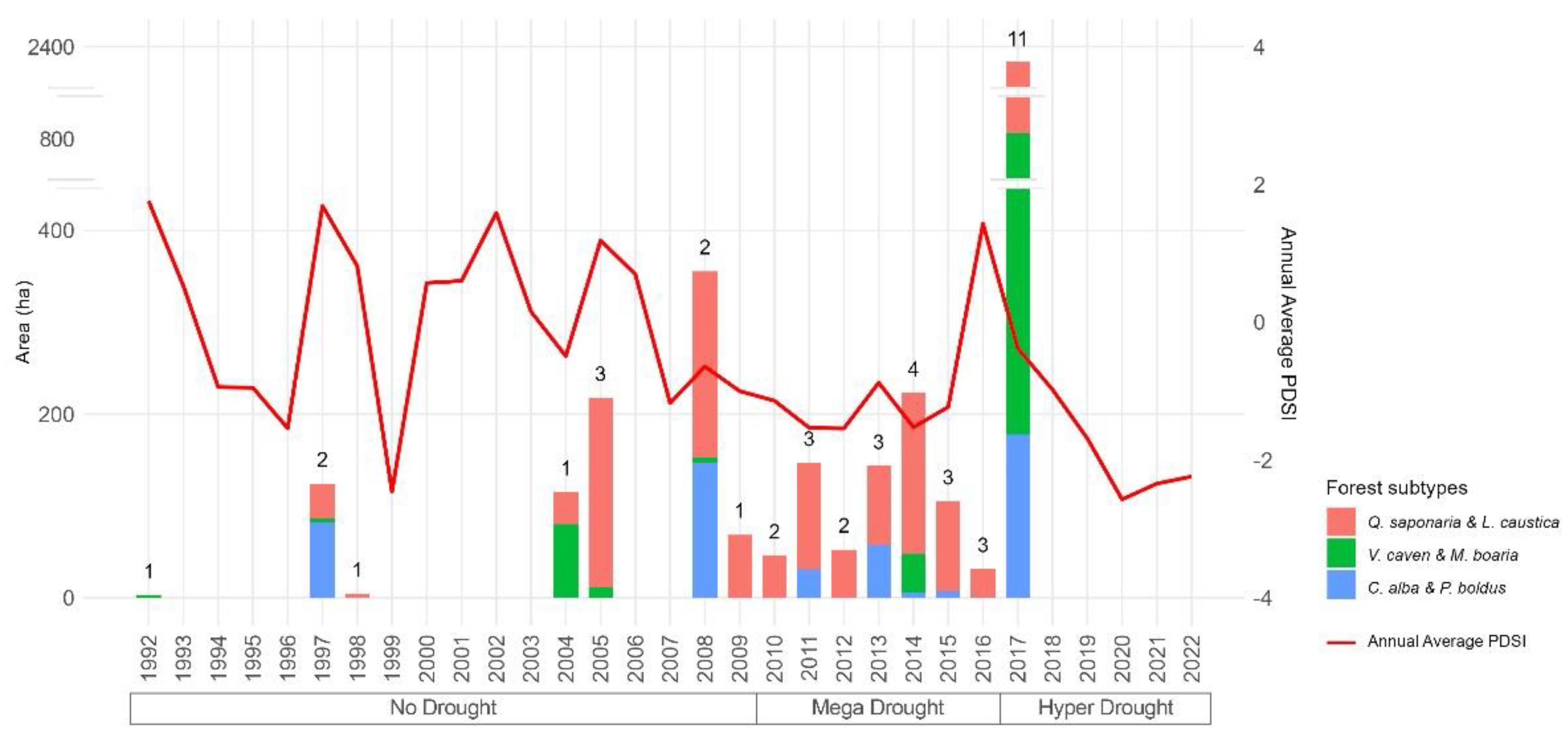
We used fire perimeter and severity data from a previous study in the area [47]. Forest spatial distribution and type were obtained from the official vegetation map provided by the National Forest Service [48,49,50]. From this data set, which for the four administrative regions of the study area included more than 5,533 wildfires, we applied a multi-criterion filtered by vegetation attributes considering natural forest, the main subtypes of Mediterranean sclerophyllous forest (Q. saponaria & L. caustica, V. caven & M. boaria, and C. alba & P. boldus), and dense canopy cover. The selection of dense canopy cover prevented the retrieval of a combined spectral response. We avoided the overlapping burned areas and ensured no change in land use (such as urban development, plantations, or agricultural activities) by evaluating five years before and after wildfires using the LandTrendr change detection algorithm (detailed later) and visually confirmed with Google Earth imagery. Consequently, 44,108 pixels in 42 wildfires from 1992 to 2017 met the vegetation, wildfire, and land use criteria (Figure 1 and Figure 2, and Table A1). The total number of burned pixels selected varied annually, with the largest being in 2017 due to the mega-fire season (Figure 2) [51].
Fire severity was classified based on the RdNBR value ranges proposed by Miller and Thode [52], with severity categories defined as low (69-315), medium (316-640), and high (≥ 641). We excluded low-severity pixels because there were fewer than 200 observations, which were concentrated in Q. saponaria & L. caustica. We categorize our analysis into three drought levels: No Drought (1992-2009) [21], the Mega Drought (2010-2016) [15,16], and the Hyper Drought (2017-2022), which represents the most severe stage of the prolonged drought, including 2019 and 2021, the driest and warmest years on record during this period [20] (Figure 2).
Figure 1.
Study Area. Locations of the wildfires analyzed in central Chile, occurring between 1992 and 2017, based on data from Miranda et al. [47] and vegetation maps from CIREN-CONAF and CONAF [48,49,50] .
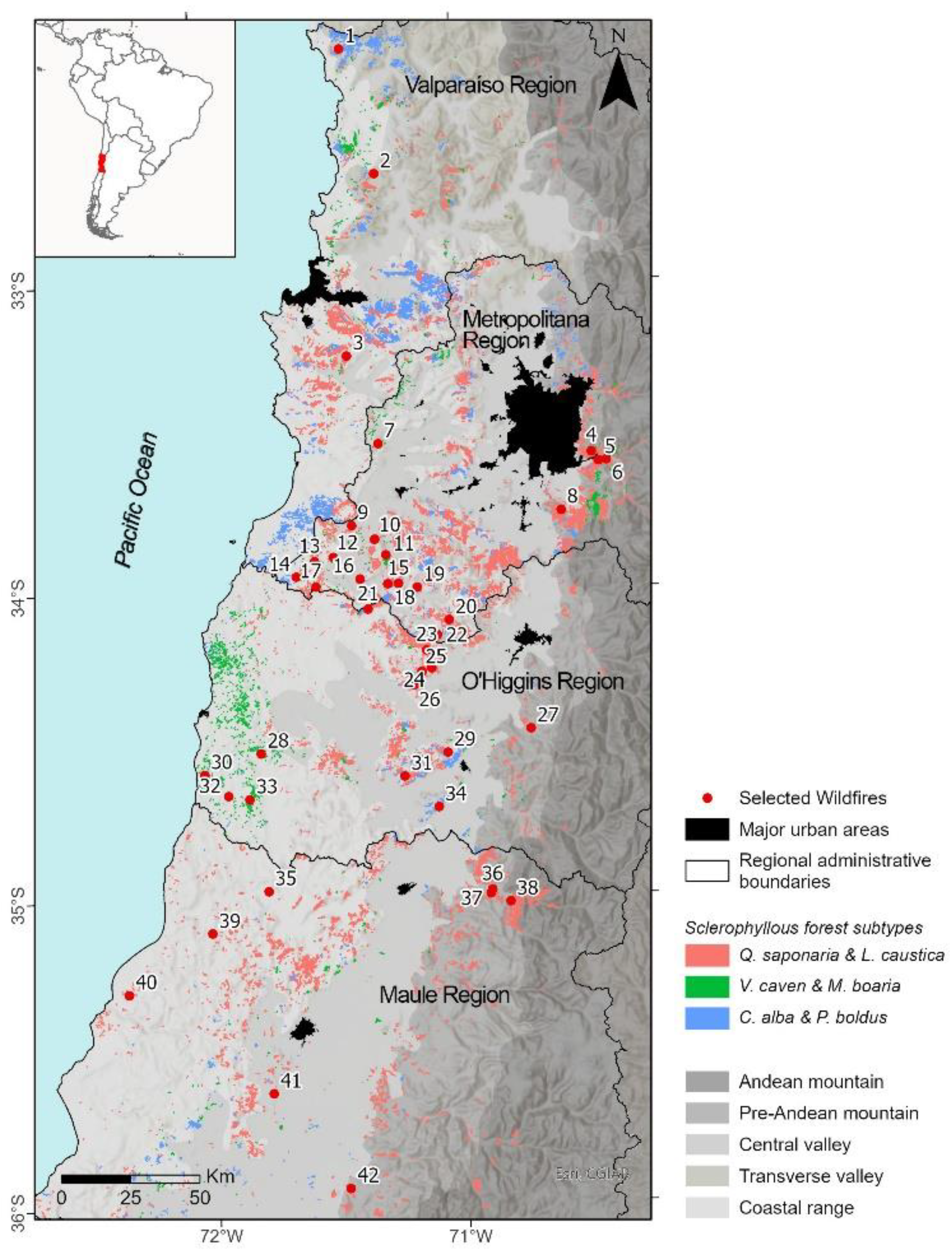
Figure 2.
Temporal distribution of burned areas analyzed covering the study area in Central Chile by forest subtype. The numbers above the bars indicate each year's selected wildfires. The red line represents the annual Palmer Drought Severity Index from TerraClimate data [53].
Figure 2.
Temporal distribution of burned areas analyzed covering the study area in Central Chile by forest subtype. The numbers above the bars indicate each year's selected wildfires. The red line represents the annual Palmer Drought Severity Index from TerraClimate data [53].
2.3. Landsat Time Series for Post-Fire Recovery
We implemented all our input data and processing in Google Earth Engine (GEE). GEE is an open cloud-computing platform tailored for geospatial analysis and offers access to a comprehensive public catalog comprising satellite imagery, topographical data, climate, and environmental datasets [54]. Prolonged periods and consistent satellite data were necessary to assess interannual changes. We used NDVI to characterize the trajectory and spectral response after the fire (post-fire). NDVI time series was built derived from the Landtrendr spectral-temporal segmentation algorithm [55]. This algorithm proposed by Kennedy et al. [35] adopts a singular perspective based on a pixel's spectral history. It undergoes a systematic procedure to discern breakpoints that delineate intervals of sustained alterations or constancy in spectral trajectories. The identified breakpoints, characterized by the year of occurrence and spectral index value, represent a pixel's spectral history as a sequence of vertices bounding line segments. The outcome is an idealized trajectory-based time series devoid of noise, where each observation is situated within the framework of a spectral-temporal trend. In this study, LandTrendr contributes to two methodological steps. The first is detecting land use changes for selecting burned areas, and the second is collecting NDVI time series without noise to evaluate the post-fire forest response. We opted for this adjusted data format instead of using unmodified surface reflectance to minimize the impact of minor fluctuations in time series data attributed to instabilities in climate, atmospheric conditions, phenology, solar angle, and the limitation of available images, on the determination of post-fire NDVI recovery.
The dataset included images that had already undergone atmospheric and geometrical corrections. We utilized all available TM/ETM+/OLI/OLI2 Level-2, Collection 2, Surface Reflectance Tier 1 images from Landsat 5, 7, 8, and 9, spanning the period from 1987 to 2022, focusing on the local summer season (December 21 - March 31). In instances where cloud-free pixels were limited, the season was extended to ensure a representative summer composition, extending up to one month beyond the typical summer season if necessary. This summer period was selected to minimize the presence of herbaceous vegetation from the wet season, thereby reducing potential confusion in post-wildfire forest recovery and concentrating on the regeneration of trees and shrubs. The result was a single, cloud-free, NDVI-band image per year, representing the median NDVI values for the summer season.
2.4. Post-Fire Recovery Percentage
To evaluate forest recovery, we calculated the relative change in NDVI for each year following the wildfire, up to year 5, between to pre-fire values (Equation 1, Figure A1). This calculation was repeated annually to create a five-year time series, using the recovery index proposed by Lloret et al. [56]. This metric accounts for the degree of recovery relative to the extent of damage experienced during a disturbance event. The usefulness of using this measure is rooted in the understanding that the ability to recover pre-disturbance performance levels is intricately linked to the degree of impact, specifically, the reduction in growth experienced during the disturbance. We explain this metric using our spectral index converted to a percentage, calculated as:
Where NDVIyi is the value of NDVI in the year of the post-fire observation. NDVIy1 is the value next to the disturbance at year 1 (we expect a lower value than pre-disturbance), and NDVIpre is the weighted NDVI value for the five years before the wildfire with greater weight given to the pre-fire years. For NDVIrec, values closer to 1 show greater recovery (1= full recovery), values close to zero show less recovery, and negative values show a decrease in recovery lower than the disturbance values. This value allows comparison of recovery between forest subtypes and wildfires. We select NDVI year one as a surrogate for post-fire vegetation conditions; this could represent both open-growing canopy recovery and the presence of a remnant or recovery of overstory tree [57,58].
2.5. Statistical Analysis of Recovery Differences in Drought Levels and Fire Severity
We examined the effects of drought and fire severity on the recovery of the three forest subtypes following fire disturbances and potential interactions between these factors. We conducted a two-way analysis of variance (ANOVA) to determine significant differences in post-fire vegetation recovery among drought levels and fire severity categories for each forest subtype and test for factor interaction. Factor interaction refers to the case where each factor not only exerts an effect on the response variable (described as main effects) but also may interact with the other factor to exert additional joint effects on the response variable (known as interaction effects). We utilized type III and II approaches for calculating sums of squares in two-way ANOVA, depending on the presence or absence of interactions between factors and the unbalance of the data [59]. When a significant difference was detected in the ANOVA, we conducted a post hoc analysis using Tukey's Honestly Significant Difference (HSD) test to identify specific differences between groups. If the interaction was significant, we explored which factor was significant at each level of the other factor. In cases where the interaction was not significant, we focused on examining the main effects. The assumptions of normality and homoscedasticity were evaluated and met before conducting the analyses.
We structured our analysis by categorizing the drought conditions into three distinct levels: No Drought (1992–2009), Mega Drought (2010–2016), and Hyper Drought (2017–2022). Additionally, we classified fire severity into two categories: moderate severity (316–640 RdNBR) and high severity (≥ 641 RdNBR). We ran a subsample selection to avoid slight differences being considered statistically significant due to the large sample sizes (Figure 2). For resampling, pixels were split into groups by combining drought level and fire severity. Within each group, pixels were drawn using stratified random sampling, ensuring samples were proportional to the strata (Table A3). All statistical analyses were performed in R software [60].
3. Results
The NDVI recovery five years post-fire exhibited differences across forest subtypes. In Q. saponaria & L. caustica, significant effects were identified for drought level (ANOVA: F = 203.518, df = 2, p < 2.2e-16), fire severity (ANOVA: F = 30.436, df = 1, p < 2.2e-16), and the interaction between these factors on NDVI recovery at year five (ANOVA: F = 3.263, df = 2, p = 0.03845). We observed the highest recovery under conditions of No Drought and High fire severity, while the lowest recovery occurred during Hyper Drought across both fire severity levels (Tukey's multiple comparison test, p < 0.01) (Figure 3, Table A4 and Table A5). Given the interaction between factors (Figure 3 and Figure A2), the significance of the interaction of fire severity for each drought level group was evaluated, showing significance for both No Drought and Mega Drought conditions (Tukey's multiple comparison test, confidence level 0.95, p < 0.01). For the V. caven & M. boaria, we noted significant effects for drought level (ANOVA: F = 173.88, df = 2, p < 2.2e-16), while fire severity did not show significant effects (ANOVA: F = 8.829, df = 1, p = 0.0030), nor did the interaction between these factors on NDVI recovery at year five (ANOVA: F = 0.2755, df = 2, p = 0.7592). The highest recovery rates were recorded under both No Drought and Mega Drought conditions, whereas the lowest recovery occurred under Hyper Drought at high severity (Tukey's multiple comparison test, p<0.01) (Figure 3). In the case of C. alba &P. boldus, significant effects were found for both drought level (ANOVA: F = 186.892, df = 2, p < 2.2e-16) and fire severity (ANOVA: F = 73.491, df = 1, p < 2.2e-16), along with their interaction (ANOVA: F = 15.102, df = 2, p = 3.312e-07). The highest recovery was observed under 'No Drought' conditions across all fire severity levels, with the lowest recovery noted during 'Hyper Drought' (Tukey's multiple comparison tests, p < 0.01). The interactions between drought and fire severity were found to be significant for each drought level, affecting NDVI recovery (Tukey's multiple comparison test, confidence level 0.95, p < 0.01) (Figure 3).
A descriptive analysis of the overall means indicated marked differences among forest subtypes (Figure 3 and Figure A3, Table A3), showed the highest recovery in high severity than medium severity. For high severity, C. alba & P. boldus achieved the highest recovery during No Drought levels, averaging approximately 100%. This was followed by Q. saponaria & L. caustica at 89.8% and V. caven & M. boaria at 63.0%. Under Mega Drought, recovery was higher for Q. saponaria & L. caustica (80.2%) and V. caven & M. boaria (65.6%), whereas for C. alba & P. boldus declined to 55.8%. As drought intensity increased to Hyper Drought, recovery converged to 39%, 30.5 and 27.9% for Q. saponaria & L. caustica, V. caven & M. boaria, and C. alba & P. boldus, respectively. For medium fire severity, variability among the three forest subtypes was reduced under No Drought and Mega Drought conditions (69.2% to 45.3%), with the lowest recovery rates observed under Hyper Drought (21.1% to 30.3%). A confidence interval is shown in Table A3.
When considering the No Drought period as a reference for post-fire vegetation recovery, the results reveal notably lower recovery percentages across all forest subtypes during the Hyper Drought level, regardless of fire severity. Hyper Drought, both in high and medium fire severity, shows a significant difference relative to No drought (Figure 4). C. alba & P. boldus exhibit significant differences relative to No drought and percentages lower relative recovery percentages compared to Q. saponaria & L. caustica and V. caven & M. boaria. Additionally, we found significant differences between Hyper Drought and Mega Drought for the three forest subtypes, both for high and medium fire severity (p < 0.01).
An analysis of the recovery trajectory from year one to year five revealed distinct differences based on drought level and fire severity. Wildfires occurring under the No Drought condition showed continuous recovery each year. In contrast, Mega Drought and Hyper Drought conditions exhibited interrupted recovery phases, particularly for Q. saponaria & L. caustica and V. caven & M. boaria. The Hyper Drought condition was notably associated with more pronounced disruptions in recovery, corresponding to the increased severity of drought and the forest browning and productivity decline observed in 2020 (Figure 6).
Figure 5.
The trajectory of mean NDVI recovery values from year 1 to year 5 for all forest subtypes, divided by burn severity. The background color represents the average of the Palmer Drought Severity Index (PDSI) for each relative year after the fire.
Figure 5.
The trajectory of mean NDVI recovery values from year 1 to year 5 for all forest subtypes, divided by burn severity. The background color represents the average of the Palmer Drought Severity Index (PDSI) for each relative year after the fire.
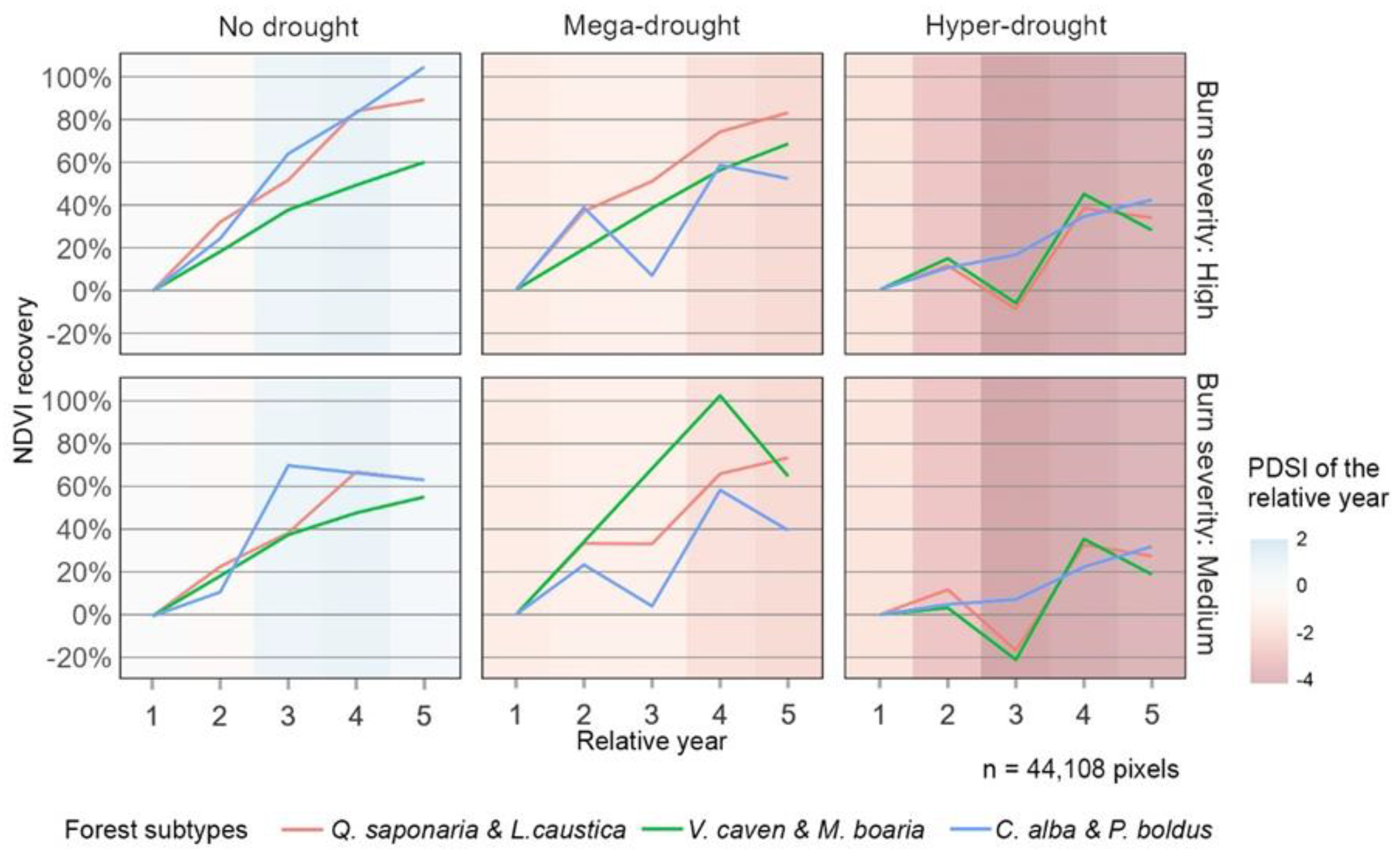
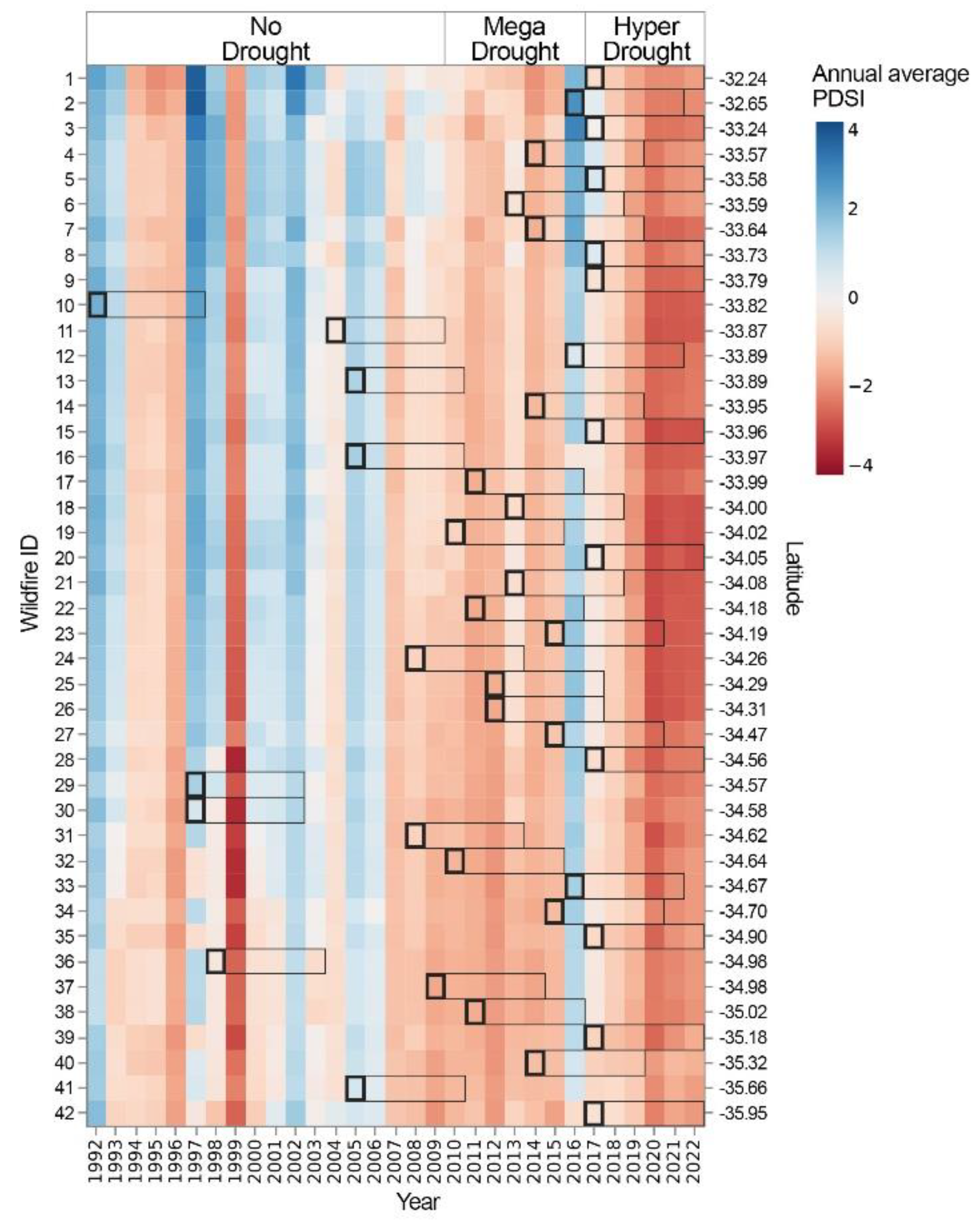
Figure 6.
Annual Palmer Drought Severity Index (PDSI) for each wildfire, derived from TerraClimate data [53]. The bold squares indicate the year of wildfire occurrence, and the rectangles represent the short-term recovery period from year 1 to year 5 post-wildfire. Wildfires are arranged by latitude, from north to south.
Figure 6.
Annual Palmer Drought Severity Index (PDSI) for each wildfire, derived from TerraClimate data [53]. The bold squares indicate the year of wildfire occurrence, and the rectangles represent the short-term recovery period from year 1 to year 5 post-wildfire. Wildfires are arranged by latitude, from north to south.
4. Discussion
Understanding ecosystem responses to environmental variables is essential for effective conservation management and planning [61,62]. In the case of Mediterranean vegetation ecosystems, evidence suggests their evolution is closely tied to a climatic history of increasing aridity throughout the Cenozoic [27]. This trend has been accompanied by persistent fire disturbances, resulting in a range of fire-adapted tolerance and reproduction mechanisms associated with fire ecology [63]. However, Chile represents an exception, where localized regional climatic changes linked to the Andean uplift during the Miocene led to a suppression of fires compared to other Mediterranean-climate regions [27,63]. Therefore, examining fire tolerance responses in the flora of central Chile provides a unique opportunity to understand the adaptive capacity of an ecosystem that evolved under the stress of climate-induced drought but now faces increasing wildfire pressure [64]. This analysis can help predict potential ecological trajectories and alternative states for this ecosystem [65].
Our analysis revealed statistically significant differences in short-term post-fire recovery based on drought levels, fire severity, and interaction effects (Figure 3). We observed a notable disparity in recovery across various forest subtypes, underscoring their distinct resilience capacities. Specifically, C. alba & P. boldus, which exhibited higher average recovery percentages under non-drought conditions, demonstrated increased vulnerability to moderate drought levels, such as the Mega Drought. In contrast, Q. saponaria & L. caustica, along with V. caven & M. boaria, showed greater resilience. However, under extreme and prolonged drought conditions, all forest subtypes experienced a significant decline in recovery, with mean recovery percentages plummeting from 39% to 21.1% during Hyper Drought (Figure 3 and Figure A3). These findings suggest that, regardless of forest subtype or fire severity (which appeared to have a lesser effect than drought) a critical threshold of drought severity substantially compromised forest recovery.
In other Mediterranean regions, such as Southern California and the Mediterranean Basin, severity and extended drought periods, especially in the early state post-fire, can significantly reduce soil moisture content, hindering seed germination and seedling establishment in vegetation [66,67,68]. This diminished water availability limits the capacity of Mediterranean plant species to initiate the regrowth process after a fire event, thereby impeding post-fire recovery[69] (Rossetti et al., 2022). Rodriguez-Blanco et al. [67] report that the effect of drought duration on post-fire recovery vegetation varied depending on the aridity level, intensifying at extremes of aridity gradient (for semi-arid and humid areas) in the western Mediterranean Basins. In arid environments, the adverse effect of drought on recovery aligns with contemporary findings suggesting that adaptations to drought (for example, embolism resistance) may act antagonistically to the post-fire recovery process [70].
The higher recovery percentages observed in Q. Saponaria & L. caustica and V. caven & M. boaria during the Mega Drought likely resulted from their superior adaptation to lower water availability compared to C. alba & P. boldus, which are typically found in more humid areas [17]. Additionally, Q. saponaria & L. caustica, particularly those in northern exposures, were often associated with xeric habitat species adapted to drought conditions. L. caustica exhibited one of the highest tolerances to water stress when compared to Q. saponaria and C. alba [71], facilitating its establishment and growth in drought-prone environments, especially during the early successional stages evaluated in this study. In this context, L. caustica may play a significant role in the recovery of sclerophyllous forests [72].
The analysis of post-fire NDVI recovery trajectories over the five years revealed significant differences based on drought levels and fire severity (Figure 5). Forests experiencing No Drought conditions demonstrated consistent and positive recovery across all forest subtypes, indicating strong resilience. Supporting field observations [73], suggested that initial increases in NDVI were primarily driven by resprouting rather than new seedling growth, particularly under No Drought conditions. However, recovery was notably disrupted during the Mega and Hyper Drought periods, with setbacks observed across every forest subtype. Consistent with the recovery patterns observed at year five, the NDVIrec trajectories during the Mega Drought showed that C. alba & P. boldus and Q. saponaria & L. caustica experienced a considerable decline in recovery by years three and five. This decline was particularly abrupt for C. alba & P. boldus, whose recovery percentages returned to similar levels in year one. These findings support that, for these forest subtypes, more severe drought conditions—evidenced by the continuity of drought—significantly impacted post-fire recovery. In Hyper Drought conditions, these setbacks became even more pronounced for Q. Saponaria & L. caustica and V. caven & M. boaria, with recovery values reaching negative levels by year three (-20%). This decline was related to the persistence of drought conditions and an increase in drought severity, as indicated by more negative precipitation anomalies across all drought levels [13,19].
As an alternative interpretation, the observed drop in NDVI recovery during the third year could be indicative of an ecological succession event. This trend might reflect a shift in community structure, where the decline in biomass at year three is more likely associated with the senescence or replacement of herbaceous vegetation rather than a loss in resprouting shrubs or trees. Consequently, this pattern may signal a transition from herbaceous-dominated to shrub or tree-dominated cover types, potentially driven by climatic events (e.g., dry episodes) that favor such successional dynamics. Similar patterns have been documented in other Mediterranean ecosystems, such as those in California [74,75]. Following this period of decline, a marked recovery was observed, which again fell by year five. This aligns with the NDVI trajectories observed, where rapid vegetative responses likely drove initial recovery under No Drought conditions. In contrast, drought conditions severely hampered these processes, resulting in less recovery (Figure 5).
Our observations revealed that only C. alba & P. boldus achieved an average recovery rate of 100% under No Drought conditions and high fire severity. In contrast, V. caven and &. boaria displayed the lowest recovery rate at 63.0% under the same conditions. The slower recovery observed in V. caven & M. boaria suggests that these species face challenges in regenerating and restoring pre-fire conditions in the short term, reflecting their lower resilience than other subtypes. This slower biomass recovery, coupled with a less complex structural response, indicates that after a fire, V. caven & M. boaria may take longer to re-establish, potentially allowing for the invasion of exotic species or loss of vegetation cover, which could negatively affect the overall recovery [76,77]. Despite being a species more adapted to extreme drought conditions, V. caven shows limited post-fire recovery capacity in short terms, hindering its regeneration.
Previous studies in the Mediterranean region of Central Chile have reported long-term post-fire coverage recovery of dense vegetation by remote sensing, spanning approximately 10 years [46]. However, these studies often included mixed observations of forest, shrubland, and forest plantation, and were based on vegetation coverage rather than photosynthetic productivity. In contrast, our study has demonstrated significant differences in recovery between forest subtypes. Similar or slower recovery rates have been reported in other Mediterranean regions through studies utilizing remote sensing-based vegetation indices [67,78,79,80,81,82] (Table A6). These findings highlight the importance of considering forest subtype-specific responses and the use of photosynthetic productivity metrics in assessing post-fire recovery dynamics.
Consistent with the findings of Meng et al. [82] and Bright [83], our analysis revealed that areas subjected to higher fire severities tended to recover at a faster rate than those experiencing lower severities (Figure 3, Figure 4, and Figure 5). This phenomenon can be attributed to the fire-adapted characteristics of Mediterranean and temperate forest types, where fire events create conditions that promote vegetation germination and regeneration [84,85]. Mechanisms such as lignotubers, which are located underground, and epicormic buds, which are latent buds found on the trunk or main branches of the plant, can activate to sprout when the plant has lost its foliage due to fire [31,34]. Field observations within the study area further supported these results, demonstrating rapid vegetation regrowth following fire, even in the face of high fire intensity and extensive damage [73]. These findings underscore the resilience of the natural Mediterranean forest to wildfires and highlight the importance of understanding the adaptive mechanisms that facilitate recovery in these ecosystems. were particularly significant for Q. saponaria & L. caustica and C. alba & P. boldus under No Drought conditions, showing substantial differences in recovery based on fire severity. In these Mediterranean ecosystems, once drought severity exceeds certain thresholds, fire severity becomes a less influential factor, likely due to the increased biomass availability prior to the fire event. Overall, the results emphasize the intricate interplay between fire severity, drought intensity, and forest type in shaping post-fire recovery dynamics.
Our analysis did not determine whether post-fire changes occurred in the structure and composition of the forests. We assumed that tree and shrub regeneration in the short term primarily occurred through resprouting [73,86] (Figure 7). Generally, studies measuring photosynthetic activity and post-disturbance forest productivity often lack field validations that explore the composition and structure of the forest. This omission may result from the high costs associated with analyzing large areas [16,17,87,88]. This limitation can lead to confusion in interpreting results, particularly given the differences in plant community composition [89].
The influence of the Pacific Ocean affects species composition variation in central Chile. Coastal regions experience more restricted temperature ranges and frequent morning fogs, which alleviate plant stress during summer droughts. As a result, these conditions lead to a species composition that is distinct from that of pre-Andean forests, particularly up to a certain drought severity threshold [90,91], as observed for C. alba and P. boldus.
Another criterion that has been attributed to the calculation of the recovery index was the selection of NDVI year 1 over immediate post-fire (year 0) NDVI, which was driven by the potential for continued declines in NDVI during the year following fire disturbance. Post-wildfire vegetation may gradually decline in the post-fire period despite surviving direct burn [69,92,93]. Substantial shifts in species composition may occur, contributing to variations in NDVI during the post-fire period [80,94]. Additionally, alterations in surface reflectance induced by leaching burned remains can further contribute to post-fire changes in NDVI [82,95].
Remote sensing tools, particularly satellite-derived spectral indices, have increasingly proven essential for assessing post-fire vegetation recovery [42]. In Mediterranean ecosystems like central Chile, on-the-ground monitoring is limited [17,86]. By selected NDVI as our primary metric due to its proven applicability in Mediterranean regions [40,80] and its effectiveness in capturing disturbances like drought [17,20], we provided valuable insights into vegetation conditions based on changes in cover density and photosynthetic activity following wildfire. However, we acknowledged that NDVI cannot detail vegetation composition and structure, highlighting the challenge of integrating field data with remote sensing for a comprehensive understanding of complex ecosystems [67,82]. Our focus on NDVI aimed to establish a foundation for future investigations that could utilize advanced remote-sensing technologies to explore specific vegetation characteristics, such as LiDAR or airborne laser scanning [96,97,98]. Recovery metrics based on remote sensing, such as NDVIrec, are particularly relevant for planning and managing conservation areas, as they inform adaptive management practices by identifying regions where post-disturbance recovery is slow or incomplete [99,100].
This study encountered unique challenges compared to other Mediterranean regions, concerning the fragmentation of natural forests in central Chile. Forest remnants' presence limited our analysis's spatial scale [17,101,102].This fragmentation resulted in a reduced number of pixels in areas with dense vegetation coverage, potentially affecting the accuracy of our analyses. Moreover, the precision of our outcomes was influenced by the inherent limitations of the official vegetation map products, which reported an overall accuracy ranging between 75% and 83% for the most recent vegetation map of the study regions [50]. However, the advent of next-generation fire remote-sensing products and model methodologies based on machine learning and artificial intelligence holds promise for enhancing the quality and efficacy of post-fire recovery studies [97,103].
Despite the projected changes in disturbance regimes due to climate change, research on post-fire vegetation recovery in the context of drought within the Chilean Mediterranean ecosystem remains scarce. To our knowledge, few studies have addressed the interaction between these factors and their effects on recovery. The results of such studies are highly dependent on climate conditions, site-specific characteristics, and the analytical methodologies employed, complicating interpretation. Understanding the thresholds of forest resilience and resistance to climate extremes, such as droughts, is crucial for forest conservation and ecological restoration efforts [100]. Extensive research has highlighted the profound impacts of climate change on forest structure, disturbance patterns, and carbon storage, particularly in Mediterranean regions [104,105,106]. This study contributes valuable insights into the complex interplay between drought conditions and fire, aiming to quantify their significance within distinct forest subtypes. Given the changes in fire regimes, post-disturbance ecosystem trajectories, and the increasing aridity associated with climate change—especially in the southern Mediterranean regions [20,107,108] —our findings underscore the critical role of post-fire climate in vegetation recovery. These factors necessitate further investigation to fully understand their implications. The results suggest that the future resilience of Mediterranean forests may be increasingly challenged by the rising frequency of extreme drought periods projected for this region.
5. Conclusions
The interaction between drought and wildfire presents significant challenges for the recovery of Mediterranean forest ecosystems, particularly in central Chile, where the frequency and intensity of these disturbances have been exacerbated in the last decades. This study highlights the urgent need for comprehensive monitoring of forest recovery dynamics in response to such disturbances. Our findings indicate that drought severity is pivotal in determining post-fire recovery rates, especially in forest subtypes C. alba & P. boldus and V. caven & M. boaria, which are more sensitive to the driest conditions. The observed differences in recovery rates among forest subtypes under varying drought conditions emphasize the necessity for further investigation into the factors that influence post-fire recovery, including species-specific traits and ecological interactions. Understanding these dynamics is essential for forecasting future forest resilience in the face of climate change, particularly as drought frequency and intensity are expected to increase.
The insights gained from this study not only enhance our understanding of Mediterranean forest recovery but also serve as a valuable resource for policymakers and land managers aiming to mitigate the impacts of climate change on these ecosystems.
Author Contributions
Conceptualization, A.H.-D., F.S., A.M., J.S.; methodology, A.H.-D.; F.S., A.M.; software, A.H.-D., E.G.; validation, A.H.-D.; formal analysis, A.H.-D and F.S.; investigation, A.H.-D., F.S., E.G.; resources, M.S.-V; data curation, A.H.-D., E.G.; writing—original draft preparation, A.H.-D.; writing—review and editing, A.H.-D., F.S., A.M., J.F., J.S., M.S.-V.; visualization, A.H.-D., F.S., J.F.; E.G.; supervision, F.S.; funding acquisition, M.S.-V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by ANID grant number ACT210080, ANID project ID22i10210 and Escuela de Postgrado Universidad de Playa Ancha, Chile. A.M. acknowledges ANID/Fondecyt Iniciación/2024 – 11240356, ANID-FONDAP N°15110009 and ANID project IT23I0109 funding. The European Unions’s Horizon 2020 research and innovation programme under the Marie Sklodowska-Curie grant agreement No 101007950 and Dirección Investigación de la Universidad de La Frontera.
Data Availability Statement
The datasets used and analyzed during the current study are available from the corresponding author upon reasonable request.
Acknowledgments
A.H-D- thanks, Dr. José González, for the statistical discussion. Pamela Ramírez and Rodrigo Villaseñor for botanical and biogeographic contributions.
Conflicts of Interest
The authors declare no conflicts of interest.
Appendix A
Figure A1, Figure A2, Table A1, Table A2, Table A3, Table A4, Table A5, Table A6.
References
- Turner, M.G. Disturbance and landscape dynamics in a changing world. Ecology 2010, 91, 2833–2849. [Google Scholar] [CrossRef] [PubMed]
- White, P.S.; Pickett, S.T.A. The Ecology of Natural Disturbance and Patch Dynamics; Academic Press: 1985.
- Scheffer, M.; Carpenter, S.; Foley, J.A.; Folke, C.; Walker, B. Catastrophic shifts in ecosystems. Nature 2001, 413, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Seidl, R.; Rammer, W.; Spies, T.A. Disturbance legacies increase the resilience of forest ecosystem structure, composition, and functioning. Ecological Applications 2014, 24, 2063–2077. [Google Scholar] [CrossRef] [PubMed]
- Frenne, D.P.; Roíguez-Sánchez, F.; Coomes, D.; Baeten, L.; Verstraeten, G.; Hommel, P.W.F.M. Microclimate moderates plant responses to macroclimate warming. Proceedings of the National Academy of Sciences - PNAS 2013, 110, 18561–18565. [Google Scholar] [CrossRef] [PubMed]
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J.; et al. Forest disturbances under climate change. Nature Climate Change 2017, 7, 395–402. [Google Scholar] [CrossRef]
- Thom, D.; Rammer, W.; Seidl, R. Disturbances catalyze the adaptation of forest ecosystems to changing climate conditions. Global change biology 2017, 23, 269–282. [Google Scholar] [CrossRef]
- Rillig, M.C.; van der Heijden, M.G.A.; Berdugo, M.; Liu, Y.-R.; Riedo, J.; Sanz-Lazaro, C.; Moreno-Jiménez, E.; Romero, F.; Tedersoo, L.; Delgado-Baquerizo, M. Increasing the number of stressors reduces soil ecosystem services worldwide. Nature Climate Change 2023, 13, 478–483. [Google Scholar] [CrossRef]
- Bendall, E.R.; Bedward, M.; Boer, M.; Clarke, H.; Collins, L.; Leigh, A.; Bradstock, R.A. Changes in the resilience of resprouting juvenile tree populations in temperate forests due to coupled severe drought and fire. Plant Ecology 2022, 223, 907–923. [Google Scholar] [CrossRef]
- Turco, M.; von Hardenberg, J.; AghaKouchak, A.; Llasat, M.C.; Provenzale, A.; Trigo, R.M. On the key role of droughts in the dynamics of summer fires in Mediterranean Europe. Scientific Reports 2017, 7, 81. [Google Scholar] [CrossRef]
- Thom, D. Natural disturbances as drivers of tipping points in forest ecosystems under climate change – implications for adaptive management. Forestry: An International Journal of Forest Research 2023, 96, 305–315. [Google Scholar] [CrossRef]
- Garreaud, R.D.; Alvarez-Garreton, C.; Barichivich, J.; Boisier, J.P.; Christie, D.; Galleguillos, M.; LeQuesne, C.; McPhee, J.; Zambrano-Bigiarini, M. The 2010–2015 megadrought in central Chile: Impacts on regional hydroclimate and vegetation. Hydrol. Earth Syst. Sci. 2017, 21, 6307–6327. [Google Scholar] [CrossRef]
- Carrasco-Escaff, T.; Garreaud, R.; Bozkurt, D.; Jacques-Coper, M.; Pauchard, A. The key role of extreme weather and climate change in the occurrence of exceptional fire seasons in south-central Chile. Weather and Climate Extremes 2024, 45, 100716. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853. [Google Scholar] [CrossRef] [PubMed]
- Boisier, J.P.; Rondanelli, R.; Garreaud, R.D.; Muñoz, F. Anthropogenic and natural contributions to the Southeast Pacific precipitation decline and recent megadrought in central Chile. Geophysical research letters 2016, 43, 413–421. [Google Scholar] [CrossRef]
- Garreaud, R.D.; Boisier, J.P.; Rondanelli, R.; Montecinos, A.; Sepúlveda, H.H.; Veloso-Aguila, D. The Central Chile Mega Drought (2010–2018): A climate dynamics perspective. International Journal of Climatology 2020, 40, 421–439. [Google Scholar] [CrossRef]
- Miranda, A.; Lara, A.; Altamirano, A.; Di Bella, C.; González, M.E.; Julio Camarero, J. Forest browning trends in response to drought in a highly threatened mediterranean landscape of South America. Ecological Indicators 2020, 115, 106401. [Google Scholar] [CrossRef]
- Alvarez-Garreton, C.; Boisier, J.P.; Garreaud, R.; Seibert, J.; Vis, M. Progressive water deficits during multiyear droughts in basins with long hydrological memory in Chile. Hydrol. Earth Syst. Sci. 2021, 25, 429–446. [Google Scholar] [CrossRef]
- Arroyo, M.T.K.; Robles, V.; Tamburrino, Í.; Martínez-Harms, J.; Garreaud, R.D.; Jara-Arancio, P.; Pliscoff, P.; Copier, A.; Arenas, J.; Keymer, J.; et al. Extreme Drought Affects Visitation and Seed Set in a Plant Species in the Central Chilean Andes Heavily Dependent on Hummingbird Pollination. Plants 2020, 9, 1553. [Google Scholar] [CrossRef]
- Miranda, A.; Syphard, A.D.; Berdugo, M.; Carrasco, J.; Gómez-González, S.; Ovalle, J.F.; Delpiano, C.A.; Vargas, S.; Squeo, F.A.; Miranda, M.D.; et al. Widespread synchronous decline of Mediterranean-type forest driven by accelerated aridity. Nature Plants 2023, 9, 1810–1817. [Google Scholar] [CrossRef]
- González, M.E.; Gómez-González, S.; Lara, A.; Garreaud, R.; Díaz-Hormazábal, I. The 2010–2015 Megadrought and its influence on the fire regime in central and south-central Chile. Ecosphere 2018, 9, e02300. [Google Scholar] [CrossRef]
- Holz, A.; Kitzberger, T.; Paritsis, J.; Veblen, T.T. Ecological and climatic controls of modern wildfire activity patterns across southwestern South America. Ecosphere 2012, 3, art103. [Google Scholar] [CrossRef]
- Urrutia-Jalabert, R.; González, M.E.; González-Reyes, Á.; Lara, A.; Garreaud, R. Climate variability and forest fires in central and south-central Chile. Ecosphere 2018, 9, e02171. [Google Scholar] [CrossRef]
- Donoso, C. Reseña Ecológica de los Bosques Mediterráneos de Chile. BOSQUE 1982, 4, 117–146. [Google Scholar] [CrossRef]
- Luebert, F.; Pliscoff, P. Sinopsis bioclimática y vegetacional de Chile 2ed.; Editorial Universitaria: Santiago, Chile, 2017. [Google Scholar]
- Rodriguez, R.; Marticorena, C.; Alarcón, D.; Baeza, C.; Cavieres, L.; Finot, V.L.; Fuentes, N.; Kiessling, A.; Mihoc, M.; Pauchard, A.; et al. Catálogo de las plantas vasculares de Chile. Gayana. Botánica 2018, 75, 1–430. [Google Scholar] [CrossRef]
- Rundel, P.W.; Arroyo, M.T.K.; Cowling, R.M.; Keeley, J.E.; Lamont, B.B.; Vargas, P. Mediterranean Biomes: Evolution of Their Vegetation, Floras, and Climate. Annual Review of Ecology, Evolution, and Systematics 2016, 47, 383–407. [Google Scholar] [CrossRef]
- Ganteaume, A.; Camia, A.; Jappiot, M.; San-Miguel-Ayanz, J.; Long-Fournel, M.; Lampin, C. A Review of the Main Driving Factors of Forest Fire Ignition Over Europe. Environmental Management 2013, 51, 651–662. [Google Scholar] [CrossRef]
- Armesto, J.J.; Bustamante-Sánchez, M.; Díaz, M.F.; González, M.E.; Holz, A.; Nuñez-Avila, M.; Smith-Ramírez, C. Fire disturbance regimes, ecosystem recovery and restoration strategies in Mediterranean and temperate regions of Chile. In Fire effects on soils and restoration strategies; CRC Press: 2009; pp. 553–584.
- Becerra, P.; Smith-Ramirez, C.; Arellano, E. Evaluación de técnicas pasivas y activas pra la recuperación del bosque esclerófilo de Chile Central Corporación Nacional Forestal Imprenta Edición, Santiago, Chile, 2018.
- Montenegro, G.; Ginocchio, R.; Segura, A.; Keely, J.E.; Gómez, M. Fire regimes and vegetation responses in two Mediterranean-climate regions. Revista Chilena de Historia Natural 2004, 77, 455–464. [Google Scholar] [CrossRef]
- Gómez-González, S.; Cavieres, L.A. Litter burning does not equally affect seedling emergence of native and alien species of the Mediterranean-type Chilean matorral. International Journal of Wildland Fire 2009, 18, 213–221. [Google Scholar] [CrossRef]
- Muñoz, M.R.; Fuentes, E.R. Does Fire Induce Shrub Germination in the Chilean Matorral? Oikos 1989, 56, 177–181. [Google Scholar] [CrossRef]
- Nolan, R.H.; Collins, L.; Leigh, A.; Ooi, M.K.J.; Curran, T.J.; Fairman, T.A.; Resco de Dios, V.; Bradstock, R. Limits to post-fire vegetation recovery under climate change. Plant, Cell & Environment 2021, 44, 3471–3489. [Google Scholar] [CrossRef]
- Kennedy, R.E.; Yang, Z.; Cohen, W.B. Detecting trends in forest disturbance and recovery using yearly Landsat time series: 1. LandTrendr—Temporal segmentation algorithms. Remote Sensing of Environment 2010, 114, 2897–2910. [Google Scholar] [CrossRef]
- Verbesselt, J.; Hyndman, R.; Newnham, G.; Culvenor, D. Detecting trend and seasonal changes in satellite image time series. Remote Sensing of Environment 2010, 114, 106–115. [Google Scholar] [CrossRef]
- Viana-Soto, A.; García, M.; Aguado, I.; Salas, J. Assessing post-fire forest structure recovery by combining LiDAR data and Landsat time series in Mediterranean pine forests. International Journal of Applied Earth Observation and Geoinformation 2022, 108, 102754. [Google Scholar] [CrossRef]
- Wang, Z.; Wei, C.; Liu, X.; Zhu, L.; Yang, Q.; Wang, Q.; Zhang, Q.; Meng, Y. Object-based change detection for vegetation disturbance and recovery using Landsat time series. GIScience & Remote Sensing 2022, 59, 1706–1721. [Google Scholar] [CrossRef]
- Veraverbeke, S.; Gitas, I.; Katagis, T.; Polychronaki, A.; Somers, B.; Goossens, R. Assessing post-fire vegetation recovery using red–near infrared vegetation indices: Accounting for background and vegetation variability. ISPRS Journal of Photogrammetry and Remote Sensing 2012, 68, 28–39. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Pérez-Cabello, F.; Lasanta, T. Pinus halepensis regeneration after a wildfire in a semiarid environment: Assessment using multitemporal Landsat images. International Journal of Wildland Fire 2011, 20, 195–208. [Google Scholar] [CrossRef]
- Pettorelli, N.; Vik, J.O.; Mysterud, A.; Gaillard, J.-M.; Tucker, C.J.; Stenseth, N.C. Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends in Ecology & Evolution 2005, 20, 503–510. [Google Scholar] [CrossRef]
- Xu, H.; Chen, H.W.; Chen, D.; Wang, Y.; Yue, X.; He, B.; Guo, L.; Yuan, W.; Zhong, Z.; Huang, L.; et al. Global patterns and drivers of post-fire vegetation productivity recovery. Nature Geoscience 2024. [Google Scholar] [CrossRef]
- Bozkurt, D.; Rojas, M.; Boisier, J.P.; Valdivieso, J. Projected hydroclimate changes over Andean basins in central Chile from downscaled CMIP5 models under the low and high emission scenarios. Climatic Change 2018, 150, 131–147. [Google Scholar] [CrossRef]
- Garreaud, R.D.; Clem, K.; Veloso, J.V. The South Pacific Pressure Trend Dipole and the Southern Blob. Journal of Climate 2021, 34, 7661–7676. [Google Scholar] [CrossRef]
- Chávez, R.O.; Castillo-Soto, M.E.; Traipe, K.; Olea, M.; Lastra, J.A.; Quiñones, T. A Probabilistic Multi-Source Remote Sensing Approach to Evaluate Extreme Precursory Drought Conditions of a Wildfire Event in Central Chile. Frontiers in Environmental Science 2022, 10, 865406. [Google Scholar] [CrossRef]
- Smith-Ramírez, C.; Castillo-Mandujano, J.; Becerra, P.; Sandoval, N.; Fuentes, R.; Allende, R.; Paz Acuña, M. Combining remote sensing and field data to assess recovery of the Chilean Mediterranean vegetation after fire: Effect of time elapsed and burn severity. Forest Ecology and Management 2022, 503, 119800. [Google Scholar] [CrossRef]
- Miranda, A.; Mentler, R.; Moletto-Lobos, Í.; Alfaro, G.; Aliaga, L.; Balbontín, D.; Barraza, M.; Baumbach, S.; Calderón, P.; Cárdenas, F.; et al. Historical Fire Scar Database - 3. Satellital images after fires. 2022. [Google Scholar]
- CIREN-CONAF. Informe técnico final proyecto: Monitoreo de cambios, corrección cartográfica y actualización del catastro de bosque nativo en las regiones de Valparaíso, Metropolitana y Libertador Bernardo O’Higgins. 2013, 130.
- CIREN-CONAF. Informe técnico final proyecto: Monitoreo de cambios, corrección cartográfica y actualización del catastro de bosque nativo de la región del Maule. 2016, 90.
- CONAF, C.N.F. Monitoreo de Cambios, Corrección Gráfica y Actualización del Catastro de los Recursos Vegetacionales de la Región de Valparaíso, año 2019; Santiago, Chile, 2022; p. 70.
- Bowman, D.M.J.S.; Moreira-Muñoz, A.; Kolden, C.A.; Chávez, R.O.; Muñoz, A.A.; Salinas, F.; González-Reyes, Á.; Rocco, R.; de la Barrera, F.; Williamson, G.J.; et al. Human–environmental drivers and impacts of the globally extreme 2017 Chilean fires. Ambio 2019, 48, 350–362. [Google Scholar] [CrossRef]
- Miller, J.D.; Thode, A.E. Quantifying burn severity in a heterogeneous landscape with a relative version of the delta Normalized Burn Ratio (dNBR). Remote sensing of environment 2007, 109, 66–80. [Google Scholar] [CrossRef]
- Abatzoglou, J.T.; Dobrowski, S.Z.; Parks, S.A.; Hegewisch, K.C. TerraClimate, a high-resolution global dataset of monthly climate and climatic water balance from 1958–2015. Scientific Data 2018, 5, 170191. [Google Scholar] [CrossRef]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sensing of Environment 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Kennedy, R.E.; Yang, Z.; Gorelick, N.; Braaten, J.; Cavalcante, L.; Cohen, W.B.; Healey, S. Implementation of the LandTrendr Algorithm on Google Earth Engine. Remote Sensing 2018, 10, 691. [Google Scholar] [CrossRef]
- Lloret, F.; Keeling, E.G.; Sala, A. Components of tree resilience: Effects of successive low-growth episodes in old ponderosa pine forests. Oikos 2011, 120, 1909–1920. [Google Scholar] [CrossRef]
- Collins, B.M.; Roller, G.B. Early forest dynamics in stand-replacing fire patches in the northern Sierra Nevada, California, USA. Landscape Ecology 2013, 28, 1801–1813. [Google Scholar] [CrossRef]
- Donato, D.C.; Fontaine, J.B.; Robinson, W.D.; Kauffman, J.B.; Law, B.E. Vegetation response to a short interval between high-severity wildfires in a mixed-evergreen forest. Journal of Ecology 2009, 97, 142–154. [Google Scholar] [CrossRef]
- Langsrud, Ø. ANOVA for unbalanced data: Use Type II instead of Type III sums of squares. Statistics and Computing 2003, 13, 163–167. [Google Scholar] [CrossRef]
- Team, R.C. R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing, 2020. [Google Scholar]
- IPBES. Global assessment report on biodiversity and ecosystem services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services (Version 1). Zenodo; 978-3-947851-20-1; 2019.
- Jolly, W.M.; Cochrane, M.A.; Freeborn, P.H.; Holden, Z.A.; Brown, T.J.; Williamson, G.J.; Bowman, D.M.J.S. Climate-induced variations in global wildfire danger from 1979 to 2013. Nature Communications 2015, 6, 7537. [Google Scholar] [CrossRef] [PubMed]
- Rundel, P.W.; Arroyo, M.T.K.; Cowling, R.M.; Keeley, J.E.; Lamont, B.B.; Pausas, J.G.; Vargas, P. Fire and Plant Diversification in Mediterranean-Climate Regions. Frontiers in Plant Science 2018, 9, 00851. [Google Scholar] [CrossRef] [PubMed]
- Keeley, J.E.; Bond, W.J.; Bradstock, R.A.; Pausas, J.G.; Rundel, P.W. Fire in Mediterranean ecosystems: Ecology, evolution and management; Cambridge University Press: 2011.
- Falk, D.A.; van Mantgem, P.J.; Keeley, J.E.; Gregg, R.M.; Guiterman, C.H.; Tepley, A.J.; Jn Young, D.; Marshall, L.A. Mechanisms of forest resilience. Forest Ecology and Management 2022, 512, 120129. [Google Scholar] [CrossRef]
- Dong, C.; MacDonald, G.; Okin, G.S.; Gillespie, T.W. Quantifying Drought Sensitivity of Mediterranean Climate Vegetation to Recent Warming: A Case Study in Southern California. Remote Sensing 2019, 11, 2902. [Google Scholar] [CrossRef]
- Blanco-Rodríguez, M.Á.; Ameztegui, A.; Gelabert, P.; Rodrigues, M.; Coll, L. Short-term recovery of post-fire vegetation is primarily limited by drought in Mediterranean forest ecosystems. Fire Ecology 2023, 19, 68. [Google Scholar] [CrossRef]
- Vidal-Macua, J.J.; Ninyerola, M.; Zabala, A.; Domingo-Marimon, C.; Pons, X. Factors affecting forest dynamics in the Iberian Peninsula from 1987 to 2012. The role of topography and drought. Forest Ecology and Management 2017, 406, 290–306. [Google Scholar] [CrossRef]
- Rossetti, I.; Cogoni, D.; Calderisi, G.; Fenu, G. Short-Term Effects and Vegetation Response after a Megafire in a Mediterranean Area. Land 2022, 11, 2328. [Google Scholar] [CrossRef]
- Resco de Dios, V.; Arteaga, C.; Hedo, J.; Gil-Pelegrín, E.; Voltas, J. A trade-off between embolism resistance and bark thickness in conifers: Are drought and fire adaptations antagonistic? Plant Ecology & Diversity 2018, 11, 253–258. [Google Scholar] [CrossRef]
- Guajardo Valdés, A.G. Respuestas ecofisiológicas de plantas de Lithraea caustica (Mol.) Hook et Arn. sometidas a restricción hídrica controlada. 2010.
- Peña-Rojas, K.; Donoso, S.; Pacheco, C.; Riquelme, A.; Gangas, R.; Guajardo, A.; Durán, S. Respuestas morfo-fisiológicas de plantas de Lithraea caustica (Anacardiaceae) sometidas a restricción hídrica controlada. Bosque (Valdivia) 2018, 39, 27–36. [Google Scholar] [CrossRef]
- Castillo, M.; Plaza, Á.; Garfias, R. A recent review of fire behavior and fire effects on native vegetation in Central Chile. Global Ecology and Conservation 2020, 24, e01210. [Google Scholar] [CrossRef]
- Keeley, J.E.; Keeley, S.C. Post-Fire Regeneration of Southern California Chaparral. American Journal of Botany 1981, 68, 524–530. [Google Scholar] [CrossRef]
- Keeley, J.E. Resilience of mediterranean shrub communities to fires. In Resilience in mediterranean-type ecosystems; Dell, B., Hopkins, A.J.M., Lamont, B.B., Eds.; Springer Netherlands: Dordrecht, 1986; pp. 95–112. [Google Scholar]
- Vita, A.; Serra, M.T.; Grez, I.; González, M.; Olivares, A. Respuesta del rebrote en espino (Acacia caven (Mol.) Mol.) sometido a intervenciones silviculturales en zona árida de Chile. Ciencias Forestales 1997, 12, 3–18. [Google Scholar]
- Montoya-Tangarife, C.; De La Barrera, F.; Salazar, A.; Inostroza, L. Monitoring the effects of land cover change on the supply of ecosystem services in an urban region: A study of Santiago-Valparaíso, Chile. PLoS ONE 2017, 12, e0188117. [Google Scholar] [CrossRef]
- Viedma, O.; Meliá, J.; Segarra, D.; Garcia-Haro, J. Modeling rates of ecosystem recovery after fires by using landsat TM data. Remote Sensing of Environment 1997, 61, 383–398. [Google Scholar] [CrossRef]
- Wittenberg, L.; Malkinson, D.; Beeri, O.; Halutzy, A.; Tesler, N. Spatial and temporal patterns of vegetation recovery following sequences of forest fires in a Mediterranean landscape, Mt. Carmel Israel. CATENA 2007, 71, 76–83. [Google Scholar] [CrossRef]
- Hope, A.; Tague, C.; Clark, R. Characterizing post-fire vegetation recovery of California chaparral using TM/ETM+ time-series data. International Journal of Remote Sensing 2007, 28, 1339–1354. [Google Scholar] [CrossRef]
- Hislop, S.; Jones, S.; Soto-Berelov, M.; Skidmore, A.; Haywood, A.; Nguyen, T.H. Using Landsat Spectral Indices in Time-Series to Assess Wildfire Disturbance and Recovery. Remote Sensing 2018, 10, 460. [Google Scholar] [CrossRef]
- Meng, R.; Dennison, P.E.; Huang, C.; Moritz, M.A.; D'Antonio, C. Effects of fire severity and post-fire climate on short-term vegetation recovery of mixed-conifer and red fir forests in the Sierra Nevada Mountains of California. Remote Sensing of Environment 2015, 171, 311–325. [Google Scholar] [CrossRef]
- Bright, B.C.; Hudak, A.T.; Kennedy, R.E.; Braaten, J.D.; Henareh Khalyani, A. Examining post-fire vegetation recovery with Landsat time series analysis in three western North American forest types. Fire Ecology 2019, 15, 8. [Google Scholar] [CrossRef]
- Agee, J.K. Fire ecology of Pacific Northwest forests; Island press Washington, DC: 1993; Volume 499.
- Meng, R.; Wu, J.; Zhao, F.; Cook, B.D.; Hanavan, R.P.; Serbin, S.P. Measuring short-term post-fire forest recovery across a burn severity gradient in a mixed pine-oak forest using multi-sensor remote sensing techniques. Remote Sensing of Environment 2018, 210, 282–296. [Google Scholar] [CrossRef]
- Smith-Ramírez, C.; Castillo-Mandujano, J.; Becerra, P.; Sandoval, N.; Allende, R.; Fuentes, R. Recovery of Chilean Mediterranean vegetation after different frequencies of fires. Forest Ecology and Management 2021, 485, 118922. [Google Scholar] [CrossRef]
- Zhou, L.; Tian, Y.; Myneni, R.B.; Ciais, P.; Saatchi, S.; Liu, Y.Y.; Piao, S.; Chen, H.; Vermote, E.F.; Song, C.; et al. Widespread decline of Congo rainforest greenness in the past decade. Nature 2014, 509, 86–90. [Google Scholar] [CrossRef]
- Hilker, T.; Lyapustin, A.I.; Tucker, C.J.; Hall, F.G.; Myneni, R.B.; Wang, Y.; Bi, J.; Mendes de Moura, Y.; Sellers, P.J. Vegetation dynamics and rainfall sensitivity of the Amazon. Proceedings of the National Academy of Sciences 2014, 111, 16041–16046. [Google Scholar] [CrossRef]
- Barbosa, J.M.; Asner, G.P. Effects of long-term rainfall decline on the structure and functioning of Hawaiian forests. Environmental Research Letters 2017, 12, 094002. [Google Scholar] [CrossRef]
- Holmgren, M.; Segura, A.M.; Fuentes, E.R. Limiting mechanisms in the regeneration of the Chilean matorral–Experiments on seedling establishment in burned and cleared mesic sites. Plant Ecology 2000, 147, 49–57. [Google Scholar] [CrossRef]
- Fuentes-Castillo, T.; Miranda, A.; Rivera-Hutinel, A.; Smith-Ramírez, C.; Holmgren, M. Nucleated regeneration of semiarid sclerophyllous forests close to remnant vegetation. Forest Ecology and Management 2012, 274, 38–47. [Google Scholar] [CrossRef]
- Solans Vila, J.P.; Barbosa, P. Post-fire vegetation regrowth detection in the Deiva Marina region (Liguria-Italy) using Landsat TM and ETM+ data. Ecological Modelling 2010, 221, 75–84. [Google Scholar] [CrossRef]
- Serra-Burriel, F.; Delicado, P.; Cucchietti, F.M. Wildfires Vegetation Recovery through Satellite Remote Sensing and Functional Data Analysis. Mathematics 2021, 9, 1305. [Google Scholar] [CrossRef]
- Justin, E.; David, V. Landscape-level interactions of prefire vegetation, burn severity, and postfire vegetation over a 16-year period in interior Alaska. Canadian Journal of Forest Research 2005, 35, 1367–1377. [Google Scholar] [CrossRef]
- Miller, R.; Chambers, J.C.; Pyke, D.A.; Pierson, F.B.; Williams, C.J. A review of fire effects on vegetation and soils in the Great Basin Region: Response and ecological site characteristics. 2013.
- Fernández-García, V.; Calvo, L.; Suárez-Seoane, S.; Marcos, E. Remote Sensing Advances in Fire Science: From Fire Predictors to Post-Fire Monitoring. Remote Sensing 2023, 15, 4930. [Google Scholar] [CrossRef]
- Pérez-Cabello, F.; Montorio, R.; Alves, D.B. Remote sensing techniques to assess post-fire vegetation recovery. Current Opinion in Environmental Science & Health 2021, 21, 100251. [Google Scholar] [CrossRef]
- Gao, Y.; Skutsch, M.; Paneque-Gálvez, J.; Ghilardi, A. Remote sensing of forest degradation: A review. Environmental Research Letters 2020, 15, 103001. [Google Scholar] [CrossRef]
- Chazdon, R.L. Tropical forest recovery: Legacies of human impact and natural disturbances. Perspectives in Plant Ecology, Evolution and Systematics 2003, 6, 51–71. [Google Scholar] [CrossRef]
- Holl, K.D.; Aide, T.M. When and where to actively restore ecosystems? Forest Ecology and Management 2011, 261, 1558–1563. [Google Scholar] [CrossRef]
- Schulz, J.J.; Cayuela, L.; Echeverria, C.; Salas, J.; Rey Benayas, J.M. Monitoring land cover change of the dryland forest landscape of Central Chile (1975–2008). Applied Geography 2010, 30, 436–447. [Google Scholar] [CrossRef]
- Armesto, J.J.; Arroyo, K.; Mary, T.; Hinojosa, L.F. The Mediterranean environment of Central Chile. In The physical geography of South America; Velben, T.T., K.R.Y., Orme, A.R., Eds.; Oxford University Press: New York, 2007; Volume 7, pp. 184–199. [Google Scholar]
- Mamadaliev, D.; Touko, P.L.M.; Kim, J.-H.; Kim, S.-C. ESFD-YOLOv8n: Early Smoke and Fire Detection Method Based on an Improved YOLOv8n Model. Fire 2024, 7, 303. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Hicke, J.A.; Fisher, R.A.; Allen, C.D.; Aukema, J.; Bentz, B.; Hood, S.; Lichstein, J.W.; Macalady, A.K.; McDowell, N.; et al. Tree mortality from drought, insects, and their interactions in a changing climate. New Phytologist 2015, 208, 674–683. [Google Scholar] [CrossRef]
- Allen, H.D. Response of past and present Mediterranean ecosystems to environmental change. Progress in Physical Geography: Earth and Environment 2003, 27, 359–377. [Google Scholar] [CrossRef]
- Lloret, F.; Escudero, A.; Iriondo, J.M.; Martínez-Vilalta, J.; Valladares, F. Extreme climatic events and vegetation: The role of stabilizing processes. Global Change Biology 2012, 18, 797–805. [Google Scholar] [CrossRef]
- Gill, N.S.; Jarvis, D.; Veblen, T.T.; Pickett, S.T.A.; Kulakowski, D. Is initial post-disturbance regeneration indicative of longer-term trajectories? Ecosphere 2017, 8, n. [Google Scholar] [CrossRef]
- Essa, Y.H.; Hirschi, M.; Thiery, W.; El-Kenawy, A.M.; Yang, C. Drought characteristics in Mediterranean under future climate change. npj Climate and Atmospheric Science 2023, 6, 133. [Google Scholar] [CrossRef]
Figure 3.
The boxplots display the mean percentage of NDVI recovery five years post-fire. Uppercase and lowercase letters denote significant group differences (p < 0.01). A white point marks significantly different interactions of fire severity with each drought level (confidence level 0.95, p<0.01).
Figure 3.
The boxplots display the mean percentage of NDVI recovery five years post-fire. Uppercase and lowercase letters denote significant group differences (p < 0.01). A white point marks significantly different interactions of fire severity with each drought level (confidence level 0.95, p<0.01).
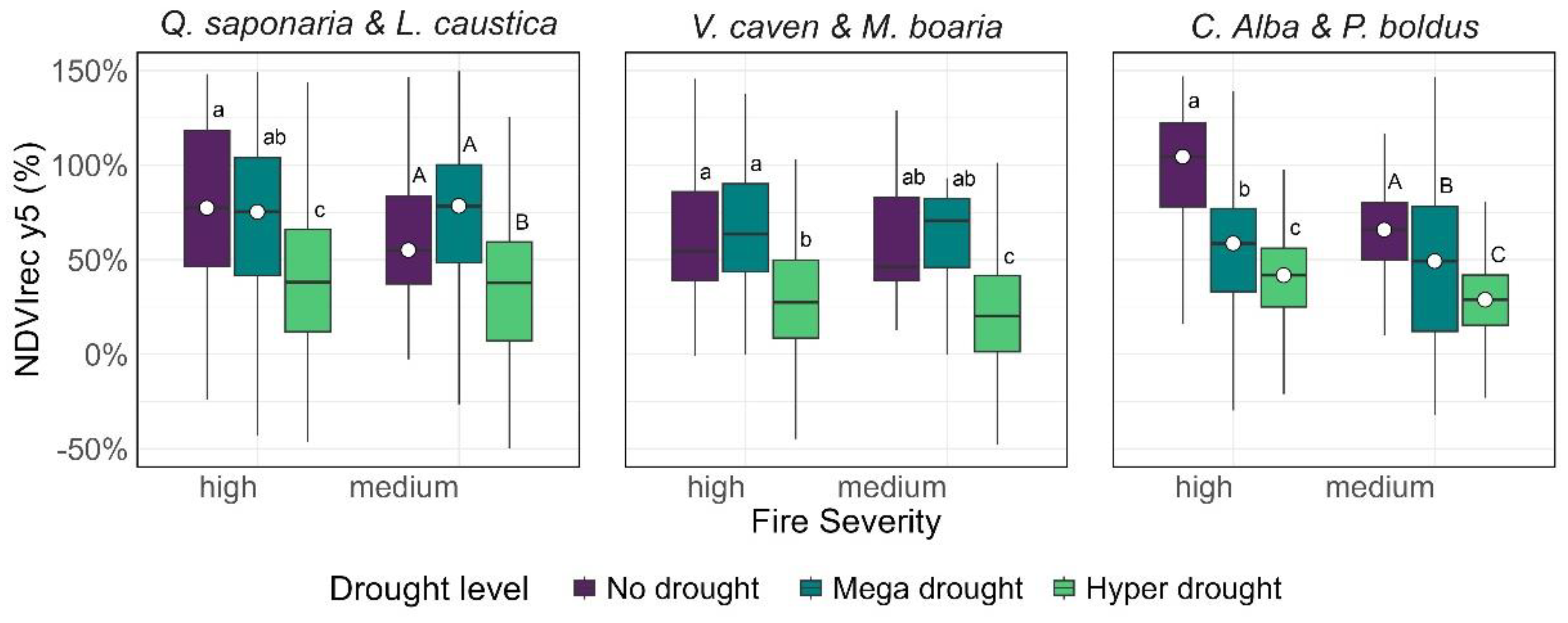
Figure 4.
Mean percentages of NDVIrec for Mega Drought and Hyper Drought relative to No Drought values as a reference period for high and medium severity. The asterisk shows significant differences concerning the No Drought.
Figure 4.
Mean percentages of NDVIrec for Mega Drought and Hyper Drought relative to No Drought values as a reference period for high and medium severity. The asterisk shows significant differences concerning the No Drought.
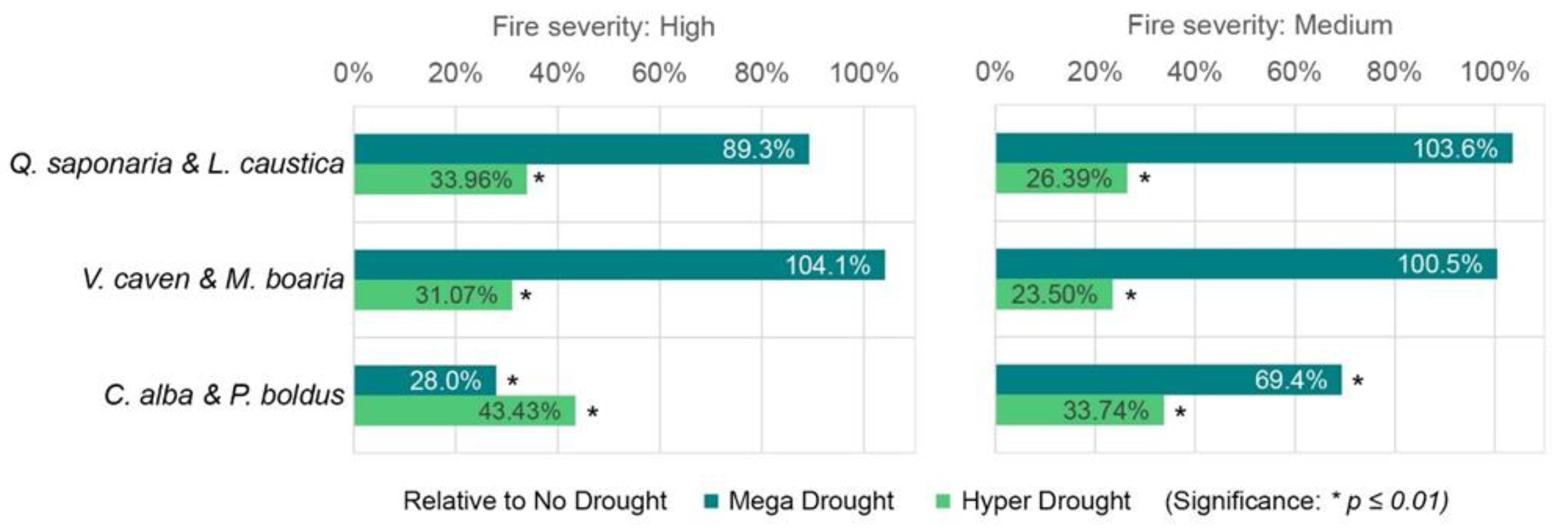
Figure 7.
Examples of post-fire resprouting and regeneration in sclerophyllous forests of Central Chile are presented. (a) shows Quillaja saponaria and three months after the wildfire that occurred in 2024; (b) Q. saponaria & Lithrea caustica forest subtype area on slope (c) Q. saponaria & Lithrea caustica forest subtype area both one year after the wildfire that occurred in 2019. Credit: a. Ana Hernández-Duarte. b.c. Jean Pierre Francois.
Figure 7.
Examples of post-fire resprouting and regeneration in sclerophyllous forests of Central Chile are presented. (a) shows Quillaja saponaria and three months after the wildfire that occurred in 2024; (b) Q. saponaria & Lithrea caustica forest subtype area on slope (c) Q. saponaria & Lithrea caustica forest subtype area both one year after the wildfire that occurred in 2019. Credit: a. Ana Hernández-Duarte. b.c. Jean Pierre Francois.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



