Submitted:
24 September 2024
Posted:
24 September 2024
You are already at the latest version
Abstract
During a survey of marine biodiversity in the deep-sea off northeastern Guam, two marine desmoscolecid nematodes belonging to the subgenus Tricoma were discovered. Tricoma (Tricoma) disparseta sp. nov. was described based on specimens collected from sponge and starfish habitats on a seamount at depths ranging from 1300 to 1500 meters. T. (T.) disparseta sp. nov. is distinguished by having 59 to 62 main rings, 9 to 10 subdorsal setae, and 14 to 18 subventral setae on each side. Notable features include the differentiation in length and insertion between subdorsal and subventral setae, and a curved, hooked gubernaculum. The specimen of T. (T.) longirostris observed in this study closely resembles previously reported specimens, characterized by 78 main rings, a long and narrow head shape, 8 to 9 subdorsal setae, 14 to 15 subventral setae, and a gubernaculum with a knobbed apophysis. The morphological characteristics and detailed illustrations of two Tricoma species from the Northwest Pacific Ocean were documented using differential interference contrast microscopy (DIC) and scanning electron microscopy (SEM). Furthermore, pictorial keys and comparative tables are presented for species exhibiting 50 to 64 main rings, aiding in their identification and differentiation.
Keywords:
marine nematodes
; Tricoma
; morphometrics
; taxonomy
; Northwest Pacific Ocean
1. Introduction
Deep-sea ecosystems, which span approximately 65% of the Earth's surface, are home to a substantial portion of global biodiversity [1,2]. Among the diverse organisms inhabiting these environments, nematodes stand out as the most prolific metazoans, representing over 90% of the benthic deep-sea population [1,2]. Recent research underscores the high species diversity within deep-sea nematode communities [3], highlighting that water depth plays a crucial role in influencing species abundance [4,5,6]. This depth-dependent variation is closely linked to nutrient availability and sediment characteristics [6]. However, investigating the diversity of deep-sea nematode communities poses significant taxonomic challenges, primarily due to the limited number of individuals available for each genus or species [3,7].
Within this diverse nematode community, the Desmoscolecida are particularly notable for their presence in deep-sea habitats. Despite their relatively low species richness, Desmoscolecida exhibit high diversity[8]. The genus Tricoma, established by Cobb in 1894, represents one of the major taxa within the Desmoscolecida. It encompasses two subgenera: Tricoma (Tricoma) and Tricoma (Quadricoma), with ongoing debates regarding their precise taxonomic classification. The subgenus Tricoma alone comprises 87 recorded species globally, with 23 of these species occurring at depths greater than 200 meters, thus representing 26% of the total [9]. Species belonging to the subgenus Tricoma are characterized by several distinct morphological features: their desmen are typically rounded or triangular in shape, they lack a pronounced reversal ring, and their head, when viewed in optical section, appears more or less triangular. Additionally, the terminal ring is predominantly cylindrical, further distinguishing them from related taxa [10].
During a survey of free-living marine nematode biodiversity in international waters around Guam in the Northwest Pacific Ocean, two species, Tricoma (T.) disparseta sp. nov. and T. (T.) longirostris (Southern, 1914), were discovered on the surface of a sponge and in sediment in the deep-sea seamount region. Seamounts, which are underwater topographic elevations of the seafloor, play a crucial role in marine biogeography [11]. These structures are known to create unique hydrological conditions, such as enhanced semi-closed circulation patterns and increased vertical mixing, which result in the formation of circulation cells above their summits [12,13,14,15]. These circulation cells enhance the local production of primary and secondary organisms and help retain larvae and pelagic production in the water column, thereby improving food resources for benthic fauna [13]. Consequently, seamounts support high biodiversity and serve as important habitats for a variety of macro- and megabenthic taxa [12,15,16]. To date, six species of the subgenus Tricoma have been recorded from the Northwest Pacific [9]. This study presents a detailed description of the two Tricoma species discovered in the seamounts off northeastern Guam, Northwest Pacific. The descriptions are supported by illustrations and photographs obtained through scanning electron microscopy (SEM) and differential interference contrast microscopy (DIC). Additionally, a comparative character table and a pictorial key are included for species with 50 to 64 main rings, offering a visual and tabulated guide for their identification and comparison.
2. Material and Methods
2.1. Field Sampling and Sample Processing
In September 2023, the oceanographic research vessel ISABU, operated by the Korea Institute of Ocean Science and Technology (KIOST), carried out a deep-sea exploration in the seamount region northeast of Guam (Figure 1; Table 1). Samples were collected from the seafloor at depths ranging from 1,300 to 1,500 meters using a suction sampler attached to a remotely operated vehicle (ROV). The collected substrate, containing meiofauna and macrofauna, was processed in the field by applying osmotic shock with tap water for 10 minutes to separate the organisms from the silt. The samples were then filtered through sieves with mesh sizes of 1 mm and 63 µm to separate microfauna from macrofauna and were fixed in a 5% formaldehyde solution for long-term preservation.
2.2. Laboratory Processing and Microscopic Analysis
In the laboratory, meiofauna was separated from detritus and sediments by flotation in Ludox HS40 and screened through a 63 µm mesh sieve. Target nematodes were manually selected using a Pasteur pipette under a dissecting microscope and transferred to a 3% glycerin solution. The solution was evaporated at room temperature over 10 days until only pure glycerol remained. Specimens were mounted in glycerin between two cover slips using the standard wax ring method. Observations and measurements were conducted using an Olympus BX53 microscope equipped with cellSens Standard software. Photographs were taken with a LEICA DM2500 LED microscope equipped with a LEICA K5C color CMOS camera (Wetzlar, Germany), and image quality was enhanced using Adobe Photoshop 2023. Drawings were produced using a 100x objective lens with immersion oil and a microscope with Nomarski differential interference contrast (DIC) from an Olympus BX53 microscope (Tokyo, Japan), equipped with a drawing tube. Line drawings were created using tracing techniques with a Wacom Cintiq 22 tablet and Adobe Illustrator.
2.3. Scanning Eelectron Microscopy (SEM) Analysis
For SEM analysis, specimens were initially fixed in 5% formaldehyde solution, rinsed three times with distilled water for 10 minutes each, freeze-dried, and mounted on aluminum stubs using pins. A thin layer of gold/palladium was applied using a high-vacuum sputter coater. SEM imaging was performed with a field emission SEM (JSM-7200F) at KOPRI, providing detailed visualization of specimen morphology.
Abbreviations used in the text are as follows:
- a: body length divided by maximum body diameter,
- b: body length divided by pharynx length,
- c: body length divided by tail length,
- V (%): vulva distance from the anterior end as a percentage of total body length.
3. Results and Discussion
Taxonomic Account
Order Desmoscolecida Filipjev, 1929
Family Desmoscolecidae Shipley, 1896
Subfamily Tricominae Lorenzen, 1969
Genus Tricoma Cobb, 1894
Subgenus Tricoma Cobb, 1894
Tricoma (Tricoma) disparseta sp. nov. (Figure 2, Figure 3, Figure 4, Figure 5, Figure 6 and Figure 7, Table 2)
Differential diagnosis: Tricoma (Tricoma) disparseta sp. nov. is characterized by a distinctive set of features that set it apart from other species. The species possesses 59 to 62 tricomoid main rings, contributing to its unique morphology. The head is triangular, with a width 1.1 to 1.4 times greater than its length, providing a notable outline. It has slender cephalic setae, which are flanked by a narrow membrane and inserted on raised peduncles, enhancing its sensory capabilities. The somatic setae are composed of 9 to 10 subdorsal setae and 14 to 18 subventral setae on each side. The subventral setae are significantly longer than the subdorsal setae and are inserted at a relatively higher point. These setae exhibit a groove distally, with a step-shaped tip, distinguishing them from other setae. The spicules have a smaller capitulum proximally, while the gubernaculum is notably bent into a hooked shape, adding to the unique sexual morphology. The tail comprises 8 to 10 main rings and terminates in a conical ring. The anterior 26 to 40% of the tail is covered by desmen, and it is marked by rounded phasmata.
Material examined: The holotype male (MNB001), paratype males (MNB002, MNB003, MNB004, MNB005, MNB006), and allotype female (MNB007), along with paratype females (MNB008, MNB009, MNB010), preserved in glycerin on HS slides, are deposited in the nematode collection at the specimen conservation room of the East Sea Research Institute, Korea Institute of Ocean Science & Technology (KIOST), Korea.
Type locality and habitat: The specimens were collected from the seamount area at Deep-Sea Station 2 (17° 02' 47.346" S, 149° 52' 50.484" E) and Station 3 (17° 05' 45.41" S, 149° 55' 07.67" E), situated in the northeastern part of Guam. The collection took place on 6-7 September 2023, conducted by J.M. Lee. Nematodes were retrieved from a sponge and starfish habitat in the deep sea using a suction sampler mounted on a remotely operated vehicle (ROV) at depths of 1,366.6 and 1,510.1 meters.
Etymology: The specific name disparseta is derived from the Latin words dispar (meaning "unequal") and seta (meaning "bristle"), highlighting the marked disparity in length between the subdorsal and subventral setae.
Measurements: All measurement data are provided in Table 2.
Description: Males. The body length ranges from 527 to 613 µm, making it relatively small and slender, tapering towards both ends (Figures 2A, 4A, 6A). The maximum body diameter at the mid-body level measures 30-35 µm.
The holotype male cuticle consists of 60 tricomoid rings ventrally and 61 dorsally, while the paratypes exhibit between 59 to 61 tricomoid rings. The cuticular layer shows secondary annulation, and the desmen are covered with secretions and fine foreign material (Figure 5A).
The head has a diameter of 19-21 µm, being 1.1 to 1.4 times wider than it is long. It tapers anteriorly from the peduncles of the cephalic setae, ending in a truncated shape that is 4–5 µm wide (Figures 2B, 4D, 6B). The anterior margin of the head cuticle is significantly thickened and sclerotized, forming a distinct rim-shaped border, except in the labial region (Figure 2D).
The labial region does not protrude, and each of the six lips bears small labial papillae. The cephalic setae measure 22–28 µm in length, making them longer than the width of the head. They are inserted on relatively high peduncles that project in front of the posterior head border. These setae taper from a broad cylindrical base toward the tip, with grooves running along their entire length, and are surrounded by a thin membrane that is difficult to observe from a lateral view (Figure 4E). The distal ends of the cephalic setae, enclosed within this delicate membrane, appear to be bifurcated (Figure 4F).
The amphids are large and vesicular, exhibiting an unusual shape that covers the lateral sides of the head. They curve slightly at their base, forming a cylindrical structure. The amphid pore is prominent and positioned at the posterior end of the head (Figure 4C). The amphids extend anteriorly toward the labial region and posteriorly to the main ring 2 or 3 (Figures 2C, 6C, 6E).
The stoma is small and cylindrical, approximately 3 µm deep. The esophagus is also cylindrical, measuring 93–97 µm in length, which constitutes about 16–18% of the total body length. It is surrounded by a nerve ring at the level of main rings 5 and 6. The esophagus-intestinal junction is located between main rings 10 and 11, corresponding to a body diameter of 24–29 µm. The ocelli are large and dark yellowish, measuring 4–8 µm in width and 5–13 µm in length. They are positioned opposite main ring 14 in the holotype and rings 12–14 or 20–24 in the paratypes. Smaller pigment spots are present along the pharynx.
Somatic setae are arranged in two distinct groups on the subdorsal and subventral sides of the body (Figure 5A). The subdorsal setae are slender and approximately half the length of the subventral setae, being mounted on relatively low peduncles (Figure 5B). In contrast, the subventral setae are longer and display a stepped appearance at the tip in optical sections (Figure 6F), and they are inserted into relatively high peduncles (Figure 5C). The subventral setae of the most anterior main ring 3 and those around the cloacal region are shorter than the other setae, gradually increasing in length toward the middle of the body. While the difference in length between the subdorsal and subventral setae in the anterior and caudal regions of the body is not substantial, it becomes approximately twofold in the central region. The somatic setae are arranged in 9 to 10 subdorsal setae and 14 to 18 subventral setae on each side, measuring 15–36 µm and 15–21 µm in length, respectively. The anterior-most pair of setae on main ring 3 is positioned laterally (Figure 2B, C). Additionally, the subventral setae located second in front of the cloacal opening and the terminal subventral setae are inserted slightly laterally (Figures 2F, 4B). In most specimens—excluding paratype male 2—the subventral setae situated on rings 44–46 (the second setae anterior to the cloacal opening) were the longest, measuring 32–36 µm. Observing the length of the setae can be challenging depending on the specimen's position or angle, which may lead to measurement errors. The somatic setae are inserted almost directly into the peduncle cuticular rings, and some setae may be severed or damaged. The arrangement of the somatic setae in the holotype male is as follows:
| subdorsal | left side: | 5,10,17,23,31,41,47,53,58 | = 9 |
| right side: | 6,10,15,19,23,33,39,46,53,59 | = 10 | |
| subventral | left side: | 3,5,8,12,14,17,20,23,27,31,34,38,42,45,47,53,55 | = 17 |
| right side: | 3,5,8,11,14,16,20,23,26,29,32,36,39,42,45,47,52,55 | = 18 |
The arrangement of the somatic setae in the paratype males are as follows (numbers in brackets indicate different positions in the paratype):
| subdorsal | left side: | 5(4),9(10),17,24(22,23),32(31),41(39),48(45,46,47),52(51,53),58(57) | = 9 |
| right side: | 5,9(10),18(17),23(25),31(30),39(38),48(46),53(51,52),58(57) | = 9 | |
| subventral | left side: | 3,5,8(7),11(9),13(12),16(14),18(17,19),21(20,22),24(23,25,26),28(27),32(30,31),36(33,34,35),40(38,39),43(41,42),46(45),48,53(52),55(54) | = 18(16,17) |
| right side: | 3,5,8(7),11,14(13),17(16),20(19),24(22,23),27(26,28),32(29,30),35(34),39(37,38),42(41),45(44),48,53(52),55 | = 17(14,16) |
The reproductive system typically contains two testes. The spicules measure 29–32 µm in length and are slightly curved, tapering distally to a pointed tip, with a relatively small capitulum at the proximal end (Figures 5D, E, 6D). The gubernaculum is hook-shaped, measuring 16–18 µm in length. Its distal side runs parallel to the spicules and exhibits a noticeable curvature, forming a hook shape in the proximal third and at the distal end. The distal portion of the gubernaculum is sclerotized and tapered, culminating in a pointed apex (Figures 2E, 6G).
The tail consists of 9–10 main rings, measuring 89–96 µm in length. The terminal ring is conical and features a slightly thicker cuticle, with the exception of the terminal spinneret. The anterior 26–33% of the terminal ring is covered with debris, while the distal end remains bare and clean (Figures 2F, 5F, 6H). Circular phasmata, measuring 2.5–2.9 µm in diameter, are located beneath the desmos of the terminal ring.
Females. They closely resemble males in most respects, apart from their sexual characteristics (Figures 3B, C, 7B, C). The body length ranges from 490 to 575 µm, with a maximum diameter of 31 to 42 µm. The cuticle of the allotype female consists of 60 tricomoid rings ventrally and 61 dorsally, while the paratypes possess 60 to 62 tricomoid rings. These rings are adorned with secretions and fine foreign material (Figures 3A, 7A). The somatic setae are arranged in 9 pairs of subdorsal setae and 16 to 18 subventral setae on each side (Figure 7F).
The arrangement of the somatic setae in the allotype female is as follows:
| subdorsal | left side: | 6,11,18,23,32,42,47,53,59 | = 9 |
| right side: | 6,11,18,24,32,40,47,53,59 | = 9 | |
| subventral | left side: | 4,5,8,10,12,15,19,22,25,29,33,37,41,45,49,52,55 | = 17 |
| right side: | 4,5,7,10,13,15,18,22,25,29,33,37,41,44,48,51,55 | = 17 |
The arrangement of the somatic setae in the paratype females are as follows (numbers in brackets indicate different positions in the paratype):
| subdorsal | left side: | 7(6),11,17(18),24(23),32,40(42),47(48),53,59(58) | = 9 |
| right side: | 6,11,18(19),23(24,26),32(33),40,48(47),54(53),59(58) | = 9 | |
| subventral | left side: | 4(3),5,8,11(12),14(15),16,19(20),22(23),25(26,27),29(30,31),33(34,35),37(38,39),41(42),44(45),49(48),55(51,53),57(56) | = 17(16) |
| right side: | 4(3),5,7(8),9(10,11),12(13),14(15),16(18),20(21),22,25(24),29(28),34(33),38(37),42(41),45,49(48),52(51),57(56) | = 18(16,17) |
The reproductive system is didelphic, with both branches fully extended. The vulva is situated in the bare medioventral part of the body wall, positioned between main rings 39 and 40 in the allotype (between rings 38 and 40 in the paratypes) (Figures 3D, 7D). The uteri may contain large and small amorphous inclusions, or may be devoid of them.
The tail consists of eight to nine main rings, measuring 71–84 µm in length. The terminal ring measures 24–26 µm in length and 10–12 µm in maximum width (Figure 3E). The terminal ring is conical, with the anterior 31–40% covered by desmos, while the remaining region is bare (Figure 7E). Circular phasmata, with a diameter of 2.5–2.8 µm, are located beneath the desmos of the terminal ring.
Discussion. The subgenus Tricoma was first described by Cobb in 1894, and to date, 87 species have been documented worldwide. However, only a limited number of researchers have extensively reported on these species. Decraemer (1978, 1979, 1983, 1987, 1996), Decraemer & Tchesunov (1996), and Soetaert & Decraemer (1989) collectively described 35 species, while Timm (1970, 1978) contributed the documentation of an additional 15 species [17,18,19,20,21,22,23,24,25,26]. Together, these contributions account for approximately 57% of the known species within the subgenus. In 1978, Decraemer made a significant contribution by describing 12 new species, one subspecies, and five previously recorded species from the Great Barrier Reef in Australia. This finding highlights the high level of species diversity within Tricoma in a single region. Despite the distinctive ringed body shape that sets Tricoma apart from other taxa, there has been relatively little research focused on comparative traits within the subgenus. Consequently, taxonomic studies on Tricoma have been limited over the past two decades, with few updates since Decraemer & Tchesunov reported two new species in 1996 [23]. Recently, however, four new species and two previously unrecorded species have been discovered in the East Sea of Korea, leading to the compilation of a literature-based species list for the subgenus Tricoma [9].
In 1975, Freudenhammer proposed a key for comparing characters within the subgenus Tricoma, categorizing it into three groups: Group A, species with prominent features in the setae; Group B, species with characteristics in the desmen, such as spines, hook-like appendages, or a tiled structure without foreign material; and Group C, species lacking distinctive features in either the setae or the desmen [27]. Group C was further subdivided into three subgroups based on the number of main rings: 36 to 40 desmens, 50 to 64 desmens, and 67 to 140 desmens. However, Decraemer (1978) later revised this classification, organizing species into six groups based on the number of main rings: 30 or fewer, 30 to 49, 50 to 60, 61 to 75, 76 to 100, and more than 100 [17]. This classification provides a more nuanced framework for grouping Tricoma species according to their main ring count, serving as a valuable taxonomic tool for preliminary species identification, despite the considerable variability in main ring numbers observed within the subgenus.
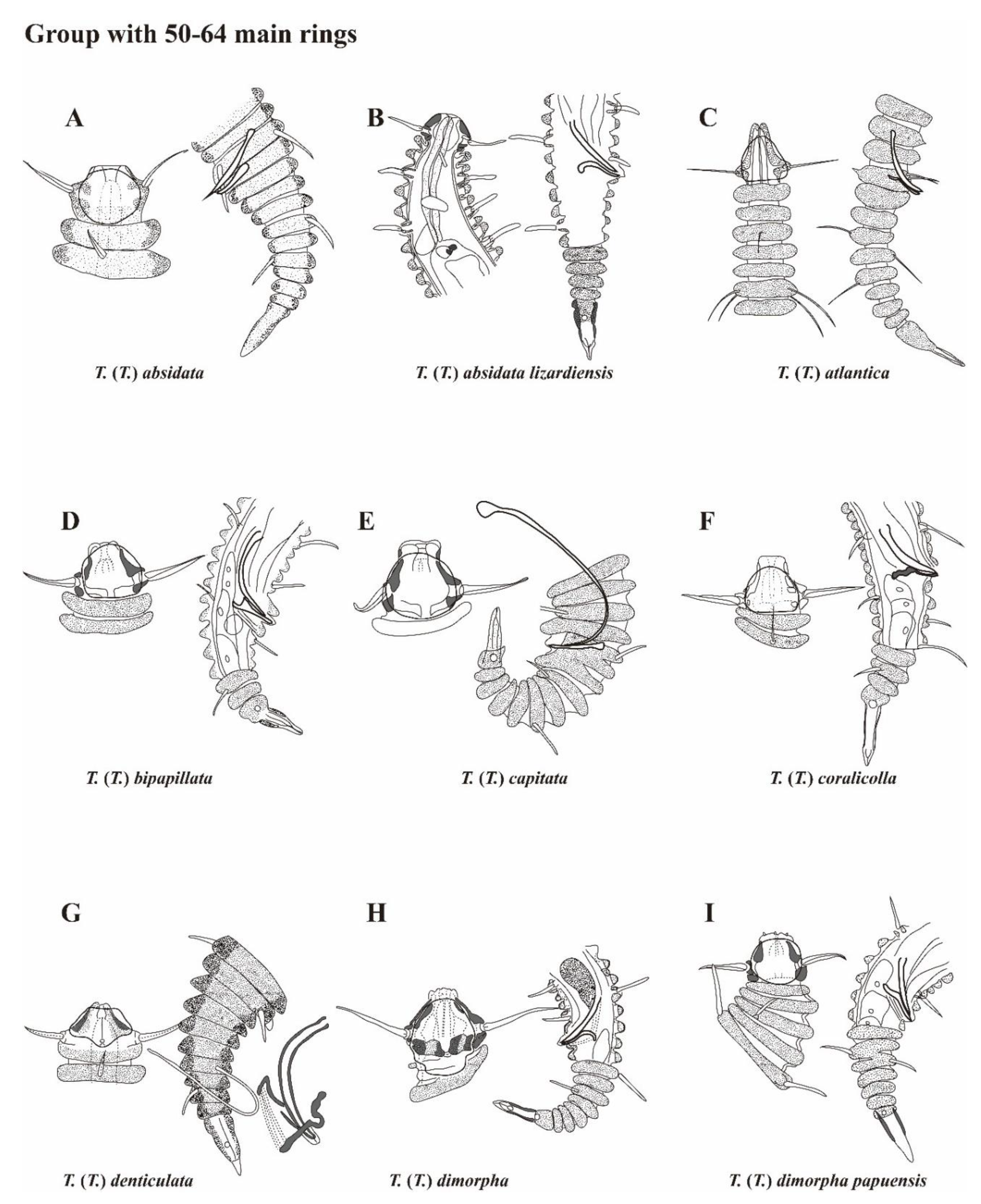
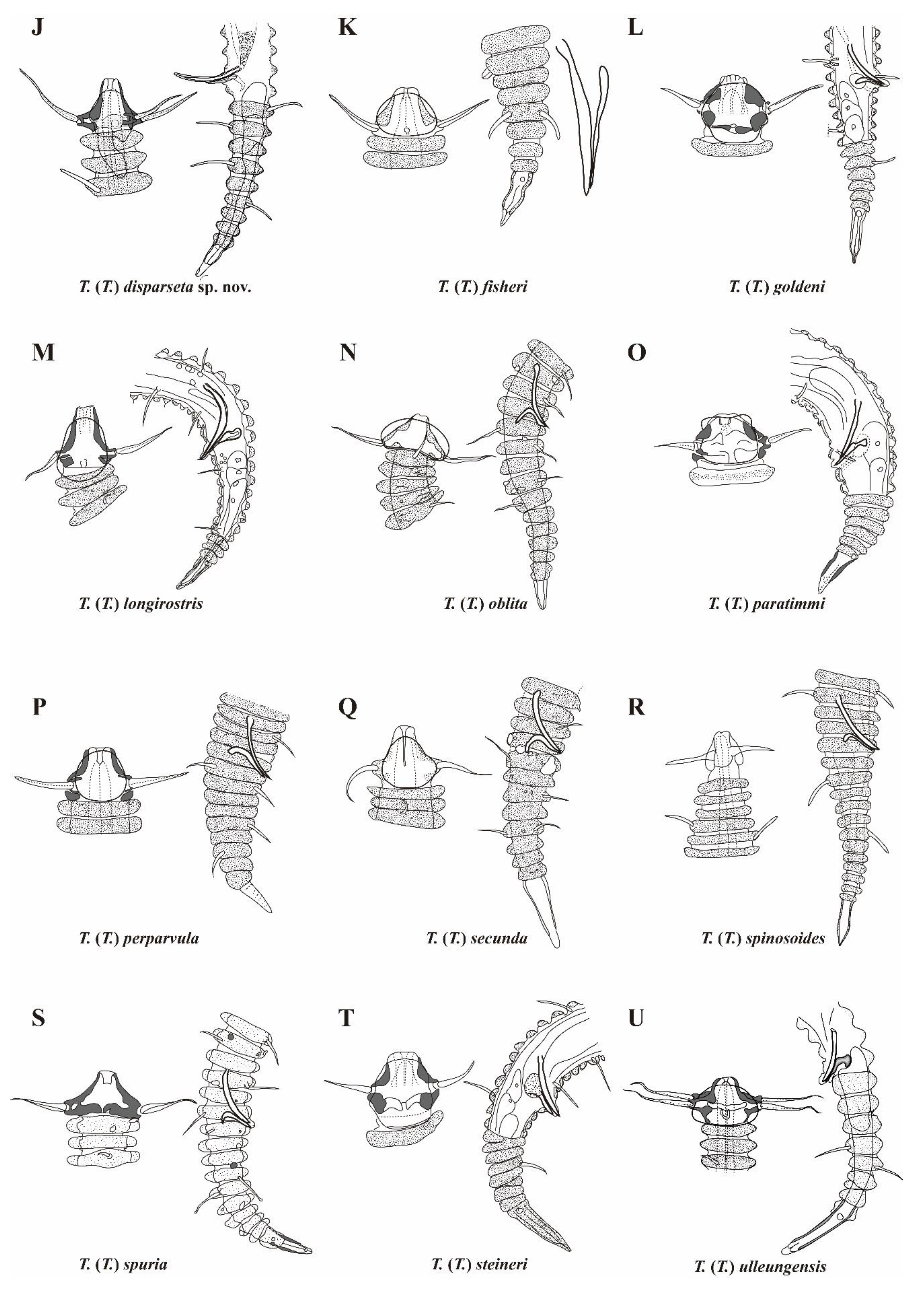
The newly described Tricoma (Tricoma) disparseta sp. nov. exhibits 59 to 62 main rings, placing it within the category of species characterized by 50 to 64 main rings, according to Freudenhammer’s classification system. This range of main rings is considered a key diagnostic feature within the subgenus Tricoma. Currently, twenty species are classified under this category, which serves as an essential criterion for distinguishing between closely related taxa. The species included in this group are: Tricoma (Tricoma) absidata Timm, 1970; T. (T.) absidata lizardiensis Decraemer, 1979; T. (T.) atlantica Freudenhammer, 1975; T. (T.) bipapillata Decraemer, 1987; T. (T.) capitata Decraemer, 1987; T. (T.) coralicolla Decraemer, 1987; T. (T.) denticulata Timm, 1970; T. (T.) dimorpha Decraemer, 1978; T. (T.) dimorpha papuensis Decraemer, 1987; T. (T.) fisher Timm, 1970; T. (T.) goldeni Decraemer, 1978; T. (T.) longirostris (Southern, 1914); T. (T.) oblita Blome, 1982; T. (T.) paratimmi Decraemer, 1987; T. (T.) perpavula Timm, 1970; T. (T.) secunda Blome, 1982; T. (T.) spinosoides Chitwood, 1951; T. (T.) spuria Inglis, 1968; T. (T.) steineri de Man, 1922, and T. (T.) ulleungensis Lee, Lee & Rho, 2023.
In this study, we provide a pictorial identification key and a comparative table of diagnostic morphological characters for species groups possessing 50 to 64 main rings (Figure 10, Table 3). The diagnostic features highlighted in these tools primarily focus on the morphology of the anterior head and posterior tail regions, with particular emphasis on the spicules and gubernaculum, which are crucial taxonomic traits for differentiating closely related nematode species. The morphological comparison table draws on data from the original species descriptions, providing a comprehensive summary of the key morphological traits that distinguish members of this species group. This approach facilitates a clear presentation and comparison of each species' unique features. For T. (T.) spuria, the information is derived from detailed reobservations of the type specimen, as documented by Decraemer (1981).
The newly discovered species, Tricoma (Tricoma) disparseta sp. nov., was found in sediment washings from habitats associated with sponges and starfish on a seamount located in the northeastern region of Guam. This species is distinguished by the presence of 59 to 62 main rings, a triangular head bearing elongated cephalic setae, and amphids that extend posteriorly to main ring 2 or 3. The somatic setae are arranged in 9 to 10 subdorsal setae and 14 to 18 subventral setae on each side, with the subventral setae being notably longer and inserted at a higher point than the subdorsal setae. Moreover, the species is characterized by a gubernaculum that curves into a distinctive hooked shape, further differentiating it from other congeners.
Within the subgenus Tricoma, only one specie, T. (T.) coralicolla Decraemer, 1987, among the 20 species with 50 to 64 main rings, exhibits a significant difference in the length of subdorsal and subventral setae. T. (T.) disparseta sp. nov. closely resembles T. (T.) coralicolla in this regard, particularly due to its subventral setae being approximately twice as long as the subdorsal setae, and the peduncles of the subventral setae being more prominently developed. T. (T.) coralicolla was originally discovered in overgrown dead coral habitats in Papua New Guinea. Despite the limited sample size, only two specimens (one male and one female), preventing a thorough assessment of variability in main ring count or seta pattern, T. (T.) disparseta sp. nov. can be clearly distinguished from T. (T.) coralicolla by the following characteristics: (1) total body length (490–613 µm vs. 185–220 µm in T. (T.) coralicolla); (2) number of main rings (59 to 62 vs. 56 to 58 in T. (T.) coralicolla); (3) arrangement and number of somatic setae (9 to 10 subdorsal setae and 14 to 18 subventral setae compared to 9 subdorsal setae and 11 to 12 subventral setae in T. (T.) coralicolla); (4) the extension of amphids to the 2nd or 3rd main rings (in contrast to T. (T.) coralicolla, where the amphids reach only the margin of the head); and (5) a gubernaculum with a distally curved, hooked shape, as opposed to the gubernaculum of T. (T.) coralicolla, which features two dorso-caudally oriented apophyses. These distinguishing features highlight the unique morphological traits of T. (T.) disparseta sp. nov., despite its superficial resemblance to T. (T.) coralicolla in seta morphology.
- Desmoscolex longirostris Sourhern, 1914, p. 62, Figure 29A-D.
- Tricoma longirostris: Steiner, 1916, p. 33.
- Tricoma glutinosa Steiner, 1916 p. 340, Figure 13; Freudenhammer, 1975, p. 25; Decraemer, 1983, p. 16.
- Tricoma septentrionalis Timm, 1978, p. 233, Figure 4EH; Decraemer, 1983, p. 16.
Material examined: The examined material comprised one male specimen (MNB011), which was mounted in anhydrous glycerin between two coverslips on an HS slide. This specimen is deposited in the nematode collection housed in the specimen conservation room of the East Sea Research Institute at the Korea Institute of Ocean Science & Technology (KIOST), Korea.
Type locality and habitat: The designated location is a seamount area in the deep sea, situated at coordinates 15° 38' 20.93" S, 151° 59' 50.35" E, in northeastern Guam. The specimen was collected on September 1, 2023, by J.M. Lee. The nematodes were retrieved from sediment in the deep sea using a suction sampler mounted on a remotely operated vehicle (ROV) at a depth of 1425.52 meters.
Description: Male. The body measures 808 µm in length and is relatively slender and elongated, tapering towards both ends. The maximum body diameter at the mid-body level is 30 µm. The cuticle is composed of 78 tricomoid rings, with the desmen covered in secretion and fine foreign material (Figures 8A, 9A).
The head is narrow and triangular in shape when viewed from the side, measuring 28 µm in width and 26 µm in height (Figure 8B). It tapers anteriorly from the peduncles of the cephalic setae, ending in a truncated manner, with the truncated end measuring 5 µm wide (Figure 8B). The edge of the head cuticle is thickly hardened, except in the labial region, and it gradually thickens towards the peduncle of the cephalic setae.
The labial region is inconspicuous, bearing six small labial papillae. The cephalic setae measure 24 µm in length, tapering to a fine, open tip, and are inserted on high peduncles. Each seta features a cuticular groove along its side and is enclosed by a thin membrane, which is challenging to observe from a lateral view.
The amphids are rounded and vesicular in shape, measuring 22 µm in width and 20 µm in height (Figure 9C). They cover the entire head region except for the labial area and extend to the posterior border of the head.
The stoma is small and cylindrical, measuring approximately 3 µm in depth. The esophagus is also cylindrical, with a length of 101 µm. It is surrounded by a nerve ring at the level of main rings 5. The esophagus-intestinal junction occurs between main rings 9 and 10, with a corresponding body diameter of 29 µm. The ocelli are very large and brownish, measuring 8-9 µm in width and 23-24 µm in length. They are positioned opposite main rings 12-13 and 14-16, respectively. Additionally, smaller pigment spots are present along the pharynx.
The somatic setae are slender and taper towards the tip, inserted into peduncles surrounded by concretions (Figure 9E). They gradually increase in length towards the middle of the body, with an overall length ranging from 14 to 22 µm. The subventral setae on the most anterior main ring (4 or 5) measure 8-11 µm, while those around the cloacal region range from 10 to 14 µm, both being shorter than the other setae. The somatic setae consist of 13 or 14 subventral setae and 8 subdorsal setae on each side, measuring 8-22 µm and 14-21 µm, respectively. The anterior-most pair on main rings 4 or 5 is sublaterally inserted. The somatic setae are nearly directly inserted into the peduncle cuticular rings, and some of the setae may be severed or damaged.
The arrangement of the somatic setae in the male is as follows:
| subdorsal | left side: | 7,14,20,28,40,50,57,67 | = 8 |
| right side: | 7,15,21,28,38,51,59,70 | = 8 | |
| subventral | left side: | 4,7,12,17,22,29,36,43,49,54,62,68,72 | = 13 |
| right side: | 5,7,11,16,19,24,30,36,43,48,54,61,68,71 | = 14 |
The reproductive system is characteristic of the genus. The distal end of the vas deferens is flanked by fine granular ejaculatory glands. The spicules measure 28 µm in length, are arcuate, and taper distally, featuring a proximal capitulum (Figure 9D). The gubernaculum, measuring 16 µm in length, is a distally sclerotized structure that runs parallel to the spicules. The dorso-caudally proximal part, marking the end of the sclerotized area, is distinguished by the presence of a conspicuous knob (Figure 8C).
The tail consists of 12 main rings, measuring 125 µm in length. The terminal ring is conical, with a length of 38 µm and a slightly thicker cuticle. The anterior 39% of the terminal ring is covered with debris, while the distal end remains bare. Circular phasmata, measuring 3 µm in diameter, are located on the desmos of the terminal ring (Figures 8D, 9F).
Figure 8.
Tricoma (Tricoma) longirostris (Southern, 1914), newly discovered in this study, male (A-D). A, habitus, lateral view; B, head region, lateral veiw; C, spicules and gubernaculum; D, spicule and tail region. T. (T.) longirostris (Southern, 1914) from the original description, male (E-G). E, head, dorsal view; F, spicules and gubernaculum; G, tail region (after Southern, 1914). T. (T.) glutinosa Steiner, 1916, male (H). H, anterior end (after Steiner, 1916). T. (T.) septentrionalis Timm, 1978, male (I-J). I, head region; J, specular apparatus (after Timm, 1978). T. (T.) longirostris (Southern, 1914), male (K-L). K, head, surface view; L, copulatory apparatus and tail (after Decraemer, 1983). Scale bars: 50 µm in A; 30 µm in L; 15 µm in K; 12 µm in I-J; 10 µm in B-D.
Figure 8.
Tricoma (Tricoma) longirostris (Southern, 1914), newly discovered in this study, male (A-D). A, habitus, lateral view; B, head region, lateral veiw; C, spicules and gubernaculum; D, spicule and tail region. T. (T.) longirostris (Southern, 1914) from the original description, male (E-G). E, head, dorsal view; F, spicules and gubernaculum; G, tail region (after Southern, 1914). T. (T.) glutinosa Steiner, 1916, male (H). H, anterior end (after Steiner, 1916). T. (T.) septentrionalis Timm, 1978, male (I-J). I, head region; J, specular apparatus (after Timm, 1978). T. (T.) longirostris (Southern, 1914), male (K-L). K, head, surface view; L, copulatory apparatus and tail (after Decraemer, 1983). Scale bars: 50 µm in A; 30 µm in L; 15 µm in K; 12 µm in I-J; 10 µm in B-D.
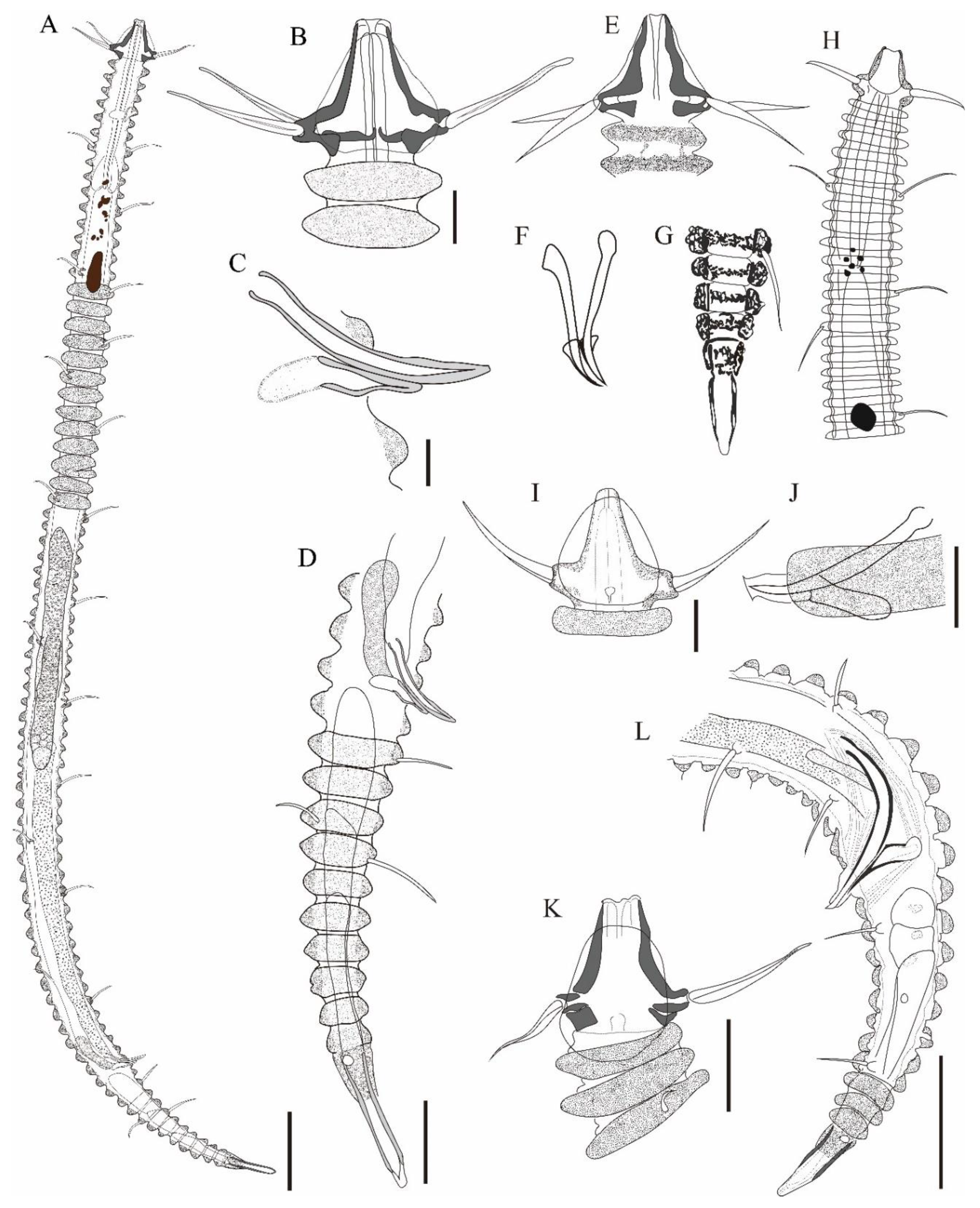
Figure 9.
Tricoma (Tricoma) longirostris (Southern, 1914), DIC photomicrographs, male. A, entire body view; B, head region; C, amphideal fovea; D, spicules and gubernaculum showing a knobbed apophysis; E, common forms of somatic setae; F, tail region. Scale bars: 50 µm in A; 10 µm in B-F.
Figure 9.
Tricoma (Tricoma) longirostris (Southern, 1914), DIC photomicrographs, male. A, entire body view; B, head region; C, amphideal fovea; D, spicules and gubernaculum showing a knobbed apophysis; E, common forms of somatic setae; F, tail region. Scale bars: 50 µm in A; 10 µm in B-F.

Figure 10.
Pictorial key to the species group with 50 to 64 main rings in the subgenus Tricoma. Source of figures: A, Timm (1970); B, Decraemer (1979); C, Freudenhammer (1975); D, Decraemer (1987); E, Decraemer (1987); F, Decraemer (1987); G, Timm (1970); H, Decraemer (1978); I, Decraemer (1987); J, T. (T.) disparseta sp. nov.; K, Timm (1970); L, Decraemer (1978); M, Decraemer (1983); N, Blome (1982); O, Decraemer (1987); P, Timm (1970); Q, Blome (1982); R, Chitwood (1951); S, Decraemer (1986); T, Decraemer (1979); U, Lee, Lee & Rho (2023).
Figure 10.
Pictorial key to the species group with 50 to 64 main rings in the subgenus Tricoma. Source of figures: A, Timm (1970); B, Decraemer (1979); C, Freudenhammer (1975); D, Decraemer (1987); E, Decraemer (1987); F, Decraemer (1987); G, Timm (1970); H, Decraemer (1978); I, Decraemer (1987); J, T. (T.) disparseta sp. nov.; K, Timm (1970); L, Decraemer (1978); M, Decraemer (1983); N, Blome (1982); O, Decraemer (1987); P, Timm (1970); Q, Blome (1982); R, Chitwood (1951); S, Decraemer (1986); T, Decraemer (1979); U, Lee, Lee & Rho (2023).
Female. Unknown.
Distribution: Tricoma (Tricoma) longirostris Southern, 1914) has been documented in a variety of geographic locations, including the Atlantic coast of Ireland [28], the west coast of Africa [29], Antarctica [26], the Mozambique Channel [20], the Bay of Bengal in Indian waters [30], and most recently in the Northwest Pacific Ocean (this study). This broad distribution suggests a high level of adaptability to diverse marine environments across different climatic zones.
Discussion. Desmoscolex longirostris was first described by Southern (1914) based on two male specimens from the Atlantic coast of Ireland (Figure 8E-G). Southern’s description lacked details on the somatic setae pattern and characterized the species by its wedge-shaped head [28]. Later, Steiner (1916) transferred this species to the genus Tricoma, noting that Southern's description was insufficiently detailed to make a definitive assessment [29]. Steiner also described a new species, Tricoma glutinosa Steiner, 1916, based on two male specimens from the west coast of Africa (Figure 8H). He distinguished it from T. longirostris based on similarities in general habitus and head shape but noted significant differences in the number of main rings (77 vs. 70), body length (250 µm vs. 650 µm), and gubernaculum morphology. However, Freudenhammer (1975) observed that Steiner's description of male individuals of Tricoma glutinosa exhibited characteristics typical of the species but considered it a 'species inquirenda' pending further investigation [27]. Subsequently, in 1983, Decraemer noted that the gubernaculum of T. longirostris had been depicted in an oblique ventral position, potentially obscuring the complete structure [20]. She also suggested that the two male specimens of T. glutinosa might correspond to T. longirostris, given the observed variability in the number of main rings and body length between the specimens (Figure 8K, L). In addition to these findings, four male specimens of T. septentrionalis collected in Antarctica by Timm in 1978 were also reclassified by Decraemer as T. longirostris (Figure 8I, J) [20]. More recently, Ansari, Lyla, and Ajmal Khan (2015) reported T. longirostris from the Bay of Bengal continental shelf in Indian waters, based on 35 specimens [30].
Discussion. Desmoscolex longirostris was first described by Southern (1914) based on two male specimens from the Atlantic coast of Ireland (Figure 8E-G). Southern’s description lacked details on the somatic setae pattern and characterized the species by its wedge-shaped head [28]. Later, Steiner (1916) transferred this species to the genus Tricoma, noting that Southern's description was insufficiently detailed to make a definitive assessment [29]. Steiner also described a new species, Tricoma glutinosa Steiner, 1916, based on two male specimens from the west coast of Africa (Figure 8H). He distinguished it from T. longirostris based on similarities in general habitus and head shape but noted significant differences in the number of main rings (77 vs. 70), body length (250 µm vs. 650 µm), and gubernaculum morphology. However, Freudenhammer (1975) observed that Steiner's description of male individuals of Tricoma glutinosa exhibited characteristics typical of the species but considered it a 'species inquirenda' pending further investigation [27]. Subsequently, in 1983, Decraemer noted that the gubernaculum of T. longirostris had been depicted in an oblique ventral position, potentially obscuring the complete structure [20]. She also suggested that the two male specimens of T. glutinosa might correspond to T. longirostris, given the observed variability in the number of main rings and body length between the specimens (Figure 8K, L). In addition to these findings, four male specimens of T. septentrionalis collected in Antarctica by Timm in 1978 were also reclassified by Decraemer as T. longirostris (Figure 8I, J) [20]. More recently, Ansari, Lyla, and Ajmal Khan (2015) reported T. longirostris from the Bay of Bengal continental shelf in Indian waters, based on 35 specimens [30].
A summary of the diagnostic characteristics of various populations tentatively identified as T. (T.) longirostris is provided in Table 4. However, apart from the number of main rings, tail rings, and body length, other morphological features are either insufficiently detailed, inconsistent, or exhibit variability. A review of the diagnostic traits described in the available literature suggests that T. (T.) longirostris can be recognized by its slender body with 63 to 78 main rings, a long and narrow head shape, the specific arrangement of the somatic setae, and a gubernaculum featuring a stout, knobbed apophysis.
The current specimen of T. (T.) longirostris was discovered in sediment collected from a deep-sea seamount in northeastern Guam. Although only a single individual was found, it exhibits key diagnostic features consistent with T. (T.) longirostris, including 78 main rings, a long and narrow head, 8 subdorsal and 13 to 14 subventral setae, relatively large pigment spots, and a gubernaculum with a knobbed apophysis. However, this specimen displays slightly shorter cephalic setae relative to the head diameter, which differs from typical observations of the species. Additionally, while Decraemer (1983) described males with spine-like preanal structures, these were not observed in the present specimen [20].

Table 3.
Comparison of diagnostic morphological characters among species groups with 50-65 main rings in the subgenus Tricoma. Morphometric values are rounded. Values marked with a dash (-) indicate unknown measurements.
Table 3.
Comparison of diagnostic morphological characters among species groups with 50-65 main rings in the subgenus Tricoma. Morphometric values are rounded. Values marked with a dash (-) indicate unknown measurements.
| Species | Characters | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| males | females | head diameter | head length | cephalic setae length | spicules length | gubernaculum length | Vulva (ring) | number of tail ring (males) |
number of tail ring (females) |
||||||
| body length | body rings | setae pattern (subdorsal/subventral) |
body length |
body rings |
setae pattern (subdorsal/subventral) |
||||||||||
| T. (T.) absidata Timm, 1970 | 485-520 | 57-60 | 11,12/19,19 | 510-550 | 59-60 | 11,12/17,19 | 22 | 16 | 18-19 | 42-45 | - | 27 | 9-10 | 9-10 | |
| T. (T.) absidata lizardiensis Decraemer, 1979 | 525-710 | 55-57 | 13,13/17,19 | 540-710 | 56-60 | 13,13/21,23 | 25-29 | 16-21 | 21-27 | 42-48 | 24-28 | 24-27 | 9 | 8 | |
| T. (T.) atlantica Freudenhammer, 1975 | 450 | 58 | 7,7/11,11 | 490 | 50 | - | 18-20 | 20-22 | 20-22 | 26 | - | 28-29 | 9 | 9 | |
| T. (T.) bipapillata Decraemer, 1987 | 185-205 | 48-55 | 9/14-15 | - | - | - | 11-12 | 8-9 | 10-11 | 25-32 | 12-13 | - | 7-9 | - | |
| T. (T.) capitata Decraemer, 1987 | 225 | 55-56 | 9/12-13 | 215-260 | 53-57 | 9/11-14 | 11-14 | 9.5-11 | 11-14 | 52 | 12 | 34-35 | 9 | 6-8 | |
| T. (T.) coralicolla Decraemer, 1987 | 185 | 56-57 | 9/12 | 220 | 58 | 9/11 | 11-13 | 11 | 12-14 | 14 | 10 | 33 | 8 | 8 | |
| T. (T.) denticulata Timm, 1970 | 645-800 | 63-64 | 10/15-17 | 760 | 65-66 | - | 32-39 | 19-26 | 20-22 | 96-104 | 50-56 | 43 | 11-12 | 9 | |
| T. (T.) dimorpha Decraemer, 1978 | 305-600 | 52-65 | 12-13/16-17 | 400 | 62-67 | 13/17 | 15-20 | 11-16 | 15-21 | 16-27 | 16-19 | 27-29 | 10-12 | 12-13 | |
| T. (T.) dimorpha papuensis Decraemer, 1987 | 175-210 | 48-55 | 9/10-11 | 180-265 | 48-56 | 8-13/11-15 | 9.5-12 | 8-11 | 9-12 | 14-17 | 8.5-10.5 | 26-33 | 8-11 | 5-10 | |
| T. (T.) disparseta sp. nov. | 527-613 | 59-61 | 9-10/14-18 | 490-575 | 60-62 | 9/16-18 | 19-22 | 15-18 | 22-28 | 29-32 | 16-19 | 38-40 | 9-10 | 8-9 | |
| T. (T.) fisheri Timm, 1970 | 300-360 | 58-61 | 8-9/10-17 | 275-425 | 58-61 | 8-11/13-16 | 18-20 | 10-14 | 13-17 | 26-38 | 13-29 | 31-32 | 8-9 | 7-8 | |
| T. (T.) goldeni Decraemer, 1978 | 310-320 | 55-59 | 12/15-16 | - | - | - | 14 | 11 | 12-13 | 21-22 | 13 | - | 11-12 | - | |
| T. (T.) longirostris (Southern, 1914) | 650 | 70 | - | - | - | - | 32 | 31 | - | - | - | - | - | - | |
| T. (T.) oblita Blome, 1982 | 477-506 | 60-63 | 8-9/15-16 | 513 | 63 | 10/18 | 25-27 | - | 21-22 | 27 | 11-13 | 37-38 | 9-10 | - | |
| T. (T.) paratimmi Decraemer, 1987 | 385-420 | 61 | 11-12/16-17 | 390-450 | 53-56 | 8-13/15-17 | 13-14 | 20-22 | 11-16 | 28-30 | 17-20 | 27-29 | 9 | 5-6 | |
| T. (T.) perparvula Timm, 1970 | 275 | 61-62 | 7,9/13,13 | - | - | - | 14 | 11 | 11 | 24 | 9 | - | 9 | - | |
| T. (T.) secunda Blome, 1982 | 328 | 56-57 | 9,9/10, 13 | - | - | - | 18 | - | 16 | 22 | 8 | - | 9 | - | |
| T. (T.) spinosoides Chitwood, 1951 | 400 | 61 | 10/17 | 380 | - | 10/14 | - | - | - | 26 | 13 | 26 | - | 12 | |
| T. (T.) spuria Inglis, 1967 | 710 | 62 | 11/21 | 735 | 62 | 10-11/18-19 | 27-28 | 18-21 | 29 | 46 | 31 | 33 | 10 | 9 | |
| T. (T.) steineri de Man, 1922 | 408 | 66 | 13/16 | 432-460 | - | - | 13.6 | 12 | - | 27 | - | - | 12 | - | |
| T. (T.) ulleungensis Lee, Lee & Rho, 2023 | 409-415 | 54-55 | 6-7/10-12 | 462-567 | 55-57 | 6-7/9-10 | 24-26 | 14-16 | 19-22 | 22-24 | 11-13 | 31 | 8-9 | 8-9 | |
Table 4.
Comparison of diagnostic features among different populations identified as Tricoma (Tricoma) longirostris. Values marked with a dash (-) indicate unknown measurements.
Table 4.
Comparison of diagnostic features among different populations identified as Tricoma (Tricoma) longirostris. Values marked with a dash (-) indicate unknown measurements.
| Characters | Southern (1914) | Steiner (1916) | Timm (1978) | Decraemer (1983) | Ansari et al. (2015) | This study |
|---|---|---|---|---|---|---|
| Specimens | 2 males | 1 male | 4 males | 2 males | 16 males, 19 females | 1 male |
| Body length in male | 650 | 250 | 780-847 | 300-420 | 600-900 | 808 |
| Body length in female | - | - | - | - | 700-1,000 | - |
| Number of body ring | 70 | 77 | 71-77 | 63-72 | 70-78 | 78 |
| Width of body | 39 | 14 | 32-39 | 19-21 | 44-57 | 30 |
| Length of head | 31 | - | 25 | 12-16 | - | 26 |
| Width of head | 32 | - | 27 | 15-19 | - | 28 |
| Cephalic seta | - | - | 28-32 | 17-21 | 21-23 | 23 |
| Esophagus ring | - | - | - | 11 | - | 10 |
| Spicule length | - | - | 30 | 32-35 | 19-23 | 28 |
| Length of gubernaculum | - | - | - | 14-15 | - | 16 |
| Number of tail rings | - | 10 | 10-11 | 11-12 | - | 12 |
| Tail length | - | 43 | 112-119 | 60-84 | - | 125 |
| Somatic setae pattern (subdorsal/subventral) |
- | - | 8-9/12-15 | 9/12-13 | - | 8,8/13,14 |
| Length of terminal ring | - | - | 22-58 | 16-23 | - | 38 |
| locality | Clew bay, Atlantic coast of Ireland, 24 fms, bottom of sand and shells, | Prampram, Gold coast, The west coast of Africa, 9 m | Mcmurdo sound, Antarctica, Hut point, 4457 m, Scott Base, 540 m, |
Mocambique Channel | Bay of Bengal continental shelf, southeast coast of India, 30-176 m, sandy silt sediment | Northeastern of Guam, Pacific Ocean, 1425.52 m, sediment |
4. Conclusions
This study expands our knowledge of marine desmoscolecid nematodes in the deep-sea environments of the Northwest Pacific Ocean, specifically off northeastern Guam. Two species belonging to the subgenus Tricoma were documented, including the newly described Tricoma (Tricoma) disparseta sp. nov. This new species, collected from sponge and starfish habitats at depths ranging from 1300 to 1500 meters, is distinguished by several unique morphological features, such as 59 to 62 main rings, asymmetrical somatic setae distribution with 9 to 10 subdorsal setae and 14 to 18 subventral setae on each side, and a distinctive curved, hooked gubernaculum. These characteristics set T. (T.) disparseta apart from other congeners, highlighting its adaptation to deep-sea habitats. The other species observed in this study, T. (T.) longirostris, exhibits morphological features consistent with previously described specimens, such as 78 main rings, a long and narrow head, and a gubernaculum with a knobbed apophysis. The comprehensive documentation of these two species through differential interference contrast microscopy (DIC) and scanning electron microscopy (SEM) provides detailed morphological illustrations and insights into their structural variations. The inclusion of pictorial keys and comparative tables for species with 50 to 64 main rings serves as a valuable taxonomic tool, facilitating the identification and differentiation of closely related species within the subgenus Tricoma. These findings contribute to our understanding of the diversity and distribution of deep-sea nematodes and underscore the importance of continued exploration and documentation of marine biodiversity in these remote and understudied habitats.
Author Contributions
Data curation and Writing-Original draft preparation., H.J.L.; Investigation, H.L. and J.-H.K.; Writing-Reviewing, Editing and Funding acquisition, H.S.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by High seas bioresources program of Korea Institute of Marine Science & Technology Promotion (KIMST) funded by the Ministry of Oceans and Fisheries (KIMST-20210646).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
All data are available in the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Danovaro, R.; Snelgrove, P.V.; Tyler, P. Challenging the paradigms of deep-sea ecology. Trends in ecology & evolution 2014, 29, 465-475. [CrossRef]
- Dell’Anno, A.; Carugati, L.; Corinaldesi, C.; Riccioni, G.; Danovaro, R. Unveiling the biodiversity of deep-sea nematodes through metabarcoding: are we ready to bypass the classical taxonomy? PLoS One 2015, 10, e0144928. [CrossRef]
- Armenteros, M.; Quintanar-Retama, O.; Gracia, A. Depth-related patterns and regional diversity of free-living nematodes in the deep-sea Southwestern Gulf of Mexico. Frontiers in Marine Science 2022, 9, 1023996. [CrossRef]
- Trebukhova, Y.A.; Miljutin, D.; Pavlyuk, O.; Mar'yash, A.; Brenke, N. Changes in deep-sea metazoan meiobenthic communities and nematode assemblages along a depth gradient (North-western Sea of Japan, Pacific). Deep Sea Research Part II: Topical Studies in Oceanography 2013, 86, 56-65. [CrossRef]
- dos Santos, G.A.; Silva, A.C.; Esteves, A.M.; Ribeiro-Ferreira, V.P.; Neres, P.F.; Valdes, Y.; Ingels, J. Testing bathymetric and regional patterns in the southwest Atlantic deep sea using infaunal diversity, structure, and function. Diversity 2020, 12, 1-25.
- Udalov, A.; Azovsky, A.; Mokievsky, V. Depth-related pattern in nematode size: what does the depth itself really mean? Progress in Oceanography 2005, 67, 1-23. [CrossRef]
- Danovaro, R.; Gambi, C. Cosmopolitism, rareness and endemism in deep-sea marine nematodes. The European Zoological Journal 2022, 89, 653-665. [CrossRef]
- Bezerra, T.N.; Pape, E.; Hauquier, F.; Vanreusel, A. Description and distribution of Erebussau nom. nov. pro Erebus Bussau, 1993 nec Erebus Latreille, 1810 with description of a new species, and of Odetenema gesarae gen. nov., sp. nov.(Nematoda: Desmoscolecida) from nodule-bearing abyssal sediments in the Pacific. Zootaxa 2021, 4903, 542-562. [CrossRef]
- Lee, H.J.; Lee, H.; Rho, H.S. Six species of Tricoma (Nematoda, Desmoscolecida, Desmoscolecidae) from the East Sea, Korea, with a bibliographic catalog and geographic information. Korean Journal of Environmental Biology 2023, 41, 570-607. [CrossRef]
- Decraemer, W.; Rho, H.S. Order Desmoscolecida. In handbook of zoology: Gastrotricha, Cycloneuralia and Gnathifera, vol. 2: Nematoda; De Gruyter: Berlin/Boston, 2013; pp. 351-372.
- Rogers, A. The biology of seamounts. In Advances in marine biology; Academic Press: 1994; Volume 30, pp. 305-350.
- Clark, M.R.; Rowden, A.A.; Schlacher, T.; Williams, A.; Consalvey, M.; Stocks, K.I.; Rogers, A.D.; O'Hara, T.D.; White, M.; Shank, T.M. The ecology of seamounts: structure, function, and human impacts. Annual Review of Marine Science 2010, 2, 253-278. [CrossRef]
- Piepenburg, D.; Müller, B. Distribution of epibenthic communities on the Great Meteor Seamount (North-east Atlantic) mirrors pelagic processes. Archive of Fishery and Marine Research 2004, 51, 55-70.
- White, M.; Bashmachnikov, I.; Arístegui, J.; Martins, A. Physical processes and seamount productivity; Blackwell Publishing: Oxford, UK 2007; pp. 65-84.
- Zeppilli, D.; Bongiorni, L.; Cattaneo, A.; Danovaro, R.; Santos, R.S. Meiofauna assemblages of the Condor Seamount (North-East Atlantic Ocean) and adjacent deep-sea sediments. Deep Sea Research Part II: Topical Studies in Oceanography 2013, 98, 87-100.
- Rowden, A.A.; Schlacher, T.A.; Williams, A.; Clark, M.R.; Stewart, R.; Althaus, F.; Bowden, D.A.; Consalvey, M.; Robinson, W.; Dowdney, J. A test of the seamount oasis hypothesis: seamounts support higher epibenthic megafaunal biomass than adjacent slopes. Marine Ecology 2010, 31, 95-106. [CrossRef]
- Decraemer, W. Morphological and taxonomic study of the genus Tricoma Cobb (Nematoda: Desmoscolecida), with the description of new species from the Great Barrier Reef of Australia. Australian Journal of Zoology 1978, 26, 1-121. [CrossRef]
- Decraemer, W. Desmoscolecids from sublittoral finesand of Pierre Noire (West Channel)(Nematoda, Desmoscolecida). Bulletin du Muséum national d'histoire naturelle 1979, 1, 299-321.
- Decraemer, W. Morphology of Tricoma absidata lizardiensis subsp. nov.(Nematoda, Desmoscolecida) with a note on its ontogeny. Biologisch Jaarboek Dodonaea 1979, 46, 101-114.
- Decraemer, W. Tricominae (Nematoda-Desmoscolecida) from the northern part of Moçambique Channel, with five new species and one new genus. Bulletin de l'Institut Royal des Sciences Naturelles de Belgique Biologie 1983, 55, 1-34.
- Decraemer, W. Tricominae (Nematoda: Desmoscolecida) from Laing Island, Papua New Guinea, with Descriptions of New Species. Invertebrate Systematics 1987, 1, 231-256. [CrossRef]
- Decraemer, W. Descriptions of two new species of Tricoma (Nematoda: Desmoscolecidae) and comments on the taxonomic status of T.(T.) tertia Blome, 1982 and T.(T.) brevirostris (Southern, 1914) Steiner, 1916. Russian Journal of Nematology 1996, 4, 107-114.
- Decraemer, W.; Tchesunov, A.V. Some Desmoscolecids from the White Sea. Russian Journal of Nematology 1996, 4, 15-130.
- Soetaert, K.; Decraemer, W. Eight new Tricoma species (Nematoda, Desmoscolecidae) from a deep-sea transect off Calvi (Corsica, Mediterranean). Hydrobiologia 1989, 183, 223-247.
- Timm, R.W. A revision of the nematode order Desmoscolecida Filipjev, 1929. University of California publications in zoology 1970, 93, 1-115.
- Timm, R.W. Marine nematodes of the order Desmoscolecida from McMurdo Sound, Antarctica. Biology of the Antarctic Seas 1978, 26, 225-236.
- Freudenhammer, I. Desmoscolecida aus der Iberischen Tiefsee, zugleich eine Revision dieser Nematoden-ordnung. 1975.
- Southern, R. Clare Island Survey. Nemathelmia, Kinorhyncha and Chaetognatha. In Proceedings of the Proceedings of the Royal Irish Academy, 1914; pp. 1-80.
- Steiner, G. Neue und wenig bekannte Nematoden von der Westküste Afrikas I; Zoologischer Anzeiger: 1916; Volume 47.
- Ansari, K.; Lyla, P.; Ajmal Khan, S. New distributional records of free-living marine nematodes from Indian waters I. Chromadorids. Indian Journal of Geo-Marine Science 2015, 44, 756-765.
Figure 1.
A map depicting the locations where samples were collected.
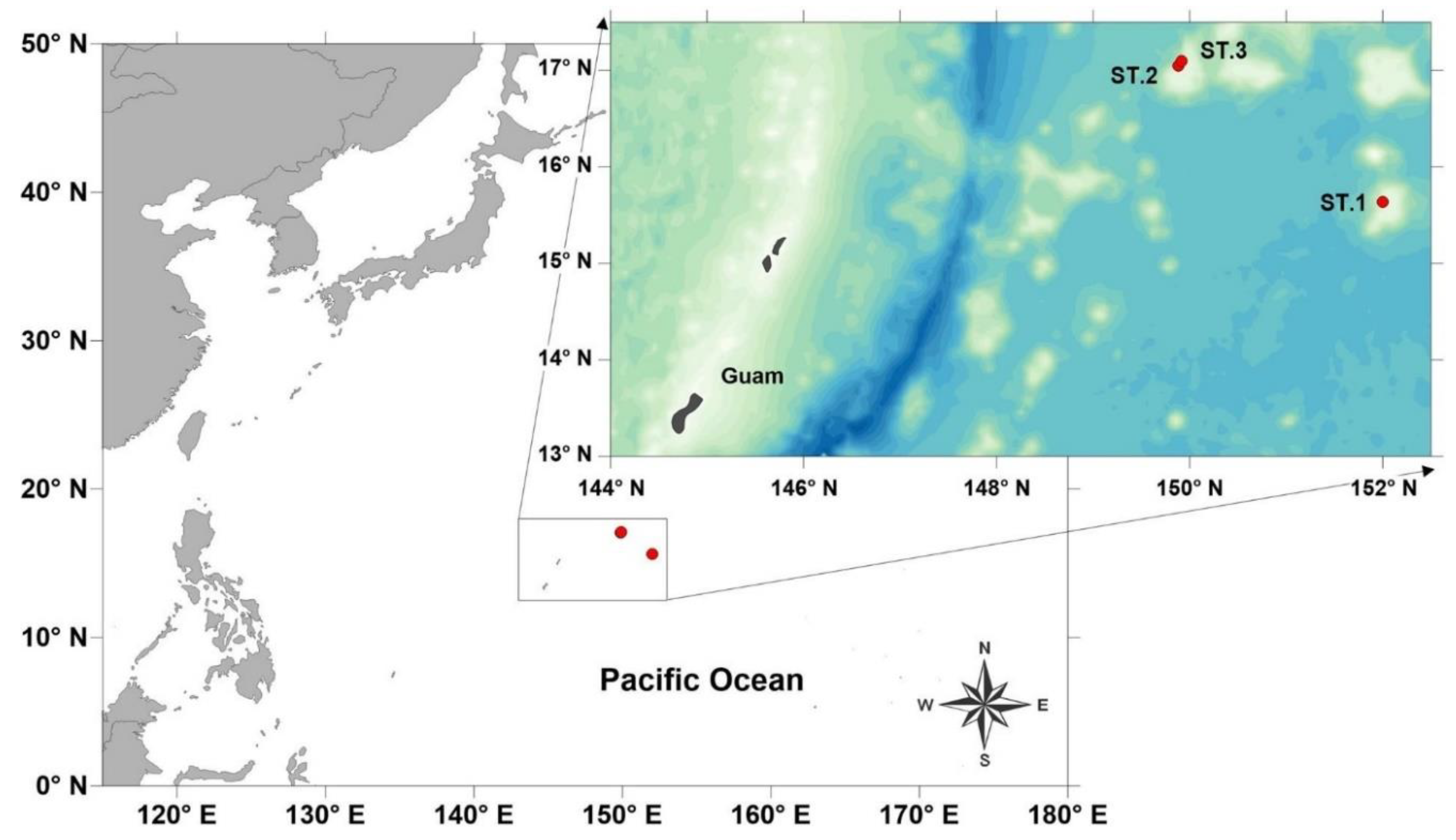
Figure 2.
Tricoma (Tricoma) disparseta sp. nov. Holotype male (MNB001). A, habitus, lateral view; B, head region, left side; C, head region, ventral view (Paratype MNB006); D, head region, right side (Paratype MNB005); E, spicules and tail region, right side (Paratype MNB005); E, posterior region showing sightly laterally inserted subventral setae. Scale bars: 50 µm in A; 10 µm in B-E; 20 µm in F.
Figure 2.
Tricoma (Tricoma) disparseta sp. nov. Holotype male (MNB001). A, habitus, lateral view; B, head region, left side; C, head region, ventral view (Paratype MNB006); D, head region, right side (Paratype MNB005); E, spicules and tail region, right side (Paratype MNB005); E, posterior region showing sightly laterally inserted subventral setae. Scale bars: 50 µm in A; 10 µm in B-E; 20 µm in F.
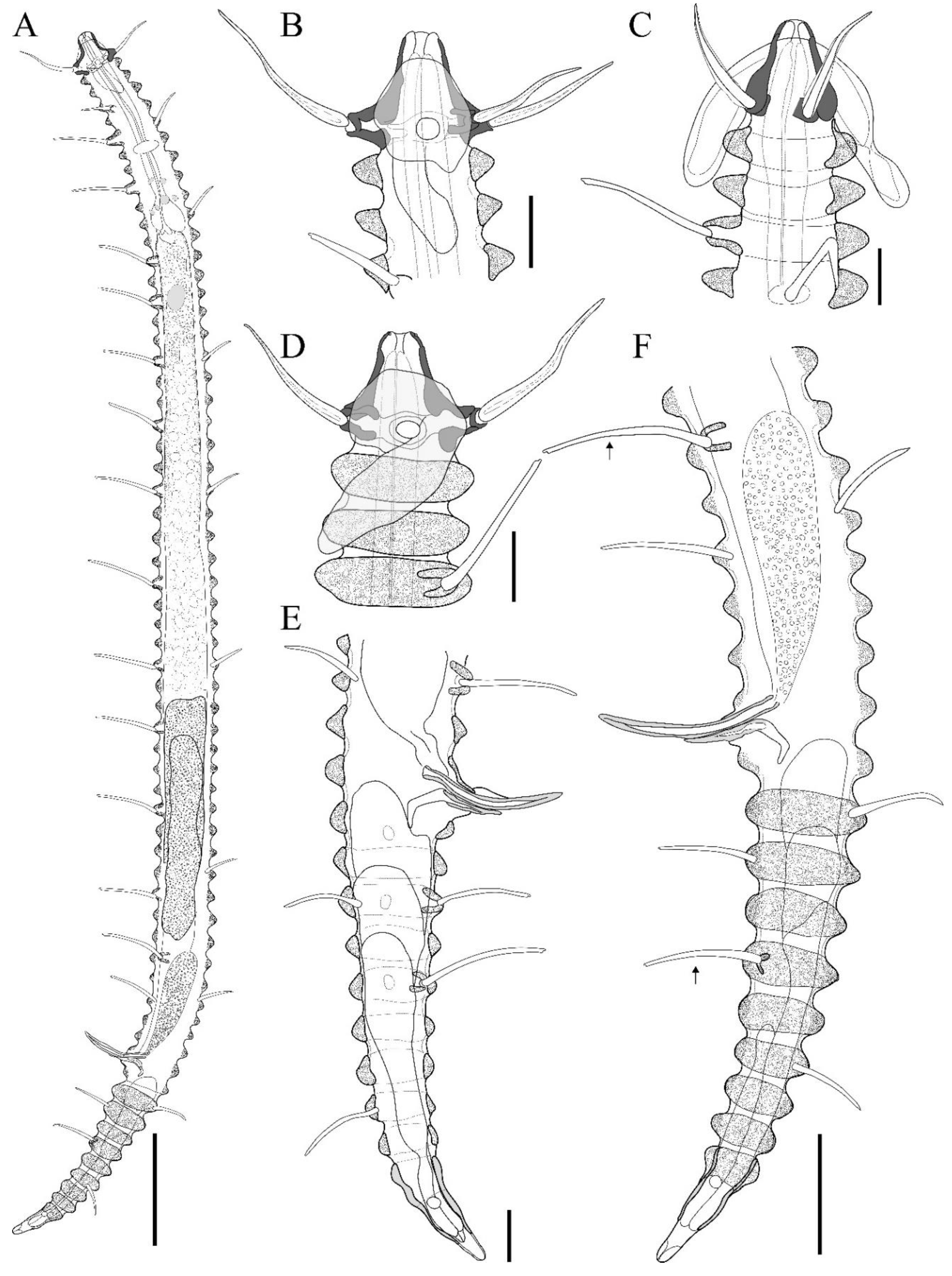
Figure 3.
Tricoma (Tricoma) disparseta sp. nov. Allotype female (MNB007). A, habitus, lateral view; B, head region, right side; C, head region, right side (Paratype MNB008); D, reproductive systems; E, tail region, right side. Scale bars: 50 µm in A; 10 µm in B-C; 20 µm in D-E.
Figure 3.
Tricoma (Tricoma) disparseta sp. nov. Allotype female (MNB007). A, habitus, lateral view; B, head region, right side; C, head region, right side (Paratype MNB008); D, reproductive systems; E, tail region, right side. Scale bars: 50 µm in A; 10 µm in B-C; 20 µm in D-E.
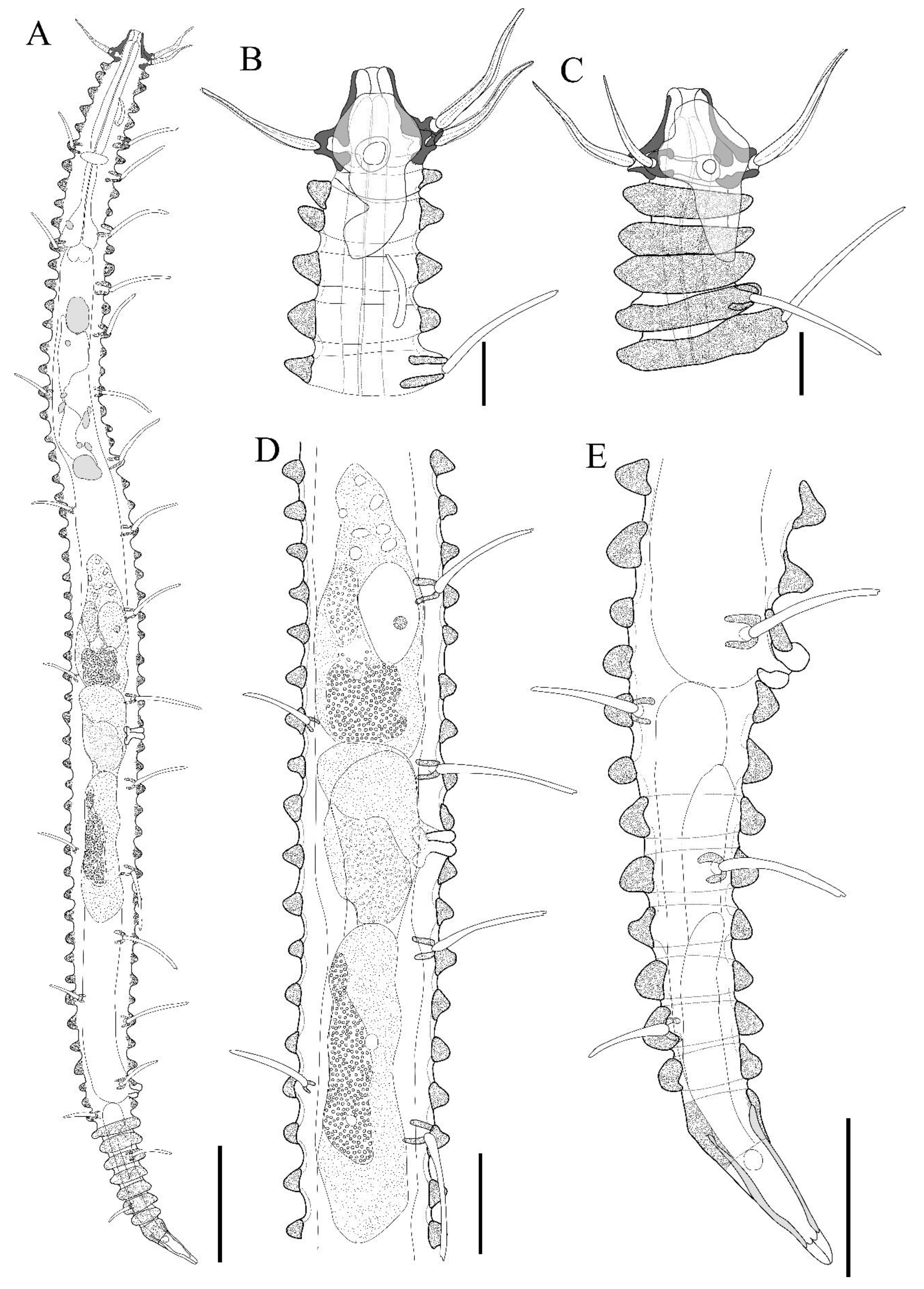
Figure 4.
Tricoma (Tricoma) disparseta sp. nov. SEM photomicrographs, male. A, entire view of the body, lateral view; B, anterior region showing laterally inserted subventral setae (white arrow); C, head showing the amphideal fovea, with an white arrow indicating the amphideal pore, dorsal view; D, head region, anterior view; E, cephalic setae enclosed by a thin membrane; F, distal end of cephalic setae, split and enclosed by a thin membrane. Scale bars: 100 µm in A; 10 µm in B; 5 µm in C-E; 500 nm in F.
Figure 4.
Tricoma (Tricoma) disparseta sp. nov. SEM photomicrographs, male. A, entire view of the body, lateral view; B, anterior region showing laterally inserted subventral setae (white arrow); C, head showing the amphideal fovea, with an white arrow indicating the amphideal pore, dorsal view; D, head region, anterior view; E, cephalic setae enclosed by a thin membrane; F, distal end of cephalic setae, split and enclosed by a thin membrane. Scale bars: 100 µm in A; 10 µm in B; 5 µm in C-E; 500 nm in F.

Figure 5.
Tricoma (Tricoma) disparseta sp. nov. SEM photomicrographs, male. A, cuticular layer showing the height difference between the dorsal and ventral peduncles; B, subdorsal setae; C, subventral setae appearing with curved tip; D, spicules region, lateral view; E, spicules region, anterior view; F, terminal ring. Scale bars: 10 µm in A, C, D; 5 µm in B, E, F.
Figure 5.
Tricoma (Tricoma) disparseta sp. nov. SEM photomicrographs, male. A, cuticular layer showing the height difference between the dorsal and ventral peduncles; B, subdorsal setae; C, subventral setae appearing with curved tip; D, spicules region, lateral view; E, spicules region, anterior view; F, terminal ring. Scale bars: 10 µm in A, C, D; 5 µm in B, E, F.

Figure 6.
Tricoma (Tricoma) disparseta sp. nov., DIC photomicrographs, Holotype male (MNB001). A, entire view of body; B, anterior region; C, amphideal fovea; D, posterior region; E, head region, ventral view (Paratype MNB006); F, somatic setae; G, spicules and gubernaculum of the specimen treated with lactic acid, which was additionally used to increase transparency for the observation of internal reproductive organs (Paratype MNB003); H, terminal ring showing phasmata (Paratype MNB004). Scale bars: 50 µm in A; 10 µm in B-H.
Figure 6.
Tricoma (Tricoma) disparseta sp. nov., DIC photomicrographs, Holotype male (MNB001). A, entire view of body; B, anterior region; C, amphideal fovea; D, posterior region; E, head region, ventral view (Paratype MNB006); F, somatic setae; G, spicules and gubernaculum of the specimen treated with lactic acid, which was additionally used to increase transparency for the observation of internal reproductive organs (Paratype MNB003); H, terminal ring showing phasmata (Paratype MNB004). Scale bars: 50 µm in A; 10 µm in B-H.

Figure 7.
Tricoma (Tricoma) disparseta sp. nov., DIC photomicrographs, Allotype female (MNB007). A, entire view of body; B, head region; C, amphideal fovea; D, midbody region showing naked vulva; E, posterior region; H, entire view of body showing somatic setae (Paratype MNB008). Scale bars: 50 µm in A, F; 10 µm in B-E.
Figure 7.
Tricoma (Tricoma) disparseta sp. nov., DIC photomicrographs, Allotype female (MNB007). A, entire view of body; B, head region; C, amphideal fovea; D, midbody region showing naked vulva; E, posterior region; H, entire view of body showing somatic setae (Paratype MNB008). Scale bars: 50 µm in A, F; 10 µm in B-E.

Table 1.
Sampling locations of the two Tricoma species collected from the Northwest Pacific Ocean.
| Stations | Date | Latitue (DMS) | Longitude (DMS) | Depth(m) | Remarks | Specimens | |
|---|---|---|---|---|---|---|---|
| St.1 | BKC90103 | September 1, 2023 | '15°38'15.16"S | '151°59'50.35"E | 1425.52 | Starfish habitat | MNB011 |
| St.2 | BKC80403 | September 6, 2023 | '17°02'47.346"S | '149°52'50.484"E | 1366.6 | Starfish habitat | MNB004,MNB005, MNB006, MNB007, MNB009, MNB010 |
| St.3 | BKC80502 | September 7, 2023 | '17°05'45.41"S | '149°55'07.67"E | 1510.1 | Sponge | MNB001, MNB002, MNB003, MNB008 |
Table 2.
Morphometric measurements of Tricoma (Tricoma) disparseta sp. nov. (in micrometers, µm).
| Holotype male | Paratype male 1 | Paratype male 2 | Paratype male 3 | Paratype male 4 | Allotype female | Paratype female 1 | Paratype female 2 | Paratype female 3 | |
|---|---|---|---|---|---|---|---|---|---|
| Total body length | 571 | 613 | 527 | 551 | 556 | 543 | 490 | 575 | 543 |
| Number of body ring | v:60 d:61 |
59 | 60 | 60 | v:60 d:61 |
v:60 d:61 |
v:62 d:61 |
60 | 61 |
| a | 19 | 18 | 15 | 18 | 16 | 16 | 12 | 14 | 18 |
| b | 6 | 6 | 6 | 6 | 6 | 6 | 7 | 7 | 6 |
| c | 6 | 6 | 6 | 6 | 6 | 7 | 7 | 7 | 6 |
| Head length | 16 | 17 | 15 | 18 | 17 | 16 | 15 | 17 | 15 |
| Head diameter at the level of cephalic setae | 21 | 20 | 20 | 19 | 21 | 19 | 21 | 21 | 19 |
| Body diameter at the level of cardia | 24 | 27 | 25 | 26 | 29 | 28 | 28 | 27 | 25 |
| Maximum body diameter | 30 | 35 | 34 | 31 | 34 | 35 | 42 | 41 | 31 |
| Cephalic setae length | 25 | 22 | 28 | 24 | 23 | 22 | 25 | 23 | 22 |
| Amphideal fovea length | 25 | 26 | 23 | 26 | 27 | 27 | 27 | 22 | 27 |
| Ocelli diameter | 8 | 5 | 8 | 7 | 4 | 10 | 7 | 7 | 4 |
| Ocelli length | 11 | 5 | 7 | 7 | 13 | 14 | 9 | 4 | 5 |
| Anterior end to ocelli | 123 | 114 | 171 | 108 | 118 | 119 | 148 | 170 | 107 |
| Pharynx length | 95 | 97 | 93 | 96 | 94 | 92 | 73 | 88 | 86 |
| Number of subventral setae (left/right) | 17/18 | 16/17 | 17/16 | 18/14 | 18/17 | 17/17 | 17/18 | 16/16 | 17/17 |
| Length of the longest subventral setae | 32 | 36 | 30 | 34 | 35 | 35 | 34 | 38 | 31 |
| Length of the shortest subventral setae | 17 | 18 | 17 | 15 | 17 | 16 | 23 | 22 | 15 |
| Number of subdorsal setae (left/rightl) | 9/10 | 9/9 | 9/9 | 9/9 | 9/9 | 9/9 | 9/9 | 9/9 | 9/9 |
| Length of the longest subdorsal setae | 20 | 20 | 19 | 18 | 21 | 19 | 22 | 19 | 18 |
| Length of the shortest subdorsal setae | 17 | 18 | 17 | 15 | 17 | 12 | 17 | 13 | 13 |
| Spicule length | 31 | 29 | 32 | 30 | 29 | - | - | - | - |
| Gubernaculum length | 18 | 18 | 16 | 16 | 18 | - | - | - | - |
| Anterior end to vulva | - | - | - | - | - | 305 | 301 | 324 | 302 |
| Body diameter at level of vulva | - | - | - | - | - | 32 | 37 | 31 | 27 |
| V(%) | - | - | - | - | - | 56 | 61 | 56 | 56 |
| Anal body diameter | 25 | 29 | 26 | 24 | 27 | 23 | 26 | 26 | 25 |
| Tail length | 96 | 95 | 89 | 92 | 92 | 82 | 71 | 79 | 84 |
| Number of tail's body ring | 10 | 9 | 10 | 10 | 10 | 9 | 9 | 8 | 9 |
| Terminal ring length | 23 | 27 | 24 | 26 | 23 | 26 | 24 | 25 | 26 |
| Desmos covering terminal ring | 7 | 8 | 8 | 7 | 6 | 9 | 9 | 10 | 8 |
| Phasmata | 2.5 | 2.5 | 2.5 | 2.5 | 2.9 | 2.7 | 2.5 | 2.8 | 2.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



