
Submitted:
18 September 2024
Posted:
18 September 2024
You are already at the latest version
Abstract
Sentinel eggs used to monitor field parasitism of stink bug pests (Hemiptera: Pentatomidae) can only be deployed for a few days to avoid releasing the pest in the monitored area. Using sterile eggs removes the risk of accidental pest introduction and extends deployment time. Freezing the eggs before deployment is one common method for sterilizing sentinel eggs. However, some egg parasitoid species have lower to no parasitism on frozen eggs. In this study, x-ray irradiation was used to sterilize Bagrada hilaris sentinel eggs intended for monitoring parasitism by Gryon aetherium (Hymenoptera: Scelionidae), the most promising biological control candidate. In this case, freezing sentinel eggs is not recommended because G. aetherium has low levels of parasitism on frozen eggs. Doses as low as 10 Gy induced 100% sterility. Irradiated eggs successfully sustained the development of G. aetherium and Ooencyrtus californicus (Hymenoptera: Encyrtidae), another egg parasitoid attacking B. hilaris, and parasitism levels were comparable to fresh eggs up to seven days. In addition, G. aetherium showed no preference for fresh non-irradiated over seven-day-old irradiated eggs. Our results indicate that x-ray irradiation is a suitable alternative to produce safe and reliable sentinel eggs to monitor egg parasitism of B. hilaris and possibly other species.
Keywords:
stink bug
; egg parasitism
; field monitoring
; sentinel eggs
; x-ray irradiation
; egg sterilization
1. Introduction
Implementation of biological control programs requires the ability to monitor the presence and impact of biocontrol agents [1,2]. Biocontrol of stink bug pests (Hemiptera: Pentatomidae) often relies on egg parasitoids [3,4,5,6]. Monitoring egg parasitism in the field is done using sentinel eggs, either naturally occurring or lab-reared eggs. Although wild sentinel eggs can lead to better estimates of parasitism levels and species richness of the indigenous parasitoid community [7,8], lab-reared eggs are more commonly used because searching for egg masses in the field is time-consuming and not always practical. Lab-reared sentinel eggs are deployed in the field for a limited period, long enough to maximize the opportunity to be discovered by naturally occurring/resident parasitoids, but not long enough that the parasitoids complete their development and emerge before retrieval of the eggs. Although more practical, lab-reared sentinel eggs come with the risk of releasing the pest in the monitored area when the unparasitized eggs hatch, thus limiting their deployment to 2 to 6 days [9,10,11,12]. Being able to extend the duration of deployment could significantly improve the detection of parasitoids and even hyperparasitoids for stink bug pests [13].
Frozen eggs are commonly used for sentinel eggs [10,14,15]. Freezing the eggs kills the developing embryo while still providing a suitable food source for parasitoid development [16,17]. However, some parasitoid species have higher parasitism rates on frozen eggs compared to living eggs, a bias caused by the fact that freezing the eggs suppresses the host's immune response to parasitism [18]. In contrast, other species have lower to no parasitism on frozen eggs, due to rejection by the ovipositing female [19], or, in some cases, lack of viability and development of the host needed for successful parasitism [20]. Furthermore, frozen eggs tend to start to rot and desiccate a few days after thawing and are suitable for parasitoid oviposition for only a few days [21].
Ionizing radiation is the most commonly used method to induce sterility in insects, a technique at the core of the Sterile Insect Technique (SIT). This pest management practice consists of mass rearing, sterilizing, and releasing a pest species to compete with the wild population [22]. Ionizing radiation can also be used to sterilize eggs directly without altering their suitability for egg parasitoid development [23]. Gamma-based irradiators have been traditionally used for insect sterilization but are becoming more difficult to procure and maintain because of economic, regulatory and safety concerns [24]. Recent studies show that x-rays are as effective as gamma rays to induce sterility in insects [25,26]. There is increasing pressure to phase out gamma irradiators in favor of x-ray. The 2019 Nuclear Defense Authorization Act (Section3141) calls for replacing all cesium-137 blood irradiators with x-ray by 2027 [27].
This study was developed to improve the sentinel eggs deployed to monitor egg parasitism of Bagrada hilaris (Burmeister) (Hemiptera: Pentatomidae), a worldwide pest of cole crops (broccoli, kale, cabbage cauliflower and all cultivated varieties of Brassica oleracea L.) found in the USA, Mexico, Chile and Europe [28,29,30,31,32]. Nymphs and adults feed on various cultivated and wild Brassicaceous species [33]. Conventional control strategies are usually ineffective because the natural habitats surrounding crops often act as a refuge for the pest. In addition, using conventional insecticide comes with economic and ecological costs, beyond the risk of insects developing resistance [34]. Therefore, biological control is considered a more sustainable control option for the bagrada bug, creating a demand for safe and reliable sentinel eggs to monitor the presence and impact of possible biocontrol agents both in the region of origin of the pest or in the invaded areas. Collecting naturally parasitized B. hilaris eggs is challenging because females lay most of their eggs under the soil surface [35], a unique strategy for a pentatomid species, most likely developed to escape egg parasitism. However, one egg parasitoid, Gryon aetherium Talamas (Hymenoptera: Scelionidae) has developed the ability to search for and parasitize eggs under the soil surface and is currently the most promising biological control candidate for this pest [36]. Although originally retrieved in Pakistan, adventive populations of this parasitoid are now recovered in California [37]. Unfortunately, using frozen eggs to monitor B. hilaris field parasitism is not recommended because G. aetherium has low levels of parasitism on frozen eggs [19,38].
In this study, we investigated the feasibility of using x-ray irradiation to sterilize B. hilaris sentinel eggs. We first looked at the doses needed to induce 100% sterility and if radiosensitivity decreased with egg age as observed in other insect species [23,39]. Next, we checked the suitability of irradiated eggs for the oviposition and development of two parasitoids: the above-described G. aetherium and Ooencyrtus californicus Girault (Hymenoptera: Encyrtidae), another egg parasitoid attacking B. hilaris found in California [37]. Finally, G. aetherium preference for irradiated eggs vs. fresh or same-age untreated eggs was investigated using choice tests. While this study was developed to enhance the field monitoring of B. hilaris parasitoids, the results could apply to other species for which frozen sentinel eggs are not a good option.
2. Materials and Methods
2.1. Insect colonies
A laboratory colony of B. hilaris was established from field-collected individuals (Solano and Monterey counties, CA in 2015 and 2018) and maintained in bugdorm cages (61 × 61 × 61 cm, BioQuip Products Inc, Rancho Dominguez, California) at 29.0 ± 2.0 °C, 41 ± 5 % RH and 16L:8D photoperiod. The colony was fed with store-bought organic broccoli and kale. Sand-filled (Quikrete Brown Play Sand, American Canyon, CA) petri dishes (diameter: 47 mm, height: 5 mm) sheltered below a laser-cut Plexiglas shade structure (14 x 7 x 2.5 cm) were provided as oviposition sites. Eggs were produced in separate oviposition boxes consisting of custom laser-cut ventilated Plexiglas boxes (28 x 16.5 x 14 cm), either oviposited in sand-filled petri dishes (sand-covered) or on strips of cheesecloth (sand-free). Eggs were collected by sieving the sand using a No. 35 sieve (Humboldt Manufacturing Co., Elgin, IL; mesh size: 500 μm) or by manually detaching the eggs from the cheesecloth using featherweight forceps.
Egg parasitoid colonies were established with adults emerging from field-parasitized B. hilaris eggs collected near Davis, California, in 2020. Adults were kept in glass vials (diameter: 25 mm, height: 95 mm) at 20-24 °C, 40-60 % RH, and 16L:8D photoperiod and fed organic raw honey spread on the inside of the stopper. About 2-4 times per week, the adults were exposed to one egg card consisting of 30-50 fresh (< 24 h old) B. hilaris eggs glued (Elmer’s School Glue, Columbus, OH) to a rectangular piece of cardstock (20 x 60 mm). These egg cards were removed 1-3 days later and incubated in the same conditions until adults emerged 21 to 30 days later. Experimental parasitoid females were assumed mated because they emerged in the presence of males and were housed with males within one day of their emergence until used for experiments.
2.2. Irradiation Process
The eggs were irradiated in a custom x-ray irradiator adapted from previously described versions [40,41] consisting of a shielded cabinet enclosing a pair of 1000 Watt fan beam x-ray tubes (MXR-100HP/20 FB, Comet Technologies USA Inc., Stamford, CT, USA) mounted side by side so that the emitted fan-shaped x-ray beams fell in a straight line on the surface of an adjacent rotating drum, parallel to the axis of rotation (Figure 1). X-ray tubes were energized with 1000-Watt (100 kV, 10 mA) power supplies (XPgN100, Matsusada Precision Inc., Kusatsu Shiga, Japan) and cooled with a portable chiller (M1-1.5A, Advantage, Greenwood, IN). The tubes were strategically spaced so that x-ray dose along the line was uniform, and thus, given many drum rotations, the dose at all points on the surface approached uniformity. The eggs were enclosed in 1-inch Ziplock bags and attached to the rotating drum with Velcro strips. The absorbed dose was measured with a radiation measurement system (Accu-Dose MNL/2086, Radcal Corporation, Monrovia, CA, USA) using a high dose rate ion chamber (10X6-0.18, Radcal Corporation) attached to the surface of the drum, allowing real-time dose determination. The drum was rotated at a rate of 25 revolutions per minute. The dose rate applied varied depending on the target dose but was generally between 5 – 6 Gy/min. Irradiator settings in terms of x-ray energy (keV) and current (ma) are given for each experiment.
2.3. Effect of X-Ray Irradiation on Egg Eclosion
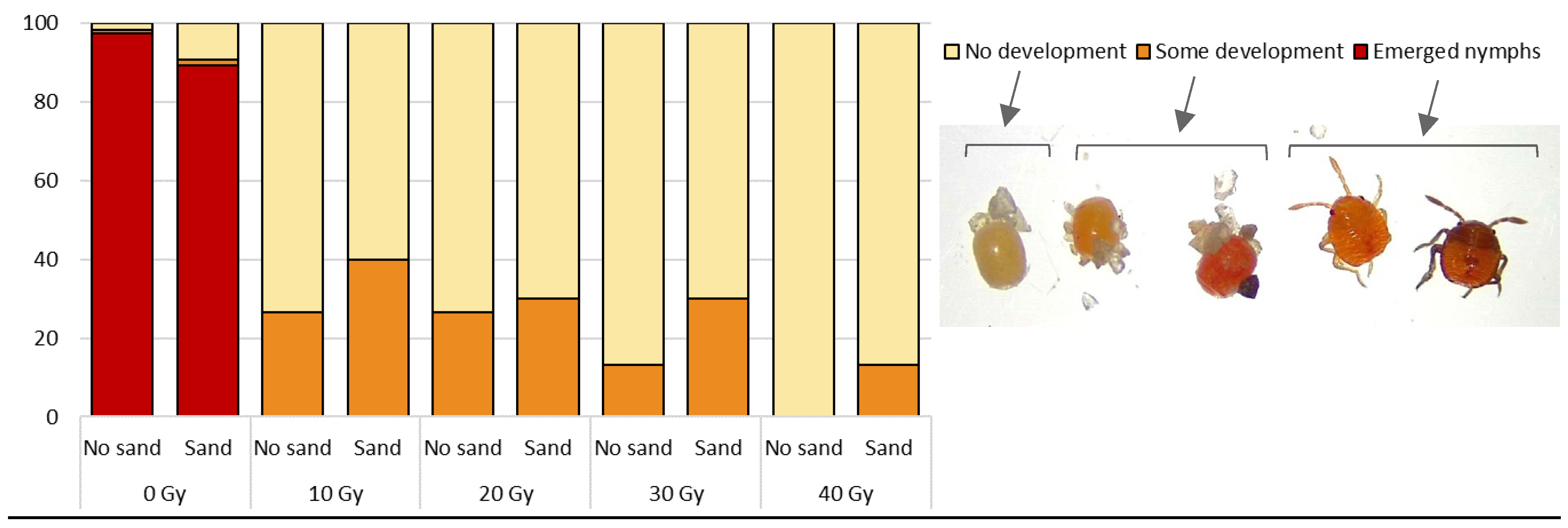
Fresh (< 24 h old) B. hilaris eggs were irradiated at 0 (control), 10, 20, 30 and 40 Gy (81 kV; 6mA; dose rate: 5.0 Gy/min). There were 30 eggs per dose. Both sieved and sand-free eggs were tested to determine if sand particles adhering to the egg surface serve as shielding against irradiation. After the irradiation, the eggs were transferred into glass vials and incubated at 23.5 ± 0.3 °C and 69 ± 9 % RH. Freshly oviposited B. hilaris eggs are whitish/cream-colored, then slowly turn orange as they develop and end up bright orange before eclosion. Freshly eclosed nymphs are bright orange (Figure 2). At 23 °C, nymphs start to emerge on day 7. In this experiment, the eggs were visually checked 12 days-post irradiation and categorized as either (1) eclosed, (2) dead and uneclosed but showing some development (orange tint/color) and (3) dead and uneclosed with no embryo development (cream color).
2.4. Effect of Egg Age on Radiosensitivity
In a similar experiment, we irradiated 1 d (<24h), 2 d (24-48h), and 3 d (48h-72h) old B. hilaris eggs, as well as eggs refrigerated at 8 °C for 7 days, with a dose of 40 Gy (81 kV; 6mA; dose rate: 5.0 Gy/min). Non-irradiated controls were also included for each treatment. After the irradiation, the eggs were transferred into glass vials and incubated in the same condition as above. The eggs were visually checked 9 days post-irradiation and categorized as either (1) eclosed, (2) dead and uneclosed but showing some development (orange color/tint) and (3) dead and uneclosed with no embryo development (cream color).
2.5. Suitability of Irradiated Eggs for Parasitoid Development
Parasitism by G. aetherium was tested in no-choice tests on fresh (< 24 h old), 7 and 12-day-old irradiated eggs (40 Gy; 81 kV; 7mA; dose rate: 5.8 Gy/min). Controls were fresh non-irradiated eggs. An additional control with 7-day old eggs was also included (no control was available for 12-day-old eggs because B. hilaris egg eclosion usually starts 7-8 days post oviposition at 23 °C). Ten egg cards with 10 sieved eggs each were prepared for each treatment and exposed to one single 1 to 5-day-old G. aetherium wasp for 24h. The numbers of emerged B. hilaris nymphs, emerged parasitoids and their sex, and dead unemerged eggs were recorded after 30 days.
Parasitism by O. californicus was tested similarly with 4- to 6-day-old females using a no-choice test with 7-day-old irradiated eggs and a control consisting of fresh (<24 h old) non-irradiated eggs. There were 16 replicates. Because parasitism rates were lower with this species, exposure was extended from 24 to 48h for the last 6 replicates.
2.6. Parasitoid preference
Gryon aetherium preference for fresh non-irradiated vs. 7-day-old, irradiated eggs was tested in choice tests, where single 2- to 4-day-old parasitoid females were presented with five fresh eggs and five 7-day-old irradiated eggs. The two types of eggs were glued in two rows of 5. After a 24 h exposure the egg cards were cut longitudinally and halves, each containing 5 eggs of one type, were incubated in separate vials. There were 10 replicates. The numbers of emerged B. hilaris nymphs, emerged parasitoids and their sex, and dead unemerged eggs were recorded after 30 days.
2.7. Data Analysis
Parasitism rates were calculated as the sums of emerged parasitoids divided by the number of eggs. We assumed that no more than one adult emerged from each host egg for both species. Indeed, G. aetherium appears to avoid conspecific superparasitism [42]. And, while some Ooencyrtus spp. can sometimes be facultatively gregarious [43], when this happens, we should expect the size of the co-emerged adults to be considerably reduced. Since no obvious difference in size has been observed among O. californicus adults in our experiments, we rejected the possibility of gregarious development. Sex ratio was expressed as the proportion of females and was calculated as the number of female parasitoids divided by the total number of parasitoids. Proportion parasitism and proportion females were compared between treatments using generalized linear models (GLM) with binomial errors, using the glm function in R version 4.2.3 [44]. For the experiment with O. californicus and the choice test with G. aetherium, treatments were compared using mixed model GLMs (GLMM), including either trial date (for the experiment with O. californicus, to account for differences in exposure time between trial dates) or replicate (for the choice test with G. aetherium, to account for non-independence of data from replicates) as a random factor, using the glmer function in R. Turkey HSD tests were used for multiple comparisons.
3. Results
3.1. Effect of X-Ray Irradiation on Egg Eclosion
X-ray doses as low as 10 Gy induced 100% sterility (no eclosion) (Figure 2). A proportion of irradiated eggs turned a light orange tint suggesting that some embryo development may have occurred. A higher proportion of these eggs showing signs of development were recorded at lower doses, and/or in sand-free eggs. Overall, the irradiated eggs started deteriorating 8 days post-irradiation, the chorion progressively separating from the shell and slowly shrinking over time. By day 14, approximately 75% of the eggs appeared desiccated and unlikely to sustain parasitoid development. Twelve days post-irradiation, most control eggs (0 Gy) had eclosed: 98% and 89% of sand-free and sieved eggs, respectively. The remaining uneclosed control eggs did not develop or contained dead unemerged nymphs (natural mortality).
Figure 2.
Percentage of sand-free and sand-covered B. hilaris eggs showing no or some embryo development or eclosed 12 days post-irradiation.
Figure 2.
Percentage of sand-free and sand-covered B. hilaris eggs showing no or some embryo development or eclosed 12 days post-irradiation.
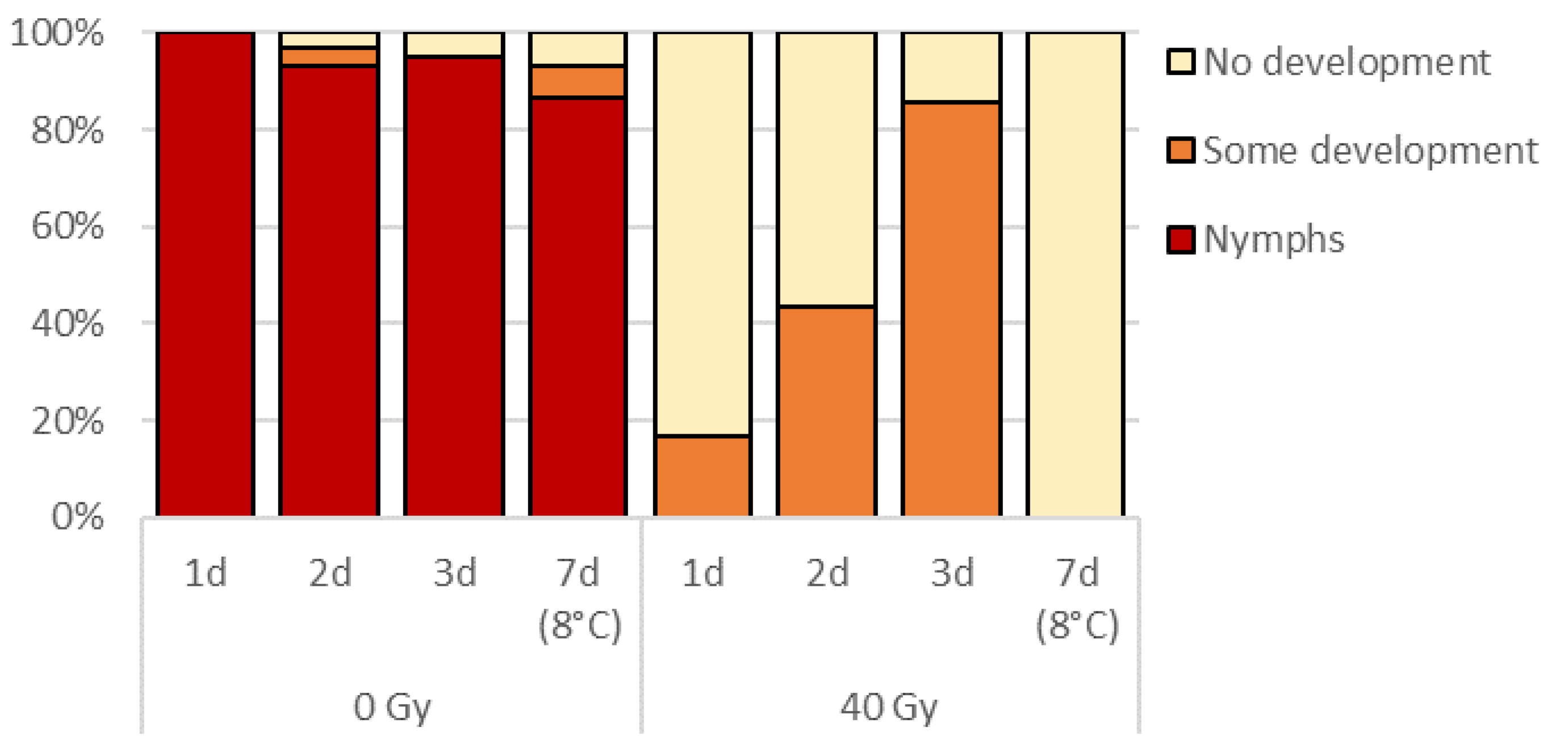
3.2. Effect of Egg Age on Radiosensitivity
Egg hatch was prevented in all irradiated 1, 2, and 3 days old treatments (Figure 3). The proportion of eggs showing some embryo development increased with egg age while no development was observed in 7-day-old irradiated refrigerated eggs. Over 85% nymphal development occurred in all non-irradiated control treatments.
3.3. Suitability of Irradiated Eggs for Parasitoid Development
There was no difference in G. aetherium parasitism rate (GLM, χ2 = 0.58; df = 1; P = 0.45) nor sex ratio (χ2 = 0.02; df = 1; P = 0.90) between fresh irradiated and non-irradiated eggs (Figure 4). Parasitism rate differed between fresh non-irradiated, 7-day-old non-irradiated eggs and 7-day-old irradiated eggs (χ2 = 195.75; df = 2, P < 0.001), and was significantly lower for 7-day-old non-irradiated compared to fresh non-irradiated or 7-day-old irradiated eggs (Tukey,s HSD, P < 0.05). There was also significant difference in sex ratio (χ2 = 7.71; df = 2; P = 0.02), although there were no significant differences between treatments in pairwise comparisons (Tukey’s HSD, P > 0.05). A proportion of the 7-day-old non-irradiated eggs had already hatched on the day of the exposure (7%) and more the next day (41%). The uneclosed eggs most likely contained fully developed nymphs ready to eclose. Surprisingly, about 15% of these eggs still provided a suitable resource for parasitoid development. Parasitism on 12-day-old irradiated eggs was lower compared to fresh eggs ( χ2 = 13.27; df = 1; P < 0.001), but there was no difference in sex ratio (χ2 < 0.01; df = 1; P = 0.95). Parasitism by O. californicus females was greater on irradiated eggs (GLMM, χ2 = 33.21; df = 1; P < 0.001)
3.4. Parasitoid Preference
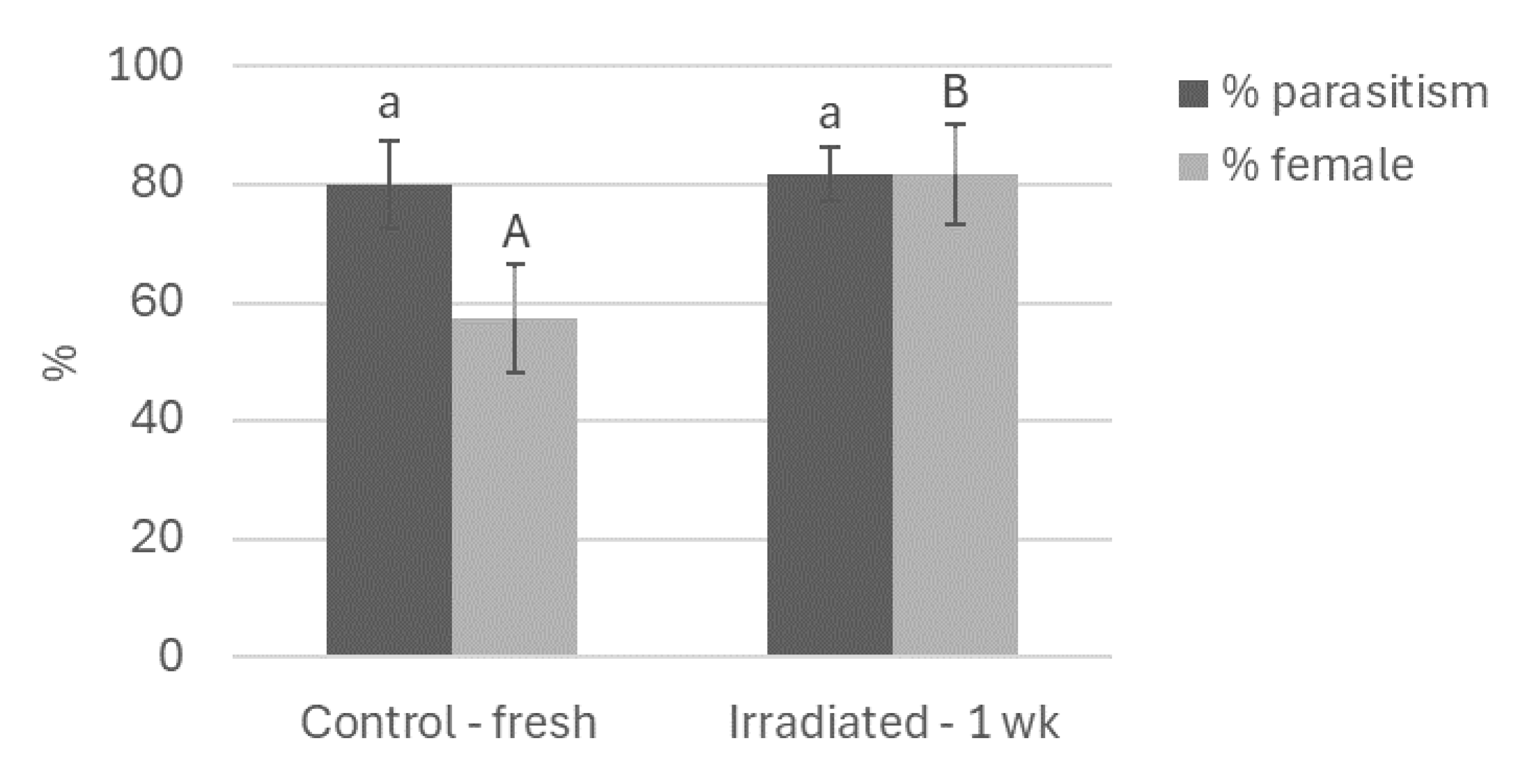
When G. aetherium females were given a choice between fresh untreated eggs and 7-day-old irradiated eggs, they did not show any preference: there was no difference in parasitism rates (GLMM, χ2 = 0.06; df = 1; P = 0.81). There was, however, a difference in sex ratio: more females were produced on irradiated eggs (χ2 = 4.48; df = 1; P = 0.03) (Figure 6).
Figure 4.
Percent parasitism and sex ratio (percent female) by G. aetherium on (a) fresh (< 24h old), (b) 7- and (c) 12-day-old irradiated (40 Gy) B. hilaris eggs. Controls were fresh B. hilaris eggs, with the addition of one control consisting of 7-day-old non-irradiated eggs when testing 7-day-old irradiated eggs. Bars with different letters are significantly (Anova or Tukey’s HSD, P < 0.05).
Figure 4.
Percent parasitism and sex ratio (percent female) by G. aetherium on (a) fresh (< 24h old), (b) 7- and (c) 12-day-old irradiated (40 Gy) B. hilaris eggs. Controls were fresh B. hilaris eggs, with the addition of one control consisting of 7-day-old non-irradiated eggs when testing 7-day-old irradiated eggs. Bars with different letters are significantly (Anova or Tukey’s HSD, P < 0.05).
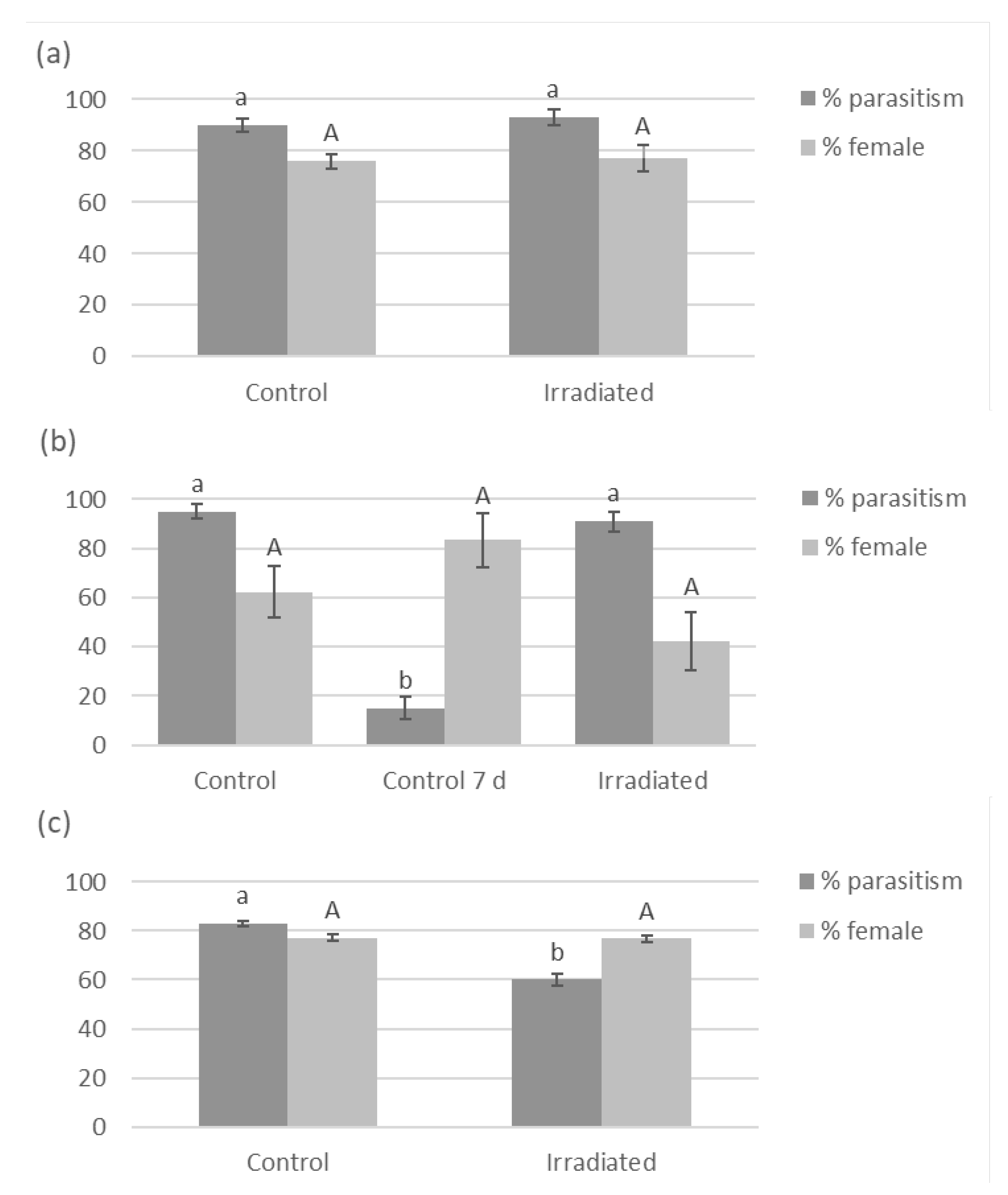
Figure 5.
Percent parasitism and sex ratio (percent female) by O. californicus on 7-day-old irradiated (40 Gy) B. hilaris eggs. Controls were fresh B. hilaris eggs. Bars with different letters are significantly (ANOVA, P < 0.05)
Figure 5.
Percent parasitism and sex ratio (percent female) by O. californicus on 7-day-old irradiated (40 Gy) B. hilaris eggs. Controls were fresh B. hilaris eggs. Bars with different letters are significantly (ANOVA, P < 0.05)
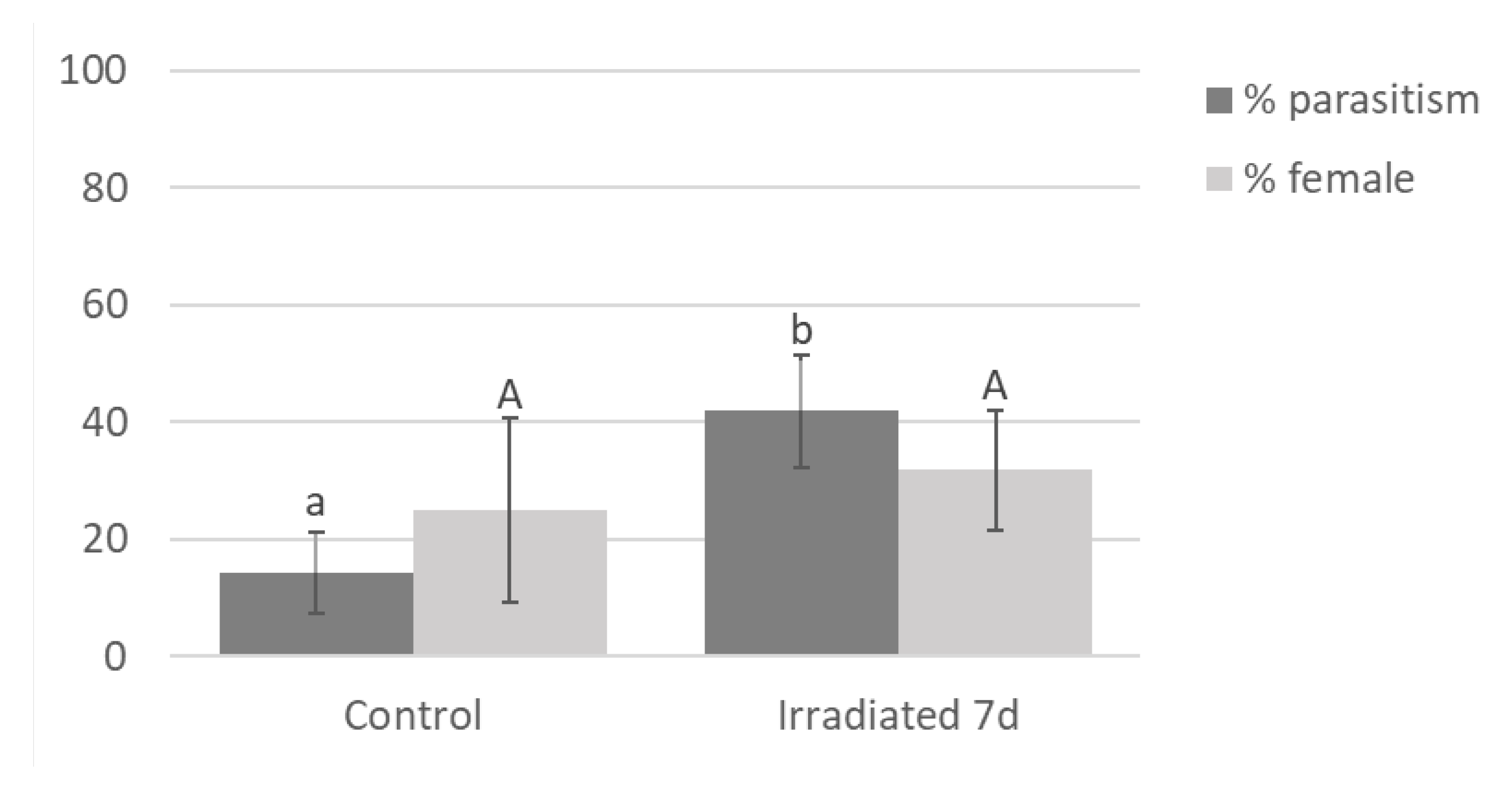
Figure 6.
Percent parasitism and female production when G. aetherium is given a choice between 7-day-old irradiated eggs (40 Gy) and fresh (<24 h old) B. hilaris eggs. Columns with similar letters do not differ significantly (paired t-test, P = 0.05).
Figure 6.
Percent parasitism and female production when G. aetherium is given a choice between 7-day-old irradiated eggs (40 Gy) and fresh (<24 h old) B. hilaris eggs. Columns with similar letters do not differ significantly (paired t-test, P = 0.05).
4. Discussion
Our results showed that x-ray doses as low as 10 Gy prevent B. hilaris eggs from hatching. When no embryo development is preferred, 40 Gy and higher doses are preferable. Our results also demonstrated that the sand particles adhering to the egg surface provide some protection from the radiation, although not enough to compromise the efficiency of the sterilization at the doses we tested. This is promising since sieved eggs are less time-consuming to collect. In a similar study undertaken with the brown marmorated stink bug (BMSB) Halyomorpha halys (Stål) (Hemiptera: Pentatomidae) using gamma rays, doses as low as 16 Gy induced 100% egg sterility [23]. More data on the radiosensitivity of insect eggs is available for Lepidopteran pests. These studies usually also use gamma radiation and show that much higher doses are needed to induce 100% sterility. For instance, irradiation doses of 300 to 350 Gy were required to inhibit the development of eggs of Ephestia cautella (Walker) [45], while eggs of Plodia interpunctella (Hübner) (Lepidoptera: Pyralidae) require 450 Gy [46]. This apparent inconsistency in the sterilization dose required for eggs of different insect species has many possible explanations, including non-equivalency in biological effects of x-rays and gamma rays, different dosimetry methods, and/or differences in the density and structure of the chorion of lepidopteran vs. hemipteran eggs [47]. For this study, all sterilization doses reported refer to the dose as measured with the reader and probe described under the x-ray irradiation parameters given for each experiment. Different dose rates, irradiation methods, and dosimetry tools could lead to different results in terms of the sterilization doses required and subsequent fitness of the irradiated eggs.
The data did not indicate a correlation between radiosensitivity with egg age. Some embryo development was observed in older eggs, but this is most likely due to the normal embryogenesis taking place before the irradiation. Indeed, no such development was observed in eggs kept at 8 °C for 7 days because the cooler temperature slowed down or prevented embryogenesis. In contrast, 2-day-old BMSB eggs were more radioresistant than fresh eggs requiring doses of 40 Gy and above to induce 100% sterility [23]. A similar decrease in egg radiosensitivity with age was also observed with lepidopteran eggs, as demonstrated for the Mediterranean flour moth, Ephestia kuehniella (Lepidoptera: Pyralidae): doses of 75, 125 and 350 Gy were needed to induce 100% sterility of 1, 2 and 3-day-old eggs, respectively [39]. Developmental changes may have provided partial protection from irradiation for BMSB and lepidopteran eggs, but not for bagrada bug eggs.
X-ray irradiated eggs were suitable for G. aetherium and O. californicus oviposition and development. Irradiated eggs performed similarly to fresh non-irradiated eggs for G. aetherium parasitism for at least 7 days, while O. californicus performed better on irradiated eggs. Irradiated BMSB eggs were suitable for the development of Trissolcus japonicus (Ashmead) (Hymenoptera: Scelionidae) the most promising candidate for classical biological control of BMSB, with parasitism rates leveling at 84-91% during the first 7 days before declining at day 10 (no control with fresh eggs was provided) [23]. This suitability of irradiated eggs to parasitoid development is not always observed. For instance, G. gnidus would not parasitize gamma irradiated eggs (10 Gy) of its host Acanthomia tomentosicollis (Stål) (Hemiptera: Coreidae) [20].
In our study, we used sex ratio as a proxy for host quality. Indeed, hymenopteran parasitoids with haplodiploid reproductive systems tend to allocate male eggs to hosts that they assess to be of lower quality [48]. In most of our experiments, the sex ratio was not significantly different in irradiated eggs vs. fresh eggs. This suggests that the host quality of irradiated eggs, or at least of the irradiated eggs accepted for oviposition, remained comparable to fresh eggs. The same observation was made with T. japonicus when parasitizing irradiated eggs of BMSB: the sex ratio hovered around 82 – 84% up to day 7, before starting to decline at day 10 [23].
5. Conclusions
Our study showed the efficiency of x-rays in producing safe and reliable sentinel eggs for B. hilaris. These irradiated eggs could be deployed for at least 7 days with an efficiency comparable to fresh eggs. After 7 days, they would still capture parasitism with no risk of accidental release. Future research will focus on field testing these irradiated sentinel eggs and comparing their efficiency in capturing resident parasitoid species with that of non-irradiated eggs.
Author Contributions
Conceptualization, E.H.; methodology, E.H. and R.P.H.; formal analysis, B.N.H.; investigation, E.H. and R.P.H.; resources, B.N.H.; writing—original draft preparation, E.H.; writing—review and editing, E.H., B.N.H. and R.P.H.; visualization, E.H. and R.P.H; supervision, E.H.; project administration, B.N.H.; funding acquisition, B.N.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by grants from the US Farm Bill and the California Specialty Crop Block Grant Program, grant number 18-0001-064-SC.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We thank Somanette Rivas and Abbie Yee for their help with insect rearing and experiments
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Heimpel, G.E.; Mills, N.J. Biological control; Cambridge University Press: 2017.
- van Lenteren, J.C. Implementation of biological control. Am. J. Altern. Agric. 1988, 3, 102-109.
- Lee, D.-H.; Short, B.D.; Joseph, S.V.; Bergh, J.C.; Leskey, T.C. Review of the biology, ecology, and management of Halyomorpha halys (Hemiptera: Pentatomidae) in China, Japan, and the Republic of Korea. Env. Entomol. 2013, 42, 627-641.
- Mahmood, R.; Jones, W.A.; Bajwa, B.E.; Rashid, K. Egg parasitoids from Pakistan as possible classical biological control agents of the invasive pest Bagrada hilaris (Heteroptera: Pentatomidae). J. Entomol. Sci. 2015, 50, 147-149. [CrossRef]
- Conti, E.; Avila, G.; Barratt, B.; Cingolani, F.; Colazza, S.; Guarino, S.; Hoelmer, K.; Laumann, R.A.; Maistrello, L.; Martel, G. Biological control of invasive stink bugs: Review of global state and future prospects. Entomol. Exp. Appl. 2021, 169, 28-51. [CrossRef]
- Ademokoya, B.; Athey, K.; Ruberson, J. Natural enemies and biological control of stink bugs (Hemiptera: Heteroptera) in North America. Insects 2022, 13, 932. [CrossRef]
- Jones, A.L.; Jennings, D.E.; Hooks, C.R.; Shrewsbury, P.M. Sentinel eggs underestimate rates of parasitism of the exotic brown marmorated stink bug, Halyomorpha halys. Biol. Control 2014, 78, 61-66. [CrossRef]
- Cornelius, M.L.; Dieckhoff, C.; Vinyard, B.T.; Hoelmer, K.A. Parasitism and predation on sentinel egg masses of the brown marmorated stink bug (Hemiptera: Pentatomidae) in three vegetable crops: importance of dissections for evaluating the impact of native parasitoids on an exotic pest. Env, Entomol. 2016, nvw134. [CrossRef]
- Costi, E.; Haye, T.; Maistrello, L. Surveying native egg parasitoids and predators of the invasive Halyomorpha halys in Northern Italy. J. Appl. Entomol. 2019, 143, 299-307. [CrossRef]
- Tillman, G.; Toews, M.; Blaauw, B.; Sial, A.; Cottrell, T.; Talamas, E.; Buntin, D.; Joseph, S.; Balusu, R.; Fadamiro, H. Parasitism and predation of sentinel eggs of the invasive brown marmorated stink bug, Halyomorpha halys (Stål)(Hemiptera: Pentatomidae), in the southeastern US. Biol. Control 2020, 145, 104247. [CrossRef]
- Ganjisaffar, F.; Talamas, E.J.; Bon, M.C.; Perring, T.M. First report and integrated analysis of two native Trissolcus species utilizing Bagrada hilaris eggs in California. J. Hymenopt. Res. 2020, 80, 49-70. [CrossRef]
- Hogg, B.N.; Grettenberger, I.M.; Borkent, C.J.; Stokes, K.; Zalom, F.G.; Pickett, C.H. Natural biological control of Bagrada hilaris by egg predators and parasitoids in north-central California. 2022, 171, 104942. [CrossRef]
- Tillman, G.; Cottrell, T.; Balusu, R.; Fadamiro, H.; Buntin, D.; Sial, A.; Vinson, E.; Toews, M.; Patel, D.; Grabarczyk, E. Effect of duration of deployment on parasitism and predation of Halyomorpha halys (Stål)(Hemiptera: Pentatomidae) sentinel egg masses in various host plants. Fla. Entomol. 2022, 105, 44-52. [CrossRef]
- Herlihy, M.V.; Talamas, E.J.; Weber, D.C. Attack and success of native and exotic parasitoids on eggs of Halyomorpha halys in three Maryland habitats. PLoS One 2016, 11, e0150275.
- Federico, R.P.; Francesco, B.; Leonardo, M.; Elena, C.; Lara, M.; Giuseppino, S.P. Searching for native egg-parasitoids of the invasive alien species Halyomorpha halys Stål (Heteroptera Pentatomidae) in Southern Europe. J. Zool. 2016, 99, 1-8.
- Power, N.; Ganjisaffar, F.; Xu, K.; Perring, T.M. Effects of parasitoid age, host egg age, and host egg freezing on reproductive success of Ooencyrtus mirus (Hymenoptera: Encyrtidae) on Bagrada hilaris (Hemiptera: Pentatomidae) eggs. Env. Entomol. 2021, 50, 58-68.
- Cantón-Ramos, J.M.; Callejón Ferre, Á.J. Raising Trissolcus basalis for the biological control of Nezara viridula in greenhouses of Almeria (Spain). Afr. J. Agric. Res. 2010, 5, 3207-3212.
- Haye, T.; Fischer, S.; Zhang, J.; Gariepy, T. Can native egg parasitoids adopt the invasive brown marmorated stink bug, Halyomorpha halys (Heteroptera: Pentatomidae), in Europe? J. Pest Sci. 2015, 88, 693-705. [CrossRef]
- Martel, G.; Auge, M.; Talamas, E.; Roche, M.; Smith, L.; Sforza, R.F.H. First laboratory evaluation of Gryon gonikopalense (Hymenoptera: Scelionidae), as potential biological control agent of Bagrada hilaris (Hemiptera: Pentatomidae). Biol. Control 2019, 135, 48-56. [CrossRef]
- Egwuatu, R.; Taylor, T.A. Development of Gryon gnidus (Nixon)(Hymenoptera: Scelionidae) in eggs of Acanthomia tomentosicollis (Stål)(Hemiptera: Coreidae) killed either by gamma irradiation or by freezing. Bull. Entom. res. 1977, 67, 31-33. [CrossRef]
- Morandin, L.A.; Long, R.F.; Kremen, C. Hedgerows enhance beneficial insects on adjacent tomato fields in an intensive agricultural landscape. Agric. Ecosyst. Environ. 2014, 189, 164-170. [CrossRef]
- Dyck, V.A.; Hendrichs, J.; Robinson, A.S. Sterile insect technique: principles and practice in area-wide integrated pest management; Taylor & Francis: 2021.
- Roselli, G.; Anfora, G.; Sasso, R.; Zapponi, L.; Musmeci, S.; Cemmi, A.; Suckling, D.M.; Hoelmer, K.A.; Ioriatti, C.; Cristofaro, M. Combining Irradiation and Biological Control against Brown Marmorated Stink Bug: Are Sterile Eggs a Suitable Substrate for the Egg Parasitoid Trissolcus japonicus? Insects 2023, 14, 654.
- Kohli, A.K. Gamma Irradiator Technology: Challenges and Future Prospects. Univers. J. Appl. Sci. 2018, 12, 47-51. [CrossRef]
- Mastrangelo, T.; Parker, A.G.; Jessup, A.; Pereira, R.; Orozco-Dávila, D.; Islam, A.; Dammalage, T.; Walder, J.M.M. A New Generation of X Ray Irradiators for Insect Sterilization. J. Econ. Entomol. 2010, 103, 85-94. [CrossRef]
- Kaboré, B.A.; Nawaj, A.; Maiga, H.; Soukia, O.; Pagabeleguem, S.; Ouédraogo/Sanon, M.S.G.; Vreysen, M.J.; Mach, R.L.; De Beer, C.J. X-rays are as effective as gamma-rays for the sterilization of Glossina palpalis gambiensis Vanderplank, 1911 (Diptera: Glossinidae) for use in the sterile insect technique. Sci. Rep. 2023, 13, 17633. [CrossRef]
- Congress.gov. Text - H.R.5515 - 115th Congress (2017-2018): An act to authorize appropriations for fiscal year 2019 for military activities of the Department of Defense, for military construction, and for defense activities of the Department of Energy, to prescribe military personnel strengths for such fiscal year, and for other purposes. Available online: https://www.congress.gov/bill/115th-congress/house-bill/5515/text (accessed on 05/28/2024).
- Bundy, C.S.; Grasswitz, T.R.; Sutherland, C. First report of the invasive stink bug Bagrada hilaris (Burmeister) (Heteroptera: Pentatomidae) from New Mexico, with notes on its biology. Southwest Entomol. 2012, 37, 411-414, doi:. [CrossRef]
- Reed, D.A.; Palumbo, J.C.; Perring, T.M.; May, C. Bagrada hilaris (Hemiptera: Pentatomidae), an invasive stink bug attacking cole crops in the southwestern United States. J. Integr. Pest Manag. 2013, 4, C1-C7, doi: . [CrossRef]
- Torres-Acosta, R.I.; Sánchez-Peña, S.R. Geographical distribution of Bagrada hilaris (Hemiptera: Pentatomidae) in Mexico. J. Entomol. Sci. 2016, 51, 165-167. [CrossRef]
- Faúndez, E.I. From agricultural to household pest: The case of the painted bug Bagrada hilaris (Burmeister)(Heteroptera: Pentatomidae) in Chile. J. Med. Entomol. 2018, 55, 1365-1368. [CrossRef]
- Colazza, S.; Guarino, S.; Peri, E. Bagrada hilaris (Burmeister)(Heteroptera: Pentatomidae) a pest of capper in the island of Pantelleria [Capparis spinosa L.; Sicily]. Inf. Fitopatol. 2004, 54.
- Palumbo, J.C.; Perring, T.M.; Millar, J.G.; Reed, D.A. Biology, ecology, and management of an invasive stink bug, Bagrada hilaris, in North America. Annu. Rev. Entomol. 2016, 61, 453-473. [CrossRef]
- Stark, J.D.; Banks, J.E. Population-level effects of pesticides and other toxicants on arthropods. Annu. Rev. Entomol. 2003, 48, 505-519.
- Taylor, M.E.; Bundy, C.S.; McPherson, J.E. Unusual ovipositional behavior of the stink bug Bagrada hilaris (Hemiptera: Heteroptera: Pentatomidae). Ann. Entomol. Soc. Am. 2014, 107, 872-877. [CrossRef]
- Tofangsazi, N.; Hogg, B.N.; Hougardy, E.; Stokes, K.; Pratt, P.D. Host searching behavior of Gryon gonikopalense and Trissolcus hyalinipennis (Hymenoptera: Scelionidae), two candidate biological control agents for Bagrada hilaris (Hemiptera: Pentatomidae). 2020, 151, 104397.
- Hogg, B.N.; Hougardy, E.; Talamas, E. Adventive Gryon aetherium Talamas (Hymenoptera, Scelionidae) associated with eggs of Bagrada hilaris (Burmeister)(Hemiptera, Pentatomidae) in the USA. J. Hymenopt. Res. 2021, 87, 481.
- Martel, G.; Sforza, R.F. Evaluation of three cold storage methods of Bagrada hilaris (Hemiptera: Pentatomidae) and the effects of host deprivation for an optimized rearing of the biocontrol candidate Gryon gonikopalense (Hymenoptera: Scelionidae). Biol. Control 2021, 163, 104759. [CrossRef]
- Mansour, M. Effects of gamma radiation on the Mediterranean flour moth, Ephestia kuehniella, eggs and acceptability of irradiated eggs by Trichogramma cacoeciae females. J. Pest Sci. 2010, 83, 243-249. [CrossRef]
- Haff, R.; Jackson, E.; Gomez, J.; Light, D.; Follett, P.; Simmons, G.; Higbee, B. Building lab-scale x-ray tube based irradiators. Radiat. Phys. Chem. 2016, 121, 43-49. [CrossRef]
- Liang, P.-S.; Haff, R.P.; Ovchinnikova, I.; Light, D.M.; Mahoney, N.E.; Kim, J.H. Curcumin and quercetin as potential radioprotectors and/or radiosensitizers for X-ray-based sterilization of male navel orangeworm larvae. 2019, 9, 2016. [CrossRef]
- Hougardy, E.; Hogg, B.N. Host patch use and potential competitive interactions between two egg parasitoids from the family Scelionidae, candidate biological control agents of Bagrada hilaris (Hemiptera: Pentatomidae). J. Econ. Entomol. 2021, 114, 611-619. [CrossRef]
- Mohammadpour, M.; Jalali, M.A.; Michaud, J.; Ziaaddini, M.; Hashemirad, H. Multiparasitism of stink bug eggs: competitive interactions between Ooencyrtus pityocampae and Trissolcus agriope. 2014, 59, 279-286. [CrossRef]
- R Development Core Team R: A language and environment for statistical computing, R Foundation for Statistical Computing: Vienna, Austria, 2023.
- Ozyardimci, B.; Cetinkaya, N.; Denli, E.; Ic, E.; Alabay, M. Inhibition of egg and larval development of the Indian meal moth Plodia interpunctella (Hübner) and almond moth Ephestia cautella (Walker) by gamma radiation in decorticated hazelnuts. J. Stored Prod. Res. 2006, 42, 183-196. [CrossRef]
- Ayvaz, A.; Albayrak, S.; Karaborklu, S. Gamma radiation sensitivity of the eggs, larvae and pupae of Indian meal moth Plodia interpunctella (Hübner)(Lepidoptera: Pyralidae). Pest Manag. Sci. 2008, 64, 505-512.
- Donoughe, S. Insect egg morphology: evolution, development, and ecology. Curr. Opin. Insect Sci. 2022, 50, 100868. [CrossRef]
- Godfray, H.C.J. Parasitoids: behavioral and evolutionary ecology; Princeton University Press: 1994.
Figure 1.
Interior of the x-ray irradiator showing samples in Ziploc bags rotating adjacent to the radiation source. Real-time dose measurements from the attached ion probe were read at an external display, and x-rays were discontinued when the desired dose was reached.
Figure 1.
Interior of the x-ray irradiator showing samples in Ziploc bags rotating adjacent to the radiation source. Real-time dose measurements from the attached ion probe were read at an external display, and x-rays were discontinued when the desired dose was reached.

Figure 3.
Percentage of 1, 2, 3, and 7 d old (refrigerated) eggs showing no or some embryo development, or eclosed 9 days post irradiation (40 Gy).
Figure 3.
Percentage of 1, 2, 3, and 7 d old (refrigerated) eggs showing no or some embryo development, or eclosed 9 days post irradiation (40 Gy).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



