
Submitted:
09 September 2024
Posted:
09 September 2024
You are already at the latest version
Abstract
This study identified the effects of substituting fishmeal (FM) with soy protein concentrate (SPC) supplemented with lysine and methionine in the olive flounder (Paralichthys olivaceus) diet on the expression of genes related to growth, stress, immunity, and digestion. Three experimental diets were prepared; the control diet (Con) contained 60% FM, and 25% (SPC25) and 50% (SPC50) FM were replaced by SPC. After the 140-day feeding period, weight gain, specific growth rate, feed availability, biological indices and chemical composition of the muscle were not significantly affected by the dietary treatments. The gene expression levels of growth-related genes (IGF, GFB-3), immune-related genes (IL-8 and caspase), stress-related genes (SOD, GPX, PRX, TRX) were higher in Con fish than in those fed the SPC25 and SPC50 diets. Chymo-TRY2, TRY2 and TRY3 in the stomach of fish fed the SPC25 diet were higher than fish fed the Con or SPC50 diets. TRY2 and lipase of fish fed the SPC25 diet were higher than those of fish fed the other diets but TRY3 was not significantly affected by the treatments in the intestine. In conclusion, substituting 25% FM with SPC was the most effective for the expression of growth, immune, stress, and digestive enzyme genes.
Keywords:
fishmeal substitution
; soy protein concentrate
; Paralichthys olivaceus
; gene expression
; growth performance
; feed utilization
1. Introduction
Olive flounder (Paralichthys olivaceus) is a popular and commercially important marine fish species in the aquaculture industry in Republic of Korea (henceforth, Korea) [1,2], accounting for 50% of aquaculture production in 2023 [2]. Feed is an important component in olive flounder farming because it makes up 30–35% of total operational costs [3]. In particular, protein is an important nutrient in the diets of carnivorous fish species such as olive flounder [4]. Fish meal (FM) is used as the main protein source and 40–70% protein is supplied with FM in carnivorous fish feeds [4]. FM has advantages, such as a balanced amino acid (AA) profile and high nutrient digestibility [5,6]. However, its production has been decreasing or plateauing in recent decades due to overfishing of fish stocks and its price is rising due to increased global demand [5,7]. Therefore, it is important to search for a replacement that is inexpensive and available year-round.
Soybeans are less expensive than FM but contain non-starch polysaccharides, which are difficult for fish to digest, and high levels of anti-nutritional factors such as trypsin inhibitors [8,9]. Soy protein concentrate (SPC) is obtained after soybean meal is processed and is almost free of anti-nutritional factors [10]. It also has higher nutrient digestibility than soybean meal and can be used to replace a substantial amount of FM in fish feed [11,12,13]. Nevertheless, Lim et al. [14] reported that plant protein sources are more likely to lack some essential amino acids (EAA) and fatty acids (FAs) compared to FM, which would adversely affect fish growth. Therefore, substituting FM with a plant protein source is limited to some extent in fish feeds [15,16]. Many attempts have been made to replace FM with various plant protein sources [16,17]. Deng et al. [13] reported that the growth performance of olive flounder fed a diet replacing 75% FM with SPC and providing an AA supplement was similar to fish fed a diet replacing 25% FM with SPC without AA supplementation but inferior to fish fed a 74% FM-based diet.
Growth performance and feed efficiency of red sea bream (Pagrus major) fed the SPC diet with AA supplementation were similar or slightly lower than the values of fish fed a diet replacing FM with SPC and providing an AA supplement [18]. Kokou et al. [19] reported that supplementing with AAs improves all growth and protein conversion ratio up to the 40 substitution levels (based on 58% FM) in the gilthead seabream (Sparus aurata L.).
Previous studies have reported changes in the expression of growth and immune-related genes when FM is replaced with other ingredients, such as SPC. In Moon et al. [20], the expression of growth hormone-releasing hormone and insulin-like growth factor-I (IGF-I) genes changed in fish when 30% of FM was replaced with SPC based on 60% FM. In another study, as a result of replacing 14.5% with SPC based on 65% FM, the expression levels of the immune-related genes interleukin (IL)-10 and IL-1β were higher in a low FM diet and induced a more balanced immune response when the low FM diet was consumed by olive flounder [21].
Therefore, this study investigated the effect of substituting FM with SPC in the diet along with providing AA supplementation on the expression of growth, stress, immune, and digestive enzyme-related genes in P. olivaceus.
2. Materials and Methods
2.1. Experimental Conditions
Olive flounder (727.8±7.89 g; mean±SE) were purchased from the Genetics and Breeding Research Center (National Fisheries Research Institute, Geoje-si, Gyeongsangnam-do, Korea) and transported to the Feed Research Center (National Fisheries Research Institute, Pohang-si, Gyeongsangbuk-do, Korea). The fish had a 2 week acclimation period by being fed commercial feed (crude protein: 54.0% and crude lipid: 10.0%) (Suhyup Feed, Uiryeong-gun, Gyeongsangnam-do, Korea) under experimental conditions. After the 2-week acclimatization period, the fish were randomly distributed into nine 8-ton flow-through tanks (water volume: 5 tons) (100 fish/tank). The water source was sand-filtered seawater. The water temperature ranged from 16.5°C to 22.4°C (19.9±3.4°C; mean±SD) and oxygen was supplied directly to each tank by an oxygen generator. Dead fish were immediately removed from each tank after measuring length and weight throughout the feeding trial.
2.2. Experimental Diet Preparation
Three experimental isonitrogenous (53.0%) and isoenergetic (3.9 kcal/g diet) diets were formulated (Table 1) to meet the dietary protein and lipid requirements of olive flounder [22,23]. The control (Con) diet contained 60% FM. In this diet, 25% (SPC25) and 50% (SPC50) FM were substituted with SPC supplemented with lysine (0.375 and 0.750%) and methionine (0.145 and 0.290%, respectively) (MP Bio, Irvine, CA, USA), respectively.
The ingredients in the experimental diets were thoroughly blended with water at a ratio of 5:1. The pressure control of the experimental feed extruder (twin screw extruder (ATX-II, Fesco, Daegu, Korea) was controlled only by the screw speed (rpm/min), while keeping the barrel temperature (115–130°C), conditioner temperature (80°C), steam (31.6 kg/h), and feed rate (50 kg/h) constant (low pressure, 885 rpm/min; high pressure: high pressure, 708 rpm/min). The speed of the discs (rpm/min) and rotors (rotor, rpm/min) from the air classifier mill (XP 50, (C) KMTECH, Gyeonggi-do, Korea) were adjusted accordingly. All experimental diets were stored at −20°C until use. The experimental diets were fed to the fish twice daily (08:00 and 17:00) to apparent satiation for 140 days. The nutrient requirements of the fish were satisfied by all of the experimental diets [22,23].

Table 1.
Ingredients and proximate composition of the experimental diets (%, DM basis).
| Experimental diet | |||
| Con | SPC25 | SPC50 | |
| Ingredient (%, DM) | |||
| Low-temperature fishmeal (LT FM)a | 60.0 | 45.0 | 30.0 |
| Soy protein concentrate (SPC)b | 9.0 | 18.0 | |
| Wheat gluten | 9.5 | 14.25 | 19.0 |
| Wheat flour | 15.5 | 15.5 | 15.5 |
| Starch | 5.6 | 3.93 | 2.26 |
| Fish oil | 4.9 | 6.25 | 7.6 |
| Mono-calcium phosphate (MCP)c | 1.05 | 2.1 | |
| Vitamin C | 0.5 | 0.5 | 0.5 |
| Mineral premixd | 2.0 | 2.0 | 2.0 |
| Vitamin premixe | 1.0 | 1.0 | 1.0 |
| Betaine | 0.5 | 0.5 | 0.5 |
| Choline | 0.5 | 0.5 | 0.5 |
| Lysine (99%) f | 0.375 | 0.75 | |
| Methionine (99%) g | 0.145 | 0.29 | |
| Nutrients (%, DM) | |||
| Dry matter | 96.3 | 96.8 | 96.3 |
| Crude protein | 53.9 | 53.1 | 52.8 |
| Crude lipid | 9.3 | 9.5 | 10.6 |
| Ash | 13.3 | 12.7 | 10.8 |
| Carbohydrateh | 23.5 | 24.7 | 25.8 |
| Gross energy (GE) (kcal/g)i | 4.800 | 4.845 | 4.992 |
a Low-temperature fishmeal (LT FM) (crude protein: 72.0%, crude lipid: 12.0%, ash: 11.0%) was purchased from FF Skagen (Havnevagtvej, Skagen, Denmark) [USD 1.75/kg FM; 1 USD = 1,300 KRW]. b Soy protein concentrate (SPC) (crude protein: 67.2%, crude lipid: 1.0%, ash: 6.3%) was purchased from ADMⓇ (Quincy, IL, USA) (USD 1.05/kg SPC). c MCP was purchased from Kunmin Chuan Jin Nuo Chemical Co., Ltd. (Kunming, Yunnan, China). d Mineral premix (g/kg mix): NaCl, 43.3; MgSO4·7H2, 136.5; NaH2PO4·2H2O, 86.9; KH2PO4, 239; CaHPO4, 135.3; Ferric citrate, 29.6; ZnSO4·7H2O, 21.9; Ca-lactate, 304; CuCl, 0.2; AlCl3·6H2O, 0.15; KI, 0.15; MnSO4·H2 O, 2.0; CoCl2·6H2O, 1.0. e Vitamin premix (g/ kg mix): L-ascorbic acid, 121.2; DL-α-tocopherol acetate, 18.8; thiamine hydrochloride, 2.7; riboflavin, 9.1; pyridoxine hydrochloride, 1.8; niacin, 36.4; Ca-D-pantothenate, 12.7; myo-inositol, 181.8; D-biotin, 0.27; folic acid, 0.68; p-aminobenzoic acid, 18.2; menadione, 1.8; retinyl acetate, 0.73; cholecalciferol, 0.003. f Lysine was purchased from MPbio (Irvine, CA, USA). g Methionine was purchased from MPbio. h Carbohydrate was calculated by the difference [100 – (crude protein + crude lipid + ash)].i Gross energy (GE) (kcal/g) was calculated by the difference [100-(crude protein + crude lipid + ash)] and 4 kcal/g for crude protein and carbohydrate, and 9 kcal/g for lipid [62].
2.3. Determination of Biological Indices
Six fish were sampled at the beginning and after the 140-day experiment to measure the biological indices. All live fish were starved for 24 h and anesthetized in 50 ppm 2-phenoxyethanol (Sigma, St. Louis, MO, USA). All live fish from each tank were counted and weighed collectively to measure survival and weight gain. The biological indices of the fish were calculated using the following equations:
Specific growth rate (SGR, %/day) = (Ln final weight – Ln initial weight) × 100/day feeding trial
Daily feed intake (%/day) = feed consumption × 100 / (initial fish weight + final fish weight + dead fish weight)
Feed efficiency (FE) = weight gain/feed consumption
Protein efficiency ratio (PER) = weight gain/protein consumption
Protein retention (PR, %) = protein gain × 100/protein consumption
Condition factor (K, g/cm3) = body weight (g) × 100/ body length (cm)3
Viscerosomatic index (VSI, %) = visceral weight × 100/body weight
Hepatosomatic index (HSI, %) = liver weight ×100/body weight
2.4. Biochemical Composition of the Experimental Diets and the Fish
The dorsal muscle of six fish at the beginning of the experiment and after completing the 140-day feeding experiment was homogenized and used to analyze the biochemical composition. The proximate composition of the experimental diets and the dorsal muscle were analyzed according to standard methods [24]. Crude protein and crude lipid content were determined using the Kjeldahl method (KjeltecTM 2100 Distillation Unit; Foss, Hillerød, Denmark) and ether-extraction (SoxtecTM 2043 Fat Extraction System; Foss), respectively. Moisture content was measured by drying in an oven for 24 h at 105°C, and the ash content of the samples was determined using a muffle furnace at 550°C for 4 h.
The AAs in the experimental feeds were hydrolyzed by adding 20 mL 6 N HCl at 110°C for 24 h in a drying oven. The solution was filtered with a glass filter and concentrated under decompression at 55°C to completely evaporate the acid and water. The sample was dissolved with sodium citrate buffer (pH 2.20) in a 25 mL flask, filtered with a 0.45 µm membrane filter, and analyzed using an automatic AA analyzer (Biochrom 30+; Biochrom Ltd., Cambridge, England).
Total lipids in the experimental feeds were extracted with a chloroform and methanol mixture (2:1) to methylate FA with a 14% BF-methanol (Sigma-Aldrich) solution and FAs were analyzed by gas chromatography (Trace 1310; Thermo Scientific, Waltham, MA, USA) equipped with a capillary column (SPTM-2560, 100 m × 0.25 mm; Supelco Inc., Bellefonte, PA, USA). A mixture of 37 FAs (PUFA 37 Component FAME Mix; Supelco) was used as the FA standard. The methods and procedures for the FA analysis are described by [25].
2.5. Quantitative Real-Time Polymerase Chain Reaction (PCR)
The brain, stomach, and middle intestine of six randomly selected fish were collected at the end of the 140-day feeding trial, immersed in Trizol reagent (Ambion, Carlsbad, CA, USA) containing five times the reagent tissue volume, and stored at –80°C until use. The tissue was thawed just before RNA extraction, and the extracted RNA was synthesized using RT and the GO Master Mix (MPbio, Irvine, CA, USA). The concentration of cDNA used was 100 ng/µL. The olive flounder 18S rRNA gene (Genbank, EF126037.1) was selected as the housekeeping gene and all primers used for gene expression are shown in Table 2. The levels of gene expression induced by the dietary treatments were compared among the growth, immune, and stress-related genes using RNA extracted from the brain. The amount of gene expression caused by the dietary treatments was compared through digestive enzyme genes using RNA extracted from the stomach and middle intestine. These genes underwent an initial denaturation step at 98°C for 2 min, followed by denaturation at 98°C for 10 s, annealing at 60°C for 10 s, and extension at 68°C for 30 s for 40 cycles.
2.6. Statistical Analysis
Statistical analysis was carried out using R program 3.5.1 (R Development Core Team, 2018) and SPSS version 24.0 software (SPSS Inc., Chicago, IL, USA) to conduct one-way analysis of variance and the Duncan’s multiple range test [26]. All values are expressed as means of triplicates±SEs. A P-value < 0.05 was considered significant.
3. Results
3.1. AA and FA Profiles of the Experimental Diets
The AA and FA profiles of the experimental diets are shown in Table 3 and Table 4, respectively. The contents of EAAs, such as lysine and methionine, were lower in the SPC than in the FM. Histidine and phenylalanine content among EAAs and glutamic acid and proline content among NEAAs increased as the amount of substituted SPC was increased, but other EAAs and NEAAs decreased. The arginine (2.04–2.10% of the diet) and lysine (1.50–2.10% of the diet) contents in all of the experimental diets met the dietary requirements of olive flounder (Alam et al., 2002; Forster and Ogata, 1998) but methionine (1.44–1.49% of the diet) was lower than the dietary requirement [27].
FAs were not detected in SPC. The total content of saturated FAs (∑SFA) increased as the amount of SPC substituted for FM increased but decreased with the total content of monounsaturated FAs (∑MUFA) and n-3 highly unsaturated FAs (∑n-3-HUFA). The ∑n-3 HUFA content in the SPC50 diet did not satisfy the dietary ∑n-3 HUFA requirement of olive flounder [28].
3.2. Survival and Growth Performance (Table 5)
The survival rate ranged from 94.8% to 96.3% but it was not affected by dietary treatment. Weight gain ranged from 390.8 to 442.3 g/fish and the SGR was 0.31–0.34%/day but these parameters were not affected by replacing FM with SPC.
Table 5.
Survival (%), weight gain (g/fish), and specific growth rate (SGR) of olive flounder fed the experimental diets for 140 days.
Table 5.
Survival (%), weight gain (g/fish), and specific growth rate (SGR) of olive flounder fed the experimental diets for 140 days.
| Experimental diet | Initial weight (g/fish) |
Final weight (g/fish) |
Survival (%) | Weight gain (g/fish) | SGR (%/day) a |
| Con | 721.4±6.95 | 1112.5±28.80 | 96.3±1.79 | 390.8±24.95 | 0.31±0.016 |
| SPC25 | 729.4±5.63 | 1170.3±90.10 | 94.7±4.09 | 442.3±86.24 | 0.34±0.048 |
| SPC50 | 732.5±3.61 | 1131.9±24.47 | 96.3±2.49 | 400.6±24.29 | 0.31±0.015 |
| P–value | P > 0.9 | P > 0.9 | P > 0.05 | P > 0.9 | P > 0.05 |
Values (mean of triplicates±SE) in the same column sharing the same superscript letter are not significantly different (P > 0.05). a Specific growth rate (SGR, %/day) = (Ln final weight of fish – Ln initial weight of fish)×100/days of feeding trial.
3.3. Feed Availability and Biological Indices (Table 6)
Daily feed intake (DFI) ranged from 0.44% to 0.48%/day and was not affected by dietary treatment. FER was 0.63–0.70, PER was 1.16–1.32, and PR was 37.36–42.90%. None of the feed availability indices changed in response to dietary treatment.
The condition factor of the fish ranged from 1.04 to 1.16 g/cm3, the VSI was 3.45– 3.99%, and the HSI was 1.44–1.57%. These parameters were not affected by dietary treatment.
Table 6.
Daily feed intake (DFI), feed efficiency (FE), protein efficiency ratio (PER), protein retention (PR), condition factor (CF), viscerosomatic index (VSI), and hepatosomatic index (HSI) of olive flounder fed the experimental diets for 140 days.
Table 6.
Daily feed intake (DFI), feed efficiency (FE), protein efficiency ratio (PER), protein retention (PR), condition factor (CF), viscerosomatic index (VSI), and hepatosomatic index (HSI) of olive flounder fed the experimental diets for 140 days.
| Experimental diet | DFI (%/day) a | FE b | PER c | PR (%) d | CF (g/cm3) e | VSI (%) f | HSI (%) g |
| Con | 0.48±0.018 | 0.63±0.024 | 1.16±0.045 | 37.36±1.510 | 1.04±0.247 | 3.99±0.574 | 1.47±0.282 |
| SPC25 | 0.46±0.004 | 0.70±0.074 | 1.32±0.139 | 42.62±3.703 | 1.16±0.283 | 3.59±0.263 | 1.57±0.344 |
| SPC50 | 0.44±0.009 | 0.69±0.021 | 1.30±0.039 | 42.90±1.256 | 1.04±0.083 | 3.45±0.356 | 1.44±0.150 |
| P–value | P > 0.05 | P > 0.05 | P > 0.5 | P > 0.9 | P > 0.6 | P > 0.8 | P > 0.9 |
Values (mean of triplicates±SE) in the same column sharing the same superscript letter are not significantly different (P > 0.05). a Daily feed intake (DFI, %/day) = feed consumption×100/[initial fish weight + final fish weight + dead fish weight]. b Feed efficiency (FE) = weight gain of fish/feed consumption. c Protein efficiency ratio (PER) = weight gain of fish/protein consumption. d Protein retention (PR, %) = protein gain of fish × 100/protein consumption. e Condition factor (CF, g/cm3) = body weight of fish (g) × 100/total length of fish (cm)3. f Viscerosomatic index (VSI, %) = viscera weight of fish × 100/body weight of fish. g Hepatosomatic index (HSI, %) = liver weight of fish ×100/body weight of fish.
3.4. Proximate Composition of Dorsal Muscle (Table 7)
The moisture content ranged from 71.95% to 72.87% and crude protein was 21.91–22.44%. These did not change after substituting SPC for FM. However, lipid levels of fish fed the SPC50 diet were significantly (P < 0.04) higher than those of fish fed the Con and SPC25 diets. Ash content of Con and SPC25 fish was significantly (P < 0.05) higher than that of SPC50 fish.
Table 7.
Proximate composition (%, wet weight) of the dorsal muscle of olive flounder fed the experimental diets for 140 days.
Table 7.
Proximate composition (%, wet weight) of the dorsal muscle of olive flounder fed the experimental diets for 140 days.
| Experimental diet | Moisture | Crude protein | Crude lipid | Ash |
| Con | 72.87±0.020 | 22.44±0.240 | 2.37±0.400b | 1.57±0.005a |
| SPC25 | 72.69±0.330 | 22.26±0.500 | 2.61±0.025b | 1.55±0.025a |
| SPC50 | 71.95±0.085 | 21.91±0.045 | 4.44±0.145a | 1.50±0.005b |
| P–value | P > 0.1 | P > 0.4 | P < 0.04 | P < 0.05 |
Values (mean of triplicates±SE) in the same column sharing the same superscript letter are not significantly different (P > 0.05).
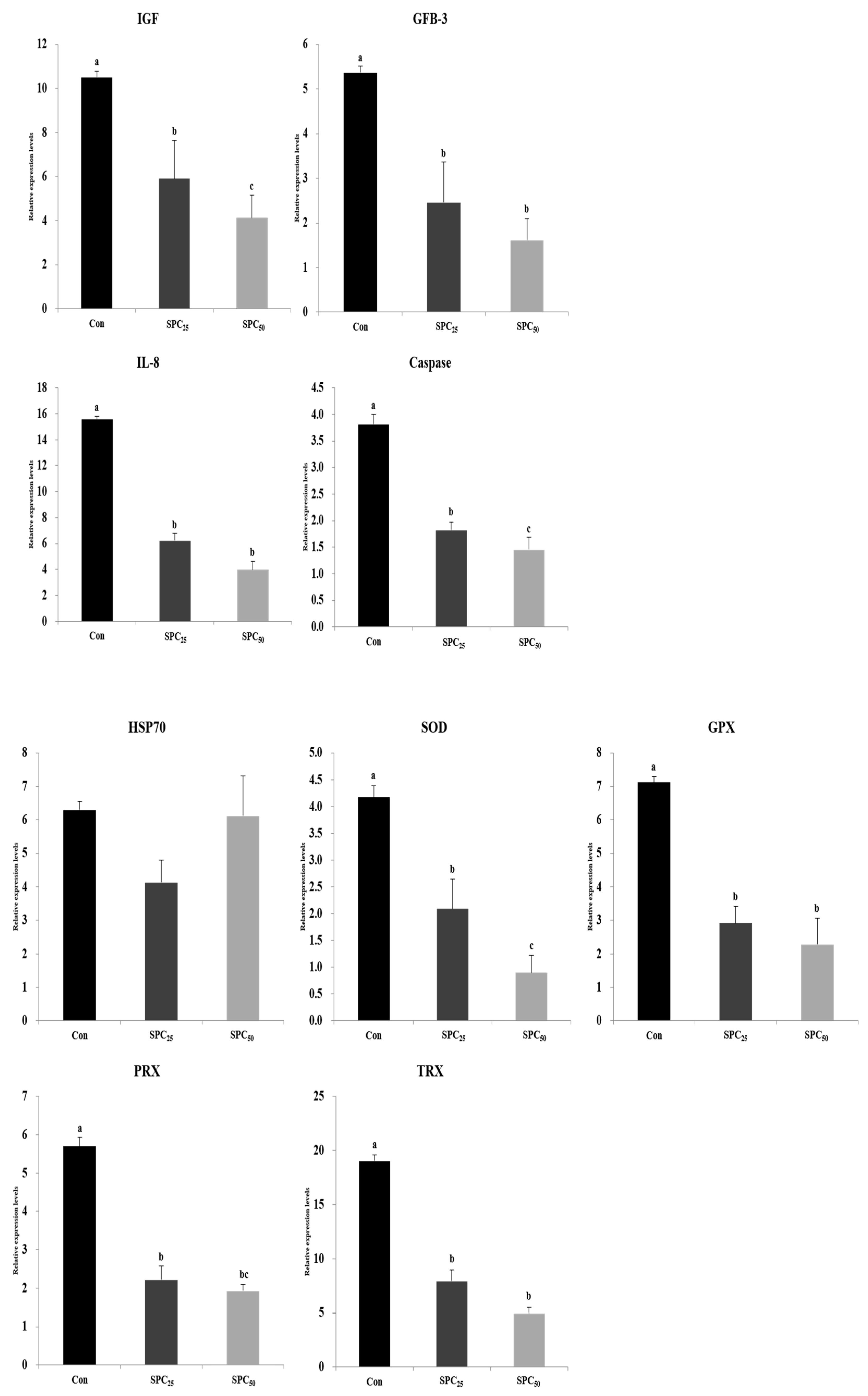
3.5. Expression Analysis of Growth, Immune, and Stress-Related Genes (Figure 1)
The expression levels of genes encoding insulin-like growth factor (IGF), growth factor beta-3-like protein (GFB-3), interleukin-8 (IL-8), and caspase in the brains of fish fed the Con diet were significantly (all P < 0.001) higher than those of fish fed the SPC25 and SPC50 diets.
The expression levels of genes related to stress, including superoxide dismutase (SOD), glutathione peroxidase (GPX), peroxiredoxin (PRX), thioredoxin (TRX), were significantly (P < 0.05) higher in Con fish than other groups. However, that of heat shock protein 70 (HSP70) was not different among the diets.
Figure 1.
Expression levels of growth, immune, stress, and antioxidant stress-related genes in the brain of olive flounder. The initial (M0-C) value for each gene was set to 1.0. (IGF, P < 0.001; GFB-3, P < 0.001; IL-8, P < 0.001; caspase, P < 0.001; HSP70, P > 0.05; SOD, P < 0.001; GPX, P < 0.05; PRX, P < 0.001; TRX, P < 0.001). IGF, insulin-like growth factor; GFB-3, growth factor beta-3-like protein; IL-8, interleukin-8; HSP70, heat shock protein 70; SOD, superoxide dismutase; GPX, glutathione peroxidase; PRX, peroxiredoxin; TRX, thioredoxin.
Figure 1.
Expression levels of growth, immune, stress, and antioxidant stress-related genes in the brain of olive flounder. The initial (M0-C) value for each gene was set to 1.0. (IGF, P < 0.001; GFB-3, P < 0.001; IL-8, P < 0.001; caspase, P < 0.001; HSP70, P > 0.05; SOD, P < 0.001; GPX, P < 0.05; PRX, P < 0.001; TRX, P < 0.001). IGF, insulin-like growth factor; GFB-3, growth factor beta-3-like protein; IL-8, interleukin-8; HSP70, heat shock protein 70; SOD, superoxide dismutase; GPX, glutathione peroxidase; PRX, peroxiredoxin; TRX, thioredoxin.
3.6. Expression Analysis of Digestive Enzyme Genes in the Stomach and Middle Intestine (Figure 2 and Figure 3)
The expression of the genes encoding α-amylase (Amy), chymotrypsinogen 2 (chymo-TRY2), trypsinogen 2 (TRY2), and trypsinogen 3 (TRY3), and lipase activity, in the stomach and middle intestine of olive flounder are presented in Figure 2 and Figure 3, respectively. Amy and lipase in the stomach were not different among the experimental diets. However, the expression levels of chymo-TRY2, TRY2, and TRY3 in the stomachs of SPC25 fish were significantly (all P < 0.02 or better) higher than those of Con and SPC50 fish.
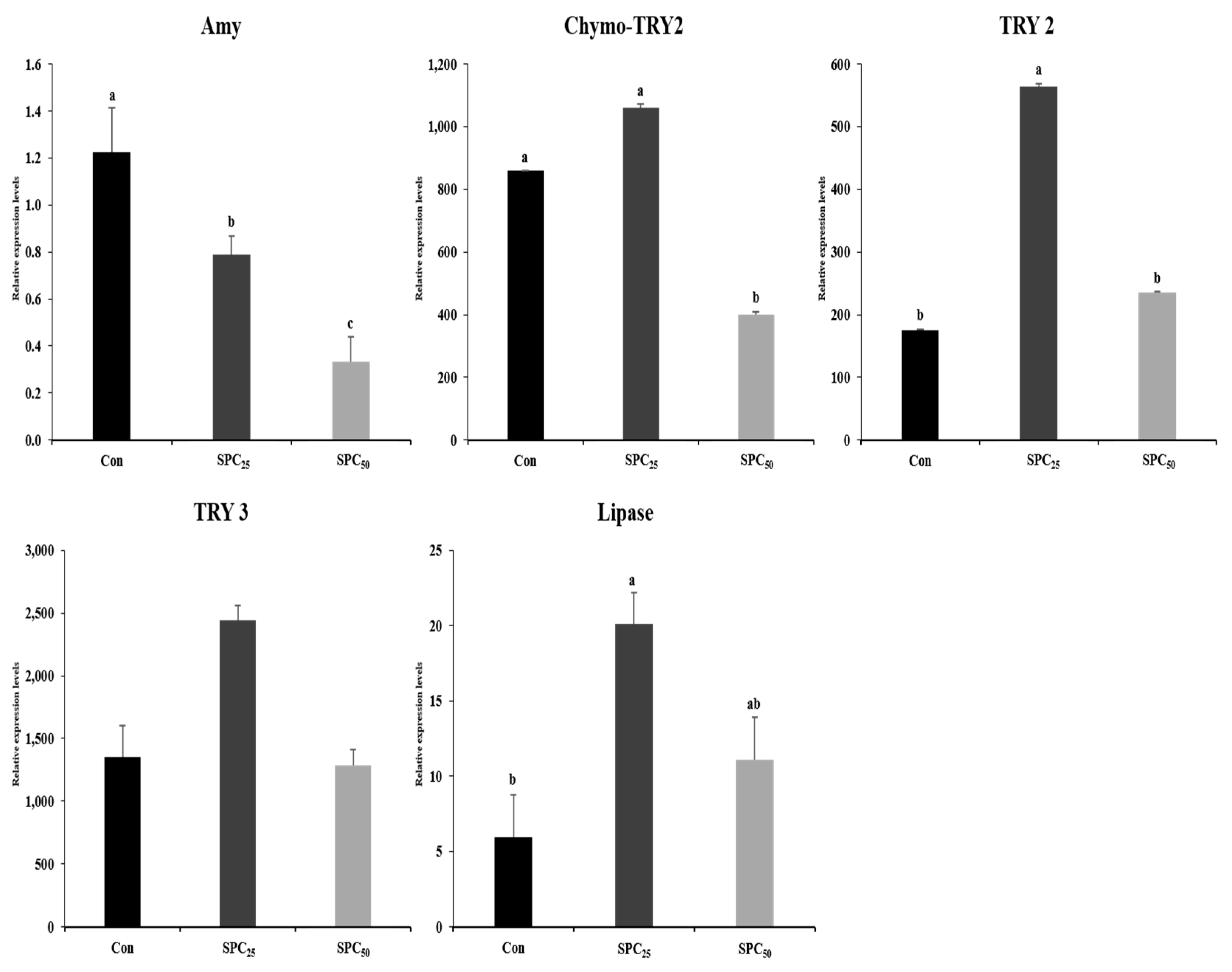
In the middle intestine, the expressions of Amy, chymo-TRY2, and TRY2 were significantly higher in Con, Con and SPC25, and Con and SPC25 fish, respectively, than in the remaining group(s) in each comparison. The TRY3 gene was not affected by dietary treatment. Lipase activity was significantly higher in SPC25 fish than in Con fish but not SPC50 fish.
Figure 2.
Expression levels of the digestive enzyme genes in the stomach of olive flounder. The initial (M0-C) value for each gene was set to 1.0. (Amy, P > 0.05; Chymo-TRY2, P < 0.02; TRY2, P < 0.02; TRY3, P < 0.001; lipase, P > 0.05). Amy, α-amylase; chymo-TRY2, chymo-trypsinogen 2; TRY2, trypsinogen 2; TRY3, trypsinogen 3.
Figure 2.
Expression levels of the digestive enzyme genes in the stomach of olive flounder. The initial (M0-C) value for each gene was set to 1.0. (Amy, P > 0.05; Chymo-TRY2, P < 0.02; TRY2, P < 0.02; TRY3, P < 0.001; lipase, P > 0.05). Amy, α-amylase; chymo-TRY2, chymo-trypsinogen 2; TRY2, trypsinogen 2; TRY3, trypsinogen 3.
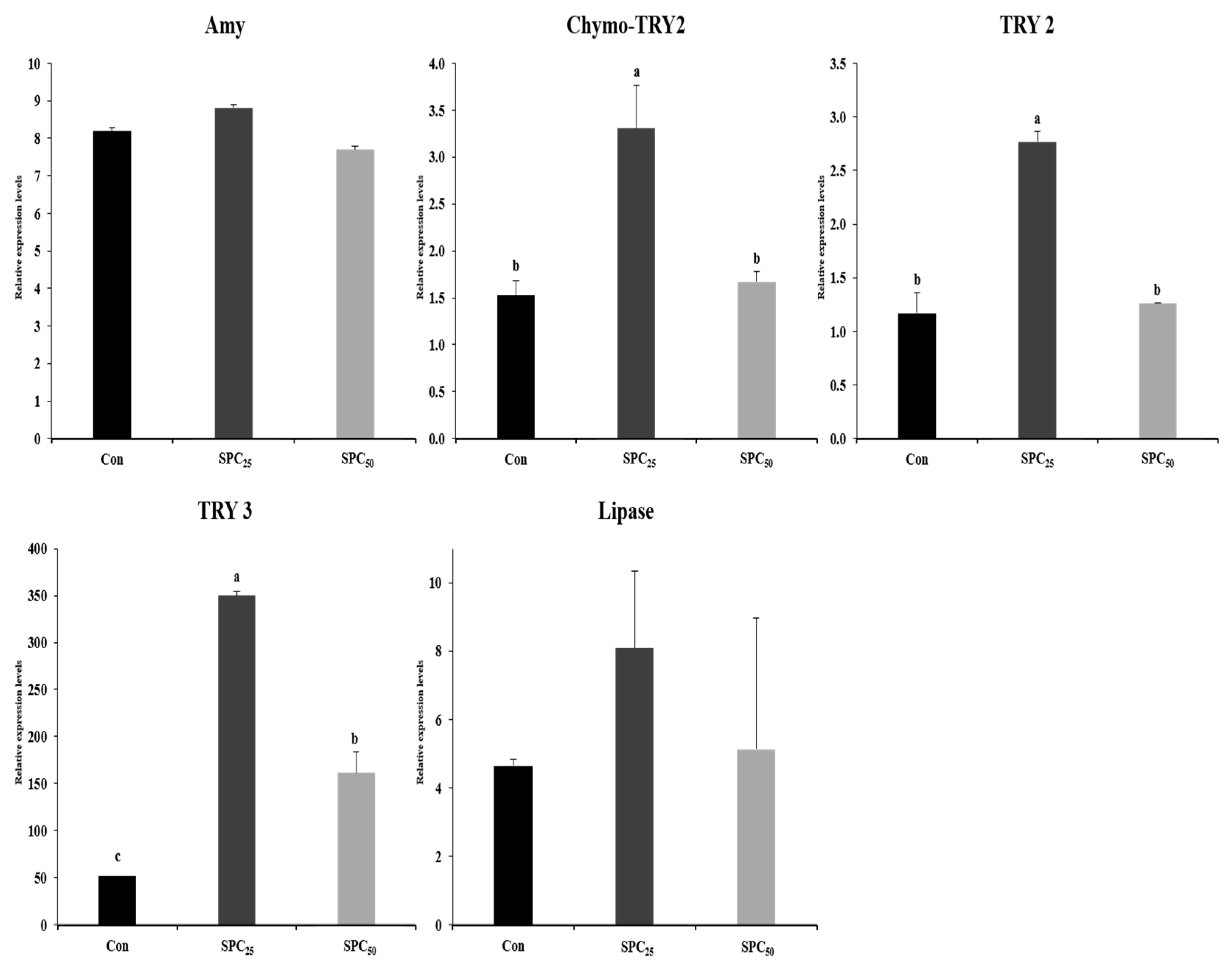
Figure 3.
Expression levels of digestive enzyme genes in the middle intestine of olive flounder. The initial (M0-C) value for each gene was set to 1.0. (Amy, P < 0.001; Chymo-TRY2, P < 0.05; TRY2, P < 0.001; TRY3, P > 0.05; lipase, P < 0.02). Amy, α-amylase; chymo-TRY2, chymo-trypsinogen 2; TRY2, trypsinogen 2; TRY3, trypsinogen 3.
Figure 3.
Expression levels of digestive enzyme genes in the middle intestine of olive flounder. The initial (M0-C) value for each gene was set to 1.0. (Amy, P < 0.001; Chymo-TRY2, P < 0.05; TRY2, P < 0.001; TRY3, P > 0.05; lipase, P < 0.02). Amy, α-amylase; chymo-TRY2, chymo-trypsinogen 2; TRY2, trypsinogen 2; TRY3, trypsinogen 3.
4. Discussion
The weight gain and SGR of olive flounder fed the SPC25 and SPC50 diets were comparable to fish fed the Con diet, suggesting that up to 50% of the FM could be replaced by SPC with AA (lysine and methionine) supplementation in a 60% FM-based diet without affecting growth performance. This contrasts with a previous study that did not include AA supplementation and found that using FM lowered growth in this species [13]. SPC can replace approximately 40% of the FM in the yellow croaker (Larimichthys crocea) diet and 60% of the FM with AA supplementation can be substituted in the gilthead sea bream diet [11,19]. Similarly, 10% and 40% FM could be substituted with corn gluten meal without AA supplementation in the Asian seabass (Lates calcarifer) diet [29] and with AA (arginine, lysine, and tryptophan) supplementation in the olive flounder diet [30] without deteriorating growth performance.
The arginine (2.04–2.10% of the diet) [31] and lysine (1.50–2.10% of the diet) [32] requirements of olive flounder were satisfied in all experimental diets in the current study. However, methionine content (1.17–1.37% of the diet) was slightly lower than the dietary methionine requirement in all experimental diets including the Con diet (1.44–1.49% of the diet in the presence of 0.06% cysteine) [27]. However, previous studies have demonstrated that cysteine can spare about 40–50% of the dietary methionine requirements of red drum (Sciaenops ocellatus) [33] and stinging catfish (Heteropneustes fossilis) [34]. Therefore, the relatively high cysteine content in our study compared to that reported by Alam et al. [27] lowered the dietary methionine requirement of olive flounder in this study.
Supplementing FM with SPC increased ∑SFA but decreased ∑MUFAs and ∑n-3 HUFAs. The levels of n-3 HUFAs, including EPA and DHA, play critical roles in the growth and health of fish [35,36]. The dietary ∑n-3 HUFA (8.16–10.20% of the total FA) requirement of olive flounder [28] was met in the Con and SPC25 diets but not the SPC50 diet. Nevertheless, lower ∑n-3 HUFA content in the SPC50 diet was not detrimental to growth performance. In a previous study, when FM was replaced with up to 25% (7.31% of total fatty acids) SPC, there was a difference in ∑n-3 HUFA content in the Con (11.4% of total fatty acids) but no difference in growth performance [37]. Similarly, Li et al. [38] reported a difference in ∑n-3 HUFA content (Con, 15.39%; SPC25, 11.30% of the total fatty acids) when up to 25% of FM was replaced with tuna or chicken byproduct with no differences in growth performance. Different levels of ∑n-3 HUFA content in the diet (0.26% diet, 6.12%; 0.52% diet, 12% of total fatty acids) did not change the growth performance of juvenile grass carp (Ctenopharyngodon idellus) [39]. Furthermore, excessive dietary ∑n-3 HUFA content does not negatively affect the growth of juvenile flounder [39,40].
DFI, FE, PER, and PR were not affected by substituting SPC for FM in this study, suggesting that substituting up to 50% of the FM with SPC in diets supplemented with limited AAs does not decrease DFI or feed utilization (FE, PER, and PR) in this species. Kim et al. [15] and Park et al. [41] reported that when 30% of the FM is replaced with SPC and the diet is supplemented with limited AAs, there are no differences in feed availability of olive flounder. In another study, no difference in feed utilization was observed when up to 40% of the FM was replaced with SPC in the yellow croaker diet [11]. In Kokou et al. [19], no difference in feed utilization of gilthead sea bream was observed when up to 60% of the FM was replaced with SPC in the diet. Similarly, in Li et al. [42], no difference in feed utilization was reported when up to 60% of the FM was replaced with SPC in the starry flounder (Platichthys stellatus) diet.
Biological indices (CF, VSI, and HSI) are commonly used to evaluate well-being, obesity, and health of fish. The VSI and HSI indicate the dietary nutritional utilization of fish and their health condition [43]. The biological indices of olive flounder remained unchanged after replacing up to 50% of the FM with SPC in a diet supplemented with limited AAs in this study, indicating that SPC can replace FM without adversely affecting the biological indices of this species. Similarly, substituting a plant protein source for FM does not affect the biological indices of fish [44,45].
Moisture and crude protein of the dorsal muscle were not affected by replacing FM with SPC in this experiment. However, higher crude lipid and lower ash content were found fish fed the SPC50 diet compared to the other groups, probably resulting from the lower levels of these components in that food preparation. Many studies have reported on the body composition, AAs, and FAs of fish fed experimental diets [38,40,42,46,47]. However, there are conflicting results on the effect of replacing FM with a plant protein source [48]. Gene expression varies according to the type, amount, and quality of the protein in the diet [49,50]. IGF plays a crucial role in regulating growth, development, and nutritional metabolism in teleost fish [49,51]. Expression of the IGF-I gene in the liver of olive flounder fed combined plant (wheat gluten and SPC) and animal (tankage meal and poultry by-product meal) protein for 20 weeks was comparable to fish fed a 65% FM-based diet, which reflected the growth of fish, but increasing the FM replacement level in the diet decreased the expression of the IGF-I gene in the liver [51]. In our study, at the end of the 140-day feeding trial, expression of growth (IGF and GFB-3) genes in the brain of fish fed the Con diet was higher than fish fed the other diets. Although the growth of fish fed the experimental diets was similar, we assumed that fish fed the Con diet would achieve higher growth than those fed the other diets as the rearing period was extended.
IL-8 presents antigens and stimulates neutrophil chemotaxis [52]. It is detectable when the immune system forms in olive flounder [53]. Caspases play an important role in the immune response and apoptosis and also maintain cell homeostasis [54,55]. A previous study reported that the immune-related gene expression levels in olive flounder were not significantly different when fish were fed a diet containing different levels of FM [44]. In this study, immune (IL-8 and caspase) gene expression in the brain of fish fed the Con diet was higher than in fish fed the other diets, indicating that immune activity might not be properly achieved and affected the activation and apoptosis of inflammatory cells of fish when fish are fed a low FM-based diet.
Jia et al. [56] reported that excessive or prolonged stress causes physiological disorders, immunosuppression, and reduced growth. Heat shock protein70 (HSP70) maintains homeostasis in response to various stressors [57] and aids the recovery of damaged cells and proteins [58]. The SOD, GPX, PRX, and TRX genes, which are involved in antioxidant responses, were used to measure antioxidant-related stress. All of these genes, except HSP70 in the brain of fish fed the Con diet, were higher than those of fish fed the SPC25 and SPC50 diets. HSP70 increased to the corresponding level in a previous study on black sea bream (Acanthopagrus schlegelii) but then decreased after HUFA supplementation [59]. Ji et al. [39] reported that excessive HUFA supplementation may be related to oxidative stress in grass carp. This may explain the results of this study so further research is needed.
Digestibility is an essential factor in the growth of fish. The expression of digestive enzyme genes (Amy, chymo-TRY2, TRY2, TRY3, and lipase) in the stomach and middle intestine of olive flounder was not different in response to the dietary treatments. However, the expression of Amy in the middle intestine decreased as the amount of FM replaced with SPC increased because carbohydrates are normally digested in the intestine [4]. The Chymo-TR2, TRY2, and TRY3 genes and lipase are involved in protein and lipid digestion, respectively. The expressions of both Chymo-TRY2 and trypsin depend on the quality of feed and increase in carnivorous marine fish with high protein availability [4,60]. The expressions of chymo-TRY2, TRY2, and TRY3 genes in the stomach of olive flounder were higher in fish fed the SPC25 diet, and the expression of the lipase gene in the stomach of fish fed the SPC25 diet was similar to that of fish fed the Con and SPC50 diets. The highest chymo-TRY2 expression was observed in the middle intestine of fish fed the Con and SPC25 diets, and the expression of TRY2 and lipase was highest in those fed the SPC25 diet. No significant difference in the growth performance of olive flounder was observed but it tended to be slightly higher in fish fed the SPC25 diet because of the higher expression of digestive-related genes in the stomach and middle intestine. Hu et al. [61] reported that lipase in the intestine of fish fed a diet in which FM was replaced with soybean meal was lower than that of fish fed a 42% FM-based diet, indicating the effect of high soybean meal content in the diet on impaired bile secretion. In this study, the lipase activity of fish fed the SPC50 diet was lower than that of fish fed the SPC25 diet, which was presumed to be affected by SPC.
5. Conclusions
Up to 50% of the FM was substituted with SPC in diets supplemented with AAs without adversely deteriorating growth performance, feed availability, or the biological indices of olive flounder. However, substituting 25% FM with SPC was most effective based on the expression of growth, immune, stress, and digestive enzyme-related genes.
Author Contributions
Sang-hyun Lee – Investigation, Writing of original draft, Reviewing and editing; Hyun Chul Kim – Conceptualization, Methodology, Project administration, Supervision, Funding acquisition.
Conflicts of Interest
The authors have no conflicts of interest to declare.
Ethics Statement
The feeding trial and handling and sampling of the experimental fish were carried out per the ethical guidelines of the Aquafeed Research Center (National Institute of Fisheries Science).
Acknowledgments
This work was supported by the National Institute of Fisheries Science, Ministry of Oceans and Fisheries, Korea (R2024032).
References
- Min, B.H.; Lee, J.H.; Noh, J.K.; Kim, H.C.; Park, C.J.; Choi, S.J.; Myeong, J.I. Hatching rate of eggs, and growth of larvae and juveniles from selected olive flounder, Paralichthys olivaceus. The Korean Soc. Dev. Biol. 2009, 14, 239–247.
- KOSIS, 2023. Korean Statistical Information Service, Korea. Available online: https://kostat.go.kr/board.es?mid=a10301010000&bid=225&act=view&list_no=430057. (Accessed 22 Mar. 2024).
- Hur, S.; Lee, J.; Lee, S.; Jeong, S.; Kim, K. Effects of worm-based extruded pellets on growth performance of olive flounder Paralichthys olivaceus in commercial aquafarms. Korean J. Fish. Aquat. Sci. 2022, 55(5), 533–540. [CrossRef]
- Lee, Y.M.; Lee, J.H.; Noh, J.K.; Kim, H.C.; Park, C.J.; Park, J.W.; Noh, G.E.; Kim, K.K. Temporal expression analyses of pancreatic and gastric digestive enzymes during early development of the olive flounder (Paralichthys olivaceus). Aquac. Res. 2017, 48, 979–989. [CrossRef]
- Kim, J.; Baek, S.I.; Cho, S.H.; Kim, T.H. Evaluating the efficacy of partially substituting fishmeal with unfermented tuna by-product meal in diets on the growth, feed utilization, chemical composition and non-specific immune responses of olive flounder (Paralichthys olivaceus). Aquac. Rep. 2022, 24, 101150. [CrossRef]
- Olsen, R.L.; Hasan, M.R. A limited supply of fishmeal: impact on future increases in global aquaculture production. Trends Food Sci. Technol. 2012, 27, 120–128. [CrossRef]
- Jeong, H.S.; Park, J.S.; Kim, H.S.; Lee, D.G.; Hwang, J.A. Comparison of dietary protein levels on growth performance of far eastern catfish silurus asotus and water quality in biofloc technology and flow-through systems. Korean J. Fish. Aquat. Sci. 2022, 55(5), 541–548.
- Médale, F.; Boujard, T.; Vallée, F.; Blanc, D.; Mambrini, M.; Roem, A.; Kaushik, S.J. Voluntary feed intake, nitrogen and phosphorus losses in rainbow trout (Oncorhynchus mykiss) fed increasing dietary levels of soy protein concentrate. Aquat. Living Resour. 1998, 11, 239–246. [CrossRef]
- Peisker, M. Manufacturing of soy protein concentrate for animal nutrition. In: Brufau, J. (Ed.), Feed Manufacturing in the Mediterranean Region. Improving Safety: From Feed to Food. Ciheam, Zaragoza, 2001, 103-107. http://om.ciheam.org/article. php?IDPDF=1600017.
- Freitas, L.E.L.; Nunes, A.J.P.; do Carmo Sá, M.V. Growth and feeding responses of the mutton snapper, Lutjanus analis (Cuvier 1828), fed on diets with soy protein concentrate in replacement of anchovy fishmeal. Aquac. Res. 2011, 42, 866–877. [CrossRef]
- Chen, Z.; Ibrahim, U.B.; Yu, A.; Wang, L.; Wang, Y. Dried porcine soluble benefits to increase fishmeal replacement with soy protein concentrate in large yellow croaker Larimichthys crocea diet. J. World Aquac. Soc. 2023, 54, 1162–1178. [CrossRef]
- Colburn, H.R.; Walker, A.B.; Breton, T.; Stilwell, J.M.; Sidor, I.F.; Gannam, A.L.; Berlinsky, D.L. Partial replacement of fishmeal with soybean meal and soy protein concentrate in diets of Atlantic Cod. N. Am. J. Aquac. 2012, 74, 330–337. 10.1080/ 15222055.2012.676008.
- Deng, J.; Mai, K.; Ai, Q.; Zhang, W.; Wang, X.; Xu, W.; Liufu, Z. Effects of replacing fishmeal with soy protein concentrate on feed intake and growth of juvenile Japanese flounder, Paralichthys olivaceus. Aquaculture 2006, 258, 503- 513. [CrossRef]
- Lim, C.H.; Lee, C.S.; Webster, C.D. Alternative protein sources in aquaculture diets. The Haworth Press, New York, NY. 2019, pp. 571.
- Kim, M.G.; Lee, C.; Shin, J.; Lee, B.J.; Kim, K.W.; Lee, K.J. Effects of fishmeal replacement in extruded pellet diet on growth, feed utilization and digestibility in olive flounder Paralichthys olivaceus. Korean J. Fish. Aquat. Sci. 2019, 52(2), 149–158. [CrossRef]
- Kim, M.G.; Shin, J.; Lee, C.; Lee, B.J.; Hur, S.W.; Lim, S.G.; Lee, K.J. Evaluation of a mixture of plant protein source as a partial fishmeal replacement in diets for juvenile olive flounder Paralichthys olivaceus. Korean J. Fish. Aquat. Sci. 2019, 52(4), 374–381. [CrossRef]
- Lim, H.; Kim, M.G.; Shin, J.; Shin, J.; Hur, S.W.; Lee, B.J.; Lee, K.J. Evaluation of three plant proteins for fishmeal replacement in diet for growing olive flounder Paralichthys olivaceus. Korean J. Fish Aquat. Sci. 2020, 53, 464–470. [CrossRef]
- Takagi S.; Shimeno, S.; Hosokawa, H.; Ukawa, M. Effect of lysine and methionine supplementation to a soy protein concentrate diet for red sea bream Pagrus major. Fish. Sci. 2001, 67, 1088–1096.
- Kokou, F.; Rigos, G.; Kentouri, M.; Alexis, M. Effects of DL-methionine-supplemented dietary soy protein concentrate on growth performance and intestinal enzyme activity of gilthead sea bream (Sparus aurata L.). Aquac. Int. 2016, 24, 257–271. [CrossRef]
- Moon, J.; Oh, D.H.; Park, S.; Seo, J.; Kim, D.; Moon, S.; Park, H.S.; Lim, S.; Lee, B.; Hur, S.; Lee, K.; Nam, T.J.; Choi, Y.H. Expression of insulin-like growth factor genes in olive flounder, Paralichthys olivaceus, fed a diet with partial replacement of dietary fishmeal. J. World Aquac. Soc. 2019, 54, 131–142. [CrossRef]
- Jang, W.J.; Hasan, M.T.; Lee, B.; Hur, S.W.; Lee, S.; Kim, K.W.; Lee, E.; Kong, I. Effect of dietary differences on changes of intestinal microbiota and immune-related gene expression in juvenile olive flounder (Paralichthys olivaceus). Aquaculture 2020, 527, 735442. [CrossRef]
- Kim, K.W.; Wang, X.J.; Bai, S.C. Optimum dietary protein level for maximum growth of juvenile olive flounder Paralichthys olivaceus (Temminck et Schlegel). Aquac. Res. 2002, 33, 673–679. [CrossRef]
- Lee, S.; Cho, S.H.; Kim, K. Effects of dietary protein and energy levels on growth and body composition of juvenile flounder Paralichthys olivaceus. J. World Aquacult. Soc. 2000, 31, 306–315. [CrossRef]
- AOAC, 1990. Association of Official Analytical Chemists. 15th ed. Arlington, VA, USA.
- Jeong, S.M.; Kim, N.L.; Hur, S.W.; Lee, S.H.; Bae J.H.; Kim, K.W. Effect of dietary inclusion of black soldier fly larvae Hermetia illucens meal on growth performance of starry flounder Platichthys stellatus and feed value. Korean J. Fish. Aquat. Sci. 2023, 56(4), 373-379. [CrossRef]
- Duncan, D.B. Multiple range and multiple F test. Biometrics 1955, 11, 1–42. [CrossRef]
- Alam, M.S.; Teshima, S.; Ishikawa, M.; Koshio, S. Methionine requirement of juvenile Japanese flounder Paralichthys olivaceus. J. World Aquacult. Soc. 2000, 31, 618–626. [CrossRef]
- Kim, K.; Lee, S. Requirement of dietary n-3 highly unsaturated fatty acids for juvenile flounder (Paralichthys olivaceus). Aquaculture 2004, 315–323, 2004. [CrossRef]
- Nandakumar, S.; Ambasankar, K.; Ali, S.S.R.; Syamadayal, J.; Vasagam, K. Replacement of fishmeal with corn gluten meal in feeds for Asian seabass (Lates calcarifer). Aquac. Int. 2017, 25, 1495–1505. [CrossRef]
- Kikuchi, K. Partial replacement of fishmeal with corn gluten meal in diets for Japanese flounder Paralichtys olivaceus. J. World Aquacult. Soc. 1999, 30, 357–363. [CrossRef]
- Alam, M.S.; Teshima, S.; Koshio, S.; Ishikawa, M. Arginine requirement of juvenile Japanese flounder Paralichthys olivaceus estimated by growth and biochemical parameters. Aquaculture 2002, 205, 127–140. [CrossRef]
- Forster, I.; Ogata, H.Y. Lysine requirement of juvenile japanese flounder Paralichthys olivaceus and juvenile red sea bream Pagrus major. Aquaculture 1998, 161, 131–142. [CrossRef]
- Goff, J.B.; Gatlin III, D.M. Evaluation of different sulfur amino acid compounds in the diet of red drum, Sciaenops ocellatus, and sparing value of cystine for methionine. Aquaculture 2004, 241, 465–477. [CrossRef]
- Farhat; Khan, M.A. Total sulfur amino acid requirement and cysteine replacement value for fingerling stinging catfish, Heteropneustes fossilis (Bloch). Aquaculture 2014, 426, 270–281. [CrossRef]
- An, W.; Dong, X.; Tan, B.; Yang, Q.; Chi, S.; Zhang, S.; Liu, H.; Yang, Y. Effects of dietary n-3 highly unsaturated fatty acids on growth, non-specific immunity, expression of some immune-related genes and resistance to Vibrio harveyi in hybrid grouper (♀ Epinephelus fuscoguttatus × ♂ Epinephelus lanceolatu). Fish. Shellfish Immunol. 2020, 96, 86–96. [CrossRef]
- Volpato, J.A.; Ribeiro, L.B.; Torezan, G.B.; da Silva, I.C.; de Oliveira Martins, I.; Genova, J.L.; de Oliveira, N.T.E.; Carvalho, S.T.; de Oliveira Carvalho, P.L.; Vasconcellos, R.S. Characterization of the variations in the industrial processing and nutritional variables of poultry by-product meal. Poult. Sci. 2022, 101, 101926. [CrossRef]
- Baek, S.I.; Jeong, H.S.; Cho, S.H. Replacement effect of fishmeal by plant protein sources in olive flounder (Paralichthys olivaceus) feeds with an addition of jack mackerel meal on growth, feed availability, and biochemical composition. Aquac. Nutr. 2023. [CrossRef]
- Li, R.; Cho, S.H. Substitution impact of tuna by-product meal for fishmeal in the diets of rockfish (Sebastes schlegeli) on growth and feed availability. Animals 2023, 13, 3586. [CrossRef]
- Ji, H.; Li, J.; Liu, P. Regulation of growth performance and lipid metabolism by dietary n-3 highly unsaturated fatty acids in juvenile grass carp, Ctenopharyngodon idellus. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2011, 159, 49–56. [CrossRef]
- Choi, J.; Aminikhoei, Z.; Kim, K.; Lee, S. Growth and fatty acid composition of juvenile olive flounder Paralichthys olivaceus fed diets containing different levels and ratios of eicosapentaenoic acid and docosahexaenoic acid. Fish Aquat. Sci. 2014, 17(1), 95-103. [CrossRef]
- Park, S.J.; Seo, B.S.; Park, H.S.; Lee, B.J.; Hur, S.W.; Nam, T.J.; Lee, K.J.; Lee, S.H.; Choi, Y.H. Effect of fishmeal content in the diet on the growth and sexual maturation of olive flounder (Paralichthys olivaceus) at a typical fish farm. Animals. 2021, 11, 2055. [CrossRef]
- Li, P.Y.; Wang, J.Y.; Song, Z.D.; Zhang, L.M.; Zhang, H.; Li, X.X.; Pan, Q. Evaluation of soy protein concentrate as a substitute for fishmeal in diets for juvenile starry flounder (Platichthys stellatus). Aquaculture 2015, 448, 578–585. [CrossRef]
- Sim, Y.J.; Cho, S.H.; Kim, K.; Jeong, S. Effect of substituting fishmeal with various by-product meals of swine-origin in diet on olive flounder (Paralichthys olivaceus). Acuac. Rep. 2023, 33, 101844 . [CrossRef]
- Niu, K.; Khosravi, S.; Kothari, D.; Lee, W.; Lim, J.; Lee, B.; Kim, K.; Lim, S.; Lee, S.; Kim, S. Effects of dietary multi-strain probiotics supplementation in a low fishmeal diet on growth performance, nutrient utilization, proximate composition, immune parameters, and gut microbiota of juvenile olive flounder (Paralichthys olivaceus). Fish. Shellfish Immunol. 2019, 93, 258–268. [CrossRef]
- Seo, B.; Park, S.; Hwang, S.; Lee, Y.; Lee, S.; Hur, S.; Lee, K.; Nam, T.; Song, J.; Kim, J.; Jang, W.; Choi, Y. Effects of decreasing fishmeal as main source of protein on growth, digestive physiology, and gut microbiota of olive flounder (Paralichthys olivaceus). Animals 2022, 12, 2043. [CrossRef]
- Fernandez-Palacios, H.; Izquierdo, M.S.; Robaina, L.; Valencia, A.; Salhi, M.; Vergara, J.M. Effect of n-3 HUFA level in broodstock on egg quality of gilthead sea bream (Sparus aurata L.). Aquaculture 1995, 132, 325–327. [CrossRef]
- Furuita, H.; Tanaka, H.; Yamamoto, T.; Suzuki, N.; Takeuchi, T. Effect of high levels of n-3 HUFA in broodstock diet on egg quality and egg fatty acid composition of Japanese flounder, Paralichthys olivaceus. Aquaculture 2002, 210, 323–333. [CrossRef]
- Bae, K.; Kim, K.; Lee, S. Evaluation of rice distillers dried grain as a partial replacement for fishmeal in the practical diet of the juvenile olive flounder Paralichthys olivaceus. Fish Aquat. Sci. 2015, 18(2), 151-158. [CrossRef]
- Fuentes, E.N.; Björnsson, B.T.; Valdés, J.A.; Einarsdottir, I.E.; Lorca, B.; Alvarez, M.; Molina, A. IGF-I/PI3K/Akt and IGF-I/ MAPK/ERK pathways in vivo in skeletal muscle are regulated by nutrition and contribute to somatic growth in the fine flounder. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300(6), R1532–R1542. [CrossRef]
- Gill, N.; Higgs, D.A.; Skura, B.J.; Rowshandeli, M.; Dosanjh, B.S.; Mann, J.; Gannam, A.L. Nutritive value of partially dehulled and extruded sunflower meal for post-smolt Atlantic salmon (Salmo salar L.) in sea water. Aquac. Res. 2006, 37(13), 1348–1359. [CrossRef]
- Park, S.; Moon, J.; Seo, J.; Nam, T.; Lee, K.; Lim, S.; Kim, K.; Lee, B.; Hur, S.; Choi, Y. Effect of fishmeal replacement on insulin-like growth factor-I expression in the liver and muscle and implications for the growth of olive flounder Paralichthys olivaceus. Korean J. Fish. Aquat. Sci. 2019, 52(2), 141–148. [CrossRef]
- Kim, J.; Jung, S. cDNA microarray analysis of viral hemorrhagic septicemia infected olive flounder, Paralichthys olivaceus: immune gene expression at different water temperature. J. Fish Pathol. 2014, 27(1), 1–9. [CrossRef]
- Kim, K.; Kim, H.C.; Park, C.; Park, J.; Lee, Y.M.; Kim, W. Interleukin-8 (IL-8) expression in the olive flounder (Paralichthys olivaceus) against viral hemorrhagic septicemia virus (VHSV) challenge. Dev. Reprod. 2019, 23(3), 231–238. [CrossRef]
- Cheng, C.; Guo, Z.; Luo, S.; Wang, A. Effects of high temperature on biochemical parameters, oxidative stress, DNA damage and apoptosis of pufferfish (Takifugu obscurus). Ecotox. Environ. Safe. 2018, 150, 190–198. [CrossRef]
- Kumari, R.; Deshmukh, R.S.; Das, S. Caspase-10 inhibits ATP-citrate lyase-mediated metabolic and epigenetic reprogramming to suppress tumorigenesis. Nat. Commun. 2019, 10, 4255. [CrossRef]
- Jia, R.; Liu, B.; Feng, W.; Han, C.; Huang, B.; Lei, J. Stress and immune responses in skin of turbot (scophthalmus maximus) under different stocking densities. Fish Shellfish Immunol. 2016, 55, 131–139. [CrossRef]
- Singh, M.K.; Shinm Y.; Ju, S.; Han, S.; Choe, W.; Yoon, K.; Kim, S.S.; Kang, I. Heat shock respose and heat shock proteins: Current understanding and future opportunities human diseases. Int. J. Mol. Sci. 2024, 25, 4209. [CrossRef]
- Currie, S.; Moyes, C.D.; Tufts, B.L. The effects of heat shock and acclimation temperature on Hsp70 and Hsp30 mRNA expression in rainbow trout: in vivo and in vitro comparisons. J. Fish Biol. 2000, 56(2), 398–408. [CrossRef]
- Jin, M.; Lu, Y.; Yuan, Y.; Li, Y.; Qiu, H.; Sun, P.; Ma, H.; Ding, L.; Zhou, Q. Regulation of growth, antioxidant capacity, fatty acid profiles, hematological characteristics and expression of lipid related genes by different dietary n-3 highly unsaturated fatty acids in juvenile black seabream (Acanthopagrus schlegelii). Aquaculture 2017, 471, 55–65. [CrossRef]
- Gwak, W.; Park, D.W. Developmental changes in digestive enzymes activity of black rockfish Sebastes inermis. J. Aquac. 2006, 19, 125–132.
- Hu, Y.; Yang, G.; Li, Z.; Hu, Y.; Zhong, L.; Zhou, Q.; Peng, M. Effect of dietary taurine supplementation on growth, digestive enzyme, immunity and resistant to dry stress of rice field eel (Monopterus albus) fed low fishmeal diets. Aquac. Res. 2018, 49(6), 2108–2118. [CrossRef]
- Garling, D.L.; Wilson, R.P. Optimum dietary protein to energy ratios for channel catfish fingerlings, Ictalurus punctatus. J Nutr. 1976, 106, 1368–1375. [CrossRef]
Table 2.
Oligonucleotide primers used for quantitative real-time PCR.
| Target | Sequence (5'-3') | Gen Bank |
| IGF | F.P = 5'-CGGCGCCTGGAGATGTACTG-3′ | AF016922.2 |
| R.P = 5'-TGTCCTACGCTCTGTGCCCT-3′ | ||
| GFB-3 | F.P = 5'-CTCAAGACCTGGAACCTCTCACTAT-3′ | KF723424.1 |
| R.P = 5'-CTCAGCTACACTTGCAAGACTTGAC-3′ | ||
| IL-8 | F.P = 5'-GTTGTTGCTGTGATGGTGCT-3′ | AB809047.1 |
| R.P = 5'-GCCGGTATCTTTCAGAGTGG-3′ | ||
| Caspase | F.P = 5'-GCACATGGACATCCTGAGTG-3′ | AB247499.1 |
| R.P = 5'-AGGCTGCTCATTTCACTGCT-3′ | ||
| HSP70 | F.P = 5'-TCCTCATGGGTGACACTTCG-3′ | AB010871.1 |
| R.P = 5'-TTGTCCTTGGTCATGGCTCT-3′ | ||
| SOD | F.P = 5'-GGGAATGTCACTGCTGGAAAA-3′ | EF681883.1 |
| R.P = 5'-CCAATAACTCCACAGGCCAGAC-3’ | ||
| GPX | F.P = 5'-GAAGGTGGATGTGAATGGGAAG-3′ | EU095498.1 |
| R.P = 5'-TCTGCCTCGATATCAATGGTAAGG-3′ | ||
| PRX | F.P = 5'-TCTCCTACAGCAAACAGCAC-3′ | DQ009987.1 |
| R.P = 5'-CCAGGAAGTGACACCATCAA-3′ | ||
| TRX | F.P = 5'-TGGACAGAGGCGAGGCTACT-3′ | XM020095833.1 |
| R.P = 5'-ACCCAAAGACCAAACCACACAC-3′ | ||
| Amy | F.P = 5'-CACTCTTCATGTGGAAGCTGGTTC -3′ | KJ908179 |
| R.P = 5'-CCATAGTTCTCAATGTTGCCACTGC -3′ | ||
| Chymo-TRY2 | F.P = 5'-ACTACACCGGCTTCCACTTC -3′ | AB029754 |
| R.P = 5'-GAACACCTTGCCAACCTTCATG -3′ | ||
| TRY2 | F.P = 5’-ATCGTCGGAGGGTATGAGTG-3′ | AB029751 |
| R.P = 5’-CATCCAGAGACTGTGCACATG-3′ | ||
| TRY3 | F.P = 5'-TATGAGTGCACGCCCTACTC -3′ | AB029752 |
| R.P = 5'-GTTCTCACAGTCCCTCTCAGAC -3′ | ||
| Lipase | F.P= 5'-ATGGGAGAAGAAAATATCTTATTTTTGA -3′ | HQ850701 |
| R.P = 5'-TACCGTCCAGCCATGTATCAC -3′ |
Insulin-like growth factor, IGF; growth factor beta-3-like protein, GFB-3; interleukin-8, IL-8; caspase; heat shock protein 70, HSP70; superoxide dismutase, SOD; glutathione peroxidase, GPX; peroxiredoxin, PRX; thioredoxin, TRX; α-Amylase, Amy; chymo-trypsinogen 2, chymo-TRY2; trypsinogen 2, TRY2; trypsinogen 3, TRY3.
Table 3.
Amino acid (g/kg of the diet) profiles of the experimental diets.
| Ingredients | Requirement | Experimental diet | ||||
| FM | SPC | Con | SPC25 | SPC50 | ||
| Essential amino acids (EAA) (g/kg) | ||||||
| Arginine | 36.6 | 45.5 | 20.4–21.01 | 29.4 | 27.2 | 26.3 |
| Histidine | 13.4 | 16.4 | 9.6 | 11.5 | 12.1 | |
| Isoleucine | 28.9 | 31.1 | 22.3 | 21.6 | 21.0 | |
| Leucine | 47.4 | 50.3 | 39.4 | 37.5 | 36.3 | |
| Lysine | 50.7 | 41.3 | 1.50–21.02 | 36.3 | 33.6 | 32.4 |
| Methionine | 17.7 | 6.7 | 14.4–14.93 | 13.7 | 12.3 | 11.7 |
| Phenylalanine | 25.3 | 32.1 | 22.6 | 22.8 | 23.2 | |
| Threonine | 25.8 | 24.9 | 21.7 | 19.2 | 17.7 | |
| Valine | 33.8 | 31.9 | 26.7 | 24.5 | 23.5 | |
| ∑EAA4 | 279.6 | 280.2 | 221.7 | 210.2 | 204.2 | |
| Non-essential amino acids (NEAA) (g/kg) | ||||||
| Alanine | 40.1 | 27.1 | 31.7 | 26.6 | 23.0 | |
| Aspartic acid | 59.0 | 72.6 | 44.8 | 40.8 | 38.5 | |
| Cysteine | 3.5 | 5.9 | 0.63 | 6.1 | 3.0 | 2.8 |
| Glutamic acid | 89.6 | 127.8 | 97.6 | 111.4 | 119.3 | |
| Glycine | 42.9 | 27.6 | 33.5 | 28.2 | 24.4 | |
| Proline | 21.8 | 33.1 | 30.9 | 35.5 | 39.3 | |
| Serine | 22.6 | 31.7 | 22.6 | 22.1 | 21.9 | |
| Tyrosine | 17.7 | 19.3 | 15.6 | 14.8 | 14.6 | |
| ∑NEAA5 | 331.5 | 384.5 | 282.8 | 282.4 | 283.8 | |
Con: 60% fishmeal-based diet; LF1: dietary replacement of 25% fishmeal with soy protein concentrate; LF2: dietary replacement of 50% fishmeal with soy protein concentrate. 1, 2, 3Data were obtained from Alam et al. (2002), Forster and Ogata (1998), and Alam et al. (2000), respectively. 4∑EAA: Total essential amino acid contents. 5∑NEAA: Total non-essential amino acid contents.
Table 4.
Fatty acid (g/kg of total fatty acids) profiles of the experimental diets.
| Ingredient | Requirement | Experimental diet | ||||
| FM | SPC | Con | SPC25 | SPC50 | ||
| C14:0 | 59.3 | 34.8 | 31.9 | 26.2 | ||
| C16:0 | 241.9 | 205.1 | 202.9 | 187.1 | ||
| C18:0 | 49.1 | 197.7 | 222.6 | 247.6 | ||
| C20:0 | 15.3 | |||||
| C22:0 | 19.4 | |||||
| ∑SFA | 384.9 | 437.6 | 457.4 | 460.9 | ||
| C17:1n-7 | 48.6 | 46.5 | 42.9 | |||
| C18:1n-9 | 28.5 | 28.0 | 27.0 | |||
| C24:1n-9 | 15.2 | |||||
| ∑MUFA2 | 15.2 | 77.1 | 74.5 | 69.9 | ||
| C18:2n-6 | 118.6 | 178.3 | 210.3 | 260.4 | ||
| C18:3n-6 | 29.3 | 31.3 | 35.2 | |||
| C20:2n-6 | 37.9 | |||||
| C20:3n-6 | 18.1 | |||||
| C20:4n-6 | 44.2 | |||||
| C20:5n-3 | 120.6 | |||||
| C22:2n-6 | 0.9 | 68.4 | 65.0 | 49.7 | ||
| C22:6n-3 | 169.5 | 117.8 | 108.3 | 77.8 | ||
| ∑n-3 HUFA3 | 491.8 | 8.16–10.204 | 117.8 | 108.3 | 77.8 | |
| Unknown | 139.7 | 73.4 | 53.2 | 46.1 | ||
Con: 60% fishmeal-based diet; LF1: dietary replacement of 25% fishmeal with soy protein concentrate; LF2: dietary replacement of 50% fishmeal with soy protein concentrate. 1∑SFA: Total saturated fatty acid contents. 2∑MUFA: Total monounsaturated fatty acid contents. 3∑n-3 HUFA: Total n-3 highly unsaturated fatty acid contents. 4Data were obtained from Kim and Lee (2004).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



