
Submitted:
18 September 2024
Posted:
18 September 2024
You are already at the latest version
Abstract
Several studies in recent years have shown that the use of non-Saccharomyces yeasts, used both in single and in mixed fermentations with Saccharomyces cerevisiae can help produce craft beers with distinctive compositional characteristics. The aim of this study was to evaluate the suitability of three Metschnikowia pulcherrima strains, isolated from Albanian vineyards, to be used as starters in the brewing process. Due to its specific enzymatic activities (protease, β-glucosidase and β-lyase) and its low production of hydrogen sulfide, M. pulcherrima 62 was selected as a starter culture for the production of craft beer. Specifically, the aptitude of this yeast to be used in sequential inoculation with S. cerevisiae S04 for the production of an American IPA-style beer and the main volatile compounds produced during fermentation were evaluated. Results showed significant differences in glycerol, isoamyl alcohol and isoamyl acetate content in beer obtained with sequential inoculum of M. pulcherrima 62 with S. cerevisiae S04 compared to beer obtained using S. cerevisiae S04 as a single starter. Therefore, these preliminary data support a candidacy of M. pulcherrima 62 as a new starter in the brewing process.
Keywords:
craft beer
; Metschnikowia pulcherrima
; Saccharomyces cerevisiae
; sequential inoculation
1. Introduction
In recent years, the craft beer production sector has increased widespread, with a growing consumer interest in new beer with distinctive organoleptic characteristics [1,2,3,4].
Among the adopted strategies, in addition to the use of unconventional ingredients (e.g. alternative grains, exotic hops) and innovative brewing techniques (e.g. spontaneous fermentation, barrel-aging) [5,6,7], the role of non-Saccharomyces yeasts is a trending topic to improve the sensory characteristics of the beers [8]. For decades, the use of starters in the brewing industry has been almost exclusively limited to pure yeast cultures belonging to the genus Saccharomyces. This well-established technique ensured better control of the fermentation process and, as a result, helped to elevate and standardize the quality of the beers. In recent years, in the brewing sector, the need to diversify final products as a business strategy has grown more and more to conquer new markets and meet the needs of increasingly demanding and quality-conscious consumers. On the basis of these considerations, especially in the craft beer industry, non-Saccharomyces yeasts represent a great source of biodiversity and opens up new possibilities compared to traditional Saccharomyces genus, in order to obtain distinctive products, with peculiar compositional and organoleptic characteristics [9,10,11,12,13,14,15,16]. Multiple studies in recent years have highlighted the potential of wild Saccharomyces and non-Saccharomyces yeasts, isolated from spontaneously fermented beers as well as from non-brewing environments (e.g. wines, vineyards, sourdoughs, honey by-products), for the production of beers with appreciable and distinctive compositional and sensory characteristics compared to those obtained with conventional brewer’s yeasts [17,18,19,20,21,22,23]. Although the use of non-Saccharomyces yeasts in beer production is relatively recent, the genus Metschnikowia is one of the most studied, due to its multiple positive contributions in winemaking processes and recently in beer production [24,25,26]. In particular, Metschnikowia pulcherrima is recommended in winemaking to obtain wines with reduced ethanol content, for the bio-control, for their contribution to the aromatic development of wines through their enzymatic activities (e.g. β-D-glucosidase, cysteine β-lyase) and the production of a wide range of metabolites (e.g. esters, higher alcohols) resulting from alcoholic fermentation [26,27,28,29].
On the basis of their low tolerance to alcohol and their enzymatic activities, recent studies have verified that selected M. pulcherrima strains are functional for obtaining beers with a low alcohol content, if used in pure culture, or can contribute to positively modifying the organoleptic properties of beers if used in co-culture with S. cerevisiae [30,31,32,33].
However, while for the wine sector there is a limited availability of commercial strains belonging to this species, in the beer sector this availability is not yet there.
The present study aims to evaluate new M. pulcherrima strains, previously isolated from vineyards located in Albania [34], to be specifically targeted for the brewing industry. After a preliminary screening, M. pulcherrima 62 was selected as a starter for craft beer production. Specifically, the aptitude of this yeast to be used in sequential inoculation with S. cerevisiae for the production of an American IPA-style beer was evaluated.
2. Materials and Methods
2.1. Yeast Strains and Growth Condition
For this study M. pulcherrima 62, 82 and 86 strains (GenBank accession numbers: PP922572.1, PP922568.1, PP922571.1), belonging to the culture collection of the Agri-Food Research Centre of the Faculty of Biotechnology and Food of Agriculture University of Tirana, were used. These strains were previously isolated from autochthonous Albanian red grapes [34]. For the preliminary characterization and for the brewing trials, the commercial S. cerevisiae S04 (Fermentis, Lesaffre, France) strain was used as a reference. For the beer fermentation, the yeasts were cultured aerobically at 28 °C in YEPD broth (Merck Millipore, Darmstadt, Germany) and after 48 hours the broth cultures were centrifuged at 8,000 rpm for 10 min at 4 °C. Finally, the cell pellet was washed twice with saline solution (0.9% w/v NaCl) and used as a starter. Cell density of inoculum was assessed using Thoma Counting Chamber (Thermo Fisher Scientific).
2.2. Pre selection Trials
2.2.1. Carbon Assimilation Profiles
Carbon sources assimilation profiles were evaluated using the API 20 C AUX system (Biomèrieux, Montalieu-Vercieu, France) based on 19 carbohydrate assimilation tests plus a negative control, read by assessing cupules for turbidity. The kit was used in accordance with the guidelines given by the manufacturer. The yeast strains before use were cultured in YEPD broth at 28 °C for 48h. Reading of the strips was done after 48 and 72 h of incubation at 30°C.
2.2.2. Cryotolerance
The cryotolerance was evaluated as reported by Iorizzo et al. [11]. Each strain pre-grown overnight in YEPD at 25 °C was inoculated at an initial concentration of 1x102 CFU/mL into Erlenmeyer flasks (100 mL capacity) containing 80 mL of YEPD, maintained under stirring using a digital orbital shaker (Heathrow Scientific, IL, USA) at 4°C. The growth was determined visually after 24 h of incubation.
2.2.3. Biogenic Amines Production
Biogenic amines production was carried as described by Barbosa et al. [35] with some modification. For this purpose, a culture media containing 3 % (wt/vol) yeast extract, 1 % (wt/vol) glucose, 2 % (wt/vol) amino acid precursor (histidine, tyrosine, ornithine, phenylalanine and histidine), and 0.015 g L−1 (w/v) bromocresol purple, was used. The pH was adjusted to 5.2. The medium without amino acid precursor was used as negative control. The decarboxylation of the amino acids to the corresponding biogenic amines results in an increase in pH, detected by the culture medium colour change. All reagents were purchased from Merck KGaA (Darmstadt, Germany).
2.2.4. Hydrogen Sulphide Production
Hydrogen sulphide (H2S) production by M. pulcherrima strains, was evaluated according to Comitini et al. [36] using BIGGY agar (Bismuth Sulphite Glucose Glycine Yeast; Thermo Fisher Scientific, Waltham, MA, USA), as medium. After 3 days, H2S-negative strains showed white colonies, while H2S-producing colonies were characterized by a brown or dark brown color. For results, the following chromatic scale was considered: 0 (white colonies, no H2S production), 1 (cream colonies), 2 (light brown colonies), 3 (brown colonies), 4 (dark brown or black colonies, very intensive H2S production).
2.2.5. Pulcherrimin Production
Pulcherrimin production, was evaluated according to the method described by Mažeika et al. [37] with some modification, using a culture medium with the following composition: 1% (wt/vol) yeast extract, 2% (wt/vol) peptone, 2% (wt/vol) glucose and 2% (wt/vol) agar. After sterilization, the medium was supplemented with sterile FeCl3 solution (0.05% wt/vol) and poured into Petri dishes. Finally, 10 µL of yeast cultures (106 cells/mL) were spotted onto the medium and the plates were incubated for 3 days at 28 °C. After incubation, colonies surrounded by reddish halos around the colonies were recorded as positive results. All reagents used in the experiment were purchased from Merck KGaA (Darmstadt, Germany).
2.3. Enzymatic Activities
2.3.1. API zym Assay
Screening of enzymatic activities was evaluated using the API ZYM system (Biomèrieux, Montalieu-Vercieu, France) according to the manufacturer’s instructions. The yeast strains were cultured in YEPD broth at 28 °C. After 48 h, 60 µL the yeast cell suspensions were transferred into the wells of the API ZYM strips and incubated at 37 °C for 4 h. The colour changes observed into the wells indicating positive enzymatic reactions and were used for evaluation of the results on the basis of the API ZYM colour chart.
2.3.2. Proteolytic Activity
The proteolytic activity was detected qualitatively by using skim milk agar hydrolysis method as previously described by Gut et al. [38] with some modifications. Briefly, 10 µL of yeast suspensions in YEPD broth (106 cells/mL), were spotted onto SDA (Sabouraud Dextrose Agar) containing 10 % (wt/vol) skim milk (Merck KGaA, Darmstadt, Germany), with a final medium pH of 7.3. Plates were incubated at 28 °C for 5 days. Proteolityc activity was indicated by the presence of a clear zone around the colony.
2.3.3. β-glucosidase Activity
Qualitative assay of β-Glucosidase activity, was performed as reported by Testa et al. [39]. For this purpose, 10 µL of yeast suspensions in YEPD broth (106 cells/mL), were spotted onto a culture medium having the following composition: 2 g/L (wt/vol) glucose, 1 g/L (wt/vol) peptone, 1 g/L (wt/vol) yeast extract, 0.3 g/L (wt/vol) esculin, 0.01g/L (wt/vol) ferric-ammonium citrate and 15 g/L (wt/vol) agar (Merck KGaA, Darmstadt, Germany). Plates were incubated at 28°C for 3 days. The β-Glucosidase activity based on appearance of black zone around the colonies indicating the hydrolysis of esculin.
2.3.4. β-lyase Activity
Qualitative screening of β-lyase activity was conducted as described by Belda et al. [40], using a medium containing 0.1 % (wt/vol) S-methyl-l-cysteine, 0.01 % (wt/vol) pyridoxal-5′-phosphate, 1.2 % (wt/vol) Yeast Carbon Base and 2 % (wt/vol) agar with a final medium pH of 3.5. 10 µL of yeast suspensions in YEPD broth (106 cells/mL), were spread onto the plate’s surface and incubated at 25 °C for 72 h. The growth of yeast cultures after 72h of incubation indicated the presence of β-lyase activity [41]. All reagents were purchased from Merck KGaA (Darmstadt, Germany).
2.4. Craft Beer Brewing Process
After the pre-selection tests, M. pulcherrima 62 was chosen as starter for the American IPA-style beer production using a Grainfather G series brewing system (Bevie Handcraft, Nelson NZ Ltd) at the Department of Agricultural, Environmental and Food Sciences (University of Molise; Campobasso, Italy). For American IPA style beer production, pale ale and cara crystal malts (Château Pale ale, Castle Malting, Lambermont, Belgium) were used. Amarillo and cascade hops (Barth-Hass, Nürnberg, Germany) were added during the boiling and dry hopping phase. The main analytical characteristics of the wort, meeting the requirements established by Beer judge certification program (BJCP) [42] for the American IPA, were as follows: pH 5.50, °Plato 11.7, density (original gravity) 1.047 g/cm3, IBU (International Bitterness Unit) 54, and FAN (free amino nitrogen) 208.67 mg/L. Fermentation tests were carried out at 20°C ± 1°C using thermoregulated stainless steel tanks (capacity 30 L), containing 20 L of wort. In detail, test A was inoculated initially with M. pulcherrima 62 and after 48 h with S. cerevisiae S04 (sequential inoculum) while test B was inoculated only with S. cerevisiae S04. The starter cultures were inoculated at an initial concentration around 106 CFU/mL and the fermentations were conducted at 20 ± 1°C. The fermentations were performed in triplicate. At the end of the primary fermentation, 3.5 g/L of sucrose was added as a primer for the secondary fermentation which took place in 330 mL dark brown glass bottles. After 40 days of maturation at 20 °C, the beers were subjected to chemical analysis.
2.5. Fermentation Kinetics
During the primary fermentation, density values (g/cm3) were monitored. For the determination of yeast cell viability (CFU/mL), the culture medium WL agar containing 100 mg/L chloramphenicol (Merck KGaA, Darmstadt, Germany) was used. The plates were incubated at 28 °C in aerobic conditions. After 72 hours, the colonies were evaluated by color and topography to distinguish S. cerevisiae from M. pulcherrima [29].
2.6. Main Chemical Parameters of Beers
A pH meter (Crison basic 20, Barcelona, Spain) was used to measure the pH. The density (g/cm3), alcohol content (% v/v), FAN and IBU were determined according to analytical procedures as described by European Brewery Convention (EBC) [43]. Glycerol (mg/L), acetic acid (mg/L), acetaldehyde (mg/L), L-malic acid (g/L) and L-lactic acid (g/L) were determined using enzymatic kits (Steroglass, Perugia, Italy) according to the manufacturer’s instructions. All measurements were conducted in triplicate.
2.7. Analysis of Volatile Compounds by GC-FID
The volatile compounds were determined by Gas Chromatography with flame ionization detection (GC-FID), using a GC2010 Plus apparatus with FID-2010 detector equipped with a headspace autosampler (HS-20) (Shimadzu Corporation, Kyoto, Japan) and a CP-WAX 57 CB column (50 m×0.32 mm ID×0.2 μm) (Agilent Technologies, Santa Clara, CA, USA). Quantification of volatile compounds was performed using external standards based on a standard curve with five calibration points (the coefficient of determination R2 was greater than or equal to 0.999). The limit of detection was 0.1 mg/l. The analysis method was conducted according to Paszkot et al [44]. All measurements were conducted in triplicate.
2.8. Statistical Analysis
Results were expressed as mean value ± standard deviation (SD). Statistical analyses were performed using t-test and one-way analysis of variance ANOVA (IBM SPSS Statistics 21) and Tukey post-hoc tests at a significance level of p < 0.05.
3. Results and Discussion
3.1. Technological and Biochemical Properties
The results of API 20 C AUX test are shown in Figure 1 as heat map. The assimilation profiles of the 19 sugars tested were similar but the inability of M. pulcherrima K191C7 to ferment D-Xylose and M. pulcherrima K185C2-9 to ferment D-Melezitose has been detected. All M. pulcherrima strains tested were able to assimilate glucose, maltose and saccharose that represent the main sugars present in beer wort. These assimilative capacities make them suitable for their possible use as a starter in beer production.
The results on the H2S and pulcherrimin production, cryotolerance and β -Glucosidase, β -Lyase and Protease activities of the three M. pulcherrima strains are reported in Table 1. M. pulcherrima 62 was able to grow at 4 °C, while M. pulcherrima 86 and 82 showed a very weak growth capacity at this temperature. Low-temperature fermentation by Saccharomyces spp. is believed to lead to the production of wines and beers with improved taste and aroma due to increased production of volatile aromatic compounds [45,46].
Therefore, the cryotolerance of M. pulcherrima 62 could be functional to production of low-temperature fermented products such as lager beers. Regarding the qualitative tests we carried out to evaluate the production of biogenic amines, our results showed that the three M. pulcherrima strains are not producers of these nitrogenous compounds. In alcoholic beverages, the biogenic amines are primarily formed by decarboxylation or transamination of precursor amino acids, which are directly affected by the activity of amino acid decarboxylase in yeasts and lactic acid bacteria (LAB) [47,48,49].
Biogenic amines in food might represent a potential public risk health because of their physiological and toxicological effects. In foods, their concentrations typically increase during processing and storage because of exposure to microorganisms that catalyze their formation [50]. In recent years there has been an increase in the number of cases of food poisoning related to biogenic amines in foods and beverages [51]. Therefore, it is extremely important to monitor the level of biogenic amines in food and alcoholic beverages such as beer. The European legislation does not specify a biogenic amines threshold, but the European Food Safety Authority (EFSA) has elaborated a scientific opinion on the risk associated with the formation of these compounds in fermented products [52].
During fermentation, yeasts are responsible for the production of several sulfur compounds including H2S. This compound can have an undesirable impact by directly affecting the flavor profile or masking other flavor compounds found in beer [20]. H2S is a highly volatile compound with a very low flavor threshold level (11–80 μg/L) reminiscent of rotten eggs [53]. Our results, pertaining semi-quantitative tests, highlighted the low production of H2S by the M. pulcherrima strains.
Pulcherrimin-producing yeast species, like M. pulcherrima, are considered effective antimicrobial agents against various microorganisms, with great potential for biocontrol applications [54]. The M. pulcherrima strains tested in this study, were results pulcherrimin producers. Pulcherrimin has inhibitory activity against several yeast species, but seems S. cerevisiae not to be affected by this antimicrobial activity [55,56,57]. This appears to be confirmed in the beer production trials discussed below, which showed that M. pulcherrima 62 did not cause any interference on alcoholic fermentation by S. cerevisiae S04.
The use of non-Saccharomyces yeasts, possessing specific enzymatic activities, is still an innovative concept in beer production and opens possibilities to produce beers with distinctive sensory characteristics, compared to the use of single cultures of Saccharomyces species [32].
The results of API ZYM test are presented in Figure 1 as heat map. All the M. pulcherrima strains exhibited the following enzymatic activities: Phosphohydrolase, α- glucosidase, β-glucosidase, acid phosphatase, valine and leucine arylamidase.
Leucine arylamidase is involved in the production of leucine, which is needed for cyclodileucine (cyclo(Leu-Leu)) formation—the precursor of pulcherriminic acid [58].
M. pulcherrima 62 and M.pulcherrima 82 possess esterase lipase (C8), and esterase (C4) activities. Esterase are the enzymes involved in the release of phenolic compounds from plant cell walls [59]. Moreover, previous studies suggesting a crucial link between esterase activity and aroma production during fermentation [16,60].
Some non-Saccharomyces yeasts, like M. pulcherrima, possess β-glucosidase enzyme, that can hydrolize glycoconjugate precursors and promote the release of active aromatic compounds [61]. In our study, it was found that all three M. pulcherrima strains possessed β-glucosidase activity. During the alcoholic fermentation of beer wort, the yeasts produced volatile compounds, as higher alcohol and esters, which directly contribute to the organoleptic characteristics of the final product (Iorizzo et al., 2021). In addition to these compounds, β-glucosidase activity results in the release of monoterpene alcohols like linalool, α-terpineol, β-citronellol, geraniol, and nerol [61]. Terpenes can have diverse flavor impacts (citrus, floral) and higher levels of these compounds are associated with greater overall hop aroma intensity. In wort, these compounds are often present in glycosidically bound forms and aromatically inactive [62]. In addition, our study detected β-lyase activity produced from the tested M. pulcherrima strains. This enzymatic activity results in the release of volatile flavour-active thiols from their conjugated (glutathionylated or cysteinylated) and therefore aroma-inactive forms present in hops [63]. Volatile thiols are active at very low flavor thresholds and impart tropical, citrus, and other fruity aromas to beers [64,65]. Our results showed that M. pulcherrima 62 and M. pulcherrima 86 possess protease activity. Yeast extracellular proteases have potential in beer stabilization, facilitating the filtration and clarification. In addition, protein degradation results in amino acids that are a source of nitrogen for yeast growth and are precursors for the biosynthesis of higher alcohols, which significantly influence the aroma and flavour of beers [66]. Based on the screening results M. pulcherrima 62 was chosen as starter for the production of an American IPA-style beer on a pilot-scale.
3.2. Main Chemical Parameters of Beers
The data on the main physical-chemical parameters of the beers are shown in Table 2. The ethanol values (% v/v) were respectively 5.2 % v/v in beer obtained in test A and 5.0 % v/v in beer obtained in test B, thus showing non-significant differences. Therefore, sequential inoculation did not affect the level of ethanol in the beer. In fact, S. cerevisiae S04 both in co-culture and as a single starter managed to complete alcoholic fermentation after 10 days. These data confirm the findings in other studies that the antimicrobial activity of M. pulcherrima does not cause any interference with the completion of alcoholic fermentation by S. cerevisiae [55,67].
In our study, the glycerol content was significantly higher in test A (1026.02 ± 28.21) than in test B (892.66 ± 9.60). Complex interactions and considerable differences have been shown in the metabolism of S. cerevisiae in single culture and in co-culture with non-Saccharomyces yeasts [68,69]. As reported in previous studies, it appears that there may be an increase in glycerol when M. pulcherrima is used in mixed culture with S. cerevisiae under controlled oxygenation conditions during the first 48 hours of fermentation and subsequently under anaerobic conditions [69,70]. These studies suggest that M. pulcherrima may have run out of oxygen during the 48 hours prior to inoculation of S. cerevisiae. Lower oxygen conditions could explain the modulation of glyceropyruvic fermentation and the orientation of metabolism towards pyruvate dehydrogenase (PDH) bypass in S. cerevisiae leading to increased glycerol production [71]. Glycerol influence beer taste because of its sweetness, improve foam stability, enhance the aroma volatility and increase the worty off-flavor retention [72]. Its content in beer is usually higher than a great many other flavor compounds (e.g. higher alcohols, esters, and organic acids), and the addition of glycerol to beer above and below the threshold level (10 g/L) was found to modify the flavor of the product [73]. No significant difference in lactic acid content was detected in the beers while lower amounts of L-malic acid (160.33 ± 2.51 mg/L) were detected in beer obtained in Test A compared to beer obtained using S. cerevisiae as a single starter (Test B) in which 202.33 ± 7.50 mg/L of L-malic acid were detected. In a previous study, it emerged that some M. pulcherrima strains are able to metabolize malic acid in wine [74]. This could explain the significant differences of these two organic acids in the beers analyzed. Finally, the analyses conducted in our study found that the quantitative values of acetaldehyde and acetic acid were not significantly different in the beers obtained in the A and B tests, thus confirming that M. pulcherrima produces low amounts of these two compounds [36,75,76]. Acetic acid is the main component of the volatile acids in beer and its threshold range is from 71 to 200 mg/L [77]. Therefore, the quantities measured in our fermentation trials (Test A: 60.66 ± 3.51 mg/L; Test B: 58.83 ± 1.25 mg/L), are lower than these values. Acetaldehyde (25 g/L threshold), is the carbonyl present in beer at the conclusion of primary fermentation in the highest concentration as a result of the decarboxylation of pyruvate and is an intermediate in the metabolic formation of ethanol during glycolysis [78,79]. In our study this compound was present in all beers and, in low concentrations such as those detected (Test A: 7.66 ± 0.80; Test B: 7.70 ± 0.19), can contribute positively to the sensory character of the beers (green apple, umpkin pulp/seed, unripe avocado) [14,80,81].
3.3. Fermentation Kinetics
The pH and density trends, monitored during alcoholic fermentation, is illustrated in Figure 2. The relative numeric data are reported in Table S1 (supplementary material).
The pH from an initial value of about 5.50 in both tests, gradually decreased, until it reaches final values between 4.35 in Test A and 4.31 in Test B without significant differences. During fermentation, the lowering of pH is due to the yeast’s consumption of compounds with buffering capacity (i.e., amino acids) and the production of organic acids.
In Test A, initially inoculated with M. pulcherrima 62, the density was decreased after 48 h of fermentation, from 1.047 to 1.042 g/cm3. After sequential inoculation with S. cerevisiae S04, density values decreased rapidly reaching 1.007 g/cm3 at the end of alcoholic fermentation (10 days). In Test B, the fermentative vigor of S. cerevisiae S04 caused a greater decrease in density after 24 hours (1.022 g/cm3). After 10 days of fermentation, at the end of alcoholic fermentation, the density values were not significantly different in the beers obtained in Tests A and B. Our results showed that the use of M. pulcherrima 62 as an initial starter did not negatively affect the fermentation activity of S. cerevisiae S04. Regarding the yeast viable cells count (Table 3), the use of WL agar medium allowed us to evaluate the populations of viable yeasts present during the alcoholic fermentation and in particular confirmed the aptitude of this culture medium to differentiate S. cerevisiae from M. pulcherrima according to colony color and colony topographic parameters (Figure 3). Validation of the taxonomic collocation of yeasts species was carried out by 26S rDNA D1/D2 domain sequence analysis, using NL1 (5′-GCA TAT CAATAA GCG GAG GAA AAG-3′) and NL4 (3′-GGT CCG TGT TTC AAG ACGG-5′) as primers [29].
In Test A, the population of M. pulcherrima averaged 6.93 log CFU/mL after 2 days of fermentation, then decreased and was not detected 5 days after inoculation. In both tests, the density of viable S. cerevisiae cells increased after inoculation to values greater than 7 log CFU/mL from day 4 and until the end of alcoholic fermentation. The fermentation kinetics reported in Table 3 confirm, as already found in previous studies, that M. pulcherrima, while possessing multiple antimicrobial activities, does not negatively affect the viability and metabolic activity of S. cerevisiae [28,30,55,82].
3.4. Volatile Compounds
The concentrations of volatile compounds identified in beers by GC-FID are shown in Table 4. The flavour and aroma of the beers originate from the raw materials (malt, hops and yeast) and the metabolic reactions that take place during fermentation that generate ethanol and other co-products, mainly volatile compounds such as higher alcohols, esters, acids and aldehydes [78,83].
Higher alcohols production by yeast occurs through the Ehrlich pathway either from amino acids transported over the cell membrane or through de novo biosynthesis of amino acids. After the initial transamination α-ketoacid intermediates are excreted in the growth medium and yeasts convert them into alcohols or acids via the Ehrlich pathway. In our tests, the identified alcohols, regardless of the initial starter culture, were isobutanol, isoamyl alcohol, 2-phenylethanol and 1-hexanol. Our results confirmed that these compounds are the most abundant higher alcohols found in beer [84,85]. However, the amounts of these compounds were significantly different in the beers obtained in the A and B Tests (Table 4). As for isoamyl alcohol, obtained using M. pulcherrima 62 as the initial starter culture (Test A), its amount (101.83 ± 1.55 mg/L) was well above its flavor threshold in beer (70 mg/L) [14]. This alcohol has also some banana flavor character and has been identified above its threshold in banana, orange, mango, pineapple and passion fruit [84]. As for isobutanol, the highest concentrations was obtained in the sequential fermentation with M. pulcherrima 62 (42.26 ± 0.75 mg/L) which has been previously reported as a high isobutanol producer [32,86]. Higher alcohols not only impact on flavour, but also provide the alcohol moiety required for the synthesis of esters, which represent the largest and possibly most important group of flavour-active compounds in beer [78].
Flavor-active esters represent an important group of compounds, which give fruity and flowery aromas to beer [87]. These volatile compounds are formed largely during the active phase of the primary fermentation by the enzymatic condensation between activate fatty acids (acyl-CoA or Acetyl-CoA) and higher alcohols [84]. Among the esters, we detected ethyl acetate and isoamyl acetate in the highest concentrations. In particular, in the beers obtained in Tests A and B were quantified respectively: ethyl acetate 1.31 ± 0.18 mg/L (Test A) and 0.68 ± 0.04 mg/L (Test B); isoamyl acetate 2.50 ± 0.14 mg/L (Test A) and 1.88 ± 0.04 mg/L (Test B). The threshold concentration of ethyl acetate in beer is 30 mg/L, but for lager-type beers the recommended concentration is <5 mg/L [88]. Therefore, the concentrations detected in the beers we produced were well below these threshold levels of perception. M. pulcherrima has been described as a good producer of isoamyl acetate in beer [86].This ester is formed by condensation of acetyl CoA and isoamyl alcohol during fermentation and its intensive ‘fruity’ aroma (banana, apple, pear) is perceived at concentrations >1.2 mg/L [78,89]. In our study, other esters are present at concentrations below their threshold value. However, the presence of different esters can have a synergistic effect on individual flavors, which means that esters as a whole can also affect the flavor of beer well below their individual threshold values [14,90].
In our study, among the volatile compounds, butyric, hexanoic, octanoic and decanoic fatty acids were detected. However, the quantities present in the beers produced were all below the sensory perception thresholds [91,92]. Fatty acids have a beneficial effect on yeast growth during fermentation, but can negatively affect the organoleptic properties of beer and the stability of the beer foam [93,94]. These compounds and their oxidized forms cause unpleasant off-flavour of beer, such as rancid, cheesy, soapy, fatty, butyric and others [95,96]. Finally, between the beers obtained in Test A and those obtained in Test B, no significant difference in the content of diacetyl and acetoin was found. This means that the use of M. pulcherrima 62 as a starter does not result in an increase in the production of these substances. Diacetyl (butanedione or butane-2,3-dione) is a vicinal diketone generated as a by-product of amino acid metabolism in yeast during wort fermentation and are secreted into beer, imparting aroma characteristics described as butter, butterscotch, or buttermilk when detected above its flavor threshold > 0.1 mg/L [79]. During maturation process, the yeast reabsorbs the diacetyl and converts it to acetoin and subsequently to 2,3-butanediol. Both acetoin and 2,3-butanediol can escape the cell, but neither contribute much in terms of flavour.
5. Conclusions
The use of non-Saccharomyces yeasts is a biotechnological strategy increasingly pursued in brewing processes to obtain a greater diversification of beers and improve their sensory characteristics. In the present study we presented the results obtained in the production of an American IPA-style beer on a pilot scale using M. pulcherrima 62 in sequential inoculation with S. cerevisiae S04.
The higher glycerol, isoamyl alcohol and isoamyl acetate contained in beer obtained by sequential inoculation of M. pulcherrima 62 with S. cerevisiae S04 compared to beer obtained using S. cerevisiae S04 as a single starter appear remarkable. These data, while supporting a valid candidacy of M. pulcherrima 62 as a new starter in beer production, need to be further validated with investigations on the volatile compounds of beer, using more performing technique such as Gas Chromatography-Mass Spectrometry (GC-MS), together with a sensory evaluation of the final product. In the future, it may also be interesting to evaluate the influence of M. pulcherrima 62 on the quality of other beer styles.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: pH and density trends monitored during alcoholic fermentation. Test A, inoculated with M. pulcherrima 62 and S. cerevisiae S04 in sequential inoculum; Test B inoculated with S. cerevisiae as a single starter.
Author Contributions
Conceptualization, M.I., B.T. and J.K.; methodology, B.T. and J.K.; software, F.L. (Francesco Letizia); validation, M.I., K.S., F.C., and R.K.; formal analysis, B.T. and J.K.; data curation, M.I., B.T. and J.K.; writing—original draft preparation, J.K.; writing—review and editing, M.I., B.T., F.L. (Francesco Letizia) and J.K.; visualization, O.K., N.X., M.R. and F.L. (Fatbardha Lamçe); supervision, M.I., R.K. and K.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding authors.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Garavaglia, C.; Swinnen, J. 216Industry Concentration and the Entry of Craft Producers into the Global Beer Market. In New Developments in the Brewing Industry: The Role of Institutions and Ownership; Madsen, E.S., Gammelgaard, J., Hobdari, B., Eds.; Oxford University Press, 2020; p. 0. ISBN 978-0-19-885460-9. [Google Scholar]
- Aquilani, B.; Laureti, T.; Poponi, S.; Secondi, L. Beer Choice and Consumption Determinants When Craft Beers Are Tasted: An Exploratory Study of Consumer Preferences. Food Qual. Prefer. 2015, 41, 214–224. [Google Scholar] [CrossRef]
- Baiano, A. Craft Beer: An Overview. Compr. Rev. Food Sci. Food Saf. 2021, 20, 1829–1856. [Google Scholar] [CrossRef] [PubMed]
- Villacreces, S.; Blanco, C.A.; Caballero, I. Developments and Characteristics of Craft Beer Production Processes. Food Biosci. 2022, 45, 101495. [Google Scholar] [CrossRef]
- De Simone, N.; Russo, P.; Tufariello, M.; Fragasso, M.; Solimando, M.; Capozzi, V.; Grieco, F.; Spano, G. Autochthonous Biological Resources for the Production of Regional Craft Beers: Exploring Possible Contributions of Cereals, Hops, Microbes, and Other Ingredients. Foods 2021, 10, 1831. [Google Scholar] [CrossRef]
- Puligundla, P.; Smogrovicova, D.; Mok, C. Recent Innovations in the Production of Selected Specialty (Non-Traditional) Beers. Folia Microbiol. (Praha) 2021, 66, 525–541. [Google Scholar] [CrossRef] [PubMed]
- Díaz, A.B.; Durán-Guerrero, E.; Lasanta, C.; Castro, R. From the Raw Materials to the Bottled Product: Influence of the Entire Production Process on the Organoleptic Profile of Industrial Beers. Foods 2022, 11, 3215. [Google Scholar] [CrossRef] [PubMed]
- Iorizzo, M.; Coppola, F.; Letizia, F.; Testa, B.; Sorrentino, E. Role of Yeasts in the Brewing Process: Tradition and Innovation. Processes 2021, 9, 839. [Google Scholar] [CrossRef]
- Larroque, M.; Carrau, F.; Fariña, L.; Boido, E.; Dellacassa, E.; Medina, K. Effect of Saccharomyces and Non-Saccharomyces Native Yeasts on Beer Aroma Compounds. Int. J. Food Microbiol. 2021, 337, 108953. [Google Scholar] [CrossRef]
- Methner, Y.; Hutzler, M.; Zarnkow, M.; Prowald, A.; Endres, F.; Jacob, F. Investigation of Non-Saccharomyces Yeast Strains for Their Suitability for the Production of Non-Alcoholic Beers with Novel Flavor Profiles. J. Am. Soc. Brew. Chem. 2022, 80, 341–355. [Google Scholar] [CrossRef]
- Iorizzo, M.; Letizia, F.; Albanese, G.; Coppola, F.; Gambuti, A.; Testa, B.; Aversano, R.; Forino, M.; Coppola, R. Potential for Lager Beer Production from Saccharomyces cerevisiae Strains Isolated from the Vineyard Environment. Processes 2021, 9, 1628. [Google Scholar] [CrossRef]
- Vanderhaegen, B.; Neven, H.; Verachtert, H.; Derdelinckx, G. The Chemistry of Beer Aging–a Critical Review. Food Chem. 2006, 95, 357–381. [Google Scholar] [CrossRef]
- Bellut, K.; Arendt, E.K. Chance and Challenge: Non-Saccharomyces Yeasts in Nonalcoholic and Low Alcohol Beer Brewing–A Review. J. Am. Soc. Brew. Chem. 2019, 77, 77–91. [Google Scholar] [CrossRef]
- Blanco, C.A.; Andrés-Iglesias, C.; Montero, O. Low-Alcohol Beers: Flavor Compounds, Defects, and Improvement Strategies. Crit. Rev. Food Sci. Nutr. 2016, 56, 1379–1388. [Google Scholar] [CrossRef]
- Basso, R.F.; Alcarde, A.R.; Portugal, C.B. Could Non-Saccharomyces Yeasts Contribute on Innovative Brewing Fermentations? Food Res. Int. 2016, 86, 112–120. [Google Scholar] [CrossRef]
- Holt, S.; Mukherjee, V.; Lievens, B.; Verstrepen, K.J.; Thevelein, J.M. Bioflavoring by Non-Conventional Yeasts in Sequential Beer Fermentations. Food Microbiol. 2018, 72, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Johansson, L.; Nikulin, J.; Juvonen, R.; Krogerus, K.; Magalhães, F.; Mikkelson, A.; Nuppunen-Puputti, M.; Sohlberg, E.; de Francesco, G.; Perretti, G. Sourdough Cultures as Reservoirs of Maltose-Negative Yeasts for Low-Alcohol Beer Brewing. Food Microbiol. 2021, 94, 103629. [Google Scholar] [CrossRef] [PubMed]
- Testa, B.; Coppola, F.; Letizia, F.; Albanese, G.; Karaulli, J.; Ruci, M.; Pistillo, M.; Germinara, G.S.; Messia, M.C.; Succi, M. Versatility of Saccharomyces cerevisiae 41CM in the Brewery Sector: Use as a Starter for “Ale” and “Lager” Craft Beer Production. Processes 2022, 10, 2495. [Google Scholar] [CrossRef]
- Sánchez-Adriá, I.E.; Sanmartín, G.; Prieto, J.A.; Estruch, F.; Randez-Gil, F. Sourdough Yeast Strains Exhibit Thermal Tolerance, High Fermentative Performance, and a Distinctive Aromatic Profile in Beer Wort. Foods 2024, 13, 1059. [Google Scholar] [CrossRef]
- Postigo, V.; García, M.; Cabellos, J.M.; Arroyo, T. Wine Saccharomyces Yeasts for Beer Fermentation. Fermentation 2021, 7, 290. [Google Scholar] [CrossRef]
- Matraxia, M.; Alfonzo, A.; Prestianni, R.; Francesca, N.; Gaglio, R.; Todaro, A.; Alfeo, V.; Perretti, G.; Columba, P.; Settanni, L. Non-Conventional Yeasts from Fermented Honey by-Products: Focus on Hanseniaspora uvarum Strains for Craft Beer Production. Food Microbiol. 2021, 99, 103806. [Google Scholar] [CrossRef]
- Rossi, S.; Turchetti, B.; Sileoni, V.; Marconi, O.; Perretti, G. Evaluation of Saccharomyces cerevisiae Strains Isolated from Non-brewing Environments in Beer Production. J. Inst. Brew. 2018, 124, 381–388. [Google Scholar] [CrossRef]
- Cubillos, F.A.; Gibson, B.; Grijalva-Vallejos, N.; Krogerus, K.; Nikulin, J. Bioprospecting for Brewers: Exploiting Natural Diversity for Naturally Diverse Beers. Yeast 2019, 36, 383–398. [Google Scholar] [CrossRef] [PubMed]
- Canonico, L.; Zannini, E.; Ciani, M.; Comitini, F. Assessment of Non-Conventional Yeasts with Potential Probiotic for Protein-Fortified Craft Beer Production. Lwt 2021, 145, 111361. [Google Scholar] [CrossRef]
- Drosou, F.; Mamma, D.; Oreopoulou, V.; Tataridis, P.; Dourtoglou, V. Metschnikowia pulcherrima: A New Yeast for Brewing. 2023. [Google Scholar]
- Canonico, L.; Agarbati, A.; Galli, E.; Comitini, F.; Ciani, M. Metschnikowia pulcherrima as Biocontrol Agent and Wine Aroma Enhancer in Combination with a Native Saccharomyces cerevisiae. LWT 2023, 181, 114758. [Google Scholar] [CrossRef]
- Puyo, M.; Simonin, S.; Bach, B.; Klein, G.; Alexandre, H.; Tourdot-Maréchal, R. Bio-Protection in Oenology by Metschnikowia pulcherrima: From Field Results to Scientific Inquiry. Front. Microbiol. 2023, 14, 1252973. [Google Scholar] [CrossRef] [PubMed]
- Hranilovic, A.; Gambetta, J.M.; Jeffery, D.W.; Grbin, P.R.; Jiranek, V. Lower-Alcohol Wines Produced by Metschnikowia pulcherrima and Saccharomyces cerevisiae Co-Fermentations: The Effect of Sequential Inoculation Timing. Int. J. Food Microbiol. 2020, 329, 108651. [Google Scholar] [CrossRef] [PubMed]
- Testa, B.; Coppola, F.; Iorizzo, M.; Di Renzo, M.; Coppola, R.; Succi, M. Preliminary Characterisation of Metschnikowia pulcherrima to Be Used as a Starter Culture in Red Winemaking. Beverages 2024, 10, 88. [Google Scholar] [CrossRef]
- Drosou, F.; Mamma, D.; Tataridis, P.; Dourtoglou, V.; Oreopoulou, V. Metschnikowia pulcherrima in Mono or Co-Fermentations in Brewing. 2022; 75, 69–76. [Google Scholar]
- Postigo, V.; Sánchez, A.; Cabellos, J.M.; Arroyo, T. New Approaches for the Fermentation of Beer: Non-Saccharomyces Yeasts from Wine. Fermentation 2022, 8, 280. [Google Scholar] [CrossRef]
- Einfalt, D. Barley-Sorghum Craft Beer Production with Saccharomyces cerevisiae, Torulaspora delbrueckii and Metschnikowia pulcherrima Yeast Strains. Eur. Food Res. Technol. 2021, 247, 385–393. [Google Scholar] [CrossRef]
- Hlangwani, E.; du Plessis, H.W.; Dlamini, B.C. The Effect of Selected Non-Saccharomyces Yeasts and Cold-Contact Fermentation on the Production of Low-Alcohol Marula Fruit Beer. Heliyon 2024, 10, e24505. [Google Scholar] [CrossRef]
- Karaulli, J.; Xhaferaj, N.; Ruci, M.; Testa, B.; Letizia, F.; Albanese, G.; Kongoli, R.; Lamçe, F.; Kyçyk, O.; Sulaj, K.; et al. Evaluation Of Saccharomyces and Non-Saccharomyces Yeasts Isolated from Albanian Autochthonous Grape Varieties for Craft Beer Production. In Proceedings of the Proceedings of V International Agricultural, Biological, Life Science Conference (AGBIOL); pp. 2023654–665.
- Barbosa, C.; Lage, P.; Esteves, M.; Chambel, L.; Mendes-Faia, A.; Mendes-Ferreira, A. Molecular and Phenotypic Characterization of Metschnikowia pulcherrima Strains from Douro Wine Region. Fermentation 2018, 4, 8. [Google Scholar] [CrossRef]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected Non-Saccharomyces Wine Yeasts in Controlled Multistarter Fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Mažeika, K.; Šiliauskas, L.; Skridlaitė, G.; Matelis, A.; Garjonytė, R.; Paškevičius, A.; Melvydas, V. Features of Iron Accumulation at High Concentration in Pulcherrimin-Producing Metschnikowia Yeast Biomass. JBIC J. Biol. Inorg. Chem. 2021, 26, 299–311. [Google Scholar] [CrossRef] [PubMed]
- Gut, A.M.; Vasiljevic, T.; Yeager, T.; Donkor, O.N. Characterization of Yeasts Isolated from Traditional Kefir Grains for Potential Probiotic Properties. J. Funct. Foods 2019, 58, 56–66. [Google Scholar] [CrossRef]
- Testa, B.; Lombardi, S.J.; Iorizzo, M.; Letizia, F.; Di Martino, C.; Di Renzo, M.; Strollo, D.; Tremonte, P.; Pannella, G.; Ianiro, M. Use of Strain Hanseniaspora Guilliermondii BF1 for Winemaking Process of White Grapes Vitis Vinifera Cv Fiano. Eur. Food Res. Technol. 2020, 246, 549–561. [Google Scholar] [CrossRef]
- Belda, I.; Ruiz, J.; Navascués, E.; Marquina, D.; Santos, A. Improvement of Aromatic Thiol Release through the Selection of Yeasts with Increased β-Lyase Activity. Int. J. Food Microbiol. 2016, 225, 1–8. [Google Scholar] [CrossRef]
- López-Enríquez, L.; Vila-Crespo, J.; Rodríguez-Nogales, J.M.; Fernández-Fernández, E.; Ruipérez, V. Screening and Enzymatic Evaluation of Saccharomyces cerevisiae Populations from Spontaneous Fermentation of Organic Verdejo Wines. Foods 2022, 11, 3448. [Google Scholar] [CrossRef] [PubMed]
- Strong, G.; England, K. Beer Judge Certification Program: 2015 Style Guidelines. Brew Assoc 2015, 47. [Google Scholar]
- European Brewery, Convention. Analytica - EBC; Hans Carl, Fachverlag: Nürnberg, 2007; ISBN 3-418-00759-7. [Google Scholar]
- Paszkot, J.; Gasiński, A.; Kawa-Rygielska, J. Evaluation of Volatile Compound Profiles and Sensory Properties of Dark and Pale Beers Fermented by Different Strains of Brewing Yeast. Sci. Rep. 2023, 13, 6725. [Google Scholar] [CrossRef] [PubMed]
- Kucharczyk, K.; Tuszyński, T. The Effect of Temperature on Fermentation and Beer Volatiles at an Industrial Scale. J. Inst. Brew. 2018, 124, 230–235. [Google Scholar] [CrossRef]
- Du, Q.; Ye, D.; Zang, X.; Nan, H.; Liu, Y. Effect of Low Temperature on the Shaping of Yeast-Derived Metabolite Compositions during Wine Fermentation. Food Res. Int. 2022, 162, 112016. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Tan, F.; Chu, R.; Li, G.; Li, L.; Yang, T.; Zhang, M. The Effect of Non-Saccharomyces Yeasts on Biogenic Amines in Wine. Trends Food Sci. Technol. 2021, 116, 1029–1040. [Google Scholar] [CrossRef]
- Nalazek-Rudnicka, K.; Wojnowski, W.; Wasik, A. Occurrence and Levels of Biogenic Amines in Beers Produced by Different Methods. Foods 2021, 10, 2902. [Google Scholar] [CrossRef] [PubMed]
- Matukas, M.; Starkute, V.; Zokaityte, E.; Zokaityte, G.; Klupsaite, D.; Mockus, E.; Rocha, J.M.; Ruibys, R.; Bartkiene, E. Effect of Different Yeast Strains on Biogenic Amines, Volatile Compounds and Sensory Profile of Beer. Foods 2022, 11, 2317. [Google Scholar] [CrossRef] [PubMed]
- Tsafack, P.B.; Tsopmo, A. Effects of Bioactive Molecules on the Concentration of Biogenic Amines in Foods and Biological Systems. Heliyon 2022, 8. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, H.; de Almeida, J.M.; Matias, A.; Saraiva, C.; Jorge, P.A.; Coelho, L.C. Detection of Biogenic Amines in Several Foods with Different Sample Treatments: An Overview. Trends Food Sci. Technol. 2021, 113, 86–96. [Google Scholar] [CrossRef]
- EFSA Panel on Biological Hazards, (BIOHAZ). Scientific Opinion on Risk Based Control of Biogenic Amine Formation in Fermented Foods. Efsa J. 2011, 9, 2393. [Google Scholar] [CrossRef]
- Wang, C.; Liu, M.; Li, Y.; Zhang, Y.; Yao, M.; Qin, Y.; Liu, Y. Hydrogen Sulfide Synthesis in Native Saccharomyces cerevisiae Strains during Alcoholic Fermentations. Food Microbiol. 2018, 70, 206–213. [Google Scholar] [CrossRef]
- Pawlikowska, E.; Kolesińska, B.; Nowacka, M.; Kregiel, D. A New Approach to Producing High Yields of Pulcherrimin from Metschnikowia Yeasts. Fermentation 2020, 6, 114. [Google Scholar] [CrossRef]
- Oro, L.; Ciani, M.; Comitini, F. Antimicrobial Activity of Metschnikowia pulcherrima on Wine Yeasts. J. Appl. Microbiol. 2014, 116, 1209–1217. [Google Scholar] [CrossRef] [PubMed]
- Morata, A.; Loira, I.; Escott, C.; del Fresno, J.M.; Bañuelos, M.A.; Suárez-Lepe, J.A. Applications of Metschnikowia pulcherrima in Wine Biotechnology. Fermentation 2019, 5, 63. [Google Scholar] [CrossRef]
- Nardi, T. Microbial Resources as a Tool for Enhancing Sustainability in Winemaking. Microorganisms 2020, 8, 507. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Yong, X.; Zhao, T.; Li, Y.; Liu, J. Research Progress of the Biosynthesis of Natural Bio-Antibacterial Agent Pulcherriminic Acid in Bacillus. Molecules 2020, 25, 5611. [Google Scholar] [CrossRef] [PubMed]
- Benoit, I.; Danchin, E.G.; Bleichrodt, R.-J.; de Vries, R.P. Biotechnological Applications and Potential of Fungal Feruloyl Esterases Based on Prevalence, Classification and Biochemical Diversity. Biotechnol. Lett. 2008, 30, 387–396. [Google Scholar] [CrossRef]
- Dank, A.; Smid, E.J.; Notebaart, R.A. CRISPR-Cas Genome Engineering of Esterase Activity in Saccharomyces cerevisiae Steers Aroma Formation. BMC Res. Notes 2018, 11, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Qin, Q.; Li, C.; Zhao, X.; Song, F.; An, M.; Chen, Y.; Wang, X.; Huang, W.; Zhan, J. Application of Non-Saccharomyces Yeasts with High β-Glucosidase Activity to Enhance Terpene-Related Floral Flavor in Craft Beer. Food Chem. 2023, 404, 134726. [Google Scholar] [CrossRef]
- He, Y.; Dong, J.; Yin, H.; Zhao, Y.; Chen, R.; Wan, X.; Chen, P.; Hou, X.; Liu, J.; Chen, L. Wort Composition and Its Impact on the Flavour-active Higher Alcohol and Ester Formation of Beer–a Review. J. Inst. Brew. 2014, 120, 157–163. [Google Scholar] [CrossRef]
- Michel, M.; Haslbeck, K.; Ampenberger, F.; Meier-Dörnberg, T.; Stretz, D.; Hutzler, M.; Coelhan, M.; Jacob, F.; Liu, Y. Screening of Brewing Yeast β-Lyase Activity and Release of Hop Volatile Thiols from Precursors during Fermentation. 2019. [Google Scholar]
- Krogerus, K.; Rettberg, N.; Gibson, B. Increased Volatile Thiol Release during Beer Fermentation Using Constructed Interspecies Yeast Hybrids. Eur. Food Res. Technol. 2023, 249, 55–69. [Google Scholar] [CrossRef]
- Bonnaffoux, H.; Roland, A.; Schneider, R.; Cavelier, F. Spotlight on Release Mechanisms of Volatile Thiols in Beverages. Food Chem. 2021, 339, 127628. [Google Scholar] [CrossRef] [PubMed]
- Buiatti, S.; Tat, L.; Natolino, A.; Passaghe, P. Biotransformations Performed by Yeasts on Aromatic Compounds Provided by Hop—A Review. Fermentation 2023, 9, 327. [Google Scholar] [CrossRef]
- Muñoz-Redondo, J.M.; Puertas, B.; Cantos-Villar, E.; Jiménez-Hierro, M.J.; Carbú, M.; Garrido, C.; Ruiz-Moreno, M.J.; Moreno-Rojas, J.M. Impact of Sequential Inoculation with the Non-Saccharomyces T. delbrueckii and M. pulcherrima Combined with Saccharomyces cerevisiae Strains on Chemicals and Sensory Profile of Rosé Wines. J. Agric. Food Chem. 2021, 69, 1598–1609. [Google Scholar] [CrossRef] [PubMed]
- Ciani, M.; Capece, A.; Comitini, F.; Canonico, L.; Siesto, G.; Romano, P. Yeast Interactions in Inoculated Wine Fermentation. Front. Microbiol. 2016, 7, 555. [Google Scholar] [CrossRef] [PubMed]
- Mencher, A.; Morales, P.; Curiel, J.A.; Gonzalez, R.; Tronchoni, J. Metschnikowia Pulcherrima Represses Aerobic Respiration in Saccharomyces cerevisiae Suggesting a Direct Response to Co-Cultivation. Food Microbiol. 2021, 94, 103670. [Google Scholar] [CrossRef] [PubMed]
- Ivit, N.N.; Longo, R.; Kemp, B. The Effect of Non-Saccharomyces and Saccharomyces Non-cerevisiae Yeasts on Ethanol and Glycerol Levels in Wine. Fermentation 2020, 6, 77. [Google Scholar] [CrossRef]
- Sadoudi, M.; Rousseaux, S.; David, V.; Alexandre, H.; Tourdot-Maréchal, R. Metschnikowia pulcherrima Influences the Expression of Genes Involved in PDH Bypass and Glyceropyruvic Fermentation in Saccharomyces cerevisiae. Front. Microbiol. 2017, 8, 1137. [Google Scholar] [CrossRef] [PubMed]
- Schreurs, M.; Piampongsant, S.; Roncoroni, M.; Cool, L.; Herrera-Malaver, B.; Vanderaa, C.; Theßeling, F.A.; Kreft, Ł.; Botzki, A.; Malcorps, P. Predicting and Improving Complex Beer Flavor through Machine Learning. Nat. Commun. 2024, 15, 2368. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Procopio, S.; Becker, T. Flavor Impacts of Glycerol in the Processing of Yeast Fermented Beverages: A Review. J. Food Sci. Technol. 2015, 52, 7588–7598. [Google Scholar] [CrossRef]
- Du Plessis, H.; Du Toit, M.; Hoff, J.; Hart, R.; Ndimba, B.; Jolly, N. Characterisation of Non-Saccharomyces Yeasts Using Different Methodologies and Evaluation of Their Compatibility with Malolactic Fermentation. South Afr. J. Enol. Vitic. 2017, 38, 46–63. [Google Scholar] [CrossRef]
- Benito, S.; Hofmann, T.; Laier, M.; Lochbühler, B.; Schüttler, A.; Ebert, K.; Fritsch, S.; Röcker, J.; Rauhut, D. Effect on Quality and Composition of Riesling Wines Fermented by Sequential Inoculation with Non-Saccharomyces and Saccharomyces cerevisiae. Eur. Food Res. Technol. 2015, 241, 707–717. [Google Scholar] [CrossRef]
- Michel, M.; Meier-Dörnberg, T.; Jacob, F.; Methner, F.; Wagner, R.S.; Hutzler, M. Pure non-Saccharomyces Starter Cultures for Beer Fermentation with a Focus on Secondary Metabolites and Practical Applications. J. Inst. Brew. 2016, 122, 569–587. [Google Scholar] [CrossRef]
- Engan, S. Organoleptic Threshold Values of Some Organic Acids in Beer. J. Inst. Brew. 1974, 80, 162–163. [Google Scholar] [CrossRef]
- Olaniran, A.O.; Hiralal, L.; Mokoena, M.P.; Pillay, B. Flavour-active Volatile Compounds in Beer: Production, Regulation and Control. J. Inst. Brew. 2017, 123, 13–23. [Google Scholar] [CrossRef]
- Stewart, G.G. The Production of Secondary Metabolites with Flavour Potential during Brewing and Distilling Wort Fermentations. Fermentation 2017, 3, 63. [Google Scholar] [CrossRef]
- Gernat, D.; Brouwer, E.; Ottens, M. Aldehydes as Wort Off-Flavours in Alcohol-Free Beers—Origin and Control. Food Bioprocess Technol. 2020, 13, 195–216. [Google Scholar] [CrossRef]
- Edgar, M.; Wycliffe, A. Investigating the Impact of Acetaldehyde Accumulation on Beer Quality: Metabolic Pathways, Yeast Health, and Mitigation Strategies. 2024; 11, 40–51. [Google Scholar]
- Comitini, F.; Agarbati, A.; Canonico, L.; Ciani, M. Yeast Interactions and Molecular Mechanisms in Wine Fermentation: A Comprehensive Review. Int. J. Mol. Sci. 2021, 22, 7754. [Google Scholar] [CrossRef] [PubMed]
- Pires, E.J.; Teixeira, J.A.; Brányik, T.; Vicente, A.A. Yeast: The Soul of Beer’s Aroma—A Review of Flavour-Active Esters and Higher Alcohols Produced by the Brewing Yeast. Appl. Microbiol. Biotechnol. 2014, 98, 1937–1949. [Google Scholar] [CrossRef] [PubMed]
- Holt, S.; Miks, M.H.; de Carvalho, B.T.; Foulquie-Moreno, M.R.; Thevelein, J.M. The Molecular Biology of Fruity and Floral Aromas in Beer and Other Alcoholic Beverages. FEMS Microbiol. Rev. 2019, 43, 193–222. [Google Scholar] [CrossRef]
- Roberts, R.; Khomenko, I.; Eyres, G.T.; Bremer, P.; Silcock, P.; Betta, E.; Biasioli, F. Online Monitoring of Higher Alcohols and Esters throughout Beer Fermentation by Commercial Saccharomyces cerevisiae and Saccharomyces pastorianus Yeast. J. Mass Spectrom. 2023, 58, e4959. [Google Scholar] [CrossRef] [PubMed]
- Postigo, V.; Sanz, P.; García, M.; Arroyo, T. Impact of Non-Saccharomyces Wine Yeast Strains on Improving Healthy Characteristics and the Sensory Profile of Beer in Sequential Fermentation. Foods 2022, 11, 2029. [Google Scholar] [CrossRef] [PubMed]
- Verstrepen, K.J.; Derdelinckx, G.; Dufour, J.-P.; Winderickx, J.; Thevelein, J.M.; Pretorius, I.S.; Delvaux, F.R. Flavor-Active Esters: Adding Fruitiness to Beer. J. Biosci. Bioeng. 2003, 96, 110–118. [Google Scholar] [CrossRef]
- Viejo, C.G.; Fuentes, S.; Torrico, D.D.; Godbole, A.; Dunshea, F.R. Chemical Characterization of Aromas in Beer and Their Effect on Consumers Liking. Food Chem. 2019, 293, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Brányik, T.; Vicente, A.A.; Dostálek, P.; Teixeira, J.A. A Review of Flavour Formation in Continuous Beer Fermentations. J. Inst. Brew. 2008, 114, 3–13. [Google Scholar] [CrossRef]
- Ocvirk, M.; Mlinarič, N.K.; Košir, I.J. Comparison of Sensory and Chemical Evaluation of Lager Beer Aroma by Gas Chromatography and Gas Chromatography/Mass Spectrometry. J. Sci. Food Agric. 2018, 98, 3627–3635. [Google Scholar] [CrossRef] [PubMed]
- Meilgaard, M.C. Prediction of Flavor Differences between Beers from Their Chemical Composition. J. Agric. Food Chem. 1982, 30, 1009–1017. [Google Scholar] [CrossRef]
- Siebert, K.J. Modeling the Flavor Thresholds of Organic Acids in Beer as a Function of Their Molecular Properties. Food Qual. Prefer. 1999, 10, 129–137. [Google Scholar] [CrossRef]
- Bravi, E.; Benedetti, P.; Marconi, O.; Perretti, G. Determination of Free Fatty Acids in Beer Wort. Food Chem. 2014, 151, 374–378. [Google Scholar] [CrossRef] [PubMed]
- Wilde, P.; Husband, F.; Cooper, D.; Ridout, M.; Muller, R.; Mills, E. Destabilization of Beer Foam by Lipids: Structural and Interfacial Effects. J. Am. Soc. Brew. Chem. 2003, 61, 196–202. [Google Scholar] [CrossRef]
- Gordon, R.; Power, A.; Chapman, J.; Chandra, S.; Cozzolino, D. A Review on the Source of Lipids and Their Interactions during Beer Fermentation That Affect Beer Quality. Fermentation 2018, 4, 89. [Google Scholar] [CrossRef]
- Olšovská, J.; Vrzal, T.; Štěrba, K.; Slabý, M.; Kubizniaková, P.; Čejka, P. The Chemical Profiling of Fatty Acids during the Brewing Process. J. Sci. Food Agric. 2019, 99, 1772–1779. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Heatmap visualization for test results of carbohydrates assimilation and enzymatic activities of M. pulcherrima strains. Red square: negative; blue square: positive.
Figure 1.
Heatmap visualization for test results of carbohydrates assimilation and enzymatic activities of M. pulcherrima strains. Red square: negative; blue square: positive.
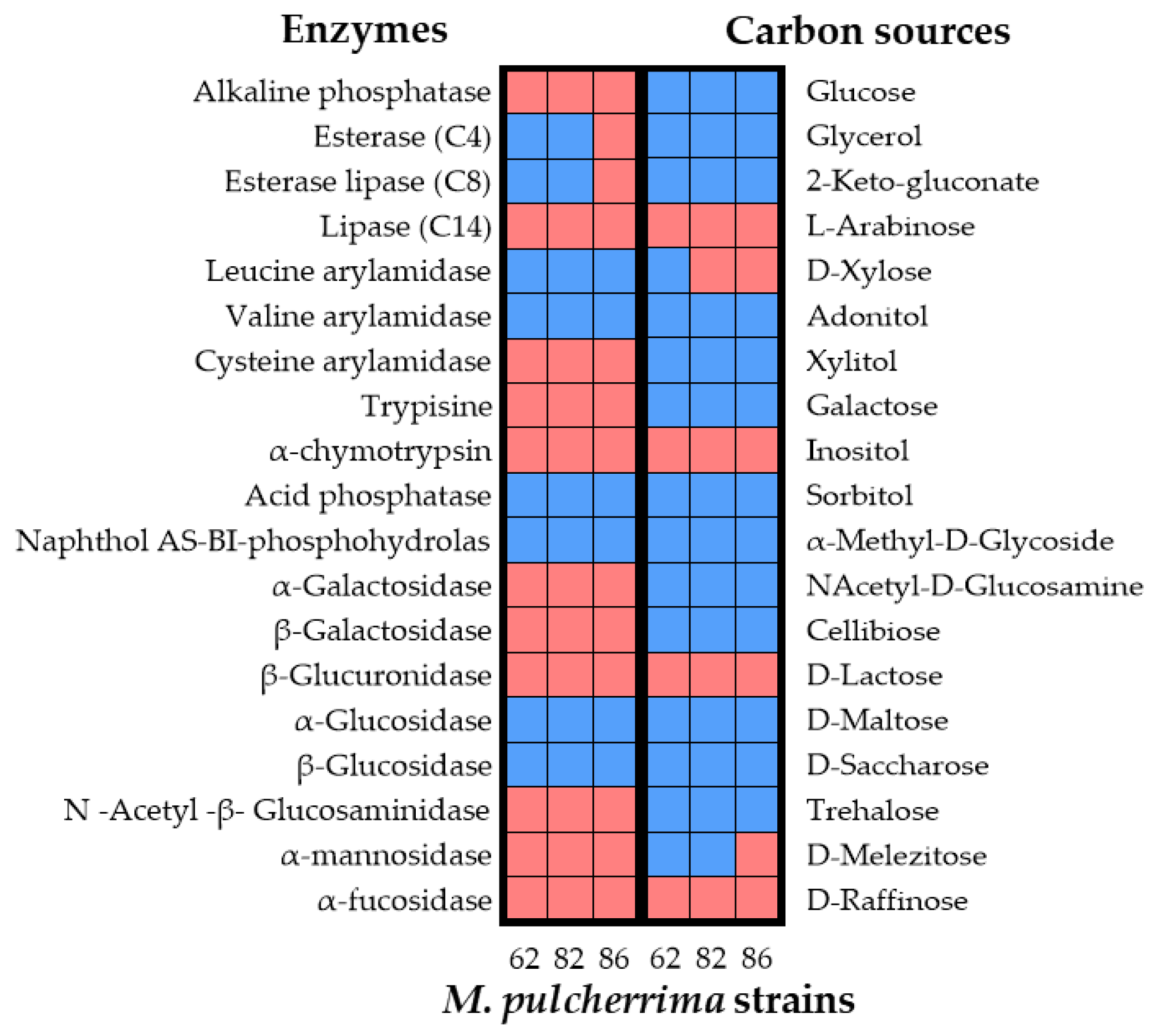
Figure 2.
pH and density (g/cm3) trends during alcoholic fermentation. Test A, inoculated with M. pulcherrima 62 and S. cerevisiae S04 in sequential inoculum; Test B inoculated with S. cerevisiae as a single starter.
Figure 2.
pH and density (g/cm3) trends during alcoholic fermentation. Test A, inoculated with M. pulcherrima 62 and S. cerevisiae S04 in sequential inoculum; Test B inoculated with S. cerevisiae as a single starter.
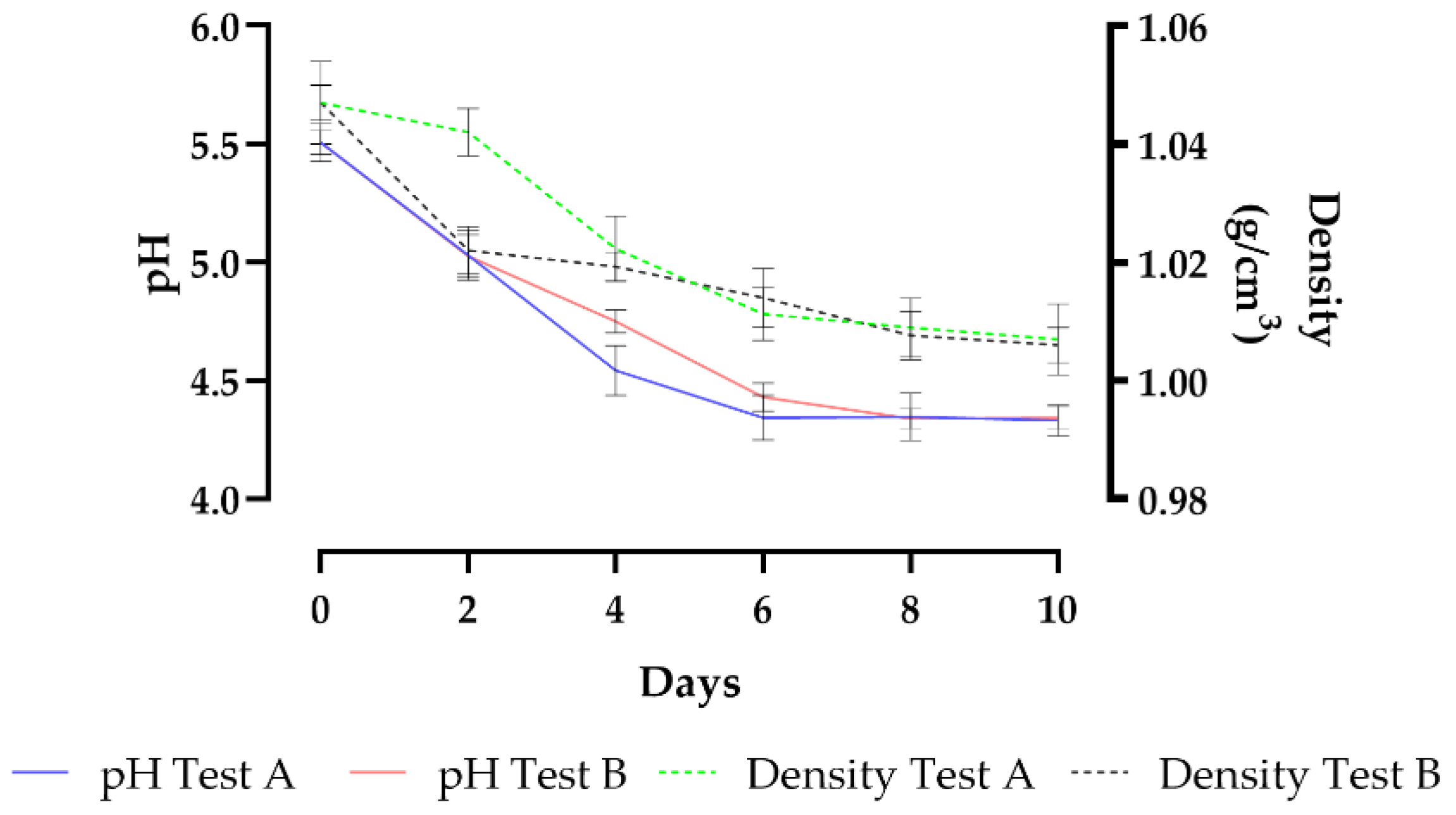
Figure 3.
Colonies morphology of S. cerevisiae (creamy white) and M. pulcherrima (light blue) using WL agar medium.
Figure 3.
Colonies morphology of S. cerevisiae (creamy white) and M. pulcherrima (light blue) using WL agar medium.
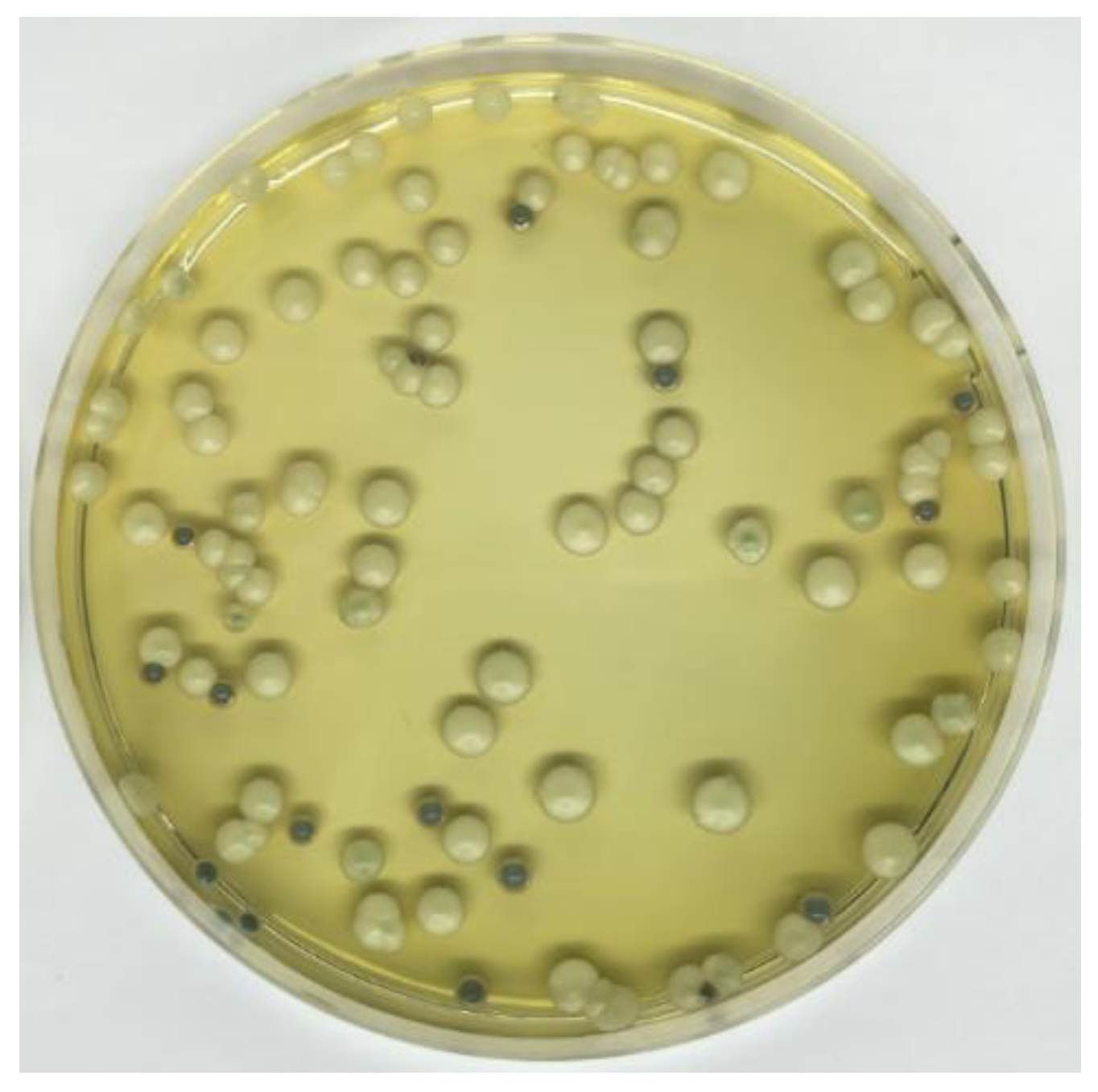

Table 1.
H2S and pulcherrimin production, cryotolerance and enzymatic activities of M. pulcherrima strains.
Table 1.
H2S and pulcherrimin production, cryotolerance and enzymatic activities of M. pulcherrima strains.
|
M. pulcherrima strains |
H2S* | β -Glucosidase** | β -Lyase** | Protease** | Pulcherrimin** | Cryotolerance** |
| 62 | 2 | + | + | + | + | + |
| 82 | 2 | weak | + | - | + | weak |
| 86 | 2 | weak | + | + | + | weak |
*H2S production: (1), white colour - no production; (2) light brown; (3) brown; (4) dark brown; (5) dark brown/black – high production. **Enzymatic activities: (β-glucosidase, β-lyase, protease), pulcherrimin production and cryotolerance (+ positive; - negative).
Table 2.
Main physical-chemical parameters of beer produced using M. pulcherrima 62 and S. cerevisiae S04 in sequential inoculation (Test A) and of beer produced using S. cerevisiae S04 as a single starter (Test B).
Table 2.
Main physical-chemical parameters of beer produced using M. pulcherrima 62 and S. cerevisiae S04 in sequential inoculation (Test A) and of beer produced using S. cerevisiae S04 as a single starter (Test B).
| physical-chemical parameters | Test A | Test B |
| pH | 4.35 ± 0.05 a | 4.31 ± 0.06 a |
| Alcohol (% v/v) | 5.20 ± 0.10 a | 5.00 ± 0.10 a |
| Acetic acid (mg/L) | 60.66 ± 3.51 a | 58.83 ± 1.25 a |
| L-malic acid (mg/L) | 160.33 ± 2.51 b | 202.33 ± 7.50 a |
| L-lactic acid (mg/L) | 115.66 ± 5.13 a | 115.50 ± 2.29 a |
| Glycerol (mg/L) | 1026.02 ± 28.21 a | 892.66 ± 9.60 b |
| Acetaldehyde (mg/L) | 7.66 ± 0.80 a | 7.70 ± 0.19 a |
All values are expressed as mean ± standard deviation (n = 3). Different superscript letters in each row indicate significant differences (p < 0.05).
Table 3.
Viability evolution (log CFU/mL) of S. cerevisiae and M. pulcherrima during alcoholic fermentation: Test A, inoculated with M. pulcherrima 62 and S. cerevisiae S04 in sequential inoculum; Test B inoculated with S. cerevisiae as a single starter.
Table 3.
Viability evolution (log CFU/mL) of S. cerevisiae and M. pulcherrima during alcoholic fermentation: Test A, inoculated with M. pulcherrima 62 and S. cerevisiae S04 in sequential inoculum; Test B inoculated with S. cerevisiae as a single starter.
| Fermentation time (days) | |||||||
| 0 | 2 | 4 | 6 | 8 | 10 | ||
| Test A | M. pulcherrima | 6.12 ± 0.13 b | 6.93 ± 0.25 a | 6.33 ± 0.15 b | 4.02 ± 0.14 c | 0.00 ±0.00 d | 0.00 ± 0.00 d |
| S. cerevisiae | 0.00 ±.0.00 d | 6.75 ± 0.25 c | 7.54 ± 0.23 b | 8.27 ± 0.30 a | 8.85 ± 0.15 a | 7.62 ± 0.20 b | |
| Test B | S. cerevisiae | 6.83 ± 0.15 c | 6.88 ± 0.20 c | 7.86 ± 0.35 b | 8.52 ± 0.33 a | 7.88 ± 0.10 a | 7.73 ± 0.15 b |
All values are expressed as mean ± standard deviation (n = 3). Different superscript letters in each row indicate significant differences (p < 0.05).
Table 4.
Volatile compounds (mg/L) detected by GC-FID in the beer produced using M. pulcherrima 62 and S. cerevisiae S04 in sequential inoculation (Test A) and in beer produced using S. cerevisiae S04 as a single starter (Test B).
Table 4.
Volatile compounds (mg/L) detected by GC-FID in the beer produced using M. pulcherrima 62 and S. cerevisiae S04 in sequential inoculation (Test A) and in beer produced using S. cerevisiae S04 as a single starter (Test B).
| Class of organic compounds | Volatile compounds | Test A | Test B |
| Higher alcohols | Isobutanol | 42.26 ± 0.75 a | 11.44 ± 0.55 b |
| Isoamyl alcohol | 101.83 ± 1.55 a | 53.02 ± 2.01 b | |
| 1-Hexanol | nd | 1.23 ± 0.10 a | |
| Esters | β-phenylethanol | 21.10 ± 1.86 b | 34.77 ±1.71a |
| Ethyl acetate | 1.31 ± 0.18 a | 0.68 ± 0.04 b | |
| Ethyl isovalerate | 0.24 ± 0.06 a | 0.17 ± 0.03 a | |
| Ethyl butirate | 0.24 ± 0.05 a | 0.34 ± 0.03 a | |
| Ethyl lactate | nd | 0.56 ± 0.06 a | |
| Isoamyl acetate | 2.50 ± 0.14 a | 1.88 ± 0.04 b | |
| Ethyl hexanoate | 0.03 ± 0.01 a | 0.05 ± 0.01 a | |
| Ethyl octanoate | 0.13 ± 0.02 b | 0.24 ± 0.01 a | |
| Diethyl succinate | 0.34 ± 0.04 a | 0.22 ± 0.02 b | |
| Fatty acids | Butyric acid | 1.80 ± 0.08 a | 0.11 ± 0.02 b |
| Hexanoic acid | 2.30 ± 0.12 a | 0.65 ± 0.05 b | |
| Octanoic acid | 1.13 ± 0.10 b | 1.94 ± 0.05 a | |
| Decanoic acid | 0.47 ± 0.08 a | 0.30 ± 0.04 a | |
| Aldehydes/Ketones | Diacetyl | 0.11 ± 0.04 a | 0.12 ± 0.03 a |
| Acetoin | 0.84 ± 0.05 a | 0.64 ± 0.08 b |
All values are expressed as mean ± standard deviation (n = 3). Different superscript letters in each row indicate significant differences (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



