
Submitted:
13 July 2023
Posted:
14 July 2023
You are already at the latest version
Abstract
The increasing demand for craft beer drives the search for novel ale yeast cultures from brew-ing-related wild environments. The focus of bioprospecting for craft cultures is to identify feral yeasts suitable to imprint unique sensorial attributes to the final product. Here, we integrated phylogenetic, genotypic, genetic, and metabolomic techniques to demonstrate that sour beer during aging in wooden barrel is a source of suitable craft ale yeast candidates. Differently from traditional lambic beer maturation phase, during aging of sour-matured production style beer, different biotypes of Saccharomyces cerevisiae dominated the cultivable in-house mycobiota, fol-lowed by Pichia membranifaciens, Brettanomyces bruxellensis, and Brettanomyces anomalus. In addi-tion, three putative S. cerevisiae x Saccharomyces uvarum hybrids were identified. S. cerevisiae feral strains sporulated, produced viable monosporic progenies, and have STA1 gene as well as the full-length promoter. During hopped wort fermentation four S. cerevisiae strains and the S. cere-visiae x S. uvarum hybrid WY213 overcame non-Saccharomyces strains in fermentative rate and ethanol production, except for P. membranifaciens WY122. This strain consumed maltose after a long lag phase, in contrast with the phenotypic profile descripted for the species. According to STA1+ genotype S. cerevisiae partially consumed dextrin, but no 4-vynil guaiacol was detected among VOCs, suggesting that these strains were phenolic off flavor-negative (POF-). In conclu-sion, the strains characterized here have relevant brewing properties and are exploitable as in-digenous craft beer starters.
Keywords:
sour beer
; craft brewing
; STA1 gene
; dextrin
; hybrid
; Saccharomyces cerevisiae
; Saccharomyces uvarum
; Pichia membranifaciens
1. Introduction
Craft beer production has gained increasing interest in many countries as an alternative to mainstream beer [1]. Craft brewing refers to relatively small breweries, that aim to produce beers with unique and original sensory characteristics based on quality and diversity [2]. Off-flavors in craft beers increase their perceived quality compared to commercial beer [3,4].
The sensory profile of beer is determined by several factors (choice of malts or adjuncts, hop varieties, water quality, etc) including yeasts, that catalyze metabolic reactions during wort fermentation which impact beer flavor profile. Other than for sugar consumption and ethanol and CO2 production, yeasts are responsible for releasing aroma-active secondary side-products of alcoholic fermentation that contribute to solvent-like and fruity notes and a sweet taste, such as higher alcohols, ethyl esters, and acetate esters [5]. They also can transform neutral precursors present in wort and hop such as glycosides, polyfunctional thiols, and terpenes, into sensorial active compounds, as reviewed by X et al. [6]. This bio-flavoring aptitude qualitatively and quantitatively varies depending on strain and species employed [7]. However, the pool of commercial Saccharomyces cerevisiae for ale beer and Saccharomyces pastorianus for lager beer is restricted and genetically redundant [8], hindering the chance to diversify beer sensorial profile through yeast bio-flavoring.
To expand the number of craft brewing starters and to diversify the portfolio of beer styles, various OGM-free strategies have been proposed. These approaches include hybridization, utilization of non-S. cerevisiae strains, and the bioprospecting for novel wild yeasts species and strains from alternative or unusual niches compared to ale and lager brewing [9-11 Krogerus et al., 2017; Cubillos et al., 2019; Iattici et al., 2020]. Recently, genetically modified yeasts were also exploited to boost hop-related flavor active molecules [12-15]. The selection of suitable yeast strains is a crucial step in producing high-quality craft beer affecting both wort fermentation and beer conditioning. Yeast strains have a significant impact on the production of volatile organic compounds (VOCs), which are essential for the beer's flavor and aroma [2, 3], and this is particularly important for craft beers, which are often unfiltered, unpasteurized, and bottle-conditioned [4, 5].
In craft brewing, the interest towards sour beer style is recently growing [16]. Traditionally fermented sour beers, such as Belgian type lambic beers, pass through spontaneous fermentation and wood maturation, which can last up to three years. This process involves conventional and unconventional yeasts, lactic acid bacteria, and, to a lesser extent, acetic acid bacteria [16-20]. These microorganisms are spontaneous beer contaminants arisen from raw materials, air, and brewery environment and equipment, such as the inner surface of barrel [21]. Recently, they have been revised as "drivers" of innovation and diversification of the sensory profiles in sour beer [22]. Other craft brewing strategies mimic Lambic-style beer in complexity, fruity, and sourness, but reduce maturation time as well as increase fermentation control (as reviewed by [16,23]). Among these methods, ale beer can be soured by a primary fermentation with brewery yeast pure cultures, followed by a maturation aging in wooden barrels, where the implantation of a complex house microbiota can lead to acidic and peculiar sensory profiles (the so-called sour-matured production styles).
In this study, we considered in-house microbiota of sour ale beers during ageing as a source for novel brewing cultures. For this purpose, we taxonomically identified and technologically profiled the cultivable microbial fraction present in sour ale beer during maturation step in wooden barrel and identified Saccharomyces wild yeast candidates to be exploited as starters in wild ale beer production.
2. Materials and Methods
2.1. Chemicals, Media, and Reference Strains
Chemicals were purchased from Sigma Aldrich (St. Louis, MO, USA), unless otherwise stated. Media and anaerobic systems were purchased from Oxoid (Cambridge, UK), while reagents for molecular biology from Thermo Scientific (Waltham, Massachusetts, United States). Oligonucleotides synthesis and Sanger sequencing were carried out by Bio-Fab Research core facility (Rome, Italy).
Reference strains were listed in Table 1.
2.2. Sour Beer Production and Sampling
The sour beer was produced at a craft brewery located in Bologna, Italy. Figure 1 illustrates the semi-spontaneous fermentation method employed by the craft brewery.
Three distinct beer products labeled as 1, 2, and 3 were blended in equal proportions (1:1:1). In detail, beer 1 was produced by a spontaneous fermentation of a wort (cereal grist: barley malt 72%, unmalted wheat 15%, rye 8%, cara malt 5%) with an adjunct of fresh whole grapes with skins (grapes/wort ratio 1:10) and an ageing of 8 months in barrel; beer 2 was produced through a wort fermentation (same grist as beer 1) with the single strain Brettanomyces bruxellensis WLP650 (White Lab), a subsequent aging of 5 months in barrels, and a final maceration (2 months) of fresh whole cherries; beer 3 was produced with a cereal grist of barley malt 62%, unmalted wheat 30%, oat 8% that underwent primary fermentation with a blend of three commercial S. cerevisiae strains (WLP565, Atecrem, and 3724; Table 1), followed by a maturation in barrel for 10 months. Samples were shipped to the laboratory under refrigerated conditions and immediately submitted to microbiological and physicochemical determinations.
2.3. Microbial Counts, Bacterial and Yeast Isolation
After ten-fold dilution in saline solution (9 g/L NaCl), appropriate sample dilutions were spread on Petri plates. Yeasts count and isolation was performed on YPDA (1% w/v yeast extract, 1% w/v peptone, 2% w/v dextrose, 2% w/v agar) and Wallerstein Laboratory (WL) media supplemented with chloramphenicol (100 μg/mL), incubated for 24-48 h at 28 °C. Man-Rogosa-Sharpe medium supplemented with maltose (5 g/l) and cycloheximide (20 mg/mL) (mMRS) was used for lactic acid bacteria (LAB) isolation setting the anaerobic incubation at 30 °C for 48-72 h.
Yeast and LAB populations were quantified in triplicate by counting plate colonies ranging from 20 to 200 and expressing the results as mean Log10 CFU/mL. Colonies were purified through at least double streaking on the same isolation medium. Cultures were microscopically examined for cellular morphology, and bacterial isolates were further screened for Gram staining and catalase activity. For long-term preservation, isolates were cryopreserved at -80°C using the corresponding isolation medium supplemented with 25% (v/v) glycerol.
2.4. Yeast and LAB Identification
Yeast genomic DNA was extracted according to Hoffman and Winston [24], while LAB DNA according to Gala et al. [25]. The integrity of DNA was checked by electrophoresis on a 0.8% (w/v) agarose gel in 0.5X TBE buffer (45 mM Tris–HCl, 45 mM boric acid, and 1 mM EDTA, pH 8.0), and DNA quality and quantity were evaluated spectrophotometrically using a NanoDrop ND-132 1000 device (Thermo Scientific, Waltham, USA). All DNA samples were diluted to a concentration of 50 ng/μL with ddH2O for further analysis.
Primers, reaction mixture, and cycling conditions of PCR amplification reactions were detailed in Supplementary Table S1. Concerning yeast identification, the region including 5.8S rRNA gene and the upstream and downstream internal transcribed spacers (globally referred to as ITS region) was PCR amplified in a final volume of 40 μL using the primers ITS1 and ITS4, as described by White et al. [26]. The restriction fragment length polymorphism (RFLP) analysis of ITS amplicons was carried out with the endonucleases HaeIII and HinfI [27] and the length of PCR amplicons and RFLP fragments were compared to those in the YeastID database (www.yeast-id.org; last accessed on 14 December 2022) using rank parameters at +/-10. Discrimination of S. cerevisiae, S. eubayanus, and S. uvarum species was carried out according to the species-specific PCR method previously described [28-29]. All primers concentrations were set up to 0.4 μM, except for primer SbayR1, which was used at the final concentration of 0.8 μM. Strains representative of each ITS RFLP and species-specific PCR profile were subjected to PCR amplification of D1/D2 region of 26S rRNA (LSU) gene according to Martini et al. [30].
Concerning LAB identification, 16S rRNA gene was PCR amplified with the primers 27f and 1490r [31]. Amplified 16S Ribosomal DNA Restriction Analysis (16S-ARDRA) was performed by the endonucleases HhaI, HinfI, and Tru1I and the resulting 16S-ARDRA profiles were compared to the in silico profiles obtained for the most common LAB beer species (Supplementary Table S2).
2.5. Phylogenetic Analysis
Phylogenetic analyses of yeast and bacteria strains were performed using D1/D2 LSU and 16S rRNA sequences, respectively. Amplicons were purified using DNA Clean & Concentrator™-5 Kit (Zymo Research, Orange, CA, USA) and sequenced using both amplification primers. Contig sequences were merged using the program SeqMan (DNASTAR, Madison, WI, USA) and the poor-quality ends were edited manually to remove primers. The resulting contig sequences were used as queries in a Blastn search against the NCBI RefSeq database [32] (last asses on 14 February 2023). Cut-offs of 98.7% and 99% gene similarity were used for bacterial and yeast species attribution, respectively [33-34]. The sequences were aligned with Muscle program [35] in MEGA X software [36] and the resulting alignments were subjected to a DNA substitution model analysis to select the best-fitting model. Phylogenetic relationships were inferred using the Neighbor joining method [37]. Confidences for the phylogenetic tree were estimated from bootstrap analysis (1000 replicates). Trees were visualized using Interactive Tree of Life (ITOL) [38] and rooted at outgroup reference species. The sequence data of D1/D2 LSU regions were deposited in the GenBank NCBI database under the accession numbers OM618137 to OM618146, while the 16S rRNA gene sequences under the accession numbers OM618118 to OM618120.
2.6. Genotyping Analysis
All the LAB and yeasts isolates were genotyped with a repetitive extragenic palindromic-polymerase chain reaction (rep-PCR) fingerprinting analysis based on (GTG)5 primer (5’-GTG GTG GTG GTG GTG-3’) [39], as previously described [40-41].
Commercial ale beer starters used in primary fermentation (Figure 1) and listed in Table 1 were genotyped by three different genotyping methods, such as (GTG)5 rep-PCR, inter-delta PCR, and R3-RAPD. (GTG)5 rep-PCR was carried out as reported above. PCR amplification of LTR regions near to TY1 and TY2 retrotransposon (referred to as inter-delta regions) was carried out with primers d12 (5’-TCAACAATGGAATCCCAAC- 3’) and d21 (5’-CATCTTAACACCGTATATGA -3’) [42], while R3-RAPD PCR amplification was carried out using the primer R3 (5’-ATG-CAGCCAC-3’) [43] (Supplementary Table S1).
(GTG)5 rep-PCR profiles were analyzed by electrophoresis on a 1.8% (w/v) agarose gel (25 cm x 27 cm) in 0.5X TBE buffer at the constant voltage of 70 V for 6 h under cold conditions; inter-delta and R3-RAPD profiles on a 1.5% (w/v) agarose gel (15 cm x 25 cm) in 0.5X TBE buffer at a constant voltage of 70 V for 150 min under cold conditions. In all genotyping tests, agarose gels were stained with SYBR™ Safe DNA Gel Stain (Invitrogen, Waltman, MA, USA) and GeneRuler 100 bp Plus DNA Ladder™ was used as a molecular weight standard. After visualization under UV light, digitalized images were analyzed using the BioNumerics software v8.10 (Applied Maths, Sint-Martens-Latem, Belgium). Pearson’s correlation similarity coefficient was used to convert computed band patterns into similarity matrix. Optimization and curve smoothening parameters were optimized with the specific script present in BioNumerics. The unweighted-pair group method analysis using the arithmetic means (UPGMA) was used for tree construction. Yeasts and LAB isolates with ≥ 92 and 85% similarity values were treated as a single strain, respectively.
2.7. Flocculation, Sporulation Assays and MAT Locus Genotyping
Flocculation assay was performed as reported in [44]. Briefly, yeasts were propagated in YPDA medium until OD600nm> 2.0. Three mL of pure liquid cultures were centrifuged at 2000 g for 5 min, and cells were resuspended in an equal volume of flocculation buffer (50 mmol/L Na acetate/acetic acid, 5 mmol/L CaSO4; pH 4.5). The OD600nm values were spectrophotometrically measured immediately after resuspension, as well as after 15 and 30 min of incubation at room temperature without agitation. To assess flocculation ability, the following equation was used:
F (%) = (OD600nm after 15 or 30 min / OD600nm at starting time) x 100,
Yeasts were classified according to F% as follows: score from 0 to 10%, “very flocculant”; from 10 to 30%, "moderately flocculant"; from 70 to 90%, "poorly flocculant"; from 90 to 100%, "non-flocculant".
To induce sporulation, exponentially grown cells were sub-cultured to acetate agar plates (2% w/v agar, 0.6% w/v sodium acetate) [45] and incubated at 28 °C for two weeks. Asci formation was checked and counted with a Bürker chamber after 3 and 14 days using an optical microscope with 400X magnification. Sporulation percentage was calculated as follows:
Sporulation (%) = [(n° asci)/(n° asci +n° cells)] x 100,
Spore viability was determined by asci dissection and monospore clone constitution as previously reported [46], according to the following equation:
Spore Viability % = (n° of vital monosporic cultures / n° of dissected spores) x 100,
2.8. Maltose Fermentation Tests
Preliminary, maltose fermentation test was carried out according to Kurtzman et al. [45]. Sterile basal medium (0.45% w/v yeast extract, 0.75% w/v peptone and 0.0048% w/v bromothymol blue as pH indicator) (4.5 mL) was supplemented with the filter-sterilized stock sugar solution (either maltose or glucose) at the final concentration of 2% (w/v) and dispensed in test tubes with screw caps with an upside-down inverted Durham tube inserted. Each yeast strain was propagated in YPD medium at 27 °C for 24 h and inoculated at the final concentration of 2 x 107 CFU/mL. Tubes were incubated at 28 °C, and gas production was checked after 3, 7, and 14 days. Scores were attributed as follows: +, strongly positive, insert filled within 7 days; s, slowly positive, insert slowly filled after more than 7 days; −, no gas production. Cells inoculated in basal medium with 2% (w/v) glucose were used as a control.
Maltose consumption tests were carried out in triplicates in 100 mL Erlenmeyer flasks containing 45 mL of YNB (0.67% w/v Yeast Nitrogen Base) medium supplemented with 2% (w/v) of either glucose or maltose and sealed with cotton plugs, as previously reported [46-47]. After 48 h of incubation at 25 °C under shaking conditions (150 rpm), optical density (OD) values at 600 nm (OD600nm) were measured using an UV-Vis photometer.
2.9. Wort Preparation and Micro-Scale Fermentation Trials
For the wort preparation, Pilsen malt (195 kg) (Bestmalz, Heidelberg, Germany), unmalted wheat (130 kg), unmalted oat (Avena sativa) (32 kg), and hop pellets Hallertau Perle (Hopsteiner, Mainburg, Germany) were mixed in a final volume of 1600 L. The “Liquor-to-grist” ratio was maintained at 3:1. The mash temperature steps involved 10 min at 45 °C, 20 min at 55 °C, 40 min at 66 °C, and 10 min at 78 °C. The boiling process was carried out for 60 min. The resulting hopped wort (11.5 °Plato original gravity; density 1.07 g/L, pH value of 5.15 ± 0.02) was pasteurized at 100 °C for 10 minutes, and subsequently frozen at -20 °C until use.
Micro-scale wort fermentation trials were conducted in duplicate using 200 mL glassware bottles equipped with airlocks and filled with 100 mL of pasteurized hopped. A single colony of each yeast candidate was propagated at 27 °C for 48 h in YPD medium under orbital shaking (150 rpm). After recovery by centrifugation for 6 minutes at 2500 g, cells were resuspended in pasteurized wort and inoculated in wort at the final concentration of 107 cells/mL. Fermentation bottles were maintained at 20 °C for 21 days and daily monitored over time by CO2 release, weighting the fermenters to track record of weight loss. The un-inoculated beer wort was used as negative control.
The fermentation curves were modeled based on the CO2 evolution, using the "grofit" package for R [48]. The maximum rate of fermentation (µ), maximum fermentation efficiency (the total amount of CO2 released at the end of the fermentation) (A), and the time of the lag phase (λ) were determined using parametric fitting methods in “grofit”.
2.10. Physicochemical Analysis
The pH was determined at the sample temperature of 25 °C (Crison Instruments, Barcelona, Spain).
Carbohydrates and organic acids in sour beer, wort, and fermented broths, clarified by centrifugation (13000 rpm for 5 min at 4°C) and filtration at 0.22 µm, were analyzed in an HPLC system equipped with a refractive index detector (1200 System, Agilent Technologies, Waldbronn, Germany). Elution in isocratic condition was carried out at 60°C with 0.6 mL/min of 5 mM H2SO4 through an ion exclusion column (Aminex HPX-87 H, Bio-Rad, Hercules, CA, USA).
Volatile organic compounds (VOCs) were traced by solid-phase microextraction (SPME) followed by gas chromatography-mass spectrometry analysis (GC–MS). Two mL of samples were spiked with 100 µL of 1-chloro-2-fluoro-benzene 5 mg/L, as internal standard, and the headspace was exposed to a divinylbenzene/carboxen/polydimethylsiloxane fiber (DVB/CAR/PDMS Supelco; Sigma-Aldrich, St. Louis, MO, United States) for 1 h at 60 °C. Analytes were desorbed in a GC–MS apparatus (7820–5975; Agilent Technologies, Santa Clara, CA, United States) equipped with a DB-624 column (30 m ✕ 250 mm ✕ 1.4 mm, Agilent Technologies). Separation was achieved with 1.3 mL/min helium flux following a thermal gradient: 2 min isotherm at 50 °C, 6 °C/min increase to 110 °C, 10 °C/min increase to 240 °C, 4 min isotherm at 240 °C. NIST 14 spectral library was utilized to infer the identity of the compounds by comparison of their mass spectra. The peak areas of compounds, normalized by the area of the inner standard, were taken to be proportional to their abundances.
2.11. Screening of STA1 Gene
The screening for STA1 gene was conducted by endpoint PCR using the SD-5A (5-CAACTACGACTTCTGTCATA-3) and SD-6B (5-GATGGTGACGCAATCACGA-3) primers [49], while the analysis of the promoter region, particularly the UAS2 portion, was conducted with the primers STA1_UAS_Fw (5'-CCTGGCTCAAATTAAACTTTCG-3') and STA1_UAS_Rv (5'-ACCACCAATAGGCAATAGAAA-3') [50]. PCR mixtures and cycling conditions were reported in Supplementary Table S1. PCR amplicon purification, sequencing and sequence analysis were carried out as reported above.
2.12. Statistical Analysis
The datasets were analyzed using GraphPad Prism 8 software (GraphPad Software Inc., San Diego, CA) through one-way or two-way ANOVA. A p-value threshold of 0.05 was set for both analyses. All values presented in this study are the means of three replicates ± standard deviation (SD).
Principal component analysis (PCA) was applied to examine the data matrix of VOCs profiles, presenting a size {20, 53}, consisting of the 53 relative areas of VOCs, determined for the two independent fermentation with the 9 yeast strains in addition to the initial wort. The screen plot was utilized to establish the number of significant principal components (PCs) based on the percentage of variance explained. Scores and loadings of the PCA model were calculated using Past version 4.12 [51], as well as UPGMA hierarchical clustering of samples based on the Bray-Curtis similarity index of VOCs profiles.
3. Results
3.1. Sour Beer Physicochemical Caracterization and Microbiological Analysis
The samples were collected during the late maturation phase. Sour beers are distinguished from ales and lagers by their elevated levels of organic acids, specifically lactic acid and acetic acid. These acids contribute to a lower pH and a more pronounced acidic taste. The beer was characterized by a pH of 3.37±0.01 and by the abundance of lactic acid (13.0 g/L), which was the major metabolite, after ethanol (51.8 g/L), likely resulting from the catabolic conversion of wort sugars operated by LAB. Other primary catabolites observed in the beer were glycerol (3.3 g/L) and acetic acid (1.0 g/L). Microbiological counts on YPDA and WL media resulted in values of 4.39±0.10 and 4.28±0.3 Log10 CFU/mL, respectively. mMRS count was 6.00±0.03 Log10 CFU/mL confirming the primary role of LAB in the production process and their predominance on the yeast population.
3.2. LAB Identification and Genotyping
All LAB isolates were cocci-shaped, Gram positive, and catalase negative. They were submitted to 16S-ARDRA with three endonucleases, namely HhaI, HinfI, and TruI. These restriction enzymes were proved to be diagnostic for most LAB species frequently encountered in sour beer fermentation (Supplementary Table S2). 16S-ARDRA patterns supported the attribution of LAB isolates to Pediococcus parvulus/Pediococcus damnosus, which are recognized as beer spoilage species and frequently found as dominant in wood aging phase of sour beer process (as reviewed by [52]).
Phylogenetic analysis of 16S rRNA gene sequences was carried out on 7 randomly selected LAB isolates. The strains formed a monophyletic group with P. damnosus strain DSM 20331T (GenBank accession number: NR_042087.1) (Figure 2, panel A). This species is critical in sour beer the production, as it is more hop-tolerant and low pH-tolerant than lactobacilli [23] and it is very persistent in wood barrels [53]. Pediococcus spp. produce diacetyl, that is responsible for a rancid and buttery flavor. However, in craft beer this effect is attenuated by some non-Saccharomyces yeasts which consume diacetyl. On the other hand, Pediococcus spp. decrease pH more slowly than lactobacilli, assuring yeasts to complete primary fermentation [55].
(GTG)5 rep-PCR was applied to analyze the genetic diversity within the group of LAB isolates. The patterns consisted of a minimum of 10 bands to a maximum of 13 bands with dimensions ranging from 150 to 2270 bp. The UPGMA dendrogram identified 5 singleton strains and one subcluster at 85% of rep-PCR profile similarity, suggesting high diversity of LAB strains isolated from sour beer (Figure 2, panel B).
3.3. Yeast Identification
A total of 50 yeast isolates were randomly selected from countable plates, 20 from YPDA and 30 from WL, respectively. Raw data about PCR-RFLP analysis of ITS regions were reported in Supplementary Table S3. Search in Yeast-ID database allowed to match five different patterns (A to E) to the following species: pattern A to S. cerevisiae (100%)/ Saccharomyces cariocanus (100%)/Saccharomyces paradoxus(100%); pattern B to Pichia membranifaciens (73%); pattern C to Saccharomyces bayanus (89%)/Saccharomyces kudriavzevii (89%)/Saccharomyces pastorianus (89%)/Saccharomyces mikatae (89%); pattern D to Kluyveromyces blattae (67%); and pattern E to Brettabomyces bruxellensis (83%).
Representative strains from each ITS PCR-RFLP pattern were submitted to sequencing of variable domain D1/D2 at the 5' end of the 26S rRNA gene. In details, strains WY104, WY115, and WY203 (pattern A) grouped monophyletically with S. cerevisiae NRRL Y-12632 (97% bootstrapping) (Figure 3). Strains WY122 and WY102 (pattern B) shared the highest D1/D2 26S rRNA sequence similarity with Pichia membranifaciens (Figure 3). However, three substitutions (60T>C, 167C>T, and 179C>T) did not allow the monophyletic clustering of these strains with P. membranifaciens strain CBS107T. This species has been identified in lambic and gueuze beers [17,55], wine [56-58], and other beverages [59,60]. Pichia membranifaciens is one of the most frequently yeast associated to mycobiome of sour beer during wood aging [20,61] due to the capability to form biofilm which accounts for the long persistence of this species on the wooden surface [62]. While it has been described that Pichia spp. can be involved in the biotransformation of hop terpenes (incl. geraniol) [63], this species also produces unpleasant compounds, such as 4-ethylphenol and 4-ethylgyaiacol [57].
Strains WY115, WY205, and WY213 (pattern C) were phylogenetically related to the complex taxon S. bayanus, which includes two well-differentiated groups of strains, such as S. bayanus var. bayanus (S. eubayanus × S. uvarum) and S. bayanus var. uvarum (Figure 3) [64,65]. Libkind et al. [66] proposed the use of S. eubayanus and S. uvarum as descriptors of species but restricted the name S. bayanus to the hybrid lineages between these pure species. S. uvarum was isolated in natural European wine [67-69] and cider fermentations [70-71], as well as in South American chicha fermentations [72]. S. bayanus/S. uvarum and the hybrid species S. pastorianus (synonym Saccharomyces carlsbergensis) are extensively used to produce lager beer through bottom fermentation at low temperature [73]. Occurrence of S. bayanus/S. uvarum was previously documented in the early stages of lambic beer fermentation; after that this species should disappear due to high ethanol content and low pH [20].
Finally strains WY59 (pattern D) and WY60 (pattern E) clustered together with Brettanomyces anomalus (homotypic synonym: Dekkera anomala) and B. bruxellensis with highest bootstrapping values, respectively (Figure 3). For strain WY59, phylogenetic results partially disagreed with ITS PCR-RFLP analysis. This strain showed an amplified ITS product of 550 bp (Supplementary Figure S1, panel A) instead of the expected 800 bp-long ITS product reported for B. anomalus in Yeast-ID database (last access on 2nd May 2023). Phylogenetic analysis of ITS sequences confirmed that strain WY59 formed a monophyletic cluster with B. anomalus CBS 77T (Supplementary Figure S1, panel B).
The presence of S. bayanus/S. uvarum is unusual in old-style sour beers which are produced at room temperature. Furthermore, hybrid strains frequently retain only one parental type of rDNA arrays in their genome, resulting in ITS restriction profiles either of S. cerevisiae type or S. eubayanus/S. uvarum type [74] (Figure 4, panel A). This loss of heterozygosity in rDNA marker makes ITS-RFLP analysis unable to discriminate hybrids from their parental lineages. Based on these considerations, we performed three species-specific PCR assays targeting S. eubayanus, S. cerevisiae, and S. uvarum species, respectively, to confirm species attribution of strains WY115, WY205, and WY213 to S. uvarum/S. bayanus and of 35 strains to S. cerevisiae. As expected, S. cerevisiae reference strains BY4743 and BY4742, as well as sour beer strains with pattern A were positive to S. cerevisiae-specific PCR and negative to S. bayanus/S. uvarum and S. eubayanus species-specific PCR assays (Figure 4, panel B). Remarkable, S. uvarum/S. bayanus sour beer strains WY115, WY205, and WY 213 resulted positive to both S. cerevisiae and S. bayanus/S uvarum species-specific PCRs but gave no PCR amplicons with S. eubayanus-specific primer pair (Figure 3, panel B). Overall, this evidence supported that the strains WY115, WY205, and WY213 could be S. cerevisiae x S. uvarum natural hybrids which only retained the S. uvarum/S. bayanus rDNA copy in their genomes.
As a whole, the culturable yeast fraction of natural sour beer considered in this study consisted of S. cerevisiae (70%), followed by P. membranifaciens (16%), S. cerevisiae x S. uvarum hybrids (6%), D. anomala (4%), and B./D. bruxellensis (4%) (Figure 3, panel B).
3.4. Yeast Genotyping
The genetic diversity of the 50 yeast isolates was estimated using the (GTG)5 rep-PCR method. The resulting patterns comprised from 4 to 23 bands with dimensions between 200 and 2830 bp. A UPGMA dendrogram, based on Pearson's similarity coefficient, was grouped the 50 isolates into two major clusters (I and II) (Figure 5). Applying a reproducibility cutoff of 92% led to the identification of 14 biotypes, consisting of 9 subclusters (S1 to S9) and 5 singletons.
Cluster I comprised 8 yeast isolates belonging to the species P. membranifaciens (subclusters S1 and 2 singletons), as well as the S. cerevisiae x S. uvarum hybrid strain WY115 and S. cerevisiae strain WY220. Within cluster II, 30 S. cerevisiae isolates were divided into 5 subclusters (S4 to S8), while S. cerevisiae isolates WY112, WY111, and WY104 were singletons. S. cerevisiae x S. uvarum hybrids WY213 and WY205 clustered together in minor cluster S3, while B. anomalus and B. bruxellensis isolates divided into two subclusters, S2 and S9, respectively.
We investigated whether the S. cerevisiae isolates, one belonging to each (GTG)5 REP-PCR subcluster, were genetically different from the commercial S. cerevisiae pure cultures used in primary fermentation, namely the strains Atecrem Belgian Saison, 3724, and WLP565 (Table 1). The (GTG)5 rep-PCR technique was not discriminatory. Conversely, two alternative techniques, the inter-delta PCR assay, which amplifies LTR regions among retrotransposable elements Ty1 and Ty2, and the R3-RAPD technique, which randomly amplifies non-coding and coding DNA using R3 sequence as a primer showed that the S. cerevisiae wild isolates were genetically distinct from the commercial starter ones (Supplementary Figure S2, panels A and B).
3.5. Maltose Consumption and Flocculation
A pool of 9 selected sour beer wild strains were screened for their ability to consume maltose and flocculate. Maltose is the primary fermentable sugar in brewer's wort, while flocculation can be desirable in removing yeasts after fermentation. As shown in Figure 6, S. cerevisiae strains WY117, WY203, and WY104 consumed maltose at the same extent of glucose (p>0.05), while B. anomalus WY59, and B. bruxellensis WY60 did not grow on maltose as unique carbon source after 48 h of incubation. This result was further confirmed by conventional fermentation test (data not shown). Differently from the conspecific strain WY102, P. membranifaciens WY122 was able to consume maltose (p<0.05). Conventional fermentation test confirmed this result (Supplementary Figure S3), which contradicted the maltose-negative phenotype described for this species [75]. Concerning flocculation assay, none of the strains showed significant flocculent phenotype (Supplementary Table S3).
3.6. Sporulation and MAT Genotyping
Four S. cerevisiae strains and 2 S. cerevisiae x S. uvarum hybrids were evaluated for their exploitability as parental lineages in hybrids construction. For this purpose, we determined sporulation efficiency and spore viability. Sporulation efficiency of S. cerevisiae strains varied from 22 to 23% after 3 days of incubation at 28 °C on agar acetate plates and slightly increased after 14 days (Table 2). S. cerevisiae x S. uvarum hybrids were not able to sporulate.
Spore viability can be defined as the ability to growth on a reach medium at 28 °C for 2 to 5 days and to produce a viable monospore colony after tetrad dissection. Despite S. cerevisiae strains were moderately sporulating, the spore viability was higher than 50%, with S. cerevisiae WY203 showing the highest spore viability (Table 2).
Since ploidy status and homothallic/heterothallic lifestyle strongly affect hybridization yield of Saccharomyces strains [46], we determined the MAT locus in each S. cerevisiae strain and their monospore progeny. All strains exhibited a MATa/MATα genotype (Table 2) and produced MATa/MATα monosporic progenies, suggesting that they are diploid and homothallic (HO/HO) strains (Supplementary Figure S4).
3.7. Micro-Scale Wort Fermentation
We performed test fermentations using both maltose consuming (S. cerevisiae WY104, WY117, WY203, and WY220; P. membranifaciens WY122; S. cerevisiae x S. uvarum WY213) and maltose non-consuming strains (P. membranifaciens WY102, D. anomala WY59, and D. bruxellensis WY60). To assess the kinetics variables during wort fermentation, we monitored the CO2 loss in the fermentations by weighing. S. cerevisiae strains exhibited the shortest lag phase and the highest fermentation efficiency values (p<0.05) (Figure 7) and did not differ significantly from S. cerevisiae x S. uvarum hybrid WY213 (p>0.05). After a long lag phase, maltose-positive P. membranifaciens WY122 showed fermentation efficiency and fermentation rate similar to S. cerevisiae. As expected, maltose-negative P. membranifaciens strain WY102 was slow to adapt to wort environment and very low in fermentation efficiency but, after a long lag phase, it exhibited high fermentative rate. B. bruxellensis WY60 was better than B. anomalus WY59 to ferment wort (p<0.05).
The consumption of carbohydrates and ethanol content were measured at the end of fermentation and were consistent with the fermentation curves (Figure 8). Specifically, Saccharomyces strains outperformed non-Saccharomyces ones in maltose and maltotriose utilization, except for P. membranifaciens WY122. Coherently, Saccharomyces strains and P. membranifaciens WY122 produced higher amounts of ethanol (ranging from 20.8 to 34.1g/L). Notably, S. cerevisiae x S. uvarum hybrid WY213 demonstrated not only the highest ethanol production, but also the highest rate of carbohydrate-to-ethanol conversion (40%w/w), considering that the maximum theoretical value is 51%.
HPLC analysis revealed a partial consumption of dextrins, which generally are unfermentable sugars for commercial (domesticated) ale yeasts (Figure 8). STA1 gene encodes extracellular glucoamylase which mostly contributes to dextrin consumption [50-76]. Recent studies showed that the mere presence of gene STA1 is not enough to be diagnostic for a “diastatic” phenotype. Krogerus et al. [50] proved that a full-length promoter assures STA1 gene expression in truly diastatic strains, while the UAS2 region of promoter is deleted in poorly diastatic STA1+ strains. Therefore, we investigated the presence of STA1 gene and the related functional promoter in S. cerevisiae strains. As shown in Figure 9, STA1 gene was present in our S. cerevisiae sour beer strains, but the promoter-specific PCR gave a PCR product longer than that expected (630 bp instead of 599 bp). Sequencing of promoter PCR amplicon showed two indels and 20 substitutions in UAS2 promoter region of S. cerevisiae WY117 compared to the truly diastatic strain S. cerevisiae 3711.
3.8. VOCs Determination
In the headspace of wort and fermented samples, SPME-GC analysis revealed a total of 53 compounds detected in at least two samples (Supplementary Figure S5). The VOCs occurring most frequently and abundantly were ethanol and other alcohols, esters, organic acids, aromatic compounds, furans, aliphatic hydrocarbons, and terpenoids. The PCA model revealed that the 2 highly informative dimensions, PC1 and PC2, accounted for 61.5% and 21.1% of data variability (Figure 10, panel A). Ethanol, 3-methyl-1-butanol, and phenylethyl alcohol presented positive loadings to PC1, resulting the major contributors to differentiate fermented samples (Figure 10, panel B). Conversely, terpenoids like humulene and beta-myrcene negatively contributed to PC1 distinguishing the profile of unfermented worth. A group of ethyl esters (i.e., hexanoic-, decanoic-, octanoic-, and ethanoic- acid ethyl esters) also contributed to characterize fermented samples but with positive loadings along PC2. Fermented samples appeared spread in two groups: samples inoculated with strains B. anomalus WY59, B. bruxellensis WY60, and P. membranifaciens WY102 were mainly located in the second quarter of the graph, at negative values of PC1 and positive PC2. This agrees with previous observation that Dekkera/Brettanomyces spp. produce fatty acid ethyl esters, such as octanoate and ethyl decanoate, responsible for fruity and grape odor, respectively, better than S. cerevisiae [77]. Samples obtained by fermentation with Saccharomyces strains and P. membranifaciens WY122 were located in the first and fourth quarters, at positive values of PC1. WY122 exhibited a VOCs profile intermediate between Saccharomyces and non-Saccharomyces strains, characterized by concomitant presence of a more complex ethyl esters profile and high amount of ethanol. Among the aroma-active esters, ethyl esters, which play a crucial role in determining the aroma of beer [78], were the second-most abundant VOCs after acetate esters. Examples of ethyl esters include ethyl hexanoate (aroma like anise seed or apple), ethyl octanoate (sour apple aroma), and ethyl decanoate (floral odor). Ethyl esters are formed intracellularly by fermenting yeast cells, the rate of formation is highly strain-dependent and is mainly affected by the concentrations of the two precursors, ethanol and acyl coenzyme A, where the acid group is a medium-chain fatty acid (MCFA) [79]. During alcoholic fermentation, MCFA intermediates are prematurely released from the cytoplasmic fatty acid synthase (FAS) complex. The key enzyme involved in the regulation of fatty acid biosynthesis can promote the release of MCFAs, which dramatically affects the formation of ethyl esters and contribute to differentiates the yeast strain's profile.
4. Discussion
Bioprospecting for native strains isolated from environments either alternative or related to brewery can contribute to diversify the portfolio of craft beer cultures. For instance, sourdough strains were proposed as S. cerevisiae parental lineage in synthetic hybrid construction [46] and as brewing cultures to produce low ethanol beer [80]. Low alcohol beer was also successfully produced with S. cerevisiae inhabiting brewery environment [81]. In this study, we applied a culturable approach to characterize the yeast fraction present in sour beer during aging in wooden barrel and proved that these indigenous yeasts can have promising brewing attributes. DNA barcoding techniques and PCR fingerprinting assured the constitution of a pool of assorted yeast candidates for subsequent phenotypic assays.
Previous studies on microbial succession during traditional lambic beer process showed that Saccharomyces species dominate the early stages, when they sequentially consume mono-, di-, and trisaccharides, releasing yeast-associated metabolites, such as ethanol, succinic acid, and methyl-1-butanol [82,83]. Depletion of maltose and maltotriose caused starvation of Saccharomyces species and was followed by the acidification phase [18]. In late stage of maturation, oxidative species such as Dekkera spp., B. bruxellensis, and P. membranifaciens should dominate the mycobiota as they possess intra- and extracellular α-glucosidases responsible for maltooligosaccharide degradation. The sour-matured production style considered here entails the inoculation with S. cerevisiae commercial starters to obtain a base beer that is then submitted to the maturation step in wood barrels. This method shortens the time of brewing and increases predictability of the process compared to traditional lambic beer process, without forgoing the crucial role of the wooden barrels as an additional microbial inoculation source. To the best of our knowledge, no studies have characterized the cultivable yeast fraction from microbrewery plants based on a sour-matured production style process. Unlike the sequential succession of yeast species documented in traditional lambic beer process, in this study S. cerevisiae dominated the cultivable mycobiota at the maturation stage in barrels, followed by P. membranifaciens, B. anomalus, and B. bruxellensis. These yeast species co-exist with LAB species P. damnosus. Using ITS1 metabarcoding approach, Shayevitz et al. [84] also demonstrated that S. cerevisiae is highly abundant in aged sour ale beer samples. As S. cerevisiae is frequently isolated from oak wood [85], it is reasonable that this species is a common resident in wood barrels used for beer souring. Furthermore, S. cerevisiae wild strains were genetically distinct from commercial cultures used for base beer fermentation, supporting that the strains are feral yeasts inhabiting the craft brewery plan. Further investigations on domestication signatures in the genome could elucidate whether they are truly wild strains or brewing strains that have evaded the domestication niche and were able to colonize natural environments [86].
Another relevant question is how these S. cerevisiae strains can survive in depletion of maltose and maltotriose. During co-fermentation experiments, Coehlo et al. [87] reported that both S. cerevisiae and B. bruxellensis partially consume ethanol when glucose was depleted, according to the make-accumulate-consume strategy [88]. However, B. bruxellensis can outperform S. cerevisiae in ethanol consumption, the latter being able to metabolize a wider range of carbon sources and turn to ethanol as a last resort. All our strains possess STA1 gene under the control of a promoter slightly longer than that found in “diastatic” reference strain 3711. This gene encodes an extra-cellular glucoamylase (EC 3.2.1.3) which cleaves α-1,4-glycosidic bonds from the non-reducing ends of soluble maltodextrin chains releasing glucose, subsequently fermented. Deletion of STA1 gene reduced maltotriose consumption in STA1-positive beer strains belonging to lineage Beer 2 [50,76], suggesting that Sta1p contributes to maltotriose consumption in these strains. The partial degradation of dextrin detected during microscale wort fermentation support the “diastatic” nature of our S. cerevisiae strains. Although we did not investigate the STA1 gene expression, we can hypothesize that the ability to survive during beer aging can be related to a “diastatic” phenotype. “Diastatic” is generally considered a negative trait in brewery as being responsible for over-attenuation and beer quality deterioration. Furthermore, “diastatic” strains generally have functional PAD1 and FDC1 genes responsible for the decarboxylation of ferulic acid into 4-vinyl guaiacol, a glove-like aroma [89]. However, in some kinds of craft beers, such as sour and Saison-style beers, 4-vinyl guaiacol and other phenolic derivative molecules, such as 4-ethyl phenol, contribute to the ‘funky’ character of these beers by imparting spicy and barnyard flavor note [81]. We did not specifically investigate POF (phenolic off-flavor) phenotype, but wort fermentation with S. cerevisiae strains did not reveal 4-vinyl guaiacol while highlighted the presence of phenylethyl alcohol which contributes to fruity-like aroma. These data indicate that these strains could contribute to formation of desirable aroma-active esters.
The intermediate sporulation efficiency associated to high viability of the spores supported that the novel isolates, as well as their monosporic derivatives, could be exploitable in synthetic hybrid construction. Interestingly, we isolated three strains positive to both S. cerevisiae and S. uvarum-specific PCR assays targeting housekeeping genes. Both ITS-RFLP analysis and D1/D2 26S sequencing indicated that these strains only retained the S. uvarum rDNA array. These results suggested that these strains could be S. cerevisiae x S. uvarum hybrids. According to this hypothesis, no sporulation was observed under the conditions used in this study, suggesting that either pre-zygotic barriers or polyploidy/aneuploidy prevent meiosis in these strains. S. cerevisiae × S. eubayanus and S. cerevisiae × S. kudriavzevii interspecies hybrids were frequently documented in craft brewing. S. pasteurianus was one of the most abundant species at the end of alcoholic fermentation phase in traditional lambic sour beer production [17,55], while many Trappist-style beers from Belgium are brewed with hybrids of S. cerevisiae × S. kudriavzevii [90]. This might be a consequence of winter season temperature which favors yeasts that are more cryotolerant, such as S. kudriavzevii and S. pastorianus. S. cerevisiae × S. uvarum hybrids were documented in brewing less frequently than S. pasteurianus and S. cerevisiae × S. kudriavzevii interspecies hybrids. Recently, a S. cerevisiae × S. uvarum hybrid, the so-called Muri isolate, was characterized from a “kveik” culture, a traditional Norwegian farmhouse brewing yeast culture [91,92]. Dextrin consumption in microscale wort fermentation suggested that, like Muri isolate, WY213 could be STA1-positive.
Pichia membranifaciens is described as a typical yeast species colonizing sour beer in late maturation step mainly due to this ability to adhere wooden surface [62]. As having a non-fermentative metabolism, it is generally considered a beer contaminant and poorly attractive as brewing cultures. Here the P. membranifaciens strain WY122 consumed maltose and exhibited an intermediate phenotype between non-S. cerevisiae species, such as the conspecific strain WY102, B. anomalus and B. bruxellensis, and the Saccaromyces strains. To the best of our knowledge only a P. membranifaciens strain from Chinese steamed bread was reported to use maltose as carbon source [93]. Genomic characterization of strain WY122 will contribute to better elucidate its taxonomic position.
5. Conclusions
During aging phase of sour-matured beer production style in a microbrewery, STA1-positive S. cerevisiae feral strains dominate the cultivable mycobiota fraction. Sour beer at the maturation stage can serve as reservoir of novel yeast cultures with interesting brewing attributes. A variety of yeast strains, including S. cerevisiae isolates, S. cerevisiae x S. uvarum putative hybrids, and P. membranifaciens WY122, possess industrially desirable phenotypic traits making them attractive candidates for both monocultures and cocultures exploitation in craft beer production.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Figure S1: Analysis of ITS region in D. anomala isolates. (A) Electrophoretic gel of ITS amplicons. D. bruxellensis strain WY60 was used as reference strain. (B) Phylogenetic tree obtained by neighbor joining (NJ) method [1] applied to a dataset of 7 rDNA sequences. The evolutionary distances were calculated by the Tamura 3-parameter method [2] considering the number of nucleotide substitutions per site. The gamma distribution was used to model the rate of change between sites. All positions containing gaps and missing data were eliminated (complete deletion option). The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test [3] (1000 replicates) are shown next to the branches. The strain collected in this study is shown in bold. The lengths of the branches are proportional to the number of nucleotide substitutions, and they have been measured using the divergence scale shown at the top left. The tree was rooted using Yarrowia lypolitica as outgroup. The tree data (Newick) were generated with MegaX [4] and exported and visualized using ITOL [5]. Abbreviation: M: molecular size marker; Bb: Brettanomyces bruxellensis; Figure S2: Figure S2. Dendrograms generated using inter-delta PCR (A) and R3-RAPD PCR (B) fingerprints of 11 S. cerevisiae strains (7 indigenous wild strains isolated in this study from sour beer, 3 commercial starter cultures commonly used in the brewery plant, and BY4743 as reference strain). Commercial starters (in red) were detailed in Table 1. Similarity percentages were calculated using Pearson correlation coefficient, while hierarchical clustering analysis was carried out using the UPGMA (unweighted pair-group method with arithmetic mean) method with Bionumerics software. Numbers near the branches represent branch lengths. The tree data (Newick) were generated with MegaX [4] and exported and visualized using ITOL [5]; Figure S3: Glucose and maltose fermentation test. Tubes containing Durham inverted tubes were photographed after 3 days of incubation at 27°C. Tests were carried out according to Kurtzamn et al. [6]. Abbreviations: G, glucose; M, maltose; Figure S4: Mating type genotyping of four S. cerevisiae sour beer wild strains and their monosporic derivatives. For each strain at least 8 meiotic events (asci) were dissected; Figure S5. Distribution of the 53 volatile organic compounds (VOCs) identified in the headspace of the fermented wort samples. The area of each VOC was mean centered and normalized by the standard deviation. Values are reported as colors ranging from the lowest (grey) to the highest (deepest red); Table S1: Primers, compound concentrations, and thermal conditions used this study. Table S2. In silico 16S-ARDRA profiles of the main beer LAB species. Table S3. ITS RFLP-PCR analysis of yeast isolates from natural sour beer.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, L.S. (Lisa Solieri); methodology, L.S. (Lisa Solieri); formal analysis, L.S. (Lisa Solieri), F.C., and S.R.; investigation, C.N., L.S. (Laura Sola), and J.R.; resource: M.D.B.; data curation, L.S. (Lisa Solieri) and S.R..; writing—original draft preparation, L.S. (Lisa Solieri); writing—review and editing, L.S. (Lisa Solieri), C.N, J.R., L.S. (Laura Sola), S.R., and F.C..; visualization, L.S. (Lisa Solieri) and L.S. (Laura Sola); supervision: L.S. (Lisa Solieri); funding acquisition, L.S. (Lisa Solieri). All authors have read and agreed to the published version of the manuscript.
Funding
L.S (Lisa Solieri) acknowledge the support of the NBFC to the University of Modena and Reggio Emilia, funded by the Italian Ministry of University and Research, PNRR, Missione 4 Componente 2, “Dalla ricerca all’impresa”, Investimento 1.4, Project CN00000033.
Data Availability Statement
The sequence data generated in this study have been submitted to NCBI database under accession numbers OM618137 to OM618146 and OM618118 to OM618120.
Conflicts of Interest
M.D.B was employed by Ca’ Del Brado Brewery. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Carbone, A.; Quici, L. Craft beer mon amour: an exploration of Italian craft consumers. Br. Food J. 2020, 122, 2671–2687. [Google Scholar] [CrossRef]
- Baiano, A. Craft beer: An overview. Compr. Rev. Food Sci. Food Saf. 2021, 20, 1829–1856. [Google Scholar] [CrossRef] [PubMed]
- Aquilani, B.; Laureti, T.; Poponi, S.; Secondi, L. Beer choice and consumption determinants when craft beers are tasted: An exploratory study of consumer preferences. Food Qual. Pref. 2015, 41, 214–224. [Google Scholar] [CrossRef]
- Viejo, C.G.; Fuentes, S. Beer aroma and quality traits assessment using artificial intelligence. Fermentation 2020, 6, 56. [Google Scholar] [CrossRef]
- Pires, E.J.; Teixeira, J.A.; Brányik, T.; Vicente, A.A. Yeast: the soul of beer’s aroma—a review of flavour-active esters and higher alcohols produced by the brewing yeast. Appl. Microbiol Biotechnol. 2014, 98, 1937–1949. [Google Scholar] [CrossRef] [PubMed]
- Svedlund, N.; Evering, S.; Gibson, B.; Krogerus, K. Fruits of their labour: biotransformation reactions of yeasts during brewery fermentation. Appl Microbiol Biotechnol. 2022, 106, 4929–4944. [Google Scholar]
- Holt, S.; Mukherjee, V.; Lievens, B. Verstrepen, K.J.; Thevelein, J.M. Bioflavoring by non-conventional yeasts in sequential beer fermentations. Food Microbiol. 2018, 72, 55–66. [Google Scholar] [CrossRef]
- Borneman, A.R.; Forgan, A.H.; Kolouchova, R.; Fraser, J.A.; Schmidt, S.A. Whole genome comparison reveals high levels of inbreeding and strain redundancy across the spectrum of commercial wine strains of Saccharomyces cerevisiae. G3: Genes, Genomes, Genetics 2016, 6, 957–971. [Google Scholar] [CrossRef]
- Krogerus, K.; Magalhães, F.; Vidgren, V.; Gibson, B. Novel brewing yeast hybrids: creation and application. Appl. Microbiol. Biotechnol. 2017, 10, 65–78. [Google Scholar]
- Cubillos, F.A.; Gibson, B.; Grijalva-Vallejos, N.; Krogerus, K.; Nikulin, J. Bioprospecting for brewers: exploiting natural diversity for naturally diverse beers. Yeast. 2019, 36, 383–3980. [Google Scholar] [CrossRef]
- Iattici, F.; Catallo, M.; Solieri, L. Designing new yeasts for craft brewing: when natural biodiversity meets biotechnology. Beverages 2020, 6, 3. [Google Scholar] [CrossRef]
- Denby, C.M.; Li RA, Vu VT. ; Costello, Z.; Lin, W.; Chan, L.J.G; Williams, J.; Donaldson, B.; Bamforth, C.W.; Petzold, C.J.; Scheller, H.V.; Martin, H.G.; Keasling, J.D. Industrial brewing yeast engineered for the production of primary flavor determinants in hopped beer. Nat. Commun. 2018, 9, 965. [Google Scholar] [CrossRef] [PubMed]
- de Ruijter, J.C.; Aisala, H.; Jokinen, I.; Krogerus, K.; Rischer, H.; Toivari, M. Production and sensory analysis of grape flavoured beer by co-fermentation of an industrial and a genetically modified laboratory yeast strain. Eur. Food Res. Technol. 2023, 30, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Astola, A.; Durán-Guerrero, E. , Díaz, A.B.; Lasanta, C.; Castro, R. Impact of the genetic improvement of fermenting yeasts on the organoleptic properties of beer. Eur. Food Res. Technol. 2023, 249, 1677–1687. [Google Scholar] [CrossRef]
- Alperstein, L.; Gardner, J.M.; Sundstrom, J.F.; Sumby, K.M.; Jiranek, V. Yeast bioprospecting versus synthetic biology—which is better for innovative beverage fermentation? Appl. Microbiol. Biotechnol. 2020, 104, 1939–1953. [Google Scholar]
- Dysvik, A.; Leanti La Rosa, S.; De Rouck, G.; Rukke, E-O. ; Westereng, B.; Wicklund, T. Microbial dynamics in traditional and modern sour beer production. Appl. Environ. Microbiol. 2020, 86, e00566–20. [Google Scholar] [CrossRef] [PubMed]
- Spitaels, F.; Wieme, A.D.; Janssens, M.; Aerts, M. Van Landschoot, A.; De Vuyst, L.; Vandamme, P. The microbial diversity of an industrially produced lambic beer shares members of a traditionally produced one and reveals a core microbiota for lambic beer fermentation. Food Microbiol. 2015, 49, 23–32. [Google Scholar] [CrossRef]
- De Roos, J.; Vandamme, P.; De Vuyst, L. Wort substrate consumption and metabolite production during lambic beer fermentation and maturation explain the successive growth of specific bacterial and yeast species. Front. Microbiol. 2018, 9, 2763. [Google Scholar] [CrossRef]
- De Roos, J.; Verce, M.; Weckx, S.; De Vuyst, L. Temporal shotgun metagenomics revealed the potential metabolic capabilities of specific microorganisms during lambic beer production. Front Microbiol. 2020, 11, 1692. [Google Scholar] [CrossRef]
- Bossaert, S.; Winne, V.; Van Opstaele, F.; Buyse, J.; Verreth, C.; Herrera-Malaver, B.; Van Geel, M.; Verstrepen, K.J.; Crauwels, S.; De Rouck, G.; Lievens, B. Description of the temporal dynamics in microbial community composition and beer chemistry in sour beer production via barrel ageing of finished beers. Int. J. Food Microbiol. 2021, 339, 109030. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Bamforth, C.W.; Mills, D.A. Brewhouse-resident microbiota are responsible for multi-stage fermentation of American coolship ale. PloS one 2012, 7, e35507. [Google Scholar] [CrossRef] [PubMed]
- Bongaerts, D.; De Roos, J.; De Vuyst, L. Technological and environmental features determine the uniqueness of the lambic beer microbiota and production process. Appl. Environment. Microbiol. 2021, 87, e00612–21. [Google Scholar] [CrossRef] [PubMed]
- Bossaert, S.; Crauwels, S.; Lievens, B.; De Rouck, G. The power of sour—A review: Old traditions, new opportunities. BrSc. 2019, 72, 78–88. [Google Scholar]
- Hoffman, C.S. , Winston, F. A ten-minute DNA preparation from yeast efficiently releases autonomous plasmids for transformaion of Escherichia coli. Gene 1987, 57, 267–272. [Google Scholar] [CrossRef]
- Gala, E.; Landi, S.; Solieri, L.; Nocetti, M.; Pulvirenti, A.; Giudici, P. Diversity of lactic acid bacteria population in ripened Parmigiano Reggiano cheese. Int. J. Food Microbiol. 2008, 125, 347–351. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Esteve-Zarzoso, B.; Belloch, C.; Uruburu, F.; Querol, A. Identification of yeasts by RFLP analysis of the 5.8 S rRNA gene and the two ribosomal internal transcribed spacers. Int. J. Syst. Evol. Microbiol. 1999, 49, 329–337. [Google Scholar] [CrossRef]
- Muir, A.; Harrison, E.; Wheals, A. A multiplex set of species-specific primers for rapid identification of members of the genus Saccharomyces. FEMS Yeast Res. 2011, 11, 552–563. [Google Scholar] [CrossRef]
- Pengelly, R.J.; Wheals, A.E. Rapid identification of Saccharomyces eubayanus and its hybrids. " FEMS Yeast Res. 2013, 13, 156–161. [Google Scholar] [CrossRef]
- Martini, S.; Bonazzi, S.; Malorgio, I.; Pizzamiglio, V.; Tagliazucchi, D.; Solieri, L. Characterization of yeasts isolated from Parmigiano Reggiano cheese natural whey starter: from spoilage agents to potential cell factories for whey valorization. Microorganisms 2021, 9, 2288. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley & Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- O’Leary, N.A.; Wright, M.W.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D.; et al. Reference sequence (RefSeq) database at NCBI: Current status, taxonomic expansion, and functional annotation. Nucleic Acids Res. 2016, 4, D733–D745. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Ebers, J. Taxonomic parameters revisited: tarnished gold standards. Microbiol. Today 2006, 33, 152–155. [Google Scholar]
- Kurtzman, C.P.; Robnett, C.J. Identification and phylogeny of ascomycetous yeasts from analysis of nuclear large subunit (26S) ribosomal DNA partial sequences. Antonie Leeuwenhoek 1998, 73, 331–371. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids. Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N; Nei M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol., 1987, 4, 406–425. [Google Scholar]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 2, 256–259. [Google Scholar] [CrossRef]
- Versalovic, J.; Schneider, M.; De Bruijn, F.J.; Lupski, J.R. Genomic fingerprinting of bacteria using repetitive sequence-based polymerase chain reaction. Meth. Mol. Cell. Biol. 1994, 5, 25–40. [Google Scholar]
- Solieri, L.; Bianchi, A.; Giudici, P. Inventory of non starter lactic acid bacteria from ripened Parmigiano Reggiano cheese as assessed by a culture dependent multiphasic approach. Syst. Appl. Microbiol. 2012, 35, 270–277. [Google Scholar] [CrossRef]
- Dakal, T.C.; Solieri, L.; Giudici, P. Evaluation of fingerprinting techniques to assess genotype variation among Zygosaccharomyces strains. Food Microbiol. 2018, 72, 135–145. [Google Scholar] [CrossRef]
- Legras, J.-L.; Karst, F. Optimisation of interdelta analysis for Saccharomyces cerevisiae strain characterisation. FEMS Microbiol. Lett. 2003, 221, 2249–2255. [Google Scholar] [CrossRef]
- Corte, L.; Lattanzi, M.; Buzzini, P.; Bolano, A.; Fatichenti, F.; Cardinali, G. Use of RAPD and killer toxin sensitivity in Saccharomyces cerevisiae strain typing. J. Appl. Microbiol. 2005, 9, 609–617. [Google Scholar] [CrossRef] [PubMed]
- Vigentini, I.; Barrera Cardenas, S.; Valdetara, F.; Faccincani, M.; Panont, C.A.; Picozzi, C.; Foschino, R. Use of native yeast strains for in-bottle fermentation to face the uniformity in sparkling wine production. Front. Microbiol. 2017, 8, 1225. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P., Fell; Robert, V. Methods for isolation, phenotypic characterization and maintenance of yeasts. In The yeasts, 5th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: Amsterdam, 2011; pp. 87–110. [Google Scholar]
- Catallo, M.; Iattici, F.; Randazzo, C.; Caggia, C.; Krogerus, K.; Magalhães, F.; Gibson, B.; Solieri, L. Hybridization of Saccharomyces cerevisiae Sourdough Strains with Cryotolerant Saccharomyces bayanus NBRC1948 as a Strategy to Increase Diversity of Strains Available for Lager Beer Fermentation. Microorganisms 2021, 9, 514. [Google Scholar] [CrossRef] [PubMed]
- Marongiu, A.; Zara, G.; Legras, J.-L.; Del Caro, A.; Mascia, I.; Fadda, C.; Budroni, M. Novel Starters for Old Processes: Use of Saccharomyces cerevisiae Strains Isolated from Artisanal Sourdough for Craft Beer Production at a Brewery Scale. J. Ind. Microbiol. Biotechnol. 2015, 42, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Kahm, M.; Hasenbrink, G.; Lichtenberg-Frate, H.; Ludwig, J.; Kschischo, M. Grofit: fitting biological growth curves with R. J. Stat. Softw. 2010, 33, 1–21. [Google Scholar] [CrossRef]
- Yamauchi, H.; Yamamoto, H.; Shibano, Y.; Amaya, N.; Saeki, T. Rapid methods for detecting Saccharomyces diastaticus, a beer spoilage yeast, using the polymerase chain reaction. J. Am. Soc. Brew. Chem. 1998, 56, 58–63. [Google Scholar]
- Krogerus, K.; Magalhães, F.; Kuivanen, J. A deletion in the STA1 promoter determines maltotriose and starch utilization in STA1+ Saccharomyces cerevisiae strains. Appl Microbiol Biotechnol. 2019, 103, 7597–7615. [Google Scholar] [CrossRef]
- Hammer, Øyvind, and David AT Harper. "Past: paleontological statistics software package for educaton and data anlysis." Palaeontologia electronica 4.1. 2001, 1.
- Bokulich, N.A.; Bamforth,C.W. The microbiology of malting and brewing. Microbiol. Mol. Biol. Rev. 2013, 77, 157–172. [CrossRef]
- De Roos, J.; Van der Veken, D.; De Vuyst, L. The interior surfaces of wooden barrels are an additional microbial inoculation source for lambic beer production. Appl. Environ. Microbiol. 2019, 85, e02226–18. [Google Scholar] [CrossRef]
- Caldwell, S.L.; McMahon, D.J.; Oberg, C.J. , Broadbent, J.R. Development and characterization of lactose-positive Pediococcus species for milk fermentation. Appl. Environ. Microbiol. 1996, 62, 936–941. [Google Scholar] [CrossRef]
- Spitaels, F.; Wieme, A.D.; Janssens, M.; Aerts, M.; Daniel, H-M. ; Van Landschoot, A.; De Vuyst, L.; Vandamme, P. The microbial diversity of traditional spontaneously fermented lambic beer. PLoS One, 2014, 9, e95384. [Google Scholar] [CrossRef] [PubMed]
- Rankine, B.C. Decomposition of L-malic acid by wine yeasts. J. Sci. Food Agric. 1966, 17, 312–316. [Google Scholar] [CrossRef] [PubMed]
- Saez, J.; Lopes, C.A; Kirs, V.E.; Sangorrín, M. Production of volatile phenols by Pichia manshurica and Pichia membranifaciens isolated from spoiled wines and cellar environment in Patagonia. Food Microbiol. 2011, 28, 503–509. [Google Scholar] [CrossRef]
- Wang, X.; Ren, X.; Shao, Q.; Peng, X.; Zou, W.; Sun, Z.; Zhang, L.; Li, H. Transformation of Microbial Negative Correlations into Positive Correlations by Saccharomyces cerevisiae Inoculation during Pomegranate Wine Fermentation. Appl. Environ. Microbiol. 2020, 86, e01847–20. [Google Scholar] [CrossRef] [PubMed]
- Lachance, M.-A. Yeast Communities in a Natural Tequila Fermentation. Antonie Van Leeuwenhoek 1995, 68, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.L.; Vianna, C.R.; Cadete. R.M.; Santos, R.O.; Gomes, F.C.; Oliveira, E.S.; Rosa, C.A. Selection, growth, and chemo-sensory evaluation of flocculent starter culture strains of Saccharomyces cerevisiae in the large-scale production of traditional Brazilian cachaça. Int J Food Microbiol. 2009, 131, 203–210.
- Bossaert, S.; Kocijan, T.; Winne, V.; Schlich, J.; Herrera-Malaver, B.; Verstrepen, K.J.; Van Opstaele, F.; De Rouck, G.; Crauwels, S.; Lievens, B. Beer ethanol and iso-α-acid level affect microbial community establishment and beer chemistry throughout wood maturation of beer. Int J Food Microbiol. 2022, 374, 109724. [Google Scholar] [CrossRef]
- Tomičić, R.; Tomičić, Z.; Thaler, N.; Humar, M.; Raspor, P. Factors Influencing Adhesion of Bacteria Escherichia coli, Pseudomonas aeruginosa, Staphylococcus aureus and Yeast Pichia membranifaciens to Wooden Surfaces. Wood Sci. Technol. 2020, 54, 1663–1676. [Google Scholar] [CrossRef]
- Ponzoni, C.; Gasparetti, C.; Goretti, M.; Turchetti, B.; Pagnoni, U.M.; Cramarossa. M.R.; Forti, L.; Buzzini, P. Biotransformation of acyclic monoterpenoids by Debaryomyces sp., Kluyveromyces sp., and Pichia sp. strains of environmental origin. Chem Biodivers. 2008, 5, 471–83. [CrossRef]
- Vaughan-Martini, A.; Martini, A. Saccharomyces Meyen ex Reess (1870). In The yeasts 5th ed; Kurtzman, C.P.; Fell, J.W.; Boekhout, T., Eds.; Elsevier, Amsterdam, 2011, pp. 733-746.
- Peŕez-Través, L.; Lopes, C.A.; Querol, A.; Barrio, E. On the complexity of the Saccharomyces bayanus taxon: Hybridization and potential hybrid speciation. PLoS ONE 2014, 9, e93729. [Google Scholar] [CrossRef]
- Libkind, D.; Hittinger, C.T.; Valério, E.; Gonçalves, C.; Dover, J.; Johnston, M.; Gonçalves, P.; Sampaio, J.P. Microbe Domestication and the Identification of the Wild Genetic Stock of Lager-Brewing Yeast. Proc. Natl. Acad. Sci. USA 2011, 108, 14539–14544. [Google Scholar] [CrossRef] [PubMed]
- Naumov, G.I.; James, S.A.; Naumova, E.S.; Louis, E.J.; Roberts, I.N. Three new species in the Saccharomyces sensu stricto complex: Saccharomyces cariocanus, Saccharomyces kudriavzevii and Saccharomyces mikatae. Int. J. Sys. Evol. Microbiol. 2000, 39050, 1931–1942. [Google Scholar] [CrossRef] [PubMed]
- Sipiczki, M. Taxonomic and physiological diversity of Saccharomyces bayanus. Biodivers. Biotechnol. Wine Yeasts 2002, 53–69. [Google Scholar]
- Demuyter, C.; Lollier, M.; Legras, J.-L.; Le Jeune, C. Predominance of Saccharomyces uvarum during spontaneous alcoholic fermentation, for three consecutive years, in an Alsatian winery. J. Appl. Microbiol. 2004, 97, 1140–1148. [Google Scholar] [CrossRef]
- Coton, E.; Coton, M.; Levert, D.; Casaregola, S.; Sohier, D. Yeast Ecology in French Cider and Black Olive Natural Fermentations. Int. J. Food. Microbiol. 2006, 108, 130–135. [Google Scholar] [CrossRef]
- Valles, B.S.; Bedriñana, R.P.; Tascón, N.F.; Simón, A.Q.; Madrera, R.R. Yeast species associated with the spontaneous fermentation of cider. Food Microbiol. 2007, 24, 25–31. [Google Scholar] [CrossRef]
- Rodríguez ME, Perez-Traves L, Sangorrín MP, Barrio E, Querol A, Lopes CA. Saccharomyces uvarum is responsible for the traditional fermentation of apple chicha in Patagonia. FEMS Yeast Res. 2017, 17, fow109.
- Rainieri, S.; Kodama, Y.; Kaneko, Y.; Mikata, K.; Nakao, Y.; Ashikari, T. Pure and mixed genetic lines of Saccharomyces bayanus and Saccharomyces pastorianus and their contribution to the lager brewing strain genome. Appl. Environ. Microbiol. 2006, 72, 3968–3974. [Google Scholar] [CrossRef]
- Gibson, B.; Dahabieh, M.; Krogerus, K.; Jouhten, P.; Magalhães, F.; Pereira, R.; Siewers, V.; Vidgren, V. Adaptive Laboratory Evolution of Ale and Lager Yeasts for Improved Brewing Efficiency and Beer Quality. Annu. Rev. Food Sci. Technol. 2020, 11, 23–44. [Google Scholar] [CrossRef]
- Kurtzman, C.P. Pichia EC Hansen (1904). In The yeasts, 5th ed; Kurtzman, C.P.; Fell, J.W.; Boekhout, T., Eds.; Elsevier, Amsterdam, 2011, pp. 273-352.
- Gallone, B.; Steensels, J.; Prahl, T.; Soriaga, L.; Saels, V.; Herrera-Malaver, B.; Merlevede, A.; Roncoroni, M.; Voordeckers, K.; Miraglia, L.; et al. Domestication and Divergence of Saccharomyces cerevisiae Beer Yeasts. Cell 2016, 166, 1397–1410. [Google Scholar] [CrossRef]
- Steensels, J.; Daenen, L.; Malcorps, P.; Derdelinckx, G.; Verachtert, H.; Verstrepen, K.J. Brettanomyces Yeasts—From Spoilage Organisms to Valuable Contributors to Industrial Fermentations. Int. J. Food Microbiol. 2015, 206, 24–38. [Google Scholar] [CrossRef]
- Verstrepen, K.J.; Derdelinckx, G.; Dufour, J.-P.; Winderickx, J.; Thevelein, J.M.; Pretorius, I.S.; Delvaux, F.R. Flavor-Active Esters: Adding Fruitiness to Beer. J. Biosci. Bioeng. 2003, 96, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Saerens, S.; Delvaux, F.; Verstrepen, K.; Van Dijck, P.; Thevelein, J.; Delvaux, F. Parameters Affecting Ethyl Ester Production by Saccharomyces cerevisiae during Fermentation. Appl. Environ. Microbiol. 2008, 74, 454–461. [Google Scholar] [CrossRef] [PubMed]
- Johansson, L.; Nikulin, J.; Juvonen, R.; Krogerus, K.; Magalhães, F.; Mikkelson, A.; Nuppunen-Puputti, M.; Sohlberg, E.; de Francesco, G.; Perretti, G.; et al. Sourdough Cultures as Reservoirs of Maltose-Negative Yeasts for Low-Alcohol Beer Brewing. Food Microbiol. 2021, 94, 103629. [Google Scholar] [CrossRef]
- Krogerus, K.; Eerikainen, R.; Aisala, H.; Gibson, B. Repurposing brewery contaminant yeast as production strains for low-alcohol beer fermentation. Yeast 2022, 39, 156–169. [Google Scholar] [CrossRef] [PubMed]
- De Keersmaecker, J. The Mystery of Lambic Beer. Sci. Am. 1996, 275, 74–80. [Google Scholar] [CrossRef]
- Piskur, J.; Rozpedowska, E.; Polakova, S.; Merico, A.; Compagno, C. How Did Saccharomyces Evolve to Become a Good Brewer? Trends Genet. 2006, 22, 183–186. [Google Scholar] [CrossRef]
- Shayevitz, A.; Harrison, K.; Curtin, C.D. Barrel-Induced Variation in the Microbiome and Mycobiome of Aged Sour Ale and Imperial Porter Beer. J. Am. Soc. Brew. Chem. 2021, 79, 33–40. [Google Scholar] [CrossRef]
- Sampaio, J.P.; Gonçalves, P. Natural Populations of Saccharomyces kudriavzevii in Portugal Are Associated with Oak Bark and Are Sympatric with S. cerevisiae and S. paradoxus. Appl. Environ. Microbiol. 2008, 74, 2144–2152. [Google Scholar] [CrossRef]
- Paraíso, F.; Pontes, A.; Neves, J.; Lebani, K.; Hutzler, M.; Zhou, N.; Sampaio, J.P. Do microbes evade domestication?-Evaluating potential ferality among diastatic Saccharomyces cerevisiae. Food Microbiol. 2023, 7, 104320. [Google Scholar] [CrossRef]
- Coelho, E.; Azevedo, M.; Teixeira, J.A.; Tavares, T.; Oliveira, J.M.; Domingues, L. Evaluation of multi-starter S. cerevisiae/D. bruxellensis cultures for mimicking and accelerating transformations occurring during barrel ageing of beer. Food Chem. 2020, 323, 126826. [Google Scholar] [CrossRef] [PubMed]
- Rozpędowska, E.; Hellborg, L.; Ishchuk, O.P.; Orhan, F.; Galafassi, S.; Merico, A.; Woolfit, M.; Compagno, C.; Piskur, J. Parallel evolution of the make-accumulate-consume strategy in Saccharomyces and Dekkera yeasts. Nat. Commun. 2011, 2, 302. [Google Scholar] [CrossRef] [PubMed]
- Meier-Dörnberg T, Kory OI, Jacob F, Michel M, Hutzler M. Saccharomyces cerevisiae variety diastaticus friend or foe?-spoilage potential and brewing ability of different Saccharomyces cerevisiae variety diastaticus yeast isolates by genetic, phenotypic and physiological characterization. FEMS Yeast Res. 2018, 18, foy023.
- Peris, D.; Pérez-Torrado, R.; Hittinger, C.T.; Barrio, E.; Querol, A. On the origins and industrial applications of Saccharomyces cerevisiae × Saccharomyces kudriavzevii hybrids. Yeast 2018, 35, 51–69. [Google Scholar] [CrossRef] [PubMed]
- Preiss, R.; Tyrawa, C.; Krogerus, K.; Garshol, L.M.; van der Merwe, G. Traditional Norwegian kveik are a genetically distinct group of domesticated Saccharomyces cerevisiae brewing yeasts. Front. Microbiol. 2018, 9, 2137. [Google Scholar] [CrossRef] [PubMed]
- Krogerus, K.; Preiss, R.; Gibson, B. A Unique Saccharomyces cerevisiae × Saccharomyces uvarum Hybrid Isolated From Norwegian Farmhouse Beer: Characterization and Reconstruction. Front Microbiol. 2018, 9, 2253. [Google Scholar] [CrossRef]
- Liu, T.; Li, Y.; Sadiq, F.A.; Yang, H.; Gu, J.; Yuan, L.; Lee, Y.K.; He, G. Predominant yeasts in Chinese traditional sourdough and their influence on aroma formation in Chinese steamed bread. Food Chem. 2018, 242, 404–411. [Google Scholar] [CrossRef]
Figure 1.
Flow-chart of semi-spontaneous craft beer fermentation. Red arrow indicates point of sampling.
Figure 1.
Flow-chart of semi-spontaneous craft beer fermentation. Red arrow indicates point of sampling.
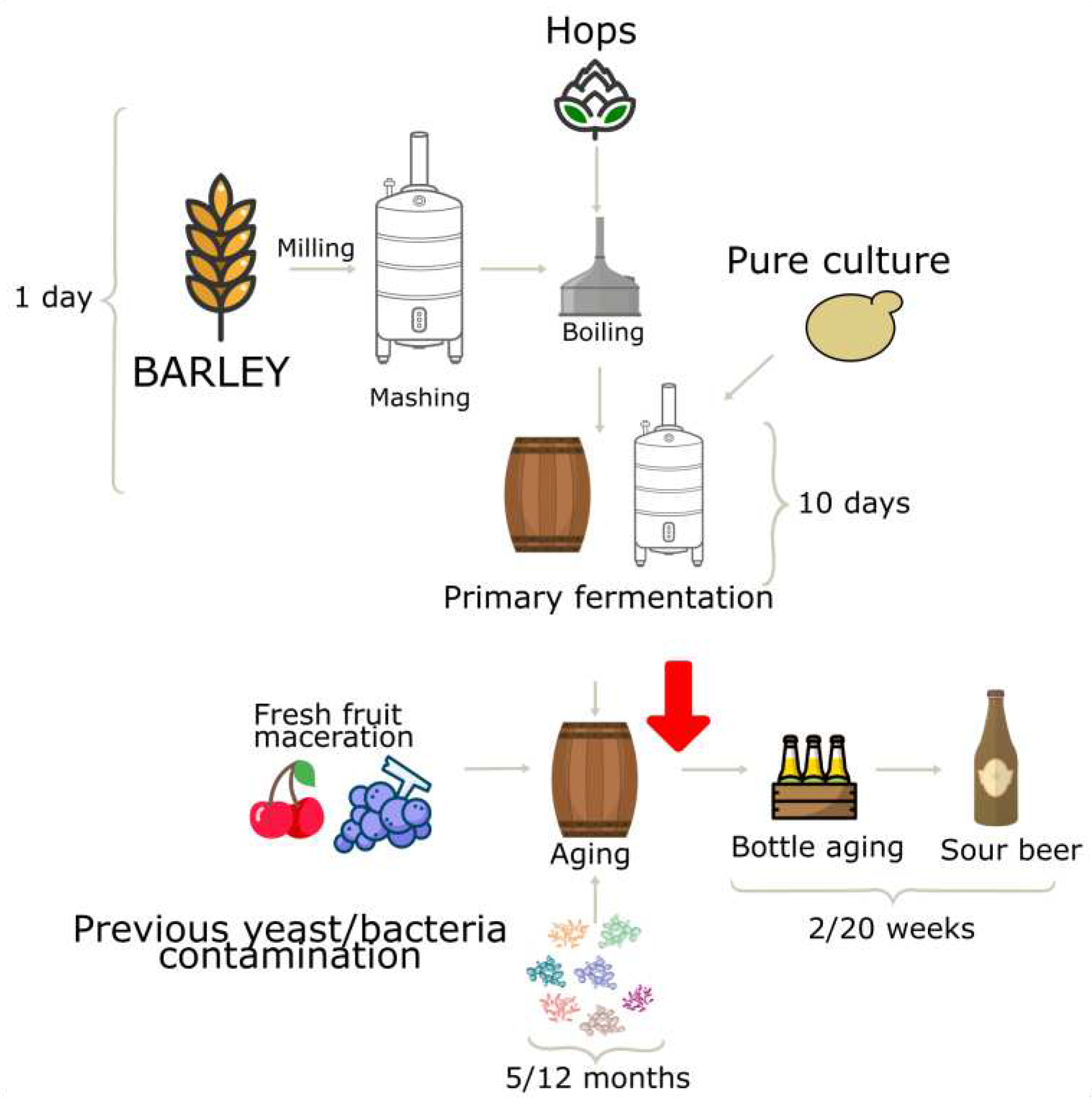
Figure 2.
Phylogenetic and genotyping analysis of LAB isolates. (A) Neighbor-joining tree based on 16S rDNA sequences showing genetic relatedness between Pediococcus damnosus isolates and related species. The evolutionary distances were computed using the Kimura 2-parameter method and the rate variation among sites was modeled with a gamma distribution. Bootstrap values are indicated beside branches (> 50%). The tree was rooted with P. acidalictici and P. pentosaceous (blue). (B) UPGMA clustering of LAB isolates based on (GTG)5 rep-PCR fingerprinting analysis. Similarities were calculated as Pearson correlation coefficient with Bionumerics software. The tree data (Newick) were exported and visualized using ITOL [38].
Figure 2.
Phylogenetic and genotyping analysis of LAB isolates. (A) Neighbor-joining tree based on 16S rDNA sequences showing genetic relatedness between Pediococcus damnosus isolates and related species. The evolutionary distances were computed using the Kimura 2-parameter method and the rate variation among sites was modeled with a gamma distribution. Bootstrap values are indicated beside branches (> 50%). The tree was rooted with P. acidalictici and P. pentosaceous (blue). (B) UPGMA clustering of LAB isolates based on (GTG)5 rep-PCR fingerprinting analysis. Similarities were calculated as Pearson correlation coefficient with Bionumerics software. The tree data (Newick) were exported and visualized using ITOL [38].
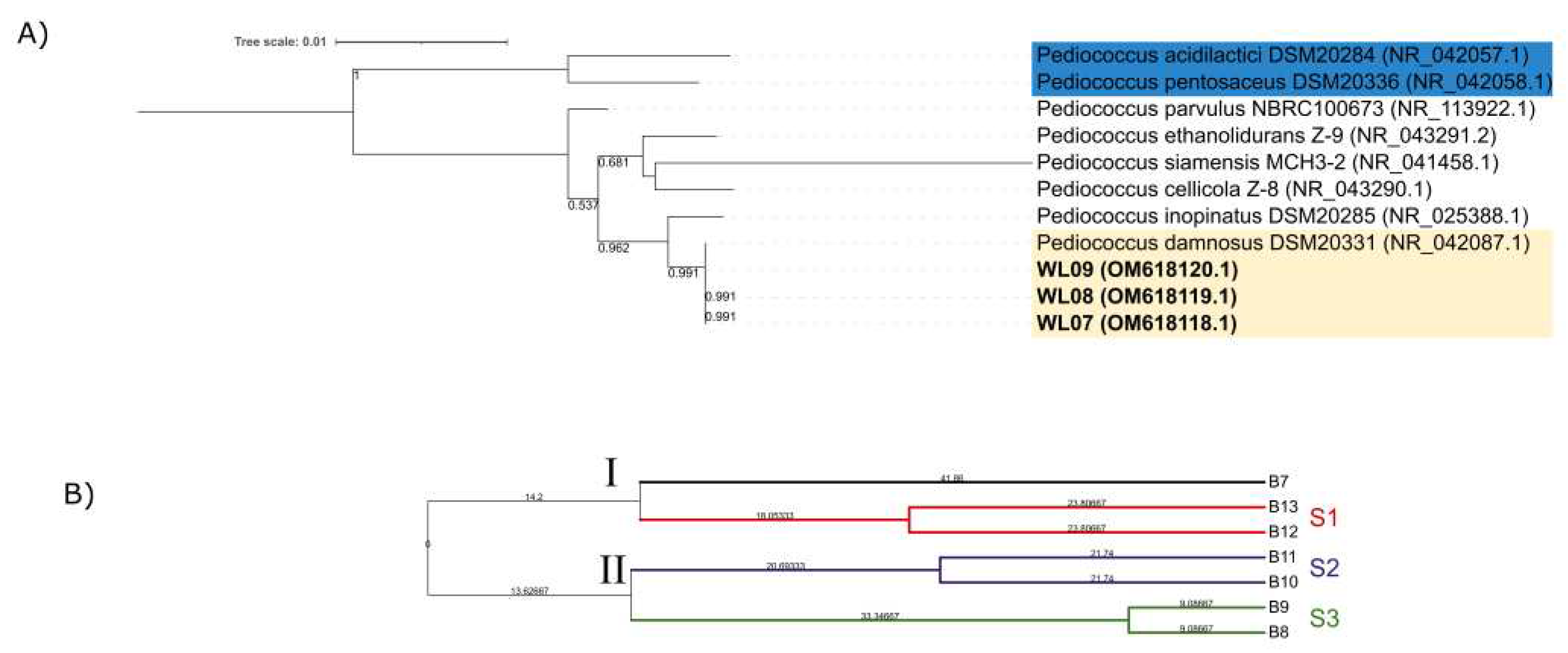
Figure 3.
Species attribution of yeast isolates (in bold). (A) Phylogenetic tree obtained by neighbor joining (NJ) method applied to a dataset of 28 D1/D2 26S rDNA sequencesThe evolutionary distances were calculated by the Tajima-Nei method. The gamma distribution was used to model the rate of change between sites. The lengths of the branches are proportional to the number of nucleotide substitutions, and they have been measured using the divergence scale shown at the top left. Bootstrap values (1000 replicates) are indicated beside branches (> 50%). The tree was rooted with Ogataea polymorpha as outgroup. The tree data (Newick) were exported and visualized using ITOL [38]. (B) Pie-chart representing species abundance in sour beer. Pink, S. cerevisiae; blue, P. membranifaciens; green, S. cerevisiae x S. uvarum hybrids; yellow, D. anomala; purple, D. bruxellensis.
Figure 3.
Species attribution of yeast isolates (in bold). (A) Phylogenetic tree obtained by neighbor joining (NJ) method applied to a dataset of 28 D1/D2 26S rDNA sequencesThe evolutionary distances were calculated by the Tajima-Nei method. The gamma distribution was used to model the rate of change between sites. The lengths of the branches are proportional to the number of nucleotide substitutions, and they have been measured using the divergence scale shown at the top left. Bootstrap values (1000 replicates) are indicated beside branches (> 50%). The tree was rooted with Ogataea polymorpha as outgroup. The tree data (Newick) were exported and visualized using ITOL [38]. (B) Pie-chart representing species abundance in sour beer. Pink, S. cerevisiae; blue, P. membranifaciens; green, S. cerevisiae x S. uvarum hybrids; yellow, D. anomala; purple, D. bruxellensis.
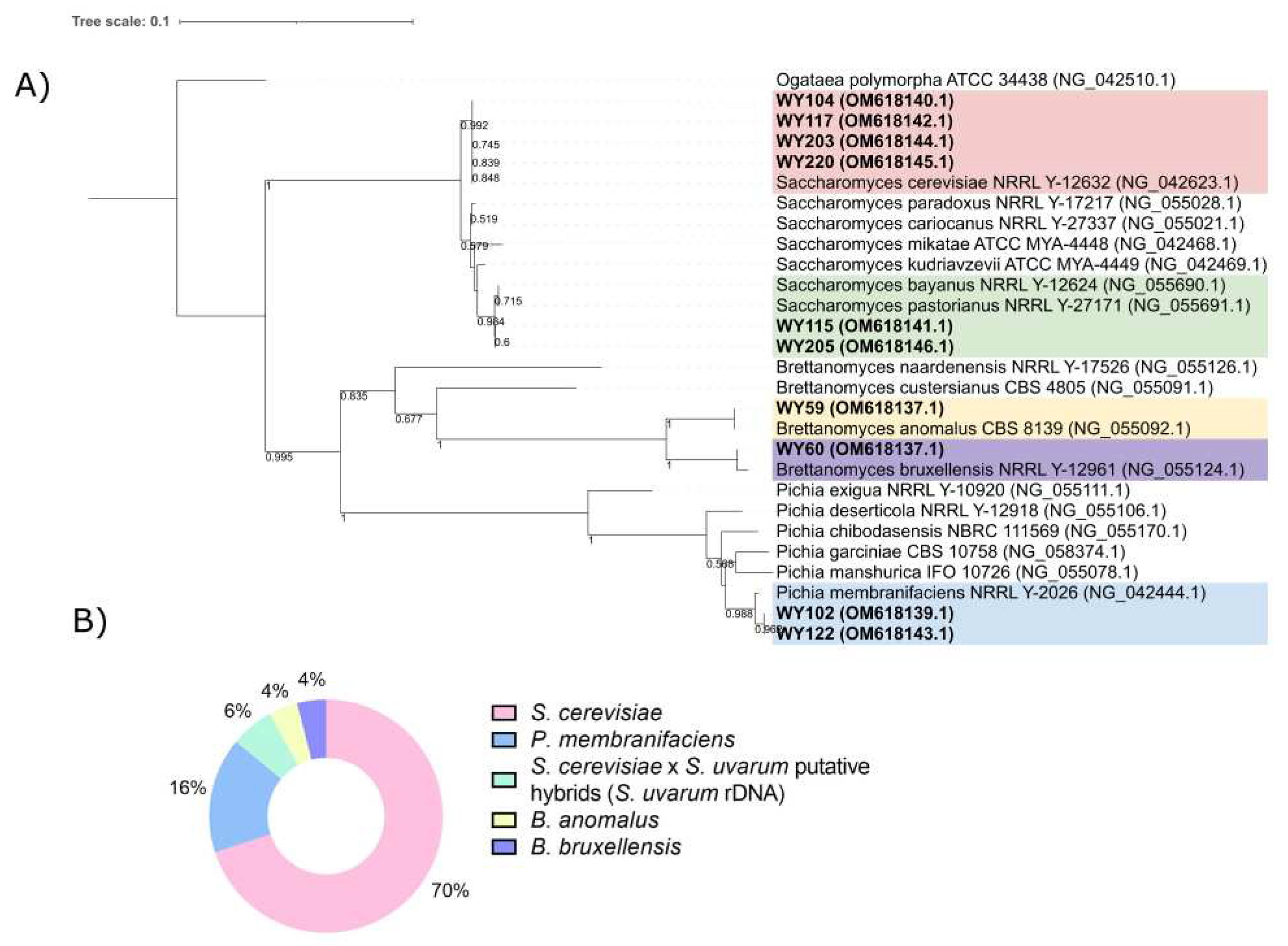
Figure 4.
Characterization of Saccharomyces strains by species-specific PCR assays. (A) Illustration of loos of heterozygosity (LOH) occurring in rDNA arrays of hybrid lineages. (B) S. cerevisiae-specific and S. bayanus/S. uvarum-specific PCR assays with Scer_F2/Scer_R2 primers targeting MEX67 gene (amplicon size 150 bp) and primers Sbay_F1/Sbay_R1 targeting DBP6 gene (amplicon size 275 bp), respectively. All tested strains were negative to PCRs targeting S. eubayanus-specific FSY1 gene (amplicon size 228 bp) (data not shown). S. cerevisiae BY4743 and S. uvarum PJP9 were used as internal control. S. cerevisiae strains WY117, WY104, WY203, and WY220 were selected as representative of pattern A. Abbreviations: M, molecular weight marker; S. cer, S. cerevisiae; Sbay, S. uvarum/S. bayanus; H. hybrid; LOH, loss of heterozygosity.
Figure 4.
Characterization of Saccharomyces strains by species-specific PCR assays. (A) Illustration of loos of heterozygosity (LOH) occurring in rDNA arrays of hybrid lineages. (B) S. cerevisiae-specific and S. bayanus/S. uvarum-specific PCR assays with Scer_F2/Scer_R2 primers targeting MEX67 gene (amplicon size 150 bp) and primers Sbay_F1/Sbay_R1 targeting DBP6 gene (amplicon size 275 bp), respectively. All tested strains were negative to PCRs targeting S. eubayanus-specific FSY1 gene (amplicon size 228 bp) (data not shown). S. cerevisiae BY4743 and S. uvarum PJP9 were used as internal control. S. cerevisiae strains WY117, WY104, WY203, and WY220 were selected as representative of pattern A. Abbreviations: M, molecular weight marker; S. cer, S. cerevisiae; Sbay, S. uvarum/S. bayanus; H. hybrid; LOH, loss of heterozygosity.
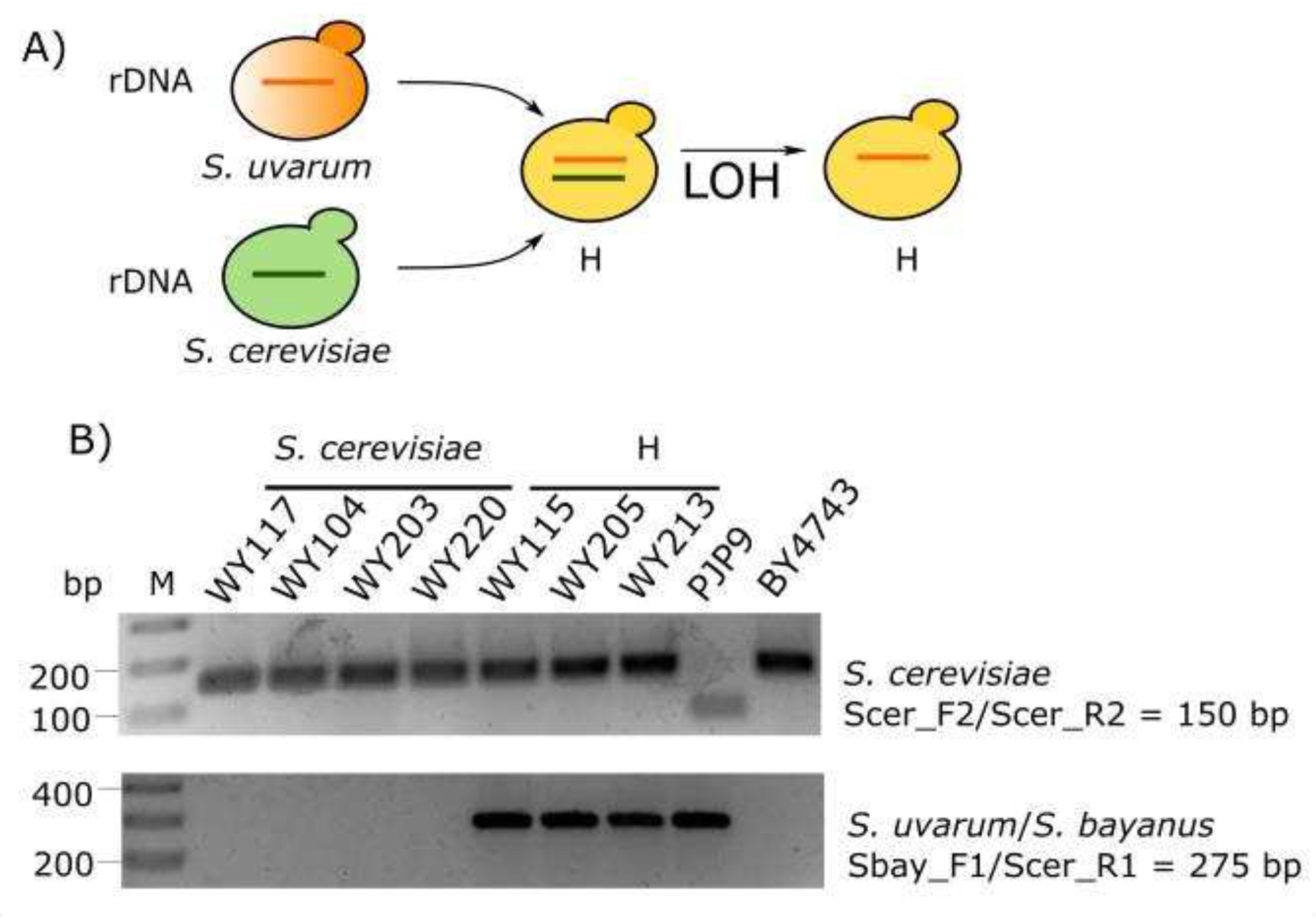
Figure 5.
Dendrogram generated from (GTG)5 rep-PCR fingerprints of 50 sour beer wild yeasts. Similarity percentages were calculated using Pearson correlation coefficient, while hierarchical clustering analysis was carried out using the UPGMA (unweighted pair-group method with arithmetic mean) method with Bionumerics software. The tree data (Newick) were exported and visualized using ITOL [38]. The similarity value above 92% was used to discriminate biotypes numbered from S1 to S9 and singletons. The color codes were attributed as follows: pink, S. cerevisiae; blue, P. membranifaciens; green, S. cerevisiae x S. uvarum hybrids; yellow, D. anomala; purple, D. bruxellensis.
Figure 5.
Dendrogram generated from (GTG)5 rep-PCR fingerprints of 50 sour beer wild yeasts. Similarity percentages were calculated using Pearson correlation coefficient, while hierarchical clustering analysis was carried out using the UPGMA (unweighted pair-group method with arithmetic mean) method with Bionumerics software. The tree data (Newick) were exported and visualized using ITOL [38]. The similarity value above 92% was used to discriminate biotypes numbered from S1 to S9 and singletons. The color codes were attributed as follows: pink, S. cerevisiae; blue, P. membranifaciens; green, S. cerevisiae x S. uvarum hybrids; yellow, D. anomala; purple, D. bruxellensis.
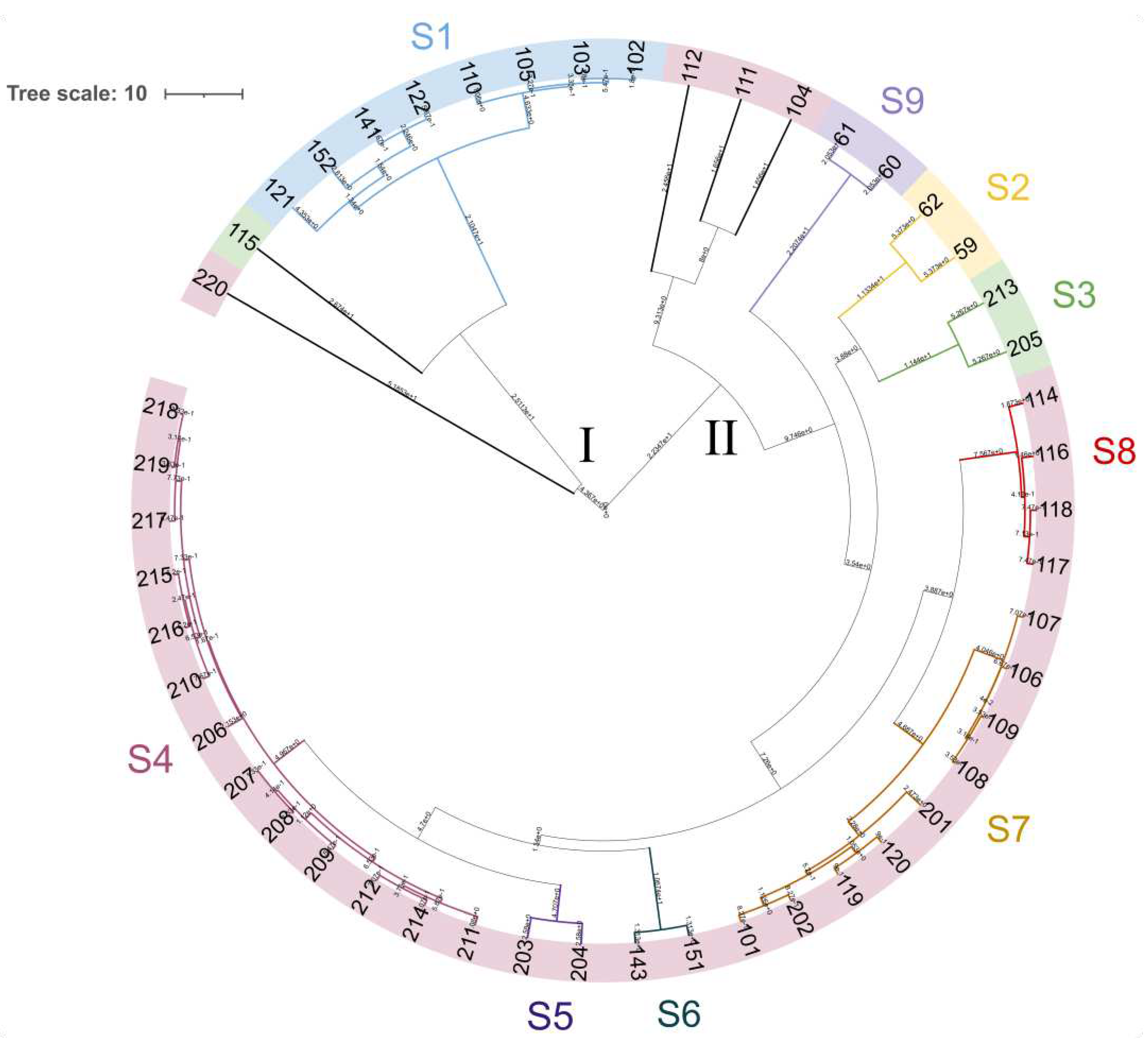
Figure 6.
Maltose (green) and glucose (grey) consumption tests. Statistical differences are indicated with different letters (two-way ANOVA; p<0.05). Abbreviation: H, S. cerevisiae x S. uvarum hybrid.
Figure 6.
Maltose (green) and glucose (grey) consumption tests. Statistical differences are indicated with different letters (two-way ANOVA; p<0.05). Abbreviation: H, S. cerevisiae x S. uvarum hybrid.
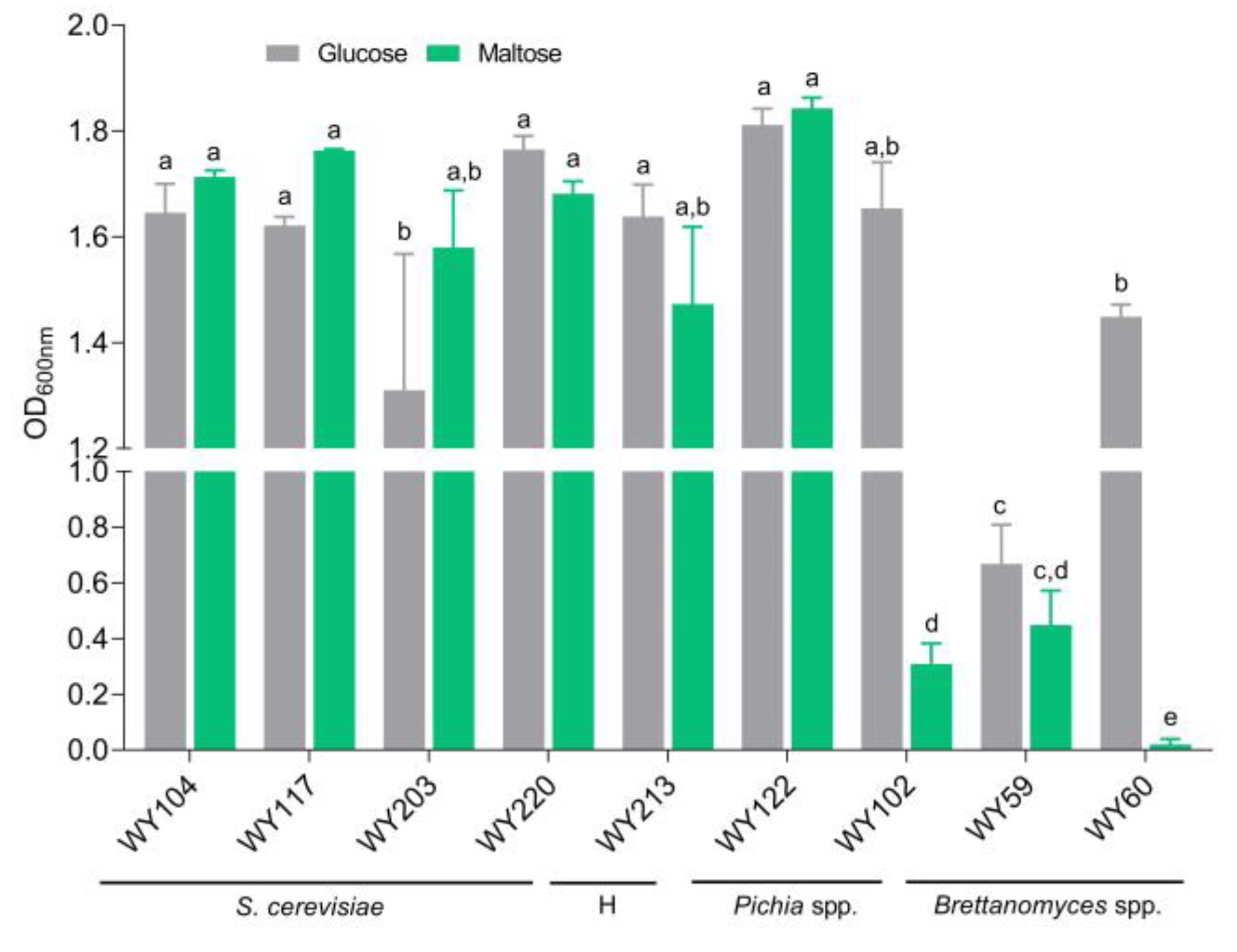
Figure 7.
Kinetics parameters (lag phase λ, fermentation rate μ, and maximum fermentation efficiency) of yeast candidates in micro-scale wort fermentation (11.5 Plato; 20 °C). Species was represented as follows: pink, S. cerevisiae; blue, P. membranifaciens; green, S. cerevisiae x S. uvarum hybrids; yellow, B. anomalus; purple, B. bruxellensis.
Figure 7.
Kinetics parameters (lag phase λ, fermentation rate μ, and maximum fermentation efficiency) of yeast candidates in micro-scale wort fermentation (11.5 Plato; 20 °C). Species was represented as follows: pink, S. cerevisiae; blue, P. membranifaciens; green, S. cerevisiae x S. uvarum hybrids; yellow, B. anomalus; purple, B. bruxellensis.
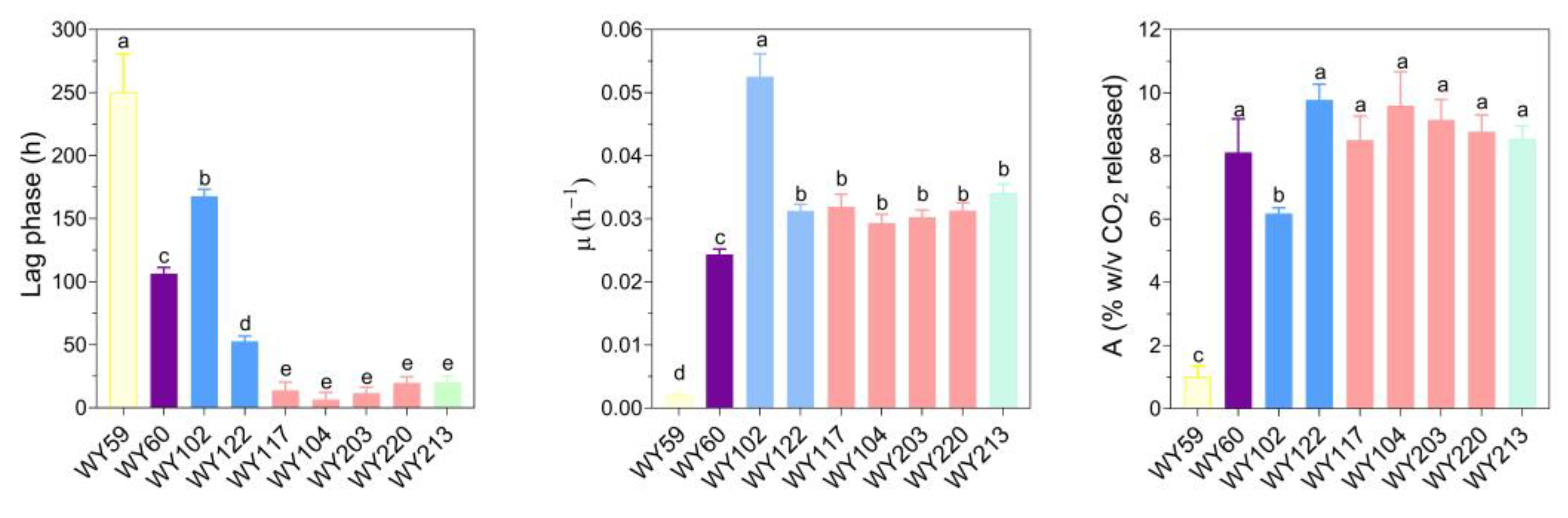
Figure 8.
HPLC analysis of microscale fermentation broths inoculated with selected yeast strains. % consumption of maltose (yellow), maltotriose (light orange) and maltodextrins (orange); ethanol formation (g/L; red). Hybrid: S. cerevisiae x S. uvarum hybrid.
Figure 8.
HPLC analysis of microscale fermentation broths inoculated with selected yeast strains. % consumption of maltose (yellow), maltotriose (light orange) and maltodextrins (orange); ethanol formation (g/L; red). Hybrid: S. cerevisiae x S. uvarum hybrid.
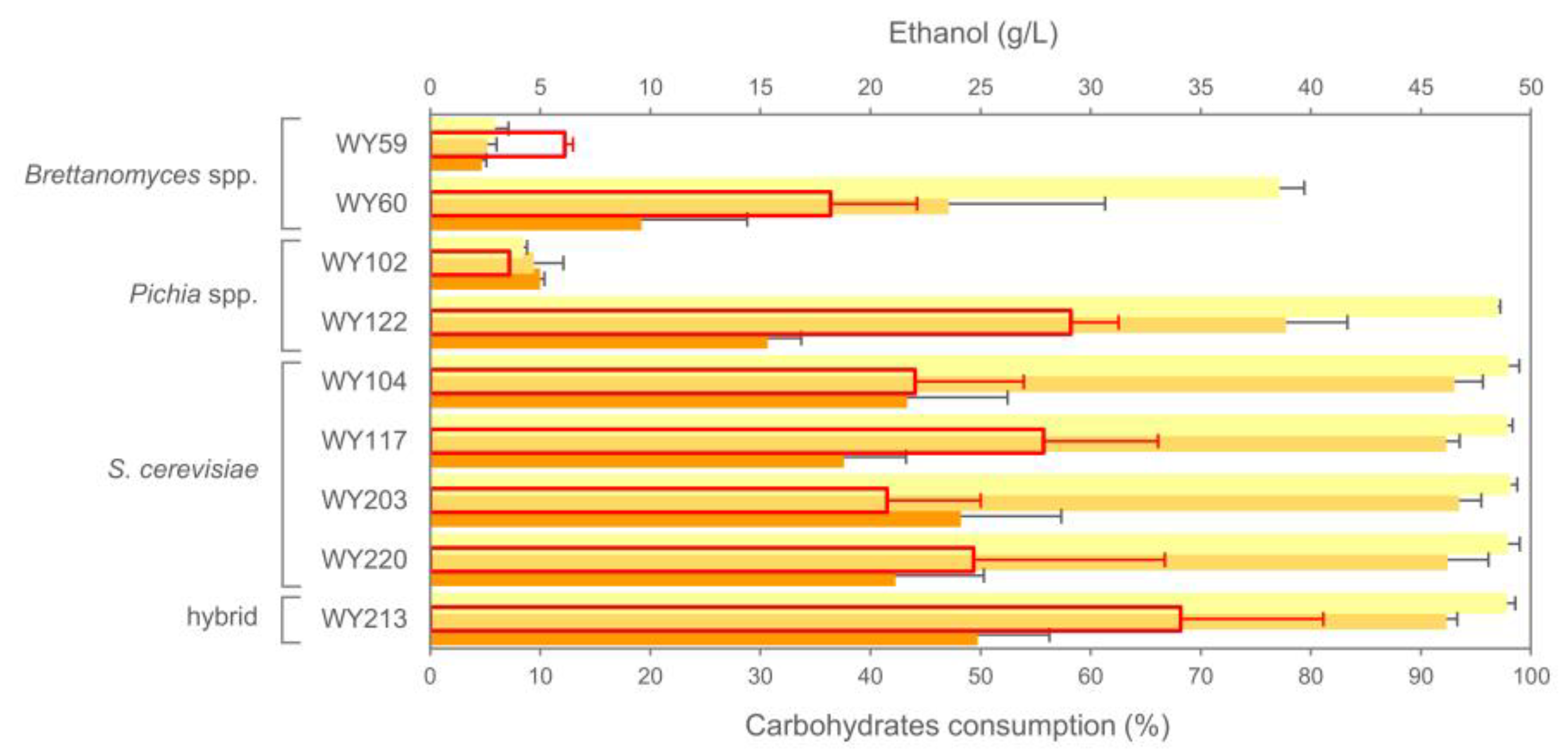
Figure 9.
Screening for STA1 gene and the related promoter in S. cerevisiae sour beer strains. (A) Strategy to PCR amplify STA1 open reading frame and the promoter region with UAS_1 and UAS_2 located upstream the activation sequence [50]. (B) Individual PCR reactions using primers SD-5A/SD-6B with 3711 as reference strain with “diastatic” phenotype (STA1+, no deletion in STA1 promoter) and WLP940 (STA1−) as reference strain with no “diastatic” phenotype. (C) Individual PCR reactions using primers STA1_UAS_Fw/STA1_UAS_Rv with 3711 as “diastatic” reference strain (STA1+, no deletion in STA1 promoter) and WLP940 reference strain with no “diastatic” phenotype (STA1−) as no “diastatic” control. (D) Sequence alignment of UAS2-2 region of WY117 and 3711. Promoter region of diastatic control strain 3711 was retrieved from [50].
Figure 9.
Screening for STA1 gene and the related promoter in S. cerevisiae sour beer strains. (A) Strategy to PCR amplify STA1 open reading frame and the promoter region with UAS_1 and UAS_2 located upstream the activation sequence [50]. (B) Individual PCR reactions using primers SD-5A/SD-6B with 3711 as reference strain with “diastatic” phenotype (STA1+, no deletion in STA1 promoter) and WLP940 (STA1−) as reference strain with no “diastatic” phenotype. (C) Individual PCR reactions using primers STA1_UAS_Fw/STA1_UAS_Rv with 3711 as “diastatic” reference strain (STA1+, no deletion in STA1 promoter) and WLP940 reference strain with no “diastatic” phenotype (STA1−) as no “diastatic” control. (D) Sequence alignment of UAS2-2 region of WY117 and 3711. Promoter region of diastatic control strain 3711 was retrieved from [50].
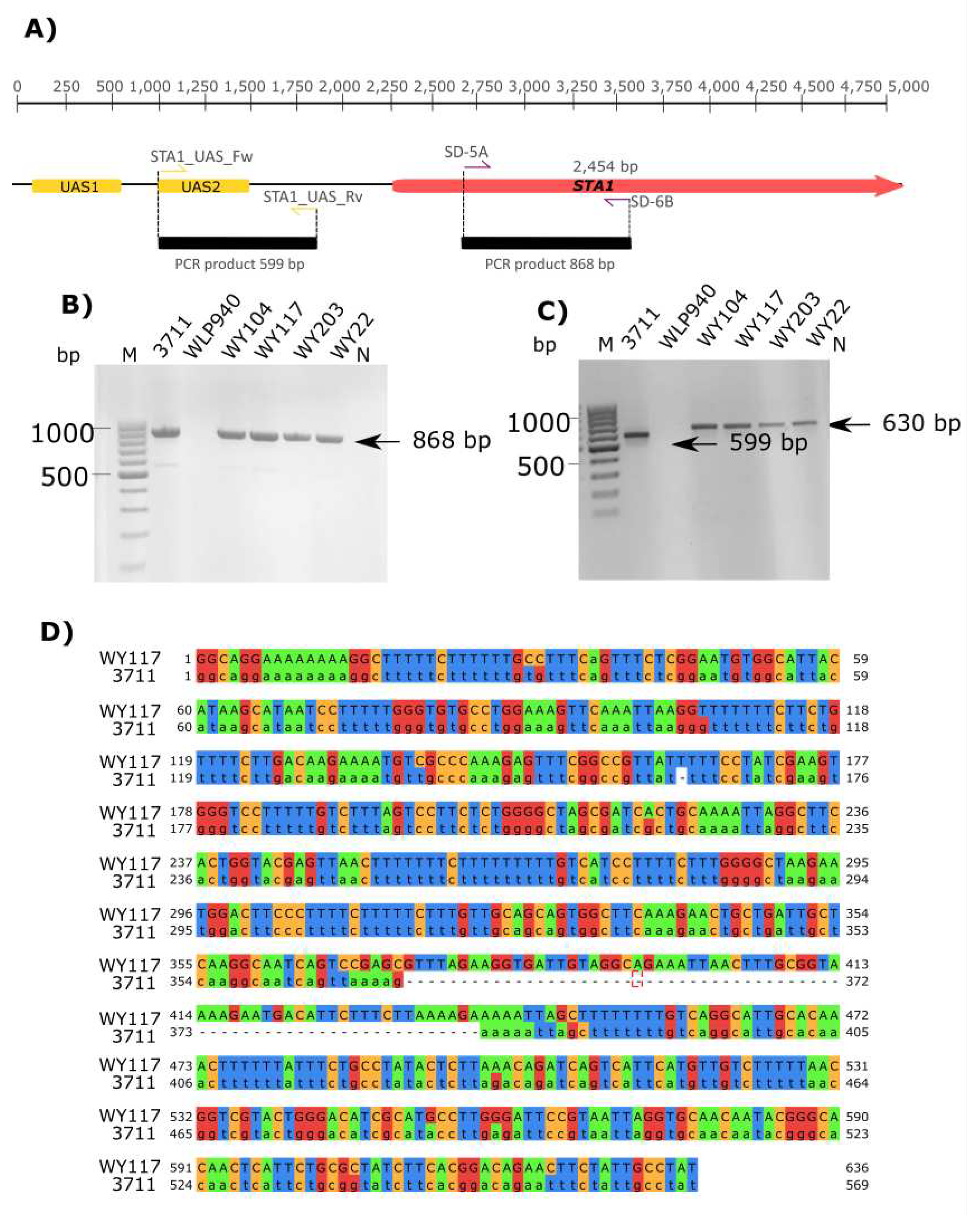
Figure 10.
VOCs profiles of wort and micro-fermented samples reconstructed by headspace SPME-GC-MS analysis, followed by principal component analysis (PCA). (A) Scores plot (PC1 vs. PC2) accounting for 61.5% and 21.1% of variability, respectively. (B) Loading plot of the volatile molecules. The compounds with the highest loading value in PC1 and PC2 are labeled. A and B were replicated samples.
Figure 10.
VOCs profiles of wort and micro-fermented samples reconstructed by headspace SPME-GC-MS analysis, followed by principal component analysis (PCA). (A) Scores plot (PC1 vs. PC2) accounting for 61.5% and 21.1% of variability, respectively. (B) Loading plot of the volatile molecules. The compounds with the highest loading value in PC1 and PC2 are labeled. A and B were replicated samples.
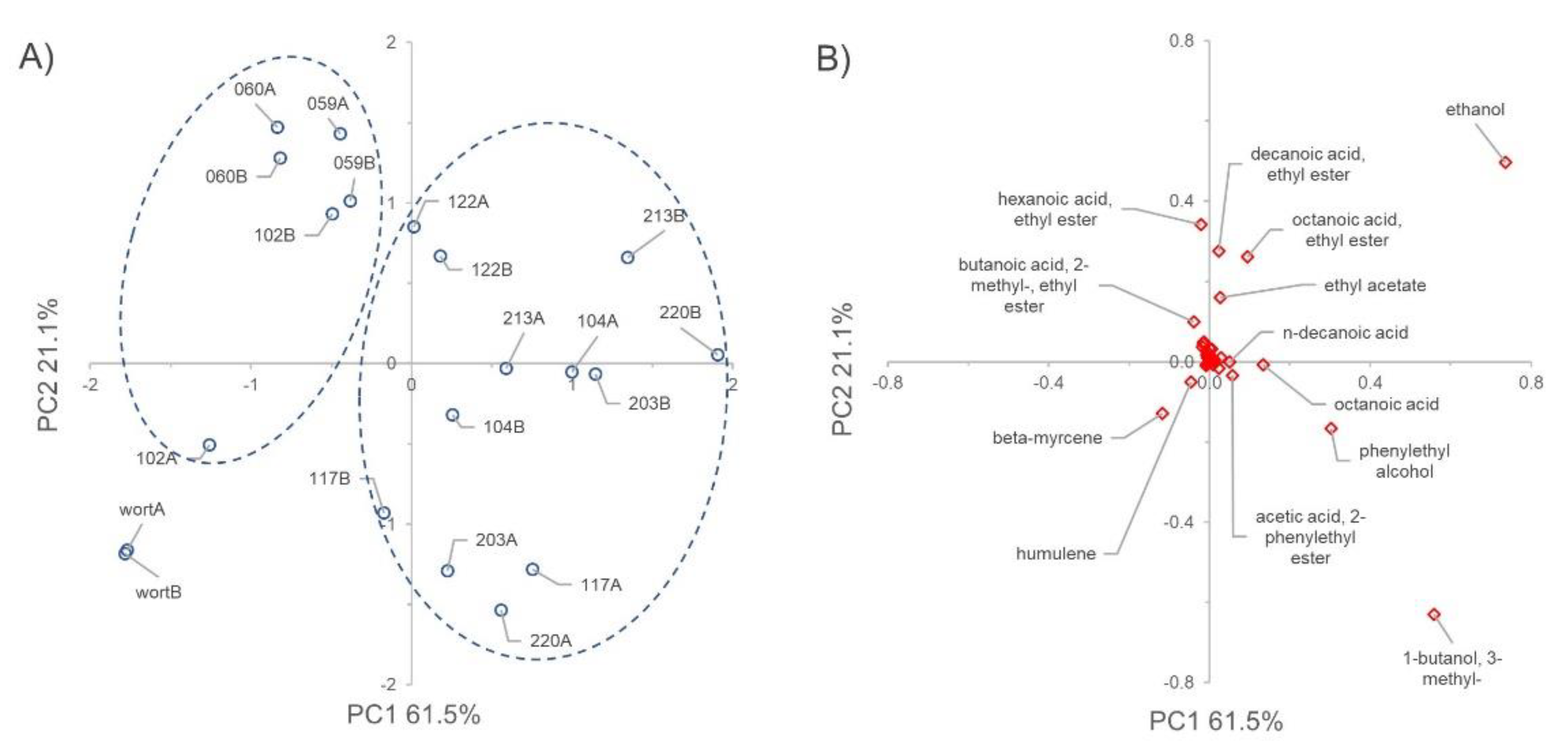

Table 1.
Reference strains used in this work.
| Strain | Species | Features | Source |
|---|---|---|---|
| BY4742 | S. cerevisiae | MATα; his3Δ1; leu2Δ0; lys2Δ0; ura3Δ0 | Euroscarf |
| BY4743 | S. cerevisiae | MATa/α; his3Δ1/ his3Δ1; leu2Δ0/ leu2Δ0; met15Δ0/MET15; LYS2/lys2Δ0; ura3Δ0/ura3Δ0 | Euroscarf |
| 3711 | S. cerevisiae var. diastaticus | STA1 positive French Saison ale yeast | Wyeast |
| WLP940 | Lager | STA1 negative lager yeast | White Lab |
| Atecrem | S. cerevisiae | Atecrem Belgian Saison | Bioenologia 2.0 |
| WLP565 | S. cerevisiae | Belgian Saison I Ale Yeast | White Lab |
| 3724 | S. cerevisiae var. diastaticus | Belgian Saison Yeast | Wyeast |
| PJP9 | S. uvarum/S. bayanus | nd | Gift from P. Marullo |
Abbreviation: nd, not determined.
Table 2.
Sporulation efficiency and spore viability of Saccharomyces wild sour beer strains. Abbreviation: nd, not determined.
Table 2.
Sporulation efficiency and spore viability of Saccharomyces wild sour beer strains. Abbreviation: nd, not determined.
| Species | Strains | Sporulation efficiency (%) | Spore viability (%) | MAT loci | |
|---|---|---|---|---|---|
| 3 d | 14 d | ||||
| S. cerevisiae | WY104 | 22.96 | 24.56 | 75 | MATa/MATα |
| WY117 | 23.84 | 32.72 | 75 | MATa/MATα | |
| WY203 | 22.70 | 27.85 | 81 | MATa/MATα | |
| WY220 | 22.78 | 32.13 | 56 | MATa/MATα | |
| hybrids | WY205 | 1.22 | 0.89 | nd | nd |
| WY213 | 2.48 | 0.41 | nd | nd | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



