
Submitted:
03 September 2024
Posted:
04 September 2024
You are already at the latest version
Abstract
Plant-Derived Extracellular Vesicles (PDEVs) contain high levels of antioxidants, especially when deriving from organic. PDEVs from different vegetal sources has shown an increased antioxidant power, compared to PDEVs from single plants, suggesting a synergistic effect of the bioactives constitutively expressed in the PDEVs from single fruits. With this study we wanted to investigate the beneficial effects of a mix of PDEVs on human skin cells. We found detectable levels of Citric Acid, Ascorbic Acid, Glutathione, Catalase and SOD in a mix of PDEVs deriving from 5 different fruits (Grape, Red Orange, Papaya, Pomegranate and Tangerine). We then treated H2O2-conditioned fibroblasts with the mix of PDEVs. The results showed that the PDEVs ‘mix reverted the H2O2-induced redox imbalance, restoring the mitochondrial homeostasis, with a strong reduction of mitochondrial anion superoxide and increase of sirtuin levels. The antioxidant action was consistent with a wound repair on a lesion produced in a Fibroblasts’ monolayer. This result was consistent with an increased level of Vimentin and Matrix Metalloproteinase-9, whose expression is directly related to the efficiency of the reparative processes. These data support a beneficial role of PDEVs in both preventing and treating skin injuries, through their potent antioxidant and reparative activities.
Keywords:
plant-derived extracellular vesicles
; antioxidants
; natural bioactives
; anti-aging
; skin repair
; fibroblasts sirtuin
; vimentin
; wound repair
1. Introduction
Skin is the largest organ in our body representing the most important barrier against external stimuli, such as pollution and UV radiation [1,2]. The most important function of stratum corneum is the protection against dryness; in fact, impairments of skin integrity led to increasing of trans-epidermal water loss and consequent insufficient skin moisture [3,4,5]. As widely demonstrated, UV can induce oxidative stress in human skin cells, generating an increased production of reactive oxidative species that can in turn affect lipid integrity, reach the genetic material in cell nuclei and trigger DNA damages, with consequent impair of cellular function and cell death. Skin aging is a complex mechanism due to both intrinsic and extrinsic factors. However, many evidences support a key role of redox imbalance as the prime cause of skin aging. Many environmental factors, take part in the skin aging, including UV-exposure and pollution [6,7,8].
Mitochondria are fundamental in the skin protection against inflammation, and local ROS, mostly through the collagen and elastin production. Mitochondria are considered the powerhouse of cells. In fact, they generate energy in form of ATP [9], which involves a series of enzyme complexes and transporters. Mitochondrial Membrane Potential derives from redox transformation with the activity of Krebs cycle and serves as an intermediate form of energy storage [10]. Mitochondrial Membrane Potential is considered a valuable indicator of mitochondria’s health, inasmuch as it maintains an active control of the ions’ transport [10,11,12]. Actually, Mitochondrial Membrane Potential is not a stable value, and the oxidative stress represents one of the most important perturbing factors leading to a reduction of Mitochondrial Membrane Potential [10,13,14]. Mitochondria generate 90% of cellular reactive oxygen species and the imbalance between mitochondrial ROS and antioxidant defense results in oxidative stress, with consequent damages to mitochondrial components that, without therapeutic intervention, inevitably leads to cellular degenerative processes [15,16]. In particular, Mitochondrial Superoxide Anion is normally converted in oxygen and water by cellular antioxidant systems, but in pathological conditions it may overcome the antioxidant apparatus leading to different levels of cellular damages [17,18]. During the oxidative stress and inflammation, passing through a mitochondrial dysfunction, different proteins are involved, including Sirtuin 1, the most extensively studied member of Sirtuins’ family. Sirtuin 1 is a NAD+-dependent deacetylase that regulates some intracellular pathways often leading to chromatin silencing and a general decrease of the energetic state of the cell [19]. Sirtuin, participates also to the regulation of several proteins involved in age-related processes at the transcriptional level [20]. It was also shown that the age-related decrease of Sirtuin 1 content is associated to a significant inhibition of cellular proliferation [21,22,23]. Natural compounds, such as resveratrol, exert beneficial effects in skin-derived cells by increasing Sirtuin 1 expression [24,25,26,27,28,29,30].
There is clear evidence that plants contain different antioxidants that have been used in treating different diseases caused by oxidative damages, including chronic wounds, carcinogenesis, and skin aging [31,32,33,34,35,36,37]. More recently, the beneficial effect on skin health of extracellular vesicles isolated from plant extracts (PDEVs) has been widely reported. In fact, PDEVs contain a variety of antioxidants, with both enzymatic that non-enzymatic activity, that efficiently reduce damages induced by oxidative stress and help wound healing in treated cells. In this regard, increased ROS at the skin level, leads to decreased proliferation of both Fibroblasts and Keratinocytes, with a reduced epidermal turnover and decreased production of elastin [8,38,39].
Wound healing is a dynamic skin process that involves different events such as inflammation, proliferation and migration of different cell types such as Fibroblasts [40,41,42]. In this process are involved some proteins, such as Matrix Metalloproteinase and Vimentin, that drive cellular proliferation and migration to the wound site [43,44,45,46,47,48,49,50].
PDEVs can mediate wound healing by modulating gene expression and/or activating signaling pathways involved in this process. For example, exosomes from wheat and grapefruit increase collagen production along with Fibroblasts proliferation and migration to the wound site. Moreover, exosomes promote the formation of tube-like structures involved in the angiogenetic processes [51,52,53].
All in all the antioxidants constitutively contained in PDEVs can efficiently contrast damages related to skin aging, reducing intracellular ROS levels and stimulating collagen production [39,54,55,56,57,58,59].
With the above background we decided to test the effect of a “PDEVs’ magic formula”, as represented by a mix fruits, on human Skin Fibroblasts in different culture conditions, in order to assess the possible use of this PDEVs’ mix as a skin-aging reversing agent.
2. Materials and Methods
2.1. Fruit Material
Tangerine (Citrus Reticulata), Blood Orange (Citrus sinensis ‘Blood Orange’), Papaya (Carica papaya L.), Pomegranate (Punica granatum) and Grape (Vitis vinifera) were purchased from several Italian farms with organic farming certification. The fruits were washed with water and bicarbonate, peeled, and extracted with a fruit juice extractor. Fruit juices were stored at −80 °C.
2.2. Nanovesicles Isolation
Fruit juices were centrifuged at 500× g × 10 min; the supernatants were filtered with 100 µm filters and serially centrifugated at 2000× g for 20 min to eliminate cell debris and then at 15,000× g for 30 min to collect the fraction enriched in microvesicles. The supernatants were subsequently ultracentrifuged in a Sorvall WX Ultracentrifuge Series (Thermo Fisher Scientific, USA) at 110,000× g for 1 h 30 min to collect the nanovesicles. The pellet was resuspended in an appropriate buffer for downstream analyses.
2.3. Total Antioxidant Activity Assay
The detection and quantification of Total Antioxidant Capacity were performed in PDEVs using a colorimetric assay, the Antioxidant Assay kit (MAK334, Sigma-Aldrich, USA). Antioxidant Assay Kit measures total antioxidant capacity in which Cu2+ is reduced by an antioxidant to Cu+. The resulting Cu+ specifically forms a colored complex with a dye reagent. The optical densities were read at 570 nm and the color intensity is proportional to TAC in the sample.
2.4. Ascorbic Acid Assay
Detection and quantification of Ascorbic Acid in PDEVs were performed using a fluorometric Ascorbic Acid Assay Kit (Sigma-Aldrich, USA). Samples were diluted in ascorbic acid buffer in a 96-well plate and subsequently to each well was added a catalyst and then reaction mix (the reaction mix is composed of an ascorbic acid buffer, ascorbic acid probe and ascorbic acid enzyme mix). After 5 min of incubation, fluorescence was read in a microplate reader at Ex/Em = 535/587 nm.
2.5. ATP Assay Kit
Total ATP was measured in PDEVs with the ATP Assay Kit (Colorimetric) (Abcam, Cambridge, UK). The assay was based on the phosphorylation of glycerol in order to generate a product that was quantified colorimetrically. After plating standard wells, sample wells, and sample background control wells at the optimal dilution, the reaction mix was added to each standard and sample well, and the background reaction mix was added to the background control sample wells. The samples were incubated at room temperature for 30 min, protected from light, and the absorbance was measured using a microplate reader at OD 570 nm.
2.6. Catalase Activity Assay
For the Catalase Activity Assay (Abcam, Cambridge, UK), a fluorometric kit was used for detection and quantification of the Catalase activity in fruit-derived nanovesicles. Briefly, samples resuspended in H2O were loaded in a 96-well plate; a stop solution was added in the control samples and incubated for 5 min at 25 °C to inhibit the Catalase activity. Catalase reaction mix (with H2O2) was added to both the control and high control samples for 30 min at 25 °C. The reaction in the high control samples and standard samples was stopped with the stop solution, the developer was added to all wells and after 10 min the fluorescence was read at Ex/Em = 535/587 nm on a microplate reader (Promega, Madison, WI, USA). Data were analyzed using the manufacturer’s instructions. One unit of Catalase corresponds to the amount of Catalase that will decompose 1 µmol of H2O2 per minute at pH 4.5 at 25 °C.
2.7. Citric Acid Assay
The Detection and quantification of Citric Acid were performed in Skin Cells Regeneration using a colorimetric assay kit, Citric Acid (CA) Colorimetric Assay Kit (MBS2563963, MyBioSource, USA).
The samples were incubated with the reaction reagents for 30 minutes at room temperature, and then the reaction product was analysed at a wavelength of 545 nm.
2.8. Reduced Glutathione (GSH) Detection and Quantification Assay
The Glutathione Colorimetric Detection Kit (Thermo Fisher, USA), a colorimetric assay, was used for detection and quantification of reduced glutathione (GSH) levels in PDEVs. Detection reagent and reaction mixture (NADPH and glutathione reductase) were added to samples and after 20 min of incubation at room temperature, the optical densities were recorded at 405 nm.
2.9. Superoxide Dismutase (SOD) Activity Assay
The Superoxide Dismutase Activity kit (Thermo Fisher, USA), a colorimetric assay, was used for detection and quantification of the superoxide dismutase activity in PDEVs’ preparations. Samples were incubated for 20 min at room temperature, after the addition of substrate and chromogenic detection reagent. The optical densities were recorded at 450 nm.
2.10. Nanoparticle Tracking Analysis
Nanoparticle Tracking Analysis (NTA) from Malvern (NanoSight NS300, UK) was used for the measurement of size distribution and concentration of extracellular vesicle samples in the liquid suspension. Five videos of typically 60 s duration were taken. Data were analyzed using the NTA 3.4 software (Malvern Instruments), which was optimized to first identify and then track each particle on a frame-by-frame basis. The Brownian motion of each particle was tracked using the Stokes–Einstein equation: D° = kT/6πηr, where D° is the diffusion coefficient, kT/6πηr = f0 is the frictional coefficient of the particle, for the special case of a spherical particle of radius r moving at a uniform velocity in a continuous fluid of viscosity η, k is Boltzmann’s constant, and T is the absolute temperature.
2.11. Dynamic Light Scattering
Dynamic Light Scattering (DLS) (Malvern Instrument, UK) was used for the evaluation of zeta potential of Plant-Derived Extracellular Vesicles. Evaluation of zeta potential allows prediction of the stability or electrostatic interactions of extracellular vesicles in dispersions.
Data from were analyzed using ZS XPLORER software (Malvern Instrument, UK).
2.12. Transmission Electron Microscopy (TEM)
The isolated PDEVs were resuspended in PBS and after a proper dilution, placed on 200 mesh carbon-coated copper grids. PDEVs were let adhere to the grids, and fixed with glutaraldehyde 2% (Electron Microscopy Sciences, USA) in PBS. Then, 3 washing steps with milliQ water were performed, followed by negative staining with phosphotungstic acid 2% solution. TEM images were captured using a Philips CM 100 Electron Microscope [60,61,62,63].
2.13. Cell line
Normal human dermal fibroblast cells (NHDFs) (Sigma-Aldrich, USA) isolated from the dermis of adult skin where cultured in Low-serum cell culture medium (Fibroblast Growth Medium 2, Sigma-Aldrich, USA) supplemented with antibiotics and antimycotics in incubator with an atmosphere of 5% CO2.
2.14. Staining Protocol of PDEVs with a fluorescent probe (DIl 1,1′-Dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine perchlorate)
To study cellular uptake of PDEVs, vesicles were marked with Dil, a fluorescent lipophilic cationic indocarbocyanine dye widely used to label membrane lipid bilayer (1,1′-Dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine perchlorate, 42364, Sigma-Aldrich, USA).
A stock solution of Dil was prepared dissolving the power in DMSO and then incubating the extracellular vesicles with a 5 µM dye solution for 30 minutes at 37°C. After incubation, the vesicles were ultracentrifugated at 110,000 rpm for one hour to remove the unlabeled dye and labelled-PDEVs were used for cellular treatment.
Fibroblasts (NHDF) were treated with Dil-labeled PDEVs and after 24 h, 48 h and 72 h of treatment, cells were fixed with paraformaldehyde (4%). The slides were counterstained with DAPI (DAPI/Antifade solution, Sigma-Aldrich, USA) and images were acquired with the Optika microscope (IM-5FLD, Optika Microscopes Italy). Mean Intensity of Fluorescence was measured using ImageJ software (National Institutes of Health of the United States).
2.15. Mitochondrial Membrane Potential Measurement
Mitochondrial membrane potential using MitoTracker® Dyes for Mitochondria Labeling (Thermo Fisher, USA), a green-fluorescent mitochondrial stain, which appears to localize to mitochondria regardless of mitochondrial membrane potential. To label mitochondria, live cells were incubated with 100 nM of MitoTracker® probes, which passively diffused across the plasma membrane and accumulated in active mitochondria. The reduced probes do not fluoresce until they enter live cells, where they are oxidized to the corresponding fluorescent mitochondrion-selective probe and then sequestered in the mitochondria. After the induction of oxidative stress (500 µM H2O2), Fibroblasts were incubated with prewarmed MitoTracker® probe staining solution at 37 °C and 5% CO2 for 30 min. After incubation cells were centrifugated and resuspended in a fresh prewarmed medium or buffer. Green fluorescence was read at Ex/Em = 490/516 using a Fluorescence Microplate Reader. Data are expressed as M.I.F. (a.u.).
2.16. Mitochondrial Superoxide Assay
To evaluate the antioxidant effect of PDEVs, NHDF were treated with 500 µM H2O2 for 24 hours to induce oxidative damages, and then with PDEVs. After 24-hour treatment, Mitochondrial superoxide was measured in cells treated with PDEVs and control untreated cells using Mitochondrial Superoxide Detection Kit (Abcam, UK). This is a sensitive fluorometric one-step assay based on MitoROS 580 dye to detect intracellular superoxide radical in live cells. The dye is cell-permeable and selectively reacts with mitochondrial superoxide present in live cells to generate a red fluorescence signal at Ex/Em = 540/590 nm. Mean Intensity of Fluorescence was measured using a Fluorescence Microplate Reader. Data are expressed as M.I.F. (a.u.).
2.17. Sirtuin quantification
Detection and quantification of Sirtuin in PDEVs were performed using an ELISA kit: SIRT1(Sirtuin1) Human SimpleStep ELISA® Kit (ab171573, Abcam, UK). To perform the assay, samples or standards are added to the wells, followed by the addiction of antibody mix. After incubation, the wells are washed to remove unbound material. TMB substrate is added and during incubation is catalyzed by HRP, generating blue coloration. This reaction is then stopped by addition of Stop Solution. Signal is generated proportionally to the amount of bound analyte and the intensity is measured at 450 nm microplate reader.
2.18. Wound healing Assay
Fibroblast cells (NHDF) were seeded in order to have a monolayer. Once the monolayer was formed, we made a scratch using a micropipette tip to mimic a wound. The cells were then treated with PDEVs and images were acquired at time 0, after 24-hour and 48-hour treatment using the Optika microscope (IM-5FLD, Optika Microscopes Italy). Wound gaps were measured using the PROview software (Optika Microscopes Italy).
2.19. Matrix Metalloproteinase-9 evaluation
As described above, we induced a scratch in the monolayer of Fibroblasts seeded on coverslips. After 24-hours treatment, cells were fixed with paraformaldehyde (4%) and stained with Monoclonal Anti-Mmp9 - FITC (SAB5200306, Sigma-Aldrich, USA). The slides were counterstained with DAPI (DAPI/Antifade solution, Sigma-Aldrich, USA) and images were acquired with the Optika microscope (IM-5FLD, Optika Microscopes Italy). Mean Intensity of Fluo-rescence was measured using ImageJ software (National Institutes of Health of the United States).
2.20. Vimentin quantification
Extracellular Vimentin was measured using VIM (Human) ELISA Kit (KA3127, Novus Biologicals, UK). This kit was based on standard sandwich enzyme-linked immune-sorbent assay technology. The purified anti-Vimentin antibody was pre-coated onto 96-well plates, then standards, test samples and anti-Vimentin HRP conjugated antibody were added to the wells subsequently, mixed and incubated. Then, unbound conjugates were washed away with wash buffer. TMB substrates (A & B) were used to visualize HRP enzymatic reaction. TMB was catalyzed by HRP to produce a blue color product that changed into yellow after adding an acidic stop solution. The density of yellow is proportional to the Vimentin amount of sample captured in plate. The O.D. absorbance were read at 450 nm in a microplate reader.
2.21. Statistical analysis
Results are reported as the means ± standard error (SE), and calculations were done using GraphPad Prism software (USA). An unpaired t-test (Student’s t-test) and one-way ANOVA Bonferroni were applied to analyze the results. Statistical significance was set at p < 0.05.
3. Results
3.1. Characterization of the PDEVs mix.
3.1.1. Bioactives’ content
We have first characterized the content in bioactives of a mix of different fruits. In our experiment we exploited a combination of PDEVs isolated from grape (Vitis vinifera), pomegranate (Punica granatum), orange (Citrus sinensis), tangerine (Citrus reticulata) and papaya (Carica papaya). The mix of these fruits has shown a very high level of bioactive compounds. PDEVs were characterized through different Colorimetric kit to detect and quantify their antioxidant cargo. In Table 1 are summarized the results obtained from the analysis of 109 PDEVs.
3.1.2. Size distribution and zeta potential analysis
Then we characterized through Nanoparticle Tracking Analysis the PDEVs’ mix in order to establish their concentration and size distribution. As reported in Figure 1a, PDEVs isolated from the above-mentioned mix of fruit extracts, have the typical size distribution of extracellular vesicles, with size equal to 189,5 ± 2,1 nm. Moreover, in order to further characterize PDEVs isolated from fruits, we used the Zetasizer to analyze the zeta potential in a liquid suspension. The zeta potential measured was -33,68 ± 1,3 mV and the results are reported in Figure 1b.
3.1.3. Morphological Characterization
PDEVs were also characterized by Transmission Electron Microscopy to better evaluate their PDEVs-like morphology [64]. In fact, this analysis, due its nanometer resolution, can efficiently distinguish Extracellular Vesicles from non-EV-particles. TEM analysis clearly showed the presence of numerous typically rounded, whole, and undamaged vesicles with a size ranging from 50-80 nm (Figure 2a) to 150-200 nm (Figure 2b) suggesting their EV-like nature. Moreover, TEM analysis demonstrated the presence of an unbroken bilayer membrane, visible as a thin white filament surrounding the electron-dense exosome content (Figure 2). Thus, our PDEVs contained a heterogeneous population in size, round in shape, and enclosed by a membrane.
3.2. PDEVs uploading into Human Skin Fibroblasts
In order to track the uptake of extracellular vesicles over time, we labelled extracellular vesicles with Dil, an orange-red lipophilic dye. This dye is usually weakly fluorescent until incorporated into membranes. The stain with lipophilic dye is usually used to label extracellular vesicles and trace their distribution in in vitro and in vivo cells [65,66,67,68,69,70].
In our experiment we stained PDEVs with Dil for 20 minutes at 37°C, then we washed unincorporated dye and used PDEVs-Dil in cells treatment for different time points. Data reported in Figure 3, showed that PDEVs were efficiently internalized in cells after 24 h (Figure 3a) and that fluorescent signal was directly proportional to treatment time. Moreover, results demonstrated that PDEVs can efficiently reach cell nucleus where they may affect the genome (Figure 3c).
3.3. Effect on mitochondrial metabolism
For the reasons reported in the introduction section, we wanted to investigate how PDEVs can mitigate progressive aging using a model of Fibroblasts treated with H2O2 [71,72]. To evaluate the effect of PDEVs in cellular metabolism, we induced an oxidative stress in in vitro Fibroblasts through H2O2 treatment and then we treated cells with the PDEVs’ mix. As shown in Figure 4a, PDEVs can restore unbalanced Mitochondrial Membrane Potential induced by oxidative stress. H2O2 treatment significantly reduced Mitochondrial Membrane Potential (447 ± 3 M.I.F.; p<0,001) compared to control cells (3434 ± 56 M.I.F.), whereas cells subjected to oxidation and then treated with PDEVs have an increased potential level (2626 ± 27 M.I.F.; p<0,01) compared to the H2O2-treated fibroblasts.
In the following series of experiments, we evaluated also the Mitochondrial Superoxide Anion levels in untreated cells, either cells undergone H2O2 or cells treated with PDEVs following H2O2. In the H2O2-treated cells we measured a 13% increase of Mitochondrial Superoxide Anion compared to control cells (p<0,05), with values equal to 578 ± 11 M.I.F. (a.u.) of untreated cells and 654 ± 6 M.I.F. (a.u.) of oxidated cells. The most significant result was observed in Fibroblasts subjected to oxidation and then treated with Extracellular Vesicles. Indeed, as shown in Figure 4b, a significant reduction (p<0,001) in superoxide anion levels was observed in the samples treated with PDEVs’ mix (65 ± 2 M.I.F.) compared to oxidated cells; in particular the anion value is 10-fold lower in PDEVs treated samples compared to human fibroblast undergone H2O2 treatment.
3.4. Effects of the PDEVs’ mix on some aging-related molecules
Sirtuin 1 is the most extensively studied member of Sirtuins family that is involved in oxidative stress response, inflammation and mitochondrial functions, as well as in the transcriptional regulation of several proteins involved in age-related processes [20]. We thus quantified the concentration of Sirtuin 1 in cell culture supernatants of human Fibroblasts treated with either H2O2 or H2O2 + PDEVs or left untreated. As reported in Figure 5, extracellular Sirtuin 1 concentration in significantly reduced after oxidation (2,32 ± 0,03 ng/ml; CTR 3,91 ± 0,07 ng/ml; p<0,0001); while the treatment of oxidated skin fibroblasts with the PDEVs’ mix efficiently increased Sirtuin 1 levels as compared to cells treated with H2O2 only (2,84 ± 0,02 ng/ml; p<0,0001).
These data showed that the PDEVs’ mix efficiently restored Sirtuin 1 levels in skin-derived Human Fibroblasts after oxidative stress, supporting the beneficial effect of PDEVs’ mix in mitigating progressive skin aging.
3.5. Skin repair: Wound healing
Fibroblasts play a crucial role in skin repair. We thus used this cellular model to investigate the effects of our PDEVs’ mix on the wound healing after the induction of a scratch in the Fibroblasts’ monolayer. Fibroblasts were seeded in 24-cell plate in order to have a uniform monolayer of cells, then we induced an injury in the monolayer and treated then with PDEVs. Images of untreated control and treated Fibroblasts were acquired after 24 and 48 hours. For in vitro evaluation of wound closure, we measured the distance between the two cell fronts. The results are summarized in Figure 6. Our experiments have shown that the PDEVs’ mix significantly speeded up wound healing at both 24 (p<0,0001) and 48 hours (p<0,0001), by reducing the distances between cells’ fronts. As showed in Figure 6, wound gap at 24 h in control cells was 412 ± 21 µm, while after the PDEVs’ mix treatment it was reduced to 165 ± 14 µm. Similarly, after 48 hours the distance between the two cells’ fronts was 148 ± 10 µm in untreated control cells compared to 16 ± 3 µm in PDEV-treated cells with an almost complete closure of the scratch after PDEVs’ mix treatment.
Wound healing repair is a complex process involving different events including mesenchymal cell differentiation, proliferation and migration to the wound site and ri-epithelization [41,73,74]. In this process Matrix Metalloproteinases (MMPs), a family of zinc-containing enzymes, play an important role in the decomposition of Extracellular Matrix after injury. Among these enzyme, MMP-9 is a fundamental protein involved in cell migration and recruitment at wound site, together with the regulation of angiogenesis process [43,44,45,46,75]. In order to investigate PDEVs’ mechanism of action in Skin Cells, we evaluated MMP-9 expression in Skin Fibroblast after the induction of an injury. Briefly, cells were seeded on coverslips in order to have a uniform monolayer, then we made a scratch and treated cells for 24 hours with PDEVs. After treatment, we evaluated MMP-9 expression through Immunofluorescence Microscopy. In Figure 7 shows images of the wound site, since it has been widely demonstrated that MMP-9 is highly overexpressed in epithelial cells at the front of the migrating epithelial sheet [43,76]. Notably, we measured a 6-fold increase of MMP-9 expression (p<0,01) after 24-hours treatment with PDEVs (4,73 ± 1,15 M.I.F.) compared to untreated control cells (0,73 ± 0,03 M.I.F).
Vimentin is another key protein involved in cellular proliferation and wound repair. In fact, Vimentin is an intermediate filament protein that participates in numerous processes including cell adhesion, migration and invasion, signaling, differentiation. In tissue repair, Vimentin regulates the migration after an injury [47,48,49,50] and its concentration is directly related to the ability of cells to close a wound [77,78,79]. We thus measured Vimentin concentration in cell culture supernatants after treatment with the PDEVs’ mix. Briefly, Fibroblasts were treated with the PDEVs’ mix for 24 hours and the concentration of Vimentin was measured in the cell culture supernatant. The results showed a significant increase of Vimentin in the supernatant of the PDEVs’ mix-treated cells compared to control cell cultures (Figure 8). Notably we found a strong increase of Vimentin in the PDEVs’ mix treated human skin fibroblasts cell cultures (CTR: 56 ± 2,4 ng/ml, PDEVs: 71 ± 1,3 ng/ml; p<0,0001).
These results highly supported the results obtained in the wound healing experiments.
4. Discussion
PDEVs naturally contain a wide array of molecules, including proteins, lipids and nucleic acids, that are all involved in the intercellular communication within the same species and between the different species and nature kingdoms. In fact, PDEVs contain antioxidant molecules that can moderate oxidative stress damages, scavenging ROS and enhancing antioxidant defense systems of cells [80]. PDEVs can efficiently influence the expression of genes involved in skin health, such as regeneration, skin barrier, moisturization and aging-related genes [81]. Taking into account these considerations, we combine different fruit extracts widely characterized to exert beneficial effect in skin regeneration and mitigation of oxidative stress damages [82,83,84,85,86,87,88,89,90,91]
First of all, we characterized PDEVs bioactive molecules, detecting a huge amount of antioxidants; including both enzymatic bioactives such Superoxide Dismutase and Catalase and non-enzymatic ones such as Glutathione and Ascorbic Acid. Moreover, ATP was detected in Extracellular Vesicles and it demonstrated not only its role as energy source but also as signaling molecule that mediates interactions between organs and systems [92]. Consistently with other works showing that ATP positively influence skin cell proliferation, DNA repair and collagen synthesis [79,93], we assume that ATP in PDEVs could positively exert beneficial regenerative action, in combination with antioxidant naturally contained into the PDEVs. Moreover, skin cells have a high turnover paying the price of a high energy consumption [93,94,95]. Thus, we hypothesized that the ATP supplementation from PDEVs could be helpful in maintaining physiological skin cell turnover, keeping cells proliferating and highly producing molecules involved in regenerative processes.
In this study we set up a PDEVs’ mix that, while combining a heterogeneous population of extracellular vesicles in term of size distribution, confirmed by NTA and TEM analysis, has proven highly stable in liquid suspension, also confirming previous reports [34,35,96]. Moreover, the PDEVs used in this study had a very negative zeta potential. Notably, the zeta potential value characterized the fusogenic properties of PDEVs with plasma membrane of target/recipient cells, and the more negative is the zeta potential of the PDEVs the more fusogenic they are. Based on this background, we analyzed the cellular uptake of our PDEVs mix by human skin fibroblasts. To this purpose we labelled PDEVs with a fluorescent probe that specifically bound the membrane phospholipids of the extracellular vesicles. The labelled PDEVs were detectable in the target Fibroblasts up to 24 hours after treatment, with an increase of the fluorescence values in the following 48 and 72 hours. Furthermore after 72 hours of treatment the signal of the PDEVs overlaps with that of the nuclei, suggesting that PDEVs are able to effectively reach the genome of the target cells, possibly influencing the gene expression of recipient cells. This set of results witnessed the ability of PDEVs to be entirely uploaded into the target cells where they fully exert their action.
Damages induced by oxidative stress can affect virtually all the cellular pathways, in turn inducing a heavy unbalance of the cellular homeostasis, of course including mitochondrial activities as well. To reproduce this condition, we treated Human Skin Cell Fibroblasts with hydrogen peroxide, that induces a redox imbalance into the cells; following the H2O2 treatment we first tested the ability of our PDEVs mix to revert the imbalanced Mitochondria Membrane Potential, that is the most important indicator of Mitochondria health. The results showed that PDEVs can efficiently restore the physiological mitochondrial membrane potential after oxidation stimuli, from the first 24 hours after treatment. Supporting this result, PDEVs reduce the production of Mitochondrial Superoxide Anion in treated cells, further demonstrating their beneficial effect on mitochondria metabolism. Also, the increase of oxidative stress damages is related to the decreased production of molecules involved in age-related processes, such as Sirtuin family of proteins. Thus, we compared the levels of extracellular Sirtuin 1 before and after treatment of oxidated cells with PDEVs, showing that that PDEVs increased the extracellular sirtuin levels in treated cultures, thus further supporting their skin anti-aging effect.
In a further series of experiments, we wanted to directly show the ability of our PDEVs mix to influence cellular mechanisms involved in the skin regenerative processes. To this purpose we used the model of wound healing in cultures of human Skin Fibroblasts. We first showed the ability of our PDEVs mix to lead to a full closure of the wound produced into the human skin fibroblasts’ monolayer. Then we showed in the same model the wound repair was associated to the increase of MMP-9 concentrations, a protein actively involved in cellular repair mechanism. Of interest, in our experiments, MMP-9 was more expressed in the fibroblasts’ cultures treated with PDEVs, particularly at the wound site, where the wound closure actually started. In the same cultures we found a higher concentration of extracellular Vimentin in Fibroblasts treated with the PDEVs mix as compared to untreated Fibroblasts. Consistently with previous reports [97] we assumed that the polyphenols contained in our PDEVs mix could well contribute to the wound healing we showed in our experiments.
All in all the results of our study provided a scientific evidence that a mix of PDEVs from different fruits from organic farming may have a clear beneficial effect on skin aging processes. This effect passes through a clear anti-oxidant action due to the high levels of anti-oxidant molecules contained into the PDEVs mix. Here we powerfully show that the PDEVs effect was due to a full uploading of the extracellular vesicles into the target cells. Our results are also supported by previous reports showing that Plant-derived Extracellular Vesicles can efficiently penetrate to recipient cells, decrease levels of oxidants species and positively regulate the regenerative processes [52,53,80,98,99,100,101,102].
5. Conclusions
Here we show that PDEVs isolated and concentrated from a mix of fruit extracts (Grape, Blood Orange, Tangerine, Papaya and Pomegranate) contain different detectable antioxidants (Superoxide Dismutase, Catalase, Glutathione and Ascorbic Acid). These vesicles are efficiently uptaken by human Skin Fibroblasts, where they mitigate aging processes related to oxidative damages, restoring the physiological mitochondrial function and increasing the concentration of Sirtuin 1, whose principal role is the regulation of age-related processes. Moreover, here we show that PDEVs are actively involved in maintaining the skin's barrier function, assisting wound repair by also modulating the expression of MMP-9 and Vimentin, essential proteins involved in tissue repair processes.
We believe that our results will contribute in positively changing the approach of local treatment of skin aging through the set-up of entirely new products, but implementing the local treatments of skin wounds. We can’t exclude that the local anti-oxidant effect of our PDEVs mixes may be improved by the systemic treatment with the same mixes, that have shown to burst an antioxidant reaction in the whole body [34].
Author Contributions
Conceptualization, Rossella Di Raimo, Stefano Fais and Mariantonia Logozzi; Data curation, Rossella Di Raimo, Davide Mizzoni, Antonella Aloi, Giulia Pietrangelo, Vincenza Dolo, Giuseppina Poppa and Mariantonia Logozzi; Formal analysis, Rossella Di Raimo, Davide Mizzoni, Antonella Aloi, Giulia Pietrangelo, Giuseppina Poppa and Mariantonia Logozzi; Funding acquisition, Stefano Fais and Mariantonia Logozzi; Investigation, Rossella Di Raimo, Davide Mizzoni, Antonella Aloi, Giulia Pietrangelo and Mariantonia Logozzi; Methodology, Rossella Di Raimo, Davide Mizzoni, Antonella Aloi, Giulia Pietrangelo, Vincenza Dolo, Giuseppina Poppa and Mariantonia Logozzi; Project administration, Stefano Fais and Mariantonia Logozzi; Resources, Stefano Fais and Mariantonia Logozzi; Software, Rossella Di Raimo; Supervision, Vincenza Dolo, Stefano Fais and Mariantonia Logozzi; Validation, Rossella Di Raimo, Davide Mizzoni and Vincenza Dolo; Visualization, Rossella Di Raimo; Writing – original draft, Rossella Di Raimo, Stefano Fais and Mariantonia Logozzi; Writing – review & editing, Rossella Di Raimo, Stefano Fais and Mariantonia Logozzi. All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
Rossella Di Raimo and Davide Mizzoni performed all the experiments at Department of Oncology and Molecular Medicine, Istituto Superiore di Sanità (Rome) and completed data analysis during their employment at ExoLab Italia (L'Aquila). Antonella Aloi and Giulia Pietrangelo are employees of ExoLab Italia that contributed to manuscript drafting.
References
- Abels, C.; Angelova-Fischer, I. Skin Care Products: Age-Appropriate Cosmetics. Curr Probl Dermatol 2018, 54, 173–182. [Google Scholar] [CrossRef]
- Chen, J.; Liu, Y.; Zhao, Z.; Qiu, J. Oxidative Stress in the Skin: Impact and Related Protection. Intern J of Cosmetic Sci 2021, 43, 495–509. [Google Scholar] [CrossRef] [PubMed]
- Yosipovitch, G.; Misery, L.; Proksch, E.; Metz, M.; Ständer, S.; Schmelz, M. Skin Barrier Damage and Itch: Review of Mechanisms, Topical Management and Future Directions. Acta Derm Venereol 2019, 99, 1201–1209. [Google Scholar] [CrossRef]
- Nilforoushzadeh, M.A.; Amirkhani, M.A.; Zarrintaj, P.; Salehi Moghaddam, A.; Mehrabi, T.; Alavi, S.; Mollapour Sisakht, M. Skin Care and Rejuvenation by Cosmeceutical Facial Mask. J of Cosmetic Dermatology 2018, 17, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Hoang, H.T.; Moon, J.-Y.; Lee, Y.-C. Natural Antioxidants from Plant Extracts in Skincare Cosmetics: Recent Applications, Challenges and Perspectives. Cosmetics 2021, 8, 106. [Google Scholar] [CrossRef]
- Gilchrest, B.A. Skin Aging and Photoaging: An Overview. J Am Acad Dermatol 1989, 21, 610–613. [Google Scholar] [CrossRef]
- Chen, X.; Yang, C.; Jiang, G. Research Progress on Skin Photoaging and Oxidative Stress. Postepy Dermatol Alergol 2021, 38, 931–936. [Google Scholar] [CrossRef] [PubMed]
- Papaccio, F.; D’Arino, A.; Caputo, S.; Bellei, B. Focus on the Contribution of Oxidative Stress in Skin Aging. Antioxidants 2022, 11, 1121. [Google Scholar] [CrossRef]
- Spinelli, J.B.; Haigis, M.C. The Multifaceted Contributions of Mitochondria to Cellular Metabolism. Nat Cell Biol 2018, 20, 745–754. [Google Scholar] [CrossRef]
- Zorova, L.D.; Popkov, V.A.; Plotnikov, E.Y.; Silachev, D.N.; Pevzner, I.B.; Jankauskas, S.S.; Babenko, V.A.; Zorov, S.D.; Balakireva, A.V.; Juhaszova, M.; et al. Mitochondrial Membrane Potential. Analytical Biochemistry 2018, 552, 50–59. [Google Scholar] [CrossRef]
- Perry, S.W.; Norman, J.P.; Barbieri, J.; Brown, E.B.; Gelbard, H.A. Mitochondrial Membrane Potential Probes and the Proton Gradient: A Practical Usage Guide. BioTechniques 2011, 50, 98–115. [Google Scholar] [CrossRef] [PubMed]
- Izyumov, D.S.; Avetisyan, A.V.; Pletjushkina, O.Y.; Sakharov, D.V.; Wirtz, K.W.; Chernyak, B.V.; Skulachev, V.P. “Wages of Fear”: Transient Threefold Decrease in Intracellular ATP Level Imposes Apoptosis. Biochimica et Biophysica Acta (BBA) - Bioenergetics 2004, 1658, 141–147. [Google Scholar] [CrossRef]
- Starkov, A.A.; Fiskum, G. Regulation of Brain Mitochondrial H 2 O 2 Production by Membrane Potential and NAD(P)H Redox State. Journal of Neurochemistry 2003, 86, 1101–1107. [Google Scholar] [CrossRef] [PubMed]
- Skulachev, V.P. Role of Uncoupled and Non-Coupled Oxidations in Maintenance of Safely Low Levels of Oxygen and Its One-Electron Reductants. Quart. Rev. Biophys. 1996, 29, 169–202. [Google Scholar] [CrossRef] [PubMed]
- Tirichen, H.; Yaigoub, H.; Xu, W.; Wu, C.; Li, R.; Li, Y. Mitochondrial Reactive Oxygen Species and Their Contribution in Chronic Kidney Disease Progression Through Oxidative Stress. Front. Physiol. 2021, 12, 627837. [Google Scholar] [CrossRef] [PubMed]
- Cadenas, E.; Davies, K.J.A. Mitochondrial Free Radical Generation, Oxidative Stress, and aging11This Article Is Dedicated to the Memory of Our Dear Friend, Colleague, and Mentor Lars Ernster (1920–1998), in Gratitude for All He Gave to Us. Free Radical Biology and Medicine 2000, 29, 222–230. [Google Scholar] [CrossRef]
- Brand, M.D.; Affourtit, C.; Esteves, T.C.; Green, K.; Lambert, A.J.; Miwa, S.; Pakay, J.L.; Parker, N. Mitochondrial Superoxide: Production, Biological Effects, and Activation of Uncoupling Proteins. Free Radical Biology and Medicine 2004, 37, 755–767. [Google Scholar] [CrossRef]
- Han, D.; Williams, E.; Cadenas, E. Mitochondrial Respiratory Chain-Dependent Generation of Superoxide Anion and Its Release into the Intermembrane Space. Biochemical Journal 2001, 353, 411–416. [Google Scholar] [CrossRef]
- Bazyluk, A.; Malyszko, J.; Hryszko, T.; Zbroch, E. State of the Art - Sirtuin 1 in Kidney Pathology - Clinical Relevance. Adv Med Sci 2019, 64, 356–364. [Google Scholar] [CrossRef]
- Bielach-Bazyluk, A.; Zbroch, E.; Mysliwiec, H.; Rydzewska-Rosolowska, A.; Kakareko, K.; Flisiak, I.; Hryszko, T. Sirtuin 1 and Skin: Implications in Intrinsic and Extrinsic Aging-A Systematic Review. Cells 2021, 10, 813. [Google Scholar] [CrossRef]
- Zerr, P.; Palumbo-Zerr, K.; Huang, J.; Tomcik, M.; Sumova, B.; Distler, O.; Schett, G.; Distler, J.H.W. Sirt1 Regulates Canonical TGF-β Signalling to Control Fibroblast Activation and Tissue Fibrosis. Ann Rheum Dis 2016, 75, 226–233. [Google Scholar] [CrossRef]
- Michan, S.; Sinclair, D. Sirtuins in Mammals: Insights into Their Biological Function. Biochemical Journal 2007, 404, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Guarente, L. Sirtuins in Aging and Disease. Cold Spring Harbor Symposia on Quantitative Biology 2007, 72, 483–488. [Google Scholar] [CrossRef]
- Caldeira, C.A.; Santos, M.A.; Araújo, G.R.; Lara, R.C.; Franco, F.N.; Chaves, M.M. Resveratrol: Change of SIRT 1 and AMPK Signaling Pattern during the Aging Process. Experimental Gerontology 2021, 146, 111226. [Google Scholar] [CrossRef] [PubMed]
- Truong, V.; Jun, M.; Jeong, W. Role of Resveratrol in Regulation of Cellular Defense Systems against Oxidative Stress. BioFactors 2018, 44, 36–49. [Google Scholar] [CrossRef]
- Najafi, M.; Nikpayam, O.; Tavakoli-Rouzbehani, O.M.; Papi, S.; Amrollahi Bioky, A.; Ahmadiani, E.S.; Sohrab, G. A Comprehensive Insight into the Potential Effects of Resveratrol Supplementation on SIRT-1: A Systematic Review. Diabetes & Metabolic Syndrome: Clinical Research & Reviews 2021, 15, 102224. [Google Scholar] [CrossRef]
- Iside, C.; Scafuro, M.; Nebbioso, A.; Altucci, L. SIRT1 Activation by Natural Phytochemicals: An Overview. Front. Pharmacol. 2020, 11, 1225. [Google Scholar] [CrossRef] [PubMed]
- Wiciński, M.; Erdmann, J.; Nowacka, A.; Kuźmiński, O.; Michalak, K.; Janowski, K.; Ohla, J.; Biernaciak, A.; Szambelan, M.; Zabrzyński, J. Natural Phytochemicals as SIRT Activators—Focus on Potential Biochemical Mechanisms. Nutrients 2023, 15, 3578. [Google Scholar] [CrossRef]
- Singh, C.K.; Chhabra, G.; Ndiaye, M.A.; Garcia-Peterson, L.M.; Mack, N.J.; Ahmad, N. The Role of Sirtuins in Antioxidant and Redox Signaling. Antioxidants & Redox Signaling 2018, 28, 643–661. [Google Scholar] [CrossRef]
- Salehi, B.; Azzini, E.; Zucca, P.; Maria Varoni, E.; Anil Kumar, N.V.; Dini, L.; Panzarini, E.; Rajkovic, J.; Valere Tsouh Fokou, P.; Peluso, I.; et al. Plant-Derived Bioactives and Oxidative Stress-Related Disorders: A Key Trend towards Healthy Aging and Longevity Promotion. Applied Sciences 2020, 10, 947. [Google Scholar] [CrossRef]
- Morgan, C.; Nigam, Y. Naturally Derived Factors and Their Role in the Promotion of Angiogenesis for the Healing of Chronic Wounds. Angiogenesis 2013, 16, 493–502. [Google Scholar] [CrossRef]
- Cui, X.; Lin, Q.; Liang, Y. Plant-Derived Antioxidants Protect the Nervous System From Aging by Inhibiting Oxidative Stress. Front. Aging Neurosci. 2020, 12, 209. [Google Scholar] [CrossRef] [PubMed]
- Roleira, F.M.F.; Tavares-da-Silva, E.J.; Varela, C.L.; Costa, S.C.; Silva, T.; Garrido, J.; Borges, F. Plant Derived and Dietary Phenolic Antioxidants: Anticancer Properties. Food Chem 2015, 183, 235–258. [Google Scholar] [CrossRef]
- Di Raimo, R.; Mizzoni, D.; Spada, M.; Dolo, V.; Fais, S.; Logozzi, M. Oral Treatment with Plant-Derived Exosomes Restores Redox Balance in H2O2-Treated Mice. Antioxidants 2023, 12, 1169. [Google Scholar] [CrossRef]
- Logozzi, M.; Di Raimo, R.; Mizzoni, D.; Fais, S. Nanovesicles from Organic Agriculture-Derived Fruits and Vegetables: Characterization and Functional Antioxidant Content. IJMS 2021, 22, 8170. [Google Scholar] [CrossRef] [PubMed]
- Logozzi, M.; Di Raimo, R.; Mizzoni, D.; Fais, S. Anti-Aging and Anti-Tumor Effect of FPP® Supplementation. Eur J Transl Myol 2020, 30, 8905. [Google Scholar] [CrossRef]
- Logozzi, M.; Mizzoni, D.; Di Raimo, R.; Macchia, D.; Spada, M.; Fais, S. Oral Administration of Fermented Papaya (FPP®) Controls the Growth of a Murine Melanoma through the In Vivo Induction of a Natural Antioxidant Response. Cancers (Basel) 2019, 11, 118. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Wan, F.; Su, W.; Xie, W. Research Progress on Skin Aging and Active Ingredients. Molecules 2023, 28, 5556. [Google Scholar] [CrossRef]
- Liu, H.-M.; Cheng, M.-Y.; Xun, M.-H.; Zhao, Z.-W.; Zhang, Y.; Tang, W.; Cheng, J.; Ni, J.; Wang, W. Possible Mechanisms of Oxidative Stress-Induced Skin Cellular Senescence, Inflammation, and Cancer and the Therapeutic Potential of Plant Polyphenols. IJMS 2023, 24, 3755. [Google Scholar] [CrossRef]
- Addis, R.; Cruciani, S.; Santaniello, S.; Bellu, E.; Sarais, G.; Ventura, C.; Maioli, M.; Pintore, G. Fibroblast Proliferation and Migration in Wound Healing by Phytochemicals: Evidence for a Novel Synergic Outcome. Int. J. Med. Sci. 2020, 17, 1030–1042. [Google Scholar] [CrossRef]
- Cialdai, F.; Risaliti, C.; Monici, M. Role of Fibroblasts in Wound Healing and Tissue Remodeling on Earth and in Space. Front Bioeng Biotechnol 2022, 10, 958381. [Google Scholar] [CrossRef] [PubMed]
- Knoedler, S.; Broichhausen, S.; Guo, R.; Dai, R.; Knoedler, L.; Kauke-Navarro, M.; Diatta, F.; Pomahac, B.; Machens, H.-G.; Jiang, D.; et al. Fibroblasts – the Cellular Choreographers of Wound Healing. Front. Immunol. 2023, 14, 1233800. [Google Scholar] [CrossRef]
- Manuel, J.A.; Gawronska-Kozak, B. Matrix Metalloproteinase 9 (MMP-9) Is Upregulated during Scarless Wound Healing in Athymic Nude Mice. Matrix Biology 2006, 25, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Caley, M.P.; Martins, V.L.C.; O’Toole, E.A. Metalloproteinases and Wound Healing. Adv Wound Care (New Rochelle) 2015, 4, 225–234. [Google Scholar] [CrossRef]
- Auf Dem Keller, U.; Sabino, F. Matrix Metalloproteinases in Impaired Wound Healing. MNM 2015, 1. [Google Scholar] [CrossRef]
- Kandhwal, M.; Behl, T.; Singh, S.; Sharma, N.; Arora, S.; Bhatia, S.; Al-Harrasi, A.; Sachdeva, M.; Bungau, S. Role of Matrix Metalloproteinase in Wound Healing. Am J Transl Res 2022, 14, 4391–4405. [Google Scholar]
- Ivaska, J.; Pallari, H.-M.; Nevo, J.; Eriksson, J.E. Novel Functions of Vimentin in Cell Adhesion, Migration, and Signaling. Experimental Cell Research 2007, 313, 2050–2062. [Google Scholar] [CrossRef]
- Mendez, M.G.; Kojima, S.-I.; Goldman, R.D. Vimentin Induces Changes in Cell Shape, Motility, and Adhesion during the Epithelial to Mesenchymal Transition. FASEB J 2010, 24, 1838–1851. [Google Scholar] [CrossRef]
- Dave, J.M.; Bayless, K.J. Vimentin as an Integral Regulator of Cell Adhesion and Endothelial Sprouting. Microcirculation 2014, 21, 333–344. [Google Scholar] [CrossRef]
- Sliogeryte, K.; Gavara, N. Vimentin Plays a Crucial Role in Fibroblast Ageing by Regulating Biophysical Properties and Cell Migration. Cells 2019, 8, 1164. [Google Scholar] [CrossRef]
- Yang, C.; Zhang, W.; Bai, M.; Luo, Q.; Zheng, Q.; Xu, Y.; Li, X.; Jiang, C.; Cho, W.C.; Fan, Z. Edible Plant-Derived Extracellular Vesicles Serve as Promising Therapeutic Systems. Nano TransMed 2023, 2, 100004. [Google Scholar] [CrossRef]
- Şahin, F.; Koçak, P.; Güneş, M.Y.; Özkan, İ.; Yıldırım, E.; Kala, E.Y. In Vitro Wound Healing Activity of Wheat-Derived Nanovesicles. Appl Biochem Biotechnol 2019, 188, 381–394. [Google Scholar] [CrossRef] [PubMed]
- Savcı, Y.; Kırbaş, O.K.; Bozkurt, B.T.; Abdik, E.A.; Taşlı, P.N.; Şahin, F.; Abdik, H. Grapefruit-Derived Extracellular Vesicles as a Promising Cell-Free Therapeutic Tool for Wound Healing. Food Funct. 2021, 12, 5144–5156. [Google Scholar] [CrossRef]
- Dunaway, S.; Odin, R.; Zhou, L.; Ji, L.; Zhang, Y.; Kadekaro, A.L. Natural Antioxidants: Multiple Mechanisms to Protect Skin From Solar Radiation. Front. Pharmacol. 2018, 9, 392. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, B.; Hota, M.; Pradhan, J. Skin Aging: Implications of UV Radiation, Reactive Oxygen Species and Natural Antioxidants. In Biochemistry; Ahmad, R., Ed.; IntechOpen, 2022; Vol. 28 ISBN 978-1-83968-281-0.
- Michalak, M. Plant-Derived Antioxidants: Significance in Skin Health and the Ageing Process. Int J Mol Sci 2022, 23, 585. [Google Scholar] [CrossRef] [PubMed]
- Varani, J.; Warner, R.L.; Gharaee-Kermani, M.; Phan, S.H.; Kang, S.; Chung, J.; Wang, Z.; Datta, S.C.; Fisher, G.J.; Voorhees, J.J. Vitamin A Antagonizes Decreased Cell Growth and Elevated Collagen-Degrading Matrix Metalloproteinases and Stimulates Collagen Accumulation in Naturally Aged Human Skin1. Journal of Investigative Dermatology 2000, 114, 480–486. [Google Scholar] [CrossRef]
- Boo, Y.C. Ascorbic Acid (Vitamin C) as a Cosmeceutical to Increase Dermal Collagen for Skin Antiaging Purposes: Emerging Combination Therapies. Antioxidants 2022, 11, 1663. [Google Scholar] [CrossRef]
- Pullar, J.; Carr, A.; Vissers, M. The Roles of Vitamin C in Skin Health. Nutrients 2017, 9, 866. [Google Scholar] [CrossRef]
- Kilasoniya, A.; Garaeva, L.; Shtam, T.; Spitsyna, A.; Putevich, E.; Moreno-Chamba, B.; Salazar-Bermeo, J.; Komarova, E.; Malek, A.; Valero, M.; et al. Potential of Plant Exosome Vesicles from Grapefruit (Citrus × Paradisi) and Tomato (Solanum Lycopersicum) Juices as Functional Ingredients and Targeted Drug Delivery Vehicles. Antioxidants 2023, 12, 943. [Google Scholar] [CrossRef]
- Nemidkanam, V.; Chaichanawongsaroj, N. Characterizing Kaempferia Parviflora Extracellular Vesicles, a Nanomedicine Candidate. PLoS ONE 2022, 17, e0262884. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, S.; Koo, Y.; Yang, A.; Dai, Y.; Khant, H.; Osman, S.R.; Chowdhury, M.; Wei, H.; Li, Y.; et al. Characterization of and Isolation Methods for Plant Leaf Nanovesicles and Small Extracellular Vesicles. Nanomedicine: Nanotechnology, Biology and Medicine 2020, 29, 102271. [Google Scholar] [CrossRef] [PubMed]
- Bruno, S.P.; Paolini, A.; D’Oria, V.; Sarra, A.; Sennato, S.; Bordi, F.; Masotti, A. Extracellular Vesicles Derived From Citrus Sinensis Modulate Inflammatory Genes and Tight Junctions in a Human Model of Intestinal Epithelium. Front Nutr 2021, 8, 778998. [Google Scholar] [CrossRef] [PubMed]
- Rikkert, L.G.; Nieuwland, R.; Terstappen, L.W.M.M.; Coumans, F. a. W. Quality of Extracellular Vesicle Images by Transmission Electron Microscopy Is Operator and Protocol Dependent. J Extracell Vesicles 2019, 8, 1555419. [Google Scholar] [CrossRef] [PubMed]
- Bao, C.; Xiang, H.; Chen, Q.; Zhao, Y.; Gao, Q.; Huang, F.; Mao, L. A Review of Labeling Approaches Used in Small Extracellular Vesicles Tracing and Imaging. Int J Nanomedicine 2023, 18, 4567–4588. [Google Scholar] [CrossRef] [PubMed]
- Nikiforova, N.; Chumachenko, M.; Nazarova, I.; Zabegina, L.; Slyusarenko, M.; Sidina, E.; Malek, A. CM-Dil Staining and SEC of Plasma as an Approach to Increase Sensitivity of Extracellular Nanovesicles Quantification by Bead-Assisted Flow Cytometry. Membranes (Basel) 2021, 11, 526. [Google Scholar] [CrossRef]
- Viveiros, A.; Kadam, V.; Monyror, J.; Morales, L.C.; Pink, D.; Rieger, A.M.; Sipione, S.; Posse De Chaves, E. In-Cell Labeling Coupled to Direct Analysis of Extracellular Vesicles in the Conditioned Medium to Study Extracellular Vesicles Secretion with Minimum Sample Processing and Particle Loss. Cells 2022, 11, 351. [Google Scholar] [CrossRef]
- Lau, S.Y.; Kang, M.; Hisey, C.L.; Chamley, L.W. Studying Exogenous Extracellular Vesicle Biodistribution by in Vivo Fluorescence Microscopy. Dis Model Mech 2023, 16, dmm050074. [Google Scholar] [CrossRef]
- Zhdanova, D.Y.; Poltavtseva, R.A.; Svirshchevskaya, E.V.; Bobkova, N.V. Effect of Intranasal Administration of Multipotent Mesenchymal Stromal Cell Exosomes on Memory of Mice in Alzheimer’s Disease Model. Bull Exp Biol Med 2021, 170, 575–582. [Google Scholar] [CrossRef]
- Aimaletdinov, A.M.; Gomzikova, M.O. Tracking of Extracellular Vesicles’ Biodistribution: New Methods and Approaches. IJMS 2022, 23, 11312. [Google Scholar] [CrossRef]
- Hahn, H.J.; Kim, K.B.; An, I.-S.; Ahn, K.J.; Han, H.J. Protective Effects of Rosmarinic Acid against Hydrogen Peroxide-Induced Cellular Senescence and the Inflammatory Response in Normal Human Dermal Fibroblasts. Molecular Medicine Reports 2017, 16, 9763–9769. [Google Scholar] [CrossRef]
- Zhang, J.; Yu, H.; Man, M.; Hu, L. Aging in the Dermis: Fibroblast Senescence and Its Significance. Aging Cell 2023, e14054. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Mobashery, S.; Chang, M. Roles of Matrix Metalloproteinases in Cutaneous Wound Healing. In Wound Healing - New insights into Ancient Challenges; Alexandrescu, V.A., Ed.; InTech, 2016 ISBN 978-953-51-2678-2.
- P., B. Wound Healing and the Role of Fibroblasts. J Wound Care 2013, 22, 407–412. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, H.; Yan, L.; Du, W.; Zhang, M.; Chen, H.; Zhang, L.; Li, G.; Li, J.; Dong, Y.; et al. MMP-2 and MMP-9 Contribute to the Angiogenic Effect Produced by Hypoxia/15-HETE in Pulmonary Endothelial Cells. Journal of Molecular and Cellular Cardiology 2018, 121, 36–50. [Google Scholar] [CrossRef] [PubMed]
- Legrand, C.; Gilles, C.; Zahm, J.M.; Polette, M.; Buisson, A.C.; Kaplan, H.; Birembaut, P.; Tournier, J.M. Airway Epithelial Cell Migration Dynamics. MMP-9 Role in Cell-Extracellular Matrix Remodeling. J Cell Biol 1999, 146, 517–529. [Google Scholar] [CrossRef] [PubMed]
- Menko, A.S.; Bleaken, B.M.; Libowitz, A.A.; Zhang, L.; Stepp, M.A.; Walker, J.L. A Central Role for Vimentin in Regulating Repair Function during Healing of the Lens Epithelium. Mol Biol Cell 2014, 25, 776–790. [Google Scholar] [CrossRef] [PubMed]
- Nieminen, M.; Henttinen, T.; Merinen, M.; Marttila-Ichihara, F.; Eriksson, J.E.; Jalkanen, S. Vimentin Function in Lymphocyte Adhesion and Transcellular Migration. Nat Cell Biol 2006, 8, 156–162. [Google Scholar] [CrossRef]
- Colucci-Guyon, E.; Portier, M.M.; Dunia, I.; Paulin, D.; Pournin, S.; Babinet, C. Mice Lacking Vimentin Develop and Reproduce without an Obvious Phenotype. Cell 1994, 79, 679–694. [Google Scholar] [CrossRef]
- Kim, M.; Jang, H.; Kim, W.; Kim, D.; Park, J.H. Therapeutic Applications of Plant-Derived Extracellular Vesicles as Antioxidants for Oxidative Stress-Related Diseases. Antioxidants 2023, 12, 1286. [Google Scholar] [CrossRef]
- Cho, J.H.; Hong, Y.D.; Kim, D.; Park, S.J.; Kim, J.S.; Kim, H.-M.; Yoon, E.J.; Cho, J.-S. Confirmation of Plant-Derived Exosomes as Bioactive Substances for Skin Application through Comparative Analysis of Keratinocyte Transcriptome. Appl Biol Chem 2022, 65, 8. [Google Scholar] [CrossRef]
- Sharafan, M.; Malinowska, M.A.; Ekiert, H.; Kwaśniak, B.; Sikora, E.; Szopa, A. Vitis Vinifera (Vine Grape) as a Valuable Cosmetic Raw Material. Pharmaceutics 2023, 15, 1372. [Google Scholar] [CrossRef]
- Apraj, V.D.; Pandita, N.S. Evaluation of Skin Anti-Aging Potential of Citrus Reticulata Blanco Peel. Pharmacognosy Res 2016, 8, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.-D.; Lu, Y.-N.; Liu, Q.; Shi, X.-M.; Tian, J. Modulation of TRPV1 Function by Citrus Reticulata (Tangerine) Fruit Extract for the Treatment of Sensitive Skin. J Cosmet Dermatol 2023, 22, 1369–1376. [Google Scholar] [CrossRef]
- Ferreira, S.S.; Silva, A.M.; Nunes, F.M. Citrus Reticulata Blanco Peels as a Source of Antioxidant and Anti-Proliferative Phenolic Compounds. Industrial Crops and Products 2018, 111, 141–148. [Google Scholar] [CrossRef]
- Kong, Y.R.; Jong, Y.X.; Balakrishnan, M.; Bok, Z.K.; Weng, J.K.K.; Tay, K.C.; Goh, B.H.; Ong, Y.S.; Chan, K.G.; Lee, L.H.; et al. Beneficial Role of Carica Papaya Extracts and Phytochemicals on Oxidative Stress and Related Diseases: A Mini Review. Biology (Basel) 2021, 10, 287. [Google Scholar] [CrossRef]
- Wattanapitayakul, S.K.; Jarisarapurin, W.; Kunchana, K.; Setthawong, V.; Chularojmontri, L. Unripe Carica Papaya Fresh Fruit Extract Protects against Methylglyoxal-Mediated Aging in Human Dermal Skin Fibroblasts. Prev Nutr Food Sci 2023, 28, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Lim, C.Y.; Jung, J.I.; Kim, T.Y.; Kim, E.J. Protective Effects of Red Orange ( Citrus Sinensis [L.] Osbeck [Rutaceae]) Extract against UVA-B Radiation-Induced Photoaging in Skh:HR-2 Mice. Nutr Res Pract 2023, 17, 641. [Google Scholar] [CrossRef]
- Nobile, V.; Burioli, A.; Yu, S.; Zhifeng, S.; Cestone, E.; Insolia, V.; Zaccaria, V.; Malfa, G.A. Photoprotective and Antiaging Effects of a Standardized Red Orange (Citrus Sinensis (L.) Osbeck) Extract in Asian and Caucasian Subjects: A Randomized, Double-Blind, Controlled Study. Nutrients 2022, 14, 2241. [Google Scholar] [CrossRef]
- Dimitrijevic, J.; Tomovic, M.; Bradic, J.; Petrovic, A.; Jakovljevic, V.; Andjic, M.; Živković, J.; Milošević, S.Đ.; Simanic, I.; Dragicevic, N. Punica Granatum L. (Pomegranate) Extracts and Their Effects on Healthy and Diseased Skin. Pharmaceutics 2024, 16, 458. [Google Scholar] [CrossRef] [PubMed]
- Akbarnejad, F. Dermatology Benefits of Punica Granatum: A Review of the Potential Benefits of Punica Granatum in Skin Disorders. Asain J. Green Chem. 2023, 7. [Google Scholar] [CrossRef]
- Lombardi, M.; Gabrielli, M.; Adinolfi, E.; Verderio, C. Role of ATP in Extracellular Vesicle Biogenesis and Dynamics. Front. Pharmacol. 2021, 12, 654023. [Google Scholar] [CrossRef]
- Sreedhar, A.; Aguilera-Aguirre, L.; Singh, K.K. Mitochondria in Skin Health, Aging, and Disease. Cell Death Dis 2020, 11, 444. [Google Scholar] [CrossRef] [PubMed]
- Chiang, B.; Essick, E.; Ehringer, W.; Murphree, S.; Hauck, M.A.; Li, M.; Chien, S. Enhancing Skin Wound Healing by Direct Delivery of Intracellular Adenosine Triphosphate. Am J Surg 2007, 193, 213–218. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Q.; Wan, R.; Mo, Y.; Li, M.; Tseng, M.T.; Chien, S. Intracellular Adenosine Triphosphate Delivery Enhanced Skin Wound Healing in Rabbits. Ann Plast Surg 2009, 62, 180–186. [Google Scholar] [CrossRef]
- Akuma, P.; Okagu, O.D.; Udenigwe, C.C. Naturally Occurring Exosome Vesicles as Potential Delivery Vehicle for Bioactive Compounds. Front. Sustain. Food Syst. 2019, 3, 23. [Google Scholar] [CrossRef]
- Comino-Sanz, I.M.; López-Franco, M.D.; Castro, B.; Pancorbo-Hidalgo, P.L. The Role of Antioxidants on Wound Healing: A Review of the Current Evidence. JCM 2021, 10, 3558. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Xu, Y.; Zhang, K.; Liu, Y.; Liang, Q.; Thakur, A.; Liu, W.; Yan, Y. Plant-Derived Extracellular Vesicles (PDEVs) in Nanomedicine for Human Disease and Therapeutic Modalities. J Nanobiotechnology 2023, 21, 114. [Google Scholar] [CrossRef]
- He, B.; Cai, Q.; Qiao, L.; Huang, C.-Y.; Wang, S.; Miao, W.; Ha, T.; Wang, Y.; Jin, H. RNA-Binding Proteins Contribute to Small RNA Loading in Plant Extracellular Vesicles. Nat Plants 2021, 7, 342–352. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-López, C.M.; Manzaneque-López, M.C.; Pérez-Bermúdez, P.; Soler, C.; Marcilla, A. Characterization and Bioactivity of Extracellular Vesicles Isolated from Pomegranate. Food Funct. 2022, 13, 12870–12882. [Google Scholar] [CrossRef]
- Kim, M.; Park, J.H. Isolation of Aloe Saponaria-Derived Extracellular Vesicles and Investigation of Their Potential for Chronic Wound Healing. Pharmaceutics 2022, 14, 1905. [Google Scholar] [CrossRef]
- Kim, M.K.; Choi, Y.C.; Cho, S.H.; Choi, J.S.; Cho, Y.W. The Antioxidant Effect of Small Extracellular Vesicles Derived from Aloe Vera Peels for Wound Healing. Tissue Eng Regen Med 2021, 18, 561–571. [Google Scholar] [CrossRef]
Figure 1.
PDEVs biophysical characterization. (a) Size and distribution of PDEVs through NTA; (b) Distribution of PDEVs’ zeta potential
Figure 1.
PDEVs biophysical characterization. (a) Size and distribution of PDEVs through NTA; (b) Distribution of PDEVs’ zeta potential
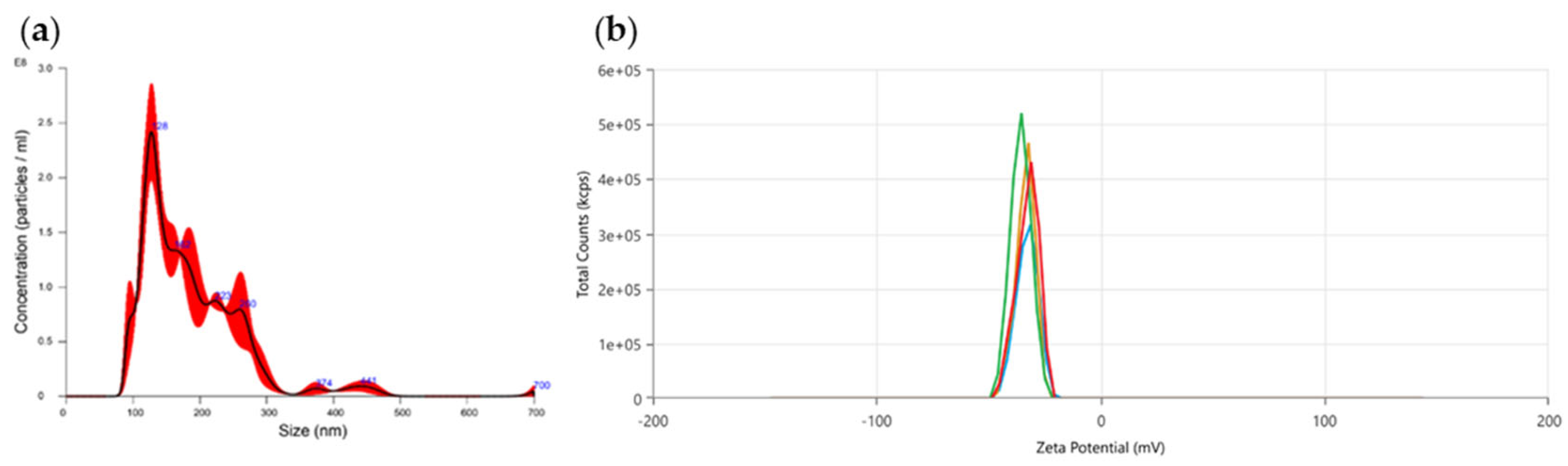
Figure 2.
PDEVs morphological characterization through TEM. (a) Round structure and membrane integrity of PDEVs with sizes between 50-80 nm; (b) Round structure and membrane integrity of PDEVs with sizes between 150-200 nm
Figure 2.
PDEVs morphological characterization through TEM. (a) Round structure and membrane integrity of PDEVs with sizes between 50-80 nm; (b) Round structure and membrane integrity of PDEVs with sizes between 150-200 nm
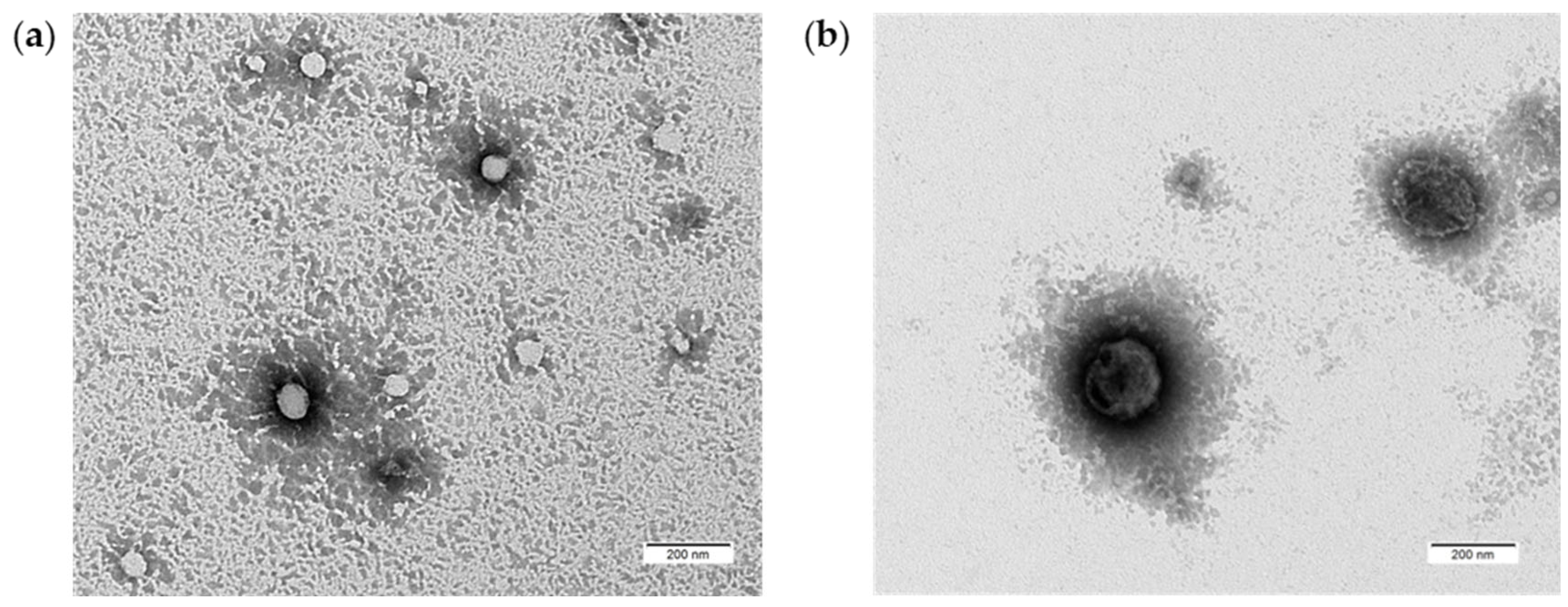
Figure 3.
Cellular uptake of Dil-PDEVs in Skin Fibroblast after (a) 24 hours of treatment; (b) 48 hours of treatment (c) 72 hours of treatment. PDEVs were labeled with Dil (red), and nuclei were counterstained with DAPI (blue).
Figure 3.
Cellular uptake of Dil-PDEVs in Skin Fibroblast after (a) 24 hours of treatment; (b) 48 hours of treatment (c) 72 hours of treatment. PDEVs were labeled with Dil (red), and nuclei were counterstained with DAPI (blue).
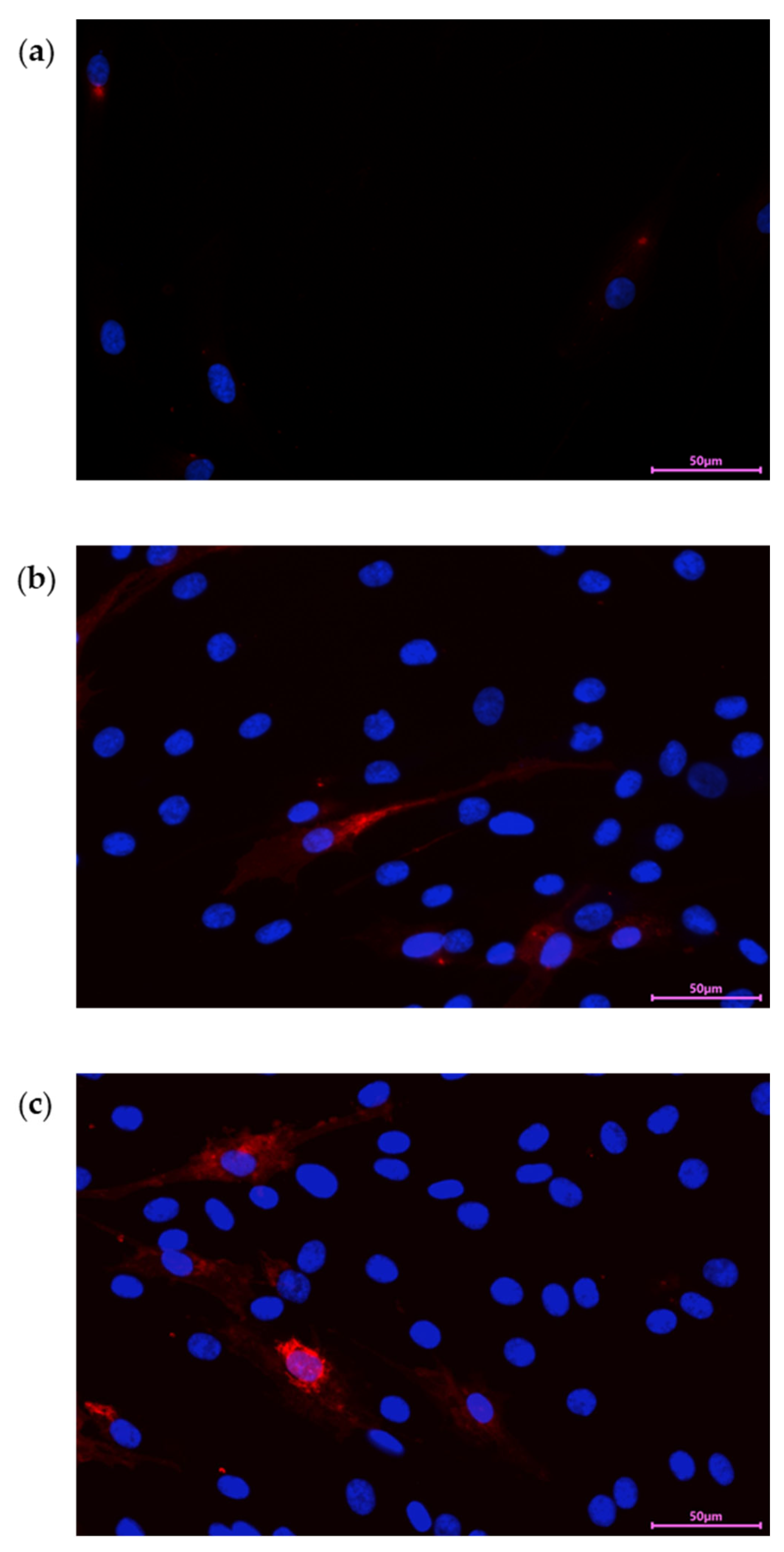
Figure 4.
PDEVs effect of mitochondrial metabolism. (a) Analysis of Mitochondrial Membrane Potential; (b) Analysis of Mitochondrial Anion Superoxide levels. Data are expressed as mean ± SE. *p<0,05, **p<0,01, ***p<0,001. Statistical analysis was performed using one-way ANOVA Bonferroni.
Figure 4.
PDEVs effect of mitochondrial metabolism. (a) Analysis of Mitochondrial Membrane Potential; (b) Analysis of Mitochondrial Anion Superoxide levels. Data are expressed as mean ± SE. *p<0,05, **p<0,01, ***p<0,001. Statistical analysis was performed using one-way ANOVA Bonferroni.
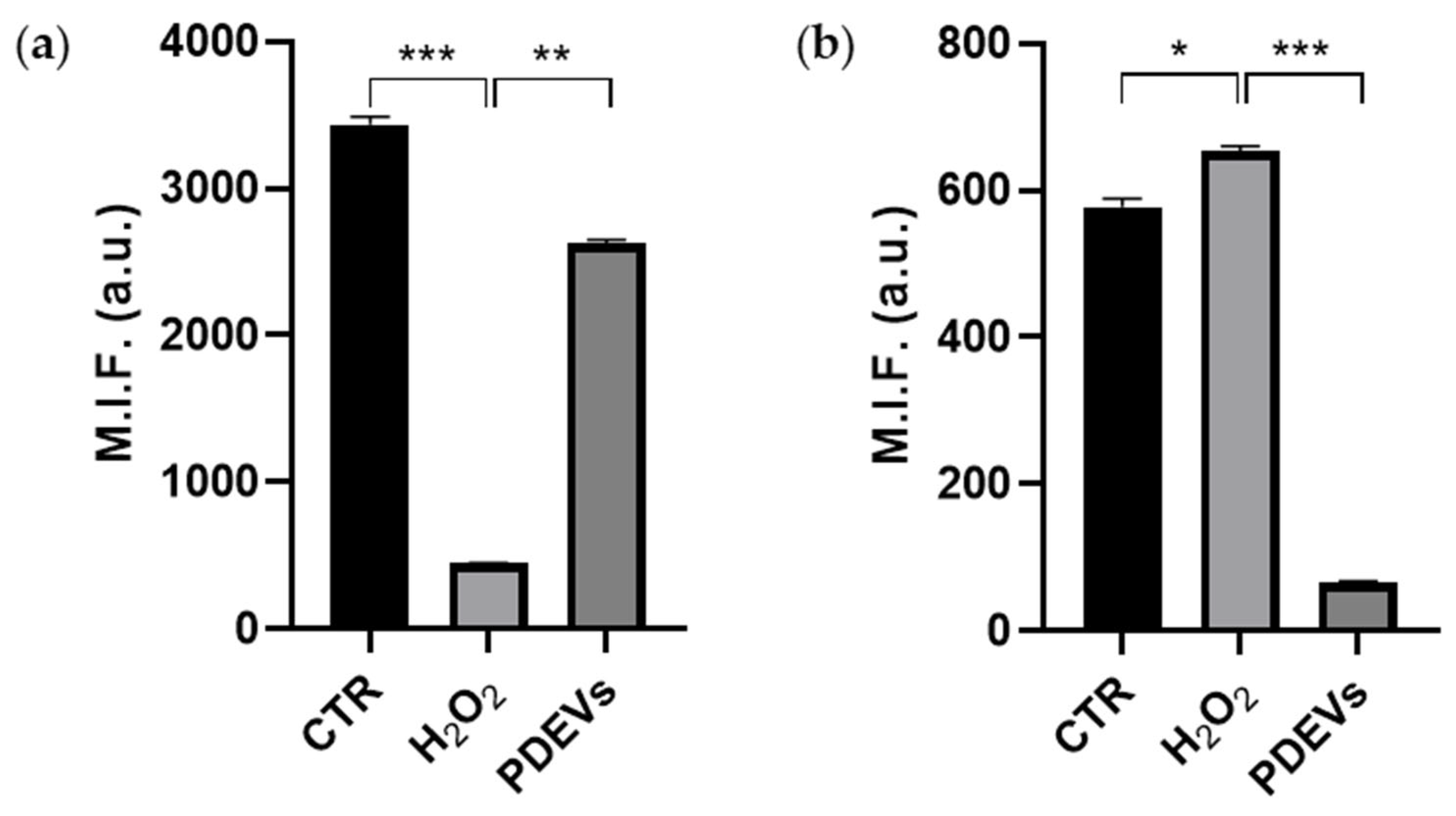
Figure 5.
Quantification of extracellular Sirtuin 1 expression. Data are expressed as mean ± SE. ****p<0,0001. Statistical analysis was performed using one-way ANOVA Bonferroni.
Figure 5.
Quantification of extracellular Sirtuin 1 expression. Data are expressed as mean ± SE. ****p<0,0001. Statistical analysis was performed using one-way ANOVA Bonferroni.
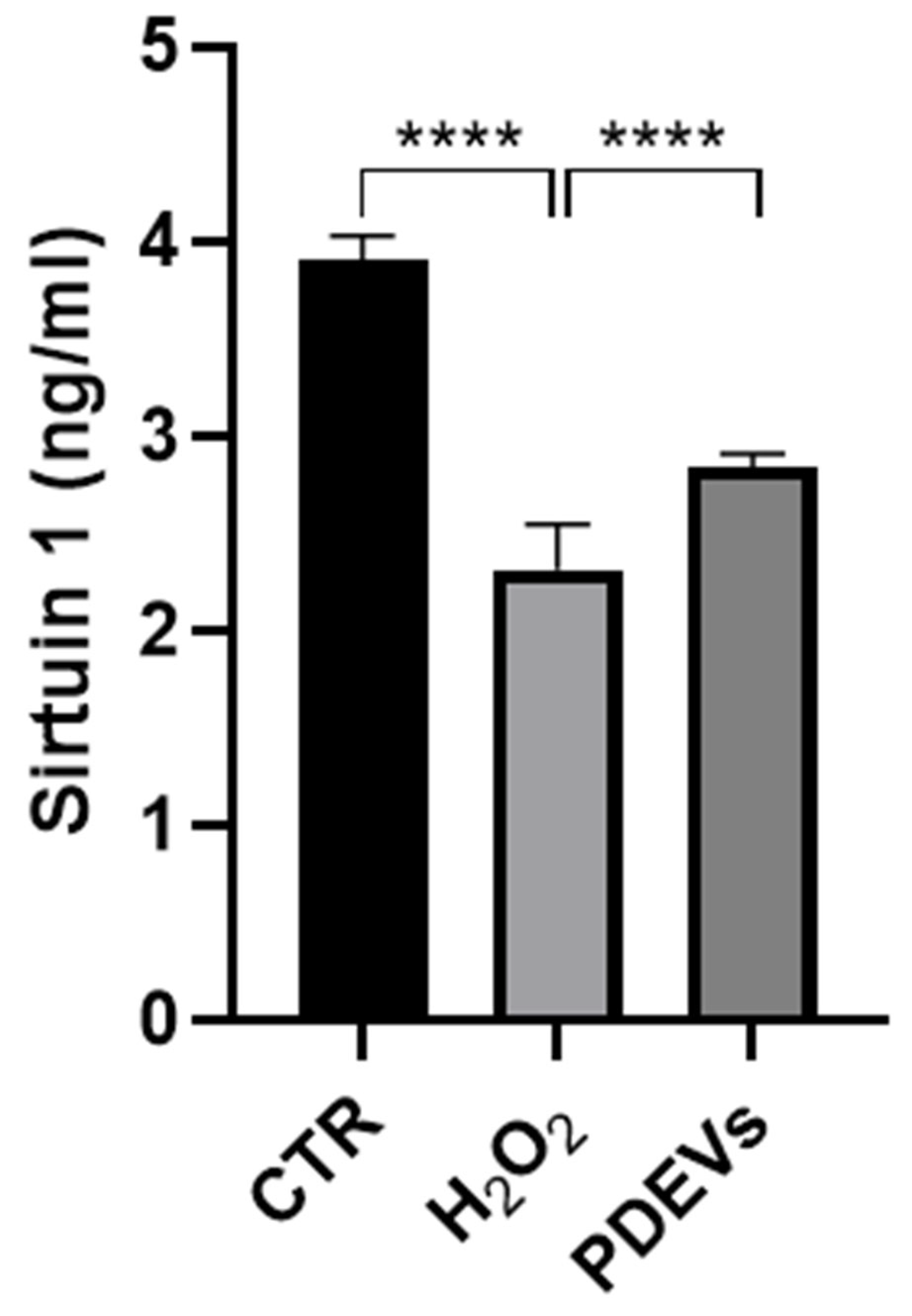
Figure 6.
Wound Healing Assay of Skin Fibroblasts. Representative images are shown from three independent experiments. Data are expressed as mean ± SE. ns: not significant; ****p<0,0001. Statistical analysis was performed using unpaired t-test (Student’s t-test).
Figure 6.
Wound Healing Assay of Skin Fibroblasts. Representative images are shown from three independent experiments. Data are expressed as mean ± SE. ns: not significant; ****p<0,0001. Statistical analysis was performed using unpaired t-test (Student’s t-test).
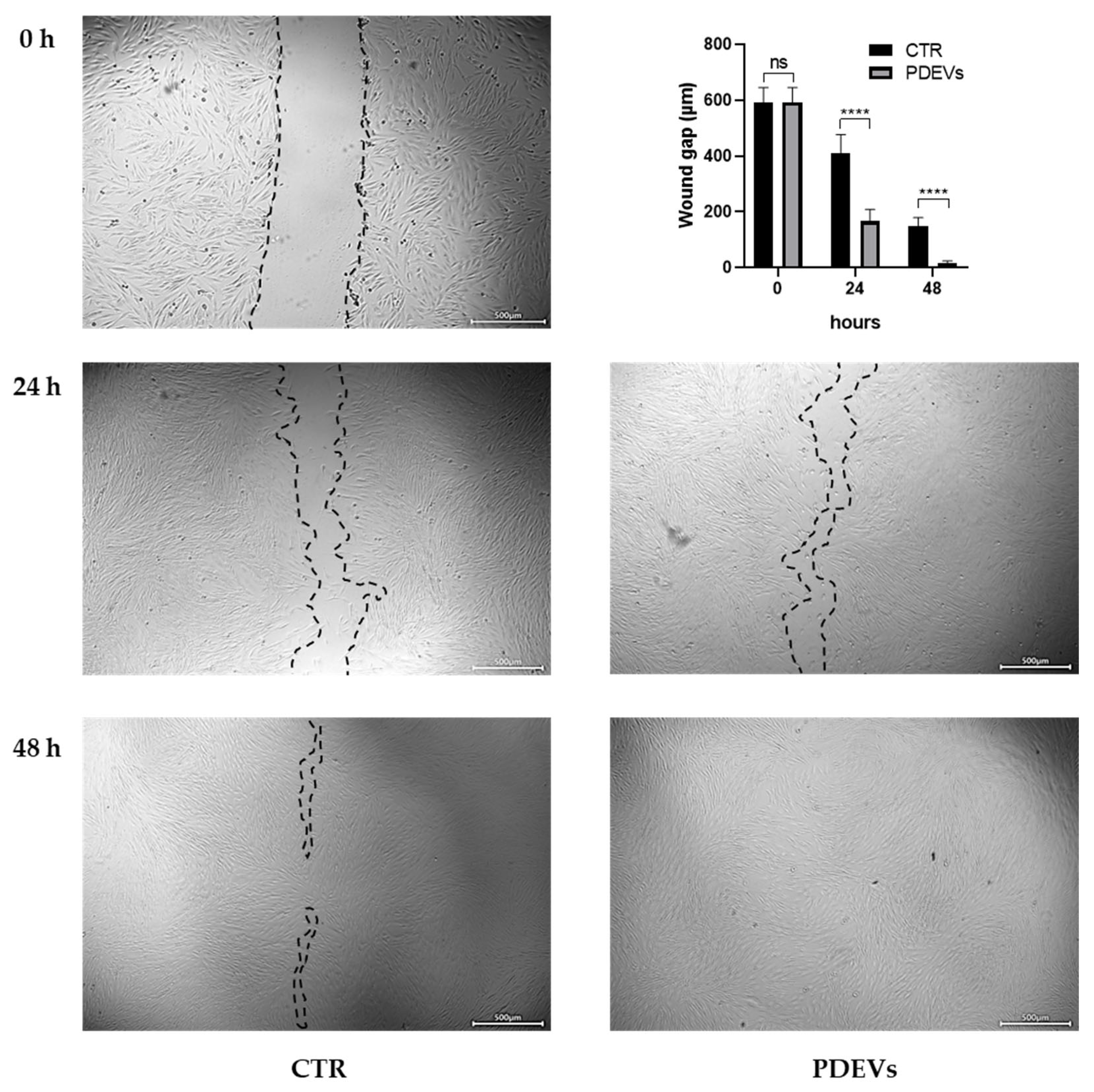
Figure 7.
MMP-9 expression in Skin Fibroblasts. Cells were stained with anti-MMP-9-FITC (green) and nuclei were counterstained with DAPI (blue).
Figure 7.
MMP-9 expression in Skin Fibroblasts. Cells were stained with anti-MMP-9-FITC (green) and nuclei were counterstained with DAPI (blue).
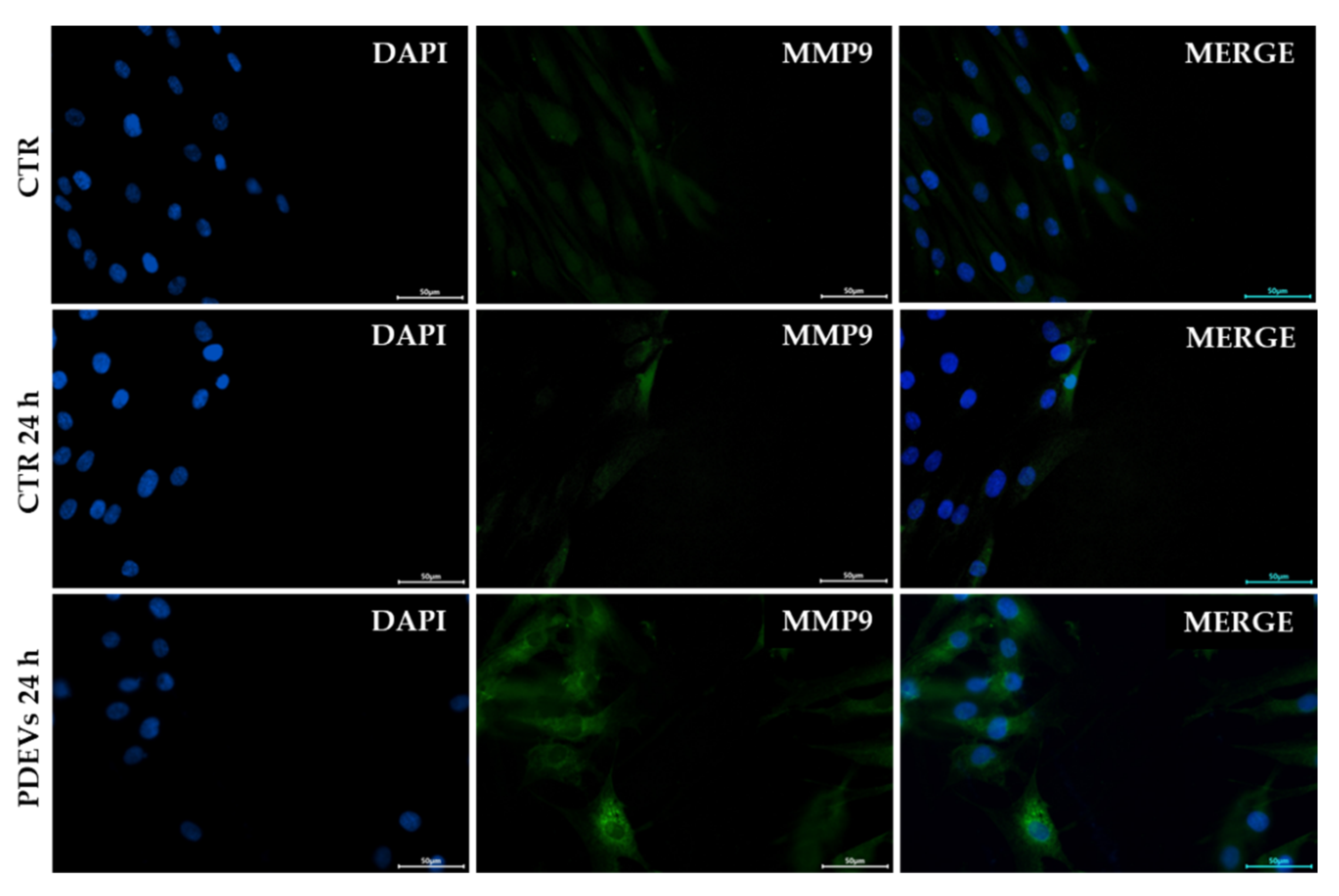
Figure 8.
Quantification of extracellular Vimentin. Data are expressed as mean ± SE. ****p<0,0001. Statistical analysis was performed using unpaired t-test (Student’s t-test).
Figure 8.
Quantification of extracellular Vimentin. Data are expressed as mean ± SE. ****p<0,0001. Statistical analysis was performed using unpaired t-test (Student’s t-test).
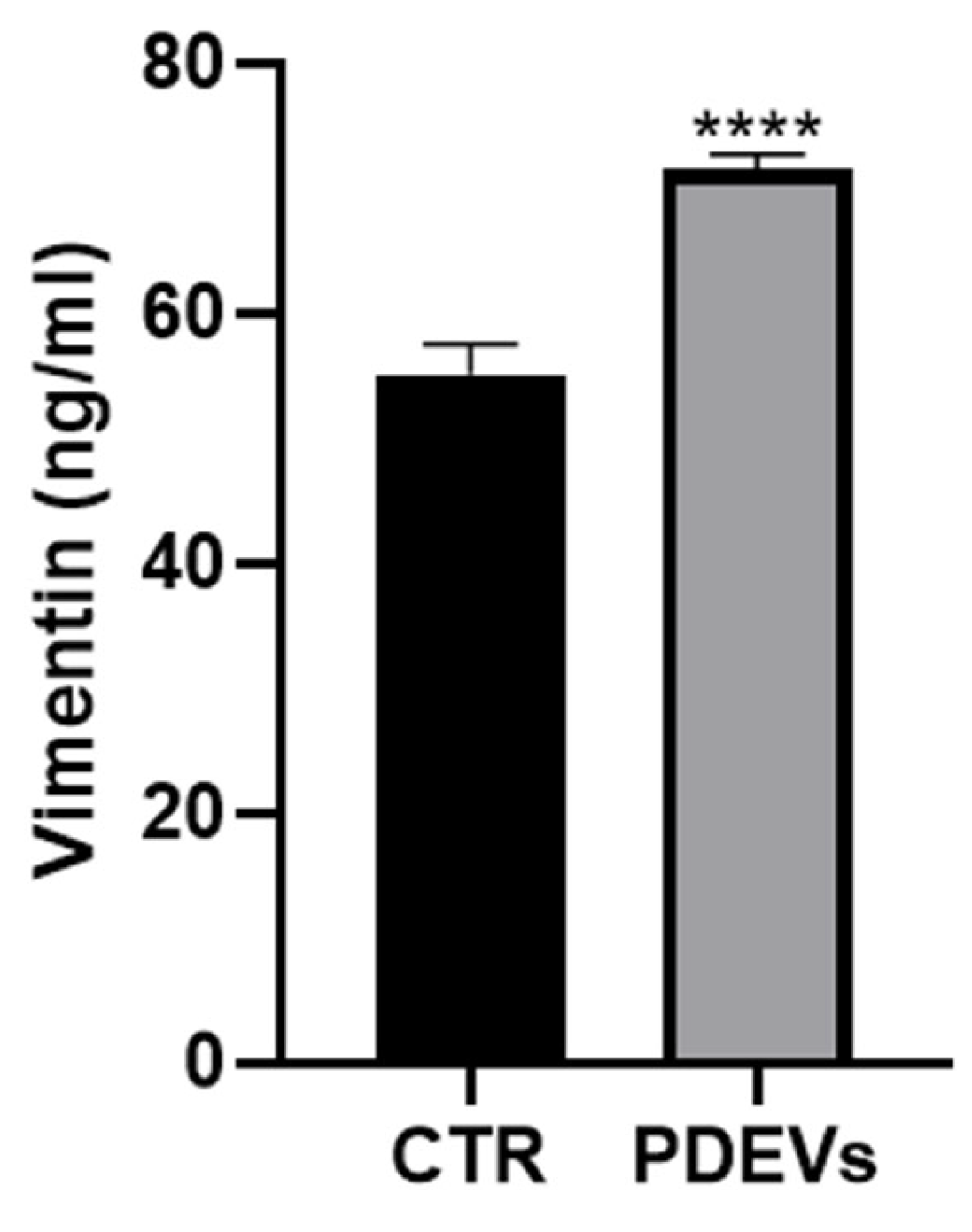

Table 1.
Quantification of bioactive compounds in PDEVs
| Bioactive compound | Concentration |
| Total Antioxidant Capacity | 2,3 ± 0,2 nMol/µL |
| Ascorbic Acid | 1910 ± 1 ng |
| ATP | 61,3 ± 16,6 mM |
| Catalase | 499,1 ± 2,2 mU/ml |
| Citric Acid | 37,67 ± 1,34 µmol/L |
| Glutathione | 11,8 ± 0,3 µM |
| SOD | 7392,00 ± 6,03 U/ml |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



