
Submitted:
16 July 2025
Posted:
17 July 2025
You are already at the latest version
Abstract
The long-horned caddisfly Brachysetodes Schmid, 1955 (Leptoceridae: Leptocerinae) is a small genus endemic to the Andean region, especially Chile. Four decades after the last taxonomic revision, a new species, Brachysetodes tricahue Desidério, Santana & Hamada sp. nov., is described and illustrated based on adult males collected from Parque Natural Tricahue in the Central Subregion of the Chilean Andes. Specimens were examined through genital dissection using heated KOH, photographed with a digital camera mounted on microscopes, and described using the DELTA system. A distribution map was produced in QGIS based on GBIF data and literature records, and an updated identification key to males of the ten known species of Brachysetodes sensu stricto is presented. B. tricahue sp. nov. closely resembles B. bifurcatus and B. nublensis, sharing key features such as paired lateral processes on tergum X and tripartite inferior appendages. However, it can be distinguished by a unique combination of genital features, including unequal lengths of the three processes of the inferior appendage. This discovery emphasizes the underexplored diversity of the southern Andes and contributes to refining the taxonomy and biogeography of the genus. It also provides a framework for future phylogenetic studies incorporating immature stages and molecular data.

Keywords:
aquatic insects
; Leptocerinae
; taxonomy
; morphology
; phylogeny
; biogeography
; Chilean Andes
; Parque Natural Tricahue
1. Introduction
Leptoceridae Leach, 1815, commonly known as long-horned caddisflies, are the second-largest family among Trichoptera [1]. Currently, four subfamilies are recognized: Grumichellinae Morse, 1981, Leptocerinae Leach, 1815, Leptorussinae Morse, 1981, and Triplectidinae Ulmer, 1906 [2]. Among the Leptoceridae of the New World, Brachysetodes is placed within the Leptocerinae, considering both morphological [3] and molecular [2,4] phylogenies, although not within any specific tribes of this subfamily.
Historically, Brachysetodes Schmid, 1955 was established by Schmid [5] to include three Chilean species: Brachysetodes trifidus Schmid, 1955, the type species, B. bifidus Schmid, 1955, and B. quadrifidus Schmid, 1955. Subsequently, Schmid [6,7] described five additional species: Brachysetodes extensus Schmid, 1958, B. forcipatus Schmid, 1964, B. major Schmid, 1958, B. spinosus Schmid, 1958, and B. tripartitus Schmid, 1964. Further contributions by Flint [8,9,10,11] expanded the genus through the description of new species in the genus (B. bifurcatus Flint, 1983, B. insularis Flint, 1968, and B. nublensis Flint, 1969) and the transfer of B. duodecimpunctata (Navás, 1916) from another genus. However, B. duodecimpunctata and B. insularis were later removed from Brachysetodes by Holzenthal [12,13] and reassigned to new Neotropical genera as Achoropsyche duodecimpunctata (Navás, 1916) and Amphoropsyche insularis (Flint, 1968), respectively.
The first comprehensive revision of Brachysetodes was conducted by Holzenthal [14], who provided a detailed diagnosis of the genus, described the immature stages, and extended its known distribution from Chile into Argentina. Furthermore, a morphological phylogenetic analysis revealed Brachysetodes to be polyphyletic, corroborating Morse’s [3] earlier hypothesis. Under this framework, Brachysetodes comprises 10 species restricted to Morrone’s [15] Andean region [16], of which eight belong to the Brachysetodes sensu stricto (Figure 1A–B), while two, B. bifidus and B. major, are classified as incertae sedis [14].
Although Brachysetodes is a member of Leptocerinae widely distributed throughout the Andean region, particularly in Chile [16], a new species of the genus has been discovered nearly four decades after the last comprehensive revision. This study, therefore, aims to describe and illustrate this new species of Brachysetodes s. str. based on male adults collected in the Chilean Andes. In addition, a key is provided for the identification of males of the nine species of Brachysetodes s. str., along with brief phylogenetic and biogeographic remarks.
2. Materials and Methods
2.1. Study Area, Specimen Collection, Preparation, and Observation
Specimens were collected using a Malaise trap [17] in a tributary ravine of the Tricahue stream, located within Parque Natural Tricahue (Figure 1C–D), which lies in the Central subregion of the Maule region in the Chilean Andes (Figure 1A–B). Parque Natural Tricahue, located in the mountainous area of San Clemente commune, Chile, is an important site for biodiversity conservation. Positioned within a nationally recognized biodiversity hotspot, this private park underwent restoration following severe forest degradation in the 1990s. Today, it serves as a crucial refuge for native species, playing a significant role in preserving regional biodiversity [18]. Specimens were preserved in 80% ethanol and will be deposited in the Museo Nacional de Historia Natural, Santiago, Chile (MNNC), Coleção de Invertebrados, Instituto Nacional de Pesquisas da Amazônia, Manaus, Brazil (INPA), and the Coleção de Insetos Aquáticos “Prof. Dr. Cláudio Gilberto Froehlich”, Universidade Estadual Paulista, Assis, Brazil (CIASP), as listed in the examined material.
Specimen preparation followed standard methods described by [19], using a hot 10% KOH solution to diaphanize the abdominal tissues. Male genital structures, once clarified, were examined under a Leica DM5500 B compound microscope and stored according to the procedures outlined in Desidério et al. [20].
2.2. Illustrations and Map
Photographs of adults were taken using a Leica DMC4500 digital video camera mounted on a Leica M205A stereomicroscope, equipped with a LED illumination dome [21]. Leica Application Suite (LAS) software (version 4.10.0) was used to capture image stacks of each structure at different focal planes, which were then automatically combined into single images with greater depth of field using Helicon Focus® Pro stacking software (version 7.6.4) employing the following rendering method and parameters: method B (depth map), radius 18, and smoothing 2.
Male genitalia were photographed with a Leica DFC295 video camera attached to a Leica DM5500 B microscope. Image stacks were captured with Leica Application Suite (LAS) software (version 4.2.0) and served as templates for creating vector illustrations in Adobe Illustrator® facilitated by a graphic tablet and pen (Intuos CTL4100, Wacom Technology Co.). All photographs and illustrations were edited and assembled into plates using Adobe Photoshop®.
Distribution maps were generated using QGIS software (version 3.34.0-Prizren) using the shapefile of Romano [22] based on the Andean biogeographical regionalization proposed by Morrone [15]. The satellite imagery was obtained from Google Earth®. Distribution data for all Brachysetodes species were compiled from the Global Biodiversity Information Facility database [23], selecting only records for SPECIES “taxonRank”. For species lacking coordinates, locality data were georeferenced in Google Earth® based on information from the original literature.
2.3. Morphological Terminology, Description, and Key
Morphological terminology for head setal warts follows Oláh and Johanson [24]. Terminology for male genitalia adheres to Nielsen [25] and Schmid [5], as interpreted for Brachysetodes by Holzenthal [14]. Wing venation terminology is based on the Comstock-Needham system, as applied to Trichoptera by Mosely and Kimmins [26]. To ensure consistency and standardization in descriptive taxonomy, the species description was generated using the DELTA editor (Description Language for Taxonomy) software (version 1.02), based on a morphological character matrix. The identification key is adapted from Holzenthal [14] and focuses exclusively on males of Brachysetodes s. str., emphasizing genital characters; all referenced figures correspond to that work, except for those of the new species.
3. Results
3.1. Species Description
Brachysetodes tricahue Desidério, Santana & Hamada, sp. nov.
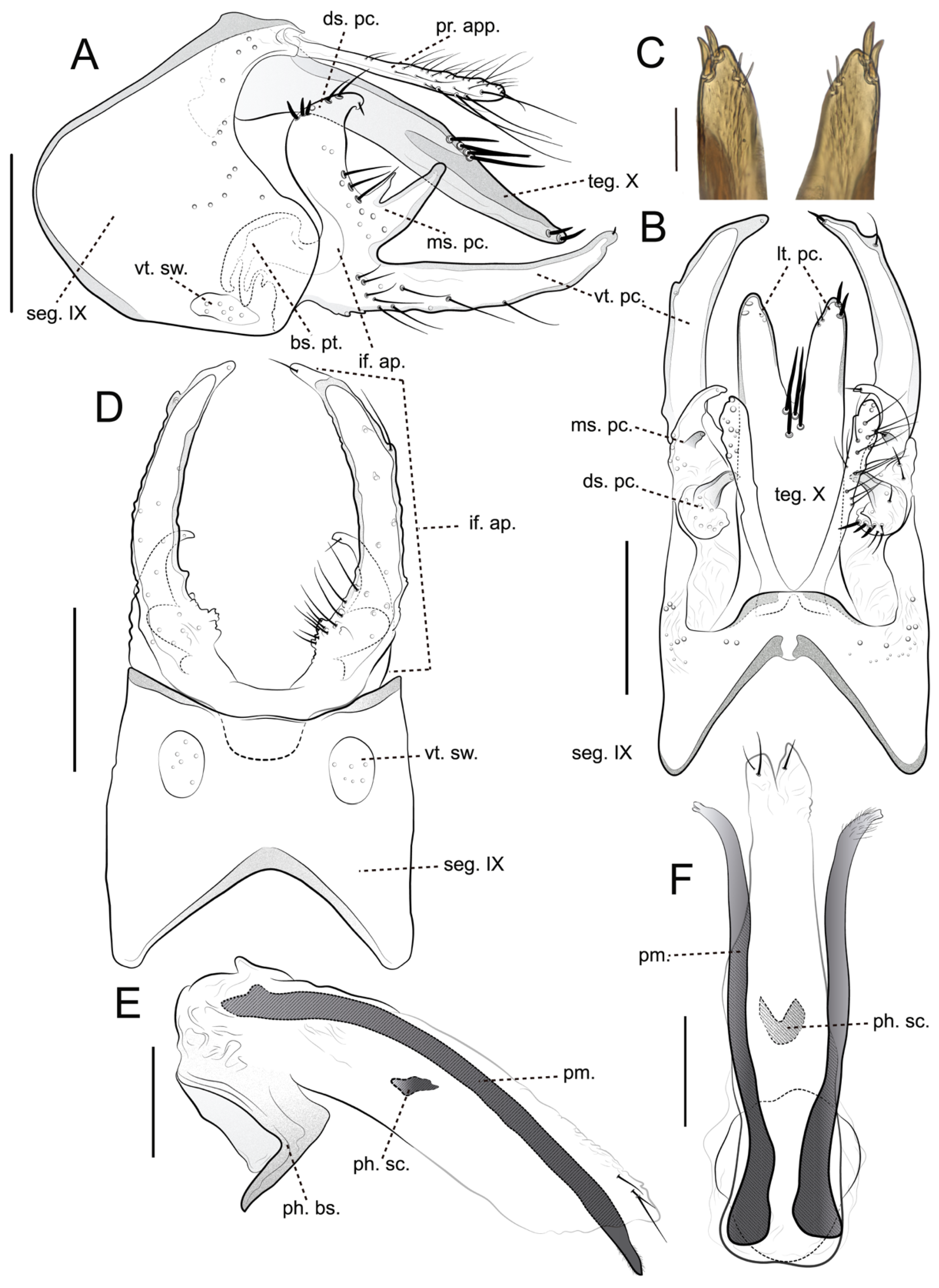
Diagnosis. Brachysetodes tricahue Desidério, Santana & Hamada, sp. nov. closely resembles B. bifurcatus and B. nublensis, particularly in the structure of tergum X and the inferior appendages. All three species share a pair of lateral processes on tergum X, each bearing a pair of medium-sized, stout apicolateral setae, as well as an inferior appendage divided into three processes. However, B. tricahue sp. nov. can be distinguished by the presence of four long, stout setae grouped at mid-length on tergum X, whereas B. bifurcatus and B. nublensis each possess only three setae, stout only in B. nublensis. In the new species, the inferior appendage bears three unequal processes, while in both B. bifurcatus and B. nublensis, these processes are subequal. Both B. tricahue sp. nov. and B. nublensis have a concave posterior margin of sternum IX, contrasting with the straight margin in B. bifurcatus. The mesal process of the inferior appendage is bifid in both B. tricahue sp. nov. and B. bifurcatus; however, in the former, the digitate lobes are unequal in length, whereas in the latter, they are equal. Another diagnostic feature of B. tricahue sp. nov. is the slender, elongate ventral process of the inferior appendage, which surpasses tergum X, with a basomesal margin that is crenulate and lacks projections or lobes. Differences in the phallic apparatus also support species separation: in B. tricahue sp. nov. and B. bifurcatus, the parameres are long, reaching the length of the phallic apparatus, whereas in B. nublensis, they are very short.
Description. Adult male: General coloration pale brown (in alcohol) (Figures 2A,D). Head dorsally with seven setal warts; medioantennal setal wart (ma. sw.) large, obovoid; lateroantennal pair (la. sw.) medium-sized, ellipsoid; occipital pair (occ. sw.) very large, obovoid; postgenal pair (pt. sw.) small, posterad to eye, rounded (Figure 2C); in frontal view, frontogenal pair (fg. sw.) very large, obovoid (Figure 2B). Frontal grooves (ft. g.) V-shaped anteromesally, well-pronounced; coronal groove (cn. g.) thread-shaped, weakly-pronounced on anterior half of head (Figure 2C). Maxillary palp length formula (I<II=III<IV<V) (damaged in holotype); labial segment length formula (I=II<III) (Figure 2B). Forewing, in alcohol, brown, with lighter anal area, without spots; forewing length 3.71–3.73 mm (n = 5); forks I and V present, both with short petiole; fork V about 2x longer than fork I; thyridial cell about as long as 2/3 of discoidal cell (Figure 2E). Hindwing with forks I and V, both with long petiole; fork V about long as fork I; r-m crossvein present (Figure 2F). Tibial spur formula 1:2:2.
Male genitalia: Segment IX, in lateral view, subtriangular, with anterodorsal margin almost straight; anteromesal margin convex, rounded, strongly produced; anteroventral margin straight; posterior margin produced basally, bearing apex truncated (Figure 3A); in dorsal view, anterior margin with shallow, wide V-shaped mesal incision, strongly sclerotized band extending posteriorly, but not touching the posterior margin; posterior margin produced mesally and straight (Figure 3B); in ventral view, anterior margin with wide V-shaped mesal incision; posterior margin slightly concave, bearing a pair of large, rounded setal warts (vt. sw.) (Figure 3D). Preanal appendages (pr. app.) originating dorsomesally, setose, slender, digitate, about ¾ length of tergum X in lateral and dorsal views (Figure 3A–B); in dorsal view, apex rounded, bearing long, thin setae (Figure 3B). Tergum X, in lateral view, narrowing apically, apex rounded (Figure 3A); in dorsal view, bearing four long, stout setae grouped at mid-length; divided by V-shaped apicomesal incision extending anteriorly about 2/3 length of the segment, originating pair of paired lateral process (lt. pc.) (Figure 3B); each lateral process (lt. pc.) slender, bearing one pair of short, stout subapical setae on mesal margin and one pair of medium-sized, stout apicolateral setae, apex rounded (Figure 3B–C). Inferior appendage (if. ap.), in lateral view, setose, base broad, divided into three processes of unequal length (Figure 3A); dorsal process (ds. pc.), in lateral and dorsal view, hook-like, medium-sized, curved mesad, apex rounded and directed mesad, bearing numerous short, stout dorsal setae (Figure 3A–B); mesal process (ms. pc.), in lateral view, bifid, with digitate lobes of unequal length (Figure 3A); in dorsal view, directed mesad (Figure 3B); ventral process (vt. pc.), in lateral and dorsal view, slender, very long, surpassing tergum X, narrowing apically slightly curved dorsad, apex rounded and directed posteromesad (Figures 3A,C); in ventral view, basomesal margin crenulate, without projections or lobes, covered by numerous long, thin setae (Figure 3D); basal plate (bs. pt.), in lateral view large, trough-like (Figure 3A). Phallic apparatus, in lateral view, tubular, almost straight along its length; phallobase large, shoe-shaped in lateral, ventral margin strongly sclerotized; endothecal membranes striate apicodorsally, apex bifid, with three medium-sized, thin setae; endothecal spines absents; parameres presents, paired, slender, strongly sclerotized, as long as phallic apparatus; apex slightly bifid, covered by numerous short, thin setae, directed posteroventrad in lateral view and posterolaterad in dorsal view; phallotremal sclerite (ph. sc.) well-developed, comma-shaped in lateral view and U-shaped in dorsal view (Figure 3E–F).
Distribution. CHILE (Región del Maule) (Figure 1).
Material examined. HOLOTYPE MALE. CHILE: Región del Maule, San Clemente, Parque Natural Tricahue, Sendero Siempreverde, 35.6701944°S, 71.0544167°W, 730 m a.s.l., ii–viii.2023, R.O. Araújo et al. legs., Malaise trap (MNNC). PARATYPES. Same data as holotype, except 739 m a.s.l., iii–viii.2023, R.O. Araújo et al. legs., Malaise trap, 3 males (MNNC), 2 males (INPA), 2 males (MZUSP), 2 males (MNRJ), 2 males (CIASP).
Etymology. The specific epithet refers to Parque Natural Tricahue, a natural sanctuary located in a transitional zone between the northern and southern forests of Chile. The park harbors ecosystems rich in biodiversity, including numerous species endemic to the Maule region, making it a unique and significant site for nature conservation. The epithet is used in apposition.
3.2. Key to Males of Brachysetodes s. str.


4. Discussion
The discovery of Brachysetodes tricahue sp. nov. highlights the ongoing potential for taxonomic and biogeographic discoveries within Brachysetodes, particularly in the southern Andes, where the genus remains insufficiently sampled. The genus may correspond to a particular biogeographic element described by Moreira-Muñoz [27] for the Chilean flora: the south-temperate element, which comprises taxa confined to regions south of 33° S in Chile and adjacent Argentina, primarily within temperate forest ecosystems (see [28]).
Although Brachysetodes is most diverse in Central Chile, where the highest number of species have been described, the majority of distributional records are concentrated in the Subantarctic region (Figure 1B). The new species, described from the Central subregion of the Chilean Andes, adds to the endemic richness of this biogeographically complex area [15,29]. This pattern may reflect either a true biogeographic signal or a collection bias resulting from uneven sampling efforts across the Andean regions.
Brachysetodes tricahue sp. nov., clearly assignable to Brachysetodes sensu stricto based on the synapomorphies identified in Holzenthal’s [14] phylogenetic analyses, particularly features of forewing venation and the male structures of tergum X and the inferior appendages, further refines our understanding of morphological variation within the genus and provides valuable data for future phylogenetic studies. Morphologically, it is most similar to B. bifurcatus and B. nublensis, especially in the trilobed structure of the inferior appendages and the lateral processes of tergum X. However, consistent and diagnostic differences in the number and robustness of setae on tergum X, the shape and proportions of the inferior appendage processes, and the structure of the phallic apparatus support recognizing B. tricahue sp. nov. as a distinct taxon. Notably, the unequal length of the bifid lobes of the median process and the elongate, crenulate ventral process are autapomorphic features distinguishing it from all known congeners.
These subtle but consistent differences reinforce the diagnostic value of male genitalia for species-level identification within Brachysetodes, a pattern also observed in other Leptocerinae genera, such as Amphoropsyche Holzenthal, 1985 (see [13,30]), Oecetis McLachlan, 1877 (see [31,32]), and Triaenodes McLachlan, 1865 (see [33]). Furthermore, such morphological distinctions may reflect not only species-level divergence but also ecological or behavioral specializations yet to be explored, potentially linked to the diversity of functional traits suspected in caddisflies [1].
The distribution of Brachysetodes species across the Andes reflects a complex biogeographic history. Although the genus occurs throughout the Chilean Andes, its presence in the Argentine Andes is limited to only three species (B. extensus, B. forcipatus, and B. quadrifidus) [14]. Notably, B. forcipatus and B. quadrifidus exhibit the broadest distributions, occurring across multiple subregions in both Argentina and Chile (Figure 1B). Their wider ranges may suggest ecological generalism or greater dispersal capacity compared to more range-restricted species. In contrast, B. tricahue sp. nov. appears to have a narrow distribution, currently known only from Parque Natural Tricahue in the Maule region, part of the Central Andean subregion [15]. This highlights the significance of local endemism and the need for further faunistic surveys in these underexplored montane habitats, employing effective sampling methods such as Malaise traps.
The occurrence of Brachysetodes species in the Subantarctic region, particularly in the Valdivian Forest province (sensu Morrone [15], aligns with findings by Sganga et al. [34], who identified this province as the most species-rich area for caddisflies in Argentina. Interestingly, the Patagonian subregion lacks any confirmed records of Brachysetodes, suggesting either ecological constraints limiting the genus’s range or a significant undersampling of aquatic insect fauna in that region.
From a phylogenetic perspective, Brachysetodes remains problematic. It is considered polyphyletic [3,14], and while its placement within the Leptocerinae is supported by both morphological and molecular data [2,4], the lack of tribal assignment highlights unresolved evolutionary relationships. The description of new species, accompanied by detailed morphological data and an updated key to males of Brachysetodes sensu stricto, provides a crucial foundation for future phylogenetic revisions, ideally integrating both adult and larval stages as well as molecular datasets.
In conclusion, B. tricahue sp. nov. contributes to the known diversity of Brachysetodes and enhances understanding of Trichoptera biogeography in the southern Andes. This study establishes a valuable baseline for future research incorporating larval stages, ecological data, and molecular analyses to reassess the monophyly of the genus and refine our understanding of leptocerid evolution and diversification in South America.
Author Contributions
Conceptualization, GRD; methodology, GRD, VS, DGP and ROA; software, GRD and VS; validation, GRD and NH; formal analysis, GRD and VS; investigation, all authors; resources, DGP, AMM and ROA; data curation, GRD; writing—original draft preparation, GRD; writing—review and editing, GRD, VS, DGP, ROA, AMM, NH, and PCB; visualization, GRD; supervision, GRD; project administration, GRD; funding acquisition, AMM, ROA and PCB. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Agencia Nacional de Investigación y Desarrollo (ANID), Chile, through the FONDECYT Regular Project No. 1221879 (ROA and AMM). Additional support was provided by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES), Finance Code 001; the INCT ADAPTA II program funded by the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), grant number 465540/2014-7; and the Fundação de Amparo à Pesquisa do Estado do Amazonas (FAPEAM) – POSGRAD 2019. GRD was supported by a postdoctoral fellowship granted by FAPEAM and CNPq through the “Programa de Apoio à Fixação de Jovens Doutores no Brasil” (grant PROFIX-JD 150625/2023-5). CNPq also awarded research fellowships to NH (grant 308970/2019-5), VS (grant 01.02.016301.04091/2022-00), and PCB (grant 306400/2022-7). GRD and PCB express their gratitude to the São Paulo Research Foundation (FAPESP) for current support through a postdoctoral fellowship (grant 2024/04714-2) and a research grant (grant 2023/03818-6), respectively.
Data Availability Statement
All available data are presented in the present study, and specimens are vouchered as indicated in the examined material section.
Acknowledgments
The authors gratefully acknowledge the directors of Parque Natural Tricahue for their support in granting research permission and facilitating access to conduct the study. GRD is grateful to Dra. Beatriz Ronchi Teles (INPA) for allowing the use of the laboratory infrastructure. Thanks are extended to the Invertebrate Collection of INPA for allowing the use of photographic equipment.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| bs. pt. | basal plate |
| cn. g. | coronal groove |
| dc | discoidal cell |
| ds. pc. | dorsal process |
| fg. sw. | frontogenal wart |
| ft. g. | frontal grooves |
| if. ap. | inferior appendage |
| la. sw. | lateroantennal wart |
| lt. pc. | lateral process |
| ma. sw. | medioantennal wart |
| ms. pc. | mesal process |
| occ. sw. | occipital wart |
| ph. bs. | phallobase |
| ph. sc. | phallotremal sclerite |
| pm. | paramere |
| pt. sw. | postgenal wart |
| tc | thyridial cell |
| vt. pc. | ventral process |
| vt. sw. | ventral setal wart |
References
- Morse, J.C.; Frandsen, P.B.; Graf, W.; Thomas, J.A. Diversity and Ecosystem Services of Trichoptera. Insects 2019, 10, 125. [Google Scholar] [CrossRef] [PubMed]
- Malm, T.; Johanson, K.A. A New Classification of the Long-Horned Caddisflies (Trichoptera: Leptoceridae) Based on Molecular Data. BMC Evol. Biol. 2011, 11. [Google Scholar] [CrossRef] [PubMed]
- Morse, J.C. A Phylogeny and Classification of Family-Group Taxa of Leptoceridae (Trichoptera). In Proceedings of the Third International Symposium on Trichoptera; Springer Netherlands: Dordrecht, 1981; pp. 257–264. [Google Scholar]
- Henriques-Oliveira, A.L.; Silva, A.L.R.; Nessimian, J.L.; Takiya, D.M. New Long-Horned Caddisfly Genus and Species of Leptoceridae (Insecta: Trichoptera) from Brazil. Zootaxa 2021, 5057, 530–544. [Google Scholar] [CrossRef] [PubMed]
- Schmid, F. Contribution à La Connaissance Des Trichopteres Néotropicaux. Mém. Soc. Vaud. Sci. Nat. 1955, 11, 117–160. [Google Scholar]
- Schmid, F. Contribution a l’etude Des Trichopteres Neotropicaux III. Mitt. Zool. Mus. Berl. 1958, 34(1), 183–217. [Google Scholar] [CrossRef]
- Schmid, F. Contribution a l’etude Des Trichopteres Neotropicaux V. Tijdschr. Entomol. 1964, 107, 307–339. [Google Scholar]
- Flint, O.S. Bredin-Archbold-Smithsonian Biological Survey of Dominica, 9. The Trichoptera (Caddisflies) of the Lesser Antilles. Proc. U.S. Natl. Mus. 1968, 125, 1–86. [Google Scholar] [CrossRef]
- Flint, O.S. Studies of Neotropical caddis flies, IX: New Genera and Species from the Chilean Subregion. Proc. Entomol. Soc. Wash. 1969, 71, 497–514. [Google Scholar]
- Flint, O.S. Studies of Neotropical caddisflies, XIV: on a collection from Northern Argentina. Proc. Biol. Soc. Wash. 1972, 85, 223–248. [Google Scholar]
- Flint, O.S. Studies of Neotropical Caddisflies, XXXIII: New Species from Austral South America (Trichoptera). Smithson. Contrib. Zool. 1983, 1–100. [Google Scholar] [CrossRef]
- Holzenthal, R.W. Studies in Neotropical Leptoceridae (Trichoptera) I: Achoropsyche, a new genus. In Proceedings of the 4th International Symposium on Trichoptera, Clemson, USA, 11-16 July 1983. [Google Scholar]
- Holzenthal, R.W. Studies in Neotropical Leptoceridae (Trichoptera) II: Amphoropsyche, a new genus and species of Leptocerinae from northern South America. Int. J. Entomol. 1985, 27, 255–269. [Google Scholar]
- Holzenthal, R.W. Studies in Neotropical Leptoceridae (Trichoptera), IV: A Revision of Brachysetodes Schmid. Trans. Am. Entomol. Soc. 1986, 111, 407–440. [Google Scholar]
- Morrone, J.J. Biogeographical Regionalisation of the Andean Region. Zootaxa 2015, 3936. [Google Scholar] [CrossRef] [PubMed]
- Holzenthal, R.W.; Calor, A.R. Catalog of the Neotropical Trichoptera (Caddisflies). ZooKeys 2017, 654, 1–566. [Google Scholar] [CrossRef] [PubMed]
- Gressitt, J.L.; Gressitt, M.K. An improved Malaise trap. Pac. Insects. 1962, 4, 87–90. [Google Scholar]
- González Lagos, N. Centro de Ecoturismo Parque Tricahue : Oportunidad de Un Nuevo Pacto Entre Arquitectura y Naturaleza. Tesis, Universidad de Chile - Facultad de Arquitectura y Urbanismo: Santiago, Chile, 2021.
- Blahnik, R.J.; Holzenthal, R.W. Collection and Curation of Trichoptera, with an Emphasis on Pinned Material. Nectopsyche, Neotropical Trichoptera Newsletter 2004, 1, 8–20. [Google Scholar]
- Desiderio, G.R.; Santana, V.; Pereira, E.S.; Pes, A.M.; Hamada, N. On the Identity of Smicridea (Smicridea) Aequalis Banks, 1920 (Trichoptera: Hydropsychidae): Morphology of Adults and Immature Stages, Bionomics, Distribution, and Male Color Dimorphism. Neotrop. Entomol. 2021, 50, 430–443. [Google Scholar] [CrossRef] [PubMed]
- Kawada, R.; Buffington, M.L. A Scalable and Modular Dome Illumination System for Scientific Microphotography on a Budget. PLOS ONE 2016, 11, e0153426. [Google Scholar] [CrossRef] [PubMed]
- Romano, G.M. A High Resolution Shapefile of the Andean Biogeographical Region. Data Brief 2017, 13, 230–232. [Google Scholar] [CrossRef] [PubMed]
- GBIF.Org. GBIF Occurrence Download. [CrossRef]
- Oláh, J.; Johanson, K.A. Trinominal Terminology for Cephalic Setose Warts in Trichoptera (Insecta). Braueria (Lunz am See, Austria) 2007, 34, 43–50. [Google Scholar]
- Nielsen, A. A Comparative Study of the Genital Segments and Their Appendages in Male Trichoptera. 1957, 8, 1–159.
- Mosely, M.E.; Kimmins, D.E. The Trichoptera (Caddis-Flies) of Australia and New Zealand; British Museum (Natural History): London, England, 1953. [Google Scholar]
- Moreira-Munoz, A. Plant Geography of Chile; Plant and Vegetation; Springer Netherlands: Dordrecht, 2011; ISBN 978-90-481-8747-8. [Google Scholar]
- Pádua, D.G.; Moreira-Muñoz, A.; Morales-Fierro, V.; Araujo, R.O. Chilean Darwin Wasps (Ichneumonidae): Biogeographic Relationships and Distribution Patterns. Insects 2024, 15, 415. [Google Scholar] [CrossRef] [PubMed]
- Morrone, J.J. Evolutionary Biogeography of the Andean Region; 1st ed.; CRC Press: Boca Raton: Taylor & Francis, 2018. | Series: CRC biogeography series, 2018; ISBN 978-0-429-48608-1.
- Rázuri-Gonzales, E.; Holzenthal, R.W.; Ríos-Touma, B. Two New Species of the Rare Neotropical Caddisfly Genus Amphoropsyche Holzenthal (Trichoptera, Leptoceridae). ZooKeys 2017, 707, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Blahnik, R.; Holzenthal, R. Review and Redescription of Species in the Oecetis Avara Group, with the Description of 15 New Species (Trichoptera, Leptoceridae). ZooKeys 2014, 376, 1–83. [Google Scholar] [CrossRef] [PubMed]
- Quinteiro, F.B.; Holzenthal, R.W. Fourteen New Species of Oecetis McLachlan, 1877 (Trichoptera: Leptoceridae) from the Neotropical Region. PeerJ 2017, 5, e3753. [Google Scholar] [CrossRef] [PubMed]
- Holzenthal, R.W.; Andersen, T. The Caddisfly Genus Triaenodes in the Neotropics (Trichoptera: Leptoceridae). Zootaxa 2004, 511. [Google Scholar] [CrossRef]
- Sganga, J.V.; Brand, C.; Santos, A.P.M.; Rueda Martín, P.A. Trichoptera. In Biodiversidad de Artrópodos Argentinos; Claps, L.E., Roig-Juñent, S., Morrone, J.J., Eds.; Editorial INSUE UNT: San Miguel de Tucumán, Argentina, 2023; Volume 5, pp. 213–228. [Google Scholar]
Figure 1.
Distribution of Brachysetodes species in the Andean region and general views of the collection site of the new species described here: (A) Map of the Andean region sensu Morrone (2015) showing its sub-regions and South American transition zone (highlighted in colors); (B) Geographic distribution map for all Brachysetodes species; (C) Image satellite from Google Earth® showing the type locality of Brachysetodes tricahue Desidério, Santana & Hamada, sp. nov.; (D) Parque Natural Tricahue, Región del Maule, Chile, site where Brachysetodes tricahue Desidério, Santana & Hamada, sp. nov. was collected.
Figure 1.
Distribution of Brachysetodes species in the Andean region and general views of the collection site of the new species described here: (A) Map of the Andean region sensu Morrone (2015) showing its sub-regions and South American transition zone (highlighted in colors); (B) Geographic distribution map for all Brachysetodes species; (C) Image satellite from Google Earth® showing the type locality of Brachysetodes tricahue Desidério, Santana & Hamada, sp. nov.; (D) Parque Natural Tricahue, Región del Maule, Chile, site where Brachysetodes tricahue Desidério, Santana & Hamada, sp. nov. was collected.

Figure 2.
Brachysetodes tricahue Desidério, Santana & Hamada, sp. nov., holotype male: (A) Lateral habitus; (B) Head, frontal view; (C) Head, dorsal view; (D) Head and thorax, dorsal view; (E) Right forewing, dorsal view (longitudinal veins highlighted in colors); (F) Right hindwing, dorsal view (longitudinal veins highlighted in colors). Scale bars in mm: A, B, D, 0.2; C, 0.05; E, F, 0.1.
Figure 2.
Brachysetodes tricahue Desidério, Santana & Hamada, sp. nov., holotype male: (A) Lateral habitus; (B) Head, frontal view; (C) Head, dorsal view; (D) Head and thorax, dorsal view; (E) Right forewing, dorsal view (longitudinal veins highlighted in colors); (F) Right hindwing, dorsal view (longitudinal veins highlighted in colors). Scale bars in mm: A, B, D, 0.2; C, 0.05; E, F, 0.1.

Figure 3.
Brachysetodes tricahue Desidério, Santana & Hamada, sp. nov., male genitalia (holotype): (A) Genitalia, left lateral view; (B) Genitalia, dorsal view; (C) Apex of lateral process of the tergum X, dorsal view (photograph); (D) Genitalia, ventral view; (E) Phallic apparatus, left lateral view; (F) Phallic apparatus, dorsal view. Scale bars in mm: A, B, D, 0.2; C, 0.05; E, F, 0.1.
Figure 3.
Brachysetodes tricahue Desidério, Santana & Hamada, sp. nov., male genitalia (holotype): (A) Genitalia, left lateral view; (B) Genitalia, dorsal view; (C) Apex of lateral process of the tergum X, dorsal view (photograph); (D) Genitalia, ventral view; (E) Phallic apparatus, left lateral view; (F) Phallic apparatus, dorsal view. Scale bars in mm: A, B, D, 0.2; C, 0.05; E, F, 0.1.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



